Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 263, No. 19, Issue of July 5 , pp. 9395-9401,1988 Prmted u1 U.S.A.
Atrial Natriuretic Peptide Clearance Receptor COMPLETE SEQUENCE AND FUNCTIONAL EXPRESSION OF cDNA CLONES*
(Received for publication, November 12, 1987)
Forrest Fuller$, J. Gordon Porter, Ann E. Arfsten, Judy Miller, James W. Schilling, Robert M. Scarborough, John A. Lewicki, and Dale B. Schenk From California Biotechnology, Inc., Mountain View, California 94043
The major class of atrial natriuretic peptide (ANP) receptors was isolated from cultured vascular smooth muscle cells, and a partial amino acid sequence was obtained. This allowed the isolation of cDNA clones from which the entire amino acid sequence was estab- lished. The smooth muscle cell ANP receptor appears to be synthesized as a 537-amino acid precursor with an N-terminal membrane translocation signal. The ma- ture form consists of 496 amino acids with a single potential transmembrane domain predicting a 37- amino acid cytoplasmic domain and a large, acidic, extracellular domain low in cysteine and probably con- taining attached carbohydrate. The receptor is there- fore similar in structure to the growth factor receptors but notably lacks repetitive cysteine-rich domains and has a relatively small intracellular domain. Expression of the cloned receptor in Xenopus oocytes elicited high affinity, membrane-associated binding sites for ANP and for truncated and internally deleted analogs of ANP. These results reflect the ligand binding specific- ity found for the major class of ANP receptors on smooth muscle cells and thus provide additional evi- dence that two distinct ANP receptors exist since ANP receptor-coupled guanylate cyclase activity exhibits a very different ANP analog specificity.
Atrial natriuretic peptide (ANP),’ a hormone which is synthesized and secreted by the atria of the heart, evokes a variety of physiological responses affecting cardiovascular homeostasis. These effects which collectively lead to reduction in blood volume include diuresis and natriuresis (1-3), inhi- bition of aldosterone secretion (4, 51, and vasorelaxation (6). Such diverse effects are reflected in the wide variety of cell types and target tissues on which specific binding sites for ANP have been identified (7-17) and in the varied responses of these cells and tissues to ANP.
Diversity of function is also indicated by the observed heterogeneity in ANP-binding sites. Cross-linking studies
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in thispaper has been submitted
503876. to the GenBankTM/EMBL Data Bank with accession number(s)
$ TO whom correspondence should be addressed California Bio- technology, Inc., 2450 Bayshore Pkwy., Mountain View, CA 94043.
The abbreviations used are: ANP, atrial natriuretic peptide; ANP C-receptor, ANP clearance receptor; rANP, rat ANP; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; BASM, bovine aortic smooth muscle; PTH, phenylthiohydantoin; HPLC, high performance liquid chromatography; CI2E~, octaethylene glycol dodecyl ether; kb, kilobase.
have identified at least two ANP-binding proteins with M, -130,000 and -60,000 as determined by SDS-PAGE analysis under reducing conditions (11, 15-28). Furthermore, a func- tional distinction between receptor types is suggested by pharmacological studies. Whereas an important receptor-me- diated effect of ANP is the stimulation of particulate guanyl- ate cyclase activity (29,30), several truncated analogs of ANP which neither stimulate guanylate cyclase nor antagonize the ANP-mediated stimulation of guanylate cyclase are neverthe- less able to bind with high affinity to the major class of receptors observed on vascular smooth muscle cells (29-33). Cross-linking studies in vascular endothelial cells suggest that such ANP analogs bind specifically to the -60-kDa subunit (30),’ whereas only analogs which activate guanylate cyclase appear to be able to bind to the -130-kDa subunit (11,16,17, 28). Takayanagi et al. (34) have recently purified two struc- turally dissimilar (35) ANP binding activities from bovine adrenal membranes to near homogeneity and have demon- strated that guanylate cyclase activity is exclusively associ- ated with an -135-kDa polypeptide, whereas truncated ANP analog binding is exclusively associated with an -62-kDa polypeptide.
These observations clearly indicate that functionally and structurally distinct ANP receptors exist. However, the func- tion and physiological role of the receptor with the -60-kDa binding subunit have not been elucidated. Recently, Maack et al. (36) have proposed a novel function for this receptor based on the observation that in vivo administration of an internally deleted ANP analog results in an increase in cir- culating ANP levels. They suggest that the primary function of this receptor is sequestration and metabolic clearance of ANP and have therefore designated it as the C-receptor. As an initial step toward further defining receptor-mediated functions of ANP and determining the molecular structures of the different ANP receptors, the major ANP receptor from cultured bovine aortic smooth muscle (BASM) cells was pu- rified to apparent homogeneity with functional binding activ- ity (37). We now report the direct determination of the N- terminal amino acid sequence of this protein and the isolation of corresponding cDNA clones. That the clones encode the ANP C-receptor found on BASM cells is demonstrated by the findings that the molecular weight and structural character- istics of the encoded polypeptide, the cell and tissue sources of homologous mRNAs, and the ANP binding imparted on membranes from Xenopus oocytes injected with the cloned sequence all reflect the known properties of the ANP C - receptor.
F. Fuller, J. G . Porter, A. E. Arfsten, J. Miller, J. W. Schilling, R. M. Scarborough, J. A. Lewicki, and D. B. Schenk, unpublished observations.
