Embed Size (px)
Citation preview

Biological Psychology 82 (2009) 192–194
Brief report
Affective modulation of conditioned eyeblinks
Suvi Karla a, Timo Ruusuvirta b,c,d, Jan Wikgren a,*a Department of Psychology, University of Jyvaskyla, P.O. Box 35, FIN-40014 Jyvaskyla, Finlandb Turku Institute for Advanced Studies, Centre for Cognitive Neuroscience, Department of Psychology, University of Turku/Turku School of Economics,
Assistentinkatu 7, FIN-20014 Turku, Finlandc Department of Psychology, University of Tampere, Kalevantie 5, FIN-33014 Tampere, Finlandd Cognitive Brain Research Unit, Department of Psychology, University of Jyvaskyla, FIN-00014, Helsinki, Finland
A R T I C L E I N F O
Article history:
Received 3 February 2009
Accepted 28 June 2009
Available online 4 July 2009
Keywords:
Reflex
Eyeblink conditioning
Emotion
IAPS
A B S T R A C T
Affective states are known to modulate reflexive actions. Aversive states potentiate defensive reflexes
while appetitive states diminish them. The present study examined whether the same holds for
associatively learned defensive eyeblinks to mild, initially neutral auditory stimuli. First, delay eyeblink
conditioning was applied to human participants while they viewed emotionally neutral images. Next,
the conditioned eyeblink responses (CRs) of the participants were tested during the viewing of
unpleasant, neutral, or pleasant images. The most vigorous CRs were found during the unpleasant
images, although they did not differ between neutral and pleasant images. The results add to the
motivational priming hypothesis by demonstrating its partial applicability to associatively learned
defensive behaviour.
� 2009 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Biological Psychology
journa l homepage: www.e lsev ier .com/ locate /b iopsycho
1. Introduction
The motivational priming hypothesis (for details see, e.g. Langet al., 1990) states that reflexive actions are augmented when theyare congruent with the ongoing motivational state. In practice, thehypothesis states that negative emotions activate the brain’saversive motivational system and prime defensive reflexes.
Experimentally, motivational states (and emotions) are typi-cally induced by presenting images varying in content fromunpleasant to pleasant. Indeed, the motivational priming hypoth-esis has withstood a plethora of tests combining picture viewingwith an acoustic startle probe. Additionally, other types ofemotional background stimuli have been utilized; for example,movies (Jansen and Frijda, 1994; Kaviani et al., 1999), words(Aitken et al., 1999), odours (Miltner et al., 1994; Ehrlichman et al.,1995 Ehrlichman et al., 1997; Prehn et al., 2006), imagery (Vranaand Lang, 1990) or stimuli previously paired with negativeconsequences (Grillon, 2002; Asli and Flaten, 2007; Mallan andLipp, 2007). Likewise, tactile eyeblink reflexes, evoked by mildairpuffs (Hawk and Cook, 1997) have been shown to be modulatedby emotional background stimuli. Only reflexes of a neutral, non-defensive nature, such as the tendon reflex, seem to be unaffectedby such modulation (Bonnet et al., 1995).
* Corresponding author. Tel.: +359 14 260 2848; fax: +358 14 260 2841.
E-mail address: [email protected] (J. Wikgren).
0301-0511/$ – see front matter � 2009 Elsevier B.V. All rights reserved.
doi:10.1016/j.biopsycho.2009.06.008
The motivational priming hypothesis (Lang et al., 1990)implies that any defensively motivated behaviour, not onlyinnate reflexes, should be modulated by emotional state.However, it is unclear whether a probe for emotional primingcould be based on associative plasticity. The present studyaimed at investigating whether conditioned eyeblinks inresponse to initially neutral non-arousing tones (e.g. Thompsonet al., 1998) could provide such a probe. The adult humanparticipants were first subjected to classical eyeblink condition-ing while they viewed emotionally neutral images. Their CRswere then assessed during the viewing of pleasant, neutral, andunpleasant images.
2. Methods
2.1. Subjects
Twenty-five university students (age range 20–28 years; mean age 21 years
male, 24 years females) participated in the experiment. As compensation for their
participation, the students received a ticket to the local cinema. The research ethics
committee of the University of Jyvaskyla approved the study.
