Embed Size (px)
Citation preview

Investigational New Drugs 14: 271-280, 1996. 271 @ 1996 Kluwer Academic Publishers. Printed in the Netherlands.
Antitumor diarylsulfonylureas: novel agents with unfulfilled promise
Pe te r J. H o u g h t o n and Jane t A. H o u g h t o n Department of Molecular Pharmacology, St. Jude Children's Research Hospital, Memphis, TN 38105-2794, USA
Key words: diarylsulfonylureas, xenografis, pharmacology
Summary
Diarylsulfonylureas represent a class of antitumor agent with significant therapeutic efficacy against rodent and human models of cancer. Despite the exciting preclinical activity of sulofenur, the prototypic agent, clinical activity was poor. Here we review the activity of sulofenur and a second generation diarylsulfonylurea, LY295501 N-[5- (2,3-dihydrobenzofuryl) sulfonyl]-W-(3,4-dichlorophenyl)urea, against colon tumor xenografts, and some of the cellular pharmacology of this class of antitumor agents.
Abbreviations. DSU - antitumor diarylsulfonylureas; Solofenur - ISCU - N-(5-indanylsulfonyl)-N-(4-chloro- phenyl) urea; LY195779 - N-(4-methylphenylsulfonyl)-NL(4-nitrophenyl)urea; LY186391 - N-(phenylsulfonyl)- N'-(3,4-dichlorophenyl)urea; MPCU - N-(4-methylphenylsulfonyl)- W-(4-chlorophenyl)urea; APCU - N- (4-azidophenylsulfonyl)-W-(4-chlorophenyl)urea; LY295501 - N-[5-(2,3-dihydrobenzofuryl)sulfonyl]-W-(3,4- dichlorophenyl)urea; FCCP - carbonylcyanide p-trifluoromethoxyphenylhydrazone; SDS-PAGE - sodium dode- cylsulfate polyacrylamide gelelectropheresis; Pgp - P-glycoprotein; MTD - maximal tolerated dose
Introduction
As part of the "extra mural" program instituted by Dr. Gerald Grindey at Eli Lilly and Company, we started to evaluate a new class of antitumor agent, the diarylsul- fonylureas (DSU), that had been identified in animal testing. The compounds have structural similarity to oral hypoglycemic agents, such as glibenclamide (Fig- ure 1), although the series was originally synthesized as potential herbicides [1]. In collaboration with Dr. Grindey, we demonstrated that N-(5-indanylsulfonyl)- W-(4-chlorophenyl)urea (sulofenur, ISCU) exhibited significant activity against advanced colon adenocar- cinoma xenografts intrinsically resistant to virtually all standard chemotherapeutic agents [2-4] and also had very significant activity against several pediatric tumors grown as xenografts [5]. Clinically, sulofenur was disappointing, and although some responses were reported in phase I trials [6, 7] this was not confirmed for ovarian cancer in the phase II setting [8]. Toxi- city in man was due mainly to methemoglobinemia and anemia [6, 9], which limited the concentrations of
drug that could be achieved to levels well below those shown to elicit a therapeutic response in the xenograft models [8]. This species specific difference was con- sidered to be a consequence of differential metabolism between man and rodents. Extensive studies by Ehlhardt [10-12] resulted in a scheme of metabolism in which sulofenur may undergo ortho-hydroxylation of the indane ring, and possibly direct metabol- ic dehydrogenation [12]. Toxicity in man is consis- tent with that associated with p-chloroaniline, which may be formed by metabolic conversion of sulofenur directly, or after initial metabolism. Consequently, the analog N-[5-(2,3-dihydrobenzofuryl)sulfonyl]-W- (3,4-dichlorophenyl)urea (LY295501; Figure 1) was synthesized with the intent of blocking the initial con- version observed with sulofenur, specifically hydroxy- lation at the 1-position of the indane ring. Toxicology in primates and rats indicated the toxicity profile of LY295501 to be different from sulofenur, being pri- marily diarrhea and radio mimetic-like lesions in tis- sues with high turnover. Intestinal toxicity was dose limiting, whereas non-limiting hematological alter-

272
~ f f
sulofenur
CI
T LY295501
Figure 1. Structure of sulofenur and LY295501.
CI
ations in monkeys were associated with oxidation of hemoglobin [13].
The mechanism by which antitumor DSU exert cytotoxicity remains unknown. Over the last five years several reports have suggested that DSU may alter intracellular calcium regulation [ 14-17], the cytoskele- tal network [18], and that introduction of activated H-ras may sensitize NRK-52E cells to these effects. Further, sulofenur has rapidly induced the expression of c-fos, c-jun and hsp70 (within 15 minutes) [19, 20]. While such observations are of interest, their signifi- cance to the pharmacologically relevant site of action of antitumor DSU are questionable. The studies cit- ed above used concentrations of sulofenur > 125 #M under serum-free conditions and most studies report- ed effects at 500 #M. Under these conditions we have shown that sulofenur causes rapid equilibration of extra cellular and intracellular pH, possibly by direct dam- age to the plasma membrane [21 ]. In plasma, sulofenur is highly protein-bound (> 99.9%), thereby yielding free-drug concentrations in the range of 0.5-1 /zM under therapeutic conditions [8, 22]. Consequently bio- chemical alterations reported using concentrations of drug 1000-fold greater than those achievable in plasma probably do not relate to our proposed "site 1" target of DSU cytotoxicity that appears relevant in vivo.
In this article we will review some of the antitumor activity of sulofenur, and the new analog, LY295501,
Table 1. Responsiveness of xenografts of childhood rhab- domyosarcoma
Dose (nag&g): Tumor 300 200 1 O0
Schedule, b.i.d. • 10 days: Rhl2 ++++a +++ Rhl2/VCR-3 ++++ ND Rhl2/ifos ++++ ND Rhl8 ++++ ND Rh 18/VCR-3 ++++++ ND Rh 18/L-PAM +++ ND Rh28 ++++++ +++++ Rh28/L-PAM +++++ +++ IRS68 ++++ ND
Schedule, b.i.d. • 5 days2~: Rhl2 ++++ +++ Rh30 ++++++ ++++++ ND IRS49 ++++++ ++++++ Rh 18/VCR-3 ++++++ ++++++ IRS68 +++++ +++++ ND
Schedule, b,i.d, x 5 days3b: Rh 18 ++++++ 4-+++++ Rhl2 +++++ ++++
a Tumor response criteria: +++, growth inhibition of > 3 tumor volume Td2; ++++, growth inhibition of > 3 • Td2 plus volume regression of _> 50%, +++++, complete regression with subsequent regrowth, ++++++, complete regression with no growth during the period of observation (_> 84 days). (Td2 mean time for tumor volume to double). b Subscript indicates the number of courses. ND, not determined. From ref. 5.
currently in clinical phase I trials, and some of the studies we have undertaken relating to the mechanism by which these agents may cause cytotoxic effects.
