Embed Size (px)
Citation preview

Biochemistry. In the article ‘‘A protein encoded by a group Iintron in Aspergillus nidulans directly assists RNA splicing andis a DNA endonuclease’’ by Yugong Ho, Sea-Jin Kim, andRichard B. Waring, which appeared in number 17, August 19,1997, of Proc. Natl. Acad. Sci. USA (94, 8994–8999), Theauthors request that the following change be noted. In Fig. 2 A,the 3 min timepoint lane headings should all read 1 min; Fig.2B is correct.
Biochemistry. In the article “ATP-dependent uptake of nat-ural product cytotoxic drugs by membrane vesicles establishesMRP as a broad specificity transporter” by Saptarshi Paul, LisaM. Breuninger, Kenneth D. Tew, Hongxie Shen, and Gary D.Kruh, which appeared in number 14, July 9, 1996, of Proc. Natl.Acad. Sci. USA (93, 6929–6934), the authors request that thefollowing be noted. “Due to the uncertain validity of the dataand the irreproducibility of some of the results, we retract thisarticle. We are currently unable to reproduce the experimentsindicating that MRP transports natural product drugs. We
apologize for any inconvenience that publication of this studymay have caused.”
Saptarshi PaulLisa M. Breuninger
Kenneth D. TewHongxie ShenGary D. Kruh
Genetics. In the article ‘‘The unique hetero-oligomeric natureof the subunits in the catalytic cooperativity of the yeast Cctchaperonin complex’’ by Paul Lin and Fred Sherman, whichappeared in number 20, September 30, 1997, of Proc. Natl.Acad. Sci. USA (94, 10780–10785), the authors request that thefollowing corrections be noted. On page 10782 in Table 1, thevalues for the cct1-5 3 cct1-1, cct1-5 3 cct1-2, and cct1-5 3cct1-3 crosses should all be 6. Lines 4–6 of the legend of Table1 should read ‘‘0–6 denotes the relative growth of the strains,where 0 denotes no growth, and 6 denotes normal or nearnormal growth.’’ We apologize for these errors.
Genetics. In the article ‘‘Neuronal death in the central nervoussystem demonstrates a non-fibrin substrate for plasmin’’ by StellaE. Tsirka, Thomas H. Bugge, Jay L. Degen, and Sidney Strick-land, which appeared in number 18, September 2, 1997, of Proc.
Natl. Acad. Sci. USA (94, 9779–9781), the authors request that thefollowing correction to Fig. 1 be noted. Due to a printer’s error,Fig. 1 was highly pixelated with extremely poor contrast. Acorrected version with higher contrast is reproduced below.
FIG. 1. Plasmin functions in neuronal cell death independently of fibrin clearance. Cresyl violet-stained coronal sections through thehippocampus reveal the neuronal degeneration generated 5 days after KA injection. (Upper Left) Hippocampus from heterozygous plg1y2;fib1y2
mouse (wt) showing substantial degeneration on the injected side (ipsilateral). CA1, CA2, and CA3 denote the hippocampal subfields. (Upper Right)Hippocampus from plg2y2 mouse showing minimal degeneration on the injected side. (Lower Left) Hippocampus from fib2y2 mouse showingextensive degeneration on the injected side. (Lower Right) Hippocampus from plg2y2;fib2y2 mouse showing minimal degeneration on the injectedside. In each case, the uninjected side (contralateral) showed no detectable degeneration. Arrows show the site of injection. The genotypes werecoded until completion of the experiments and scoring of the results.
14976 Corrections and Retraction Proc. Natl. Acad. Sci. USA 94 (1997)

Proc. Natl. Acad. Sci. USAVol. 93, pp. 6929-6934, July 1996Biochemistry
ATP-dependent uptake of natural product cytotoxic drugs bymembrane vesicles establishes MRP as a broadspecificity transporterSAPrARSHI PAUL*, LISA M. BREUNINGER*, KENNETH D. TEWt, HONGXIE SHENt, AND GARY D. KRUH*tDepartments of *Medical Oncology and tPharmacology, Fox Chase Cancer Center, Philadelphia, PA 19111
Communicated by Robert P. Perry, Fox Chase Cancer Center, Philadelphia, PA, March 4, 1996 (received for review August 28, 1995)
ABSTRACT MRP is a recently isolated ATP-binding cas-sette family transporter. We previously reported transfectionstudies that established that MRP confers multidrug resis-tance [Kruh, G. D., Chan, A., Myers, K., Gaughan, K., Miki,T. & Aaronson, S. A. (1994) Cancer Res. 54, 1649-1652] andthat expression of MRP is associated with enhanced cellularefflux of lipophilic cytotoxic agents [Breuninger, L. M., Paul,S., Gaughan, K., Miki, T., Chan, A., Aaronson, S. A. & Kruh,G. D. (1995) Cancer Res. 55, 5342-5347]. To examine thebiochemical mechanism by which MRP confers multidrugresistance, drug uptake experiments were performed usinginside-out membrane vesicles prepared from NIH 3T3 cellstransfected with an MRP expression vector. ATP-dependenttransport was observed for several lipophilic cytotoxic agentsincluding daunorubicin, etoposide, and vincristine, as well asfor the glutathione conjugate leukotriene C4 (LTC4). However,only marginally increased uptake was observed for vinblastineand Taxol. Drug uptake was osmotically sensitive and satu-rable with regard to substrate concentration, with Km valuesof 6.3 ,IM, 4.4 ,uM, 4.2 FM, 35 nM, and 38 ,uM, for dauno-rubicin, etoposide, vincristine, LTC4, and ATP, respectively.The broad substrate specificity ofMRP was confirmed by theobservation that daunorubicin transport was competitivelyinhibited by reduced and oxidized glutathione, the glutathioneconjugates S-(p-azidophenacyl)-glutathione (APA-SG) andS-(2,4-dinitrophenyl)glutathione (DNP-SG), arsenate, andthe LTD4 antagonist MK571. This study establishes that MRPpumps unaltered lipophilic cytotoxic drugs, and suggests thatthis activity is an important mechanism by which the trans-porter confers multidrug resistance. The present study alsoindicates that the substrate specificity ofMRP is overlappingbut distinct from that of P-glycoprotein, and includes both theneutral or mildly cationic natural product cytotoxic drugs andthe anionic products of glutathione conjugation. The wide-spread expression ofMRP in tissues, combined with its abilityto transport both lipophilic xenobiotics and the products ofphase II detoxification, indicates that the transporter repre-sents a widespread and remarkably versatile cellular defensemechanism.