9395
9396 ANP C-receptor cDNA Sequence and Expression
Purified Receptor Mr x
Jll. 200 .. I16 - 92
.- 68
- 45
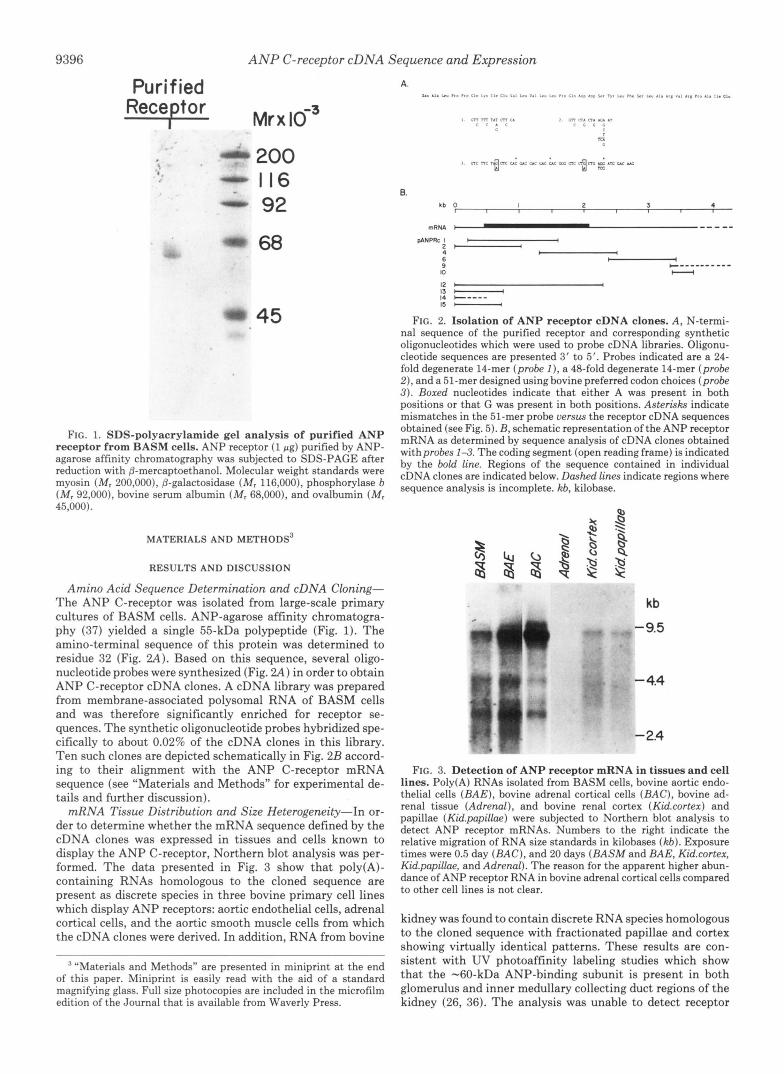
FIG. 1. SDS-polyacrylamide gel analysis of purified ANP receptor from BASM cells. ANP receptor (1 pg) purified by ANP- agarose affinity chromatography was subjected to SDS-PAGE after reduction with ,!?-mercaptoethanoL Molecular weight standards were myosin ( M , 200,000), 0-galactosidase ( M , 116,000), phosphorylase b ( M , 92,000), bovine serum albumin ( M y 68,000), and ovalbumin ( M , 45,000).
MATERIALS AND METHODS3
RESULTS AND DISCUSSION
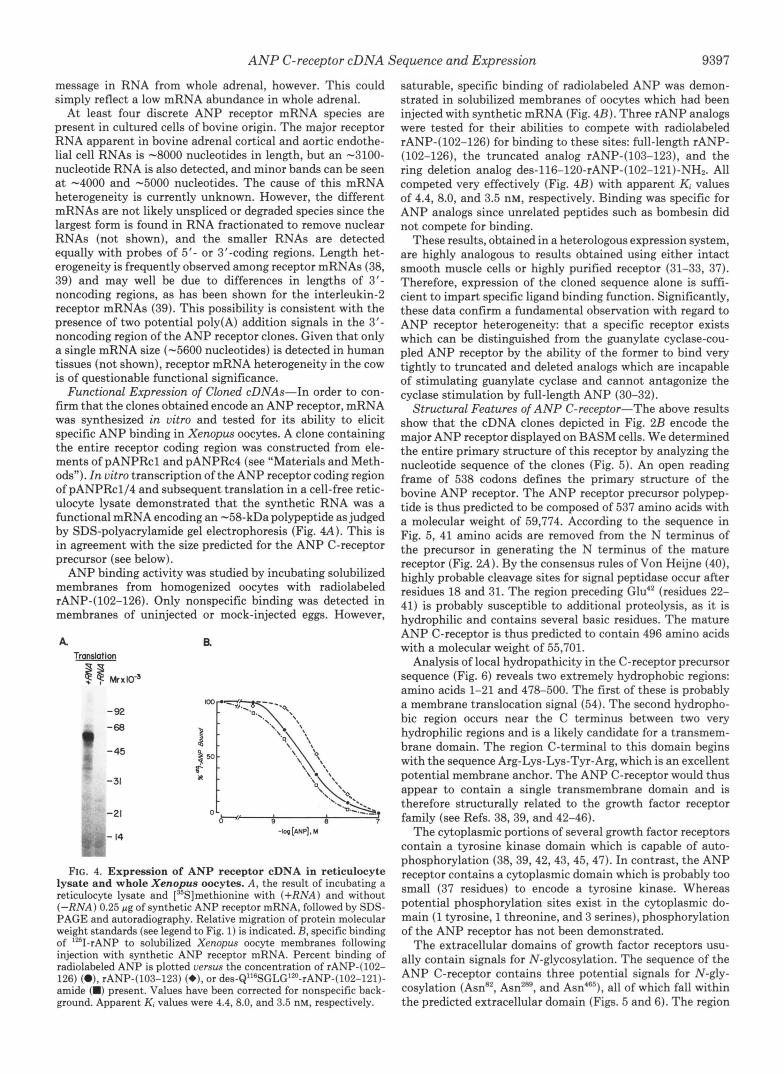
Amino Acid Sequence Determination and cDNA Cloning- The ANP C-receptor was isolated from large-scale primary cultures of BASM cells. ANP-agarose affinity chromatogra- phy ( 3 7 ) yielded a single 55-kDa polypeptide (Fig. 1). The amino-terminal sequence of this protein was determined to residue 32 (Fig. 2 A ) . Based on this sequence, several oligo- nucleotide probes were synthesized (Fig. 2 A ) in order to obtain ANP C-receptor cDNA clones. A cDNA library was prepared from membrane-associated polysomal RNA of BASM cells and was therefore significantly enriched for receptor se- quences. The synthetic oligonucleotide probes hybridized spe- cifically to about 0.02% of the cDNA clones in this library. Ten such clones are depicted schematically in Fig. 2B accord- ing to their alignment with the ANP C-receptor mRNA sequence (see "Materials and Methods" for experimental de- tails and further discussion).
