Embed Size (px)
Citation preview

THE JOURNAL OF EXPERIMENTAL ZOOLOGY 275:239-248 (1996)
Autoregulatory Index, Adrenergic Responses, and Interaction Between Adrenoreceptors and Prostacyclin in the Coronary System of Rainbow Trout
C . AGNISOLA, T. MUSTAFA, AND J.K. HANSEN Dipartimento di Fisiologia Generale ed Ambientale, Universita di Napoli "Federico II", I-80134 Naples, Italy (C.A.); and Institute of Biology, University of Odense, Campusvej 55, DK-5230 Odense M, Denmark (TM., J.K.H.)
ABSTRACT Some salient regulatory properties of the coronary system in nonworking heart preparation of trout are studied, in particular: (1) the pressure dependence of coronary resistance; (2) the reactivity to adrenergic compounds; and (3) the interaction between prostacyclin and spe- cific adrenergic agonists. Coronary resistance was not influenced by variation in ventricle weight or heart rate among various preparations used throughout this study. A positive autoregulatory index (ArI) was determined in the pressure range of 2.5-3.5 kPa, suggesting the existence of an active vasoconstrictive response probably involving local regulatory mechanisms. Noradrenaline was a powerful vasoconstrictor and prostacyclin (PG12) was a vasodilator when perfused sepa- rately. Both the ul-agonist phenylephrine and the u2-agonist clonidine induced dose-dependent vasoconstriction, while the P-agonist isoproterenol elicited a dose-dependent vasodilation. The P- antagonists propranolol (P-aspecific) and atenolol (P1-specific) induced vasoconstriction, with atenolol as the more potent (minimal effective concentration M; maximal effect > 50% vasoconstric- tion). These results suggest the existence of a P-adrenergic tone in the preparation. The vasoactiv- ity of PGIz was not affected by the u-agonists, while a significant vasoconstriction was observed at lo4 M in presence of 10" M of isoproterenol. This suggests the occurrence of a specific interaction between prostacyclin and P-adrenoreceptors in the coronary system of trout. 0 1996 Wiley-Liss, Inc.
A common view on the fish heart function is that the oxygen delivery from luminal blood to compact myocardium is not diffusion-limited and that coronary blood supply is only essential in con- ditions of high cardiac work (Davie and Farrell, '91a,b; Davie et al., '92; Gamperl et al., '94), with coronary perfusion pressure playing only a minor role in controlling the coronary flow (Davie and Farrell, '91a). However, recent studies on the coro- nary perfusion in vivo in salmonids indicate that coronary flow is related to the myocardial oxygen demand under various conditions, implying a fine control of coronary supply in which both perfu- sion pressure and coronary resistance appear to be involved (Axelsson and Farrell, '93; Gamperl et al., '94).
Part of the in vivo control of coronary flow may involve adrenergic as well as local mechanisms including pressure, temperature, metabolites and autocoids (Levick, '91). In vivo injection of adrena- line induces an increase in coronary flow, despite a coronary vasoconstriction, because of a significant increase in the dorsal aorta pressure (Axelsson and 0 1996 WILEY-LISS, INC.
Farrell, '93; Gamperl et al., '94). On the other hand, the a-antagonist phentolamine has scarce effects on coronary resistance in vivo, and local control mechanisms of coronary resistance have been proposed to explain the exercise-induced in- crease in coronary flow in coho salmon, which can- not be eliminated by adrenergic blockade (Axelsson and Farrell, '93). The possibility exists that a-adr- energic coronary vasoconstriction competes with metabolic vasodilation, a phenomenon well-known in the mammalian coronary system (Baumgart et al., '93; Miyashiro and Feigl, '93). Local mecha- nisms have been also proposed to be involved in active responses t o pressure changes in vitro (Agnisola, '93; Mustafa and Agnisola, '94). The mechanical factors associated with perfusion pres- sure modification are known to induce liberation of various factors, including prostanoids, from the vas- cular wall (Zusman, '86; Karwatowska-Prokopczuk
Address reprint requests to T. Mustafa, Biologisk Institut, Odense University, Campusvej 55, DK-5230 Odense M, Denmark.

240 C. AGNISOIA ET AL.
et al., '89). However, it8 is not clear if a real auto- regulative response occurs in the trout coronary system, and a more significant evaluation of this phenomena, e.g., in terms of autoregulation in- dex (Norris et al., '79) is necessary.
A putative role of prostanoids in the local con- trol of coronary resistance in trout has been re- cently proposed (Muetafa et al., ,921, and the vasoactivity of prostanoids normally found in the trout blood, like prostacyclin and thromboxane Az (Mustafa and Jensen, '92), has been reported to- gether with the interesting evidence of significant interactions between these compounds and nora- drenaline (Mustafa and Agnisola, '94). In particu- lar, the vasoconstriction induced by noradrenaline was relieved by lo-' htl carbocyclic thromboxane A2 (CTA,, a stable analogue of TXA2 and a potent positive agonist in the mammalian vasculature, Nicolaou et al., '80), which by itself elicited vaso- constriction at higher concentrations, and was po- tentiated by lo-' M prostacyclin (PGIz), which by itself elicited vasodilation at higher concentra- tions. A main question concerns the type of adreno- receptors involved in tihis interaction, if any. In contrast with the coronary artery ring prepara- tions, where noradrenaline is vasodilator (Small et al., 'go), the vasoconstriction induced by this catecholamine in the whole coronary system, to- gether with its sensitivity to both a- and P-adren- ergic antagonists (Mustafa and Agnisola, '94), suggests the presence of both a- and P-adrenore- ceptors with the predoniinance of a-receptors. The use of specific a- and P-agonists would address the above question.
Aims of the present study are: (1) to reevaluate the in vitro coronary pressure-flow relationship in the nonworking heart preparation of trout, by calculating the autoregulation index using data previously published (Mustafa and Agnisola, '94); and (2) to investigate the involvement of a- and P-adrenoreceptors in their interaction with the prostanoid prostacyclin .
MATERJALS AND METHODS Animals
Forty-nine specimens of rainbow trout (Onco- rhynchus rnykiss) of both sexes, obtained from a local fish farm, were held in tanks with running tap water at 12 2 0.5"C, fed twice a week with commercial trout pellets and allowed to acclimate for 10 days before the experiments were per- formed. Their mean weight was 0.33 2 0.01 kg (mean k S.E.).