Results and discussion
Therapeutic activity in human tumor xenografis. We initially demonstrated that sulofenur had significant activity against a panel of xenografts derived from childhood rhabdomyosarcoma, a tumor of myogenic origin [5]. As shown in Table 1, when administered by oral gavage twice daily for 10 days, or 5 days per week for two or three consecutive weeks, sulofenur caused partial or complete regressions in most of the tumors examined. Of significance was a lack of cross- resistance in tumors selected in situ for resistance to vincristine or bifunctional alkylating agents.
The activity of sulofenur against colon adenocar- cinoma xenografts is presented in Table 2. In this

Table 2. Activity o f sulofenur against adult derived colon adenocarc inomas
Dose Days to Growth delay %
Tumor (b.i.d. mg/kg) 4 x 4- SE (days) PR CR MCR Survival
HCt Control t6.7 4- 1.6 (100 x 5)2 27.4 4- 5.6 10 0 0 0 100 (200 x 5)2 29.4 4- 2.2 12 0 0 0 100
(100 x 5)3 30.7 4- 5.4 14 0 0 0 100 (300 x 5)2 Toxic
GC3 Control 20.1 4- 4.5
(200 x 5)2 63.6 4- 4.1 43 33 0 0 83 (300 x 5)2 64.1 4- 5 44 92 8 0 80
(400 • 5)2 >84 >64 41 42 17 86 (100 x 5)3 67.2 4- 3.7 47 i6 0 0 100
(200 x 5)3 >84 >64 30 30 20 80
VRC5 Control 10.9 • 0.5 (100 x 5)2 59.9 4- 4.0 49 21 79 14 100
(200 x 5)2 68.8 4- 9.9 58 7 93 14 100 (400 x 5)2 71.2 4- 5.6 61 43 50 29 50
(600 • 5)2 Toxic
ELC2 Control 36.5 4- 4.6 (200 x 5)2 67.9 4- 3.7 31 0 0 0 100 (300 • 5)2 79.0 4- 2.2 43 7 0 0 86
(400 x 5)2 82.9 4- 1.2 46 0 0 0 86 (200 x 5)3 >84 >47 0 0 0 100
GC3/cl Control 49.9 4- 6.4 (200 x 5)2 74.1 4- 4.7 24 50 33 33 I00
(400 x 5)2 76.1 4- 1.7 26 75 25 25 57
GC3/LYC5 Control 38.5 4- 2.6 (200 x 5)3 41.1 4- 3.6 2 0 0 0 100 (400 x 5)3 34.1 4-0,4 -4 0 0 0 i00
PR, partial regression ( > 5 0 % ) ; CR, complete regression; MCR, maintained CR at 12 weeks. F rom reference 4.
273
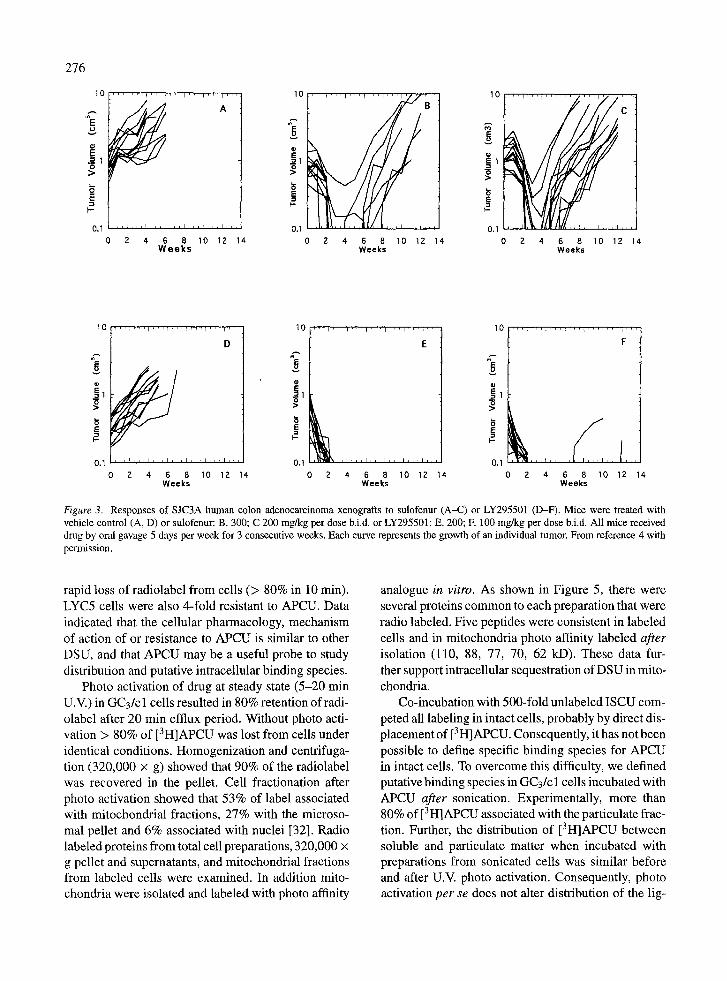
study, sulofenur was administered for two or three consecutive 5-day courses, and resulted in a signif- icant frequency of partial and complete regressions. Of note, the line GC3/LYC5, selected in vitro for resistance to sulofenur, was completely resistant when established as a xenograft. Representative responses of parental GC3/c 1 and GC3/LYC5 xenografts are shown in Figure 2. The activity of LY295501 against colon tumors derived from adult and juvenile patients are summarized in Table 3. Although more toxic than sulofenur, LY295501 demonstrated equal or superi- or efficacy against SJC3A tumors (Figure 3). These results indicate that LY295501 has a similar spectrum of activity as sulofenur in a panel of colon adenocarci- noma xenografts, and this analog has recently entered phase I clinical trial. In colon tumor xenografts, at the minimum tolerated dose intensity (MTD), LY295501 caused objective regressions in 3 or 6 tumor lines with significant growth stasis in two additional tumor lines.