Cellular resistance to cytotoxic drugs is a major obstacle to thetreatment of disseminated cancers. Resistant cancer cells candisplay a multidrug resistant phenotype that involves simulta-neous resistance to a spectrum of natural product agents,including anthracyclines, vinca alkaloids, epipodophyllotoxins,and dactinomycin. One form of multidrug resistance resultsfrom overexpression of P-glycoprotein (Pgp), the 170-kDaproduct of the MDR1 gene. Pgp, an ATP-binding cassettefamily transporter, functions as an ATP-dependent effluxpump that reduces the intracellular concentrations of chemo-therapeutic agents by transporting them across the plasma
membrane (1, 2). Recently, MRP, another ATP-binding cas-sette transporter, was isolated from a drug-resistant cell line(3). We and others have reported transfection studies thatestablished that MRP.confers resistance to lipophilic cytotoxicagents (4, 5). However, the biochemical mechanism by whichthe transporter confers resistance is unknown. The detectionof increased cellular drug efflux in MRP transfectants (6-8)and in some (9-12), but not all (13), drug-selected cell linesthat overexpress MRP suggests that it may function as a pumpfor natural product drugs. However, reports that MRP inmembrane vesicles pumps glutathione conjugates (14-16) andthe observation that the glutathione depleting agent butathi-onine sulfoximine (BSO) reverses MRP-conferred resistance(17-20) have led to consideration of the idea thatMRP transportseither the glutathione conjugates or anionic metabolites of drugs,as opposed to transporting unaltered drugs (21).
In the present study, we examined the biochemical mecha-nism of MRP action by analyzing drug uptake into membranevesicles prepared from NIH 3T3 cells transfected with an MRPexpression vector. Our results indicate that MRP is an ATP-dependent pump for a spectrum of natural product cytotoxicdrugs, including anthracyclines, vincristine, and etoposide.Thus, this study provides the first direct evidence that MRPpumps unaltered lipophilic cytotoxic drugs, and suggests thatthis activity is an important component of MRP-conferredmultidrug resistance.
MATERIALS AND METHODSMaterials. Antipain, creatine phosphate, ATP, 5'-AMP,
pepstatin A, benzamidine, glutathione, oxidized glutathione,S-(p-azidophenacyl)-glutathione (APA-SG), RNase A, andDNase I were purchased from Sigma. Arsenate was purchasedfrom J. T. Baker. Creatine kinase, leupeptin A, and aprotininwere purchased from Boehringer Mannheim. [3H]LeukotrieneC4 (LTC4) (140 Ci/mmol; 1 Ci = 37 GBq) and [G-3H]dauno-mycin (3.7 Ci/mmol) were purchased from DuPont/NEN.[3H]Etoposide (900 mCi/mmol), [3H]vinblastine sulfate (9Ci/mmol), and [3H]taxol (7.4 Ci/mmol) were purchased fromMoravek Biochemicals (Brea, CA). [3H]Vincristine sulfate(10.6 Ci/mmol) was purchased from Amersham. MK571 wasa gift of A. W. Ford-Hutchinson (Merck-Frost Center forTherapeutic Research, Quebec).
S-(2,4-dinitrophenyl)glutathione (DNP-SG) was preparedas described (22). Briefly, 5 mmol of glutathione, 10 mmol of1-chloro-2,4-dinitrobenzene, and 100 mg of rat liver glutathi-one S-transferase (Sigma) were mixed in a final volume of 3 mlof 50 mM Tris-HCl (pH 7.4), and incubated for 1 hr at 37°C.The incubation mixture was diluted 2-fold with cold waterapplied onto a QAE-Sephadex column. The column waswashed with 10 mM Tris-HCl (pH 7.4) to remove unreacted
Abbreviations: LTC4, leukotriene C4; Pgp, P-glycoprotein; BSO, bu-tathione sulfoximine; APA-SG, S-(p-azidophenacyl)-glutathione;DNP-SG, S-(2,4-dinitrophenyl)glutathione.4To whom reprint requests should be addressed.
6929
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 93 (1996)
precursors and the DNP-SG was eluted with 0.5 N formic acid.The product was collected, lyophilized, and stored at -20°C.
Preparation of Membrane Vesicles and Drug Uptake Anal-ysis. Inside-out membrane vesicles were prepared as described(23, 24) from the previously reported NIH 3T3 MRP trans-fectant pSRa-MRP-32 cells and the control transfectant pSRacells (6). Drug uptake by inside-out membrane vesicles wasmeasured by the rapid filtration technique (25). Invertedmembrane vesicles were quickly thawed at 37°C shortly beforeuse and kept on ice. Radiolabeled drugs were mixed with ATPreaction buffer (10 mM Tris-HCl, pH 7.5/4 mM ATP/10 mMMgCl2/10 mM creatine phosphate/0.25 M sucrose/100 jig/mlcreatine kinase) and membrane vesicles (20 ,ug of protein)were added (ATP concentration of 1.2 mM in the final reactionmixture). The suspension was incubated at 37°C and 25-,ulaliquots were removed at various times and diluted in 1.0 mlof ice-cold incubation buffer. The suspensions were immedi-ately applied to 0.45 ,um Millipore nitrocellulose filters (pre-soaked in incubation buffer) under suction and washed twicewith 5.0 ml of ice-cold incubation buffer. The filters weredissolved in scintillation fluid and the radioactivity measuredin a Beckman liquid scintillation counter. In parallel controlexperiments, ATP in the reaction buffer was replaced with thesame concentration of 5'-AMP. ATP-dependent transport wascalculated by subtracting the uptake values obtained with5'-AMP from those obtained with ATP. Osmotic sensitivityexperiments were performed by incubating the membranevesicles in incubation buffer containing various concentrationsof sucrose for 15 min at 25°C. Drug uptake was then measuredin the presence of either ATP or 5'-AMP as described above,except that the sucrose concentration of the ATP reactionbuffer was adjusted to that of the incubation buffer. ForLineweaver-Burk and sucrose inhibition analyses, initial re-action rates were measured at 60 sec. All experiments wereperformed in triplicate.
