Embed Size (px)
Citation preview

Author's personal copy
Environmental and Experimental Botany 69 (2010) 343–352
Contents lists available at ScienceDirect
Environmental and Experimental Botany
journa l homepage: www.e lsev ier .com/ locate /envexpbot
Bacillus pumilus ES4: Candidate plant growth-promoting bacterium to enhanceestablishment of plants in mine tailings
Luz E. de-Bashana,b, Juan-Pablo Hernandezb, Yoav Bashana,b,∗, Raina M. Maiera
a Department of Soil, Water and Environmental Science, University of Arizona, Tucson, AZ, USAb Environmental Microbiology Group, Northwestern Center for Biological Research (CIBNOR), La Paz, B.C.S., Mexico
a r t i c l e i n f o
Article history:Received 23 December 2009Received in revised form 13 April 2010Accepted 22 April 2010
Keywords:BacillusDGGEFISHPhytostabilizationMine tailings
a b s t r a c t
Three plant growth-promoting bacteria (PGPB; Bacillus pumilus ES4, B. pumilus RIZO1, and Azospirillumbrasilense Cd) were tested for their ability to enhance plant growth and development of the native SonoranDesert shrub quailbush (Atriplex lentiformis) and for their effect on the native bacterial community inmoderately acidic, high-metal content tailings (AHMT) and in neutral, low-metal content natural tailings(NLMT) in controlled greenhouse experiments. Inoculation of quailbush with all three PGPB significantlyenhanced plant growth parameters, such as germination, root length, dry weight of shoots and roots, androot/shoot ratio in both types of tailings. The effect of inoculation on the indigenous bacterial communityby the most successful PGPB B. pumilus ES4 was evaluated by denaturating gradient gel electrophoresis(PCR-DGGE) fingerprinting and root colonization was followed by specific fluorescent in situ hybridization(FISH). Inoculation with this strain significantly changed the bacterial community over a period of 60days. FISH analysis showed that the preferred site of colonization was the root tips and root elongationarea. This study shows that inoculation of native perennial plants with PGPB can be used for developingtechnologies for phytostabilizing mine tailings.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
In the deserts of the southwestern United States and northernMexico, large mounds of mine tailings, the main waste product ofmineral ore processing of abandoned and productive mines, arepotentially a health hazard to nearby urban populations in theform of wind-blown dust and ground water sources. This hap-pens because tailings, lacking plant cover and soil structure, easilydegrade by wind and rain action and serve as a continuous sourceof metal pollution (Pilon-Smits, 2005). Phytostabilization, usingnative plants has been proposed as an economic strategy to reducethese hazards (McCutcheon and Schnoor, 2003). This phytoreme-diation approach involves creation of a plant cover on the tailingsthat is sufficient to prevent erosion. A major challenge is that someof these tailings cannot serve, in their natural state, as growth sub-strate for most plant species because of metal toxicity, low pH, lackof essential minerals, lack of clay and organic matter to retain water,lack of soil structure, lack of a seed bank of nearby native plants,or some combination of these environmental parameters (Mendezand Maier, 2008). Thus, tailings can remain devoid of plants fordecades or have only a slight plant cover (Gonzalez-Chavez et al.,
∗ Corresponding author at: Environmental Microbiology Group, CIBNOR, MarBermejo 195, Colonia Playa Palo de Santa Rita, La Paz, B.C.S. 23090, Mexico.
E-mail address: [email protected] (Y. Bashan).
2009). A partial solution and an upgrade of this inhospitable sub-strate to a “soil-like” status can be accomplished by adding largequantities of compost, biosolids, and irrigation (Ye et al., 2001; Chiuet al., 2006). The drawback of the compost-water strategy is thatapplying large quantities of compost is often not economically fea-sible because of the extensive area of tailings and the remotenessof the sites. Furthermore, water and irrigation facilities are largelyabsent in deserts, especially at long-abandoned mines.
Inoculation with plant growth-promoting bacteria (PGPB;Bashan and Holguin, 1998) has been proposed to aid in estab-lishment of plants on tailings at reduced compost concentrations(Petrisor et al., 2004; Zhuang et al., 2007; Mendez and Maier,2008). Successful attempts have been made to isolate and usepotential PGPB from tailings (Grandlic et al., 2008). The premisefor this approach is that PGPB isolated from tailings may aid inplant survival and growth through several mechanisms including:increasing nutrient availability to plants, increasing plant resis-tance to metal toxicity, or decreasing toxic metal bioavailabilityin the rhizosphere (Burd et al., 1998, 2000; Pishchik et al., 2002;Glick, 2003; Belimov et al., 2004; Reed and Glick, 2005; Reed et al.,2005; Vivas et al., 2006; Wu et al., 2006a; Li et al., 2007; Rajkumarand Freitas, 2008a,b).
Alternatively, there are many well-studied PGPB used in otherapplications that could be useful in helping plant establishmentin mine tailings. For example, strains from the genus Bacillushave been shown to enhance the growth of agricultural crops,
0098-8472/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.envexpbot.2010.04.014

Author's personal copy
344 L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352
wild plants, trees, microalgae, and model plants, through differ-ent mechanisms of plant growth-promotion (Bashan et al., 2000;Enebak et al., 1998; Hernandez et al., 2009; Kloepper et al., 2004a,b;Ryu et al., 2005; Vessey, 2003). Bacillus is also a genus com-monly found in various types of mine tailings (Natarajan, 1998;Vijayalakshmi and Raichur, 2003; Wu et al., 2006b; Tsuruta, 2007;Zhang et al., 2007). A second genus, Azospirillum, is also commonlyused for its PGP activities. Azospirillum is perhaps the best studiedof the PGPB except for rhizobia (Bashan et al., 2004).
The hypothesis of this study is that inoculation with classi-cal agricultural PGPB can support establishment and growth ofthe common, native Sonoran Desert shrub, quailbush (Atriplexlentiformis) in mine tailings at reduced compost concentrations. Thespecific objectives were to: (1) Evaluate the growth of quailbushin two types of tailings: an acidic, high-metal content tailing andneutral low-metal content tailing supplemented with a suboptimallevel of compost and inoculated with one of three PGPB (Bacilluspumilus ES4, B. pumilus RIZO1, and A. brasilenese Cd), comparingthe performance of the three PGPB on plant growth; (2) Monitorroot colonization capacity of the most successful PGPB using thetechnique of fluorescent in situ hybridization (FISH) and confocallaser microscopy; and (3) Assess whether there is an effect in struc-ture on the tailings bacterial community by PGPB inoculation usingPCR-DGGE fingerprint analysis.
2. Material and methods
2.1. Organisms, bacterial growth conditions and inoculantpreparation
The plant growth-promoting bacteria Azospirillum brasilense Cd(DSM 1843, the type strain of A. brasilense; Braunschweig, Ger-many), B. pumilus ES4, and B. pumilus RIZO1 (Puente et al., 2004) andthe evergreen shrub quailbush (A. lentiformis) native to southwest-ern North America, served as model organisms. The two B. pumilusstrains were identified by sequencing their entire 16S rRNA gene(100% similarity to deposited B. pumilus) and their sequences weredeposited in the GenBank (B. pumilus RIZO1 accession number isFJ032016 and B. pumilus ES4 is FJ032017). Both B. pumilus strainsare diazotrophs, where strain ES4 is also a phosphate solubilizer.Both strains can degrade various types of rocks, were isolated fromthe rhizoplane of a cactus, and promoted growth of the giant car-don cactus. Strain ES4 also promoted the growth of the microalgaeChlorella vulgaris by supplying it with fixed atmospheric nitrogen(Hernandez et al., 2009; Puente et al., 2004). Nomenclature of PGPBin the latter reference differs from the current nomenclature as B.pumilus.
