Embed Size (px)
Citation preview

Association Mapping for Epistasis and EnvironmentalInteraction of Yield Traits in 323 Cotton Cultivars under 9Different EnvironmentsYinhua Jia1., Xiwei Sun2., Junling Sun1., Zhaoe Pan1, Xiwen Wang1, Shoupu He1, Songhua Xiao3,
Weijun Shi4, Zhongli Zhou1, Baoyin Pang1, Liru Wang1, Jianguang Liu3, Jun Ma4, Xiongming Du1*,
Jun Zhu2*
1 Institute of Cotton Research of Chinese Academy of Agricultural Sciences (ICR, CAAS), State Key Laboratory of Cotton Biology, Key Laboratory of Cotton Genetic
Improvement, Ministry of Agriculture, Anyang, China, 2 Key Laboratory of Crop Germplasm Resource of Zhejiang Province, Zhejiang University, Hangzhou, China,
3 Institute of industrial Crops, Jiangsu Academy of Agricultural Sciences, Nanjing, China, 4 Economic Crop Research Institute, Xinjiang Academy of Agricultural Science,
Urumqi1, China
Abstract
Improving yield is a major objective for cotton breeding schemes, and lint yield and its three component traits (boll number,boll weight and lint percentage) are complex traits controlled by multiple genes and various environments. Associationmapping was performed to detect markers associated with these four traits using 651 simple sequence repeats (SSRs). Amixed linear model including epistasis and environmental interaction was used to screen the loci associated with these fouryield traits by 323 accessions of Gossypium hirsutum L. evaluated in nine different environments. 251 significant loci weredetected to be associated with lint yield and its three components, including 69 loci with individual effects and all involvedin epistasis interactions. These significant loci explain , 62.05% of the phenotypic variance (ranging from 49.06% , 72.29%for these four traits). It was indicated by high contribution of environmental interaction to the phenotypic variance for lintyield and boll numbers, that genetic effects of SSR loci were susceptible to environment factors. Shared loci were alsoobserved among these four traits, which may be used for simultaneous improvement in cotton breeding for yield traits.Furthermore, consistent and elite loci were screened with 2Log10 (P-value) .8.0 based on predicted effects of loci detectedin different environments. There was one locus and 6 pairs of epistasis for lint yield, 4 loci and 10 epistasis for boll number,15 loci and 2 epistasis for boll weight, and 2 loci and 5 epistasis for lint percentage, respectively. These results providedinsights into the genetic basis of lint yield and its components and may be useful for marker-assisted breeding to improvecotton production.
Citation: Jia Y, Sun X, Sun J, Pan Z, Wang X, et al. (2014) Association Mapping for Epistasis and Environmental Interaction of Yield Traits in 323 Cotton Cultivarsunder 9 Different Environments. PLoS ONE 9(5): e95882. doi:10.1371/journal.pone.0095882
Editor: Baohong Zhang, East Carolina University, United States of America
Received October 31, 2013; Accepted March 31, 2014; Published May 8, 2014
Copyright: � 2014 Jia et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricteduse, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This work has been funded by the National Science and Technology Support Program of China (2006BAD13B04), and National Basic Research Programof China (2010CB12600, 2011CB109306). The funders had no rule in study design, data collection and analysis, decision to publish, or preparation of themanuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected] (JZ); [email protected] (XD)
. These authors contributed equally to this work.
Introduction
Cotton is one of the most important economic crops in the
world and provides large amounts of raw materials for textile
industry. High yield is always remained the primary focus of
cotton breeding programs. Boll number, boll weight, and lint
percentage are three yield component traits, which are complex
traits controlled by multiple genes with gene 6 gene interaction
and gene6environment interaction [1–2]. Understanding genetic
architecture of lint yield and its component traits at the molecular
level is very important for efficiently improving cotton yield traits.
Identifying and charactering the key QTLs and genes affecting
yield traits in cotton has been a research frontier over the past few
decades. On the basis of linkage analysis in the QTL mapping
populations, several QTLs controlling lint yield and its component
traits have been obtained [1–8]. However, these detected QTLs
involved in large regions of the chromosome, where candidate
genes or genetic variants are difficult to distinguish. In addition,
the mapping results have only limit utilization because of lower
genetic diversity in the special population used for QTL mapping
and can not fully elucidate the genetic basis of lint yield and its
component traits.
Genome-wide association studies (GWAS), which are based on
the linkage disequilibrium (LD), provide the opportunity to
systematically identify the genetic components of complex traits
of crops in recent years [9–14]. It has the potential to detect single
nucleotide polymorphisms (SNPs) or other types of molecular
markers such as single sequence repeats (SSRs) within or nearby a
gene. It can extensively exploit historical recombination and
natural genetic diversity in natural population, and overcome the
limitations of conventional linkage mapping for crop breeding.
Recently, Zhang et al performed association mapping using 81
PLOS ONE | www.plosone.org 1 May 2014 | Volume 9 | Issue 5 | e95882

accessions of Upland cotton based on 121 SSRs to map 12
agronomical and fiber quality traits, and identified 180 loci only
with additive effects significantly associated with the traits [14].
However, current GWAS predominately focus on examination of
SNPs at a single environment and fails to detect the gene 6 gene
interaction as well as gene 6 environment interaction across
multiple environments. The genetic variation of complex traits is
contributed in part by epistasis and environmental interaction
effects. Therefore, searching for only major effects may miss other
key genetic variants specific to environment factors and cannot
provide reliable estimates for genetic effects [3].
Due to a heavy computational burden, current methods of
GWAS have not been used for detecting gene 6 gene interaction
and gene 6 environment interaction for breeding populations in
multiple environments. In this study, we used mixed linear model
approach and a newly developed mapping software of QTXNet-work based on GPU parallel computation to identify individual
and epistasis loci and their environmental interactions for lint yield
and its component traits. We also identified candidate genes
related to markers of these target traits. These results and genetic
architecture identified in this study could provide more precise and
reliable information for marker-assisted selection in crop breeding.
Materials and Methods
Plant MaterialsMapping populations representing Upland cotton cultivars and
genetic resources were mainly obtained from China and some
from foreign countries including the United States of America,
Russia, Australia etc (Table S1), which are available from the
National Mid-term Genebank of the Institute of Cotton Research,
Chinese Academy of Agricultural Sciences (ICR-CAAS) after
signing the Material Transfer Agreement (MTA). These cultivars
are suitable for association mapping since they have abundant
genetic resources. 323 sampled cultivars of Upland cotton were
planted and evaluated for lint yield and yield components during
2007 to 2009 in three locations: 1) Anyang of Henan province
(annual frost free period = 180–230 days, average active accumu-
lated temperature = 3800–4900uC, annual rainfall = 500–
1000 mm) in the Yellow River valley cotton growing region; 2)
Kuche of Xinjiang province (annual frost free period = 170–230
days, average active accumulated temperature = 3000–5400uC,
annual rainfall = 15–380 mm) in the northwestward cotton
growing region; 3) Nanjing of Jiangsu province (annual frost free
period = 227–278 days, average active accumulated tempera-
ture = 3500–5500uC, annual rainfall = 1000–1600 mm) in the
Yangtze River valley cotton growing region. This research is
regular cotton regional trial test conducted in farm field of cotton
research institute, and not related with protected area of land and
protection of wildlife. It does not need specific official permission.
We regarded combination of each location and each year as an
individual environment, setting a total of nine environments. A
randomized complete block design with three replications was
employed in the filed trials, and each block was settled with two
rows and every row was kept in a plot with 5 m long and 0.7 m
wide. The field management utilized conventional field production
management techniques adjusted to local practice. Lint yield and
three yield components were evaluated: number of bolls per plant,
boll weight, and lint percentage.
DNA Extraction and Marker AnalysisThe genomic DNA was extracted from fresh, young leaves of
the 323 cultivars using CTAB method. SSR primers sequences
including BNL, JESPR, TMB, CIR, HAU, DPL, GH, NAU,
MGHES etc. can be available from Cotton Microsatellite
Database (CMD, http://www.cottonssr.org). We selected 20
cultivars of large morphological variations from the research
natural population to screen for polymorphisms primers. In
total, 5,600 SSR primers were chosen to survey the polymor-
phisms of the 323 cultivars. Only 651 polymorphisms SSR
alleles were detected, which were used for association mapping
(Table S2).
Genetic Models and Statistical MethodsAssociation mapping were performed using the mixed linear
model, including environment (e) as fixed effects, SSR loci effects
(a, aa) and loci by environment interaction (ae, aae) as random
effects. The genetic model for phenotypic value of the k-th
genotypes in the h-th environment (yhk ) can be expressed by the
following mixed linear model,
yhk~mzehzX
i
aiuikzX
ivj
aaijuikujkzX
i
aehiuhik
zX
ivj
aaehijuhikuhjkzehk
ð1Þ
where m is the population mean; eh is the fixed effect of the h-th
environment; ai is the i-th additive effect with coefficient uik; aaij is
the i-th additive by j-th additive epistasis effect with coefficient
uikujk; aehi is the i-th additive by the h-th environment interaction
effect with coefficient uhik; aaehij is the aaij by the h-th
environment interaction effect with coefficient uhikuhjk; and ehk is
the random residual effect of the k-th breeding line in the h-th
environment.
The mixed linear model can be presented in matrix notation,
y~XbzUAeAzUAAeAAzUAEeAEzUAAEeAAEzee
~XbzX4
v~1
Uvevzee
*MVN(Xb,X4
v~1
s2vUvUT
v zIs2e )
ð2Þ
where y is an n|1 column vector of phenotypic values and n is the
sample size of observations; b is a column vector of u and
environments in the experiment; X is the known incidence matrix
relating to the fixed effects; Uv is the known coefficient matrix
relating to the v-th random vector ev; ee~MMVN(0, Is2
e ) is an n|1
column vector of residual effects.
The phenotypic variance Vp is considered as the sum of additive
variance VA, epistasis variance VAA, additive by environment
interaction variance VAE , epistasis by environment interaction
variance VAAE , and residual variance Ve.
Vp~VAzVAAzVAEzVAAEzVe ð3Þ
Heritability is defined as the relative contribution of genetic
variance to phenotypic variance with the following estimation
model:
h2GzGE~h2
Azh2AAzh2
AEzh2AAE
~X
h2az
Xh2
aazX
h2aez
Xh2
aae
ð4Þ
Detecting Epistasis and GE Effect of Yield Traits
PLOS ONE | www.plosone.org 2 May 2014 | Volume 9 | Issue 5 | e95882

