Embed Size (px)
Citation preview

Food Chemistry 120 (2010) 673–678
Contents lists available at ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Antioxidant and polyphenol oxidase activity of germinated barleyand its milling fractions
Paras Sharma, Hardeep Singh Gujral *
Department of Food Science and Technology, Guru Nanak Dev University, Amritsar 143005, India
a r t i c l e i n f o a b s t r a c t
Article history:Received 2 June 2009Received in revised form 22 September2009Accepted 20 October 2009
Keywords:Antioxidant activityBarleyPolyphenol oxidase activityTotal phenolic content
0308-8146/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.foodchem.2009.10.059
* Corresponding author. Tel.: +91 183 2258802.E-mail address: [email protected] (H.S. Gujral
Eight different hulled barley cultivars were evaluated before and after germination for total phenolic con-tent (TPC), antioxidant activity (AOA) and polyphenol oxidase (PPO) activity. Germination for 12 h signif-icantly decreased the TPC in whole flour (WF), bran (B) and refined flour (RF), however, further increase ingermination to 24 h significantly increased the TPC in the fractions. Germination for 12 h duration signif-icantly increased the AOA for WF, B and RF and further increase in germination to 24 h significantlyincreased the AOA in all the barley cultivars tested. A strong positive correlation was observed betweenAOA and TPC. Upon germination for 12 h the polyphenol oxidase activity of WF, B and RF significantlydecreased by 13.31–59.22, 1.93–59.46 and 7.08–45.87%, respectively, however, the PPO activityincreased upon germination for 24 h. The highest level of TPC and AOA was observed in the bran fractionof all the barley cultivars.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Cereals such as wheat, rice, corn, and barley in addition to beingprimary sources of carbohydrates, also provide trace minerals, die-tary fibre and bioactive compounds (Madhujith & Shahidi, 2006).Positive physiological role of cereal grains is mainly attributed todietary fibre. However, recent findings revealed that the cerealsalso contain phenolic compounds that have antioxidant activityand prevent diseases in humans (Madhujith & Shahidi, 2007). Bar-ley is consumed around the world mostly in the malted form inbrewing and bakery industry. In recent years barley has gainedpopularity due to the functional properties of its bioactive com-pounds such as b-glucan, arabinoxylan, oligosaccharides, tocolsand phenolic compounds. At present only 2% of barley is used forhuman food (Baik & Ullrich, 2008). Barley contains many phenoliccompounds in the free and bound forms including benzoic and cin-namic acid derivatives, proanthocyanidins, quinines, flavonols,chalcones, flavones, flavanones, and amino phenolic compoundswhich are concentrated in outer layer of barley grain (Goupy,Hugues, Boivin, & Amiot, 1999). The natural antioxidants in cerealsmay function as reducing agents, free radical scavengers, singletoxygen quencher and potential complexer of prooxidants.
The enzyme, polyphenol oxidase (PPO) catalyses the oxidativereaction of polyphenols that imparts discoloration in barley. PPOreacts with phenolic compounds to produce o-quinones whichfurther react with other phenolic compounds or amino acids to
ll rights reserved.
).
give discoloration in different foods made from barley (Saper,1993). The discoloration of foods by PPO limits the use of barleyin food (Lagasse, Hatcher, Dexter, Rossnagel, & Izydorczyk, 2006).Germination is a process in which physical modification of endo-sperm is carried out to increase the bioactive compounds. Theobjective of the present study was to determine the effect ofgermination duration on total phenolic content (TPC), antioxi-dant activity (AOA) and polyphenol oxidase (PPO) activity ineight Indian hulled barley cultivars and their roller millingfractions.
2. Materials and methods
2.1. Barley samples
Eight commonly grown barley hulled cultivars namely, PL-172,PL-426, RD-2503, RD-2508, RD-2035, RD-2052, RD-2552 andDWR-28, were procured from Central State Seed Farm, Sri Gang-anagar, Rajasthan, India. The grain of each variety was cleanedand stored for evaluation. All tests were performed in triplicateson a dry weight basis.