Tumors intrinsically resistant to sulofenur were also resistant to LY295501. Thus, LY295501 ranks as one of the more efficacious agents tested against this panel of xenografts, and provides an important lead in devel- oping a new therapeutic agent with potentially novel mechanism of action.
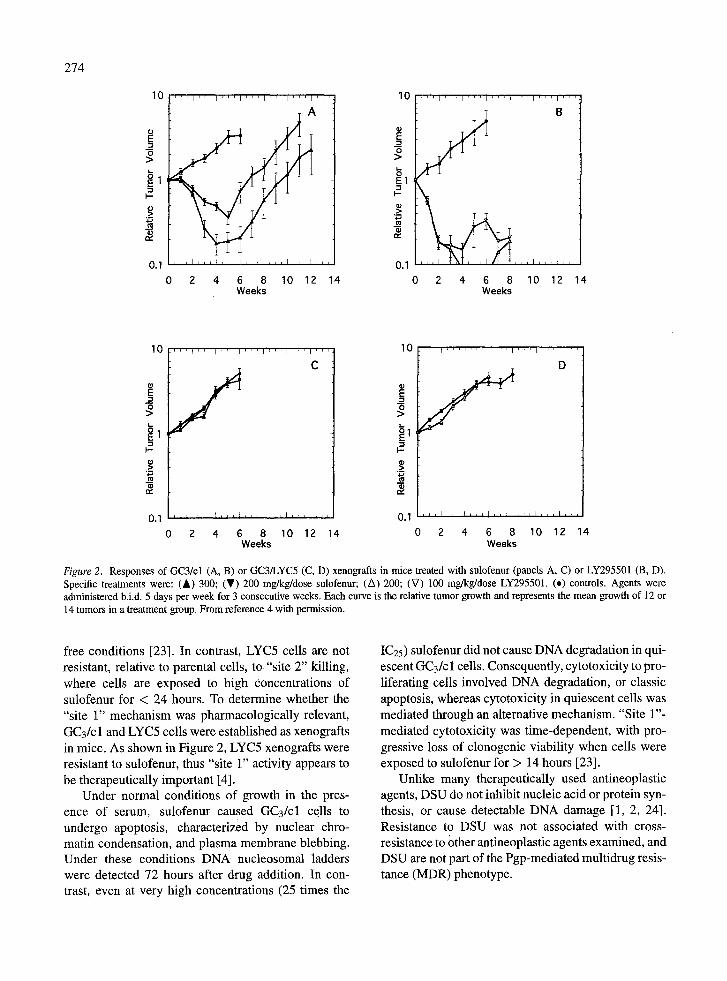
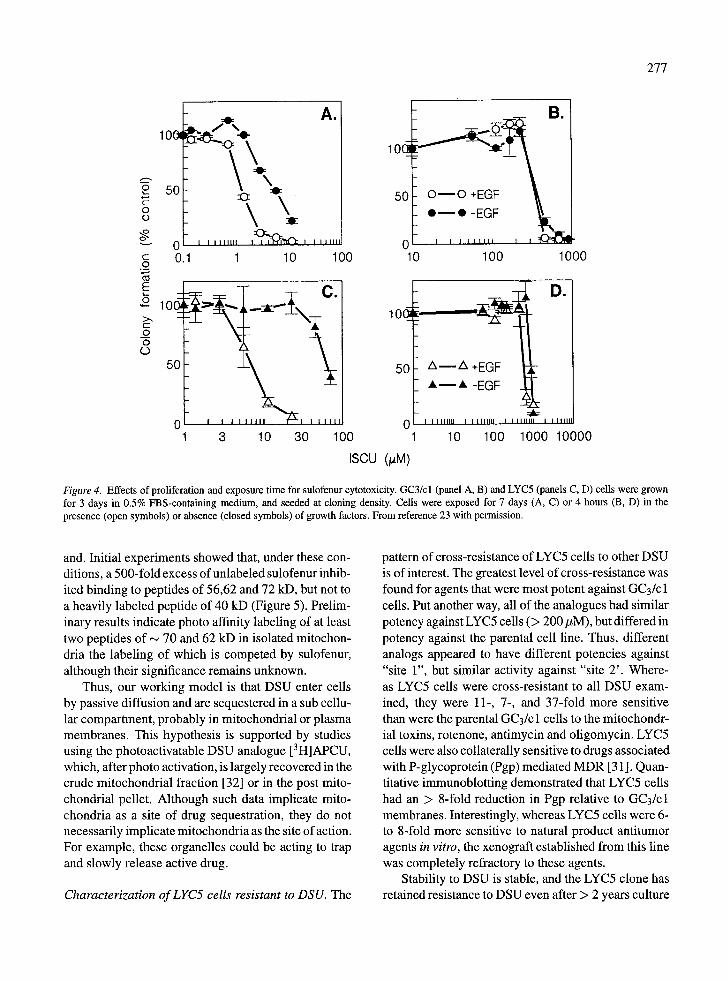
Mechanisms of cytotoxicity. Our studies have sug- gested at least two mechanisms of drug action based on studies with GC3/cl parental sensitive colon ade- nocarcinoma cells, and a clone selected for resis- tance to sulofenur (LYC5) [23]. The pharmacologi- cally relevant mechanism has been designated "site 1", and pertains to cytotoxicity at low drug concentra- tions when cells are exposed continuously for 7 days. Under these conditions cytotoxicity is proliferation- dependent (Figure 4). LYC5 cells were approximately 13-fold resistant to "site 1" cytotoxicity under serum-
274
10
_= o
>
-n
0.1
, , i , , . i , , , i , , , i , , , i , , , 1 1
2 4 6 8 10 12 Weeks
10
E _= o
>
E1 .-I I -
>o
e~
' 1 ' ' 1 ' '
B
0 2 4 6 8 Weeks
0 . 1 ~ ~ ' '
14 10 12 14
10
==
E 1
0.1
0
' r ' ' i ~ ' l ' ' ' l '
/ , , , i ~
C
2 4 6 8 1 0 1 2
W e e k s
10
~o E _.= o
>
t -
"D
'1 ' 1 ' '
D
0 . 1 ~ , 1 , ~ ~ 1 ~ , ~ r ~ , I ,
0 2 4 6 8 Weeks
14 14
l i i I r
10 12
Figure 2. Responses of GC3/cl (A, B) or GC3/LYC5 (C, D) xenografts in mice treated with sulofenur (panels A, C) or LY295501 (B, D). Specific treatments were: (&) 300; (T) 200 mg/kg/dose sulofenur; (A) 200; (V) 100 mg/kg/dose LY295501. (.) controls. Agents were administered b.i.d. 5 days per week for 3 consecutive weeks. Each curve is the relative tumor growth and represents the mean growth of 12 or 14 tumors in a treatment group. From reference 4 with permission.
