Embed Size (px)
Citation preview

Biochemical Pharmacology xxx (2014) xxx–xxx
G Model
BCP-12021; No. of Pages 12
Review
Autophagy: An adaptive metabolic response to stress shaping theantitumor immunity
Elodie Viry, Jerome Paggetti, Joanna Baginska, Takouhie Mgrditchian,Guy Berchem, Etienne Moussay 1, Bassam Janji 1,*
Laboratory of Experimental Hemato-Oncology, Department of Oncology, Public Research Center for Health, Luxembourg City, Luxembourg
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
2. Autophagy induction as adaptive metabolic response under stresses. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
2.1. Autophagy activation under hypoxia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
2.2. Autophagy activation under nutrient starvation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3. Effect of autophagy on modulating immune cells function in cancer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.1. Innate immunity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.1.1. Mast cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.1.2. Macrophages . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.1.3. Neutrophils . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.1.4. Dendritic cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.2. Adaptive immunity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.2.1. B lymphocytes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
3.2.2. T lymphocytes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
A R T I C L E I N F O
Article history:
Received 27 May 2014
Accepted 9 July 2014
Available online xxx
Keywords:
Autophagy
Immune response
Hypoxia
Tumor resistance
Metabolism
A B S T R A C T
Several environmental-associated stress conditions, including hypoxia, starvation, oxidative stress, fast
growth and cell death suppression, modulate both cellular metabolism and autophagy to enable cancer
cells to rapidly adapt to environmental stressors, maintain proliferation and evade therapies. It is now
widely accepted that autophagy is essential to support cancer cell growth and metabolism and that
metabolic reprogramming in cancer can also favor autophagy induction. Therefore, this complex
interplay between autophagy and tumor cell metabolism will provide unique opportunities to identify
new therapeutic targets. As the regulation of the autophagic activity is related to metabolism, it is
important to elucidate the exact molecular mechanism which drives it and the functional consequence
of its activation in the context of cancer therapy. In this review, we will summarize the role of autophagy
in shaping the cellular response to an abnormal tumor microenvironment and discuss some recent
results on the molecular mechanism by which autophagy plays such a role in the context of the anti-
tumor immune response. We will also describe how autophagy activation can behave as a double-edged
sword, by activating the immune response in some circumstances, and impairing the anti-tumor
immunity in others. These findings imply that defining the precise context-specific role for autophagy in
cancer is critical to guide autophagy-based therapeutics which are becoming key strategies to overcome
tumor resistance to therapies.
� 2014 Elsevier Inc. All rights reserved.
Contents lists available at ScienceDirect
Biochemical Pharmacology
jo u rn al h om epag e: ww w.els evier .c o m/lo cat e/b io c hem p har m
* Corresponding author.
E-mail address: [email protected] (B. Janji).1 Co-principal investigators.
Please cite this article in press as: Viry E, et al. Autophagy: An adaptive metabolic response to stress shaping the antitumor immunity.Biochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.006
http://dx.doi.org/10.1016/j.bcp.2014.07.006
0006-2952/� 2014 Elsevier Inc. All rights reserved.

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx2
G Model
BCP-12021; No. of Pages 12
4. Effect of autophagy on tumor cell response to immune system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
4.1. Autophagy induction modulates cancer cell secretome and surface proteome . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
4.1.1. Damage associated molecular patterns (DAMPs) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
4.1.2. Antigen presentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
4.1.3. Cytokines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
4.1.4. Extracellular vesicles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
4.2. Autophagy induction in cancer cell provides an intrinsic mechanism of resistance to immune response. . . . . . . . . . . . . . . . . . . . . . 000
5. Autophagy modulators as therapeutic strategy for improving anticancer immunotherapy. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
6. Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 000
1. Introduction
Cancer cells display multiple levels of metabolic alterationswhich sustain cell proliferation and survival through theactivation of oncogenic pathways and/or the inhibition of tumorsuppressors. It is becoming increasingly clear that rapid cellproliferation creates areas within the tumor site with limiteddiffusion of oxygen and nutrients, leading to the establishment ofabnormal microenvironmental conditions such as hypoxia, lowpH and/or nutrient deprivation. In addition, the structurally andfunctionally abnormal tumor vasculature, combined to intrinsi-cally altered tumor cell metabolism, can also create spatial andtemporal heterogeneity in oxygenation, pH, concentrations ofglucose and many other metabolites. In turn, such abnormaltumor microenvironment has a major role in determining themetabolic phenotype of tumor cells [1,2]. In this context, tumorcells activate additional adaptive mechanisms which driveprofound metabolic reprograming in order to overcome theenvironmental constraints. These adaptations optimize tumorcell metabolism for proliferation by providing support for thethree basic needs of dividing cells: (i) rapid ATP generation tomaintain energy status, (ii) increased biosynthesis of macro-molecules, and (iii) tightened maintenance of appropriatecellular redox status [2]. It is well described that cancer cellsutilize several mechanisms to adapt stress conditions. The tumorcell response to hypoxic stress is the best studied, as hypoxiaalters several molecular pathways, among which autophagy, thatinfluence cellular metabolism [3–5].
Macroautophagy (hereafter referred to as autophagy) is one ofthe main mechanisms by which tumor cells adapt unfavorabletumor microenvironment [6,7]. The term of autophagy refers to theprocess by which cellular components are degraded through thelysosomal enzymatic pathway providing cells with essential aminoacids, nucleotides, and fatty acids that enable production ofelements required for energy and macromolecule biosynthesis[8,9]. Autophagy is a highly conserved catabolic process withcritical functions in maintenance of cellular homeostasis undernormal growth conditions and in preservation of cell viabilityunder stress [10,11]. Autophagy is characterized by the sequestra-tion of cytoplasmic constituents and organelles into double-membraned vacuoles called autophagosomes, followed by selec-tive degradation in lysosomes, thereby recycling energy andbuilding blocks for the synthesis of new biomolecules. Whileselective autophagy requires cargo proteins, autophagy can benon-selective, leading to lysosomal processing of bulk cytoplasmicmaterial. The implication of autophagy in the regulation of cellmetabolism has been recently reported, thus highlighting a newfunction of autophagy as a regulator of cell metabolism [12].Indeed, the observation that autophagy regulates lipid metabolismhas extended our understanding of the delicate and indispensablerole that autophagy plays in cellular metabolism and holdspromise of a new and exciting strategy for treating metabolicdisorders [13]. Furthermore, the intimate link between autophagy
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
and cancer cell metabolism has received widespread attention andis considered a topic of growing interest.
While the role of autophagy in the chemo-resistance isrelatively well described [14], the present review will focus onthe emerging role of autophagy as a mechanism of adaptation totumor microenvironment stresses and therefore its involvement inshaping the anti-tumor immune response. We will also discuss thecomplex and the context-dependent role of autophagy in tumorimmunity by providing an overview of its role in either theactivation or the inhibition of the anti-tumor immune response.Furthermore, we will briefly address how the development ofautophagy modulators is becoming a highly clinically relevanttopic in translational cancer therapies.
2. Autophagy induction as adaptive metabolic response understresses
Autophagy is activated in response to multiple stresses duringcancer progression, including hypoxia, nutrient deprivation,extracellular matrix detachment, and endoplasmic reticulum(ER) stress [15,16]. The role of autophagy within the tumormicroenvironment is still controversial. Thus, it has been reportedthat autophagy can be correlated with an increase [17] or adecrease [18] of tumor cell survival.
Briefly, the first step of the autophagic process, callednucleation, consists in the formation of the phagophore or isolationmembrane which depends mainly on Beclin 1 (BECN1)-Vps15 corecomplexes. Subsequently, the phagophore is elongated by severalAutophagy-related proteins (ATG). During this elongation step,microtubule-associated protein 1 light chain 3 (LC3)-I is lipidatedto LC3-II. Next, the phagophore is maturated primarily upon theaction of LC3-II and BECN1 proteins. This maturation leads to theformation of an enclosed vesicle called autophagosome. Finally,the autophagosome fuses with the lysosome to form an autolyso-some. Following this fusion, the internal material of the autophagicvacuole is degraded by the lysosomal hydrolases [19].
2.1. Autophagy activation under hypoxia
Autophagy is activated in cells as response to hypoxicmicroenvironment. Hypoxia can stimulate autophagy in a hypox-ia-inducible factor-1 (HIF-1)-dependent or -independent manner.HIF-1 is a transcription factor formed by two subunits, HIF-1a and-b, that regulates a large number of genes involved in themetabolic and vascular adaptation of cells to oxygen restriction[20]. Under hypoxia, HIF-1a is stabilized and its heterodimeriza-tion with HIF-1b allows the binding of the transcription factor tohypoxia response-elements (HREs) in target genes [21]. The genecoding for the Bcl-2/adenovirus E1B 19 kDa protein-interactingprotein 3 (BNIP3) contains two HRE sites in its promoter region,HRE1 and HRE2. It has been demonstrated that HIF-1 directly bindsto HRE2 site to induce the expression of BNIP3 [22]. The induction
e metabolic response to stress shaping the antitumor immunity.006

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 3
G Model
BCP-12021; No. of Pages 12
of BNIP3 leads to the disruption of the complex BECN1-Bcl-2, andthe subsequent release of BECN1 promotes the activation ofautophagy.
HIF-1-independent induction of autophagy can be mediatedthrough different signaling pathways. The mammalian target ofrapamycin (mTOR) signaling, that regulates cell growth andmetabolism in response to hypoxic environment, regulatesautophagy in a HIF-1a independent manner. Upon hypoxia, mTORsignaling is inhibited and protein synthesis is downregulated.Indeed, mTOR signaling consists of two major pathways mediatedby the specific mTOR complexes mTORC1 and mTORC2 [23]. It hasbeen reported that mTORC1 negatively controls autophagy by theinhibition of protein kinase ATG1 involved in the formation ofautophagosomes [24].