2.2. Reagents
Standard ferulic acid and 2,2-diphenyl-1-picrylhydrazyl (DPPH)were procured from Sigma–Aldrich (Steinheim, Germany). Sodiumcarbonate (analytical grade) was procured from Qualikens India

674 P. Sharma, H.S. Gujral / Food Chemistry 120 (2010) 673–678
Ltd., Folin Ciocalteu’s (analytical grade) reagent was procured fromSRL, Mumbai, India.
2.3. Germination of barley
Barley samples were steeped for 24 h at 25 �C and water waschanged at 2 h intervals. After soaking, barley seeds were allowedto germinate in a BOD incubator maintained at 25 �C and 100% RHfor 12 and 24 h. The germinated barley was dried in a dryer (LaParmigiana, Fidenza, Italy) at 40 �C to a moisture content of 10%.The dried samples were then packed in air tight bags and storedfor further analysis.
2.4. Dehusking and milling of barley
Dehusking of control and germinated barley samples was car-ried out using a rice miller (McGill, Brookshire, TX, USA) as re-ported by Singh, Singh, Kaur, and Bakshi (2000). Hulled barley(150 g) was placed in the polishing chamber and the polisherwas run till the husk was completely removed from the grain.The whole barley flour was prepared by grinding dehusked barleyin a Super Mill (Newport, Australia) and flour was sieved through250 lm sieve. The dehusked barley was further subjected to rollermilling in a Brabender Quadrument Junior Mill, (Brabender,Germany). Two fractions were obtained which were defined as re-fined flour and bran.
The whole barley flour samples were labelled as control (WF0),12 h germinated (WF12) and 24 h germinated (WF24), the refinedbarley flour was labeled as control (RF0), 12 h germinated (RF12)and 24 h germinated (RF24). The barley bran was labeled as control(B0), 12 h germinated (B12) and 24 h germinated (B24). The branwas ground in an electrical grinder (Sujata, India) to pass through250 lm sieve before analysis.
2.5. Total phenolic content (TPC)
The total phenolic content was determined according theFolin–Ciocalteu specterophotometric method explained by Gao,Wang, Oomah, and Mazza (2002). Barley flour samples (200 mg)were extracted with 4 ml acidified methanol (HCl/methanol/water, 1:80:10, v/v/v) at room temperature (25 �C) for 2 h usingwrist action shaker (Narang Scientific, Delhi, India). The mixturewas centrifuged at 3000g for 10 min on a centrifuge (REMI, C24, Mumbai, India). The supernatant was used for determinationof total phenolic content. Aliquot of extract (200 ll) was addedto 1.5 ml freshly diluted (10-fold) Folin–Ciocalteu reagent. Themixture was allowed to equilibrate for 5 min and then mixedwith 1.5 ml of sodium carbonate solution (60 g/l). After incuba-tion at room temperature (25 �C) for 90 min, the absorbance ofthe mixture was read at 725 nm. Acidified methanol was usedas a blank. The results were expressed as lg of ferulic acid equiv-alents per gram of flour.
2.6. Antioxidant activity
Antioxidant activity was measured using a modified versionof the method explained by Brand-Williams, Cuvelier, andBerset (1995). This involved the use of free radical 2,2-diphe-nyl-1-picrylhydrazyl (DPPH) solution in the methanol. Groundbarley samples (100 mg) were extracted with 1 ml methanolfor 2 h and centrifuged at 3000g for 10 min. The supernatant(100 ll) was reacted with 3.9 ml of a 6 � 10�5 mol/l of DPPHsolution. Absorbance (A) at 515 nm was read at 0 and 30 minusing a methanol blank. Antioxidant activity was calculated as% discoloration.
% Antioxidant activity ¼ ð1� ðA of samplet¼30=A of controlt¼0ÞÞ� 100:
2.7. Polyphenol oxidase activity
Polyphenol oxidase activity was determined using the standardmethod 22-85 of the AACC (2000) by measuring the absorbance at475 (A475) nm using a spectrophotometer (Shimadzu, UV-2450,Kyoto, Japan). Control barley samples were assayed without L-3-(3,4-dihydroxyphenyl) alanine substrate. The PPO activity was cal-culated as difference in the absorbance of sample and control andexpressed as D475/min g flour.