2.2. Stimuli and apparatus
The tone CS (70 dB, 500 ms, 1000 Hz) was presented via a loudspeaker above the
participant’s head. The US was an airpuff (0.5 bar source pressure, 100 ms) to the
outer corner of the right eye, delivered via a plastic tube attached to specially
designed goggles.
The emotional stimuli were 50 pictures, 40 neutral (e.g. household objects and
neutral faces), 10 pleasant (e.g. food and animals), and 10 unpleasant (e.g. violence
and spiders) randomly selected for each participant from a subset of the

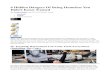
Fig. 1. CR magnitudes were modulated as a function of image content. Unpleasant
(UN) images evoked the largest and neutral pictures (NE) the smallest CRs. Paired
comparisons (t-tests) on standardized CR magnitude showed significant differences
between pleasant (PL) and unpleasant (UN) and also between neutral (NE) and
unpleasant (UN) images. UR magnitudes did not differ significantly between image
contents.
S. Karla et al. / Biological Psychology 82 (2009) 192–194 193
International Affective Picture System1 (IAPS; The Center for Research in
Psychophysiology, 2001). The mean normative valence ratings for the pictures
used were 7.53, 5.00 and 2.67, respectively (1–9 scale, low values representing
unpleasantness) and arousal ratings were 4.85, 2.97, and 5.91 (1–9 scale) for the
pleasant, neutral, and unpleasant categories, respectively.
2.3. Procedure
The participants were informed that their task was to sit in a comfortable chair in
a dimly lit room and to watch images presented on a 17 in. computer monitor
approximately 1 m in front of them, while occasional tones and airpuffs to the
corner of the eye were administered.
Eyeblinks were recorded as EMG activity of the orbicularis oculi muscle beneath
the right eye. The ground electrode was attached to the forehead. The raw EMG
signal was fed into custom-built amplifier, digitized (sampling rate 1000 Hz),
rectified, and digitally off-line filtered with low-pass (<20 Hz) filter.
The experiment consisted of two phases. In the acquisition phase, 80 CS–US pairs
were presented during viewing of pictures, presented in a random order and
selected from among 40 neutral Images.1 In the test phase, 30 CS–US pairs were
presented during presentation of pictures varying in emotional content (10
pleasant, 10 neutral, and 10 unpleasant pictures in presented pseudorandom
order). During both phases, the pictures were visible for 6 s and CS–US pairs were
presented within that time-frame so that the onset of the CS varied randomly
between 1 and 5 s from the image onset. The CS and US were presented in a delayed
fashion with an inter-stimulus interval (onset-to-onset) of 400 ms. The inter-trial
intervals varied between 15 and 24 s, during which the monitor was black. E-Prime
software was used to control the experiment.
2.4. Data analysis
To minimize the between-subject variability in the EMG magnitude, the signal
was standardized for each participant by expressing each data point as a percentage
of the average response magnitude across the first five unconditioned responses. All
off-line signal processing was carried out using Matlab with the Signal Processing
Toolbox.
Three periods were determined for the purpose of the analyses: baseline (200 ms
before the onset of the CS), CR (200 ms before the onset of the US) and UR (200 ms
after the onset of the US). For each trial, the maximum CR and UR magnitudes were
determined from corresponding 200 ms periods using the standardized EMG
signals and then averaged for eight conditioning blocks (10 trials each) in the
acquisition phase, and for each image content in the test phase.
Variation in spontaneous EMG activity was determined from the baseline
activity. An eyeblink was considered to have occurred when the maximum EMG
activity in a given period exceeded the spontaneous activity by more than 5
standard deviations. Blink probability was thus determined in each trial for baseline
and CR periods. The CR probability was then calculated by subtracting the blink
probability in the baseline period from that of the CR period in each trial block.
Analysis of variance for repeated measures and subsequent paired samples
t-tests with Bonferroni-correction were used to analyse the effects of conditioning
and image content. Greenhouse–Geisser corrected degrees of freedom were used if
the sphericity assumption was violated.