free conditions [23]. In contrast, LYC5 cells are not resistant, relative to parental cells, to "site 2" killing, where cells are exposed to high concentrations of sulofenur for < 24 hours. To determine whether the "site 1" mechanism was pharmacologically relevant, GC3/c 1 and LYC5 cells were established as xenografts in mice. As shown in Figure 2, LYC5 xenografts were resistant to sulofenur, thus "site 1" activity appears to be therapeutically important [4].
Under normal conditions of growth in the pres- ence of serum, sulofenur caused GC3/cl cells to undergo apoptosis, characterized by nuclear chro- matin condensation, and plasma membrane blebbing. Under these conditions DNA nucleosomal ladders were detected 72 hours after drug addition. In con- trast, even at very high concentrations (25 times the
IC25) sulofenur did not cause DNA degradation in qui- escent GC3/c 1 cells. Consequently, cytotoxicity to pro- liferating cells involved DNA degradation, or classic apoptosis, whereas cytotoxicity in quiescent cells was mediated through an alternative mechanism. "Site l"- mediated cytotoxicity was time-dependent, with pro- gressive loss of clonogenic viability when cells were exposed to sulofenur for > 14 hours [23].
Unlike many therapeutically used antineoplastic agents, DSU do not inhibit nucleic acid or protein syn- thesis, or cause detectable DNA damage [1, 2, 24]. Resistance to DSU was not associated with cross- resistance to other antineoplastic agents examined, and DSU are not part of the Pgp-mediated multidrug resis- tance (MDR) phenotype.
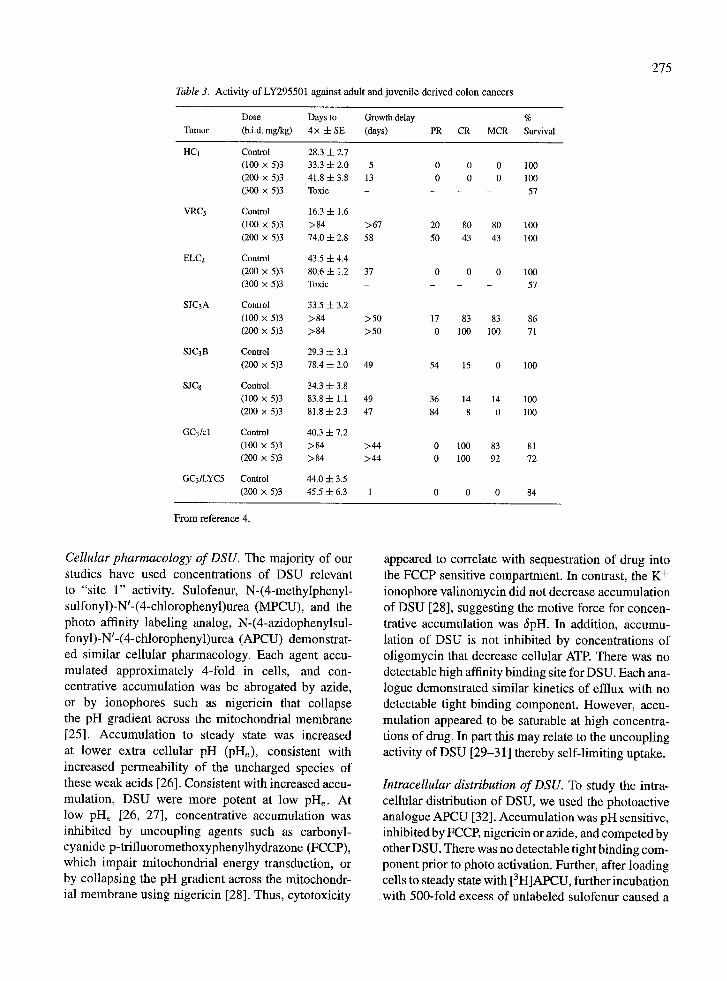
Table 3. Activity of LY295501 against adult and juvenile derived colon cancers
Dose Days to Growth delay %
Tumor (b.i.d. mg/kg) 4 x -4- SE (days) PR CR MCR Survival
HC1 Control 28.3 -I- 2.7
(100 x 5)3 33.3 4- 2.0 5 0 0 0 100
(200 x 5)3 41.8 4- 3.8 13 0 0 0 100
(300 x 5)3 Toxic - - - 57
VRC5 Control 16.3 4- 1.6
(100 x 5)3 >84 >67 20 80 80 100
(200 x 5)3 74.0 4- 2.8 58 50 43 43 100
ELC2 Control 43.5 4- 4.4
(200 x 5)3 80.6 4- 1.2 37 0 0 0 100 (300 x 5)3 Toxic - - - 57
SJC3A Control 33.5 4- 3.2
(100 • 5)3 >84 >50 17 83 83 86
(200 x 5)3 >84 >50 0 100 100 71
SJC3B Control 29.3 4- 3.3
(200 x 5)3 78.4 4- 2.0 49 54 15 0 100
SJC8 Control 34.3 4- 3.8
(100 • 5)3 83.8 4- 1.1 49 36 14 14 100
(200 x 5)3 81.8 4- 2.3 47 84 8 0 100
GC3/cl Control 40.3 4- 7.2
(100 x 5)3 >84 >44 0 100 83 81
(200 x 5)3 >84 >44 0 100 92 72
GC3/LYC5 Control 44.0 4- 3.5
(200 x 5)3 45.5 4- 6.3 1 0 0 0 84
From reference 4.