2.8. Statistical analysis
Analysis of variance was carried out and Fishers least significantdifference test was used to describe means with 95% confidence.The Pearson correlation coefficients were calculated by SPSS soft-ware at a probability level of p < 0.05.
3. Results and discussion
3.1. Total phenolic content
Total phenolic content (TPC) in whole barley flour (WF0) variedsignificantly among cultivars (Table 1). The highest TPC was ob-served in PL-426 cultivar (control) with 4480 lg (ferulic acidequivalents, FAE)/g while DWR-28 showed the lowest TPC(3070 lg FAE/g). Madhujith and Shahidi (2006) reported TPC rang-ing from 0.81 to 1.38 mg of ferulic acid equiv/g in defatted barleyflour. The total phenolic content significantly decreased in all thecultivars upon germination for 12 h. Barley cultivar PL-172 showedthe highest reduction in TPC whereas the lowest reduction was ob-served for RD-2503. A similar result was observed for different bar-ley cultivars during steeping and germination by Lu et al. (2007).The TPC of whole flour from barley germinated for 12 h (WF12) var-ied significantly among the cultivars. The highest TPC was ob-served for PL-426 (2718 lg FAE/g) and it was the lowest forDWR-28 (2359 lg FAE/g). When the duration of germination in-creased to 24 h the TPC significantly (p < 0.05) increased for allthe cultivars (Table1). The highest increase in TPC of 24 h germi-nated whole flour (WF24) was observed for DWR-28 and the lowestwas for PL-172. The TPC varied significantly (p < 0.05) among cul-tivars for the bran from control barley (Table 2) and ranged from3574 to 5021 lg FAE/g. The highest and the lowest TPC was ob-served in PL-426 and DWR-28, respectively. The total phenoliccontent in the bran from barley germinated for 12 h (B12) signifi-cantly decreased among all cultivars with the highest decrease ob-served for PL-426 while RD-2503 showed the lowest decrease.DWR-28 showed a minimum TPC (3410 lg FAE/g) among cultivarsin the bran from B12. Further increase in duration of germination to24 h significantly increased the total phenolic content in barleybran (B24). Barley cultivar DWR-28 showed a maximum increasein TPC while the