Isolated heart preparation and coronary perfusion
The heart was isolated and cannulated accord- ing to Mustafa and Agnisola ('94) and mounted in a jacketed chamber maintained at 10 k 0.1"C with a water bath. The coronary artery was perfused with a constant head pressure. The perfusate draining into the atrium from the coronary sys- tem was the only flow entering the atrium while output pressure was set to zero. Hearts were spon- taneously beating at constant rate. There was no signifcant change in heart rate in the various pro- tocols used.
The perfusate solution was the Cortland saline as modified by Farrell et al. ('86). Its composition was (in g 1-'1: NaCl, 7.25; KCl, 0.23; MgS04.7H20, 0.23; NaH2P04.H20, 0.016; Na2HP04.2H20, 0.41; CaCl2-2H2O, 0.37; glucose, 1.0; polyvinylpyrroli- done (PVP), 10.0. The saline was gassed with 99.5% O2 and 0.5% C02, and the pH adjusted to 7.9 at 10°C with NaHC03. Throughout the experi- mental protocols the perfusate was gassed with the above mixture.
Chemicals PVP (Mw 40000), noradrenaline, phenyleph-
rine, clonidine, propranolol, isoproterenol, salbuta- mol and atenolol were from Sigma Chemical Co., St. Louis, MO. Solutions were freshly prepared in distilled water and diluted in the above Ringer as required. Care was taken to avoid photodegra- dation of light-sensitive drugs by covering contain- ers with foil. Carba prostacyclin (carba-PG12, a stable analogue of prostacyclin) was from Cayaman Chemicals, Ann Arbor, MI. Desired dilution of this prostanoid from the stock solution (1 mg ml-I in ethanol) contained less than 0.03% ethanol. This ethanol concentration does not affect the coronary resistance (Mustafa and Agnisola, '94). All other reagents were the highest grade available.
Experimental protocols Dose-response curves of adrenergic agonists and antagonists
A constant head pressure of 3.0 kPa was main- tained throughout the experiment. After about 30 min of stabilization period, the basal flow was measured and the perfusion was shifted to the drug-containing perfusate. The following are the adrenergic agonists tested: noradrenaline (a$ ago- nist), phenylephrine (a, agonist), clonidine (a2 ago- nist), isoproterenol (P,,P2 agonist), and salbutamol (p2 agonist), while the antagonists were: propra-

ADRENORECEPTORS AND PROSTACYCLIN 241
nolol (p antagonist) and atenolol (PI antagonist). A stepwise increase of concentration in the range 10-9-10-5 M was performed for all the agonists, while the ranges 10-6-10-3 M and 10-19-10-5 M were used for propranolol and atenolol, respec- tively. Each step was 10 min in duration and coro- nary flow was measured after 5 rnin and at the end of the step. At any given concentration, 5 rnin of perfusion were generally sufficient to elicit a measurable effect.
Effects of carba-PGI, in presence of adrenergic agonists
The dose-response curve of prostacyclin (10-l'- M) was determined in absence and in pres-
ence of M of each of the above listed agonists. After the initial setting and stabilization as in the first protocol, the perfusion medium was replaced with the agonist-containing saline. After 10 min of perfwion and flow measurement, the dose-response curve of carba-PG12 was determined through a step- wise increase of concentration. Carba-PG12 solu- tions contained M of the appropriate agonist.
Measurement and calculations The atrial pressure and the input coronary pres-
sure were continuously measured through saline- filled sidearms connected to a Uniflow pressure transducer (Baxter, Morton Grove, IL, USA). The transducer was connected to a computer for di- rect data acquisition. Pressure measurements were referenced to the level of saline in the per- fusion chamber and expressed in kPa. Coronary pressure was corrected for cannula resistance. Atrial pressure was negligible and not considered in the calculation of coronary resistance. Heart rate was determined by visual direct counting.
The coronary flow was determined by measur- ing the time it took for 0.05 ml of perfusate to pass through the coronary artery, according to Agnisola et a]. ('94). The coronary resistance (TPa s m-3, corresponding to 16.7 kPa min ml-l) was determined as: mean coronary pressure (kPa) x 0.06koronary flow (ml min-l).
The autoregulation index (ArI) was calculated according to Norris et al. ('79) as:
ArI l-[(AF/Fi)/(AP/PJ]
where F is the flow at pressure P, and Fi and Pi are the initial flow and pressure, respectively. The autoregulation index as a function of perfusion pressure has been calculated using the data pre- viously published by Mustafa and Agnisola ('94),
using the pressure range 1.43.8 kPa, and with an initial pressure of 2.5 kPa, giving an initial flow of 0.62 ml m i d kg-l.
In the dose-response curves, the response was expressed in terms of percent change in coronary resistance. One-way ANOVA test has been used to evaluate the significance of the curves. Two- way ANOVA was used to compare different per- fusion conditions. Student's t-test (P < 0.05) was used to compare means. The arcsin data trans- formation was used to perform statistics on the percentage values.