The PGPB were cultivated on nutrient broth (Sigma) for 24 hat 30 ◦C and stirred at 120 rpm. The resulting bacteria cultureswere formulated into a dry, microbead inoculant made of alginate,as described by Bashan et al. (2002), using specialized equip-ment (http://www.bashanfoundation.org/bead.html, accessed 8April, 2010). Alginate inoculants, during numerous greenhouseand field experiments, proved to be an inert and reliable car-rier for inoculation of PGPB for crops (Bashan, 1986a,b; Bashanet al., 2002) and various desert plants (Bashan et al., 2009a,b;Grandlic et al., 2009). Bacterial microbead inoculant was mixedwith wild quailbush seeds in the same planting hole in the green-house experiment at a level of 1.2 × 106 CFU g−1 soil, as describedbelow.
2.2. Tailings characterization
Two types of substrate were used: AHMT and NLMT. The phys-iochemical properties of AHMT were previously characterized
(Grandlic et al., 2008). A single composite sample of NLMT tailingswas oven-dried at 105 ◦C, sieved through a 2-mm mesh, and ana-lyzed for texture (Soil Survey Laboratory Methods Manual, 2004).Total organic carbon (TOC) and total nitrogen (TN) were measuredin the solid phase using a Shimadzu high temperature combustionTOC/TN analyzer (organic and inorganic nitrogen). Electrical con-ductivity (EC), pH, dissolved organic carbon (DOC), and dissolvednitrogen (DN, organic plus inorganic) were measured in a saturatedpaste extract (1:1) in deionized water. Samples were prepared bymicrowave acid digestion (EPA method 3051; EPA, 2004) for totalelement (As, Cd, Cu, Fe, K, Mn, Na, Pb, Zn) analysis by ICP–MS. Anal-ysis was done as a service by a laboratory in the Department of Soil,Water, and Environmental Science University of Arizona, Tucson,AZ. Total heterotrophic bacterial counts were done by suspending1 g of tailing material in sterile 0.85% saline solution with shakingat 120 rpm for 1 h; serial dilution were done in 0.85% saline solu-tion and plated on nutrient agar; the plates were incubated at 37 ◦Cand developed colonies were counted. Enumerated bacteria wereexpressed as CFU g−1 soil.
2.3. Greenhouse cultivation
Three treatments were tested in this experiment. Treatment 1evaluated the effect of three individual PGPB on the growth of quail-bush in composted tailings. Treatment 2 evaluated the growth ofquailbush in composted tailings in the absence of PGPB (the uninoc-ulated control). Treatment 3 was an unplanted control consistingof pots of composted tailings that did not receive either plants orPGPB inoculation. Mature compost with relatively low levels of res-ident bacteria (de-Bashan et al., unpublished data) was applied. Thephysico-chemical compositions of the compost were previouslydescribed (Mendez et al., 2007).
In this experiment, quailbush was grown from seeds(Carter Seeds, Vista, CA, USA) in black plastic commercial pots(15 cm × 10 cm dia.) with a drainage hole, each containing a thor-oughly mixed substrate of 530 g made of 9:1 (non-sieved tailings:dairy compost, w/w). As discussed by Grandlic et al. (2008), this isa suboptimal compost concentration for these tailings (the optimaltailings:compost ratio is 17:3). Initially, 20 seeds per pot wereplanted at ∼0.5 cm with a sterile tweezers and were checked forgermination 5 days later (day 0). Then, each pot was thinned to 10seedlings. At day 5, one seedling was removed from each pot. Thisprocedure was repeated at days 15, 30, and 60.
At the end of the experiment, 6 plants per pots were left andwere used for measuring growth parameters and for FISH analy-sis of their roots, as explained below. A total of 50 pots were used(10 per treatment in a randomized layout). All plants were grownin a greenhouse on elevated metal net beds at controlled temper-ature (20–30 ◦C, n/d) and drip irrigated with tap water every 3 hwith 93.4 ml pot−1. These are optimal irrigation conditions for thisspecies. The experiment lasted 60 days, after which plant growthparameters were measured.
2.4. Extraction of DNA from soil and PCR amplification
“Rhizosphere tailing soil” was defined according to the originaldefinition of rhizosphere soil by Hiltner (Hartmann et al., 2008),namely, tailing particles adhere to the roots after the plant wasremoved from the tailing substrate. Rhizosphere samples from eachtreatment (inoculated plants, non-inoculated plants, soil amendedwith compost without plant, and compost) were collected in ster-ile 1.5-ml snap cap tubes and stored at −80 ◦C until analysis. DNAwas extracted using a kit (Fast DNA SPIN for soils, MP Biomedi-cals, Solon, OH), following the manufacturer’s protocol. In samplescontaining humic acids, the binding matrix–DNA complex was fur-ther rinsed with saturated 5.5 M guanidine thiocyanate (Flucka

Author's personal copy
L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352 345
Sigma–Aldrich, Buchs, Switzerland) until the supernatant lost itsbrown tint (Rosario et al., 2007). Samples (0.4 g soil) were usedfor each extraction and three replicates for each treatment wereanalyzed.
Once extracted, DNA was quantified in a mini-fluorometer (TBS380, Turner BioSystems, Sunnyvale, CA), using the protocol forQuant-iT PicoGreen dsDNA (Molecular Probes, Eugene, OR). About100 pg �l−1 of each sample was used for PCR reaction.
2.4.1. Polymerase chain reaction (PCR)The V9 variable region of the 16S rRNA gene was PCR ampli-
fied using the bacteria primers 1070F (5′-ATG GCT GTC GTC AGCT-3′) and 1406R (5′-ACG GGC GGT GTG TAC-3′) with a 40-bpGC clamp (Ferris et al., 1996). PCR for DGGE was done using amodification of the method of Colores et al. (2000). Briefly, 25 �lof the reaction mix contained 2.5 �l of 10× buffer with 15 mMMgCl2 (Qiagen Sciences, Germantown, MD), 200 �M dNTP, 0.4 �Mof each primer, 5% dimethyl sulfoxide (Sigma, St. Louis, MO),0.4 g l−1 unacetylated bovine serum albumin (Sigma), 0.5 units �l−1
HotStarTaq DNA polymerase (Qiagen Sciences), and 100 pg of com-munity DNA extract. Reaction was run on an automated PCRthermoblock (Peltier thermal cycler 200, MJ Research, Waltham,MA) for 95 ◦C for 15 min and 30 cycles (94 ◦C for 45 s, 55 ◦C for45 s. 72 ◦C for 30 s), followed by a 72 ◦C extension for 7 min.PCR products were viewed after electrophoresis by running a2% agarose gel (GenePure LE ISC BioExpress, Kaysville, UT), fol-lowed by ethidium bromide (EMD Chemicals, Darmstat, Germany)staining.