lowest increase was observed for RD-2503. A sig-nificant change was noticed in total phenolic content among culti-vars in the bran from barley germinated for 24 h and it ranged from4180 to 4689 lg FAE/g. The highest TPC was observed in RD-2552and the lowest in RD-2503. Madhujith and Shahidi (2007) reportedthe TPC value of 0.36–6.26 mg FAE/g in different pearling fractionsof barley. In the present study the bran obtained after roller millingwas contaminated with shorts or fragment of endosperm as barleydoes not have true bran as in wheat (Bhatty, 1993). Madhujith andShahidi (2008) reported a TPC of 4.16 and 6.26 mg ferulic acidequivalents/g in the outer most layers of two different barley
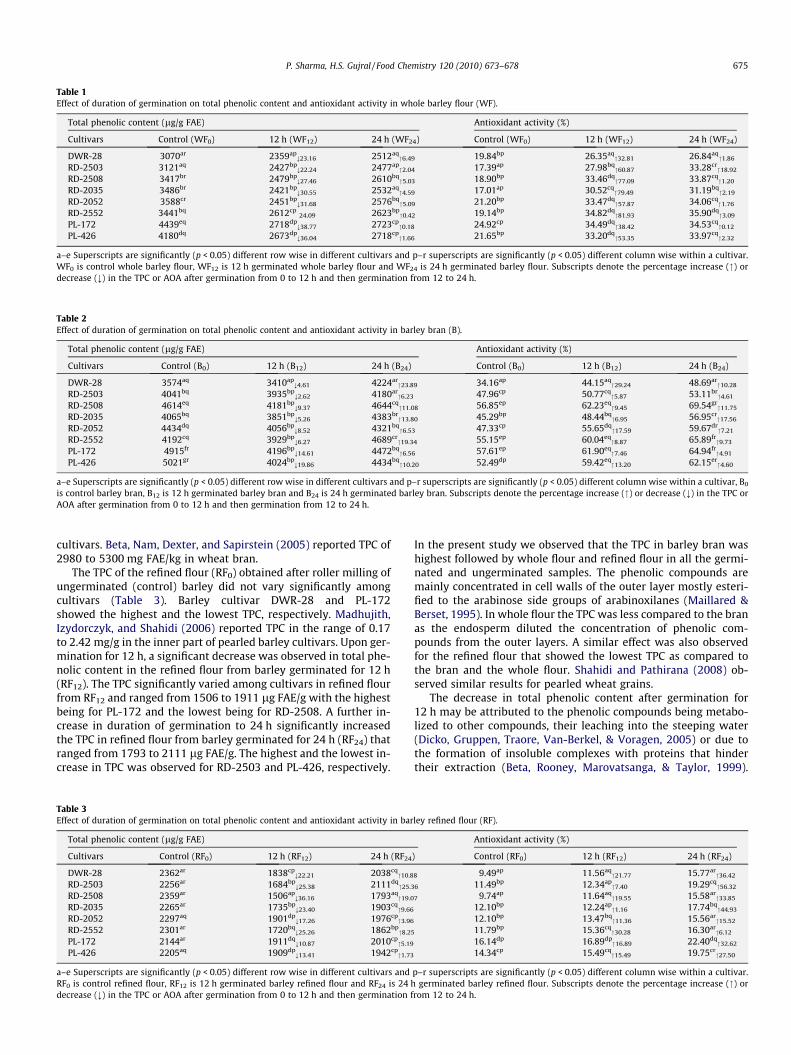
Table 1Effect of duration of germination on total phenolic content and antioxidant activity in whole barley flour (WF).
Total phenolic content (lg/g FAE) Antioxidant activity (%)
Cultivars Control (WF0) 12 h (WF12) 24 h (WF24) Control (WF0) 12 h (WF12) 24 h (WF24)
DWR-28 3070ar 2359ap;23.16 2512aq
"6.49 19.84bp 26.35aq"32.81 26.84aq
"1.86
RD-2503 3121aq 2427bp;22.24 2477ap
"2.04 17.39ap 27.98bq"60.87 33.28cr
"18.92
RD-2508 3417br 2479bp;27.46 2610bq
"5.03 18.90bp 33.46dq"77.09 33.87cq
"1.20
RD-2035 3486br 2421bp;30.55 2532aq
"4.59 17.01ap 30.52cq"79.49 31.19bq
"2.19
RD-2052 3588cr 2451bp;31.68 2576bq
"5.09 21.20bp 33.47dq"57.87 34.06cq
"1.76
RD-2552 3441bq 2612cp24.09 2623bp
"0.42 19.14bp 34.82dq"81.93 35.90dq
"3.09
PL-172 4439eq 2718dp;38.77 2723cp
"0.18 24.92cp 34.49dq"38.42 34.53cq
"0.12
PL-426 4180dq 2673dp;36.04 2718cp
"1.66 21.65bp 33.20dq"53.35 33.97cq
"2.32
a–e Superscripts are significantly (p < 0.05) different row wise in different cultivars and p–r superscripts are significantly (p < 0.05) different column wise within a cultivar.WF0 is control whole barley flour, WF12 is 12 h germinated whole barley flour and WF24 is 24 h germinated barley flour. Subscripts denote the percentage increase (") ordecrease (;) in the TPC or AOA after germination from 0 to 12 h and then germination from 12 to 24 h.
Table 2Effect of duration of germination on total phenolic content and antioxidant activity in barley bran (B).