3. Results
3.1. Development of the CR
As a function of training, average CR magnitude increased from12.59 (SEM = 1.54) to 19.02 (2.34) standardized units and CRprobability from 40.0 (7.64) to 74.0 (4.43)%. There was a significantmain effect of conditioning for both CR magnitude [F(7,168) = 6.52,p < 0.001, h2
partial = 0.214] and probability [F(7,168) = 10.96,p < 0.05, h2
partial = 0.313]. Subsequent paired comparisons verifiedthat both CR magnitude and CR probability in blocks 2–8 weresignificantly higher than in the first block [CR magnitude:
1 IAPS-slide numbers:
Neutral: 2190, 2200, 2210, 2393, 2514, 2600, 5130, 5500, 5510, 5740, 5750,
5900, 7000, 7002, 7004, 7009, 7010, 7020, 7030, 7031, 7050, 7060, 7080, 7090,
7100, 7110, 7130, 7150, 7160, 7170, 7180, 7182, 7184, 7211, 7217, 7233, 7500,
7550, 7700, 7705.
Pleasant: 1440, 1460, 1600, 1603, 1610, 1620, 1750, 1920, 2040, 2050, 2071,
2080, 2091, 2208, 2250, 2299, 2331, 4650, 4660, 4680, 5200, 7200, 7230, 7270,
7280, 7330, 7350, 8030, 8080, 8120, 8200, 8510.
Unpleasant: 1070, 1090, 1110, 1114, 1120, 1205, 1274, 1280, 1300, 2053, 2095,
2120, 2141, 2205, 2276, 2683, 2710, 2800, 3000, 3010, 3030, 3100, 3120, 3130,
3140, 3150, 3530, 6020, 6190, 6200, 6230, 6370, 6415, 6555, 7380, 9001, 9040,
9041, 9050, 9340, 9342, 9490.
t(24) = 3.39–4.47, p < 0.01; CR probability: t(24) = 2.92–4.47,p < 0.01]. Comparison of CR magnitude and probability in blocks3–8 with that in block 2 only yielded a statistically significantdifference for CR probability [block 2 vs. block 8: t(24) = 2.17,p < 0.05], suggesting that the level of conditioned responding wasrelatively stable from the block 2 onwards.
3.2. Effect of image content on CR and UR magnitudes
Fig. 1 shows the CR and UR magnitudes as a function of picturecontent in the test phase. The picture content had a significantmain effect on CR magnitude [F(2,48) = 6.76, p < 0.01, h2
par-
tial = 0.220], but not on CR probability [F(2,48) = 0.63, p = 0.54,h2
partial = 0.026] or UR magnitude [F(2,48) = 1.80, p = 0.178,h2
partial = 0.069]. Subsequent paired comparisons showed higherCR magnitudes during unpleasant relative to pleasant [t(24) = 2.68,p < 0.05] and relative to neutral pictures [t(24) = 2.82, p < 0.01].No difference was observed between neutral and pleasant pictures[t(24) = 0.15, p = 0.879].
4. Discussion
We investigated the applicability of the motivational priminghypothesis (Lang et al., 1990) to associatively learned, defensiveeyeblinks. We found that defensive eyeblink CRs were augmentedduring unpleasant pictures. This augmentation is analogous to thatof startle responses in similar contexts (Lang et al., 1990). Ourfinding, therefore, suggests that emotional state also affectsassociatively learned, motivationally driven, yet emotionallyrelatively neutral, behavioural acts. This adds to the generalvalidity of the motivational priming hypothesis (Lang et al., 1990).
However, the lack of observable CR diminution for pleasantrelative to neutral pictures was in contrast to the previous findingsof such diminution in startle eyeblinks (Lang et al., 1990). Thecause of this discrepancy remains unclear. Nevertheless, we mustacknowledge that CRs and URs recruit partially distinct neuralnetworks and mechanisms. In other words, whereas eyeblinkreflexes are driven by simple brain-stem pathways, conditioned

S. Karla et al. / Biological Psychology 82 (2009) 192–194194
eyeblinks also depend on a distinct cerebellar learning circuit (e.g.Steinmetz, 2000), which might be differently affected by experi-enced valence, specifically concerning pleasant states.