where h2GzGE = total heritability; h2
A = heritability contributed by
sum of individual locus, h2AA = heritability contributed by sum of
pair-wise epistasis, h2AE = additive by environment interaction
heritability contributed by sum of individual additive by environ-
ment interaction effects, h2AAE = epistasis by environment interac-
tion heritability contributed by sum of pair-wise epistasis by
environment interaction effects; h2a = additive heritability of
individual locus, h2aa = epistasis heritability of pair-wise loci,
h2ae = heritability of additive by environment interaction effect,
h2aae = heritability of epistasis by environment interaction effect.
Association MappingA GPU parallel computing software QTXNetwork (http://
ibi.zju.edu.cn/software/QTXNetwork/) was used to dissect the
genetic architecture of lint yield and its component traits. There
were 651 SSR markers used for association mapping for yield
traits of 323 representative Upland cotton cultivars in nine
different environments. Significant SSR markers were screened
and the estimation of fixed effects (e) and prediction of random
effects (a, aa, ae, and aae) of loci were obtained.
Gene Locations Based on the Associated Markers byReference D Genome
Based on the experiment-wise type I error (aEW v0:05) setting
by QTXNetwork, the SSR loci associated with the fiber yield
traits were selected. The primer information was acquired from
cotton marker database (http://www.cottonmarker.org/cmd_
downloads/ssr_project_data/CMD_PRIMER_ALL.xls). Only
one SSR location for each marker on the genome D (ftp://ftp.
ncbi.nlm.nih.gov/genbank/genomes/Eukaryotes/plants/Gossypium_
raimondii/) was acquired by bowtie method which 2 bases not
matched was permitted (bowtie -a -v 2–fr cottonPD -f -1
primerForward.fasta -2 primerReverse.fasta –best –strata bow-
tieResult_PD.txt). The SSR motifs of the searched SSR sequences
on the D genome were scanned by MISA. The location between
SSR locus and the related gene was decided based the star location
of the motif on the genome.
Results
In total, there were 94 significant SSRs (14 loci with individual
effects and 93 loci involved in epistasis interactions) associated with
lint yield, 95 SSRs (28 individual loci) associated with boll number,
91 SSRs (21 individual loci) associated with boll weight, and 88
SSRs (19 individual loci) associated with lint percentage. All
identified loci associated with lint yield components were involved
in epistasis interactions.
As to the lint yield, the total heritability of genotype 6environment interaction effects (h2
GEo38.26%) was mainly con-
tributed due to epistasis 6 environment interaction
(h2AAEo34.05%), and was much larger than the total genotype
heritability (h2Go10.80%) (Table 1). We listed highly significant
genetic effects of loci (2Log10P.8.0 and h2GE.1.0%, h2
G.0.5%)
and consistent loci with environment interaction which were stable
in a particular ecological area across at least two years for lint yield
and three yield traits (Figure 1, Table 2–5). Among the loci
associated with lint yield (Table 2), 1 locus (NAU3011-2) and 6
pairs of epistasis loci were highly significant with environmental
interaction at Nanjing in 2007 and 2009. It was suggested that
improving lint yield could be expected by selecting major-allele
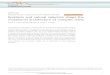
Figure 1. Network plot of highly significant loci detected for yield and yield components. Notes: The bottom axis is the SNP ID for QTS.Red dot = loci with additive effect, green dot = loci with additive 6 environment interaction effect, blue dot = loci with both additive andenvironment-specific effects, black dot with a line = loci with epistasis but no individual effect. Red line between two dot = aa epistasis, Green linebetween two dot = aae environment-specific epistasis.doi:10.1371/journal.pone.0095882.g001
Detecting Epistasis and GE Effect of Yield Traits
PLOS ONE | www.plosone.org 3 May 2014 | Volume 9 | Issue 5 | e95882

genotype QQ (NAU30112197) and QQ6QQ (MUCS1012
3306MGHES182228, NAU332522386TMB19892255,
NAU360822456HAU7732170), as well as minor-allele geno-
type qq6qq (HAU179423206TMB102375, NAU13622
2256NAU37742248, and NAU332522386HAU4232175) at
Nanjing (Table 2).
For boll number trait, the heritability of epistasis6environment
interaction effects (h2AAEo41.93%) was larger than other trait
heritability (Table 1), which indicated that boll number was mainly
controlled by environment-specific epistasis effects. 3 loci
(HAU1385-3, NAU1102-3, MGHES41-6) with additive heritabil-
ity were found to be more than 0.5%, and 10 pair epistasis loci
with environment-specific epistasis heritability showed more than
1.0% genetic effects (Figure 1, Table 3), indicating that the
improvement of boll number in all 9 environments could be
predictable (h2Ao1.83%) by selecting genotype QQ for
HAU13852150, and qq for other 2 loci (MGHES412333,
NAU11022241). Further improving boll number could be
expected (h2AE3
o11.31%) through selecting HAU10292197 6NAU11252243 in both Anyang and Nanjing areas and other 9
pair epistasis loci only in Nanjing area, among these 7 epistasis loci
with genotype of QQ6QQ showed positive effects, and 3 pair
epistasis loci with genotype of qq6qq showed negative effects. The
total heritability (h2GzGEo 62.85%) of boll weight was mostly due
to additive effects (h2Ao21.48%), epistasis effects (h2
AAo22.01%)
and environment-specific additive effects (h2AAEo18.73%). It was
suggested that most genetic effects were fairly stable across
environments (h2Go43.49%) for boll weight. There were 15 loci
with heritability of additive effect larger than 0.5% and 2 pair
epistasis loci with heritability of environment-specific epistasis
effects larger than 1.0% for boll weight (Figure 1, Table 4). With
regard to little impact of environmental factors on boll weight,
relevant robust loci could be selected for improvement of lint yield
across different ecological areas.
For lint percentage, the total heritability (h2GzGEo72.29%) was
mostly attributed to epistasis effects (h2AAo57.78%). Thus, it was
concluded that lint percentage was very stable across various
environments and mainly controlled by epistasis effects. Com-
pared with lint yield and other yield components, most genetic
effects of individual loci and epistasis loci explained a small
proportion of total phenotypic variance for lint percentage. Only 2
loci with additive effects and 5 pair epistasis loci with epistasis
effects had heritability larger than 0.5% (Figure 1 and Table 5). It
was apparent that lint percentage was mostly controlled by many
loci with smaller effects as compared with lint yield and other
component traits. In this population, further genetic gain could be
projected (h2AzAAo 6.3%) based on selecting 2 individual loci with
additive effects (NAU33252238, NAU35192200) and 5 pair loci
with epistasis effects for increasing lint percentage across different
environments.
It has been observed that some loci were associated with more
than one trait. 4 loci (DPL910-120, HAU639-175, NAU1155-205,
NAU40422175) were found to be associated with all of the four
traits, 22 loci were detected to be associated with three of the four
traits, and 61 loci were associated with two of the four traits.
Generally, these common loci made it possible for us to
simultaneously improve multiple traits via breeding programs.
However, 41 loci controlling lint yield were not detected in other
component traits, which suggested that lint yield, as the result of
complex biosynthesis pathways, may comprise some unknown
components affected by polygenes.
It had been observed that epistasis is the genetic base of lint
yield and yield components. In the results, most of the loci with
individual effects were involved in epistasis interactions for lint
yield and yield components. Altogether 293 digenic loci with
epistasis effect (aa) and/or epistasis 6 environment interaction
effect (aae) were identified to be associated with lint yield. There
were 193 digenic epistasis interactions with their total heritability
(h2AAzAAEo 30.71%) of epistasis effects and environment-specific
epistasis effects involved in the loci without individual effects. We
also observed this phenomenon in yield components. It was shown
that many loci, even without controlling yield and yield
components on their own, could affect the traits in combination
with other loci.
It was observed that many loci could interact with several other
loci for controlling phenotypic variation of lint yield and its
component traits. For example, NAU2126-195 could affect lint
yield in combination with other 4 loci (HAU7732170, NAU3377-
180, TMB102375, GH501-265), respectively. Magnitude of
epistasis effects of them differed from each other. NAU2126-195
may be regarded as core locus of epistasis interaction jointly with
other casual genes for controlling target traits. This phenomenon
might indicate that higher order interaction could also exist for
association with lint yield and its components.
Discussion
Association mapping is a powerful method to detect loci
underlying complex traits and more efficient compared to linkage
analysis, because it exploited abundant genetic variation in diverse
genetic backgrounds [15]. However, in most of the current
GWAS, the epistasis and environmental interaction effects were
not detectable due to absence of appropriate statistical methods
and a heavy computational burden [16–18]. The models without
epistasis and environmental interaction for GWAS may cause
some problems: 1) It may result in biased estimation of effects on
loci and decrease the precision and power of loci detection [18–
Table 1. Estimated heritability for lint yield and yield components.
Trait h2A(%) h2
AA(%) h2AE (%) h2
AAE (%) h2GzGE (%)
Lint Yield 0.20 10.6 4.21 34.05 49.06
Boll Number 3.87 9.28 8.89 41.93 63.98
Boll Weight 21.48 22.01 0.63 18.73 62.85
Lint Percentage 6.58 57.78 0.29 7.64 72.29
Note: h2A = heritability for additive effects, h2
AA = heritability for additive epistasis effects, h2AE = heritability for additive by environment interaction effects,
h2AAE = heritability for additive epistasis by environment effects, h2
GzGE = total heritability.
doi:10.1371/journal.pone.0095882.t001
Detecting Epistasis and GE Effect of Yield Traits
PLOS ONE | www.plosone.org 4 May 2014 | Volume 9 | Issue 5 | e95882