Total phenolic content (lg/g FAE) Antioxidant activity (%)
Cultivars Control (B0) 12 h (B12) 24 h (B24) Control (B0) 12 h (B12) 24 h (B24)
DWR-28 3574aq 3410ap;4.61 4224ar
"23.89 34.16ap 44.15aq"29.24 48.69ar
"10.28
RD-2503 4041bq 3935bp;2.62 4180ar
"6.23 47.96cp 50.77cq"5.87 53.11br
"4.61
RD-2508 4614eq 4181bp;9.37 4644cq
"11.08 56.85ep 62.23eq"9.45 69.54gr
"11.75
RD-2035 4065bq 3851bp;5.26 4383br
"13.80 45.29bp 48.44bq"6.95 56.95cr
"17.56
RD-2052 4434dq 4056bp;8.52 4321bq
"6.53 47.33cp 55.65dq"17.59 59.67dr
"7.21
RD-2552 4192cq 3929bp;6.27 4689cr
"19.34 55.15ep 60.04eq"8.87 65.89fr
"9.73
PL-172 4915fr 4196bp;14.61 4472bq
"6.56 57.61ep 61.90eq"7.46 64.94fr
"4.91
PL-426 5021gr 4024bp;19.86 4434bq
"10.20 52.49dp 59.42eq"13.20 62.15er
"4.60
a–e Superscripts are significantly (p < 0.05) different row wise in different cultivars and p–r superscripts are significantly (p < 0.05) different column wise within a cultivar, B0
is control barley bran, B12 is 12 h germinated barley bran and B24 is 24 h germinated barley bran. Subscripts denote the percentage increase (") or decrease (;) in the TPC orAOA after germination from 0 to 12 h and then germination from 12 to 24 h.
P. Sharma, H.S. Gujral / Food Chemistry 120 (2010) 673–678 675
cultivars. Beta, Nam, Dexter, and Sapirstein (2005) reported TPC of2980 to 5300 mg FAE/kg in wheat bran.
The TPC of the refined flour (RF0) obtained after roller milling ofungerminated (control) barley did not vary significantly amongcultivars (Table 3). Barley cultivar DWR-28 and PL-172showed the highest and the lowest TPC, respectively. Madhujith,Izydorczyk, and Shahidi (2006) reported TPC in the range of 0.17to 2.42 mg/g in the inner part of pearled barley cultivars. Upon ger-mination for 12 h, a significant decrease was observed in total phe-nolic content in the refined flour from barley germinated for 12 h(RF12). The TPC significantly varied among cultivars in refined flourfrom RF12 and ranged from 1506 to 1911 lg FAE/g with the highestbeing for PL-172 and the lowest being for RD-2508. A further in-crease in duration of germination to 24 h significantly increasedthe TPC in refined flour from barley germinated for 24 h (RF24) thatranged from 1793 to 2111 lg FAE/g. The highest and the lowest in-crease in TPC was observed for RD-2503 and PL-426, respectively.
Table 3Effect of duration of germination on total phenolic content and antioxidant activity in bar
Total phenolic content (lg/g FAE)
Cultivars Control (RF0) 12 h (RF12) 24 h (RF24
DWR-28 2362ar 1838cp;22.21 2038cq
"10.8
RD-2503 2256ar 1684bp;25.38 2111dq
"25.3
RD-2508 2359ar 1506ap;36.16 1793aq
"19.0
RD-2035 2265ar 1735bp;23.40 1903cq
"9.66
RD-2052 2297aq 1901dp;17.26 1976cp
"3.96
RD-2552 2301ar 1720bq;25.26 1862bp
"8.25
PL-172 2144ar 1911dq;10.87 2010cp
"5.19
PL-426 2205aq 1909dp;13.41 1942cp
"1.73
a–e Superscripts are significantly (p < 0.05) different row wise in different cultivars andRF0 is control refined flour, RF12 is 12 h germinated barley refined flour and RF24 is 24 hdecrease (;) in the TPC or AOA after germination from 0 to 12 h and then germination f
In the present study we observed that the TPC in barley bran washighest followed by whole flour and refined flour in all the germi-nated and ungerminated samples. The phenolic compounds aremainly concentrated in cell walls of the outer layer mostly esteri-fied to the arabinose side groups of arabinoxilanes (Maillared &Berset, 1995). In whole flour the TPC was less compared to the branas the endosperm diluted the concentration of phenolic com-pounds from the outer layers. A similar effect was also observedfor the refined flour that showed the lowest TPC as compared tothe bran and the whole flour. Shahidi and Pathirana (2008) ob-served similar results for pearled wheat grains.
The decrease in total phenolic content after germination for12 h may be attributed to the phenolic compounds being metabo-lized to other compounds, their leaching into the steeping water(Dicko, Gruppen, Traore, Van-Berkel, & Voragen, 2005) or due tothe formation of insoluble complexes with proteins that hindertheir extraction (Beta, Rooney, Marovatsanga, & Taylor, 1999).
ley refined flour (RF).