2.5. Denaturing gradient gel electrophoresis (DGGE) analysis
DGGE of the 16s rRNA gene products was performed usinga D-Code Universal Mutation Detection System (Bio-Rad Labora-tories, Hercules, CA). Acrylamide gels (7%) were prepared witha 45–70% urea-formamide denaturing gradient, according to themanufacturer’s protocol. Lanes were loaded with 10–15 �l of PCRproduct.
Electrophoresis was run at a constant 50 V for 16.5 h at 60 ◦C.Gels were stained with SYBR Green I nucleic acid gel stain (Lonza,Rockland, ME) and gel images were recorded with an AlphaImagerHP (Alpha Innotech, San Leandro, CA) gel documentation system.DGGE profiles were analyzed using Quantity One 4.5.2 software(Bio-Rad Laboratories).
2.6. Probes for fluorescent in situ hybridization (FISH)
Two probes were used: EUB338 (Amman et al., 1990), which isspecific for the domain Bacteria and the specific probe that tar-gets the complementary region of 16S rRNA in Bacillus (BAC07;Probe Base accession number: pB00403; Liu et al., 2001). BAC07was checked with the Probe Match tool of the Ribosomal DatabaseProject and with Greengenes (http://greengenes.ibl.gov). Oligonu-cleotides probes were labeled at the 5′ end with the fluorescentdyes Cy3 (specific probe) and Cy5 (universal probe) (IntegratedDNA Technologies, Coralville, IA).
2.6.1. Preparation of roots for fluorescent in situ hybridizationRoot samples were taken at 15, 30, and 60 days. Roots were
carefully separated from the soil, washed with 1 × PBS (200 mMNaH2PO4, 200 mM Na2HPO4), and fixed with 4% paraformalde-hyde (Acros Organics, Geel, Belgium) for 2 h at −4 ◦C. After fixation,roots were washed with 1 × PBS and stored in a mix of 1 × PBS/96%Ethanol (1:1 v/v) at −20 ◦C until hybridization.
2.6.2. In situ hybridizationFISH analysis was performed as described by Iverson and Maier
(2009). Briefly, slices of the fixed root were thawed and placedon a gelatin (0.1%, w/v, 0.01%, w/v chromium potassium sulfate)-coated microscope slides and fixed to the slide by adding 1 dropof warm, low-melt agarose solution (0.2%, w/v; Agarose LMP,Promega, Madison, WI) and dried at 37 ◦C for 45 min. Samples weredehydrated by successive 50%, 80%, and 96% ethanol washes (3 mineach), then air dried. Prior to hybridization, dehydrated roots weretreated with 10 mg ml−1 lysozyme (Sigma) for 15 min to enhancepermeation of the probe into the Gram-positive Bacillus (Mogge etal., 2000).
Hybridization was performed at 15% stringency at 46 ◦C for2 h (Daims et al., 2005). The final concentration of the probe was3 ng �l−1. A careful washing was performed at 48 ◦C for 15 min with50 ml warm washing buffer. The slides were then rinsed for a fewseconds with ice-cold deionized water and then air dried. Slides notvisualized immediately were stored at −20 ◦C in the dark.
2.6.3. Visualization of root colonization by confocal laserscanning microscopy
Before visualization, the slides were mounted in AF1 anti-fading reagent (Citifluor, Electron Microscopy Sciences Hatfield,PA). Microscopy was performed with a confocal scanning lasermicroscope (LMS 510, Carl Zeiss MicroImaging, Thornwood, NY). B.pumilus and other bacteria were viewed simultaneously by usingtwo helium neon lasers for excitation of the dyes Cy3 at 543 nm andCy5 at 633 nm. The pinhole was set for both channels at 1.2 Airyunits. Samples were observed with a 63× water immersion objec-tive (Carl Zeiss MicroImaging). For visualization, fluorescence fromCy3 and Cy5-labeled oligonucleotide probes were assigned greenand red colors, respectively. B. pumilus cells were identified whenthe two single-color images were merged into one picture that pro-duced a yellow color. Other bacterial colonies appeared red in thephotomicrographs.
The images were combined and analyzed with a software (LSM510, version 4.2 (Carl Zeiss International, Oberkochen, Germany).Images were processed with Adobe Photoshop, version 8.0 (AdobeSystems, Mountain View, CA).
2.7. Measurement of plant parameters
Plant growth parameters measured at day 60 of cultivationwere: shoot and root length, number of developing leaves, anddry weight of shoot and roots. Once harvested, roots and shootswere separated and rinsed gently under running water to removeall substrate particles. Plant roots and shoots were then placed inindividual aluminum foil packets, dried for 48 h in a 60 ◦C oven, andimmediately weighed (Bashan and de-Bashan, 2005b).
2.8. Experimental design and statistical analysis
Greenhouse experiments were conducted in a completely ran-domized design with final six plants per pot (30 plants pertreatment and 150 plants per experiment). The experiment wasanalyzed statistically and repeated. The presented data is from oneexperiment. After normalization of the data, results of all experi-ments were analyzed by one-way ANOVA and then by Tukey’s HSDpost hoc analysis. Significance was set at P < 0.05. Data in percentagewas converted to arcsine before analysis. These statistical measuresused Statistica, ver. 6.0 (StatSoft, Tulsa, OK).
The profiles obtained from DGGE gels were analyzed for simi-larity using the Dice coefficient. A dendrogram was formed fromthe unweighted pair group matching band average (UPGMA). Sim-ilarity varies from 0 to 1, where 1 indicates 100% similarity.Additionally, similarities among profiles for all treatments at each
Author's personal copy
346 L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352
time frame were evaluated by the Kruskal’s isotonic multidimen-sional scaling analysis (KNMDS) (Venables and Ripley, 2002) usingthe statistical software R (R Foundation for Statistical Computing,Vienna, Austria), as described for mine tailings by Rosario et al.(2007) at the significance level of P < 0.05. A stress factor was cal-culated to reflect goodness-of-fit of the models. Values <0.1 areconsidered a good fit.