Generation ofMRP Monoclonal Antibody.A segment of theMRP cDNA (6) encoding aa 609-761 was generated by PCRand inserted into the bacterial expression vector pDlO (a giftof Frank Raucher, Wistar Institute, Philadelphia). The recom-binant histidine fusion protein was expressed in bacteria andpurified using a nickel column. Immunization of BALB/c micewith the recombinant protein, ELISA assays, and splenicfusion were performed as described (26). Culture medium wastested for the presence of antibodies to the recombinant fusionprotein and several positive cultures were tested for reactivityto MRP in immunoblots containing lysates prepared fromMRP-overexpressing cell lines. One supernatant detected MRPbut not Pgp in immunoblots and was designated MRPmAb-1.Immunoblot Analysis. Protein samples (50 ,ug) of cell lysates
or membrane vesicles were separated on 6% SDS-polyacryl-amide gels and transferred to nitrocellulose membranes(Schleicher and Schuell). Filters were blocked in PBS contain-ing 0.1% Tween-20 (PBST) and 5% bovine serum albumin,and MRP was detected using MRPmAb-1 and a secondaryanti-mouse antibody. Blots were washed several times withPBST before and after the secondary antibody treatment andthree times in PBS alone before development with an En-hanced Chemiluminescence kit (Amersham).
RESULTSATP-Dependent Transport of Daunorubicin by Inside-Out
Membrane Vesicles Prepared from MRP Transfectants. Wepreviously characterized pSRa-MRP-32, a G418-selected NIH3T3 MRP transfectant that displays 8-12 fold resistance toetoposide, daunorubicin, and doxorubicin, -3 fold resistanceto vincristine, and marginally increased resistance to vinblas-tine and Taxol (6). To examine the substrate specificity ofMRP, inside-out membrane vesicles were prepared from theMRP transfectant and pSRa, a G418-selected control cell line
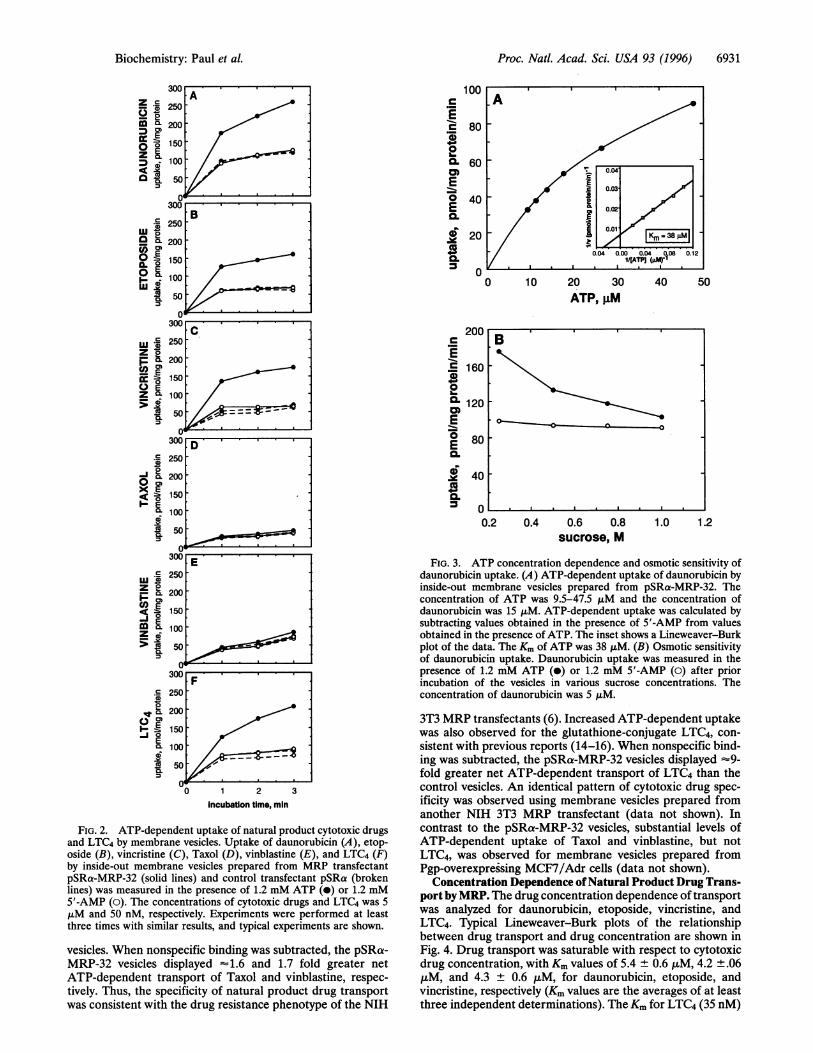
obtained by transfection of expression vector lacking the MRPcDNA (6). Fig. 1 shows the detection of MRP in lysates andinside-out membrane vesicles prepared from the two cell lines.The membrane vesicles were used to analyze daunorubicintransport. Fig. 2A shows a time course of daunorubicin uptakeover 3 min. To distinguish between ATP-dependent transportof drug into membrane vesicles and nonspecific binding,vesicle-associated drug was measured in the presence of AMPor ATP. As has been previously reported for vesicle transportstudies of lipophilic cytotoxic drugs (25, 27, 28), substantialnonspecific binding of daunorubicin was detected (AMP con-trols). Comparable levels of nonspecific binding were detectedfor the pSRa-MRP-32 and control vesicles. However, in thepresence ofATP, substantial drug uptake was observed for thepSRa-MRP-32 vesicles but not for the control vesicles. Whennonspecific binding was subtracted, the pSRa-MRP-32 vesiclesdisplayed -15 fold greater net uptake than the control vesiclesat 3 min.Osmotic Sensitivity and ATP Concentration Dependence of
Daunorubicin Transport. To confirm that the observeddaunorubicin uptake was the result of transport into mem-brane vesicles, the ATP concentration dependence and os-motic sensitivity of uptake were analyzed. Transport shouldincrease with increasing concentrations of ATP in the incu-bation mixture if uptake is energy dependent. As shown in Fig.3A, daunorubicin uptake by inside-out vesicles prepared frompSRa-MRP-32 increased with increasing ATP concentrations.Lineweaver-Burk analysis indicated that daunorubicin trans-port was saturable with respect to ATP concentration, with aKm of 38 ,uM for ATP (Fig. 3A Inset). A biochemical hallmarkof transport into membrane vesicles, as opposed to nonspecificbinding, is inhibition at high osmotic pressures (25, 27, 28).High osmotic pressure reduces the intravesicular space byshrinking the membrane vesicles, the result ofwhich is reduceduptake capacity. As expected for transport into membranevesicles, ATP-dependent uptake of daunorubicin dramaticallydecreased as sucrose concentrations were increased above thestandard assay conditions af 0.25 M, whereas nonspecificbinding in the presence of AMP was unchanged (Fig. 3B). At1.0 M sucrose, vesicle associated daunorubicin in the presenceof ATP was comparable with the AMP values.