275
Cellular pharmacology of DSU. The majority of our studies have used concentrations of DSU relevant to "site 1" activity. Sulofenur, N-(4-methylphenyl- sulfonyl)-N~-(4-chlorophenyl)urea (MPCU), and the photo affinity labeling analog, N-(4-azidophenylsul- fonyl)-Nl-(4-chlorophenyl)urea (APCU) demonstrat- ed similar cellular pharmacology. Each agent accu- mulated approximately 4-fold in cells, and con- centrative accumulation was be abrogated by azide, or by ionophores such as nigericin that collapse the pH gradient across the mitochondrial membrane [25]. Accumulation to steady state was increased at lower extra cellular pH (pile), consistent with increased permeability of the uncharged species of these weak acids [26]. Consistent with increased accu- mulation, DSU were more potent at low pile. At low pH~ [26, 27], concentrative accumulation was inhibited by uncoupling agents such as carbonyl- cyanide p-trifluoromethoxyphenylhydrazone (FCCP), which impair mitochondrial energy transduction, or by collapsing the pH gradient across the mitochondr- ial membrane using nigericin [28]. Thus, cytotoxicity
appeared to correlate with sequestration of drug into the FCCP sensitive compartment. In contrast, the K + ionophore valinomycin did not decrease accumulation of DSU [28], suggesting the motive force for concen- trative accumulation was ~pH. In addition, accumu- lation of DSU is not inhibited by concentrations of oligomycin that decrease cellular ATR There was no detectable high affinity binding site for DSU. Each ana- logue demonstrated similar kinetics of efflux with no detectable tight binding component. However, accu- mulation appeared to be saturable at high concentra- tions of drug. In part this may relate to the uncoupling activity of DSU [29-31 ] thereby self-limiting uptake.
Intracellular distribution of DSU. To study the intra- cellular distribution of DSU, we used the photoactive analogue APCU [32]. Accumulation was pH sensitive, inhibited by FCCP, nigericin or azide, and competed by other DSU. There was no detectable tight binding com- ponent prior to photo activation. Further, after loading cells to steady state with [3H]APCU, further incubation with 500-fold excess of unlabeled sulofenur caused a
276
r E --=1 o >
=E I-
0.1 : , t L , , l , , , l , , , l , , , ~ , 0 2 4 6 8 10
Weeks
r l , ,
A
, i , ,
12 14
10 ,,,~. i ' " t ' " , ' , ' t ' , B," 10 ,,,~ ,~,, , . . . . . . , . . . . C
0.I 0.1 " 0 2 4 6 8 10 12 14 0 2 4 6 8 10 12 14
Weeks Weeks
10
_~I
h=
0.1 0 2 4 6 8
Weeks
t ' 1 "
D
i , , , i , .
10 12 14
10
I-
0.1 0 2
E
4 (5 8 10 12 14 Weeks
i O I . . . . .
0 2 4
F
6 8 10 12 14 Weeks
Figure 3. Responses of SJC3A human colon adenocarcinoma xenografts to sulofenur (A-C) or LY295501 (D-F). Mice were treated with vehicle control (A, D) or sulofenur: B. 300; C 200 mg/kg per dose b.i.d, or LY295501: E. 200; E 100 mg/kg per dose b.i.d. All mice received drug by oral gavage 5 days per week for 3 consecutive weeks. Each curve represents the growth of an individual tumor. From reference 4 with permission.
rapid loss of radiolabel from cells (> 80% in 10 rain). LYC5 cells were also 4-fold resistant to APCU. Data indicated that the cellular pharmacology, mechanism of action of or resistance to APCU is similar to other DSU, and that APCU may be a useful probe to study distribution and putative intracellular binding species.
Photo activation of drug at steady state (5-20 min U.V.) in GC3/cl cells resulted in 80% retention ofradi- olabel after 20 min efflux period. Without photo acti- vation > 80% of [3H]APCU was lost from cells under identical conditions. Homogenization and centrifuga- tion (320,000 • g) showed that 90% of the radiolabel was recovered in the pellet. Cell fractionation after photo activation showed that 53% of label associated with mitochondrial fractions, 27% with the microso- mal pellet and 6% associated with nuclei [32]. Radio labeled proteins from total cell preparations, 320,000 x g pellet and supernatants, and mitochondrial fractions from labeled cells were examined. In addition mito- chondria were isolated and labeled with photo affinity
analogue in vitro. As shown in Figure 5, there were several proteins common to each preparation that were radio labeled. Five peptides were consistent in labeled cells and in mitochondria photo affinity labeled after isolation (110, 88, 77, 70, 62 kD). These data fur- ther support intracellular sequestration of DSU in mito- chondria.
Co-incubation with 500-fold unlabeled ISCU com- peted all labeling in intact cells, probably by direct dis- placement of [3H]APCU. Consequently, it has not been possible to define specific binding species for APCU in intact cells. To overcome this difficulty, we defined putative binding species in GC3/c 1 cells incubated with APCU after sonication. Experimentally, more than 80% of [3H]APCU associated with the particulate frac- tion. Further, the distribution of [3H]APCU between soluble and particulate matter when incubated with preparations from sonicated cells was similar before and after U.V. photo activation. Consequently, photo activation per se does not alter distribution of the lig-
277
I0(~
~ 50 c - O 0
0 "- 0.1 .o_
E
>,., t -
O
O
o 5(
:X2[
I I I I I I I I I
1
A.
I I I P I " r T ~ I I I I I I I I
10 100
lO~
50
I
3 10 30 100
10(~
5(
ISCU (p.M)
0 10 100 1000
A - - A +E
~, - - '~ ' -EGF Z
I I I I I I I I I
10
~
I I l l l l l l l I I I I I I I I I I I I I l l l
100 1000 10000
Figure 4. Effects of proliferation and exposure time for sulofennr cytotoxicity. GC3/cl (panel A, B) and LYC5 (panels C, D) cells were grown for 3 days in 0.5% FBS-containing medium, and seeded at cloning density. Cells were exposed for 7 days (A, C) or 4 hours (B, D) in the presence (open symbols) or absence (closed symbols) of growth factors. From reference 23 with permission.
and. Initial experiments showed that, under these con- ditions, a 500-fold excess of unlabeled sulofenur inhib- ited binding to peptides of 56,62 and 72 kD, but not to a heavily labeled peptide of 40 kD (Figure 5). Prelim- inary results indicate photo affinity labeling of at least two peptides of ~ 70 and 62 kD in isolated mitochon- dria the labeling of which is competed by sulofenur, although their significance remains unknown.