An additional level of regulation of autophagy via the inhibitionof mTOR signaling implicates the tuberous sclerosis complex (TSC).TSC is a heterodimeric complex formed by TSC1 and TSC2 whichacts as inhibitor of mTORC1 activity [25]. The 50AMP-activatedprotein kinase (AMPK) is a heterotrimeric complex encoded by severalgenes and is the primary energy sensor in cells. Upon its activationthrough the increase in the AMP/ATP ratio, AMPK phosphorylatesmany downstream targets, including TSC2. TSC2 phosphorylation onserine residues 1270 and 1388 enhances the activity of the TSC1/TSC2complex, and thereby blocks the Ras homolog enriched in brain(RHEB)-dependent activation of mTOR [26,27].
Another signaling pathway that activates autophagy in a HIF-1a-independent manner is the endoplasmic reticulum (ER) stresswhich activates the unfolded protein response (UPR). The UPR ismediated by three ER stress sensors: PKR-like ER kinase (PERK), ERto nucleus signaling 1 (ERN1), and activating transcription factor(ATF) 6 [28]. For example, by inducing PERK-dependent phos-phorylation of the eukaryotic translation initiation factor 2 alpha(elF2a), hypoxia activates autophagy by transcriptional inductionof LC3 through the expression of ATF4 [29]. Furthermore, UPRactivation leads to the induction of the transcription of Unc51-likekinase 1 (ULK1), which is an important regulator of autophagyinitiation and progression [30].
2.2. Autophagy activation under nutrient starvation
In response to nutrient deprivation, cancer cells activateautophagy as an alternative metabolic pathway to sustain cancerprogression. Autophagy allows the catabolism of existing cyto-plasmic components that provide essential components, thusserving to maintain cell survival [31].
Several studies have demonstrated that starvation-inducedautophagy is regulated by reactive oxygen species (ROS) whichcomprises superoxide ion (O2
�), hydrogen peroxide (H2O2), andhydroxyl radical (OH). ROS activate AMPK [32] leading to inhibitionof mTOR pathway through ULK1 which subsequently activatesautophagy flux [33]. Furthermore, it has been recently reportedthat starvation-induced autophagy can be regulated by microRNAs(miRNAs). Indeed, autophagy induction by serum starvation seemsto be inhibited by the overexpression of miR-376a which targets keyautophagy proteins ATG4C and BECN1 in MCF-7 breast adenocarci-noma and Huh7 hepatocellular carcinoma cell lines [34]. In addition,it has been demonstrated that the overexpression of miR-181a leadsto the attenuation of starvation- and rapamycin-induced autophagyin MCF-7, Huh-7 and K562 cells, by targeting ATG5 which is involvedin the formation of autophagosomes [35].
3. Effect of autophagy on modulating immune cells function incancer
Autophagy is implicated in the regulation of multiple immu-nological functions and influences inflammation, infection and
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
immunity. By controlling antigen presentation and modulatinglymphocyte homeostasis, autophagy represents a central pathwayof immune regulation [36]. In this section, we will brieflysummarize the functional role of autophagy in cells involved inthe innate and adaptive immunity.
3.1. Innate immunity
3.1.1. Mast cells
Mast cells (MCs) are immune cells residing in all vascularizedtissues [37] as well as solid tumors in which they play a role intumor angiogenesis [38,39]. MCs, that function as sentinel cells,recruit innate and adaptive immune effector cells and playmultifaceted roles in regulating inflammatory processes, tissueremodeling and host defense [40]. Increased number of MCscorrelates with poor prognosis in numerous tumors [41]. It hasbeen shown that conversion of LC3-I to LC3-II is constitutivelyinduced in MCs, and LC3-II localizes at their secretory granules.Moreover, LC3-II co-localizes with the cluster of differentiation(CD) 63, a secretory lysosomal marker, and its upregulation on cellsurface is impaired in ATG7-deficient MCs. LC3-II is released duringthe degranulation process in ATG7-competent MCs while it isimpaired in ATG7-deficient MCs. These results highlight thatautophagy is constitutively induced in MCs and plays a crucial rolein the degranulation process [42].
3.1.2. Macrophages
Macrophages are cells of the innate immunity implicated indefense against pathogens and wound healing. Tumor-associatedmacrophages (TAMs) are the major component of tumor-infiltrating leukocytes [43] which promote disease progressionby supporting cancer cell survival, proliferation and invasion [44].It has been widely described that autophagy plays a crucial role inmacrophage homeostasis by different mechanisms involving theregulation of hematopoietic stem cell maintenance, the recruit-ment of monocytes and their differentiation into macrophages[45]. It has been reported that toll-like receptors (TLR) signalingmodulates the phagocytic function of macrophages by autophagyand TLR ligands engagement triggers autophagy [46]. Indeed, TLRdeficiency decreases the number of liver-infiltrating macrophagesand correlates with suppressed autophagy flux and p62 aggregatesin hepatocellular carcinoma model [47]. Furthermore, mTORpathway, which inhibits autophagy, is a critical element in theregulation of monocyte differentiation to TAM. It has been shownthat the knockdown of the TSC2 is critical for macrophagepolarization toward the pro-tumoral M2 phenotype [48].
3.1.3. Neutrophils
Neutrophils are the most abundant granulocytes in blood whichexpressed Fc receptors for both immunoglobulin (Ig) G and IgA anddisplayed potent cytotoxic ability [49]. Neutrophils are newpromising effectors of antibody (Ab)-mediated immunotherapybased on the fact that they execute potent cytotoxic functions inthe presence of tumor antigen-specific monoclonal antibodies(mAbs) [50]. In the Ab-dependent cellular cytotoxicity (ADCC)mediated by neutrophils, cancer cells killing is enhanced in thepresence of tumor-specific Fc alpha RI bispecific or IgA antibodies.It has been shown that neutrophils-mediated ADCC of cancer cellsis accompanied by characteristics of autophagy, such as formationof autophagosomes. Furthermore, inhibition of autophagy abol-ished cell death of cancer cells, indicating that autophagy can be amechanism of cell death in ADCC [51].
3.1.4. Dendritic cells
Dendritic cells (DCs) are cells of the innate immune system ableto detect invading pathogens and cancer cells through the
e metabolic response to stress shaping the antitumor immunity.006

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx4
G Model
BCP-12021; No. of Pages 12
expression of sensors on the plasma membrane such as TLR, or inthe cytosol such as nucleotide-binding oligomerization domain(NOD)-like receptors. DCs are highly efficient in antigen presenta-tion to the adaptive immune system thereby initiating long lasting,antigen-specific responses [52]. It is well documented thatDC-mediated cross-presentation of antigens obtained fromphagocytosis of infected or cancerous cells is an important stepin the T lymphocytes activation process [53]. DCs display increasedautophagic activity compared to other cell types due to intensiveprocessing of extra- and intracellular antigens for the majorhistocompatibility complex (MHC) class-I and -II presentation[54]. Autophagy is an important mechanism influencing multiplefunctions of DCs such as (i) direct elimination of pathogen assistedby sequestosome 1-like receptors (SLRs), (ii) regulation of effectorfunctions of pattern recognition receptors (PRRs), and (iii)activation of inflammasome and secretion of alarmins. Moreover,autophagy plays a major role in facilitating the recognition ofdanger signals, owing to its implication in cytoplasmic antigenprocessing for MHC-II presentation, and thereby orchestratesadaptive immune response mediated by T cells [55]. Autophagy isalso involved in the presentation of MHC class-II restrictedpeptides at the cell surface because it allows peptide access tothe MHC class-II loading compartment [56]. In addition, autophagyparticipates in the cross-presentation of MHC class-I restrictedantigens. Captured and degraded by autophagosomes, antigens aresubsequently reprocessed in proteasome and transferred to theendosomal pathway, leading to their loading into recycled MHCclass-I molecules [53]. Overall, autophagy is involved in the uptake,digestion and loading of peptides to MHC class-I or -II moleculesand in their presentation to T cells.
3.2. Adaptive immunity
3.2.1. B lymphocytes
B lymphocytes are part of the adaptive immune system able toproduce antibodies against a broad range of antigens. Due to theirantigen-presenting capacity, B cells also mediate and regulatenumerous essential functions for immune homeostasis including Tcell immune responses [57]. Increase in number of B cells has beendescribed in various human cancers [58] and their presence,together with CD8+ T cells, has been correlated with enhancedsurvival in patients with ovarian [59] and non-small cell lungcancer (NSCLC) [60]. Autophagy plays an important role in theinteraction of B cells with CD4+ T cells by promoting thepresentation of antigens by MHC class-II molecules through amechanism similar to that in DCs [56] [61]. It has been reportedthat MHC class-II loading compartments can fuse with autophago-somes [62] and therefore promote the interaction of CD4+ T cellreceptors (TCRs) with MHC class-II molecules of B cells. Antigenstimulation induces signaling through the B cell antigen receptor(BCR) and triggers apoptosis [63]. Indeed, activation of antigen-stimulated B cells requires costimulatory signals mediated byCD40 ligand and TLR ligands, for example, to rescue B cells fromapoptosis and induce their activation and proliferation. It has beenshown that autophagy is implicated in this rescue-machinery, asBCR signaling rapidly induces extensive autophagosome formationin B cells [64]. Another important implication of autophagy in B cellbiology includes costimulation pathway involving TLR9 signaling[65]. Upon antigen binding, BCR ligation leads to the translocationof both BCR and TLR9 to autophagosomes, which allows theinteraction of TLR9 with the internalized antigen, a process thatcan be blocked with autophagy inhibitors [65].