19]; 2) The heritability of complex traits would be missed in the
condition of reduced models [20–21].
Our proposed methodology is based on mixed linear model
including epistasis and environmental interaction, but it can also
be used for reduced models ignoring epistasis and environmental
interaction. We also performed GWAS of various reduced models
for lint yield and yield components. As compared with the full
model (Table 1), the total heritability of lint yield and yield
components decreased in the additive model (h2Go18.35% for lint
yield, 6.53% for boll number, 28.65% for boll weight, and 22.95%
for lint percentage), and also in the additive with environmental
interaction model (h2GzGEo17.31% for lint yield, 23.76% for boll
number, 32.25% for boll weight, and 25.08% for lint percentage).
The results also showed steep decrease in numbers and effects of
loci associated lint yield and yield components between reduced
models and full model. This may explain the reasons for missing
heritability reported recently [21].
Because of low LD associated with higher frequencies of cross-
pollination, cotton species are amenable for association mapping
of agronomic traits with a relatively large numbers of markers.
Previous studies involving nearly 1000 polymorphic markers
would ensure the coverage of whole cotton genome [14]. A total
of 651 SSR markers and 323 Upland cotton cultivars under 9
environments in this study can significantly improve the power of
association mapping. However, with the rapid development of
next generation sequencing, a substantial number of single
nucleotide polymorphisms (SNPs) will facilitate the accurate and
fine association analysis.
It is shown by the results of GWAS that lint yield, boll number
and lint percentage are inherited in a mainly epistasis manner,
whereas boll weight is predominately controlled by both additive
and epistasis effects. In addition, lint yield and boll number are
particularly susceptible to a broad range of environments, as
compared to boll weight and lint percentage, which has also been
observed in previous studies [5,22–23]. Both lint yield and
component traits were complex traits controlled by many genes
and gene-network cascades interacting with each other and also
with the environment factors (Figure 1). Many loci (ten or more in
the present study) associated with lint yield, boll number, and boll
weight showed much larger effects and heritability (additive or
epistasis effects), while few loci associated with lint percentage
could have large effects. This was indicated that improving cotton
lint yield could be difficult through increasing lint percentage. All
these information obtained from GWAS could provide useful
reference for MAS in cotton breeding programs.
Although the effects of many loci appeared to be consistent
across various environments, the magnitude of effects and even
direction of loci may vary depending on environmental factors
because of interactions between loci and environment [3,18,24].
We can classify loci identified in the present study into three types.
The first category of loci is called constituted loci with large major
effects (a, aa) and can be consistently detected across different
environments. This type of loci is very useful for improving cotton
traits across different ecological locations. Another type of loci is
called environment-specific loci with only environment-special
effects identified in a special ecological region but stable in
different years. For example, TMB3122212 6 GH132-180 was
stably found in the Northwestward region and NAU30132245 6NAU51632216 was stably scanned in the Yangtze River valley
region. This type of loci could be used for maker-assisted breeding
in a particular ecological area. The third type of loci is called
environment-sensitive loci that could not be consistently detected
in various grown regions. This kind of loci could only be used in
specific conditions.
SSRs widely used in major crop were highly reliable,
polymorphic, simple and cheap. The SSRs associated with nearby
genes were extremely useful for MAS. In the present study, we
successfully detected 251 significant loci associated with lint yield
and its three components, including 69 loci with individual effects.
Most of these significant loci generally had small impacts on cotton
Table 2. Predicted genetic effects with significance and heritability of lint yield for cotton regional trials in three locations andthree years.
Locus Effect Predict 2Log10P h2 (%) Candidate Gene
NAU30112197 ae3y1 1.01 11.32 1.04 IPR000679
ae3y3 1.56 25.93
HAU179423206TMB102375 aae3y1 21.58 26.17 0.96 – 6Gorai.004G126800
aae3y3 20.99 10.82
MUCS10123306MGHES182228 aae3y1 0.98 10.97 0.65 Cotton_A_293946IPR001128
aae3y3 1.06 12.52
NAU136222256NAU37742248 aae3y1 20.84 8.05 0.79 IPR0017646IPR002913
aae3y2 21.15 14.61
aae3y3 21.04 12.03
NAU332522386HAU4232175 aae3y1 21.03 11.93 0.72 IPR0007786IPR001461
aae3y3 21.25 17.12
NAU332522386TMB19892255 aae3y1 1.44 21.74 1.00 IPR0007786Cotton_A_24169
aae3y3 1.15 14.23
NAU360822456HAU7732170 aae3y1 1.10 13.26 0.73 IPR0007196IPR007087
aae3y3 0.89 8.99
Note: ae3y1 = additive by environment interaction effect at Nanjing in 2007; ae3y3 = additive by environment interaction effect at Nanjing in 2009; aae3y1 = epistasis byenvironment interaction effect at Nanjing in 2007; aae3y2 = epistasis by environment interaction effect at Nanjing in 2008; aae3y3 = epistasis by environment interactioneffect at Nanjing in 2009; 2Log10P = minus log10 (P-value), h2 (%) = heritability (%).doi:10.1371/journal.pone.0095882.t002
Detecting Epistasis and GE Effect of Yield Traits
PLOS ONE | www.plosone.org 5 May 2014 | Volume 9 | Issue 5 | e95882