Specificity of Natural Product Drug Transport. Drug trans-port by MRP was analyzed for a spectrum of natural productagents. As shown in Fig. 2, markedly enhanced ATP-dependent uptake was observed for etoposide and vincristine.When nonspecific binding was subtracted, the pSRa-MRP-32vesicles displayed -11 and 14 fold greater net ATP-dependent uptake for etoposide and vincristine, respectively,than the control vesicles. Similar levels of ATP-dependentuptake were also observed for doxorubicin (data not shown).In contrast, only slight increases in ATP-dependent uptake ofTaxol and vinblastine were detected for the pSRa-MRP-32
1 2 3 4
200 kDa-
MRP--p-
FIG. 1. Detection of MRP in cell lysates and membrane vesicles.Inside-out membrane vesicles were prepared from control transfectantpSRa or MRP transfectant pSRa-MRP-32 and 50 ,ug samples oflysates (lanes 1 and 2, respectively) or membrane vesicles (lanes 3 and4, respectively) were separated by 6% SDS/PAGE. MRP was detectedusing monoclonal antibody MRPmAb-1. The position of a 200-kDamolecular mass marker is shown.
6930 Biochemistry: Paul et al.
Proc. Natl. Acad. Sci. USA 93 (1996) 6931
100C
c 80
Si100. 60
E0. 4E
l 20
0.
0
0 10 20 30ATP, FM
25
F~ 2C
Z O 15E
Z o- lF. d
t 5
(I)9
0.
0
~.0
wJ
0c.9
0.
,o
30U
250
200
150r
100i
50
25u
250
2001
150~
101
Om
Incubation time, min
FIG. 2. ATP-dependent uptake of natural product cytotoxic drugsand LTC4 by membrane vesicles. Uptake of daunorubicin (A), etop-oside (B), vincristine (C), Taxol (D), vinblastine (E), and LTC4 (F)by inside-out membrane vesicles prepared from MRP transfectantpSRa-MRP-32 (solid lines) and control transfectant pSRa (brokenlines) was measured in the presence of 1.2 mM ATP (-) or 1.2 mM5'-AMP (0). The concentrations of cytotoxic drugs and LTC4 was 5,uM and 50 nM, respectively. Experiments were performed at leastthree times with similar results, and typical experiments are shown.
vesicles. When nonspecific binding was subtracted, the pSRa-MRP-32 vesicles displayed -1.6 and 1.7 fold greater netATP-dependent transport of Taxol and vinblastine, respec-tively. Thus, the specificity of natural product drug transportwas consistent with the drug resistance phenotype of the NIH
200 B 'gC B
3 1600.1r-L 120-
o 80E0.
l. 40
0o . . . I
0.2 0.4 0.6 0.8sucrose, M
1.0 1.2
FIG. 3. ATP concentration dependence and osmotic sensitivity ofdaunorubicin uptake. (A) ATP-dependent uptake of daunorubicin byinside-out membrane vesicles prepared from pSRa-MRP-32. Theconcentration of ATP was 9.5-47.5 ,uM and the concentration ofdaunorubicin was 15 ,uM. ATP-dependent uptake was calculated bysubtracting values obtained in the presence of 5'-AMP from valuesobtained in the presence of ATP. The inset shows a Lineweaver-Burkplot of the data. The Km of ATP was 38 ,uM. (B) Osmotic sensitivityof daunorubicin uptake. Daunorubicin uptake was measured in thepresence of 1.2 mM ATP (-) or 1.2 mM 5'-AMP (0) after priorincubation of the vesicles in various sucrose concentrations. Theconcentration of daunorubicin was 5 ,uM.
3T3 MRP transfectants (6). Increased ATP-dependent uptakewas also observed for the glutathione-conjugate LTC4, con-sistent with previous reports (14-16). When nonspecific bind-ing was subtracted, the pSRa-MRP-32 vesicles displayed -9-fold greater net ATP-dependent transport of LTC4 than thecontrol vesicles. An identical pattern of cytotoxic drug spec-ificity was observed using membrane vesicles prepared fromanother NIH 3T3 MRP transfectant (data not shown). Incontrast to the pSRa-MRP-32 vesicles, substantial levels ofATP-dependent uptake of Taxol and vinblastine, but notLTC4, was observed for membrane vesicles prepared fromPgp-overexpressing MCF7/Adr cells (data not shown).