RESULTS
Basal coronary resistance and its dependence from heart size and heart rate
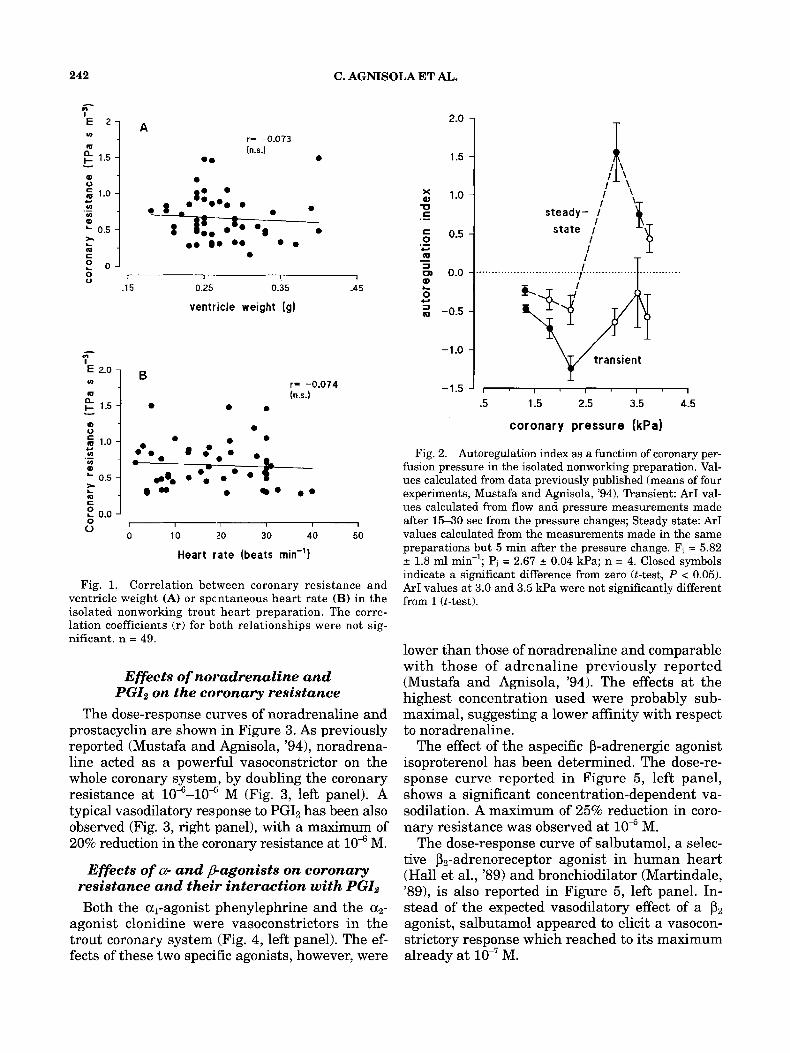
In Figure 1A the correlation between control coronary resistance and ventricle weight is pre- sented. It is evident that in our preparation and in the range of animal size used the resistance was independent from the weight of the ventricle. On this basis the resistance values in this study were not normalized. The mean basal resistance calculated from all preparations was 0.66 L 0.05 TPa s m-3.
Heart rate did not significantly affect the coro- nary resistance (Fig. 1B). Isolated preparations were spontaneously and regularly beating and the rate was highly variable among preparations. However, the absence of correlation, which can easily be explained by the fact that the heart is nonworking, allows the comparison between dif- ferent groups of preparations, excluding second- ary effects of chronotropism.
Autoregulation index
Figure 2 reports the autoregulatory index (ArI) as a function of coronary pressure calculated re- evaluating the data reported by Mustafa and Agnisola ('94) in the same kind of preparation here used. Both during transient and under steady state, the ArI increased when the pressure was higher than 2.5 kPa. Interestingly, at all the pressure values tested, ArI increased within 5 min from the pressure change. The pressure de- pendence of ArI under transient and steady- state conditions was significantly different (two-factor ANOVA with replication, P < 0.01). However, ArI values positive and not signifi- cantly different from 1 only occurred in a rela- tively narrow range of pressures (3.0-3.5 kPa) under steady-state conditions.
242 C. AGNISOLA ET AL.
- 0
a r= -0.073 1n.s.I
1.5 .. . - . al f 1
i j 2 0
. ~
0 0
I I I I
.15 0.25 0.35 A5
ventricle weight Igl
e i . -
2 0.0 1 0
I I I t
O E , 10 20 30 40 50
Heart rate (beats rnin-'1
Fig. 1. Correlation between coronary resistance and ventricle weight (A) or spc'ntaneous heart rate (B) in the isolated nonworking trout heart preparation. The corre- lation coefficients (r) for both relationships were not sig- nificant. n = 49.
Effects of noradrenaline and PG12 on the coronary resistance
The dose-response curves of noradrenaline and prostacyclin are shown in Figure 3. As previously reported (Mustafa and Agnisola, '94), noradrena- line acted as a powerful vasoconstrictor on the whole coronary system, by doubling the coronary resistance at 104-104' M (Fig. 3, left panel). A typical vasodilatory response to PG12 has been also observed (Fig. 3, right panel), with a maximum of 20% reduction in the coironary resistance at lo4 M.
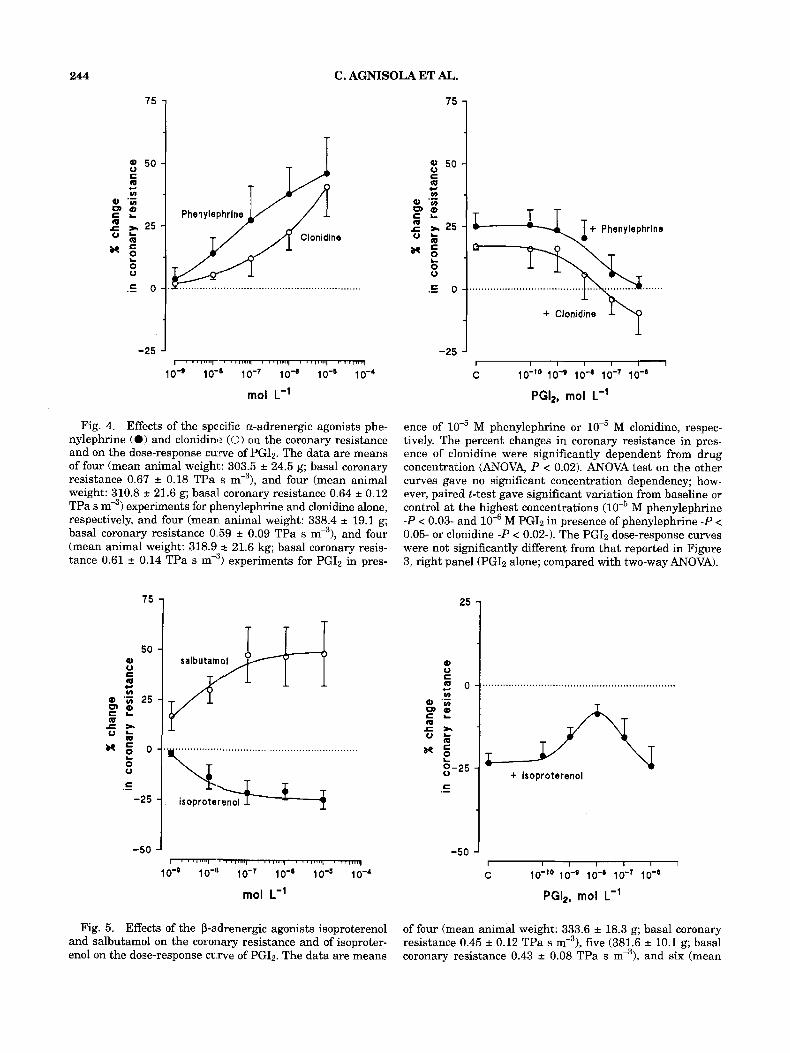
Effects of a- and P-agonists on coronary resistance and their interaction with PG12 Both the al-agonist ;phenylephrine and the a2-
agonist clonidine were vasoconstrictors in the trout coronary system (Fig. 4, left panel). The ef- fects of these two specific agonists, however, were
2-o 1 1.5 4 T
I \ 1 il ', I ' 2 1.0 - I '
-0 e .- state I
I 0.5 - .- CI Q =I -
transient
L
CI 0
-0.5 -
-1.0 -
-1.5 I I I ' I I
.5 1.5 2.5 3.5 4.5
coronary pressure (kPa1
Fig. 2. Autoregulation index as a function of coronary per- fusion pressure in the isolated nonworking preparation. Val- ues calculated from data previously published (means of four experiments, Mustafa and Agnisola, '94). Transient: ArI val- ues calculated from flow and pressure measurements made after 15-30 sec from the pressure changes; Steady state: ArI values calculated from the measurements made in the same preparations but 5 min after the pressure change. Fi = 5.82 2 1.8 ml min-'; Pi = 2.67 2 0.04 Wa; n = 4. Closed symbols indicate a significant difference from zero (t-test, P < 0.05). ArI values a t 3.0 and 3.5 W a were not significantly different from 1 (t-test).