breeding due to small effects and easily affected by environment
[23]. In order to develop effective MAS schemes, reliable and elite
loci associated with lint yield and yield components have been
screened. These loci, which consist of constituted loci and
environment-specific loci, are reliable to predict corresponding
phenotype. Constituted loci (such as NAU35882315 and
NAU33252238) can be applied for MAS in various cotton-
growing region, whereas application of environment-specific loci
(examples are NAU11022241 and NAU13622225 6NAU37742248) are limited to specific cotton-growing region.
Both of them have substantial potential to improve the efficiency
and precision of conventional cotton breeding.
With the information from Cotton Microsatellite Database
(CMD) and published report, several markers identified in our
present study had been located within or nearby the reported
QTL in previous linkage analysis and association mapping [1–
8,14]. For example, Zhang et al [14] independently detected
QTLs called qLP-A1-1 (between JESPR101 and BNL3590) using
linkage method and a locus called JESPR101 using association
method, which affected lint percentage. We also identified this
marker (2Log10P-value = 15.09) associated with the same
(Table 5). In addition, most of these consistent loci were involved
in interaction in our present results (examples are NAU54332330
for numbers of boll in Table 3). It indicated that these loci were
stably transferred and can be further used in MAS.
The traditional QTL was mainly located between two markers
with the scale of cM (centi-Mogan). Based on the physical
mapping of reference genome, the fine mapping of the markers
and the genes related to the fiber yield traits become possible. In
this study, candidate gene locations of yield fiber traits were
identified based on the associated SSR markers analyzed by the
QTXNetwork. For the lint yield, one major-allele genotype QQ
(NAU30112197) related to zinc finger (GATA-type) gene
(IPR000679) was identified. 3 epistasis loci with positive effects
(QQ6QQ genotype) could be related to the interactions with the
genes of cytochrome P450 (IPR001128), Cytochrome b245
(IPR000778), zinc finger with C2H2-type (IPR007087), and
protein kinase (IPR000719) etc. 3 minor-allele with negative
effects (qq6qq genotype) was linked to the interactions with the
genes of glycoside hydrolase (IPR001764), lipid-binding START
Table 3. Predicted genetic effects with significance and heritability of boll number for cotton regional trials in three locations andthree years.
Locus Effect Predict 2Log10P h2 (%) Candidate Gene
HAU13852150 a 0.3 42.29 0.68 IPR003245
MGHES412330 a 20.27 34.72 0.55 IPR000297
NAU11022241 a 20.28 37.61 0.6 IPR003329
ae3y1 20.38 8.35 0.63
ae3y2 20.46 11.99
ae3y3 20.52 14.8
GH11122456NAU8742215 aae3y1 0.37 8.25 0.37 – 6IPR001128
aae3y3 0.39 8.93
GH11122456NAU54332330 aae3y1 0.55 16.75 0.96 – 6IPR001865
aae3y3 0.84 37.43
HAU102921976NAU11252243 aae1y2 0.47 12.16 1.59 IPR0010126IPR016196
aae1y3 0.55 16.85
aae3y1 0.93 46.13
aae3y3 0.63 21.32
HAU196923756JESPR422128 aae3y1 1.08 60.86 2.43 IPR0001816Gorai.009G322400
aae3y3 1.27 83.72
HAU163923206NAU50992233 aae3y1 0.38 8.55 0.53 IPR0002176IPR001214
aae3y3 0.5 14.18
NAU271521846JESPR422128 aae3y2 20.63 22.12 0.65 IPR0040226Gorai.009G322400
aae3y3 20.38 8.43
NAU287323526NAU54332330 aae3y1 20.50 13.93 0.77 IPR0033116IPR001865
aae3y3 20.77 32.56
NAU404221756GH1112245 aae3y1 0.47 12.53 0.64 Cotton_A_040136–
aae3y3 0.68 25.62
STV61-1316NAU33052155 aae3y1 20.91 43.35 1.49 DUF16856IPR002913
aae3y2 20.65 22.48
TMB118122206NAU35632149 aae1y1 0.6 19.26 1.25 Gorai.002G0885006IPR003311
aae1y2 0.42 9.91
Note: a = additive effect; ae3y1, ae3y3, aae3y1, aae3y2, aae3y3 as defined in Table 2; ae3y2 = additive by environment interaction effect at Nanjing in 2008,aae1y1 = epistasis by environment interaction effect at Anyang in 2007, aae1y2 = epistasis by environment interaction effect at Anyang in 2008, aae1y3 = epistasis byenvironment interaction effect at Anyang in 2009; 2Log10P = minus log10 (P-value), h2 (%) = heritability (%).doi:10.1371/journal.pone.0095882.t003
Detecting Epistasis and GE Effect of Yield Traits
PLOS ONE | www.plosone.org 6 May 2014 | Volume 9 | Issue 5 | e95882

(IPR002913), and peptidase aspartic (IPR021109) at Nanjing areas
(Table 2). It seems that the genes catalyzed the oxidation of
organic substances will have positive effects on cotton lint yield,
but the decompose genes of glycoside, lipid and protein maybe
have negative effects for lint yield.
For boll number, there were 3 additive SSR loci (HAU1385-
150, NAU11022241, MGHES412333) related to genes of
peptidyl-prolyl cis-trans isomerases (PPIase, IPR000297), plasto-
cyanin-like (IPR003245), and acylneuraminate cytidylyltransferase
(IPR003329). 7 epistasis loci with positive effects for boll number
(QQ6QQ genotype) might be related to gene interactions of
cytochrome P450 (IPR001128), UBX domain-containing protein
(IPR001012), tubulin (IPR000217), ribosomal protein
(IPR001865), major facilitator superfamily (IPR016196), formyl-
methionine deformylase (IPR000181), and SET domain of
SUVH4 (IPR001214) etc. There were 3 epistasis loci with
negative effects (qq6qq genotype) related to gene interactions of
DDT domain superfamily (IPR004022), AUX/IAA protein
(IPR003311), ribosomal protein (IPR001865), protein of unknown
function (DUF1685), lipid-binding START (IPR002913) etc. It
seems that the gene interactions for the metabolism and transport
of cysteine, prolyl and formylmethionine could increase the boll
number. But the regulation of the gene interactions of the lipid-
binding DNA binding and auxin-binding protein could also affect
the boll number.
Table 4. Predicted genetic effects with significance and heritability of boll weight for cotton regional trials in three locations andthree years.
Locus Effect Predict 2Log10P h2(%) Candidate Gene
NAU37442200 a 20.07 66.02 1.29 IPR020828
TMB3122212 a 20.06 54.75 1.07 IPR011335
TMB12962230 a 0.07 60.15 1.18 IPR002591
BNL1313-180 a 0.08 95.32 1.89 –
NAU35882315 a 20.15 299.91 6.14 –
GH1322180 a 20.06 47.17 0.92 IPR001611
JESPR2742129 a 20.06 43.33 0.84 –
JESPR1012122 a 0.05 42.82 0.83 IPR004813
NAU33052155 a 20.05 42.63 0.83 IPR002913
NAU50992280 a 0.05 42.79 0.83 IPR001214
TMB19632243 a 20.05 37.99 0.73 –
HAU13852155 a 0.05 36.04 0.7 IPR003245
BNL30332175 a 20.05 33.68 0.65 IPR008540
NAU29312250 a 20.04 27.91 0.53 IPR001841
NAU1163-160 a 20.04 26.63 0.51 –
TMB31222126GH1322180 aae2y1 20.08 11.95 0.35 IPR0113356IPR001611
aae2y2 20.07 8.79
NAU301322456NAU51632216 aae3y1 20.09 12.53 0.66 Cotton_A_345106IPR002917
aae3y3 20.08 10.01
Note: a = additive effect, aa = additive by additive epistasis effect; aae2y1 = epistasis by environment interaction effect at Kuche in 2007; aae2y2 = epistasis byenvironment interaction effect at Kuche in 2008; aae3y1 = epistasis by environment interaction effect at Nanjing in 2007. aae3y3 = epistasis by environment interactioneffect at Nanjing in 2009; 2Log10P = minus log10 (P-value), h2 (%) = heritability (%).doi:10.1371/journal.pone.0095882.t004
Table 5. Predicted genetic effects with significance and heritability of lint percentage for cotton regional trials in three locationsand three years.
Loci Effect Predict 2Log10P h2 (%) Candidate Gene
NAU33252238 a 20.44 164.11 2.47 IPR000778
NAU35192200 a 20.23 46 0.67 IPR000719
NAU351922206 TMB12682157 aa 0.29 71.31 1.04 IPR0007196 IPR002048
HAU118521746TMB17912217 aa 20.21 38.63 0.55 IPR000194 6IPR000209
TMB163821896DPL5132320 aa 0.21 37.87 0.54 IPR002109 6–
NAU311022926NAU28622248 aa 20.20 36.04 0.52 IPR003329 6IPR001841
JESPR10121226BNL12312228 aa 20.20 35.61 0.51 IPR004813 6 –
Note: Genetic effect: a = additive effect, aa = additive by additive epistasis effect; 2Log10P = minus log10 (P-value), h2 (%) = heritability (%).doi:10.1371/journal.pone.0095882.t005
Detecting Epistasis and GE Effect of Yield Traits
PLOS ONE | www.plosone.org 7 May 2014 | Volume 9 | Issue 5 | e95882