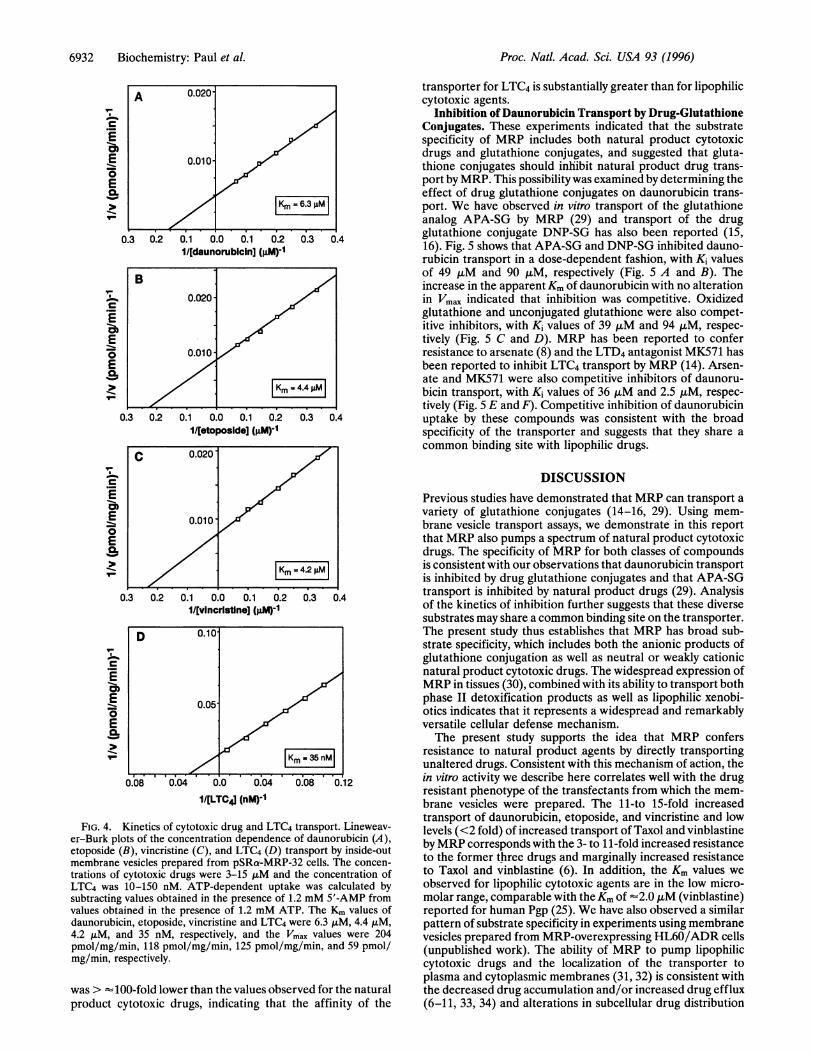
Concentration Dependence ofNatural Product Drug Trans-port by MRP. The drug concentration dependence of transportwas analyzed for daunorubicin, etoposide, vincristine, andLTC4. Typical Lineweaver-Burk plots of the relationshipbetween drug transport and drug concentration are shown inFig. 4. Drug transport was saturable with respect to cytotoxicdrug concentration, with Km values of 5.4 ± 0.6 ,uM, 4.2 ±.06tLM, and 4.3 ± 0.6 ,uM, for daunorubicin, etoposide, andvincristine, respectively (Km values are the averages of at leastthree independent determinations). The Km for LTC4 (35 nM)
300Z ,, 250 A
m n200
o o 150
100
O5 50
)u"U
*,, 25wo.
20
1U cm 5is10
'B
ioGSK0
40 50
50)o An0
n
--
-
u -
0
n.
Biochemistry: Paul et al.
nM
Proc. Natl. Acad. Sci. USA 93 (1996)
E
-
Ecm
E
0
E0.
A 0.020-
0.010/
0.3 0.2 0.1 0.0 0.1 0.2 0.3 0.1/[daunorubicin] (M)-1
B0.020-
0.010
IKm = ,. M
0.3 0.2 0.1 0.0 0.1 0.21/[etoposide] (pM)-l
EcmE
0
E
:0
I-
1-.
0.08
0.3
.4
transporter for LTC4 is substantially greater than for lipophiliccytotoxic agents.
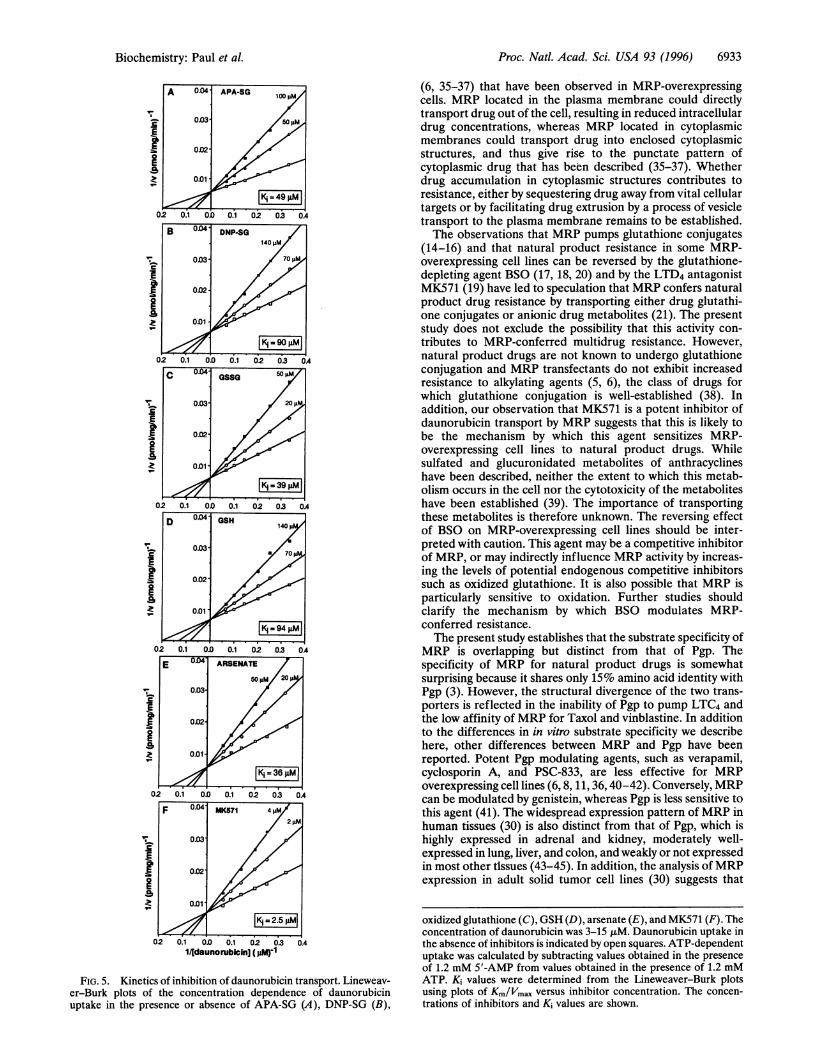
Inhibition of Daunorubicin Transport by Drug-GlutathioneConjugates. These experiments indicated that the substratespecificity of MRP includes both natural product cytotoxicdrugs and glutathione conjugates, and suggested that gluta-thione conjugates should inhibit natural product drug trans-port by MRP. This possibility was examined by determining theeffect of drug glutathione conjugates on daunorubicin trans-port. We have observed in vitro transport of the glutathioneanalog APA-SG by MRP (29) and transport of the drugglutathione conjugate DNP-SG has also been reported (15,16). Fig. 5 shows that APA-SG and DNP-SG inhibited dauno-rubicin transport in a dose-dependent fashion, with Ki valuesof 49 ,tM and 90 ,tM, respectively (Fig. 5 A and B). Theincrease in the apparent Km of daunorubicin with no alterationin Vma, indicated that inhibition was competitive. Oxidizedglutathione and unconjugated glutathione were also compet-itive inhibitors, with Ki values of 39 ,uM and 94 ,uM, respec-tively (Fig. 5 C and D). MRP has been reported to conferresistance to arsenate (8) and the LTD4 antagonist MK571 hasbeen reported to inhibit LTC4 transport by MRP (14). Arsen-ate and MK571 were also competitive inhibitors of daunoru-bicin transport, with Ki values of 36 ,M and 2.5 ,tM, respec-tively (Fig. 5 E and F). Competitive inhibition of daunorubicinuptake by these compounds was consistent with the broadspecificity of the transporter and suggests that they share acommon binding site with lipophilic drugs.