3. Results
3.1. Mine tailing analysis
The NLMT tailings have the following physiochemical prop-erties: pH 7.0; 0.2% total carbon; 0.02% total organic carbon;<0.01% total nitrogen; 450 mg kg−1 total phosphorus; 1.4 mg kg−1
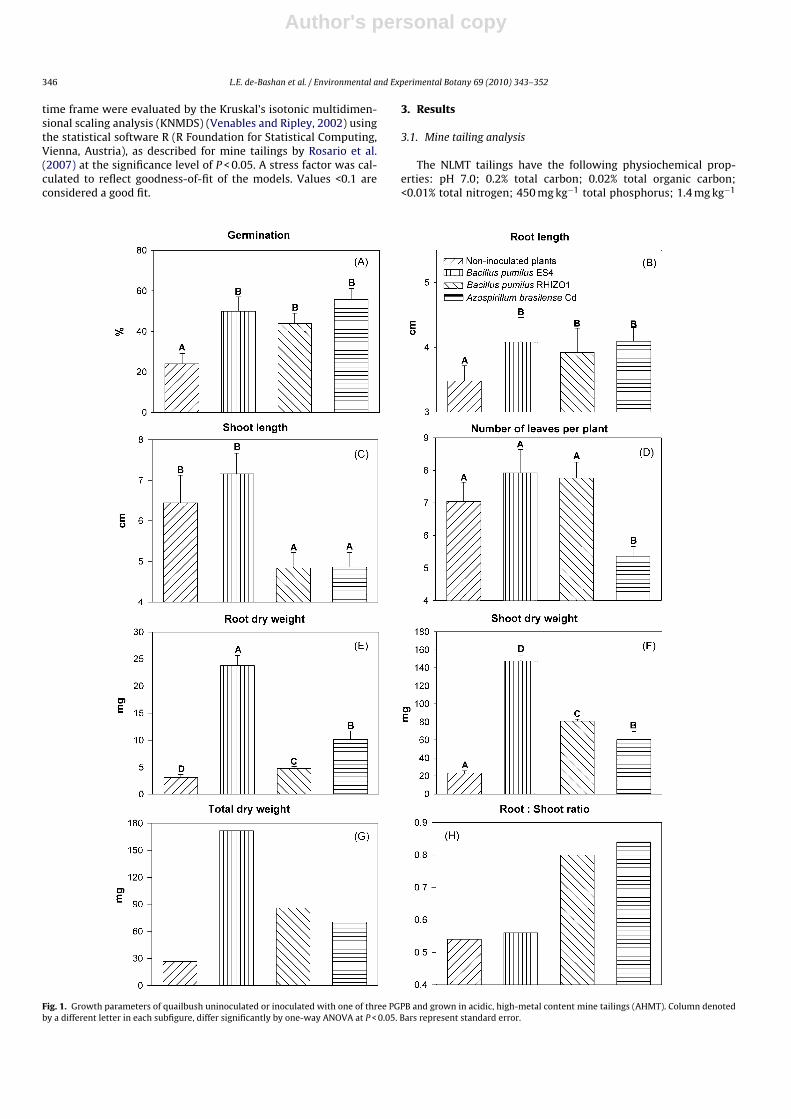
Fig. 1. Growth parameters of quailbush uninoculated or inoculated with one of three PGPB and grown in acidic, high-metal content mine tailings (AHMT). Column denotedby a different letter in each subfigure, differ significantly by one-way ANOVA at P < 0.05. Bars represent standard error.
Author's personal copy
L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352 347
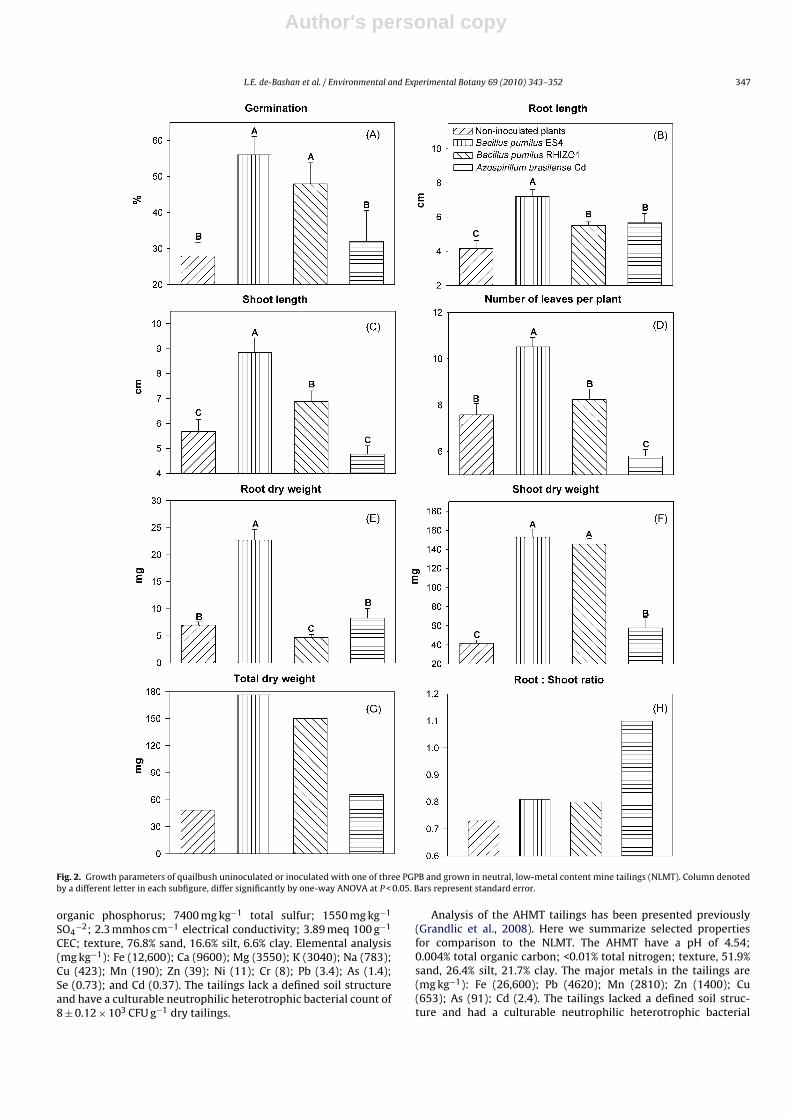
Fig. 2. Growth parameters of quailbush uninoculated or inoculated with one of three PGPB and grown in neutral, low-metal content mine tailings (NLMT). Column denotedby a different letter in each subfigure, differ significantly by one-way ANOVA at P < 0.05. Bars represent standard error.
organic phosphorus; 7400 mg kg−1 total sulfur; 1550 mg kg−1
SO4−2; 2.3 mmhos cm−1 electrical conductivity; 3.89 meq 100 g−1
CEC; texture, 76.8% sand, 16.6% silt, 6.6% clay. Elemental analysis(mg kg−1): Fe (12,600); Ca (9600); Mg (3550); K (3040); Na (783);Cu (423); Mn (190); Zn (39); Ni (11); Cr (8); Pb (3.4); As (1.4);Se (0.73); and Cd (0.37). The tailings lack a defined soil structureand have a culturable neutrophilic heterotrophic bacterial count of8 ± 0.12 × 103 CFU g−1 dry tailings.
Analysis of the AHMT tailings has been presented previously(Grandlic et al., 2008). Here we summarize selected propertiesfor comparison to the NLMT. The AHMT have a pH of 4.54;0.004% total organic carbon; <0.01% total nitrogen; texture, 51.9%sand, 26.4% silt, 21.7% clay. The major metals in the tailings are(mg kg−1): Fe (26,600); Pb (4620); Mn (2810); Zn (1400); Cu(653); As (91); Cd (2.4). The tailings lacked a defined soil struc-ture and had a culturable neutrophilic heterotrophic bacterial

Author's personal copy
348 L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352
Fig. 3. A comparison of the bacterial community structure from the three treatments studied (composted tailings planted and inoculated with Bacillus pumilus ES4, uninocu-lated planted composted tailings, unplanted uninoculated composted tailings at 15 (A), 30 (B), and 60 (C) days. The tailings used for this analysis was the neutral, low-metalcontent mine tailings (NLMT). Each subfigure contains the original DGGE gel and a dendrogram and Kruskal’s isotonic multidimentional scaling analysis derived from thedata of that gel. Data in circles are grouped together and differ from data from other groups at the indicated level of significance.
count of 2.95 ± 0.97 × 103 CFU g−1 dry tailings (de-Bashan et al.,unpublished data).