For boll weight, there were 15 additive SSR loci with positive
effects relate to following genes: Type I phosphodiesterase
(IPR002591), SET domain (IPR001214) of SUVH4, plastocya-
nin-like (IPR003245), oligopeptide transporter (OPT) superfamily
(IPR004813). While the genes could have negative effects, such as
glyceraldehyde 3-phosphate dehydrogenase (IPR020828), restric-
tion endonuclease (IPR011335), leucine-rich repeat (IPR001611),
STAR-related lipid-transfer (START) domain (IPR002913), bras-
sinosteroid signalling positive regulator (BZR1) family protei-
n(IPR008540), and RING-type zinc finger (IPR001841), respec-
tively. Except the main additive effects, there are also 2 epistasis
loci with negative effects (qq6qq genotype) which maybe resulted
from the interactions of leucine-rich repeat (IPR001611), GTP-
binding protein (IPR002917) etc. It was inferred that boll weight is
mainly controlled by the genes with the phosphorus, lipid and
signalling transport, which may be further regulated by the zinc
finger, leucine-rich repeat, and the methylation protein.
For lint percentage, 2 SSR loci with negative additive effects
could be related to protein kinase (IPR000719) and cytochrome
b245 (IPR000778), respectively. There were 2 epistasis loci with
positive effects (QQ6QQ genotype) related to the interactions of the
genes of protein kinase (IPR000719), calcium-binding EF-hand
(IPR002048), glutaredoxin (IPR002109). Three epistasis loci with
negative effects (qq6qq genotype) relate to the interactions of the
genes of peptidase S8/S53 (IPR000209), zinc finger with RING-
type (IPR001841), and oligopeptide transporter OPT superfamily
(IPR004813).
Supporting Information
Table S1 Variety germplasm of Upland cotton used inassociation mapping for fiber traits. Note: New lines were
new breeding lines and genetic stocks bred by Institute of Cotton
Research of Chinese Academy of Agricultural Sciences (ICR,
CAAS) in recent years.
(DOCX)
Table S2 651 primer alleles that were polymorphic inthe tested population.
(XLS)
Author Contributions
Conceived and designed the experiments: XMD. Performed the
experiments: YHJ XWS JLS ZEP XWW SPH SHX WJS ZLZ BYP
LRW JGL JM. Analyzed the data: JZ. Contributed reagents/materials/
analysis tools: XMD JZ. Wrote the paper: XWS JZ XMD.
References
1. He DH, Lin ZX, Zhang XL, Nie YC, Guo XP, et al. (2005) Mapping QTLs of
traits contributing to yield and analysis of genetic effects in tetraploid cotton.Euphytica 144: 141–149.
2. Shen XL, Zhang TZ, Guo WZ, Zhu XF, Zhang XY (2006) Mapping Fiber andYield QTLs with Main, Epistatic, and QTL 6Environment Interaction Effects
in Recombinant Inbred Lines of Upland Cotton. Crop Science 46: 61.3. Murcray CE, Lewinger JP, Gauderman WJ (2008) Gene-Environment
Interaction in Genome-Wide Association Studies. American Journal of
Epidemiology 169: 219–226.4. Chen H, Qian N, Guo WZ, Song QP, Li BC, et al. (2010) Using three selected
overlapping RILs to fine-map the yield component QTL on Chro.D8 in Uplandcotton. Euphytica 176: 321–329.
5. Liu RZ, Wang BH, Guo WZ, Qin YS, Wang LG, et al. (2011a) Quantitative
trait loci mapping for yield and its components by using two immortalizedpopulations of a heterotic hybrid in Gossypium hirsutum L. Mol Breeding 29:
297–311.6. Yadava SK, Arumugam N, Mukhopadhyay A, Sodhi YS, Gupta V, et al. (2012)
QTL mapping of yield-associated traits in Brassica juncea: meta-analysis andepistatic interactions using two different crosses between east European and
Indian gene pool lines. Theor Appl Genet 125: 1553–1564.
7. Yu JW, Zhang K, Li SY, Yu SX, Zhai HH, et al. (2012) Mapping quantitativetrait loci for lint yield and fiber quality across environments in a Gossypium
hirsutum 6 Gossypium barbadense backcross inbred line population. TheorAppl Genet 126: 275–287.
8. Lacape JM, Gawrysiak G, Cao TV, Viot C, Llewellyn D, et al. (2013) Mapping
QTLs for traits related to phenology, morphology and yield components in aninter-specific Gossypium hirsutum6G. barbadense cotton RIL population. Field
Crops Research 144: 256–267.9. Huang XH, Wei XH, Sang T, Zhao Q, Feng Q, et al. (2010) Genome-wide
association studies of 14 agronomic traits in rice landraces. Nature Genetics 42:961–967.
10. Poland JA, Bradbury PJ, Buckler ES, Nelson RJ (2011) Genome-wide nested
association mapping of quantitative resistance to northern leaf blight in maize.Proc Natl Acad Sci USA 108: 6893–6898.
11. Kump KL, Bradbury PJ, Wisser RJ, Buckler ES, Belcher AR, et al. (2011)Genome-wide association study of quantitative resistance to southern leaf blight
in the maize nested association mapping population. Nature Genetics 43: 163–
168.
12. Tian F, Bradbury PJ, Brown PJ, Hung H, Sun Q, et al. (2011) Genome-wide
association study of leaf architecture in the maize nested association mapping
population. Nature Genetics 43: 159–162.
13. Li H, Peng ZY, Yang XH, Wang WD, Fu JJ, et al. (2012) Genome-wide
association study dissects the genetic architecture of oil biosynthesis in maize
kernels. Nature Genetics: 1–10.
14. Zhang TZ, Qian N, Zhu XF, Chen H, Wang S, et al. (2013) Variations and
Transmission of QTL Alleles for Yield and Fiber Qualities in Upland Cotton
Cultivars Developed in China. PLoS ONE 8: e57220.
15. Ziegler A, Konig IR, Thompson JR (2008) Biostatistical Aspects of Genome-
Wide Association Studies. Biom J 50: 8–28.
16. Carlborg O, Haley CS (2004) Epistasis: too often neglected in complex trait
studies? Nat Rev Genet 5: 618–625.
17. Phillips PC (2008) Epistasis – the essential role of gene interactions in the
structure and evolution of genetic systems. Nature Publishing Group 9: 855–867.
18. van Os J, Rutten BPF (2009) Gene-environment-wide interaction studies in
psychiatry. Am J Psychiatry 166: 964–966.
19. Culverhouse R, Suarez BK, Lin J, Reich T (2002) A perspective on epistasis:
limits of models displaying no main effect. Am J Hum Genet 70: 461–471.
20. Brachi B, Morris GP, Borevitz JO (2011) Genome-wide association studies in
plants: the missing heritability is in the field. Genome Biol 12: 232.
21. Zuk O, Hechter E, Sunyaev SR, Lander ES (2012) The mystery of missing
heritability: genetic interactions create phantom heritability. Proc Natl Acad Sci
USA 109: 1193–1198.
22. Godoy S, Palomo G (1999) Genetic analysis of earliness in upland cotton. II.
Yield and fiber properties. Euphytica 105: 161–166.
23. Campbell BT, Jones MA (2005) Assessment of genotype 6 environment
interactions for yield and fiber quality in cotton performance trials. Euphytica
144: 69–78.
24. Collard BCY, Mackill DJ (2008) Marker-assisted selection: an approach for
precision plant breeding in the twenty-first century. Philosophical Transactions
of the Royal Society B: Biological Sciences 363: 557–572.
Detecting Epistasis and GE Effect of Yield Traits
PLOS ONE | www.plosone.org 8 May 2014 | Volume 9 | Issue 5 | e95882

1
Table S1. Variety germplasm of Upland cotton used in association mapping for fiber traits Serial No. Reg. No. Germplasm Name Geographical Origin
V9 110313 HuaiMian4Hao Anhui, China V17 110644 WanJiu828 Anhui, China V62 110737 XiaoXianDaLing Anhui, China V324 112240 SuMian9108 Anhui, China V266 112572 WangJiangChangRongMian Anhui, China V152 110761 86-1 (72-100) BeiJing, China V132 111550 ZhongZhiBD13 BeiJing, China V133 111552 ZhongZhiBD27 BeiJing, China V134 111553 ZhongZhiBD89 BeiJing, China V210 112795 GK20 BeiJing, China V116 112796 GK22 BeiJing, China V85 111027 NaShangQuDaHua Guizhou, China V55 110070 JiHan2HaoXuanXi Hebei, China V4 110083 Bao6722 Hebei, China V284 110097 353DaLingDi3Xi Hebei, China V278 110730 HanDanChangRong Hebei, China V286 110733 ChengDingDaLing Hebei, China V3 110810 Bao6716 Hebei, China V41 110861 Jimian11 (CK) Hebei, China V86 110862 JiMian12Hao (Handan177) Hebei, China V93 110938 XingTai6 Hebei, China V227 111018 HanDan568 Hebei, China V96 111081 XingTai79-11 Hebei, China V21 111346 Han8901 Hebei, China V123 111373 Han8944 Hebei, China V22 111381 Han8959 Hebei, China V326 112019 JiYuan12-13 Hebei, China V260 112022 JiYuan55 (91Han14) Hebei, China V279 112023 JiHaiLu6 (91Han6) Hebei, China V24 112043 Ji91-31 Hebei, China V252 112046 Ji91-33 Hebei, China V237 112123 Ji85-3 Hebei, China V235 112126 Ji91-12 Hebei, China V301 112128 Ji91-18 Hebei, China V231 112132 Ji91-22 Hebei, China V238 112135 Ji91-32 Hebei, China V194 112144 JiA-1-7 (33Xi) Hebei, China V120 112145 JiA-7-8 (33Xi) Hebei, China