3.2.2. T lymphocytes
Cytotoxic T lymphocytes (T cells) provide potent defenceagainst virus infection, intracellular pathogens [66] and are also
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
implicated in immunosurveillance of tumors [67]. Autophagicpathway is essential for the development, maintenance, andsurvival of T cells [68–70]. Using nude mice transplanted withATG5�/� thymi, it has been reported that autophagy participatesin CD4+ T cell selection and generation of a self-tolerant T cellrepetoire [71]. Autophagy regulates different aspects of T cellfunction. Indeed, the number of peripheral naive CD4+ and CD8+cells deficient in ATG3, ATG5, or ATG7 was reduced and theirproliferation was impaired upon stimulation [68,72–74]. Thisphenomenon was related to the fact that mitochondrial contentis reduced upon the transition from thymocyte to peripheral Tcell, indicating that loss of autophagy impairs mitochondrialclearance leading to an increase in ROS production [68,72,74].Furthermore, impaired autophagy in T cells can lead to ER stressand alteration of calcium homeostasis [68]. Moreover, inautophagy-deficient T cells, eukaryotic translation initiationfactor 2a (eIF2a) kinases, including PERK, were unable tostimulate autophagy in response to ER stress signaling or alteredcalcium homeostasis. The role of autophagy in maintaining ERand mitochondrial homeostasis and in controlling ROS produc-tion in T cells could explain the importance of autophagyproteins in T cell survival [75,76].
4. Effect of autophagy on tumor cell response to immunesystem
4.1. Autophagy induction modulates cancer cell secretome and
surface proteome
Following stressful conditions, cancer cells privilege thedevelopment of autophagy to adapt stress and maintain a certainhomeostasis. Cells that fail such adaptation will die and pre-mortem autophagy will be used to generate or transport mediatorsnecessary for the induction of immune response to clear damagedcells [77]. Alternatively, these signaling molecules can also induceprotective autophagy to resist and evade immune clearance.Subsequently, autophagy plays an important immunomodulatoryrole by regulating the cancer cell secretome and surface proteomeafter cell stress or death induction. It is now well established thatstressed or dying cells influence their neighboring cells bysignaling the environmental danger through presenting andsecreting soluble factors [78]. The role of autophagy in thisprocess is multiple as autophagy is required for cell-to-cellcommunication and for the release of mediators which in turninduce autophagy in adjacent cancer cells. However, depending onthe cell death inducer or stress condition, tumor cell autophagy hasbeen shown to either induce tumor cell elimination through theincrease of cancer cell immunogenicity or help tumor cells to evadeimmune system under these harmful conditions. Stressed cellsrelease damage associated molecular patterns (DAMPs), cytokinesand extracellular vesicles that mediate danger signals modulatingthe immune response [79]. These factors will be described in moredetails in the following section.
4.1.1. Damage associated molecular patterns (DAMPs)
DAMPs are pro-inflammatory molecules released by dying cellsfollowing chemotherapeutic treatment or metabolic stress (e.g.
starvation, hypoxia). DAMPs elicit antitumor immunity after theimmunogenic cell death (ICD) of targeted cancer cells [77]. DAMPsexist under multiple forms and constitute both short-rangeantitumor immunity mediators (adenosine 50-triphosphate andcalreticulin) and systemic pro-inflammatory molecules (highmobility group box 1) [78]. Autophagy is involved in DAMPssignaling through the induction or the release of DAMPs in theextracellular space. It is worthy to note that autophagy can also beinduced by DAMPs [80]. Below, we will briefly describe DAMPs
e metabolic response to stress shaping the antitumor immunity.006

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 5
G Model
BCP-12021; No. of Pages 12
mediators which exhibit short-range antitumor immunity effect,and those displaying systemic pro-inflammatory function.
4.1.1.1. Adenosine 50-triphosphate (ATP). Since autophagy canpromote the survival of tumor cells through nutrients recoveryfrom degrading and recycling damaged organelles, it has beenrecently proposed that chemotherapy-induced autophagy causesthe release of adenosine 50-triphosphate (ATP) from tumor cells,thereby stimulating antitumor immune response (Fig. 1). ATPfunctions as a ‘‘find me’’ signal for phagocytes when it accumulatesin the extracellular space [81]. ATP is a low-molecular-massimmune mediator secreted after an unconventional fusion ofautophagosomes and vesicle-soluble N-ethylmaleimide-sensitivefactor attachment protein receptor (V-SNARE)-positive vacuoles.Targeting autophagy blunted the release of ATP by tumor cells inresponse to chemotherapy. Autophagy-dependent extracellularATP can bind to ionotropic P2X purinoceptors 7 (P2RX7) andstimulate the differentiation of myeloid precursors into IL-1b-producing DCs [82], recruits them into tumors and activates a T cellresponse toward tumor cells. Extracellular ATP also contributes totumorigenesis through P2 receptors signaling to induce adhesion,proliferation and inflammation [83].
Fig. 1. Autophagy activation in cancer cell stimulates anticancer immune response. P
enhance the release of ‘‘find me’’ signal, such as adenosine 50-triphosphate (ATP), which
activation. Cancer cell autophagy favors the release of ‘‘eat me’’ signal such as calreticulin
also release autophagosomes containing tumor antigens that can be taken up and proce
While the role of autophagy in the regulation of exosomes secretion is still controvers
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
4.1.1.2. Calreticulin (CRT). The best characterized cell surfaceDAMP characterizing immunogenic dying cells consists in pre-apoptotically exposed calreticulin (or ecto-CRT). The exposure ofCRT, which requires autophagy, generates an ‘‘eat-me’’ signaltriggering engulfment by macrophages. Indeed, CRT surfaceexposure is abrogated in cells lacking lysosomal-associatedmembrane protein 2 (LAMP2), a gene essential for chaperone-mediated autophagy (Fig. 1) [84]. However, in the specific case ofROS-induced ICD, autophagy was shown to suppress pre-apoptoticCRT exposure on cancer cells and to reduce DC-based IL-6production and T cell stimulation, thus helping in evading fromICD (Fig. 2) [85].
4.1.1.3. High mobility group box 1 (HMGB1). Autophagy canmodulate inflammation during metastasis by different ways.Indeed, the pro-inflammatory role of autophagy also extends to theunconventional secretion of the HMGB1 protein in a Golgireassembly stacking protein- (GRASP) and autophagy-dependentnon-classical membrane trafficking process (Fig. 2) [86]. HMGB1 isa DNA-binding protein mainly located in the nucleus and acting asa DNA chaperone necessary for nuclear homeostasis [87]. HMGB1exhibits opposite roles in promoting both cell survival and cell
re-mortem autophagic response and/or stress-induced autophagy in cancer cells
attracts myeloids precursors, induces their differentiation and finally leads to T cell
, which stimulates the phagocytic functions of macrophages. Dying cancer cells can
ss by antigen-presenting cells, and finally resulting in activation of T cell response.
ial, their potent immunostimulatory functions are now well established.
e metabolic response to stress shaping the antitumor immunity.006
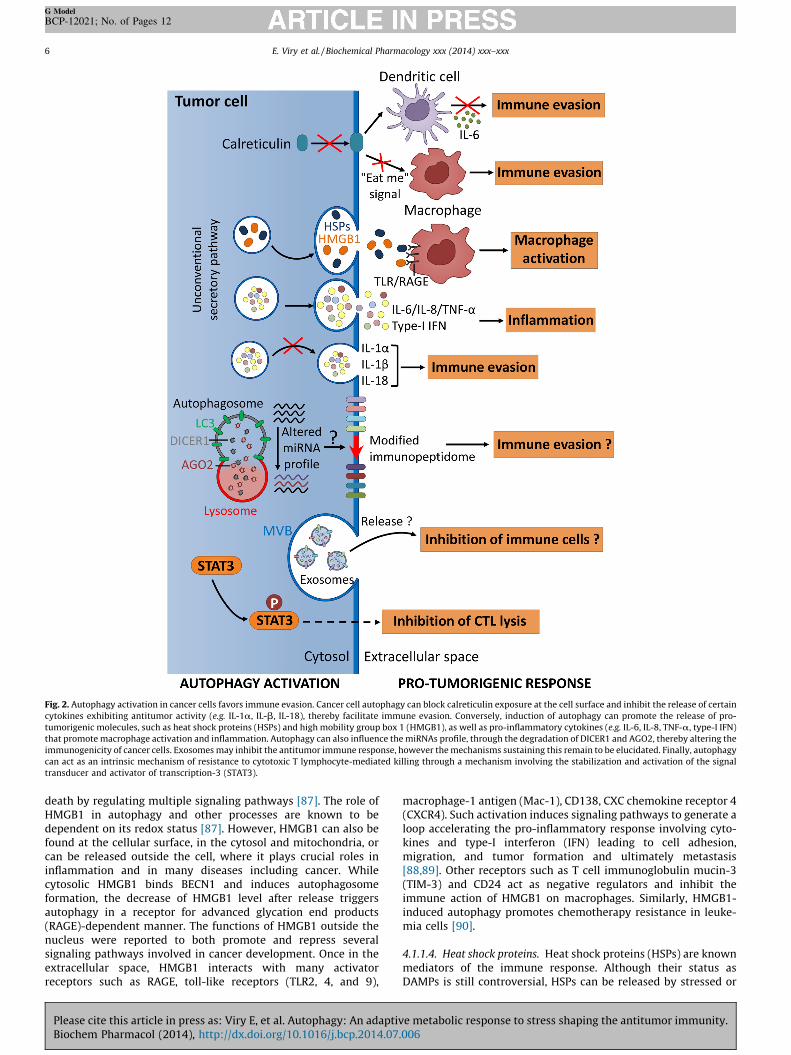
Fig. 2. Autophagy activation in cancer cells favors immune evasion. Cancer cell autophagy can block calreticulin exposure at the cell surface and inhibit the release of certain
cytokines exhibiting antitumor activity (e.g. IL-1a, IL-b, IL-18), thereby facilitate immune evasion. Conversely, induction of autophagy can promote the release of pro-
tumorigenic molecules, such as heat shock proteins (HSPs) and high mobility group box 1 (HMGB1), as well as pro-inflammatory cytokines (e.g. IL-6, IL-8, TNF-a, type-I IFN)
that promote macrophage activation and inflammation. Autophagy can also influence the miRNAs profile, through the degradation of DICER1 and AGO2, thereby altering the
immunogenicity of cancer cells. Exosomes may inhibit the antitumor immune response, however the mechanisms sustaining this remain to be elucidated. Finally, autophagy
can act as an intrinsic mechanism of resistance to cytotoxic T lymphocyte-mediated killing through a mechanism involving the stabilization and activation of the signal
transducer and activator of transcription-3 (STAT3).