3.2. Effect of inoculation with three PGPB on growth parametersof quailbush
Promotion of growth of quailbush after inoculation with PGPBwas significant, and variable in the two tailings. In the AHMT, thethree PGPB increased seed germination (Fig. 1A) and root length(Fig. 1B). None of the PGPB significantly increased shoot length(Fig. 1C) or the number of leaves per plant (Fig. 1D). All PGPB strainsenhanced root, shoot, and total dry weight of the plants (Fig. 1E–G);however, B. pumilus ES4 had the greatest effect in these parameters.The root:shoot ratio was enhanced only by B. pumilus RIZO1 and A.brasilense Cd.
In the NLMT, seed germination, shoot length, and the number ofleaves per plant were enhanced by the two B. pumilus strains butnot by A. brasilense Cd (Fig. 2A, C and D). Root length was enhancedby all PGPB strains (Fig. 2B). Root dry weight was enhanced only by
B. pumilus ES4 (Fig. 2E). Shoot dry weight and total dry weight ofplants were enhanced by the three PGPB (Fig. 2G). The root:shootratio was enhanced only by A. brasilense Cd (Fig. 2H).
3.3. Effect of B. pumilus ES4 inoculation on tailings bacterialcommunity structure
PCR-DGGE gels revealed that the rhizosphere bacterial commu-nity appeared to be different among the treatments, compostedmine tailings inoculated with B. pumilus ES4 and planted, uninoc-ulated planted controls, and unplanted controls at each time pointexamined (15, 30, and 60 days) (Fig. 3). Multidimensional scalinganalysis was used to evaluate the relationship among the repli-cate sample profiles from each treatment. At 15 days, the sampleprofiles grouped into three clusters that were significantly differ-ent from each other (P < 0.05, Fig. 3A). One cluster contained thetwo unplanted control samples, the second cluster contained twoof the three uninoculated control samples, and the third containedall of the inoculated samples as well as one uninoculated control

Author's personal copy
L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352 349
Fig. 4. Colonization of roots of quailbush by Bacillus pumilus ES4 detected by fluorescent in situ hybridization (FISH) using double hybridization with the specific probeBAC07 and the universal probe EUB338 and viewed by confocal laser microscopy. (A) Non-inoculated roots; (B and C) root tip and elongation zone 15 days after inoculation,respectively, (D) root elongation zone 60 days after inoculation (E) root tip of the same root 60 days after inoculation, and (F) root tip and elongation zone 60 days afterinoculation. Yellow colonies and yellow patches are labeled cells of Bacillus pumilus ES4 (marked with white arrows). Red colonies and red patches are labeled cells ofunidentified bacteria on the roots (marked with green arrows). Because all non-inoculated roots showed the same bacterial population, only one control microphotographis presented. RS = root surface; RT = root tip.
sample. At 30 days, there were three clusters (P = 0.006, Fig. 3B).The first contained the two unplanted control samples, the secondcluster contained the three uninoculated control samples, and thethird contained the three inoculated samples. Finally, at 60 daysthere were three clusters (P = 0.04, Fig. 3C). The first contained thetwo unplanted control samples and one uninoculated control sam-ple. The second contained the remaining two uninoculated controlsamples. The third contained all three inoculated samples.
3.4. Root colonization of quailbush plants by B. pumilus ES4
Bacterial colonization of same-age quailbush roots was exam-ined using two sets of FISH primers, one specific for B. pumilus ES4and one for the domain Bacteria. Originally, tailings had very lim-ited populations of unidentified bacteria and most root surfaceswere devoid of bacteria. This continued for the 60 days of the exper-iment (Fig. 4A). Inoculation with B. pumilus ES4 significantly andconsistently changed the visual rhizosphere profile of these roots.Specific detection of microcolonies of B. pumilus ES4 after day 15was observed exclusively in the root tip and elongation zone areaof the root (Fig. 4B and C) and this was observed up to day 60 of the
experiment (Fig. 4D and E). With time, the population of bacteriaof other species also increased and became dominant over the rootsurfaces (Fig. 4D–F).
4. Discussion
Phytostabilization of mine tailings is a type of phytoremediationthat uses plants capable of surviving on these stressed substrates,with the objective of eliminating dispersal of contaminant and par-ticulates from the site. In general, this has been done with largequantities of organic matter coupled with irrigation. Alternatively,the use of PGPB in phytostabilization of mine tailings is proposed(Petrisor et al., 2004; Zhuang et al., 2007; Mendez and Maier, 2008).There are two possibilities to consider in the search for PGPB com-patible with mine tailings. One is to isolate native, potential PGPBfrom tailings or from plants already growing in the tailings, prop-agate them, and use them as inoculants. This is a labor intensiveprocedure that has been successfully tried (Grandlic et al., 2008,2009). The other approach, described in this study, is to examineisolates from the large number of available and proven PGPB strains,mainly used in agriculture (Bashan and de-Bashan, 2005a).

Author's personal copy
350 L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352
All three PGPB used in this study proved to be growth promotersof quailbush in the two types of mine tailings tested. Although thetwo types of tailings were very different, the same small amountof compost was added to give the same baseline for comparisonof the performance of the PGPB. Addition of compost is essentialfor normal development of native plants in the desert on toxic andeven less toxic mine tailings (Iverson and Maier, 2009; Mendez andMaier, 2008; Grandlic C., pers. comm.). Inoculation with PGPB canreduce the amount of compost from 15% to 10% for raising quail-bush but in buffalo grass PGPB enhanced growth in the absence ofcompost for a period of time (Grandlic et al., 2008). Yet, for practi-cal revegetation of mine tailings, and for most native plant speciestested, complete elimination of compost was impossible (GrandlicC., pers. comm.); a significant difficulty for this technology, so far.Of these isolates, B. pumilus ES4 performed the best, as measured byplant biomass produced. Increasing plant mass at the whole plantlevel is a common effect of PGPB (Bashan and Dubrovsky, 1996).Consequently, it was chosen as the candidate PGPB for detailedstudies of root colonization and its effect on the native substratecommunity of bacteria. It is notable that the PGPB stimulated plantgrowth in both types of tailings even though the pH and metalcontent were very different.
Inoculation with B. pumilus ES4 significantly changed the tail-ings bacterial community structure as early as 15 days into theexperiment. The triplicate B. pumilus ES4 inoculated treatmentsamples were completely distinguishable from the unplanted con-trol by day 15 and from the uninoculated control by day 30. Theseresults are in contrast to three studies using the PGPB A. brasilensethat examined maize grown in pots in a controlled greenhouse.Analysis of the structure of the bacterial community in these exper-iment, using two fingerprinting methods, DGGE and automatedribosomal intergenic spacer analysis (ARISA), showed that inocu-lation with A. brasilense strains Cd and Sp 245 had no effect or verymarginal effect, at best, on the size or the structure of the bacterialcommunities and that plant age was far more significant in affect-ing bacterial communities (Herschkovitz et al., 2005a,b; Lerner etal., 2006). Another study employing tomatoes and A. brasilense andB. subtilis in a peat-based substrate and evaluated by DGGE showedsimilar trends (Felici et al., 2008).