2
V45 112194 ShiYuan638 Hebei, China V257 112233 JiZi123 (Jimian25) Hebei, China V117 112714 Han109 Hebei, China V247 112714 HanDan109 Hebei, China V268 112733 Ji668 Hebei, China V270 112751 HanDan333 Hebei, China V217 112773 SGKShiXuan321 Hebei, China V243 113561 GK99-1 Hebei, China V81 114008 JiFeng197 Hebei, China V5 110033 ZhangDe184 Henan, China V144 110323 ZhongYuan9114 Henan, China V276 110477 ZhengZhouZhangRongMian Henan, China V90 110508 Yu284 Henan, China V149 110544 Zhong521 Henan, China V80 110722 DaTaoMian Henan, China V58 110728 NeiHuangDaZi Henan, China V68 110729 MeiFuDaLing Henan, China V295 110839 ZhongMianSuo12 Henan, China V103 110855 Zhong2108 Henan, China V175 110856 ZhongMianSuo17Hao (Zhong117) Henan, China V176 110919 ZhongMianSuo19Hao (Zhong7886) Henan, China V92 110921 Zhong85271 Henan, China V293 110959 XinXiang89S-210 Henan, China V70 110961 XinDaLing Henan, China V203 111006 Zhong1276 Henan, China V229 111037 ZhongZi9103 Henan, China V135 111117 ZhongZi640 Henan, China V226 111180 Zhong932906 Henan, China V91 111184 Zhong5913-2 Henan, China V228 111210 ZhongZi9102 Henan, China V19 111225 ChangRongMuZiMian Henan, China V163 111336 Zhong89-1 Henan, China V261 111395 RTBaiXu Henan, China V143 111421 ZhongYuan911 Henan, China V150 111422 ZhongYuan9112 Henan, China V146 111424 ZhongYuan9116 Henan, China V145 111425 ZhongYuan9115 Henan, China V147 111431 ZhongYuanHAS-1 Henan, China V259 111434 ZhongYuanHST1 Henan, China V290 111895 ZhongMianSuo16 Henan, China V211 111900 YuMian9Hao (Yuzao1109) Henan, China V251 111944 Zhong07 Henan, China

3
V130 111950 Zhong870203 Henan, China V131 111961 Zhong961716 Henan, China V151 112196 YuMian2067 Henan, China V212 112241 ZhongMianSuo27 Henan, China V215 112247 ZhongMianSuo35 Henan, China V265 112570 YuMian19 Henan, China V292 112686 ZhongMianSuo41 (SGK9708) Henan, China V104 112690 ZhongJi926 Henan, China V206 112698 ZhongR03 Henan, China V287 112726 XinYan96-48 Henan, China V118 112746 ZhongMianSuo32 Henan, China V124 112782 KangHuangWei164 Henan, China V289 112798 ZhongZi9196 (Jiguang) Henan, China V306 112799 Zhong2220 Henan, China V102 113229 ZhengKang01-505 Henan, China V299 113693 ZhongMianSuo50 Henan, China V61 113697 ZhongZi4480 Henan, China V298 113939 ZhongMianSuo49 Henan, China V172 113943 ZhongZi04184 Henan, China V321 114019 Zhong2201 Henan, China V325 114152 ZhongZi10Hao Henan, China V312 114483 ZhongG5 Henan, China V37 130293 ZaoShuChangRong7 Henan, China V106 131404 CZA(70)33 Henan, China V262 131624 MSCO-11 Henan, China V282 131625 MSCO-12 Henan, China V59 131676 NongJiu Henan, China V153 140135 ZiSeMeiMian Henan, China V288 140152 YuMian2Hao (YuWu302) Henan, China V38 140338 YaHuang9103 Henan, China V323 140444 LangHuangF10 (ZongXu) Henan, China V51 New lines ZhongARR40682 Henan, China V52 New lines ZhongARNnXu Henan, China V72 New lines ZhongR773-72 Henan, China V73 New lines ZhongARR40681 Henan, China V74 New lines ZhongARR40683 Henan, China V75 New lines ZhongR773-75 Henan, China V76 New lines ZhongArc-76 Henan, China V77 New lines ZhongAR683-77 Henan, China V78 New lines RTN78 Henan, China V79 New lines RTN79 Henan, China V105 New lines ZhongArc-105 Henan, China

4
V119 New lines ZhongAR40772 Henan, China V177 New lines Su08B2-177 Henan, China V185 New lines ZhongArc-185 Henan, China V187 New lines RNX187 Henan, China V188 New lines RNX188 Henan, China V189 New lines RNX189 Henan, China V190 New lines RNX190 Henan, China V202 New lines Yu17-202 Henan, China V204 New lines Zhong31-204 Henan, China V205 New lines Zhong4612YaH Henan, China V221 New lines Zhong9708-221 Henan, China V222 New lines Zhong9708-222 Henan, China V223 New lines TM1-IPR Henan, China V307 New lines Zhong507145 Henan, China V308 New lines ZhongArc-308 Henan, China V309 New lines ZhongR773-309 Henan, China V310 New lines ZhongR773-310 Henan, China V311 New lines ZhongAR-RNX311 Henan, China V313 New lines ZhongAR-RNX313 Henan, China V314 New lines ZhongR773-314 Henan, China V315 New lines ZhongArc-315 Henan, China V316 New lines ZhongAR681-316 Henan, China V317 New lines 99633 Henan, China V320 New lines Ari-320 Henan, China V322 New lines HongTao Henan, China V327 New lines Ari-327 Henan, China V329 New lines ZhongR773-329 Henan, China V6 110639 ChangRong67-12 Hubei, China V83 110746 HuBeiSongZiDaLing Hubei, China V71 110747 XinZhouDaLing Hubei, China V26 111664 Jing55263 Hubei, China V20 111709 E408 Hubei, China V42 112272 Jing55168 Hubei, China V23 112527 HuaZhong91-0102 Hubei, China V267 112705 EKangMian9Hao Hubei, China V35 110235 XiangMian2Hao Hunan, China V28 110743 LiXianDaTao Hunan, China V67 110744 DaLingMian69Hao Hunan, China V13 110286 ShaMaoChangTao Jiangsu, China V272 110381 601ChangRongMian Jiangsu, China V218 110614 SiMian2Hao Jiangsu, China V14 110646 SiChang2Xi Jiangsu, China

5
V69 110731 QiFengDaLing Jiangsu, China V56 110738 JiangSuDaTao Jiangsu, China V285 110740 XuShiDaTao Jiangsu, China V98 111022 XuZhouBanBanMian Jiangsu, China V277 111086 SuMian2Hao (Xuzhou553) Jiangsu, China V15 111398 Su7036YuanYuan Jiangsu, China V200 111401 SuYuan7235 Jiangsu, China V99 111409 YanCheng1115 Jiangsu, China V47 112189 SuXu137 (Zaoshu) Jiangsu, China V48 112190 SuXu138 (Zaoshu) Jiangsu, China V121 112581 JCG59 Jiangsu, China V122 112593 JCG94 Jiangsu, China V148 112697 ChangKangMian Jiangsu, China V258 112734 XuZhou261 Jiangsu, China V269 112734 XuZhou261 Jiangsu, China V95 112736 Si168 Jiangsu, China V97 112755 XuZhou244 Jiangsu, China V198 113189 SuQ1 Jiangsu, China V199 113190 SuTKH-1 Jiangsu, China V16 113368 SuNong6Hao Jiangsu, China V319 113675 SuYuan04-129 Jiangsu, China V49 113676 J02-247 Jiangsu, China V50 113677 J02-508 Jiangsu, China V318 113691 SuYuan04-162 Jiangsu, China V65 130452 DaLingFuZiMian Jiangsu, China V281 110600 GaoYiMian Jiangxi, China V7 110176 ZhaoYang70Hao Liaoning, China V154 111762 LiaoMian-5Hao Liaoning, China V18 111772 Liao61107 Liaoning, China V10 111783 Jin444 Liaoning, China V246 112337 Liao96-63-70 Liaoning, China V241 112341 Liao96-23-30 Liaoning, China V248 112372 Liao823-834-23 Liaoning, China V138 113790 LiaoMian16 Liaoning, China V139 113791 LiaoMian17 Liaoning, China V140 113792 LiaoMian18 Liaoning, China V296 113793 LiaoMian19 Liaoning, China V137 113811 Liao4853 Liaoning, China V136 113842 Liao4835 Liaoning, China V274 110131 DaZeMian Shandong, China V94 110486 ShanNong3Hao Shandong, China V36 110671 YangFen31Hao Shandong, China