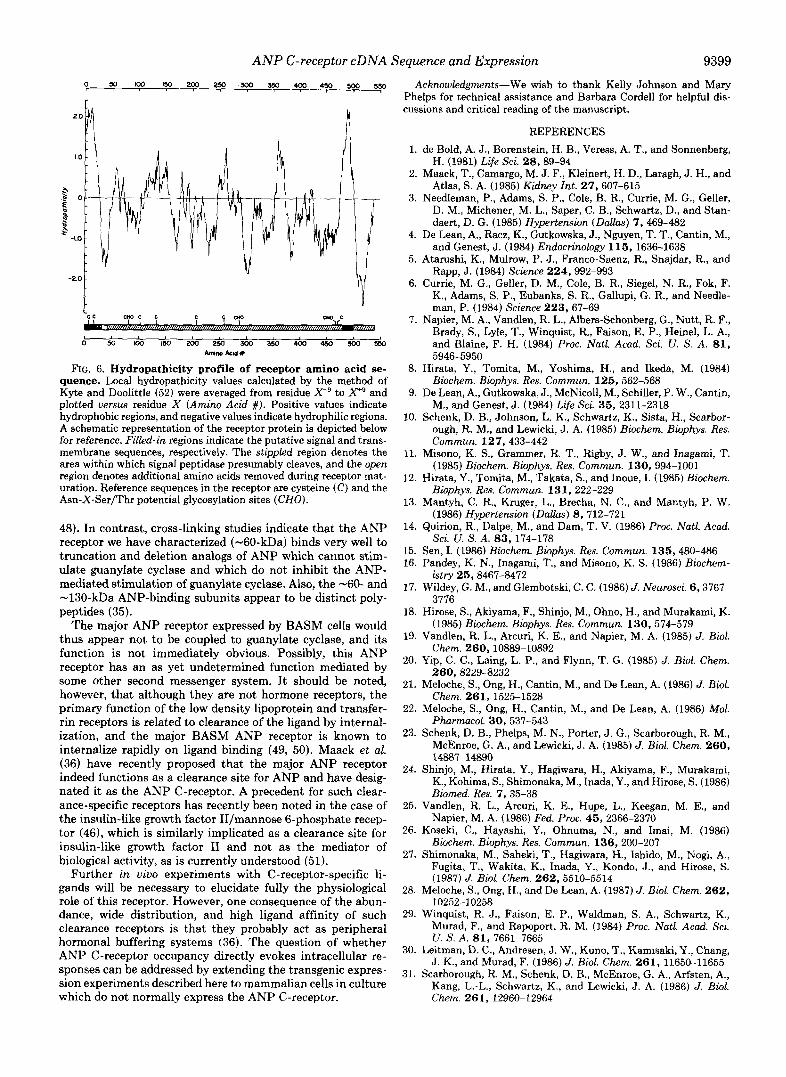
mRNA Tissue Distribution and Size Heterogeneity-In or- der to determine whether the mRNA sequence defined by the cDNA clones was expressed in tissues and cells known to display the ANP C-receptor, Northern blot analysis was per- formed. The data presented in Fig. 3 show that poly(A)- containing RNAs homologous to the cloned sequence are present as discrete species in three bovine primary cell lines which display ANP receptors: aortic endothelial cells, adrenal cortical cells, and the aortic smooth muscle cells from which the cDNA clones were derived. In addition, RNA from bovine
' "Materials and Methods" are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press.
A. I" .I. Lu PC* F ro i t " L,. I,. C I " "1, Y" "./ L." 1." .rO //I A.0 .,I *, I,. Ll" m. *, 8," AI. j..: ",, ,,-A,. I,. C,"
I L7l m 7.7 m c. ? Gn n. 0 1 u "7 r r . r < < < I .
m
1 ~~,,~,;,,,;,,~,,~.~,,
6. kb 0 I 2 3 4
I
mRNA "_" pANPRc I b I
2-
6 - 9 lo - 12 I I 13 - 14 C---- IS -
b 4
+ """"_ -
FIG. 2. Isolation of ANP receptor cDNA clones. A , N-termi- nal sequence of the purified receptor and corresponding synthetic oligonucleotides which were used to probe cDNA libraries. Oligonu- cleotide sequences are presented 3' to 5'. Probes indicated are a 24- fold degenerate 14-mer (probe I), a 48-fold degenerate 14-mer (probe 2) , and a 51-mer designed using bovine preferred codon choices (probe 3 ) . Boxed nucleotides indicate that either A was present in both positions or that G was present in both positions. Asterisks indicate mismatches in the 51-mer probe versus the receptor cDNA sequences obtained (see Fig. 5). B, schematic representation ofthe ANP receptor mRNA as determined by sequence analysis of cDNA clones obtained withprobes 1-3. The coding segment (open reading frame) is indicated by the bold line. Regions of the sequence contained in individual cDNA clones are indicated below. Dashed lines indicate regions where sequence analysis is incomplete. kb, kilobase.
FIG. 3. Det .ection of ANP re
kb
-9.5
- 4.4
- 2.4
ceptor mRNA in tissues and cell lines. Poly(A) RNAs isolated from BASM cells, bovine aortic endo- thelial cells ( B A E ) , bovine adrenal cortical cells (BAC) , bovine ad- renal tissue (Adrenal), and bovine renal cortex (Kidxortex) and papillae (Kid.papilhe) were subjected to Northern blot analysis to detect ANP receptor mRNAs. Numbers to the right indicate the relative migration of RNA size standards in kilobases ( k b ) . Exposure times were 0.5 day ( B A C ) , and 20 days (BASM and BAE, Kidxortex, Kid.papilhe, and Adrenal). The reason for the apparent higher abun- dance of ANP receptor RNA in bovine adrenal cortical cells compared to other cell lines is not clear.
kidney was found to contain discrete RNA species homologous to the cloned sequence with fractionated papillae and cortex showing virtually identical patterns. These results are con- sistent with UV photoaffinity labeling studies which show that the -60-kDa ANP-binding subunit is present in both glomerulus and inner medullary collecting duct regions of the kidney (26, 36). The analysis was unable to detect receptor
ANP C-receptor cDNA Sequence and Expression 9397
message in RNA from whole adrenal, however. This could simply reflect a low mRNA abundance in whole adrenal.