The two tailings used in this study have an extremely low pop-ulation of heterotrophic bacteria (Mendez et al., 2007, this study),comparable in size to other tailings from the same area (Iversonand Maier, 2009). Even with the addition of 10% compost, the totalpopulation of heterotrophic bacteria in these tailings is relativelylow, depending on the type of tailings (103–105 CFU g−1, Iversonand Maier, 2009); 104–106 CFU g−1, Mendez et al., 2007). The lowinitial heterotrophic numbers is a possible and plausible explana-tion for the changes that B. pumilus ES4 induced in the bacterialcommunity of the tailings. In comparison to previous studies onchanging of bacterial community structure, maize,in those studies,is a large and fast-growing plant that produces a large amount ofroots in the pots and consequently transformed the entire soil of thepot to a rhizosphere-like mini-ecosystem where a large populationof bacteria exists, normally 108–109 CFU culturable bacteria g−1
soil. Similarly, this occurs with microbial populations in the tomatorhizosphere (Meyer and Linderman, 1986; de Brito-Alvarez et al.,1995). On this elevated native bacterial background, inoculationwith Azospirillum, normally done at the level of 106 CFU g−1 or about0.1–1% of the background population (Bashan, 1986a,b), has onlymarginal, if any, effect on the general population observed in theabove studies (Felici et al., 2008; Herschkovitz et al., 2005a,b; Lerneret al., 2006) and is below the limits of the molecular techniquesused. In complex microflora, PCR-DGGE does not detect microor-ganisms present at a level lower than 1% of the total microbialpopulation (Felske et al., 1998). However, our experiments weredone with quailbush, a relatively slow growing small plant with
roots that did not completely colonize the substrate even after 60days; this might have had a small effect on the relatively low initialbacterial community. For relatively low community size of bac-teria, as occurs in the tailings (Iverson and Maier, 2009; Mendezet al., 2007), inoculation of B. pumilus ES4 (106 CFU g−1) created alarge shift in the populations, allowing the molecular techniques todetect it.
To verify that the effects on plant growth resulted from inoc-ulation, it is essential to demonstrate that seeds inoculated withbacterial strains yield colonized roots. Numerous methods foridentifying root colonization by PGPB have been developed. Theyinclude traditional microbiology, immunology, molecular insertionof specific marker genes (Jacoud et al., 1998), insertion of the gfpgreen fluorescent protein (Ramos et al., 2002; Bacilio et al., 2004),lacZ markers (Arsene et al., 1994), and FISH (Kirchhof et al., 1997).Because the tailings were non-sterile, we employed a modified FISHtechnique to follow root colonization by the PGPB. FISH monitoredwith confocal laser microscopy is a powerful tool to detect PGPBin soils and rhizosphere samples (Assmus et al., 1995). B. pumiluscells were specifically detected by the FISH technique on the col-onized root tips and the elongation zone areas at days 15 and 60.Furthermore, the PGPB changed the bacterial colonization of thetailings by inducing proliferation of other rhizosphere bacteria onthe root surfaces, as detected by DGGE and FISH analyses. Prolif-eration of other bacteria may have been the result of an increasein exudation from the roots induced by the PGPB on the roots.Enhanced proton and organic acid extrusion are one of the mech-anisms proposed for PGPB (Bashan and Levanony, 1991; Carrillo etal., 2002; Bashan and de-Bashan, 2005a). Taken together, this sug-gests that this PGPB has sufficient rhizosphere fitness to serve asan effective PGPB. Rhizosphere fitness is a major condition for thesuccess of any PGPB (Lugtenberg and Dekkers, 1999; Bashan et al.,2004).
In summary, this study identified an efficient PBPB strain to pro-mote growth of a native plant in two distinct types of mine tailingsand its capacity to modify the community of bacteria of the tailings.
Acknowledgements
We thank Julia Neilsen, Antje Legatzki, Sadie Iverson, andFernando Solis-Dominguez from our research group at the Uni-versity of Arizona for advice and discussion, Anton Hartmann andMichael Schmid of the Hemholtz Zentrum, München, Germany foradvice on FISH development, and Christopher Grandlic at SyntheticGenomics, California, for advice on compost application in tailings.This research was supported by the National Institute of Environ-mental Health Sciences Superfund Basic Research Program, NIH,USA (grant 2 P42 ES04940-11) and by Consejo Nacional de Cienciay Tecnologia of Mexico (CONACYT basic research grant 50052-Z).
References
Amman, R.I., Binder, B.J., Olson, R.J., Chisholm, S.W., Devereux, R., Stahl, D.A., 1990.Combination of 16S rRNA-targeted oligonucleotide probes with flow cytom-etry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 56,1919–1925.
Arsene, F., Katupitiya, S., Kennedy, I.R., Elmerich, C., 1994. Use of LacZ fusions tostudy the expression of nif genes of Azospirillum brasilense in association withplants. Mol. Plant: Microbe Int. 7, 748–757.
Assmus, B., Hutzler, P., Kirchhof, G., Amann, R., Lawrence, J.R., Hartmann, A., 1995. Insitu localization of Azospirillum brasilense in the rhizosphere of wheat with fluo-rescently labeled rRNA-targeted oligonucleotide probes and scanning confocallaser microscopy. Appl. Environ. Microbiol. 61, 1013–1019.
Bacilio, M., Rodriguez, H., Moreno, M., Hernandez, J.-P., Bashan, Y., 2004. Mitigationof salt stress in wheat seedlings by a gfp-tagged Azospirillum lipoferum. Biol.Fertil. Soils 40, 188–193.
Bashan, Y., 1986a. Significance of timing and level of inoculation with rhizospherebacteria on wheat plants. Soil Biol. Biochem. 18, 297–301.
Bashan, Y., 1986b. Alginate beads as synthetic inoculant carriers for the slow releaseof bacteria that affect plant growth. Appl. Environ. Microbiol. 51, 1089–1098.

Author's personal copy
L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352 351
Bashan, Y., de-Bashan, L.E., 2005a. Bacteria/plant growth-promotion. In: Hillel, D.(Ed.), Encyclopedia of Soils in the Environment, vol. 1. Elsevier, Oxford, U.K., pp.103–115.
Bashan, Y., de-Bashan, L.E., 2005b. Fresh-weight measurements of roots provideinaccurate estimates of the effects of plant growth-promoting bacteria on rootgrowth: a critical examination. Soil Biol. Biochem. 37, 1795–1804.
Bashan, Y., Dubrovsky, J.G., 1996. Azospirillum spp. participation in dry matter par-titioning in grasses at the whole plant level. Biol. Fertil. Soils 23, 435–440.
Bashan, Y., Hernandez, J.-P., Leyva, L.A., Bacilio, M., 2002. Alginate microbeads asinoculant carrier for plant growth-promoting bacteria. Biol. Fertil. Soils 35,359–368.
Bashan, Y., Holguin, G., 1998. Proposal for the division of plant growth-promoting rhizobacteria into two classifications: biocontrol-PGPB (plantgrowth-promoting bacteria) and PGPB. Soil Biol. Biochem. 30, 1225–1228.
Bashan, Y., Holguin, G., de-Bashan, L.E., 2004. Azospirillum-plant relationships:physiological, molecular, agricultural, and environmental advances (1997-2003). Can. J. Microbiol. 50, 521–577.
Bashan, Y., Levanony, H., 1991. Alterations in membrane potential and in protonefflux in plant roots induced by Azospirillum brasilense. Plant Soil 137, 99–103.
Bashan, Y., Moreno, M., Troyo, E., 2000. Growth promotion of the oilseed halophyteSalicornia bigelovii in seawater inoculated with mangrove rhizosphere bacteriaand Azospirillum. Biol. Fertil. Soils 32, 265–272.
Bashan, Y., Salazar, B., Puente, M.E., 2009a. Responses of native legume deserttrees used for reforestation in the Sonoran Desert to plant growth-promotingmicroorganisms in screen house. Biol. Fertil. Soils 45, 655–662.