6
V89 110942 LuMian11Hao (3389) Shandong, China V88 110947 LinQing201 Shandong, China V264 112552 Lu458 Shandong, China V82 112952 Lu21 Shandong, China V273 112952 LuMianYan21(Lu1138) Shandong, China V44 113219 ShanNong6Hao Shandong, China V195 113756 LuNong9648 Shandong, China V8 111620 Hu749513 Shanghai, China V201 110182 ShenMian5Hao ShanXi, China V127 111181 Shan3184 ShanXi, China V234 111274 Tai83158 ShanXi, China V236 111493 JinKang157 ShanXi, China V232 111495 Jin90Kang282 ShanXi, China V128 111937 Shan960329-2Yuan3 ShanXi, China V125 112106 Shan2747 ShanXi, China V126 112108 Shan2800 ShanXi, China V100 112295 Yun3060 ShanXi, China V129 112322 Yun92Kang124 ShanXi, China V242 112324 Yun93Kang354 ShanXi, China V244 112325 Yun93Kang393 ShanXi, China V101 112331 Yun92A-260 ShanXi, China V275 112550 QinYuan4Hao (Qinyuan91406) ShanXi, China V271 112897 QinLi514 ShanXi, China V141 112924 YongJi1Hao ShanXi, China V142 112925 YongJi2Hao ShanXi, China V60 New lines NangFenDaTao ShanXi, China V29 110014 MianYang73-39 Sichuan, China V30 110821 QingKang1Hao Sichuan, China V39 111593 ChuanGaoYiFen58 Sichuan, China V57 113374 KangSanXingDaTao Sichuan, China V27 111015 KuiTun80-2056W(XinLuZao-3Hao) Xijiang, China V328 111517 BaZhou7416 Xijiang, China V33 111525 Tu188 Xijiang, China V34 112227 Tu83-161 Xijiang, China V249 112387 KuChe96518 Xijiang, China V11 112432 KuCheT94-1 Xijiang, China V239 112435 KuCheT94-4 Xijiang, China V12 112437 KuCheT94-6 Xijiang, China V233 112439 KuChe93551 Xijiang, China V87 112596 KuChe96486 Xijiang, China V43 112647 KuiTunXi96-167 Xijiang, China V2 112889 BaZhou5628 Xijiang, China

7
V31 140182 Sha24-3 Xijiang, China V53 New lines S-050019 Xijiang, China V54 New lines S-050031 Xijiang, China V283 New lines 0102X-10-1 Xijiang, China V1 110452 70-29-5 Yunnan, China V66 110723 DaLingMian Zhaoyang, China V256 131391 Cook 310-5110 Australia V191 131628 Ao152 Australia V192 131632 AoC Australia V193 131643 AoL23/757 Australia V179 131063 LAPAR45 Brazil V183 130423 ZhaDeMian Chad V225 130440 ZhaDe3Hao Chad V186 130267 L142-9 Cote d'ivoire V182 130403 Europe 40 Europe V178 140246 HG-BR-8 Europe V180 130164 PAR-51 Pakistan V294 131368 M11 Pakistan V32 130541 USSR8908 Russia V46 131251 USSR21Xi (91-133) Russia V219 131292 USSR91Xi Russia V230 131309 USSR116Xi Russia V25 131509 AnJiYan6Hao Russia V181 130233 Sudan2 Sudan V220 130391 BPA68 Uganda V250 111530 Bu3363 US V209 112566 DP33B US V184 130237 Acala SJ-1 US V297 130238 AcalaSJ-1-9 US V173 130241 Acala SJ-4 US V300 130242 Acala SJ-5 US V216 130261 BELLSIRO US V291 130294 LineF US V168 130300 AC239 US V167 130543 USSR8911 US V169 130581 M-8124-1159 US V280 130721 PD6186 US V240 130797 FB20 US V171 130831 Mei8123 US V263 130836 Upland US V112 130838 Mei28114-313 US V166 130841 MeiD#1 US

8
V164 130866 Miscot78-27 US V109 130867 Miscot7803-52 US V174 130939 Arcot-1 US V157 130943 Arcot438 US V156 130944 Arcot436 US V155 130947 Arcot402bne US V158 130951 Coker139 US V107 130961 DES926 US V224 130974 GP67 US V84 130977 GP70 US V161 130989 GP83 US V108 130998 GP93 US V162 131000 GP95 US V159 131039 GP137 US V160 131040 GP138 US V110 131076 N73DeltapineNGF US V111 131081 N74-250 US V63 131164 Acala (DaLing)B US V208 131166 Acala 927 US V253 131395 DP2156 US V165 131408 MM-2 US V207 131439 UA887(You) US V213 131441 ST474 US V245 131613 DP410B US V170 131662 AoSiv2 US V113 131664 Qik US V196 131739 MeiF-18 US V115 131747 MeiG-84 US V197 131847 MeiF-19 US V114 131848 MeiG-82 US V330 New lines Acala1517-2 US V254 131468 r-3149 Vietnam V255 131470 r4136 Vietnam Note: New lines were new breeding lines and genetic stocks bred by Institute of Cotton Research of Chinese Academy of Agricultural Sciences (ICR, CAAS) in recent years.

Table S2. 651 primer alleles that were polymorphic in the tested population.Allell Name Allell Name Allell Name Allell Name Allell Name BNL1040-102 HAU1016-153 JESPR101-115 NAU1042-225 STV31-160BNL1040-80 HAU1016-160 JESPR101-122 NAU1042-242 STV31-185BNL1040-85 HAU1029-180 JESPR101-135 NAU1042-318 STV31-190BNL1040-89 HAU1029-197 JESPR101-145 NAU1042-352 STV31-190BNL119-215 HAU1168-250 JESPR114-101 NAU1043-220 STV33-150BNL119-230 HAU1168-300 JESPR114-110 NAU1043-230 STV33-160BNL119-242 HAU1168-315 JESPR158-120 NAU1048-240 STV33-165BNL119-251 HAU1168-375 JESPR158-124 NAU1048-265 STV33-169BNL1227-220 HAU1185-174 JESPR165-92 NAU1071-168 STV33-175BNL1231-200 HAU1185-178 JESPR165-99 NAU1071-180 STV33-180BNL1231-208 HAU1185-300 JESPR274-109 NAU1085-262 STV33-195BNL1231-215 HAU1185-310 JESPR274-117 NAU1085-275 STV67-110BNL1231-228 HAU1185-368 JESPR274-129 NAU1093-175 STV67-114BNL1317-175 HAU1384-270 JESPR274-157 NAU1093-177 STV67-118BNL1317-180 HAU1384-285 JESPR274-170 NAU1093-179 STV69-165BNL1317-192 HAU1384-290 JESPR274-198 NAU1102-230 STV69-90BNL1317-199 HAU1385-125 JESPR42-128 NAU1102-241 TMB10-330BNL1421-212 HAU1385-130 JESPR42-150 NAU1103-191 TMB1120-165BNL1421-248 HAU1385-150 JESPR50-155 NAU1103-208 TMB1120-178BNL1421-260 HAU1385-155 JESPR50-195 NAU1119-135 TMB1120-215BNL1694-235 HAU1413-125 MGHES18-190 NAU1119-140 TMB1120-328BNL1694-415 HAU1413-130 MGHES18-228 NAU1119-200 TMB1181-220BNL2449-120 HAU1413-150 MGHES18-248 NAU1119-235 TMB1181-242BNL2449-139 HAU1632-260 MGHES41-242 NAU1119-315 TMB1181-253BNL2449-156 HAU1632-270 MGHES41-280 NAU1119-370 TMB1181-256BNL2449-233 HAU1794-285 MGHES41-333 NAU1125-239 TMB1268-152BNL3033-175 HAU1794-300 MGHES41-335 NAU1125-243 TMB1268-154BNL3033-186 HAU1794-320 MGHES70-214 NAU1155-205 TMB1268-157BNL3383-172 HAU1794-330 MGHES70-222 NAU1155-260 TMB1296-230BNL3383-180 HAU1952-225 MUCS101-155 NAU1155-270 TMB1296-235BNL3383-190 HAU1952-235 MUCS101-330 NAU1167-191 TMB1296-240BNL3383-270 HAU1952-247 MUCS127-227 NAU1167-193 TMB1296-250BNL3646-156 HAU1952-250 MUCS127-234 NAU1167-196 TMB1638-154BNL3646-162 HAU1969-242 MUSS138-212 NAU1167-210 TMB1638-157BNL830-108 HAU1969-247 MUSS138-230 NAU1169-230 TMB1638-183BNL830-125 HAU1969-375 MUSS422-221 NAU1169-235 TMB1638-189CIR307-188 HAU2056-366 MUSS422-229 NAU1186-165 TMB1667-275CIR307-192 HAU2056-405 MUSS440-345 NAU1186-172 TMB1667-288CIR307-225 HAU2056-410 MUSS440-358 NAU1190-228 TMB1791-190CIR328-217 HAU2067-270 MUSS440-401 NAU1190-235 TMB1791-190CIR328-235 HAU2067-280 NAU1190-241 TMB1791-195CIR328-244 HAU2067-285 NAU1190-375 TMB1791-203CM45-140 HAU250-135 NAU1196-238 TMB1791-203CM45-150 HAU250-138 NAU1196-320 TMB1791-210CM45-168 HAU250-148 NAU1200-225 TMB1791-212CSHES128-176 HAU2511-201 NAU1200-235 TMB1791-217CSHES128-199 HAU2511-205 NAU1200-236 TMB1791-217CSHES128-215 HAU2543-370 NAU1200-242 TMB1791-220CSHES133-222 HAU2543-380 NAU1200-340 TMB1963-203CSHES133-235 HAU2543-418 NAU1200-368 TMB1963-207