0.4
0.12
1/[LTC4J (nM)l
FIG. 4. Kinetics of cytotoxic drug and LTC4 transport. Lineweav-er-Burk plots of the concentration dependence of daunorubicin (A),etoposide (B), vincristine (C), and LTC4 (D) transport by inside-outmembrane vesicles prepared from pSRa-MRP-32 cells. The concen-
trations of cytotoxic drugs were 3-15 ,uM and the concentration ofLTC4 was 10-150 nM. ATP-dependent uptake was calculated bysubtracting values obtained in the presence of 1.2 mM 5'-AMP fromvalues obtained in the presence of 1.2 mM ATP. The Km values ofdaunorubicin, etoposide, vincristine and LTC4 were 6.3 ,uM, 4.4 ,uM,4.2 ,LM, and 35 nM, respectively, and the Vmax values were 204pmol/mg/min, 118 pmol/mg/min, 125 pmol/mg/min, and 59 pmol/mg/min, respectively.
was > 100-fold lower than the values observed for the naturalproduct cytotoxic drugs, indicating that the affinity of the
DISCUSSIONPrevious studies have demonstrated that MRP can transport avariety of glutathione conjugates (14-16, 29). Using mem-brane vesicle transport assays, we demonstrate in this reportthat MRP also pumps a spectrum of natural product cytotoxicdrugs. The specificity of MRP for both classes of compoundsis consistent with our observations that daunorubicin transportis inhibited by drug glutathione conjugates and that APA-SGtransport is inhibited by natural product drugs (29). Analysisof the kinetics of inhibition further suggests that these diversesubstrates may share a common binding site on the transporter.The present study thus establishes that MRP has broad sub-strate specificity, which includes both the anionic products ofglutathione conjugation as well as neutral or weakly cationicnatural product cytotoxic drugs. The widespread expression ofMRP in tissues (30), combined with its ability to transport bothphase II detoxification products as well as lipophilic xenobi-otics indicates that it represents a widespread and remarkablyversatile cellular defense mechanism.The present study supports the idea that MRP confers
resistance to natural product agents by directly transportingunaltered drugs. Consistent with this mechanism of action, thein vitro activity we describe here correlates well with the drugresistant phenotype of the transfectants from which the mem-brane vesicles were prepared. The 11-to 15-fold increasedtransport of daunorubicin, etoposide, and vincristine and lowlevels (<2 fold) of increased transport of Taxol and vinblastineby MRP corresponds with the 3- to 11-fold increased resistanceto the former three drugs and marginally increased resistanceto Taxol and vinblastine (6). In addition, the Km values weobserved for lipophilic cytotoxic agents are in the low micro-molar range, comparable with the Km of -2.0 ,uM (vinblastine)reported for human Pgp (25). We have also observed a similarpattern of substrate specificity in experiments using membranevesicles prepared from MRP-overexpressing HL60/ADR cells(unpublished work). The ability of MRP to pump lipophiliccytotoxic drugs and the localization of the transporter toplasma and cytoplasmic membranes (31, 32) is consistent withthe decreased drug accumulation and/or increased drug efflux(6-11, 33, 34) and alterations in subcellular drug distribution
6932 Biochemistry: Paul et al.
Proc. Natl. Acad. Sci. USA 93 (1996) 6933
0.