E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx6
G Model
BCP-12021; No. of Pages 12
death by regulating multiple signaling pathways [87]. The role ofHMGB1 in autophagy and other processes are known to bedependent on its redox status [87]. However, HMGB1 can also befound at the cellular surface, in the cytosol and mitochondria, orcan be released outside the cell, where it plays crucial roles ininflammation and in many diseases including cancer. Whilecytosolic HMGB1 binds BECN1 and induces autophagosomeformation, the decrease of HMGB1 level after release triggersautophagy in a receptor for advanced glycation end products(RAGE)-dependent manner. The functions of HMGB1 outside thenucleus were reported to both promote and repress severalsignaling pathways involved in cancer development. Once in theextracellular space, HMGB1 interacts with many activatorreceptors such as RAGE, toll-like receptors (TLR2, 4, and 9),
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
macrophage-1 antigen (Mac-1), CD138, CXC chemokine receptor 4(CXCR4). Such activation induces signaling pathways to generate aloop accelerating the pro-inflammatory response involving cyto-kines and type-I interferon (IFN) leading to cell adhesion,migration, and tumor formation and ultimately metastasis[88,89]. Other receptors such as T cell immunoglobulin mucin-3(TIM-3) and CD24 act as negative regulators and inhibit theimmune action of HMGB1 on macrophages. Similarly, HMGB1-induced autophagy promotes chemotherapy resistance in leuke-mia cells [90].
4.1.1.4. Heat shock proteins. Heat shock proteins (HSPs) are knownmediators of the immune response. Although their status asDAMPs is still controversial, HSPs can be released by stressed or
e metabolic response to stress shaping the antitumor immunity.006

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 7
G Model
BCP-12021; No. of Pages 12
injured cells. HSPs stimulate cytokine release leading to immunecells activation and generate pro-inflammatory response favoringresistance to chemotherapy, cancer proliferation, metastasis andfinally a poor prognosis (Fig. 2). Moreover, acetylated HSP70 isrequired for autophagosome formation [91] and can probably beexported by autophagosomes in the extracellular space.
4.1.2. Antigen presentation
Endogenous peptides presented by MHC class-I moleculesconstitute the immunopeptidome. Through its cytoprotectivecharacter, autophagy strongly impacts the composition of thispool of antigens exposed at the cell surface mainly by its influenceon translation [92] leading to the appearance of multiple antigenson cell surface [93]. The immunopeptidome is also influenced bythe relative abundance of certain miRNAs [94]. In particular, miR-155 knockdown was shown to suppress antigen presentation andto decrease MHC-class II expression [95]. In addition, by using thenuclear dot protein 52 kDa (NDP52) as a particular cargo,autophagy targets DICER1 and AGO2, two crucial regulators ofmiRNAs processing and function, for degradation [96,97]. Thisautophagy-dependent degradation could modify the cellularmiRNAs profile and therefore potentially impact the antigenprocessing and presentation on cell surface, and as a resultrecognition by immune cells (Fig. 2).
Moreover, autophagosomes also play a role in cancer cellclearance by efficiently carrying antigens for cross-presentation toT cells [98]. Autophagy facilitates the development of inflamma-tory responses, as the release of autophagosomes by the antigendonor cell triggers the production of type I IFNs in phagocytic DCs[99]. In this regard, it has been reported that autophagy inductionin vitamin E analog alpha-tocopheryloxyacetic acid (a-TEA)-treated tumor cells leads to the release of autophagosomescontaining antigens, which efficiently cross-primed CD8+ T cells[100] (Fig. 2).
4.1.3. Cytokines
It is now clearly established that the activation of pro-tumoralpathways and cancer progression are closely related to theexpression of many cytokines. Indeed, the activation of the signaltransducer and activator of transcription-3 (STAT3) pathway hasbeen shown in cancer cells to be strongly dependent on severalcytokines and growth factors such as IL-6, IL-11, IL-22, hepatocytegrowth factor (HGF), and epidermal growth factor (EGF), and ononcogenic tyrosine kinases (c-Met and Src) [101]. The activation ofthe activator protein (AP)-1 pathway is also largely dependent onTNF signaling in skin cancer cells [102]. In response to DAMPs, therelease of cytokines necessary for tumor development is now welllinked to autophagy. Although autophagy has been shown topositively regulate the transcription and secretion of TNF-a, IL-6,IL-8, and possibly the type-I IFN, it can in contrast negativelyregulate the release of IL-1a, IL-1b, and IL-18 in certain conditions(Fig. 2). Indeed, recent data confirmed that autophagy supports IL-6 secretion necessary for oncogenic RAS-driven invasion in 3Dculture [103].
It has been reported that other cytokines can be released byunconventional secretion through autophagy. Indeed, severalcytokines (IL-1b and IL-18) lack signal peptides which facilitatetheir entry to the endoplasmic reticulum and their subsequentrelease through the conventional secretory pathway. In thisregard, autophagy can have opposite roles in different cellularmodels. Indeed, autophagy inhibits the secretion of IL-1b andIL-18 by macrophages as the depletion of the autophagic proteinsLC3-II and BECN1 was shown to enhance the activation ofcaspase-1 and secretion of these cytokines [104]. In contrast, therelease of IL-1b was found to be performed through autophagy-based unconventional secretory pathway by a mechanism
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
involving Atg5-GRASP-Rab8. This result demonstrates thatautophagy can be required for IL-1b secretion [105]. The exactmechanisms by which autophagy contributes to the release ofthese pro-inflammatory factors are currently not fully under-stood. The function of autophagy as a switch, allowing thesecretion of cytokines in certain conditions (starvation i.e. stress,or infection) and the repression of the inflammasome impairingactive cytokine secretion under basal conditions, is now wellestablished [106].
Autophagy and TLR play important roles in cancer developmentand immune escape. The induction of autophagy by TLR3 and TLR4was shown to be an important mechanism driving lung cancerprogression through the increased production of cytokines andfactors necessary for enhanced migration and invasion of lungcancer cells including IL-6, CC chemokine ligand (CCL)-2/monocytechemoattractant protein (MCP)-1, CCL20/macrophage inflamma-tory protein (MIP)-3a, VEGFA, and matrix metalloproteinase(MMP)-2 [107].
4.1.4. Extracellular vesicles
Extracellular vesicles are produced and released by all types ofcells and exhibit potent immunomodulatory functions. Consider-able investigations have been mostly focused on a subtype of 30–100 nm vesicles called exosomes that contain small RNAs,cytokines, HSPs, and many other mediators which can stimulateor repress immune reactions. How exosomes contribute to cancerdevelopment, metastasis and their impact on the immune systemhas been extensively reviewed by others [108,109]. Hypoxic cancercells release exosomes into their microenvironment to promotetheir own survival and invasion [110]. However, the intrinsicconnection between autophagy and exosome secretion is complexand not fully understood. In this regard, the role of autophagy inthe regulation of exosomes secretion is still controversial (Figs. 1and 2). Indeed, autophagy induction was reported to completelyblock exosome secretion, suggesting that multivesicular bodies(MVBs) are diverted to the autophagic pathway [111] leading to animpairment of exosome release in cells displaying high autophagicactivity. In contrast, a concomitant autophagy and exosomesecretion have been recently observed in the retinal epithelium[112], indicating that models displaying different metabolic andcatabolic capabilities might not respond to autophagy-inducingstresses in similar ways.
4.2. Autophagy induction in cancer cell provides an intrinsic
mechanism of resistance to immune response
Although mounting evidence suggests that induction ofautophagy in tumor cell modulates antitumor immune response[78], relatively little attention has been given to its intrinsic role ininducing resistance to immune effectors. Given the role ofautophagy as a common cellular response to stress, it stands toreason that autophagy may protect tumor cell from immune cell-mediated killing by operating as an intrinsic tumor cell-resistancemechanism against both innate and adaptive immunity (Fig. 2).