lower than those of noradrenaline and comparable with those of adrenaline previously reported (Mustafa and Agnisola, '94). The effects at the highest concentration used were probably sub- maximal, suggesting a lower affinity with respect to noradrenaline.
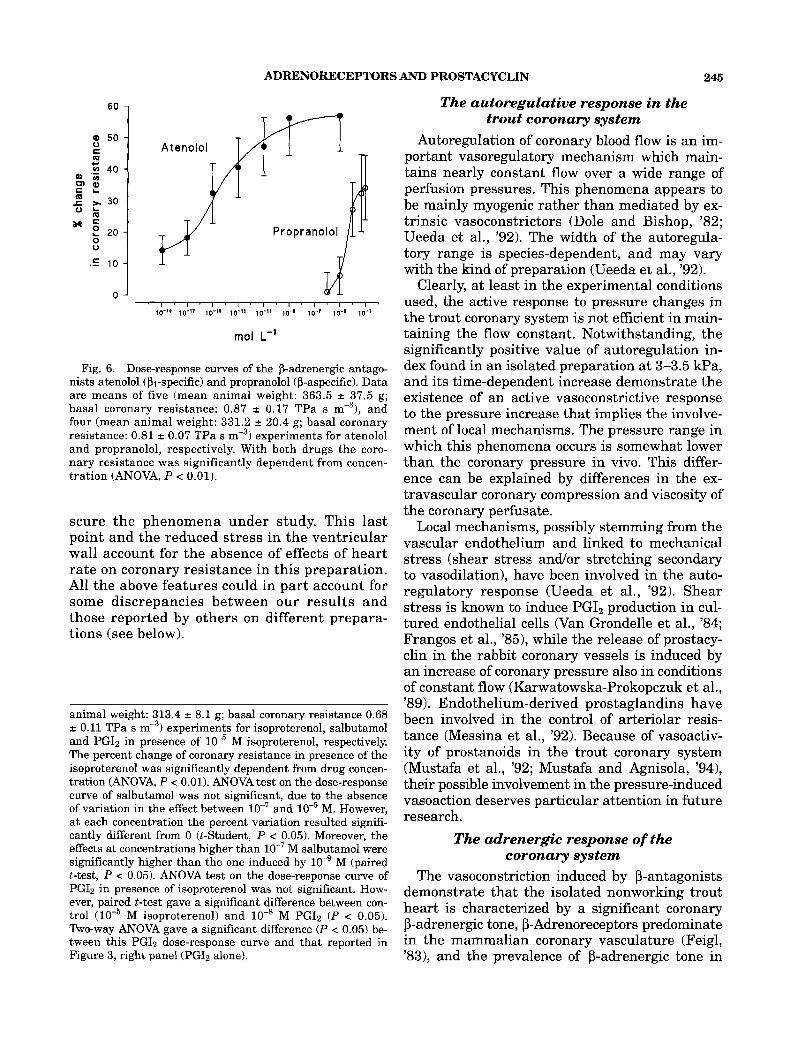
The effect of the aspecific P-adrenergic agonist isoproterenol has been determined. The dose-re- sponse curve reported in Figure 5, left panel, shows a significant concentration-dependent va- sodilation. A maximum of 25% reduction in coro- nary resistance was observed at 10" M.
The dose-response curve of salbutamol, a selec- tive P2-adrenoreceptor agonist in human heart (Hall et al., '89) and bronchiodilator (Martindale, '89), is also reported in Figure 5, left panel. In- stead of the expected vasodilatory effect of a P2 agonist, salbutamol appeared to elicit a vasocon- strictory response which reached to its maximum already at M.

ADRENORECEPTORS AND PROSTACYCLIN 243
150 1 125 4
20 1 Q 0 C $100 - ul
u .G 0 0 C L
2 2 7 5 - 0 I U = 2
50- c .-
25 -
0 -
u 0 e $ 0 ul
0) ' E m a e L m
L
$-20 c .-
PGI?, rnol L"
Fig. 3. Dose-response curves of noradrenaline and PGIz in the coronary system of the isolated nonworking trout heart. The data are the means (2 S.E.) of five (mean animal weight: 290.5 2 13.1 g; basal coronary resistance 0.77 2 0.11 TPa s m-3), and four (mean animal weight: 325.0 2 11.8 g; basal coronary resistance 0.35 2 0.05 TPa s m-3) experiments for
noradrenaline and PGI2, respectively. A sigmoidal regression was applied to the dose-response curve of noradrenaline and a X50 of 5.5 x mol L-l was calculated. The percent change in coronary resistance was significantly de- pendent from drug concentration (one-way ANOVA norad- renaline, P < 0.01; PGI2, P < 0.05).