A t least four discrete ANP receptor mRNA species are present in cultured cells of bovine origin. The major receptor RNA apparent in bovine adrenal cortical and aortic endothe- lial cell RNAs is -8000 nucleotides in length, but an -3100- nucleotide RNA is also detected, and minor bands can be seen a t -4000 and -5000 nucleotides. The cause of this mRNA heterogeneity is currently unknown. However, the different mRNAs are not likely unspliced or degraded species since the largest form is found in RNA fractionated to remove nuclear RNAs (not shown), and the smaller RNAs are detected equally with probes of 5'- or 3"coding regions. Length het- erogeneity is frequently observed among receptor mRNAs (38, 39) and may well be due to differences in lengths of 3'- noncoding regions, as has been shown for the interleukin-2 receptor mRNAs (39). This possibility is consistent with the presence of two potential poly(A) addition signals in the 3'- noncoding region of the ANP receptor clones. Given that only a single mRNA size (-5600 nucleotides) is detected in human tissues (not shown), receptor mRNA heterogeneity in the cow is of questionable functional significance.
Functional Expression of Cloned cDNAs-In order to con- firm that the clones obtained encode an ANP receptor, mRNA was synthesized in vitro and tested for its ability to elicit specific ANP binding in Xenopus oocytes. A clone containing the entire receptor coding region was constructed from ele- ments of pANPRcl and pANPRc4 (see "Materials and Meth- ods"). In vitro transcription of the ANP receptor coding region of pANPRcl/4 and subsequent translation in a cell-free retic- ulocyte lysate demonstrated that the synthetic RNA was a functional mRNA encoding an -58-kDa polypeptide as judged by SDS-polyacrylamide gel electrophoresis (Fig. 4A). This is in agreement with the size predicted for the ANP C-receptor precursor (see below).
ANP binding activity was studied by incubating solubilized membranes from homogenized oocytes with radiolabeled rANP-(102-126). Only nonspecific binding was detected in membranes of uninjected or mock-injected eggs. However,
A R Tmlotion
3 M r x l O "
-92 -68
-45 v
-31
-21
FIG. 4. Expression of ANP receptor cDNA in reticulocyte lysate and whole Xenopus oocytes. A , the result of incubating a reticulocyte lysate and [35S]methionine with (+RNA) and without ( - R N A ) 0.25 pg of synthetic ANP receptor mRNA, followed by SDS- PAGE and autoradiography. Relative migration of protein molecular weight standards (see legend to Fig. 1) is indicated. B, specific binding of '*'I-rANP to solubilized Xenopus oocyte membranes following injection with synthetic ANP receptor mRNA. Percent binding of radiolabeled ANP is plotted versus the concentration of rANP-(102- 126) (O), rANP-(103-123) (+), or de~-Q"~SGLG'*~-rANP-(102-121)- amide (W) present. Values have been corrected for nonspecific back- ground. Apparent Ki values were 4.4, 8.0, and 3.5 nM, respectively.
saturable, specific binding of radiolabeled ANP was demon- strated in solubilized membranes of oocytes which had been injected with synthetic mRNA (Fig. 4B). Three rANP analogs were tested for their abilities to compete with radiolabeled rANP-( 102-126) for binding to these sites: full-length rANP- (102-126), the truncated analog rANP-(103-123), and the ring deletion analog des-116-120-rANP-(102-121)-NH2. All competed very effectively (Fig. 4B) with apparent Ki values of 4.4, 8.0, and 3.5 nM, respectively. Binding was specific for ANP analogs since unrelated peptides such as bombesin did not compete for binding.
These results, obtained in a heterologous expression system, are highly analogous to results obtained using either intact smooth muscle cells or highly purified receptor (31-33, 37). Therefore, expression of the cloned sequence alone is suffi- cient to impart specific ligand binding function. Significantly, these data confirm a fundamental observation with regard to ANP receptor heterogeneity: that a specific receptor exists which can be distinguished from the guanylate cyclase-cou- pled ANP receptor by the ability of the former to bind very tightly to truncated and deleted analogs which are incapable of stimulating guanylate cyclase and cannot antagonize the cyclase stimulation by full-length ANP (30-32).
Structural Features of ANP C-receptor-The above results show that the cDNA clones depicted in Fig. 2B encode the major ANP receptor displayed on BASM cells. We determined the entire primary structure of this receptor by analyzing the nucleotide sequence of the clones (Fig. 5). An open reading frame of 538 codons defines the primary structure of the bovine ANP receptor. The ANP receptor precursor polypep- tide is thus predicted to be composed of 537 amino acids with a molecular weight of 59,774. According to the sequence in Fig. 5, 41 amino acids are removed from the N terminus of the precursor in generating the N terminus of the mature receptor (Fig. 2A). By the consensus rules of Von Heijne (40), highly probable cleavage sites for signal peptidase occur after residues 18 and 31. The region preceding Glu4' (residues 22- 41) is probably susceptible to additional proteolysis, as it is hydrophilic and contains several basic residues. The mature ANP C-receptor is thus predicted to contain 496 amino acids with a molecular weight of 55,701.
Analysis of local hydropathicity in the C-receptor precursor sequence (Fig. 6) reveals two extremely hydrophobic regions: amino acids 1-21 and 478-500. The first of these is probably a membrane translocation signal (54). The second hydropho- bic region occurs near the C terminus between two very hydrophilic regions and is a likely candidate for a transmem- brane domain. The region C-terminal to this domain begins with the sequence Arg-Lys-Lys-Tyr-Arg, which is an excellent potential membrane anchor. The ANP C-receptor would thus appear to contain a single transmembrane domain and is therefore structurally related to the growth factor receptor family (see Refs. 38,39, and 42-46).