Noman et al. provided the first demonstration that hypoxictumor cells evade T cell-mediated lysis through several mecha-nisms allowing adaptation to hypoxic stress. Indeed, activation ofHIF-1a enhances the phosphorylation of STAT3 [113], inducesautophagy [114], upregulates the expression of the embryonicmarker NANOG [115], and increases the expression of miR-210that targets the non-receptor type protein tyrosine phosphatase(PTPN)-1, homeobox (HOX) A1, and tumor protein-p53 inducibleprotein 11 (TP53I11) [116]. It should be emphasized that thecrucial role of autophagy was highlighted by showing that itstargeting in hypoxic lung carcinoma cells compromises HIF-1a-dependent activation and stabilization of STAT3 and restores
e metabolic response to stress shaping the antitumor immunity.006
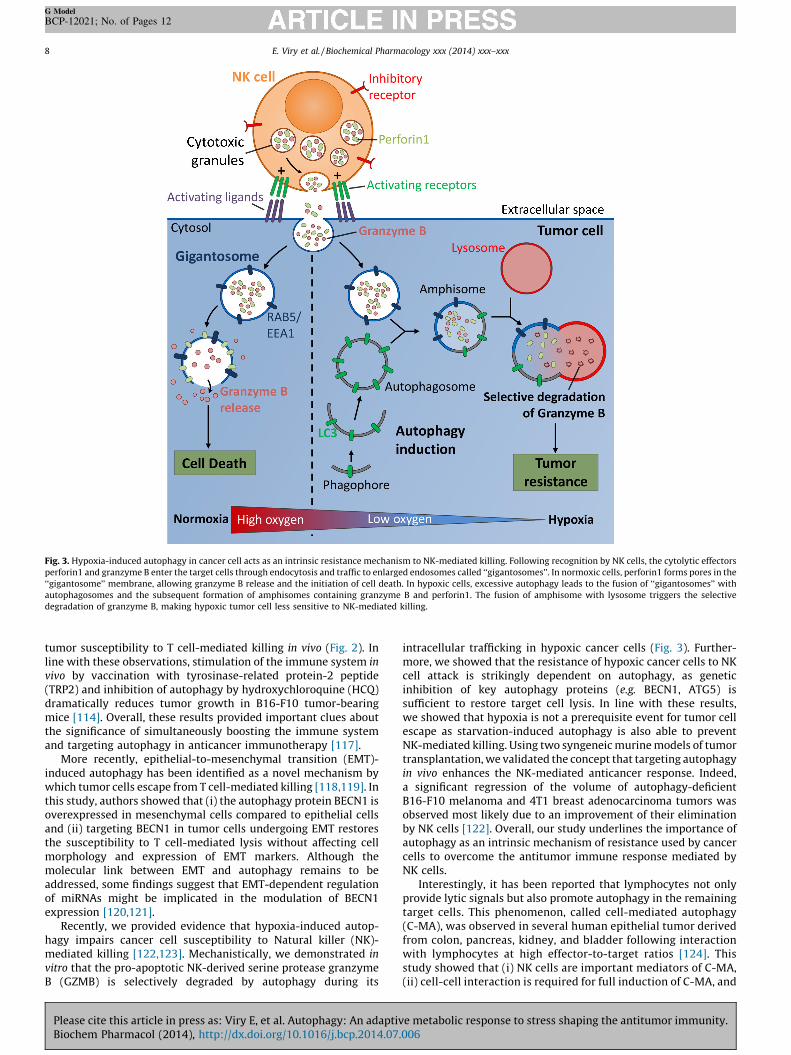
Fig. 3. Hypoxia-induced autophagy in cancer cell acts as an intrinsic resistance mechanism to NK-mediated killing. Following recognition by NK cells, the cytolytic effectors
perforin1 and granzyme B enter the target cells through endocytosis and traffic to enlarged endosomes called ‘‘gigantosomes’’. In normoxic cells, perforin1 forms pores in the
‘‘gigantosome’’ membrane, allowing granzyme B release and the initiation of cell death. In hypoxic cells, excessive autophagy leads to the fusion of ‘‘gigantosomes’’ with
autophagosomes and the subsequent formation of amphisomes containing granzyme B and perforin1. The fusion of amphisome with lysosome triggers the selective
degradation of granzyme B, making hypoxic tumor cell less sensitive to NK-mediated killing.
E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx8
G Model
BCP-12021; No. of Pages 12
tumor susceptibility to T cell-mediated killing in vivo (Fig. 2). Inline with these observations, stimulation of the immune system in
vivo by vaccination with tyrosinase-related protein-2 peptide(TRP2) and inhibition of autophagy by hydroxychloroquine (HCQ)dramatically reduces tumor growth in B16-F10 tumor-bearingmice [114]. Overall, these results provided important clues aboutthe significance of simultaneously boosting the immune systemand targeting autophagy in anticancer immunotherapy [117].
More recently, epithelial-to-mesenchymal transition (EMT)-induced autophagy has been identified as a novel mechanism bywhich tumor cells escape from T cell-mediated killing [118,119]. Inthis study, authors showed that (i) the autophagy protein BECN1 isoverexpressed in mesenchymal cells compared to epithelial cellsand (ii) targeting BECN1 in tumor cells undergoing EMT restoresthe susceptibility to T cell-mediated lysis without affecting cellmorphology and expression of EMT markers. Although themolecular link between EMT and autophagy remains to beaddressed, some findings suggest that EMT-dependent regulationof miRNAs might be implicated in the modulation of BECN1expression [120,121].
Recently, we provided evidence that hypoxia-induced autop-hagy impairs cancer cell susceptibility to Natural killer (NK)-mediated killing [122,123]. Mechanistically, we demonstrated in
vitro that the pro-apoptotic NK-derived serine protease granzymeB (GZMB) is selectively degraded by autophagy during its
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
intracellular trafficking in hypoxic cancer cells (Fig. 3). Further-more, we showed that the resistance of hypoxic cancer cells to NKcell attack is strikingly dependent on autophagy, as geneticinhibition of key autophagy proteins (e.g. BECN1, ATG5) issufficient to restore target cell lysis. In line with these results,we showed that hypoxia is not a prerequisite event for tumor cellescape as starvation-induced autophagy is also able to preventNK-mediated killing. Using two syngeneic murine models of tumortransplantation, we validated the concept that targeting autophagyin vivo enhances the NK-mediated anticancer response. Indeed,a significant regression of the volume of autophagy-deficientB16-F10 melanoma and 4T1 breast adenocarcinoma tumors wasobserved most likely due to an improvement of their eliminationby NK cells [122]. Overall, our study underlines the importance ofautophagy as an intrinsic mechanism of resistance used by cancercells to overcome the antitumor immune response mediated byNK cells.
Interestingly, it has been reported that lymphocytes not onlyprovide lytic signals but also promote autophagy in the remainingtarget cells. This phenomenon, called cell-mediated autophagy(C-MA), was observed in several human epithelial tumor derivedfrom colon, pancreas, kidney, and bladder following interactionwith lymphocytes at high effector-to-target ratios [124]. Thisstudy showed that (i) NK cells are important mediators of C-MA,(ii) cell-cell interaction is required for full induction of C-MA, and
e metabolic response to stress shaping the antitumor immunity.006

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 9
G Model
BCP-12021; No. of Pages 12
(iii) C-MA limits the response of tumor cells to subsequent stressorsuch as g-radiation. These findings highlighted autophagy inductionin target cells not only as a mechanism of resistance to immuneeffectors but also as an important target to be considered for theimprovement of the efficacy of anticancer therapies.
5. Autophagy modulators as therapeutic strategy for improvinganticancer immunotherapy
Accumulating evidence suggests that one important conse-quence of autophagy activation in cancer cells under stresscondition is the ability to resist the impact of chemotherapy[125,126], radiotherapy [127,128] and immunotherapy[114,116,122,123]. Indeed, either through supporting metabolismdirectly, or by limiting cell death pathway, autophagy may have apro-survival role in aggressive cancer, leading to anticancertherapy resistance. In line with this, and given the critical roleof autophagy in tumorigenesis as well as in tumor and cancer stemcell maintenance [129,130], various preclinical and clinical studieshave been undertaken to develop therapeutic agents to inhibitautophagy. Pharmacological inhibitors of autophagy identified sofar can be classified as early- or late-stage inhibitors of theautophagic pathway. 3-Methyladenine (3-MA), wortmannin, andLY294002 target the class III PI3K (Vps34) and are classified as anearly-stage inhibitor, whereas chloroquine (CQ), HCQ, bafilomycinA1, and monensin interfere with lysosomal function and aretherefore classified as late-stage inhibitors of autophagy. Knowingthat autophagosomes and lysosomes move along microtubules,microtubule-disrupting agents such as taxanes, nocodazole,colchicine, and vinca alkaloids has been identified as anotherclass of autophagy inhibitors. CQ, HCQ and quinacrine have beenevaluated in humans as promising anti-autophagy agents.Currently, more than 50 clinical trials registered in the NationalCancer Institute to evaluate effect of HCQ, CQ or quinacrine as anautophagy inhibitor in a variety of human cancers (http://clinicaltrials.gov). Because only a subpopulation of tumor cellsundergoes autophagy, it is unlikely that autophagy inhibitors areused in cancer therapy as single agents. Indeed, most of the clinicaltrials described above used HCQ in combination with otheranticancer therapies. While the majority of clinical trials arecurrently evaluating the combination of autophagy inhibitors withchemotherapy, new data support that autophagy inhibition canalso improve the antitumor immune response. Such statement hasbeen proposed based on the fact that increased autophagy withinthe cancer cell suppressed the antitumor immune response andthat several cancer immunotherapeutic strategies based onadoptive transfer of T cells, DC vaccines, administration ofantibodies or recombinant cytokines such as IL-2, could only beeffective if the inhibition of the autophagic process is achieved[131]. In addition, based on our recent findings, autophagyinhibition may increase the cytotoxicity of effector T cells andNK cells once they have been activated to lyse the tumor. Thecombination of high dose of IL-2 with CQ increased long termsurvival, decreased toxicity associated with vascular leakage, andenhanced immune cell proliferation and infiltration in the liver andspleen [132].
While experimental and preclinical data are generallysupportive of incorporating anti-autophagy therapies in cancertreatment trials, it has been observed in some circumstances thatinhibition of autophagy decreases therapeutic efficacy. It is nowclearly established that autophagy plays also a crucial role inincreasing the immunogenicity of the tumor cell, and activelyparticipates in the antigen processing and the subsequentactivation of the effectors T cells. Thus, induction of autophagymay be exploited as adjuvant strategy to stimulate theantitumor immune response. For example, it has been shown
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
that tumor-derived autophagosomes vaccine, or autophago-somes loaded onto DCs, in combination with agents that reversethe immune suppression (e.g. IFN-g, TLR agonist) enhanced theantitumor response against 3LL Lewis lung carcinoma andB16-F10 melanoma [133]. A phase I/II is currently ongoing toinvestigate the efficacy of such vaccine in patients with NSCLCcancer.
Given the dual role of autophagy in modulating the anticancerimmune response, development of innovative therapeuticapproaches may consider, for example, the use of ex vivo inductionof autophagy within professional antigen-presenting cells (APC)concomitantly with in vivo inhibition of tumor-induced autophagyin order to both activate the immune system and overcome tumorresistance to lysis. Thus, understanding to which extent and underwhich circumstances inducers and/or inhibitors of autophagyaffect the therapeutic efficacy of anticancer treatments will be ofgreat importance to improve the rational use of such modulators.
6. Conclusion
Abnormal tumor microenvironment induces a collection ofcellular stress responses that further contribute to the distortedmetabolic phenotype of tumor cells and influence tumor progres-sion, anti-tumor immunity and response to therapies. However,the relationship between the tumor microenvironment and cancercell metabolism should not be considered as a simple cause andeffect.