Bashan, Y., Salazar, B., Puente, M.E., Bacilio, M., Linderman, R.G., 2009b. Enhancedestablishment and growth of giant cardon cactus in an eroded field in theSonoran Desert using native legume trees as nurse plants aided by plantgrowth-promoting microorganisms and compost. Biol. Fertil. Soils 45, 585–594.
Belimov, A.A., Kunakova, A.M., Safronova, V.I., Stepanok, V.V., Yudkin, L.V., Alekseev,Y.V., Kozhemyakov, A.P., 2004. Employment of rhizobacteria for the inocula-tion of barley plants cultivated in soil contaminated with lead and cadmium.Microbiology 73, 99–106.
Burd, G.I., Dixon, D.G., Glick, B.R., 1998. A plant growth-promoting bacterium thatdecreases nickel toxicity in seedlings. Appl. Environ. Microbiol. 64, 3663–3668.
Burd, G.I., Dixon, D.G., Glick, B.R., 2000. Plant growth-promoting bacteria thatdecrease heavy metal toxicity in plants. Can. J. Microbiol. 46, 237–245.
Carrillo, A.E., Li, C.Y., Bashan, Y., 2002. Increased acidification in the rhizosphereof cactus seedlings induced by Azospirillum brasilense. Naturwissenschaften 89,428–432.
Chiu, K.K., Ye, Z.H., Wong, M.H., 2006. Growth of Vetiveria zizanioides and Phragmitiesaustralis on Pb/Zn and Cu mine tailings amended with manure compost andsewage sludge: a greenhouse study. Bioresour. Technol. 97, 158–170.
Colores, G.M., Macur, R.E., Ward, D.M., Inskeep, W.P., 2000. Molecular analysisof surfactant-driven microbial population shifts in hydrocarbon-contaminatedsoil. Appl. Environ. Microbiol. 66, 2959–2964.
Daims, H., Stoecker, K., Wagner, M., 2005. Fluorescence in situ hybridization for thedetection of prokaryotes. In: Osborn, A.M., Smith, C.J. (Eds.), Molecular MicrobialEcology. Taylor and Francis Group, New York.
de Brito-Alvarez, M.A., Gagne, S., Antoun, H., 1995. Effect of compost on rhizospheremicroflora of the tomato and on the incidence of plant growth-promoting rhi-zobacteria. Appl. Environ. Microbiol. 61, 194–199.
EPA, 2004. Test Methods for Evaluating Solid Waste. EPA SW-846, US EnvironmentalProtection Agency. Washington, DC.
Enebak, S.A., Wei, G., Kloepper, J.W., 1998. Effects of plant growth-promoting rhi-zobacteria on loblolly and slash pine seedlings. For. Sci. 44, 139–144.
Felici, C., Vettori, L., Giraldi, E., Forino, L.M.C., Toffanon, A., Tagliasacchi, A.M., Nuti, M.,2008. Single and co-inoculation of Bacillus subtilis and Azospirillum brasilenseon Lycopersicon esculentum: Effects on plant growth and rhizosphere microbialcommunity. Appl. Soil Ecol. 40, 260–270.
Felske, A., Akkermans, A.D.L., de Vos, W.M., 1998. Quantification of 16S rRNA incomplex bacterial communities by multiple competitive reverse transcription-PCR in temperate gradient gel electrophoresis fingerprintings. Appl. Environ.Microbiol. 64, 4581–4587.
Ferris, M.J., Muyzer, G., Ward, D.M., 1996. Denaturing gradient gel electrophoresisprofiles of 16S rRNA-defined populations inhabiting a hot spring microbial matcommunity. Appl. Environ. Microbiol. 62, 340–346.
Glick, B.R., 2003. Phytoremediation: synergistic use of plants and bacteria to cleanup the environment. Biotechnol. Adv. 21, 383–393.
Gonzalez-Chavez, M.C., Carrillo-Gonzalez, R., Gutierrez-Castorena, M.C., 2009. Nat-ural attenuation in a slag heap contaminated with cadmium: The role of plantsand arbuscular mycorrhizal fungi. J. Hazard Mater. 161, 1288–1298.
Grandlic, C.J., Mendez, M.O., Chorover, J., Machado, B., Maier, R.M., 2008. Plantgrowth-promoting bacteria for phytostabilization of mine tailings. Environ. Sci.Technol. 42, 2079–2084.
Grandlic, C.J., Palmer, M.W., Maier, R.M., 2009. Optimization of plant growth-promoting bacteria-assisted phytostablization of mine tailings. Soil Biol.Biochem. 41, 1734–1740.
Hartmann, A., Rothballer, M., Schmid, M., 2008. Lorenz Hiltner, a pioneer in rhizo-sphere microbial ecology and soil bacteriology research. Plant Soil 312, 7–14.
Hernandez, J.-P., de-Bashan, L.E., Rodriguez, D.J., Rodriguez, Y., Bashan, Y., 2009.Growth promotion of the freshwater microalga Chlorella vulgaris by thenitrogen-fixing, plant growth-promoting bacterium Bacillus pumilus from aridzone soils. Eur. J. Soil Biol. 45, 88–93.
Herschkovitz, Y., Lerner, A., Davidov, Y., Okon, Y., Jurkevitch, E., 2005a. Azospiril-lum brasilense does not affect population structure of specific rhizobacterialcommunities of inoculated maize (Zea mays). Environ. Microbiol. 7, 1847–1852.
Herschkovitz, Y., Lerner, A., Davidov, Y., Rothballer, M., Hartmann, A., Okon, Y.,Jurkevitch, E., 2005b. Inoculation with the plant growth promoting rhizobac-terium Azospirillum brasilense causes little disturbance in the rhizosphere andrhizoplane of maize (Zea mays). Microbial Ecol 50, 277–288.
Iverson, S.L., Maier, R.M., 2009. Effects of compost on colonization of roots ofplants grown in metalliferous mine tailings, as examined by fluorescence insitu hybridization. Appl. Environ. Microbiol. 75, 842–847.
Jacoud, C., Faure, D., Wadoux, P., Bally, R., 1998. Development of a strain-specificprobe to follow inoculated Azospirillum lipoferum CRT1 under field conditionsand enhancement of maize root development by inoculation. FEMS Microbiol.Ecol. 27, 43–51.
Kirchhof, G., Schloter, M., Assmus, B., Hartmann, A., 1997. Molecular microbial ecol-ogy approaches applied to diazotrophs associated with non-legumes. Soil Biol.Biochem. 29, 853–862.
Kloepper, J.W., Reddy, M.S., Rodríguez-Kabana, R., Kenney, D.S., Kokalis-Burelle,N., Martinez-Ochoa, N., Vavrina, C.S., 2004a. Application for rhizobacteria intransplant production and yield enhancement. Acta Horticulturae 631, 217–229.
Kloepper, J.W., Ryu, C.-M., Zhang, S., 2004b. Induced systemic resistance and pro-motion of plant growth by Bacillus spp. Phytopathology 94, 1259–1266.
Lerner, A., Herschkovitz, Y., Baudoin, E., Nazaret, S., Moenne-Loccoz, Y., Okon, Y.,Jurkevitch, E., 2006. Effect of Azospirillum brasilense inoculation on rhizobac-terial communities analyzed by denaturing gradient gel electrophoresis andautomated ribosomal intergenic spacer analysis. Soil Biol. Biochem. 38, 1212–1218.