The cytoplasmic portions of several growth factor receptors contain a tyrosine kinase domain which is capable of auto- phosphorylation (38,39,42,43,45,47). In contrast, the ANP receptor contains a cytoplasmic domain which is probably too small (37 residues) to encode a tyrosine kinase. Whereas potential phosphorylation sites exist in the cytoplasmic do- main (1 tyrosine, 1 threonine, and 3 serines), phosphorylation of the ANP receptor has not been demonstrated.
The extracellular domains of growth factor receptors usu- ally contain signals for N-glycosylation. The sequence of the ANP C-receptor contains three potential signals for N-gly- cosylation (AmR2, AsnZR9, and AS^^^'), all of which fall within the predicted extracellular domain (Figs. 5 and 6). The region

9398 A N P C-receptor cDNA Sequence and Expression 150
300
450
5 1 6
6 9 )
1 2 2
9 4 5
1 0 6 1
1191
1114
1417
1560
1 6 a 1
1106
1 9 1 9
2 0 5 2
ATC ACA TCC CAC TTT TCG CTC GCT TAA MCCAACTCTC TTCTTTTCCC TTGAGATTCI TTAACGAGAT ACArOCGATC AAAGACATCA ATCCAATACA AGGGQCCCTC Tac-cT CATTCTTTTA AGCACTTAGT AATTTTC 2191 11. A r g 5.r "1s Ph. S.r V a l Ala End
5 1 7
I T A TAAAATTTCT TTAGAAGCIC AGGAACTATI AlYAATCACC ATAT'OCCCGC TCCCCTCTCA TCTCAICACA AACAAACACA ICAATATAAC ATCACTCCTA AATCITGATT CTGTTTCAAG aCCATATCAT TAGATTTATG T T C T G M 1 1 4 7
ACT CTCAGCTCTC CATATCTTGT TCCCCTTTGG CCCCATTTTA CACAAGCCTA TAAAATCTGT TTTACIIAAC TGAGATGTTT T A T A O C T M ~ T C A T TTTTACAICT AGGATATTAT T C I I U G G A T TTAACCCCAA OAAGAAC 2 4 9 7
AAA A'XTAAlGGA A M C C T C M G G r I C A M A T O C A G E A m C ? CTCTCTACAG CIPCTTOGAG G G A T C I C I I G TCAACCCCCT TCTATCTGAT ATATGGCAITC ACATCC1OIC TYrArGTTTC -GUT- TCCCCCACCT CTTTCAC 2647
ICT CTTGTAAGAG CTACTTTGGA AAGTTGTAAT TATCAATCAO ATAACACCAT T T A T C C A G M AAACCAAATC TAACTATTTC ACTTTTTMA ATATAAAAAA CCCTATTTCA CACTAACATT T T A T T T T T M OTATTTTAAT CTTATAT 1797
TTT CCTATTAGAA AATGTGTCTA TTTTTTCAYT TICAAGATTA AATTTCACTT ATATTTTAAA AACATGGGTA ATGTCTACAG CAAACCCAAT AATCATCAM GCA?CCCCTC TCZTTTTITC TCCCTCIITC CCTCTTCCCT GTCCCCA 2 9 4 7
TIC CCCAATACGA ATTaCTCC7.T CAACTACACA CATCTACAAA T C I C I T C C G A TTCIACACTC TACACCAATA CATCACTTTA CTTOTTTTAA ATGCAAGCYA TTTTAGCAIA GTCYCCTTCC A ~ T T C T C G C C AAACCATGAA AT=&?= IO97
A C I ATTAAGTCAC C T T T T A T A M CCGAGGCAAC CTTTTTTTCI CMCAACAAC TTTATAGACA GTTACAAETI GGCACTACCC ATACAAATCA TAATTT=TGACATTT TACCAAAATT CACCATTITG ATrTTaGTTA -GAG0 3 2 4 7
OM ICTIUCACC TACTAICTTC CCTTTTACTA AACACCCCIC AACTTTGCAC TCATCCACTA ATAACTTCTC r m c A G G a o z rGccTAccaA TATCACGAAT TCGTCAACCT rr.UT*.LrTA TTTCCCCAAT CTCAACTTTG CICCAAC 1 1 9 7
TAC TCTGTGTATA GGTACCTTTA ACCTTACGTT GCCACTTTCA TOTATAAT'IT BTCCACAAAA CAC3ACACICA GCGAAGAATC OGAAGTTCCC ICATCGGTCT ACTATCTTAI AATCATATCA TCAA(IGTaCT TCCICAATAA 7 C I I I C B 3541
ACC ATCTCaAA 15511
FIG. 5. ANP receptor cDNA sequence and predicted amino acid sequence. Numbers to the right indicate the nucleotide position in the sequence. Numbers below refer to the amino acid sequence of the preproreceptor predicted by the uninterrupted reading frame. Potential signal peptidase cleavage sites (arrowhends) and the beginning of the mature receptor N terminus (solid triangle) are shown. Potential N-linked glycosylation sites are boxed, and the putative transmembrane domain is denoted by the bur. Potential poly(A) addition signals in the 3'- noncoding region are overlined.
preceding the predicted transmembrane domain is also rich in serine and threonine and could contain 0-linked carbohy- drate. The major class of ANP receptor on smooth muscle cells is composed of subunits with M, -60,500-66,000 (23-25, 27, 30, 37). Since this is 5,000-10,000 daltons larger than predicted for the mature ANP C-receptor, it seems likely that carbohydrate is in fact added at some of these sites.
The amino acid composition of the purified ANP C-receptor indicates that it contains 4.6 cysteine residues/500 amino acids, which is in good agreement with the 5 cysteines pre- dicted by the cDNA sequence to be present in the mature receptor. A common feature of the extracellular domains of many growth factor receptors is that they contain repeated substructures rich in cysteine (38, 42, 43). The functional significance of the lack of cysteine clusters in the ANP recep- tor is unclear; however, the interleukin-2 (39) and transferrin (47) receptors similarly lack this feature. The ANP C-receptor is known to be a disulfide-linked homodimer (37). In this context, the odd number of cysteines and the presence of Cys"' so close to the transmembrane domain are worth not- ing.