The stresses present in the tumor microenvironment includinghypoxia, low nutrient availability, immune infiltrates are nowrecognized as physiological selective pressures that promotetumor aggressiveness. Undoubtedly, significant advances havebeen made in our understanding of the molecular pathwaysactivated in response to microenvironmental stresses. There arenumerous studies supporting that autophagy activation in theabnormal tumor microenvironment is an important stressresponse mechanism that promotes tumor dormancy and therapyresistance. Autophagy represents an important modulator of anti-tumor immune response by acting both in immune effectors andtarget cancer cells. In accordance with this, modulation ofautophagy may represent an important therapeutic opportunityto enhance efficacy several immune-based anticancer therapies. Itshould be emphasized that a better understanding of themechanism by which autophagy modulates cell metabolism inresponse to environmental stresses and how this impacts theresponse to anticancer therapies will help developing moreeffective therapeutic strategies.
Acknowledgments
This work was supported by grants from CRP-Sante (REC-LHCE-2013-1105), Kriibskrank Kanner Foundation (FON-LHCE-2013-1005), Caloust Gulbenkian Foundation, Televie (7.4628.12F,7.4606.13 and 7.4504.11F) and ‘‘Fonds National de la Recherche’’(FNR) Luxembourg (C12/BM/3962058 and AFR 7842786).
References
[1] Lunt SJ, Chaudary N, Hill RP. The tumor microenvironment and metastaticdisease. Clin Exp Metastasis 2009;26:19–34.
[2] Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nat RevCancer 2011;11:85–95.
[3] Denko NC, Hypoxia. HIF1 and glucose metabolism in the solid tumour. NatRev Cancer 2008;(8):705–10.
[4] Wouters BG, Koritzinsky M. Hypoxia signalling through mTOR and theunfolded protein response in cancer. Nat Rev Cancer 2008;8:851–60.
[5] Koritzinsky M, Magagnin MG, van den Beucken T, Seigneuric R, Savelkouls K,Dostie J, et al. Gene expression during acute and prolonged hypoxia isregulated by distinct mechanisms of translational control. EMBO J 2006;25:1114–20.
e metabolic response to stress shaping the antitumor immunity.006

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx10
G Model
BCP-12021; No. of Pages 12
[6] Janji B, Noman MZ, Viry E, Hasmim M, Messai Y, Berchem G, et al. Emergingrole of hypoxia-induced autophagy in cancer immunotherapy. In: Hayat MA,editor. Autophagy: cancer, other pathologies, inflammation, immunity, in-fection, and aging. Elsevier/Academic Press; 2014. p. 247–50.
[7] Janji B, Viry E, Baginska J, Van Moer K, Berchem G. Role of autophagy in cancerand tumor progression. In: Bailly Y, editor. Autophagy: a double-edged sword– cell survival or death?. Intech; 2013. p. 189–90.
[8] Corcelle EA, Puustinen P, Jaattela M. Apoptosis autophagy: targetingautophagy signalling in cancer cells – ‘trick or treats’? FEBS J2009;276:6084–90.
[9] Glick D, Barth S, Macleod KF. Autophagy: cellular and molecular mechanisms.J Pathol 2010;221:3–12.
[10] Mizushima N. Autophagy: process and function. Genes Dev 2007;21:2861–2870.
[11] Yang Z, Klionsky DJ. Eaten alive: a history of macroautophagy. Nat Cell Biol2010;12:814–20.
[12] Mathew R, White E. Autophagy, stress, and cancer metabolism: what doesn’tkill you makes you stronger. Cold Spring Harb Symp Quant Biol 2011;76:389–390.
[13] Czaja MJ. Autophagy in health and disease. 2. Regulation of lipid metabolismand storage by autophagy: pathophysiological implications. Am J Physiol CellPhysiol. 2010;298:C973–80.
[14] White E. Deconvoluting the context-dependent role for autophagy in cancer.Nat Rev Cancer 2012;12:401–10.
[15] Rosenfeldt MT, Ryan KM. The role of autophagy in tumour development andcancer therapy. Expert Rev Mol Med 2009;11:e36.
[16] Yang Z, Klionsky DJ. An overview of the molecular mechanism of autophagy.Curr Top Microbiol Immunol 2009;335:1–32.
[17] Bellot G, Garcia-Medina R, Gounon P, Chiche J, Roux D, Pouyssegur J, et al.Hypoxia-induced autophagy is mediated through hypoxia-inducible factorinduction of BNIP3 and BNIP3L via their BH3 domains. Mol Cell Biol2009;29:2570–80.
[18] Azad MB, Chen Y, Henson ES, Cizeau J, McMillan-Ward E, Israels SJ, et al.Hypoxia induces autophagic cell death in apoptosis-competent cells througha mechanism involving BNIP3. Autophagy 2008;4:195–200.
[19] Kang R, Zeh HJ, Lotze MT, Tang D. The Beclin 1 network regulates autophagyand apoptosis. Cell Death Differ 2011;18:571–80.
[20] Greijer AE, van der Groep P, Kemming D, Shvarts A, Semenza GL, Meijer GA,et al. Up-regulation of gene expression by hypoxia is mediated predominant-ly by hypoxia-inducible factor 1 (HIF-1). J Pathol 2005;206:291–300.
[21] Harris AL. Hypoxia—a key regulatory factor in tumour growth. Nat Rev Cancer2002;2:38–47.
[22] Kothari S, Cizeau J, McMillan-Ward E, Israels SJ, Bailes M, Ens K, et al. BNIP3plays a role in hypoxic cell death in human epithelial cells that is inhibited bygrowth factors EGF and IGF. Oncogene 2003;22:4734–40.
[23] Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism.Cell 2006;124:471–80.
[24] Kamada Y, Funakoshi T, Shintani T, Nagano K, Ohsumi M, Ohsumi Y. Tor-mediated induction of autophagy via an Apg1 protein kinase complex. J CellBiol 2000;150:1507–10.
[25] Inoki K, Guan KL. Tuberous sclerosis complex, implication from a rare geneticdisease to common cancer treatment. Hum Mol Genet 2009;18:R94–100.
[26] Papandreou I, Lim AL, Laderoute K, Denko NC. Hypoxia signals autophagy intumor cells via AMPK activity, independent of HIF-1, BNIP3, and BNIP3L. CellDeath Differ 2008;15:1572–80.
[27] Kwiatkowski DJ, Manning BD. Tuberous sclerosis: a GAP at the crossroads ofmultiple signaling pathways. Hum Mol Genet 2005;14(Spec no. 2):R251–60.
[28] Ron D, Walter P. Signal integration in the endoplasmic reticulum unfoldedprotein response. Nat Rev Mol Cell Biol 2007;8:519–20.
[29] Rzymski T, Milani M, Pike L, Buffa F, Mellor HR, Winchester L, et al. Regulationof autophagy by ATF4 in response to severe hypoxia. Oncogene2010;29:4424–30.
[30] Pike LR, Singleton DC, Buffa F, Abramczyk O, Phadwal K, Li JL, et al. Transcrip-tional up-regulation of ULK1 by ATF4 contributes to cancer cell survival.Biochem J 2013;449:389–90.
[31] Lum JJ, DeBerardinis RJ, Thompson CB. Autophagy in metazoans: cell survivalin the land of plenty. Nat Rev Mol Cell Biol 2005;6:439–40.
[32] Emerling BM, Weinberg F, Snyder C, Burgess Z, Mutlu GM, Viollet B, et al.Hypoxic activation of AMPK is dependent on mitochondrial ROS butindependent of an increase in AMP/ATP ratio. Free Radic Biol Med2009;46:1386–90.
[33] Li L, Chen Y, Gibson SB. Starvation-induced autophagy is regulated bymitochondrial reactive oxygen species leading to AMPK activation. Cell Signal2013;25:50–65.
[34] Korkmaz G, Tekirdag KA, Ozturk DG, Kosar A, Sezerman OU, Gozuacik D.MIR376A is a regulator of starvation-induced autophagy. PLOS ONE2013;8:e82556.
[35] Tekirdag KA, Korkmaz G, Ozturk DG, Agami R, Gozuacik D. MIR181A regulatesstarvation- and rapamycin-induced autophagy through targeting of ATG5.Autophagy 2013;9:374–80.
[36] Deretic V, Saitoh T, Akira S. Autophagy in infection, inflammation andimmunity. Nat Rev Immunol 2013;13:722–30.
[37] Marichal T, Tsai M, Galli SJ. Mast cells: potential positive and negative roles intumor biology. Cancer Immunol Res 2013;1:269–70.
[38] Ribatti D, Crivellato E, Molica S. Mast cells and angiogenesis in haematolo-gical malignancies. Leuk Res 2009;33:876–80.
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
[39] Ribatti D, Guidolin D, Marzullo A, Nico B, Annese T, Benagiano V, et al. Mastcells and angiogenesis in gastric carcinoma. Int J Exp Pathol 2010;91:350–60.
[40] Oldford SA, Marshall JS. Mast cells as targets for immunotherapy of solidtumors. Mol Immunol 2014.
[41] Ribatti D. Mast cells and macrophages exert beneficial and detrimentaleffects on tumor progression and angiogenesis. Immunol Lett 2013;152:83–88.
[42] Ushio H, Ueno T, Kojima Y, Komatsu M, Tanaka S, Yamamoto A, et al. Crucialrole for autophagy in degranulation of mast cells. J Allergy Clin Immunol2011;127:1267–70. e6.
[43] Mantovani A, Schioppa T, Porta C, Allavena P, Sica A. Role of tumor-associatedmacrophages in tumor progression and invasion. Cancer Metastasis Rev2006;25:315–20.
[44] Allavena P, Mantovani A. Immunology in the clinic review series; focus oncancer: tumour-associated macrophages: undisputed stars of the inflamma-tory tumour microenvironment. Clin Exp Immunol 2012;167:195–200.