0.1 0.0 0.1 02
14[daunorublcin] (jlM)1
FIG. 5. Kinetics of inhibition of daunorubicin transport. Lineweav-er-Burk plots of the concentration dependence of daunorubicinuptake in the presence or absence of APA-SG (A), DNP-SG (B),
(6, 35-37) that have been observed in MRP-overexpressingcells. MRP located in the plasma membrane could directlytransport drug out of the cell, resulting in reduced intracellulardrug concentrations, whereas MRP located in cytoplasmicmembranes could transport drug into enclosed cytoplasmicstructures, and thus give rise to the punctate pattern ofcytoplasmic drug that has been described (35-37). Whetherdrug accumulation in cytoplasmic structures contributes toresistance, either by sequestering drug away from vital cellulartargets or by facilitating drug extrusion by a process of vesicletransport to the plasma membrane remains to be established.The observations that MRP pumps glutathione conjugates
(14-16) and that natural product resistance in some MRP-overexpressing cell lines can be reversed by the glutathione-depleting agent BSO (17, 18, 20) and by the LTD4 antagonistMK571 (19) have led to speculation that MRP confers naturalproduct drug resistance by transporting either drug glutathi-one conjugates or anionic drug metabolites (21). The presentstudy does not exclude the possibility that this activity con-tributes to MRP-conferred multidrug resistance. However,natural product drugs are not known to undergo glutathioneconjugation and MRP transfectants do not exhibit increasedresistance to alkylating agents (5, 6), the class of drugs forwhich glutathione conjugation is well-established (38). Inaddition, our observation that MK571 is a potent inhibitor ofdaunorubicin transport by MRP suggests that this is likely tobe the mechanism by which this agent sensitizes MRP-overexpressing cell lines to natural product drugs. Whilesulfated and glucuronidated metabolites of anthracyclineshave been described, neither the extent to which this metab-olism occurs in the cell nor the cytotoxicity of the metaboliteshave been established (39). The importance of transportingthese metabolites is therefore unknown. The reversing effectof BSO on MRP-overexpressing cell lines should be inter-preted with caution. This agent may be a competitive inhibitorof MRP, or may indirectly influence MRP activity by increas-ing the levels of potential endogenous competitive inhibitorssuch as oxidized glutathione. It is also possible that MRP isparticularly sensitive to oxidation. Further studies shouldclarify the mechanism by which BSO modulates MRP-conferred resistance.The present study establishes that the substrate specificity of
MRP is overlapping but distinct from that of Pgp. Thespecificity of MRP for natural product drugs is somewhatsurprising because it shares only 15% amino acid identity withPgp (3). However, the structural divergence of the two trans-porters is reflected in the inability of Pgp to pump LTC4 andthe low affinity of MRP for Taxol and vinblastine. In additionto the differences in in vitro substrate specificity we describehere, other differences between MRP and Pgp have beenreported. Potent Pgp modulating agents, such as verapamil,cyclosporin A, and PSC-833, are less effective for MRPoverexpressing cell lines (6, 8, 11, 36, 40-42). Conversely, MRPcan be modulated by genistein, whereas Pgp is less sensitive tothis agent (41). The widespread expression pattern of MRP inhuman tissues (30) is also distinct from that of Pgp, which ishighly expressed in adrenal and kidney, moderately well-expressed in lung, liver, and colon, and weakly or not expressedin most othertissues (43-45). In addition, the analysis of MRPexpression in adult solid tumor cell lines (30) suggests that
oxidized glutathione (C), GSH (D), arsenate (E), and MK571 (F). Theconcentration of daunorubicin was 3-15,uM. Daunorubicin uptake inthe absence of inhibitors is indicated by open squares. ATP-dependentuptake was calculated by subtracting values obtained in the presenceof 1.2 mM 5'-AMP from values obtained in the presence of 1.2 mMATP. Ki values were determined from the Lineweaver-Burk plotsusing plots of Km/Vmax versus inhibitor concentration. The concen-trations of inhibitors and Ki values are shown.
A 0.04 APA-SG 100
0.03-
0.02
0.01
'S= 49iLMl
12 0.1 0.0 0.1 0.2 0. 04
Biochemistry: Paul et al.

Proc. Natl. Acad. Sci. USA 93 (1996)
MRP is likely to be widely expressed in common adult tumorssuch as colon, breast and lung, and ovary, whereas Pgp isexpressed in colon, renal, and liver tumors, but poorly or notexpressed in breast, lung, and ovarian tumors (46, 47). Thesedifferences in substrate specificity and expression may haveimportant clinical correlates and possibly afford opportunitiesfor novel treatment strategies. For example, it is possible thatthe sensitivity of ovarian and breast tumors to Taxol may relateto both the low levels of Pgp expressed in these tumors, and theweak affinity of MRP for Taxol, although MRP is likely to bewell-expressed in these tumors (30). In addition, our observa-tion that glutathione conjugates and MK571 can competitivelyinhibit lipophilic drug transport by MRP suggests that novelcompounds may modulate MRP but not Pgp activity, andraises the possibility that a combination of circumventingagents may be required to achieve effective modulation ofmultidrug resistance in tumors in which both transporters areexpressed.
We thank Irwin Rose, Naomi Laing, and Abhijit Raha for theiradvice on kinetic studies. This work was supported by NationalInstitutes of Health Grant CA63173 and a Betz Foundation grant toG.D.K. Additional support was provided by U.S. Public Health ServiceGrant CA06927 and by an appropriation from the Commonwealth ofPennsylvania to the Fox Chase Cancer Center.
1. Gottesman, M. M. & Pastan, I. (1993) Annu. Rev. Biochem. 62,385-427.
2. Endicott, J. A. & Ling, V. (1989) Annu. Rev. Biochem. 58,137-171.
3. Cole, S. P., Bhardwaj, G., Gerlach, J. H., Mackie, J. E., Grant,C. E., Almquist, K. C., Stewart, A. J., Kurz, E. U., Duncan, A. M.& Deeley, R. G. (1992) Science 258, 1650-1654.
4. Kruh, G. D., Chan, A., Myers, K., Gaughan, K., Miki, T. &Aaronson, S. A. (1994) Cancer Res. 54, 1649-1652.
5. Grant, C. E., Valdimarsson, G., Hipfner, D. R., Almquist, K. C.,Cole, S. P. & Deeley, R. G. (1994) Cancer Res. 54, 357-361.
6. Breuninger, L. M., Paul, S., Gaughan, K., Miki, T., Chan, A.,Aaronson, S. A. & Kruh, G. D. (1995) CancerRes. 55,5342-5347.
7. Zaman, G. J. R., Flens, M. J., van Leusden, M. R., de Haas, M.,Mulder, H. S., Lankelma, J., Pinedo, H. M., Scheper, R. J., Baas,F., Broxterman, H. J. & Borst, P. (1994) Proc. Natl. Acad. Sci.USA 91, 8822-8826.
8. Cole, S. P., Sparks, K. E., Fraser, K., Loe, D. W., Grant, C. E.,Wilson, G. M. & Deeley, R. G. (1994) CancerRes. 54,5902-5910.
9. Slovak, M. L., Hoeltge, G. A., Dalton, W. S. & Trent, J. M.(1988) Cancer Res. 48, 2793-2797.
10. Versantvoort, C. H., Broxterman, H. J., Pinedo, H. M., de Vries,E. G., Feller, N., Kuiper, C. M. & Lankelma, J. (1992) CancerRes. 52, 17-23.