Li, W.C., Ye, Z.H., Wong, M.H., 2007. Effects of bacteria on enhanced metal uptake ofthe Cd/Zn-hyperaccumulating plant, Sedum alfredii. J. Exper. Bot. 58, 4173–4182.
Liu, W.-T., Mirzaberok, A.D., Stahl, D.A., 2001. Optimization of an oligonucleotidemicrochip for microbial identification studies: a non-equilibrium dissociationapproach. Environ. Microbiol. 3, 619–629.
Lugtenberg, B.J.J., Dekkers, L.C., 1999. What makes Pseudomonas bacteria rhizo-sphere competent? Environ. Microbiol. 1, 9–13.
McCutcheon, S.C., Schnoor, J.L., 2003. Phytoremediation: Transformation and Con-trol of Contaminants. Wiley-Interscience, Hoboken, NJ.
Mendez, M.O., Glenn, E.P., Maier, R.M., 2007. Phytostabilization potential of quail-bush for mine tailings: Growth, metal accumulation, and microbial communitychanges. J Environ. Qual. 36, 245–253.
Mendez, M.O., Maier, R.M., 2008. Phytostabilization of mine tailings in arid andsemiarid environments—an emerging remediation technology. Environ. HealthPerspect. 116, 278–283.
Meyer, J.R., Linderman, R.G., 1986. Selective influence on populations of rhizosphereor rhizoplane bacteria and actinomycetes by mycorrhizas formed by Glomusfasciculatum. Soil Biol. Biochem. 18, 191–196.
Mogge, B., Loferer, C., Agerer, R., Hutzler, P., Hartmann, A., 2000. Bacterial communitystructure and colonization patterns of Fagus sylvatica L. ectomycorrhizospheresas determined by fluorescence in situ hybridization and confocal laser scanningmicroscopy. Mycorrhiza 9, 271–278.
Natarajan, K.A., 1998. An integrated biotechnological approach to gold processing—an Indian experience. Miner. Process. Extr. Metall. Rev. 19, 235–251.
Petrisor, I., Dobrota, S., Komnitsas, K., Lazar, I., Kuperberg, J.M., Serban, M., 2004. Arti-ficial inoculation—perspectives in tailings phytostabilization. Int. J. Phytorem. 6,1–15.
Pilon-Smits, E., 2005. Phytoremediation. Annu. Rev. Plant Biol. 56, 15–39.Pishchik, V.N., Vorobyev, N.I., Chernyaeva, I.I., Timofeeva, S.V., Kozhemyakov, A.P.,
Alexeev, Y.V., Lukin, S.M., 2002. Experimental and mathematical simulation ofplant growth promoting rhizobacteria and plant interaction under cadmiumstress. Plant Soil. 243, 173–186.
Puente, M.E., Bashan, Y., Li, C.Y., Lebsky, V.K., 2004. Microbial populations and activ-ities in the rhizoplane of rock-weathering desert plants. I. Root colonization andweathering of igneous rocks. Plant Biol. 6, 629–642.
Ramos, H.J.O., Roncato-Maccari, L.D.B., Souza, E.M., Soares-Ramos, J.R.L., Hungria, M.,Pedrosa, F.O., 2002. Monitoring Azospirillum-wheat interactions using the gfpand gusA genes constitutively expressed from a new broad-host range vector. J.Biotechnol. 97, 243–252.
Reed, M.L.E., Glick, B.R., 2005. Growth of canola (Brassica napus) in the presenceof plant growth-promoting bacteria and either copper or polycyclic aromatichydrocarbons. Can. J. Microbiol. 51, 1061–1069.
Reed, M.L.E., Warner, B.G., Glick, B.R., 2005. Plant growth-promoting bacteria facil-itate the growth of the common reed Phragmites australis in the presence ofcopper or polycyclic aromatic hydrocarbons. Curr. Microbiol. 51, 425–429.
Rajkumar, M., Freitas, H., 2008a. Effects of inoculation of plant growth-promotingbacteria on Ni uptake by Indian mustard. Bioresour. Technol. 99, 3491–3498.
Rajkumar, M., Freitas, H., 2008b. Influence of metal-resistant plant growth-promoting bacteria on the growth of Ricinus communis in soil contaminatedwith heavy metals. Chemosphere 71, 834–842.
Rosario, K., Iverson, S.L., Henderson, D.A., Chartrand, S., McKeon, C., Glenn, E.P., Maier,R.M., 2007. Bacterial community changes during plant establishment at the SanPedro River mine tailings site. J. Environ. Qual. 3, 1249–1259.
Ryu, C.-M., Hu, C.-H., Locy, R.D., Kloepper, J.W., 2005. Study of mechanisms for plantgrowth promotion elicited by rhizobacteria in Arabidopsis thaliana. Plant Soil268, 285–292.

Author's personal copy
352 L.E. de-Bashan et al. / Environmental and Experimental Botany 69 (2010) 343–352
Laboratory Methods Manual, 2004. In: Burt, R. (Ed.), Soil Survey Laboratory Inves-tigations Report No. 42, Version 4.0. National Resources Conservation Service,Lincoln, NE.
Tsuruta, T., 2007. Removal and recovery of uranium using microorganisms iso-lated from North American uranium deposits. Am. J. Environ. Sci. 3, 60–66.
Venables, W.N., Ripley, B.D., 2002. Modern Applied Statistics with S, 4th ed. Springer,New York, pp. 385–389.
Vessey, J.K., 2003. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil.255, 571–586.
Vijayalakshmi, S.P., Raichur, A.M., 2003. The utility of Bacillus subtilis as a biofloccu-lant for fine coal. Colloids Surf. B: Biointerfaces 29, 265–275.
Vivas, A., Biro, B., Ruiz-Lozano, J.M., Barea, J.M., Azcon, R., 2006. Two bacterial strainsisolated from a Zn-polluted soil enhance plant growth and mycorrhizal effi-ciency under Zn-toxicity. Chemosphere 62, 1523–1533.
Wu, S.C., Cheung, K.C., Luo, Y.M., Wong, H.M., 2006a. Effects of inoculation of plantgrowth-promoting rhizobacteria on metal uptake by Brassica juncea. Environ.Pollut. 140, 124–135.
Wu, S.C., Luo, Y.M., Cheung, K.C., Wong, M.H., 2006b. Influence of bacteria on Pb andZn speciation, mobility and bioavailability in soil: a laboratory study. Environ.Pollut. 144, 765–773.
Ye, Z.H., Yang, Z.Y., Chan, G.Y.S., Wong, M.H., 2001. Growth response of Sesbania ros-trata and S. cannabina to sludge-amended lead/zinc mine tailings. A greenhousestudy. Environ. Int. 26, 449–455.
Zhang, H.-B., Yang, M.-X., Shi, W., Zheng, Y., Sha, T., Zhao, Z.-W., 2007. Bacterialdiversity in mine tailings compared by cultivation and cultivation-independentmethods and their resistance to lead and cadmium. Microb. Ecol. 54, 705–712.
Zhuang, X., Chen, J., Shim, H., Bai, Z., 2007. New advances in plant growth-promotingrhizobacteria for bioremediation. Environ. Int. 33, 406–413.





