[45] Chen P, Cescon M, Bonaldo P. Autophagy-mediated regulation of macro-phages and its applications for cancer. Autophagy 2014;10:192–200.
[46] Xu Y, Jagannath C, Liu XD, Sharafkhaneh A, Kolodziejska KE, Eissa NT. Toll-likereceptor 4 is a sensor for autophagy associated with innate immunity.Immunity 2007;27:135–40.
[47] Lin H, Yan J, Wang Z, Hua F, Yu J, Sun W, et al. Loss of immunity-supportedsenescence enhances susceptibility to hepatocellular carcinogenesis andprogression in Toll-like receptor 2-deficient mice. Hepatology 2013;57:171–80.
[48] Chen W, Ma T, Shen XN, Xia XF, Xu GD, Bai XL, et al. Macrophage-inducedtumor angiogenesis is regulated by the TSC2-mTOR pathway. Cancer Res2012;72:1363–70.
[49] Nathan C. Neutrophils and immunity: challenges and opportunities. Nat RevImmunol 2006;6:173–80.
[50] van Egmond M, Bakema JE. Neutrophils as effector cells for antibody-basedimmunotherapy of cancer. Semin Cancer Biol 2013;23:190–200.
[51] Bakema JE, Ganzevles SH, Fluitsma DM, Schilham MW, Beelen RH, Valerius T,et al. Targeting FcalphaRI on polymorphonuclear cells induces tumor cellkilling through autophagy. J Immunol 2011;187:726–30.
[52] Mellman I. Dendritic cells: master regulators of the immune response. CancerImmunol Res 2013;1:145–50.
[53] Fiegl D, Kagebein D, Liebler-Tenorio EM, Weisser T, Sens M, Gutjahr M, et al.Amphisomal route of MHC class I cross-presentation in bacteria-infecteddendritic cells. J Immunol 2013;190:2791–800.
[54] Ma Y, Galluzzi L, Zitvogel L, Kroemer G. Autophagy and cellular immuneresponses. Immunity 2013;39:211–20.
[55] Deretic V. Autophagy: an emerging immunological paradigm. J Immunol2012;189:15–20.
[56] Dengjel J, Schoor O, Fischer R, Reich M, Kraus M, Muller M, et al. Autophagypromotes MHC class II presentation of peptides from intracellular sourceproteins. Proc Natl Acad Sci U S A 2005;102:7922–30.
[57] Fremd C, Schuetz F, Sohn C, Beckhove P, Domschke C. B cell-regulatedimmune responses in tumor models and cancer patients. Oncoimmunology2013;2:e25443.
[58] Nelson BHCD20. B cells: the other tumor-infiltrating lymphocytes. J Immunol2010;185:4977–80.
[59] Milne K, Kobel M, Kalloger SE, Barnes RO, Gao D, Gilks CB, et al. Systematicanalysis of immune infiltrates in high-grade serous ovarian cancerreveals CD20, FoxP3 and TIA-1 as positive prognostic factors. PLoS ONE2009;4:e6412.
[60] Al-Shibli KI, Donnem T, Al-Saad S, Persson M, Bremnes RM, Busund LT.Prognostic effect of epithelial and stromal lymphocyte infiltration in non-small cell lung cancer. Clin Cancer Res Off J Am Assoc Cancer Res2008;14:5220–30.
[61] Nimmerjahn F, Milosevic S, Behrends U, Jaffee EM, Pardoll DM, BornkammGW, et al. Major histocompatibility complex class II-restricted presentationof a cytosolic antigen by autophagy. Eur J Immunol 2003;33:1250–60.
[62] Schmid D, Pypaert M, Munz C. Antigen-loading compartments for majorhistocompatibility complex class II molecules continuously receive inputfrom autophagosomes. Immunity 2007;26:79–92.
[63] Wang LD, Clark MR. B-cell antigen-receptor signalling in lymphocyte devel-opment. Immunology 2003;110:411–20.
[64] Watanabe K, Tsubata T. Autophagy connects antigen receptor signaling tocostimulatory signaling in B lymphocytes. Autophagy 2009;5:108–10.
[65] Chaturvedi A, Dorward D, Pierce SK. The B cell receptor governs the subcel-lular location of Toll-like receptor 9 leading to hyperresponses to DNA-containing antigens. Immunity 2008;28:799–800.
[66] Barry M, Bleackley RC, Cytotoxic. T lymphocytes: all roads lead to death. NatRev Immunol 2002;2:401–10.
[67] Noman MZ, Messai Y, Carre T, Akalay I, Meron M, Janji B, et al. Microenvi-ronmental hypoxia orchestrating the cell stroma cross talk, tumor progres-sion and antitumor response. Crit Rev Immunol 2011;31:357–60.
[68] Jia W, He YW. Temporal regulation of intracellular organelle homeostasis inT lymphocytes by autophagy. J Immunol 2011;186:5313–20.
[69] Nedjic J, Aichinger M, Emmerich J, Mizushima N, Klein L. Autophagy in thymicepithelium shapes the T-cell repertoire and is essential for tolerance. Nature2008;455:396–400.
[70] Arsov I, Adebayo A, Kucerova-Levisohn M, Haye J, MacNeil M, PapavasiliouFN, et al. A role for autophagic protein beclin 1 early in lymphocyte develop-ment. J Immunol 2011;186:2201–10.
e metabolic response to stress shaping the antitumor immunity.006

E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx 11
G Model
BCP-12021; No. of Pages 12
[71] Klein L, Munz C, Lunemann JD. Autophagy-mediated antigen processing inCD4(+) T cell tolerance and immunity. FEBS Lett 2010;584:1405–10.
[72] Hubbard VM, Valdor R, Patel B, Singh R, Cuervo AM, Macian F. Macroauto-phagy regulates energy metabolism during effector T cell activation. JImmunol 2010;185:7349–50.
[73] Pua HH, Dzhagalov I, Chuck M, Mizushima N, He YW. A critical role for theautophagy gene Atg5 in T cell survival and proliferation. J Exp Med2007;204:25–31.
[74] Pua HH, Guo J, Komatsu M, He YW. Autophagy is essential for mitochondrialclearance in mature T lymphocytes. J Immunol 2009;182:4046–50.
[75] Liang SH, Zhang W, McGrath BC, Zhang P, Cavener DRPERK. (eIF2alphakinase) is required to activate the stress-activated MAPKs and induce theexpression of immediate-early genes upon disruption of ER calcium homo-eostasis. Biochem J 2006;393:201–10.
[76] Lepine S, Allegood JC, Park M, Dent P, Milstien S, Spiegel S. Sphingosine-1-phosphate phosphohydrolase-1 regulates ER stress-induced autophagy. CellDeath Differ 2011;18:350–60.
[77] Krysko DV, Garg AD, Kaczmarek A, Krysko O, Agostinis P, Vandenabeele P.Immunogenic cell death and DAMPs in cancer therapy. Nat Rev Cancer2012;12:860–70.
[78] Maes H, Rubio N, Garg AD, Agostinis P. Autophagy: shaping the tumormicroenvironment and therapeutic response. Trends Mol Med 2013;19:428–430.
[79] de Jong OG, Verhaar MC, Chen Y, Vader P, Gremmels H, Posthuma G, et al.Cellular stress conditions are reflected in the protein and RNA content ofendothelial cell-derived exosomes. J Extracell Vesicles 2012;2012.
[80] Liu L, Yang M, Kang R, Wang Z, Zhao Y, Yu Y, et al. DAMP-mediated autophagycontributes to drug resistance. Autophagy 2011;7:112–20.
[81] Elliott MR, Chekeni FB, Trampont PC, Lazarowski ER, Kadl A, Walk SF, et al.Nucleotides released by apoptotic cells act as a find-me signal to promotephagocytic clearance. Nature 2009;461:282–90.
[82] Michaud M, Martins I, Sukkurwala AQ, Adjemian S, Ma Y, Pellegatti P, et al.Autophagy-dependent anticancer immune responses induced by chemother-apeutic agents in mice. Science 2011;334:1573–80.
[83] Corriden R, Insel PA. Basal release of ATP: an autocrine-paracrine mechanismfor cell regulation. Sci Signal 2010;3:re1.
[84] Garg AD, Dudek AM, Agostinis P. Calreticulin surface exposure is abrogated incells lacking, chaperone-mediated autophagy-essential gene, LAMP2A. CellDeath Dis 2013;4:e826.
[85] Garg AD, Dudek AM, Ferreira GB, Verfaillie T, Vandenabeele P, Krysko DV,et al. ROS-induced autophagy in cancer cells assists in evasionfrom determinants of immunogenic cell death. Autophagy 2013;9:1292–1300.
[86] Thorburn J, Horita H, Redzic J, Hansen K, Frankel AE, Thorburn A. Autophagyregulates selective HMGB1 release in tumor cells that are destined to die. CellDeath Differ 2009;16:175–80.
[87] Kang R, Zhang Q, Zeh 3rd HJ, Lotze MT, Tang D. HMGB1 in cancer: good, bad,or both? Clin Cancer Res Off J Am Assoc Cancer Res 2013;19:4046–57.
[88] Gebhardt C, Riehl A, Durchdewald M, Nemeth J, Furstenberger G, Muller-Decker K, et al. RAGE signaling sustains inflammation and promotes tumordevelopment. J Exp Med 2008;205:275–80.
[89] Mittal D, Saccheri F, Venereau E, Pusterla T, Bianchi ME, Rescigno M. TLR4-mediated skin carcinogenesis is dependent on immune and radioresistantcells. EMBO J 2010;29:2242–50.
[90] Liu L, Yang M, Kang R, Wang Z, Zhao Y, Yu Y, et al. HMGB1-inducedautophagy promotes chemotherapy resistance in leukemia cells. Leukemia2011;25:23–31.