Physiological Relevance of ANP C-receptor-The conclusion that the cDNA sequence which we have reported encodes the major subclass of ANP receptors (C-receptor) found on bovine smooth muscle cells is supported by several observations. The molecular weight of the encoded polypeptide, its amino acid composition and charge, and its structural characteristics are consistent with those of the major ANP receptor isolated by ANP affinity chromatography (27,33). Also, mRNA encoding this protein is present in cells which express the ANP C- receptor, and expression of this sequence alone is sufficient to elicit the appearance of specific ANP-binding sites on membranes of Xenopus oocytes. These sites specifically com- pete for binding of truncation and deletion analogs of ANP, which is the major characteristic of the ANP C-receptor (36).
At least two pharmacologically distinct populations of ANP receptors exist (31, 32, 34). Increasing evidence suggests that the receptor which is coupled to guanylate cyclase contains a binding subunit which is -130 kDa under reducing conditions. This polypeptide may in fact exhibit both high affinity ANP binding as well as intrinsic guanylate cyclase activity, as suggested by the observations of other investigators (34, 35,
n
‘cc cm c c c m c
0 x ) m l ~ m z w ~ 3 s o 4 o o w 5 o o ~ h i m Acld #
FIG. 6. Hydropathicity profile of receptor amino acid se- quence. Local hydropathicity values calculated by the method of Kyte and Doolittle (52) were averaged from residue X-’ to X+’ and plotted versus residue X (Amino Acid #). Positive values indicate hydrophobic regions, and negative values indicate hydrophilic regions. A schematic representation of the receptor protein is depicted below for reference. Filled-in regions indicate the putative signal and trans- membrane sequences, respectively. The stippled region denotes the area within which signal peptidase presumably cleaves, and the open region denotes additional amino acids removed during receptor mat- uration. Reference sequences in the receptor are cysteine (C) and the Asn-X-Ser/Thr potential glycosylation sites (CHO).
48). In contrast, cross-linking studies indicate that the ANP receptor we have characterized (-60-kDa) binds very well to truncation and deletion analogs of ANP which cannot stim- ulate guanylate cyclase and which do not inhibit the ANP- mediated stimulation of guanylate cyclase. Also, the -60- and -130-kDa ANP-binding subunits appear to be distinct poly- peptides (35).
The major ANP receptor expressed by BASM cells would thus appear not to be coupled to guanylate cyclase, and its function is not immediately obvious. Possibly, this ANP receptor has an as yet undetermined function mediated by some other second messenger system. It should be noted, however, that although they are not hormone receptors, the primary function of the low density lipoprotein and transfer- rin receptors is related to clearance of the ligand by internal- ization, and the major BASM ANP receptor is known to internalize rapidly on ligand binding (49, 50). Maack et al. (36) have recently proposed that the major ANP receptor indeed functions as a clearance site for ANP and have desig- nated it as the ANP C-receptor. A precedent for such clear- ance-specific receptors has recently been noted in the case of the insulin-like growth factor II/mannose 6-phosphate recep- tor (461, which is similarly implicated as a clearance site for insulin-like growth factor XI and not as the mediator of biological activity, as is currently understood (51).
Further in vivo experiments with C-receptor-specific li- gands will be necessary to elucidate fully the physiological role of this receptor. However, one consequence of the abun- dance, wide distribution, and high ligand affinity of such clearance receptors is that they probably act as peripheral hormonal buffering systems (36). The question of whether ANP C-receptor occupancy directly evokes intracellular re- sponses can be addressed by extending the transgenic expres- sion experiments described here to mammalian cells in culture which do not normally express the ANP C-receptor.
Sequence and Expression 9399
Acknowledgments-We wish to thank Kelly Johnson and Mary Phelps for technical assistance and Barbara Cordell for helpful dis- cussions and critical reading of the manuscript.
REFERENCES 1. de Bold, A. J., Borenstein, H. B., Veress, A. T., and Sonnenberg,
H. (1981) Life Sci. 28,89-94 2. Maack, T., Camargo, M. J. F., Kleinert, H. D., Laragh, J. H., and
Atlas, S. A. (1985) Kidney Int. 27,607-615 3. Needleman, P., Adams, S. P., Cole, B. R., Currie, M. G., Geller,
D. M., Michener, M. L., Saper, C. B., Schwartz, D., and Stan- daert, D. G. (1985) Hypertension (Dallas) 7, 469-482
4. De Lean, A., Racz, K., Gutkowska, J., Nguyen, T. T., Cantin, M., and Genest, J. (1984) Endocrinology 115, 1636-1638
5. Atarushi, K., Mulrow, P. J., Franco-Saenz, R., Snajdar, R., and Rapp, J. (1984) Science 224,992-993
6. Currie, M. G., Geller, D. M., Cole, B. R., Siegel, N. R., Fok, F. K., Adams, S. P., Eubanks, S. R., Gallupi, G. R., and Needle- man, P. (1984) Science 223, 67-69
7. Napier, M. A., Vandlen, R. L., Albers-Schonberg, G., Nutt, R. I?., Brady, S., Lyle, T., Winquist, R., Faison, E. P., Heinel, L. A., and Blaine, F. H. (1984) Proc. Natl. Acad. Sci. U. S. A. 81,
8. Hirata, Y., Tomita, M., Yoshima, H., and Ikeda, M. (1984) Biochem. Biophys. Res. Commun. 125,562-568
9. De Lean, A., Gutkowska, J., McNicoll, M., Schiller, P. W., Cantin, M., and Genest, J. (1984) Life Sci. 35, 2311-2318
10. Schenk, D. B., Johnson, L. K., Schwartz, K., Sista, H., Scarbor- ough, R. M., and Lewicki, J. A. (1985) Biochem. Biophys. Res. Commun. 127,433-442
11. Misono, K. S., Grammer, R. T., Rigby, J. W., and Inagami, T. (1985) Biochem. Biophys. Res. Commun. 130, 994-1001
12. Hirata, Y., Tomita, M., Takata, S., and Inoue, I. (1985) Biochem. Biophys. Res. Commun. 131,222-229
13. Mantyh, C. R., Kruger, L., Brecha, N. C., and Mantyh, P. W. (1986) Hypertension (Dallas) 8 , 712-721
14. Quirion, R., Dalpe, M., and Dam, T. V. (1986) Proc. Natl. Acad. Sci. U. S. A. 8 3 , 174-178
15. Sen, I. (1986) Biochem. Biophys. Res. Commun. 135,480-486 16. Pandey, K. N., Inagami, T., and Misono, K. S. (1986) Biochem-