Thus, our working model is that DSU enter cells by passive diffusion and are sequestered in a sub cellu- lar compartment, probably in mitochondrial or plasma membranes. This hypothesis is supported by studies using the photoactivatable DSU analogue [3H]APCU, which, after photo activation, is largely recovered in the crude mitochondrial fraction [32] or in the post mito- chondrial pellet. Although such data implicate mito- chondria as a site of drug sequestration, they do not necessarily implicate mitochondria as the site of action. For example, these organelles could be acting to trap and slowly release active drug.
Characterization of LYC5 cells resistant to DSU. The
pattern of cross-resistance of LYC5 cells to other DSU is of interest. The greatest level of cross-resistance was found for agents that were most potent against GC3/c 1 cells. Put another way, all of the analogues had similar potency against LYC5 cells (> 200 #M), but differed in potency against the parental cell line. Thus, different analogs appeared to have different potencies against "site 1", but similar activity against "site 2'. Where- as LYC5 cells were cross-resistant to all DSU exam- ined, they were 11-, 7-, and 37-fold more sensitive than were the parental GC3/c 1 ceils to the mitochondr- ial toxins, rotenone, antimycin and oligomycin. LYC5 cells were also collaterally sensitive to drugs associated with P-glycoprotein (Pgp) mediated MDR [31]. Quan- titative immunoblotting demonstrated that LYC5 cells had an > 8-fold reduction in Pgp relative to GC3/cl membranes. Interestingly, whereas LYC5 cells were 6- to 8-fold more sensitive to natural product antitumor agents in vitro, the xenograft established from this line was completely refractory to these agents.
Stability to DSU is stable, and the LYC5 clone has retained resistance to DSU even after > 2 years culture

278
Figure 5. Radiolabeling of particulate fractions and mitochondria of GC3/cl cells by [3H]LY219703. Monolayer GC3/cl cells were incu- bated with 7,5/tM [3H]LY219703 for 60 rain, and U.V. irradiated for 20 min. Monolayers were extensively washed, trypsinized, homog- enized, and centrifuged. Pellets were suspended in RIPA buffer (containing 2% aprotin and l mM PMSF), and kept on ice until par- ticulate matedai had dissolved. After addition of 10 x SDS sample buffer (40% v/v), samples were mixed and allowed to denature for 2 hours at room temperature. Samples were analyzed by SDS-PAGE, and autoradiograms of gels prepared. A: Radiolabeling pattern for proteins in the 320,000 x g pellet; B lane 1: Coomassie stained gel of a fraction enriched for mitochondria; lane 2: Autoradiogram from the gel shown in lane 1; C: Radiolabeling pattern for proteins in samples of mitochondria isolated from cells prior to incubation and photoactivation of [3H]LY219703. Molecular weight markers indi- cate the mobilities for each set of results. Results are representative of several individual experiments.
in the absence of drug selection pressure. No sponta- neous revertant clones have been identified. To deter- mine whether resistance was dominant or recessive,. hybrid lines fused between parental cells and LYC5 were made using a selective system using irreversible biochemical inhibitors [33]. In these experiments [31], one cell line was treated with diethylpyroearbonate (DEPC) and the other with iodoacetamide. Only cells fused with those from the alternative treatment sur- vived under the conditions used; unfused cells or cells fused to other cells that received the same treatment (e.g., DEPC/DEPC) were not viable. Four indepen- dently derived GC3/LYC5 hybrid clones were char- acterized for sensitivity to sulofenur, and mitochon- drial toxins. Each hybrid demonstrated sensitivity to sulofenur similar to parental GC3/cl cellsl suggest- ing that resistance to sulofenur was recessive. Hybrids
demonstrated resistance to rotenone, antimycin and oligomycin, indicating that hypersensitivity to these agents in LYC5 was also a recessive trait. Thus, from these experiments we may conclude that the phenotype of LYC5, characterized by resistance to DSU and col- lateral sensitivity to mitochondrial toxins, is recessive, and hence both traits may be genetically linked. Com- plicating this were two observations, i) LYC5 cells were collaterally sensitive to agents associated with Pgp-mediated MDR, and these cells demonstrated a significant decrease in detectable Pgp; ii) the MDR modulator, verapamil, markedly increased the sensitiv- ity of parental GC3/c 1 cells to each mitochondrial tox- in, but had significantly less effect in potentiating the toxicity of these toxins in LYC5 cells. Thus, although from structural and charge considerations it seemed unlikely that agents such as rotenone, antimycin and oligomycin were substrates for Pgp-mediated efflux, this potential did exist.
Resistance to DSU in LYC5 cells is stable, conse- quently it was not possible to check linkage between DSU-resistance and mitochondrial toxin-senstivity in revertants. Thus, a genetic approach was undertak- en to transfer rotenone resistance to LYC5 cells with the primary objective of determining whether this restored sensitivity to antitumor DSU [34]. A sec- ondary aim was to determine whether resistance to rotenone was associated with Pgp-mediated MDR. A size-selected cDNA library from parental GC3/cl ceils was constructed, and transfected into LYC5 cells. Clones selected for G418 resistance were pooled and placed under positive selection for rotenone resistance. Four colonies (designated T5LR) growing in 100 nM rotenone were expanded. Integration of plasmid DNA was confirmed in each clone by RT-PCR and Southern analysis for detection of the n e t gene of the pcDNA3 vector. Using primers designed to allow recovery of cDNA inserts, a ,,~ 1200 bp insert was identified in 3 T5LR clones. This cDNA is being characterized.