CSHES3-165 HAU2543-421 NAU1225-242 TMB1963-218CSHES3-179 HAU2543-496 NAU1225-325 TMB1963-243CSHES51-178 HAU2631-315 NAU1225-370 TMB1963-248CSHES51-205 HAU2631-330 NAU1225-401 TMB1989-220CSHES62-215 HAU2631-345 NAU1225-410 TMB1989-235CSHES62-228 HAU2753-1100 NAU1255-225 TMB1989-255DPL101-183 HAU2753-300 NAU1255-243 TMB1989-265DPL101-195 HAU2753-540 NAU1255-435 TMB1989-266DPL101-207 HAU2753-700 NAU1255-458 TMB2945-246DPL196-100 HAU2753-820 NAU1269-145 TMB2945-269DPL196-105 HAU2753-900 NAU1269-345 TMB2945-288DPL196-130 HAU2759-200 NAU1302-205 TMB2945-320DPL196-135 HAU2759-215 NAU1302-213 TMB312-192DPL196-160 HAU2759-225 NAU1302-220 TMB312-205DPL196-95 HAU2759-235 NAU1322-180 TMB312-212DPL209-223 HAU2759-245 NAU1322-190 TMB312-232DPL209-245 HAU2759-253 NAU1322-202 TMB426-198DPL212-103 HAU2759-262 NAU1322-241 TMB426-202DPL212-99 HAU2786-220 NAU1362-225 TMB436-180DPL249-121 HAU2786-225 NAU1362-240 TMB436-191DPL249-132 HAU2786-230 NAU2026-178 TMB436-203DPL26-210 HAU2786-240 NAU2026-189 TMB436-228DPL26-215 HAU2792-135 NAU2083-163 TMB436-228DPL26-220 HAU2792-201 NAU2126-195 TMB436-232DPL26-240 HAU2792-230 NAU2265-375 TMB436-250DPL431-192 HAU2824-248 NAU2265-382 TMB799-150DPL431-250 HAU2824-275 NAU2272-163 TMB799-160DPL443-120 HAU2837-370 NAU2272-169 TMB799-175DPL443-125 HAU2837-390 NAU2274-105 TMB799-180DPL443-130 HAU2893-245 NAU2274-110 TMB799-185DPL461-165 HAU3061-258 NAU2277-60 TMB799-190DPL461-170 HAU3236-246 NAU2277-68DPL461-175 HAU3236-255 NAU2277-72DPL46-240 HAU3356-180 NAU2277-78DPL46-245 HAU3356-220 NAU2336-198DPL46-255 HAU3356-245 NAU2336-205DPL46-260 HAU3356-270 NAU2343-247DPL513-228 HAU3356-280 NAU2343-255DPL513-298 HAU423-165 NAU2437-240DPL513-320 HAU423-175 NAU2437-245DPL513-332 HAU639-175 NAU2437-252DPL715-128 HAU639-210 NAU2508-150DPL715-130 HAU639-220 NAU2508-156DPL715-145 HAU717-340 NAU2508-278DPL715-152 HAU717-350 NAU2631-193DPL715-161 HAU773-130 NAU2631-312DPL751-115 HAU773-140 NAU2679-218DPL752-178 HAU773-150 NAU2679-227DPL752-200 HAU773-160 NAU2715-182DPL752-205 HAU773-170 NAU2715-184DPL752-238 HAU878-113 NAU2715-198DPL799-138 HAU878-120 NAU2715-265

DPL910-120 HAU878-125 NAU2730-340DPL910-135 HAU878-130 NAU2730-345DPL910-150 HAU878-295 NAU2730-370DPL910-195 HAU878-302 NAU2730-410GH111-194 HAU880-178 NAU2741-250GH111-245 HAU880-180 NAU2741-252GH111-252 HAU880-190 NAU2741-282GH132-158 HAU880-210 NAU2776-260GH132-163 HAU940-145 NAU2776-263GH132-177 HAU940-150 NAU2820-158GH132-180 HAU940-170 NAU2820-163GH268-128 HAU940-175 NAU2820-178GH268-135 NAU2826-165GH354-130 NAU2826-180GH354-150 NAU2862-243GH354-165 NAU2862-248GH354-183 NAU2865-360GH354-208 NAU2865-370GH354-230 NAU2873-328GH354-236 NAU2873-352GH454-118 NAU2931-189GH454-122 NAU2931-191GH501-180 NAU2931-250GH501-265 NAU2954-203GH501-268 NAU2954-212GH594-105 NAU2980-256GH594-130 NAU2980-385GH594-170 NAU3011-197GH594-90 NAU3011-235GH60-154 NAU3013-145GH60-160 NAU3013-155GH72-139 NAU3013-158GH72-145 NAU3013-245GH72-149 NAU3013-255GH72-165 NAU3013-280GH72-170 NAU3013-288
NAU3013-294NAU3018-355NAU3018-365NAU3018-395NAU3018-400NAU3018-405NAU3036-185NAU3036-208NAU3036-210NAU3036-213NAU3096-187NAU3096-189NAU3096-241NAU3096-243NAU3096-245NAU3110-224

NAU3110-268NAU3110-292NAU3110-318NAU3254-265NAU3254-288NAU3305-155NAU3305-162NAU3307-230NAU3307-235NAU3307-240NAU3307-398NAU3307-490NAU3325-232NAU3325-238NAU3325-249NAU3325-268NAU3325-275NAU3377-150NAU3377-165NAU3377-170NAU3377-173NAU3377-180NAU3419-240NAU3419-252NAU3486-195NAU3486-210NAU3486-230NAU3486-290NAU3499-215NAU3499-220NAU3499-225NAU3499-230NAU3519-200NAU3519-210NAU3563-149NAU3563-178NAU3563-228NAU3588-290NAU3588-315NAU3588-321NAU3588-325NAU3588-332NAU3602-188NAU3602-195NAU3608-242NAU3608-245NAU3665-237NAU3665-285NAU3668-215NAU3676-185NAU3676-195NAU3736-201

NAU3736-240NAU3744-175NAU3744-200NAU3744-220NAU3774-241NAU3774-248NAU3797-206NAU3797-225NAU3839-370NAU3839-385NAU3995-192NAU3995-193NAU3995-194NAU3995-196NAU4030-145NAU4030-150NAU4030-155NAU4030-175NAU4030-235NAU4030-255NAU4030-298NAU4042-175NAU4042-180NAU4042-185NAU4042-203NAU4042-208NAU4042-222NAU4047-365NAU4047-375NAU4047-458NAU4047-476NAU4047-484NAU4956-240NAU4956-250NAU4956-260NAU5099-233NAU5099-280NAU5128-316NAU5128-329NAU5128-348NAU5128-516NAU5129-140NAU5129-143NAU5129-147NAU5129-175NAU5163-216NAU5163-221NAU5306-155NAU5306-160NAU5306-170NAU5306-180NAU5306-189

NAU5306-193NAU5368-140NAU5368-142NAU5368-145NAU5373-218NAU5418-240NAU5418-248NAU5418-253NAU5433-231NAU5433-330NAU895-175NAU895-178NAU895-182NAU895-188NAU899-220NAU905-168NAU905-172NAU934-195NAU934-200NAU990-352