17. Wildey, G. M., and Glembotski, C. C. (1986) J. Neurosci. 6,3767- 3776
18. Hirose, S., Akiyama, F., Shinjo, M., Ohno, H., and Murakami, K. (1985) Biochem. Biophys. Res. Commun. 130,574-579
19. Vandlen, R. L., Arcuri, K. E., and Napier, M. A. (1985) J . Biol. Chem. 260,10889-10892
20. Yip, C. C., Laing, L. P., and Flynn, T. G. (1985) J. Biot. Chem.
21. Meloche, S., Ong, H., Cantin, M., and De Lean, A. (1986) J. Biol. Chem. 261,1525-1528
22. Meloche, S., Ong, H., Cantin, M., and De Lean, A. (1986) Mol. Pharmacol. 30, 537-543
23. Schenk, D. B., Phelps, M. N., Porter, J. G., Scarborough, R. M., McEnroe, G. A., and Lewicki, J. A. (1985) J . Biol. Chem. 260,
24. Shinjo, M., Hirata, Y., Hagiwara, H., Akiyama, F., Murakami, K., Kohima, S., Shimonaka, M., Inada, Y., and Hirose, S. (1986) Biomed. Res. 7,35-38
25. Vandlen, R. L., Arcuri, K. E., Hupe, L., Keegan, M. E., and Napier, M. A. (1986) Fed. Proc. 45, 2366-2370
26. Koseki, C., Hayashi, Y., Ohnuma, N., and Imai, M. (1986) Biochem. Biophys. Res. Commun. 136, 200-207
27. Shimonaka, M., Saheki, T., Hagiwara, H., Ishido, M., Nogi, A,, Fugita, T., Wakita, K., Inada, Y., Kondo, J., and Hirose, S. (1987) J. Biol. Chem. 262,5510-5514
28. Meloche, S., Ong, H., and De Lean, A. (1987) J. Bid. Chem. 262,
29. Winquist, R. J., Faison, E. P., Waldman, S. A., Schwartz, K., Murad, F., and Rapoport, R. M. (1984) Proc. Natl. Acad. Sci. U. S. A. 8 1 , 7661-7665
30. Leitman, D. C., Andresen, J. W., Kuno, T., Kamisalti, Y., Chang, 3. K., and Murad, F. (1986) J. Biol. Chem. 261, 11650-11655
31. Scarborough, R. M., Schenk, D. B., McEnroe, G. A., Arfsten, A., Kang, L.-L., Schwartz, K., and Lewicki, J. A. (1986) J. Biol. Chem. 261, 12960-12964
5946-5950
istry 25,8467-8472
260,8229-8232
14887-14890
10252-10258

9400 ANP C-receptor cDNA Sequence and Expression 32. Leitman, D. C., and Murad, F. (1986) Biochim. Biophys. Acto
33. Hayashi, Y., Iwasa, F., Furuya, M., Kanai, Y., Minamitake, Y., Kubota, I., Ohnuma, N., Kangawa, K., and Matsuo, H. (1986) Peptide Chem. 1986,27-32
34. Takayanagi, R., Snajdar, R. M., Imada, T., Tamura, M., Pandey, K. N., Misono, K. S., and Inagami, T. (1987) Biochem. Biophys. Res. Commun. 144,244-250
35. Takayanagi, R., Inagami, T., Snajdar, R. M., Imada, T., Tamura, M., and Misono, K. S. (1987) J. Biol. Chem. 262,12104-12113
36. Maack, T., Suzuki, M., Almeida, F. A., Nussenzveig, D., Scarbor- ough, R. M., McEnroe, G. A., and Lewicki, J. A. (1987) Science
37. Schenk, D. B., Phelps, M. N., Porter, J . G., Fuller, F., Cordell, B., and Lewicki, J. (1987) Proc. Natl. Acad. Sci. U. S. A. 84,
38. Ullrich, A., Coussens, L., Hayflick, J. S., Dull, T. J., Gray, A., Tam, A. W., Lee, J., Yarden, Y., Libermann, T. A., Schlessin- ger, J., Downward, J., Mayes, E. L. V., Whittle, N., Waterfield, M. D., and Seeburg, P. H. (1984) Nature 309,418-425
39. Leonard, W. J., Depper, J. M., Crabtree, G. R., Rudikoff, S., Pumphrey, J., Robb, R. J., Kronke, M., Svetlik, P. B., Peffer, N. J., Waldmann, T. A., and Greene, W. C. (1984) Nature 31 1 ,
886,74-79
238,675-678
1521-1525
626-631 40. Von Heijne, G. (1983) Eur. J. Biochem. 133,17-21 41. Walter, P., Gilmore, R., and Blobel, G. (1986) Cell 38, 5-8 42. Ullrich, A., Bell, J. R., Chen, E. Y., Herrera, R., Petruzzell, L.