Unexpectedly, however, the emotional state did not modulatethe magnitude of airpuff URs. The reason for this was most likelymethodological. The high US intensity used to assure robustlearning was very likely the cause of a ceiling effect for URmagnitude, even with pleasant pictures. In line with this,emotional state has been shown to modulate airpuff-inducedeyeblinks in the case of milder airpuffs (Hawk and Cook, 1997).
To conclude, our findings add to the motivational priminghypothesis (Lang et al., 1990) by showing that unpleasant affectivestates also augment associatively learned defensive responses.Future studies should address the lack of their diminution bypleasant affective states.
Acknowledgements
The study was funded by a grant from the Academy of Finlandto JW (114258). The authors wish to thank Lauri Viljanto fortechnical help, Dave Lavond for commenting an earlier version ofthe manuscript and Michael Dutton for revising our English.
References
Aitken, C.J., Siddle, D.A., Lipp, O.V., 1999. The effects of threat and nonthreat wordlead stimuli on blink modification. Psychophysiology 36, 699–705.
Asli, O., Flaten, M.A., 2007. Conditioned facilitation of the unconditioned reflexafter classical eyeblink conditioning. International Journal of Psychophysiology67, 17–22.
Bonnet, M., Bradley, M.M., Lang, P.J., Requin, J., 1995. Modulation of spinal reflexes:arousal, pleasure, action. Psychophysiology 32, 367–372.
Ehrlichman, H., Brown, S., Zhu, J., Warrenburg, S., 1995. Startle reflex modulationduring exposure to pleasant and unpleasant odors. Psychophysiology 32,150–154.
Ehrlichman, H., Kuhl, S.B., Zhu, J., Warrenburg, S., 1997. Startle reflex modulation bypleasant and unpleasant odors in a between-subjects design. Psychophysiology34, 726–729.
Grillon, C., 2002. Associative learning deficits increase symptoms of anxiety inhumans. Biological Psychiatry 51, 851–858.
Hawk, L.W., Cook 3rd, E.W., 1997. Affective modulation of tactile startle. Psycho-physiology 34, 23–31.
Jansen, D.M., Frijda, N.H., 1994. Modulation of the acoustic startle response by film-induced fear and sexual arousal. Psychophysiology 31, 565–571.
Kaviani, H., Gray, J.A., Checkley, S.A., Kumari, V., Wilson, G.D., 1999. Modulation ofthe acoustic startle reflex by emotionally-toned film-clips. International Journalof Psychophysiology 32, 47–54.
Lang, P.J., Bradley, M.M., Cuthbert, B.N., 1990. Emotion, attention and startle reflex.Psychological Review 97, 377–395.
Lang, P.J., Bradley, M.M., Cuthbert, B.N., 2001. International affective picture system(IAPS). Instruction manual and affective ratings. Technical report A-5, TheCenter for research in psychophysiology, University of Florida.
Mallan, K.M., Lipp, O.V., 2007. Does emotion modulate the blink reflex in humanconditioning? Startle potentiation during pleasant and unpleasant cues in thepicture-picture paradigm. Psychophysiology 44, 737–748.
Miltner, W., Matjak, M., Braun, C., Diekmann, H., Brody, S., 1994. Emotional qualitiesof odors and their influence on the startle reflex in humans. Psychophysiology31, 107–110.
Prehn, A., Ohrt, A., Sojka, B., Ferstl, R., Pause, B.M., 2006. Chemosensory anxietysignals augment the startle reflex in humans. Neuroscience Letters 394,127–130.
Steinmetz, J.E., 2000. Brain substrates of classical conditioning: a highly localizedbut also distributed system. Behavioral Brain Research 110, 13–24.
Thompson, R.F., Thompson, J.K., Kim, J.J., Krupa, D.J., Shinkman, P.G., 1998. Thenature of reinforcement in cerebellar learning. Neurobiology of Learning andMemory 70, 150–176.
Vrana, S.R., Lang, P.J., 1990. Fear imagery and startle-probe reflex. Journal ofAbnormal Psychology 99, 189–197.