The sensitivity of T5LR clones to rotenone, antimycin and oligomycin was next examined. Each clone demonstrated resistance to each of these tox- ins, hence T5LR clones were similar to parental GC3/c 1 cells. We next tested whether T5LR rotenone- resistant clones had reverted to DSU-sensitivity using two analogs of sulofenur. As reported previously [31 ], LYC5 cells were 45- to 50-fold resistant to LY186391 and LY195779 relative to GC3/cl cells, whereas the level of resistance to sulofenur is ~ 5-fold. The rea- son for the greater differential toxicity of these analogs is their greater potency against GC3/cl cells, whereas

each analogue inhibited growth of LYC5 cells at simi-
lar concentration. Each of the rotenone-selected T5LR
clones retained the same level of resistance to these DSU analogs as did LYC5. Consequently, we con-
clude that rotenone resistance and collateral sensitivity
to antitumor DSU are not genetically linked.
F u t u r e direct ions
In retrospect, we can be relatively certain that failure to demonstrate activity in clinical trials may relate to
lower drug concentrations being achieved in patients
relative to that in mice. In part, this appears to be a con- sequence of differential metabolism between species,
leading to methemaglobinemia and anemia that was
not observed in rodents. The new analogue, LY295501,
designed to overcome these problems appears to have similar activity in the xenograft models, and studies in
both rodents and other species suggest that hemolyt- ic anemia and methemoglobinemia should be demi- nished. However, protein binding appears to be even greater than with sulofenur, which may present prob-
lems in achieving meaningful levels of free drug in
patients. Clearly the future of this class of antitumor
agents depends on some clinical activity of LY295501.
From a mechanistic standpoint, antitumor DSU remain
interesting. Our studies have been of some value in understanding the cellular pharmacology, but have not
identified a specific target, or mechanism of action. Perhaps it is fair to say that many actions of antitumor DSU have been proposed, but as yet none have been
shown to be altered or deminished in DSU-resistant clones. Such demonstration is imperative if we are to understand the basis for activity and therapeutic selec-
tivity of these novel agents.
Acknowledgements
This work was initiated by Jerry Grindey, and it has
been a privilege for us to be associated with his pro- gram at Eli Lilly. His untimely death has robbed us
of a mentor and a friend. We acknowledge also the resources made available by Eli Lilly and Company that have made these studies possible.
Supported in part by PHS award CA51949 from the
National Cancer Institute, and by American, Lebanese, Syrian Associated Charities (ALSAC).
279
References
I. Howbert JJ, Grossman CS, Crowell TA, Rieder B J, Harper RW, Kramer KE, Tao EV, Aikins J, Poore GA, Rinzel SM, Grindey GB, Shaw WN, Todd GC: Novel agents effective against solid tumors: The diarylsulfonylureas, synthesis, and activities and analysis of quantitative structure-activity relationships. J Med Chem 33:2393-2407, 1990
2. Grindey GB: Identification of diarylsulfonylureas on novel anticancer agents. Proc Am Assoc Cancer Res 29:535a, 1988
3. Houghton PJ, Sosinski J, Thakar JH, Houghton JA: Diarylsu- lonyluras: new anticancer agents with novel activities, toxic- ities and mechanism of action. Proc. Int. Congress on neoad- juvant chemotherapy. In: Banzet P, Holland JF, Kayat D, Well M (eds) Neo-adjuvant Chemotherapy. Springer Verlag, Paris, 1991, pp 405-408
4. Houghton PJ, Cheshire PJ, Myers L, Lutz L, Toth JE, Grindey GB, Houghton JA: Efficacy of sulofenur and a second gen- eration diarylsulfonylurea, N-[5-(2,3-dihydrobenzofuryl)sul- fonyl]-N_(3,4-dichlorophenyl)urea (LY295501), against colon adenocarcinoma xenografts. Anti-Cancer Drugs 6:317-323, 1995
5. Houghton PJ, Houghton JA, Myers L, Cheshire P J, How- bert JJ, Grindey GB: Evaluation of 5-(indanylsulfonyl)-N_- (4-chlorophenyl)-urea against xenografts of pediatric rhab- domyosarcoma. Cancer Chemother Pharmaco125:84-88, 1989
6. Taylor CW, Alberts DS, Ketchum MA, Satterlee WG et al.: Clinical pharmacology of a novel diarylsulfonylurea anticancer agent. J Clin Oncol 7:1733-1740, 1989
7. Brown TD, O'Rourke TJ, Kuhn JG, Craig JB, Harlin K et al.: Phase I trial of sulofenur (LY186641) given orally on a daily • schedule. Anti-Cancer Drugs 5:151-159, 1994
8. O'Brien MER, Hardy JH, Tan S, Walling J, Peters B et al.: A phase II study of sulofenur, a novel sulfonylurea, in recur- rent epithelial ovarian cancer. Cancer Chemother Pharmacol 30:245-248, 1992
9. Hainsworth J, Hande K, Satterlee W et al.: Phase I clini- cal study of N-[(4-chlorophenyl)amino]carbonyl-2,3,dihydro- 1H-indane-5-sulfonamide (LY186641). Cancer Res 49:5217- 5220, 1989
10. Ehlhardt WJ, Howbert JJ: Metabolism and disposition of p- chloroaniline in rat, mouse, and monkey. Drug Metab Disp 19:366-369, 1991
11. Ehlhardt WJ: Metabolism and disposition of the anticancer agent sulofenur in mouse, rat, monkey, and human. Drug Metab Disp 19:370--375, 1991
12. Ehlhardt WJ, Zimmerman JL, Ray JE, Toth JE: Formation of indanesulfonylureas in the metabolism of the anticancer against sulofenur in rats~ monkeys and humans. Drug Metab Disp 20:958-961, 1992
13. Zimmerman J: Primate and rat toxicology studies conducted on a new oncolytic sylfonylurea compound, LY295501. Proc Am Assoc Cancer Res 35:460a, 1994