M., Dull, T. J., Gray, A., Coussens, L., Liao, Y.-C., Tsubokawa, M., Mason, A,, Seeburg, P. H., Grunfeld, C., Rosen, 0. M., and Ramachandran, J. (1985) Nature 313 , 756-761
43. Yamamoto, T., Davis, C. G., Brown, M. S., Schneider, W. J., Casey, M. L., Goldstein, J. L., and Russell, D. W. (1984) Cell
44. Yarden, Y., Escobedo, J. A., Kuang, W.-J., Yang-Feng, T. L., Daniel, T. O., Tremble, P. M., Chen, E. Y., Ando, M. E., Harkins, R. N., Francke, U., Fried, V. A., Ullrich, A., and
39,27-38
James V . Schilling, Robert 1. Scarborough. John A . Levicki and Dale 8. Schcnk Forrest Ful ler* . 1. Gordon Porlec,Ann Arfrfen, Judy Uillcr,
Williams, L. T. (1986) Nature 323 , 226-232 45. Johnson, D., Lanahan, A., Buck, C. R., Sehgal, A., Morgan, C.,
Mercer, E., Bothwell, M., and Chao, M. (1986) Cell 47, 545- 554
46. Morgan, D. O., Edman, J. C., Standring, D. N., Fried, V. A., Smith, M. C., Roth, R. A., and Rutter, W. J. (1987) Nature
47. McClelland, A., Kuhn, L., and Ruddle, F. H. (1984) Cell 39,267- 274
48. Kuno, T., Andresen, J. W., Kamisaki, Y., Waldman, S. A., Chang, L. Y., Saheki, S., Leitman, D. C., Nakane, M., and Murad, F. (1986) J. Bwl. Chem. 261,5817-5823
49. Hirata, Y., Takata, S., Tomita, M., and Takaichi, S. (1985) Biochem. Biophys. Res. Commun. 132,976-984
50. Napier, M. A., Arcuri, K. E., and Vandlen, R. L. (1986) Arch. Biochem. Biophys. 248,516-522
51. Kiess, W., Haskell, J. F., Lee, L., Greenstein, L. A., Miller, B. E., Aarons, A. L., Rechler, M. M., and Nissley, S. P. (1987) J. Biol. Chem. 262,12745-12751
329,301-307
52. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Bwl. 157 , 105-132 53. Deleted in proof 54. Land, H., Grez, M., Hauser, H., Werner, L., and Schutz, G. (1981)
Nucleic Acids Res. 9,2251-2266 55. Wood, W. I., Capon, D. J., Simonsen, C. C., Eaton, D. L.,
Gitschier, J., Keyt, B., Seeburg, P. H., Smith, D. H., Hollings- head, P., Wion, K. L., Delwart, E., Tuddenham, E. G. D., Vehar, G. A., and Lawn, R. M. (1984) Nature 312,330-333
56. Chargwin, J . M., Przybyla, A. E., MacDonald, R. J., and Rutter, W. J. (1979) Biochemistry 18,5294-5299
57. Gurdon, J. B., and Wickens, M. P. (1983) Methods Ertzymol.
58. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.
59. Messing, J., and Vieira, J. (1982) Gene (Amst.) 1 9 , 269-276 60. Lomedico, P., Rosenthal, N., Efstratiadis, A., Gilbert, W., and
61. Kozak, M. (1986) Cell 44,283-292
101,370-386
Acad. Sci. U. S. A. 74,5463-5469
Tizard, R. (1979) Cell 18,545-558

ANP C-receptor cDNA Sequence and Expression 9401 varying concenlrallons of vnlabled ANP 01 ANP m a l o g for 1 hr at 21°C. Reactions were
terminated and free peptide nepsrafed from hound by preciplraflon vlth aeelme (&OX v l v
nonspecific binding I c m o r s bound in the presence Of lWn1 unlabeled rANP(102-126)l "ere
f i n a l ) . Counts hound i n the absence of unlabled ligand were 34 ,824 +I- 514 cpm while
10.212 + / - 457 cpm. Bornbesin (Peninsula Labs) had no effect on binding even at 1WuU.
DNA Sequence of e C-receptor CON& Clones
Thlr revealed over 3558 nuchofides of mRNA sequence. An open reading frame of 1611
ANP C-receptor DDNA clones "ere subjected t o sequence analysis by rhe 113 method (58.59).
nucleotides is depicred by the solid line In Figure 28. This Includes an in-frame segment
encoding the N-fcmmnal amino acid sequence of Figure 2A. A reverse Iranrcriptare strong stop. vhlch may be the 5' end of the receptoc m W A , vas defin-d by the sequence of four
an the 5 , end of pANPRc2. The sequencer of the 5' ends af clones pANPRc2. pANPRcl2 and additional clones obcained by rescreening the library vlth a nondegenerafe 2Omer probe bared
pANPRcl5 ere Identical while those o f pANPRcl3 and pANPRcl4 differ by 4 and 3 nucleatides respectirely. Thc 5' end of the &NP receptor m E N A chur appears t o contain at leas t 465
and probes hared on fhe plWPRc-6 sequence used I o obtain pANPRc-9 and 10. Nearly 1500
noncoding nucleotides. Probes based on the pANPRc-4 sequence "ere used t o obtain pANPRc-6
nwcleoflder of 3' noncoding sequence h = w been obcained with over 500 more defined by partial sequence and mapping of pANPRc9. None of the clones contain a polyIA1 nequenee indicative of
an nRNA 3' terminus, although two po~enfial poly(A) addition ~ignals ( M U M ) ace present a t
2640 and 1191 nucleotlder. The cecepior appear., therefore, 10 be encoded az the 5' end of s
Single nucleotide differences befveen some of the different clones "ere noted, four of
Ihese in the coding region. Nuclcolides 552 . 1010, 1636. and 1558 were G in pANPRc2. G in
pANPRc2, C I n pANfRel and A in pANPRc4 respectively. Since t h i s frequency is sinllar fa the error frequency associated with Reverre Transcriptase (60) 11 is llkely chat there
differences are cloning arrifacfs. The sequence prcsenr in Figure 5 represents the cansenws of a t least 2 clones i n each of these m r l f i o o l .
initiation codon rhavn in f igure 5 are correct. Although the A X shorn a= the lnlliafioo Several obrervafions lead us t o conclude that the reading frame and translation
cadon i n Pigure 5 i s preceded by four additional ATGs which could be s t a r t codons, the latter
four are followed by stop codons In each reading frame as vel1 at; by a T rich region which would encode an unlikely aligaphcnylalanine stretch i n any fcame. A good Irsnrlatlon
initiation signal (GCACC) as defrned by Korak lCCA/~CC) 161) precedes the predicted ATC and
this ATG is in-frame with an oligopepfide sequence identical t o the N-terminal sequence of
the isolated receptor. In addition, the size predicted for the receptor precursar Is i n
excellent agreement vIth the observed size of the 2 translation product ( n r -58,OW)
af RNA nynfhesired w i n g the cDNA3 as template (Figure & A ) . and rhe m i n o acid COmpOSitiOn of
the purified reccpfor (not shown) and if. acidic PI (37) are a l s o i n good agreement with the
medicfed sewonce.





