11. Coley, H. M., Workman, P. & Twentyman, P. R. (1991) Br. J.Cancer 63, 351-357.
12. McGrath, T. & Center, M. S. (1988) Cancer Res. 48, 3959-3963.13. Cole, S. P., Chanda, E. R., Dicke, F. P., Gerlach, J. H. & Mirski,
S. E. (1991) Cancer Res. 51, 3345-3352.14. Leier, I., Jedlitschky, G., Buchholz, U., Cole, S. P., Deeley, R. G.
& Keppler, D. (1994) J. Biol. Chem. 269, 27807-27810.15. Jedlitschky, G., Leier, I., Buchholz, U., Center, M. & Keppler, D.
(1994) Cancer Res. 54, 4833-4836.16. Muller, M., Meijer, C., Zaman, G. J., Borst, P., Scheper, R. J.,
Mulder, N. H., de Vries, E. G. & Jansen, P. L. (1994) Proc. Natl.Acad. Sci. USA 91, 13033-13037.
17. Meijer, C., Mulder, N. H., Timmer-Bosscha, H., Peters, W. H. &de Vries, E. G. (1991) Int. J. Cancer 49, 582-586.
18. Versantvoort, C. H., Broxterman, H. J., Bagrij, T., Scheper, R. J.& Twentyman, P. R. (1995) Br. J. Cancer 72, 82-89.
19. Gekeler, V., Ise, W., Sanders, K. H., Ulrich, W. R. & Beck, J.(1995) Biochem. Biophys. Res. Commun. 208, 345-352.
20. Schneider, E., Yamazaki, H,, Sinha, B. K. & Cowan, K. H. (1995)Br. J. Cancer 71, 738-743.
21. Ishikawa, T., Akimaru, K., Kuo, M. T., Priebe, W. & Suzuki, M.(1995) J. Natl. Cancer Inst. 87, 1639-1640.
22. Ishikawa, T. (1989) J. Bio. Chem. 264, 17343-17348.23. Ishikawa, T., Muller, M., Klunemann, C., Schaub, T. & Keppler,
D. (1990) J. Bio. Chem. 265, 19279-19286.24. Leier, I., Jedlitschky, G., Buchholz, U. & Keppler, D. (1994) Eur.
J. Biochem. 220, 599-606.25. Horio, M., Gottesman, M. M. & Pastan, I. (1988) Proc. Natl.
Acad. Sci. USA 85, 3580-3584.26. Bizub-Bender, D., Kulkosky, J. & Skalka, A. M. (1994)AIDSRes.
Hum. Retroviruses 10, 1105-1115.27. Lelong, I. H., Padmanabhan, R., Lovelace, E., Pastan, I. &
Gottesman, M. M. (1992) FEBS Lett. 304, 256-260.28. Kamimoto, Y., Gatmaitan, Z., Hsu, J. & Arias, I. M. (1989)
J. Biol. Chem. 264, 11693-11698.29. Shen, H., Paul, S., Breuninger, L. M., Ciaccio, P., Laing, M., Helt,
M., Tew, K. D. & Kruh, G. D. (1996) Biochemistry 35, 5719-5725.30. Kruh, G. D., Gaughan, K. T., Godwin, A. K. & Chan, A. (1995)
J. Natl. Cancer Inst. 87, 1256-1258.31. Krishnamachary, N. & Center, M. S. (1993) Cancer Res. 53,
3658-3661.32. Flens, M. J., Izquierdo, M. A., Scheffer, G. L., Fritz, J. M., Mei-
jer, C. J., Scheper, R. J. & Zaman, G. J. (1994) Cancer Res. 54,4557-4563.
33. Zijlstra, J. G., de Vries, E. G. & Mulder, N. H. (1987) CancerRes.47, 1780-1784.
34. McGrath, T., Latoud, C., Arnold, S. T., Safa, A. R., Felsted, R. L.& Center, M. S. (1989) Biochem. Pharmacol. 38, 3611-3619.
35. Marquardt, D. & Center, M. S. (1992) CancerRes. 52,3157-3163.36. Barrand, M. A., Rhodes, T., Center, M. S. & Twentyman, P. R.
(1993) Eur. J. Cancer 29A, 408-415.37. Coley, H. M., Amos, W. B., Twentyman, P. R. & Workman, P.
(1993) Br. J. Cancer 67, 1316-1323.38. Tew, K. D. (1994) Cancer Res. 54, 4313-4320.39. Pratt, W. B., Ruddon, R. W., Ensminger, W. D. & Maybaum, J.
(1994) The Anticancer Drugs (Oxford Univ. Press, New York).40. Twentyman, P. R., Fox, N. E. & Bleehen, N. M. (1986) Int. J.
Radiat. Oncol. Biol. Phys. 12, 1355-1358.41. Versantvoort, C. H., Schuurhuis, G. J., Pinedo, H. M., Eekman,
C. A., Kuiper, C. M., Lankelma, J. & Broxterman, H. J. (1993)Br. J. Cancer 68, 939-946.
42. Cole, S. P., Downes, H. F. & Slovak, M. L. (1989) Br. J. Cancer59, 42-46.
43. Fojo, A. T., Ueda, K., Slamon, D. J., Poplack, D. G., Gottesman,M. M. & Pastan, I. (1987) Proc. Natl. Acad. Sci. USA 84,265-269.
44. Sugawara, I., Kataoka, I., Morishita, Y., Hamada, H., Tsuruo, T.,Itoyama, S. & Mori, S. (1988) Cancer Res. 48, 1926-1929.
45. Cornwall, M. M. (1991) in Molecular and Clinical Advances inAnticancer Drug Resistance, ed. Ozols, R. F. (Kluwer, Boston),pp. 37-56.
46. Goldstein, L. J., Galski, H., Fojo, A., Willingham, M., Lai, S. L.,Gazdar, A., Pirker, R., Green, A., Crist, W., Brodeur, G. M.,Lieber, M., Cossman, J., Gottesman, M. M. & Pastan, I. (1989)J. Natl. Cancer Inst. 81, 116-124.
47. Goldstein, L. J., Gottesman, M. M. & Pastan, I. (1991) CancerTreat. Res. 57, 101-119.
6934 Biochemistry: Paul et al.





