[91] Yang Y, Fiskus W, Yong B, Atadja P, Takahashi Y, Pandita TK, et al. Acety-lated hsp70 and KAP1-mediated Vps34 SUMOylation is required forautophagosome creation in autophagy. Proc Natl Acad Sci U S A 2013;110:6841–6850.
[92] Thakor N, Holcik M. IRES-mediated translation of cellular messenger RNAoperates in eIF2alpha- independent manner during stress. Nucleic Acids Res2012;40:541–50.
[93] Caron E, Vincent K, Fortier MH, Laverdure JP, Bramoulle A, Hardy MP, et al.The MHC I immunopeptidome conveys to the cell surface an integrative viewof cellular regulation. Mol Syst Biol 2011;7:533.
[94] Granados DP, Yahyaoui W, Laumont CM, Daouda T, Muratore-Schroeder TL,Cote C, et al. MHC I-associated peptides preferentially derive from transcriptsbearing miRNA response elements. Blood 2012;119:e181–90.
[95] Li J, Gong J, Li P, Li M, Liu Y, Liang S, et al. Knockdown of microRNA-155 inKupffer cells results in immunosuppressive effects and prolongs survival ofmouse liver allografts. Transplantation 2014;97:626–30.
[96] Gibbings D, Mostowy S, Voinnet O. Autophagy selectively regulates miRNAhomeostasis. Autophagy 2013;9:781–90.
[97] Frankel LB, Lund AH. MicroRNA regulation of autophagy. Carcinogenesis2012;33:2018–20.
[98] Li Y, Wang LX, Yang G, Hao F, Urba WJ, Hu HM. Efficient cross-presentationdepends on autophagy in tumor cells. Cancer Res 2008;68:6889–90.
[99] Uhl M, Kepp O, Jusforgues-Saklani H, Vicencio JM, Kroemer G, Albert ML.Autophagy within the antigen donor cell facilitates efficient antigencross-priming of virus-specific CD8+ T cells. Cell Death Differ 2009;16:991–1000.
[100] Li Y, Hahn T, Garrison K, Cui ZH, Thorburn A, Thorburn J, et al. The vitamin Eanalogue alpha-TEA stimulates tumor autophagy and enhances antigencross-presentation. Cancer Res 2012;72:3535–40.
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
[101] Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell2010;140:883–90.
[102] Balkwill F. Tumour necrosis factor and cancer. Nat Rev Cancer 2009;9:361–370.
[103] Lock R, Kenific CM, Leidal AM, Salas E, Debnath J. Autophagy-dependentproduction of secreted factors facilitates oncogenic RAS-driven invasion.Cancer Discov 2014;4:466–70.
[104] Nakahira K, Haspel JA, Rathinam VA, Lee SJ, Dolinay T, Lam HC, et al.Autophagy proteins regulate innate immune responses by inhibiting therelease of mitochondrial DNA mediated by the NALP3 inflammasome. NatImmunol 2011;12:222–30.
[105] Dupont N, Jiang S, Pilli M, Ornatowski W, Bhattacharya D, Deretic V. Autop-hagy-based unconventional secretory pathway for extracellular delivery ofIL-1beta. EMBO J 2011;30:4701–10.
[106] Harris J. Autophagy and cytokines. Cytokine 2011;56:140–4.[107] Zhan Z, Xie X, Cao H, Zhou X, Zhang XD, Fan H, et al. Autophagy facilitates
TLR4- and TLR3-triggered migration and invasion of lung cancer cellsthrough the promotion of TRAF6 ubiquitination. Autophagy 2014;10:257–260.
[108] Robbins PD, Morelli AE. Regulation of immune responses by extracellularvesicles. Nat Rev Immunol 2014;14:195–200.
[109] Thery C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immuneresponses. Nat Rev Immunol 2009;9:581–90.
[110] King HW, Michael MZ, Gleadle JM. Hypoxic enhancement of exosome releaseby breast cancer cells. BMC Cancer 2012;12:421.
[111] Fader CM, Sanchez D, Furlan M, Colombo MI. Induction of autophagy pro-motes fusion of multivesicular bodies with autophagic vacuoles in k562 cells.Traffic 2008;9:230–40.
[112] Wang AL, Lukas TJ, Yuan M, Du N, Tso MO, Neufeld AH. Autophagy andexosomes in the aged retinal pigment epithelium: possible relevance todrusen formation and age-related macular degeneration. PLoS ONE2009;4:e4160.
[113] Noman MZ, Buart S, Van Pelt J, Richon C, Hasmim M, Leleu N, et al. Thecooperative induction of hypoxia-inducible factor-1 alpha and STAT3 duringhypoxia induced an impairment of tumor susceptibility to CTL-mediated celllysis. J Immunol 2009;182:3510–20.
[114] Noman MZ, Janji B, Kaminska B, Van Moer K, Pierson S, Przanowski P, et al.Blocking hypoxia-induced autophagy in tumors restores cytotoxic T-cellactivity and promotes regression. Cancer Res 2011;71:5976–80.
[115] Hasmim M, Noman MZ, Lauriol J, Benlalam H, Mallavialle A, Rosselli F, et al.Hypoxia-dependent inhibition of tumor cell susceptibility to CTL-mediat-ed lysis involves NANOG induction in target cells. J Immunol 2011;187:4031–4040.
[116] Noman MZ, Buart S, Romero P, Ketari S, Janji B, Mari B, et al. Hypoxia-inducible miR-210 regulates the susceptibility of tumor cells to lysis bycytotoxic T cells. Cancer Res 2012;72:4629–30.
[117] Noman MZ, Janji B, Berchem G, Mami-Chouaib F, Chouaib S. Hypoxia-induced autophagy: a new player in cancer immunotherapy. Autophagy2012;8:704–10.
[118] Akalay I, Janji B, Hasmim M, Noman MZ, Andre F, De Cremoux P, et al.Epithelial-to-mesenchymal transition and autophagy induction in breastcarcinoma promote escape from T-cell-mediated lysis. Cancer Res2013;73:2418–20.
[119] Akalay I, Janji B, Hasmim M, Noman MZ, Thiery JP, Mami-Chouaib F, et al. EMTimpairs breast carcinoma cell susceptibility to CTL-mediated lysis throughautophagy induction. Autophagy 2013;9:1104–10.
[120] Siemens H, Jackstadt R, Hunten S, Kaller M, Menssen A, Gotz U, et al. miR-34and SNAIL form a double-negative feedback loop to regulate epithelial-mesenchymal transitions. Cell Cycle 2011;10:4256–60.
[121] Yu Y, Yang L, Zhao M, Zhu S, Kang R, Vernon P, et al. Targeting microRNA-30a-mediated autophagy enhances imatinib activity against human chronicmyeloid leukemia cells. Leukemia 2012;26:1752–60.
[122] Baginska J, Viry E, Berchem G, Poli A, Noman MZ, van Moer K, et al. GranzymeB degradation by autophagy decreases tumor cell susceptibility to naturalkiller-mediated lysis under hypoxia. Proc Natl Acad Sci U S A 2013;110:17450–17460.
[123] Viry E, Baginska J, Berchem G, Noman MZ, Medves S, Chouaib S, et al.Autophagic degradation of GZMB/granzyme B: a new mechanism of hypoxictumor cell escape from natural killer cell-mediated lysis. Autophagy2014;10:173–80.
[124] Buchser WJ, Laskow TC, Pavlik PJ, Lin HM, Lotze MT. Cell-mediated autophagypromotes cancer cell survival. Cancer Res 2012;72:2970–80.
[125] Sui X, Chen R, Wang Z, Huang Z, Kong N, Zhang M, et al. Autophagy andchemotherapy resistance: a promising therapeutic target for cancer treat-ment. Cell Death Dis 2013;4:e838.
[126] Li JL, Han SL, Fan X. Modulating autophagy: a strategy for cancer therapy.Chin J Cancer 2011;30:655–60.
[127] He WS, Dai XF, Jin M, Liu CW, Rent JH. Hypoxia-induced autophagy confersresistance of breast cancer cells to ionizing radiation. Oncol Res2012;20:251–60.
[128] Chaachouay H, Ohneseit P, Toulany M, Kehlbach R, Multhoff G, RodemannHP. Autophagy contributes to resistance of tumor cells to ionizing radiation.Radiother Oncol 2011;99:287–90.
[129] Gong C, Bauvy C, Tonelli G, Yue W, Delomenie C, Nicolas V, et al. Beclin 1 andautophagy are required for the tumorigenicity of breast cancer stem-like/progenitor cells. Oncogene 2013;32:2261–70. 72e 1–11.
e metabolic response to stress shaping the antitumor immunity.006
E. Viry et al. / Biochemical Pharmacology xxx (2014) xxx–xxx12
G Model
BCP-12021; No. of Pages 12
[130] Guan JL, Simon AK, Prescott M, Menendez JA, Liu F, Wang F, et al. Autophagyin stem cells. Autophagy 2013;9:830–40.
[131] Amaravadi RK, Lippincott-Schwartz J, Yin XM, Weiss WA, Takebe N,Timmer W, et al. Principles and current strategies for targeting autophagyfor cancer treatment. Clin Cancer Res Off J Am Assoc Cancer Res 2011;17:654–660.
Please cite this article in press as: Viry E, et al. Autophagy: An adaptivBiochem Pharmacol (2014), http://dx.doi.org/10.1016/j.bcp.2014.07.
[132] Liang X,De Vera ME, BuchserWJ, RomodeVivar ChavezA, LoughranP, BeerStolz D,et al. Inhibiting systemic autophagy during interleukin 2 immunotherapy pro-motes long-term tumor regression. Cancer Res 2012;72:2791–801.
[133] Li Y, Wang LX, Pang P, Cui Z, Aung S, Haley D, et al. Tumor-derived autopha-gosome vaccine: mechanism of cross-presentation and therapeutic efficacy.Clin Cancer Res Off J Am Assoc Cancer Res 2011;17:7047–50.
e metabolic response to stress shaping the antitumor immunity.006





