2 1.36 x
In Figure 4, right panel, the dose-response curves of PG12 in presence of M of the cx-ago- nists phenylephrine and clonidine are reported. In presence of both phenylephrine and clonidine, PG12 elicited the typical vasodilative response, completely relieving, at the highest concentration (10" M), the vasoconstriction induced by the a- agonists. In presence of M isoproterenol, the effect of PG12 appeared to be completely different (Fig. 5 , right panel). A significant vasoconstriction at lo-* M PGIz was observed, which was replaced by vasodilation at the highest concentration. This result suggests the existence of a specific interac- tion between prostacyclin and P-adrenoreceptors in the coronary system of trout.
PAdrenergic tone in the coronary system The dose-response curves of the aspecific p-an-
tagonist propranolol and of the specific P,-antago- nist atenolol are reported in Figure 6. Both antagonists displayed a significant vasoconstric- tory action. Atenolol, in particular, was very po- tent, being significantly effective at concentrations as low as M and producing a maximal effect higher than 50%. The vasoconstriction induced by these inhibitors was much higher than the vasodi- lation induced by isoproterenol (25%, see Fig. 5,
left panel). These results clearly indicate the pres- ence of a P-adrenergic tone in the preparation.
DISCUSSION There are two main observations reported in
this study: (1) there is a narrow range of coro- nary perfusion pressure in which a positive auto- regulatory index can be evaluated in the trout coronary system; and (2) the interaction between prostacyclin and adrenergic systems is confirmed; apparently, this interaction involves the P-adreno- receptors.
The present study used the same experimental preparation previously used to demonstrate the occurrence of interaction between noradrenaline and prostanoids in the coronary system of trout (Mustafa and Agnisola, '94), i.e., the isolated non- working heart. Advantages of this preparation, which does no external work, are the following: (1) resistance vessels are included in the prepa- ration; (2) the oxygen demand of the heart is low, which increases its ability to tolerate the perfu- sion with a perfusate whose oxygen content is lower than that of the blood; and (3) the low work performed by the heart reduces the probability that changes in oxygen consumption will cause metabolic coronary vasodilation that would ob-
244 C. AGNISOLA ET AL.
75
50 E Q
...........................................................
I
-25
." I" I " I" I "
ma1 L"
Fig. 4. Effects of the specific a-adrenergic agonists phe- nylephrine (0) and clonidine (0) on the coronary resistance and on the dose-response curve of PG12. The data are means of four (mean animal weight: 303.5 i 24.5 g; basal coronary resistance 0.67 f 0.18 TPa s m-3), and four (mean animal weight: 310.8 f 21.6 g; basall coronary resistance 0.64 f 0.12 TPa s m3) experiments for phenylephrine and clonidine alone, respectively, and four (mean animal weight: 338.4 19.1 g; basal coronary resistance 0.59 i 0.09 TPa s m-3), and four (mean animal weight: 318.9,* 21.6 kg; basal coronary resis- tance 0.61 2 0.14 TPa s m-'? experiments for PGIz in pres-
75
50 a 0 e Q c ul
Q '5 25 m a l C L rn z *
0 b 0
c .- -25
L - 4 I+ Phenylephrine
'7 + Clonidine
I I I I I I 1
C 10-o 10-e 1 0 - ~ lo-' PGlz, mol L"
ence of M phenylephrine or M clonidine, respec- tively. The percent changes in coronary resistance in pres- ence of clonidine were significantly dependent from drug concentration (ANOVA, P < 0.02). ANOVA test on the other curves gave no significant concentration dependency; how- ever, paired t-test gave significant variation from baseline or control at the highest concentrations ( M phenylephrine - P < 0.03- and M PGIz in presence of phenylephrine -P < 0.05- or clonidine -P < 0.02-1. The PGIz dose-response curves were not significantly different from that reported in Figure 3, right panel (PGI2 alone; compared with two-way ANOVA).
l.
? T
.........
-50 ' I I I I I I I
C 10-0 10-8 lo-' lo-'
PGI,, rnol L"
Fig. 5. Effects of the p-adrenergic agonists isoproterenol and salbutamol on the coronary resistance and of isoproter- enol on the dose-response c u v e of PGI2. The data are means
of four (mean animal weight: 333.6 f 18.3 g; basal coronary resistance 0.45 2 0.12 TPa s m-3), five (381.6 i 10.1 g; basal coronary resistance 0.43 f 0.08 TPa s m3), and six (mean
ADRENORECEPTORS AND PROSTACYCLIN 245
rnol L-'
Fig. 6. Dose-response curves of the P-adrenergic antago- nists atenolol (PI-specific) and propranolol (P-aspecific). Data are means of five (mean animal weight: 363.5 -c 37.5 g; basal coronary resistance: 0.87 2 0.17 TPa s m-3), and four (mean animal weight: 331.2 = 20.4 g; basal coronary resistance: 0.81 2 0.07 TPa s m-3) experiments for atenolol and propranolol, respectively. With both drugs the coro- nary resistance was significantly dependent from concen- tration (ANOVA, P < 0.01).
scure the phenomena under study. This last point and the reduced stress in the ventricular wall account for the absence of effects of heart rate on coronary resistance in this preparation. All the above features could in part account for some discrepancies between our results and those reported by others on different prepara- tions (see below).
animal weight: 313.4 -c 8.1 g; basal coronary resistance 0.68 2 0.11 TPa s mF3) experiments for isoproterenol, salbutamol and PGIz in presence of M isoproterenol, respectively. The percent change of coronary resistance in presence of the isoproterenol was significantly dependent from drug concen- tration (ANOVA, P i 0.01). ANOVA test on the dose-response curve of salbutamol was not significant, due to the absence of variation in the effect between and M. However, at each concentration the percent variation resulted signifi- cantly different from 0 @-Student, P < 0.05). Moreover, the effects at concentrations higher than M salbutamol were significantly higher than the one induced by M (paired t-test, P < 0.05). ANOVA test on the dose-response curve of PGI2 in presence of isoproterenol was not significant. How- ever, paired t-test gave a significant difference between con- trol M isoproterenol) and lo-' M PGI2 (P < 0.05). Two-way ANOVA gave a significant difference ( P < 0.05) be- tween this PGI2 dose-response curve and that reported in Figure 3, right panel (PGI2 alone).