14. Trump BF, Jain PT, Phelps PC, Berezesky IK, Boder GB: Effects of sulofenur LY186641 on the intracelhilar ionized cal- cium [Ca 2+] in colon carcinoma cells. Proc Am Assoc Cancer Res 33:177a, 1992
15. Morris CJ, Phelps PC, Berezesky IK, Merriman RL, Boder GB, Trump BF: Digital imaging fluorescence microscopy of cytosolic calcium [Ca2+], in NRK-52E and H/l-2 NRK -52E following treatment with the antineoplastic agent sulofenur (LY186641). Proc Am Assoc Cancer Res 34:1743a, 1993

280
16. Phelps PC, Morris CJ, Merriman RL, Berezesky IK, Boder GB, Trump BF: Comparative studies on the effect of two anti- neoplastic agents LY186641 (sulofenur) and LY295501, on cytosolic calcium [Ca2+], deregulation and cell death in par- ent and H-ras transfected rat kidney cell lines, NRK-52E and H/1.2 NRK-52E. Proc Am Assoc Cancer Res 35:1915a, 1994
17. Phelps PC, Jain PT, Berezesky IK, Boder GB, Trump BF: Sulofenur cytotoxicity and changes in cytosolic calcium and mitochondrial membrane potential in human colon adenocar- cinoma cell lines. Cancer Lett 88:27-35, 1995
18. Phelps PC, Morris CJ, Elliget KA, Boder GB, Trump BF: The effects of the antitumor compound sulofenur (LY186641) on the cytoskeletal network of rat kidney cell lines NRK-52E and H/1.2 NRK-52E. Proc Am Assoc Cancer Res 35:1916a, 1994
19. Gu H, Merriman RL, Berezesky IK, Boder GB, Trump BF: Induction of Fos and Juu proteins in normal rat kidney NRK- 52E and H-ras transfected NRK-52E (H/1.2 NRK-52E) cells following sulofenur treatment. Proc Am Assoc Cancer Res 35:1917a, 1994
20. Gu H, Merriman RL, Berezesky IK, Boder GB, Trump BF: Role of immediate early and stress gene expression in NRK- 52E and H/1.2 NRK-52E cells following treatment with the antineoplastic agent sulofenur (LY186641). Proc Am Assoc Cancer Res 35:1918a, 1994
21. Tannock I, Lee C, Thakar J, Houghton P: The pH-dependent activity of the diarylsulfonylurea N-(5-indanylsulfonyl)-N-(4- chlorophenyl)urea. Cell Pharmacol 2:193-198, 1995
22. Schultz RM, Andis SL, Toth JE, Boder GB, Rimzel SM, Grindey GB: Effect of albumin on antitumor activity of diaryl- sulfonylureas. Anticancer Res 13:1939-1944, 1993
23. Sosinski J, Thakar JH, Germain GS, Harwood FC, Houghton PJ: Proliferation-dependent and -independent cytotoxicity by antitumor diarylsulfonylureas may indicate multiple mecha- nisms of drug action. Biochem Pharmaco145:2135-2142, 1993
24. Houghton PJ, Bailey FC, Gerrnaln GS, Grindey GB, Houghton JA: Cytotoxic activity of N-(5-indanylsulfonyl)N_- (4-chlorophenyl)-urea: A novel antitumor agent active against non-proliferating cells. Cancer Res 50:318-322, 1990
25. Houghton PJ, Bailey FC, Germain GS, Grindey GB, Howbert JJ, Houghton JA: Studies on the cellular pharmacology of N- (4-methylphenylsulfonyl)-N-(4-chlorophenyl)-urea. Biochem Pharmacol 39:1187-1192, 1990
26. Sosinski J, Chapin C, Thakar JH, Houghton PJ: Influence of extracellular pH upon the accumulation and cytotoxici-
ty of N-(4-methylphenylsulfonyl)-N-(4-chlorophenyl)urea in human cell lines. Cancer Commun 3:373-381, 1991
27. Rockwell S, Grindey GB: Diarylsulfonyhireas and radio- therapy. I. In vitro studies with N-(5-indanylsulfonyl-N_-(4- chlorophenyl)urea (sulofenur). Cancer Commun 3:373-381, 1991
28. Houghton PJ, Bailey FC, Houghton JA, Murti KG, How- bert JJ, Grindey GB: Evidence for localization of N- (4-methylphenylsulfonyl)-N_-(4-chlorophenyl)-urea in mito- chondria of colon adenocarcinoma cells. Cancer Res 50:664- 668, 1990
29. Thakar JH, Chapin C, Berg RH, Ashmun RA, Houghton PJ: Effect of antitumor diarylsulfonylureas on in vivo and in vitro mitochondrial structure and functions. Cancer Res 51:6286- 6291, 1991
30. Rush GF, Rinzel S, Boder GB, Heim A, Toth JE, Ponsler GD: Effects of diarylsulfonylurea antitumor agents on the func- tions of mitochondria isolated from rat liver and GC3/cl cells. Biochem Pharmacol 44:2387-2394, 1992
31. Sosinski J, Thakar JH, Germaln GS, Dias P, Harwood FC, Kuttesch JF, Houghton PJ: Cross-resistance of antitumor diarylsulfonyhireas, and collateral sensitivity to mitochondrial toxins in a human cell line selected for resistance to the anti- tumor agent N-(5-indanylsulfonyl)-N_-(4-chlorophenyl)urea. Mol Pharmacol 45:962-970, 1994
32. Houghton PJ, Sosinski J, Thakar JH, Boder GB, Grindey GB: Characterization of the intracellular distribution and binding in human adenocarcinoma cells of N-(4-azidophenylsulfonyl)- N-(4-chlorophenyl)urea LY219703) a photoaffinity analogue of the antitumor diarylsulfonylurea sulofenur. Biochem Phar- macol 49:661-668, 1995
33. Wright WE: Isolation of heterokaryotes and hybrids by a selec- tive system using irreversible biochemical inhibitors. Exp Cell Res 112:395-407, 1978
34. Shu L, Houghton PJ: Separation of resistance to antitu- mor diarylsulfonylurea agents from collateral sensitivity to mitochrondrial toxins. Mol Pharm (1996, in press)
Address for offprints: Peter J. Houghton, Dept. Molecular Phar- macology, St Jude Children's Research Hospital, N. Lauderdale, Memphis, TN 38105-2794, USA. (Tel.: (901) 945 3440; Fax: (901) 521 1668; E-mail [email protected])















![EGFR as a Target for Glioblastoma Treatment: An Unfulfilled Promise … · 2017. 8. 28. · which there is only preclinical promise [34]. Brain pene-trance is difficult to measure](https://img.pdfslide.us/doc/110x75/60589686b1bd3b19386788e4/egfr-as-a-target-for-glioblastoma-treatment-an-unfulfilled-promise-2017-8-28.jpg)