The autoregulative response in the trout coronary system
Autoregulation of coronary blood flow is an im- portant vasoregulatory mechanism which main- tains nearly constant flow over a wide range of perfusion pressures. This phenomena appears to be mainly myogenic rather than mediated by ex- trinsic vasoconstrictors (Dole and Bishop, '82; Ueeda et al., '92). The width of the autoregula- tory range is species-dependent, and may vary with the kind of preparation (Ueeda et al., '92).
Clearly, at least in the experimental conditions used, the active response to pressure changes in the trout coronary system is not efficient in main- taining the flow constant. Notwithstanding, the significantly positive value of autoregulation in- dex found in an isolated preparation at 3-3.5 kPa, and its time-dependent increase demonstrate the existence of an active vasoconstrictive response to the pressure increase that implies the involve- ment of local mechanisms. The pressure range in which this phenomena occurs is somewhat lower than the coronary pressure in vivo. This differ- ence can be explained by differences in the ex- travascular coronary compression and viscosity of the coronary perfusate.
Local mechanisms, possibly stemming from the vascular endothelium and linked to mechanical stress (shear stress andor stretching secondary to vasodilation), have been involved in the auto- regulatory response (Ueeda et al., '92). Shear stress is known to induce PGIz production in cul- tured endothelial cells (Van Grondelle et al., '84; Frangos et al., '851, while the release of prostacy- clin in the rabbit coronary vessels is induced by an increase of coronary pressure also in conditions of constant flow (Karwatowska-Prokopczuk et al., '89). Endothelium-derived prostaglandins have been involved in the control of arteriolar resis- tance (Messina et al., '92). Because of vasoactiv- ity of prostanoids in the trout coronary system (Mustafa et al., '92; Mustafa and Agnisola, '94), their possible involvement in the pressure-induced vasoaction deserves particular attention in future research.
The adrenergic response of the coronary system
The vasoconstriction induced by P-antagonists demonstrate that the isolated nonworking trout heart is characterized by a significant coronary P-adrenergic tone, 0-Adrenoreceptors predominate in the mammalian coronary vasculature (Feigl, '831, and the prevalence of P-adrenergic tone in

246 C. AGNISOLA ET AL.
the coronary arterial microvessels has also been reported (Sekiguchi et al., '92). Interestingly, in the trout coronary system the vasoconstrictive ef- fect of P-antagonists is much higher than the va- sodilation induced in the same preparation by the aspecifk a-adrenergic antagonist phentolamine (Mustafa and Agnisola. '94). On the other hand, the highly vasoconstrirtive action of noradrena- line suggests that the a-adrenoreceptors dominate the P-receptors when thle adrenergic receptor sys- tem of coronaries is aspecifically stimulated. Ap- parently, both al-(phenylephrine treatment) and a2-receptors (clonidine treatment) appear to con- tribute to this vasoconstriction. This contrasts with the results found in the coronary artery rings (Small et al., ,901, in which a vasodilation induced by both catecholamines has been reported, with no effect of phenylephrine. It is well-known from mammalian studies, however, that the response of isolated vessels may be affected by in vitro con- ditions such as ionic content, pH, temperature, or the condition of the endothelium and changes in the prostaglandin synt'hesis. The discrepancy be- tween the results of the present study and those on isolated coronary arteries may also be due to differences in the size of the vessels. In the prepa- ration used here, resistance vessels are included. In mammals, large and small coronary arteries re- spond differently to caitecholamines (Zuberbuhler and Bohr, '65).
The higher effect of atenolol in respect to pro- pranolol, suggests that, in contrast with the coro- nary arterial microvessels of mammals (Sekiguchi et al., '921, the P,-adrenergic tone predominates, although the difference in the potency of the two antagonists (see Fig. 6) may limit this conclusion. Surprisingly, salbutamol, a known &-agonist, acts as P-antagonist in the trout coronary system by inducing a strong vasoconstriction. This result may be due to: (i) P2-adlrenoreceptors in fish have different structure than those occurring in mam- mals; and (ii) salbutamd may have different prop- erties at 10°C than 2.t 37°C normally used in mammalian studies. The unusual response of salbutamol in trout coronary system confirms the existence of species specificity in the characteris- tics of the adrenoreceptor system of fishes and stresses caution when using specific mammalian- derived pharmacological tools in lower vertebrates (Bjenning et al., '93). A.t present, spatial distribu- tion (i.e., receptor presence in endothelium and vascular smooth muscle) and subclassification of P-adrenoreceptors even in mammalian coronary vasculature is a controversial issue. Radioligand re-
ceptor binding studies indicated the presence of P1- and P,-adrenoreceptor subtypes in pig (Schwartz and Velly, '83), canine (Vatner et al., ,861, and bo- vine (Nakane et al., '88) coronary arteries. In con- trast, in vitro experiments have confirmed only P,-adrenoreceptors in the coronary artery of dog (O'Donnel and Wanstall, '84), rabbits (De et al., '74), cows (Purdy et al., '88) and rats (Nyborg and Mikkelson, '85). Inconsistencies in the above re- sults may be due either to the fact that P-agonist and antagonist drugs used in different studies were not selective enough for one class of receptors or may have resulted from the use of different spe- cies and different preparation and methodologies.
Interaction between isoproterenol and PGIz The results reported here indicate that the in-
crease in vasoconstriction noradrenaline depen- dence in the presence of prostacyclin, previously reported (Mustafa and Agnisola, '941, is due to the reduced p-activation. The interesting aspect of the interaction between isoproterenol and prostacy- clin is that a vasoconstriction is the result of the simultaneous action of two vasodilators. Indeed, the interaction can be seen as a PGI,-induced at- tenuation of the vasodilation mediated by P-adreno- receptors. The mechanism of this phenomena could be complex. A simple direct inhibition of P-adreno- receptors by prostacyclin is probably not involved, as PGIB alone does not induce vasoconstriction at any of the concentrations tested despite the fact that the preparation is characterized by a signifi- cant P-adrenergic tone (see above).
Two possibilities can be suggested: (1) prosta- cyclin alters the sensitivity of P-receptors to spe- cific agonists; in this case the results available imply that, while in the basal tone P1-adrenore- ceptors appear to be dominating, the P,-adreno- receptors should be those involved in both the activation by isoproterenol and the block by pros- tacyclin; and (2) the activation of P-adrenorecep- tors alters the response of coronary vessels to prostacyclin. The fact that only the lower concen- trations of prostacyclin affect the isoproterenol re- sponse, while higher concentrations induce the typical vasodilation, could indicate the involve- ment of competitive mechanisms.
P-Adrenoreceptors activate adenylate cyclase through Gs proteins (Kobilka, '92). P-Adrenergic relaxation in mammalian coronary arteries is en- hanced by inhibition of prostaglandin metabolism and inhibited by its stimulation (Rubanyi and Paul, '84; Rubanyi and Vanhoutte, '85). However, the relationship between this effect and CAMP lev-

ADRENORECEPTORS AND PROSTACYCLIN 247
els is not clear and a compartmentalization of CAMP responses in vascular smooth muscle has been proposed (Rubanyi et al., '86), similar to that occurring in the myocardial cell, where there are two pools of CAMP-dependent protein kinase A that are also differently distributed between cy- tosolic and particulate fractions (Hayes et al., '80). A selective activation of these two pools by iso- proterenol and PGEl has been observed (Hayes et al., '80), and their role in the differential re- sponses to PI and P2 receptors has been suggested (Xiao and Lakatta, '93). Prostaglandins like PGEl and PG12 are known to activate adenylate cyclase and relaxation mechanisms mediated by CAMP (Metsa-Ketela, '81). Alteration in the mobilization of intracellular calcium stores has been involved in the synergy between the thromboxane A2 re- ceptors and several vasoactive substances (Cocks et al., '93). Thromboxane A2 is also able to inter- fere with the adrenergic stimulation of coronary vessels (Mustafa and Agnisola, '94). Clearly, in- terference between prostacyclin and P-receptors in trout coronary vessels can occur at different lev- els and hrther study is necessary for elucidation.
ACKNOWLEDGMENTS During part of the course of this study, C.A. was
at the Institute of Biology, Odense University, as a Visiting Scientist. The stay of C.A. and the study were funded by a research program from the Dan- ish Natural Science Research Council 9401633.
LITERATURE CITED Agnisola, C. (1993) The coronary circulation in teleosts and
elasmobranchs: Open problems. In: The Vertebrate Gas Transport Cascade: Adaptation to Environment and Mode of Life. E. Bicudo and M. Glass, eds. CRC Press, Boca Ra- ton, pp. 224-232.
Agnisola, C., R. Venzi, D. Houlihan, and B. Tota (1994) Coro- nary flow-pressure relationship in the working isolated fish heart: trout (Oncorhynchus mykiss) versus torpedo (Torpedo marmorata). Phil. Trans. Roy. SOC. Lond. B, 343:189-198.
Axelsson, M., and A.P. Farrell(1993) Coronary blood flow in vivo in the coho salmon (Oncorhynchus kisutchj. Am. J. Physiol., 264:R963-R97 1.
Baumgart, D., T. Ehring, P. Kowallik, B.D. Guth, M. Krajcar, and G. Heusch (1993) Impact of a-adrenergic coronary vasoconstriction on the transmural myocardial blood flow dis- tribution during humoral and neuronal adrenergic activa- tion. Circ. Res., 73:869-886.
Bjenning, C., S. Holmgren, and A.P. Farrell (1993) Neu- ropeptide Y potentiates contractile response to norepi- nephrine in skate coronary artery. Am. J. Physiol.,
Cocks, T.M., B.K. Kemp, D. Pruneau, and J.A. Angus (1993) Comparison of contractile responses to 5-hydroxytryptamine and sumatriptan in human isolated coronary artery: Syn- ergy with the thromboxane A2-receptor agonist, U46619. Br. J. Pharmacol., 110:360-368.
265:H661-H665.
Davie, P.S., and A.P. Farrell (1991a) The coronary and lumi- nal circulations of the myocardium of fishes. Can. J. Zool.,
Davie, P.S., and A.P. Farrellr1991b) Cardiac performance of an isolated heart preparation from the dogfish (Squalus acanthias): The effects of hypoxia and coronary artery per- fusion. Can. J. Zool., 69:1822-1828.
Davie, P.S., A.P. Farrell, and C.E. Franklin (1992) Cardiac performance of an isolated eel heart: Effects of hypoxia and responses to coronary artery perfusion. J. Exp. Zool.,
De, L.A., I.S. Lande, J.A. Harvey, and S. Holt (1974) Response of the rabbit coronary arteries to autonomic agents. Blood Vessels, 11:319-337.
Dole, W.P., and V.S. Bishop (1982) Influence of autoregula- tion and capacitance on diastolic coronary artery pressure- flow relationships in the dog. Circ. Res., 51~261-270.
Farrell, A.P., K.R. MacLeod, and B. Chancey (1986) Intrinsic mechanical properties of the perfused rainbow trout heart and the effects of catecholamines and extracellular calcium under control and acidotic conditions. J. Exp. Biol., 125:3 19-345.
Feigl, E.O. (1983) Coronary physiolom. Physiol. Rev., 63:l-205. Frangos, J.A., S.G. Eskin, L.V. McIntire, and C.L. Ives (1985)
Flow effects on prostacyclin production by cultured human endothelial cells. Science, 227:1477-1479.
Gamperl, A.K., A.W. Pinder, R.R. Grant, and R.G. Boutilier (1994) Influence of hypoxia and adrenaline administration on coronary blood flow and cardiac performance in seawa- ter rainbow trout (Oncorhynchus mykiss). J. Exp. Biol.,
Hall, J.A., M.C. Petch, and M.J. Brown (1989) Intracoronary injections of salbutamol demonstrate the presence of func- tional Pa-adrenoreceptors in the human heart. Circ. Res., 65546-553.
Hayes, J.S., L.L. Brunton, and S.E. Mayer (1980) Selective activation of particulate CAMP-dependent protein kinase by isoproterenol and prostaglandin El. J. Biol. Chem., 255:5113-5119.
Karwatowska-Prokopczuk, E., G. Ciabattoni, and A. Wennmalm (1989) Effects of hydrodynamic forces on coronary pro- duction of prostacyclin and purines. Am. J. Physiol., 256:H1532-1538.
Kobilka, B. (1992) Adrenergic receptors as models for G pro- tein-coupled receptors. Ann. Rev. Pharmacol. Toxicol.,
Levick, J.R. (1991) An Introduction to Cardiovascular Physi- ology. Butterworths, London.
Martindale (1989) The Extra Pharmacopoeia. J.E.F. Reynolds, ed. The Pharmaceutical Press, London, pp. 1480-1483.
Messina, E.J., D. Sun, A. Koller, M.S. Wolin, and G. Kaley (1992) Role of endothelium-derived prostaglandins in hy- poxia-elicited arteriolar dilation in rat skeletal muscle. Circ. Res., 71:790-796.
Metsa-Ketela, T. (1981) Cyclic AMP-dependent and -indepen- dent effects of prostaglandins on the contraction-relaxation cycle of spontaneously beating isolated rat atria. Acta Physiol. Scand., 112:481-485.
Miyashiro, J.K., and E.O. Feigl (1993) Feedforward control of coronary blood flow via coronary a-receptor stimulation. Circ. Res., 73:252-263.
Mustafa, T., and C. Agnisola (1994) Vasoactivity of prostanoids in the trout (Oncorhynchus mykiss) coronary system: Modification by noradrenaline. Fish Physiol. Hiochem., 13:249-261.
69~1993-200 1.
262:113-121.
193:209-232.
32:87-114.

248 C. AGNISOLA ET AL.
Mustafa, T., and F.B. Jensen (1992) Effect of hypoxia on in uiuo thromboxane and prostacyclin levels in arterial blood of rainbow trout, Oncorhync,hus mykiss (Walbaum). J. Fish. Biol., 40:303-306.
Mustafa, T., C. Agnisola, and B. Tota (1992) Myocardial and coronary effects of exogenous arachidonic acid on the iso- lated and perfused heart pi-eparation and its metabolism in the heart of trout ( O ~ C O F ~ ~ ~ C ~ U S mykiss). Comp. Bio- chem. Physiol., 103C:163-167.
Nakane, T., G. Tsujimoto, K. Hashimoto, and S. Chiba (1988) P-adrenoreceptors in the cartine large coronary arteries: PI adrenoreceptors predominate in vasodilation. J . Pharmacol. Exp. Ther., 245:936-943.
Nicolaou, K.C., R.L. Magolda, ,and D.A. Claremon (1980) Syn- thesis of thromboxane A2 analogs. In: Advances in Prostag- landin and Thromboxane Research. Vol. 6. B. Samuelsson, P. Ramwell, and P. Paoletti, eds. Raven Press, New York, pp. 481483.
Norris, C.P., G.E. Barnes, E.E. Smith, and H.J. Granger (1979) Autoregulation of superior mesenteric flow in fasted and fed dogs. Am. J. Physiol., 237:H174-H177.
Nyborg, N.C.B., and E.O. Mikkelson (1985) Characterization of P-adrenoreceptor subtypes in isolated ring preparations of intramural rat coronary arteiies. J . Cardiovasc. Pharmacol.,
O'Donnell, S.R., and J.C. Waristall (1984) The classification of P-adrenoreceptors in isolai,ed ring preparations of canine coronary arteries. Br. J. Phalmacol., 81:637-644.
Purdy, R.E., G.L. Stupecky, and RR. Coulombe (1988) Fur- ther evidence for a homogeneous population of @I-adreno- receptors in bovine coronary artery. J . Pharmacol. Exp. Theor., 245:67-71.
Rubanyi, G., and R.J. Paul (1994) 02-sensitivity of P-adrener- gic responsiveness in isolated bovine and porcine coronary arteries. J. Pharmacol. Exp. ?her., 230:692-698.
Rubanyi, G., and P.M. Vanhoiitte (1985) Inhibitors of pros- taglandin synthesis augmen ; P-adrenergic responsiveness in canine coronary arteries. Circ. Res., 56:117-125.
7:1113-1117.
Rubanyi, G., P. Galvas, J. DiSalvo, and R.J. Paul (1986) Eicosanoid metabolism and P-adrenergic mechanisms in coro- nary arterial smooth muscle: Potential compartmentation of CAMP. Am. J. Physiol., 250:C406-C412.
Schwartz, J . , and J. Velly (1983) The P-adrenoreceptor of pig coronary arteries: Determination of and P2 sub- types by radioligand binding. Br. J. Pharmacol., 79: 409-414.
Sekiguchi, N., H. Kanatsuka, T. Komaru, K. Akai, K. Sato, Y. Wang, M. Sugi, K. Ashikawa, and T. Takishima (1992) Ef- fects of a and P adrenergic blockade on coronary arterial microvessels in the beating canine heart. Cardiovasc. Res.,
Small, S.A., C. MacDonald, and A.P. Farrell (1990) Vascular reactivity of the coronary artery in rainbow trout (Onco- rhynchus mykiss). Am. J. Physiol., 258:R1402-R1410.
Ueeda, M., S.K. Silvia, and R.A. Olsson (1992) Nitric oxide modulates coronary autoregulation in the guinea pig. Circ. Res., 70:1296-1303.
Van Grondelle, A., G.S. Worhten, D. Ellis, M.M. Mathias, R.C. Murphy, R.J. Strife, J.T. Reeves, and N.F. Voelkel (1984) Altering hydrodynamic variables influences PGI2 produc- tion by isolated lung and endothelial cells. J. Appl. Physiol.,
Vatner, D.E., D.R. Knight, C.J. Homcy, S.F. Vatner, and M.A. Young (1986) Subtypes of P-adrenergic receptors in bovine coronary arteries. Circ. Res., 59:463-473.
Xiao, R.-P., and E.G. Lakatta (1993) PI-adrenoceptor stimu- lation and P2-adrenoceptor stimulation differ in their effects on contraction, cytosolic Ca", and Ca2+ current in single rat ventricular cells. Circ. Res., 73:286300.
Zuberbuhler, R.C., and D.F. Bohr (1965) Responses of coronary smooth muscle to catecholamines. Circ. Res., 16:431440.
Zusman, R.M. (1986) Eicosanoids-Prostaglandins, thrombox- ane and prostacyclin. In: Heart and Cardiovascular Sys- tem, Vol. 2. H.A. Fozzard, R.B. Jennings, E. Haber, A.M. Katz, and H.E. Morgan, eds. Raven Press, New York, pp.
26:4 15-42 1,
57:388-395.
1613-1630.





