Antioxidant activity (%)
) Control (RF0) 12 h (RF12) 24 h (RF24)
8 9.49ap 11.56aq"21.77 15.77ar
"36.42
6 11.49bp 12.34ap"7.40 19.29cq
"56.32
7 9.74ap 11.64aq"19.55 15.58ar
"33.85
12.10bp 12.24ap"1.16 17.74bq
"44.93
12.10bp 13.47bq"11.36 15.56ar
"15.52
11.79bp 15.36cq"30.28 16.30ar
"6.12
16.14dp 16.89dp"16.89 22.40dq
"32.62
14.34cp 15.49cq"15.49 19.75cr
"27.50
p–r superscripts are significantly (p < 0.05) different column wise within a cultivar.germinated barley refined flour. Subscripts denote the percentage increase (") or
rom 12 to 24 h.

676 P. Sharma, H.S. Gujral / Food Chemistry 120 (2010) 673–678
The increase in TPC upon germination for 24 h could be attributedto the bound phenolic compounds becoming free by the action ofenhanced hydrolytic enzyme activity (Maillard, Soum, Boivin, &Berset, 1996). Shibuya (1984) reported that the increase in pheno-lic compounds in germinated brown rice could be due to the in-crease in the free phenolics forms due to dismantling of the cellwall during germination. Tian, Nakamura, and Kayahara (2004) re-ported that during germination of rice the carbohydrase enzymeshydrolysed the starch to release the bound phenolic compoundsincreasing the TPC. They also reported that during germination,free phenolic acid content increased significantly; the ferulic acidcontent of brown rice also increased and became the most abun-dant phenolic compound in germinated brown rice.
3.2. Antioxidant activity
The DPPH is a stable free radical widely used to determine theantioxidant properties or radical scavenging activity. The antioxi-dant activity of barley flour significantly varied among cultivarsfor whole flour from control barley (WF0) (Table 1) and rangedfrom 17.01% to 24.92%. Barley cultivar PL-172 showed the highestantioxidant activity while RD-2035 exhibited the lowest. Therewas a significant (p < 0.05) positive correlation (R2 = 0.82) betweenTPC and antioxidant activity for WF0. Lu et al. (2007) reported a po-sitive correlation between the total phenolic content and antioxi-dant activity of wheat. Beta et al. (2005) also reported a positivecorrelation between TPC and AOA of different roller milling frac-tions of wheat.
The germination of barley up to 12 h increased the antioxidantactivity in the whole flour from barley germinated for 12 h (WF12)of all the cultivars the highest increase was observed in RD-2552and the lowest in DWR-28. A significant positive (p < 0.05) correla-tion (R2 = 0.757) was found between TPC and antioxidant activity.
There was no significant increase in the AOA upon further ger-mination for 24 h except in the case of RD-2503. However, the AOAof WF24 significantly varied among the cultivars and ranged from26.84% to 35.90%. The highest AOA was observed in RD-2552 andthe lowest in DWR-28. Liu and Yao (2007) reported that theDPPH-radical scavenging activity ranged from 15% to 50% whenbarley flour was extracted with 70% methanol. Lu et al. (2007) re-ported that the raw barley showed DPPH-radical scavenging activ-ity of 11.90 to 12.56 lmol trolox equivalents/g dry weight; theyalso reported that germination for a longer time increased theAOA. A correlation coefficient of R2 = 0.494 was observed betweentotal phenolic content and antioxidant activity in whole flour frombarley germinated for 24 h.
The antioxidant activity significantly varied among cultivars forbran, B0 which ranged from 34.16% to 57.61% (Table 2). The highestAOA was observed for PL-172 whereas the lowest was for DWR-28.A significant (p < 0.05) positive correlation (R2 = 0.801) was ob-served between TPC and AOA in B0. A similar correlation betweenTPC and AOA also existed in sorghum (Dicko et al., 2005). When theduration of germination was increased up to 12 h the antioxidantactivity significantly increased by 5.86–29.24% in bran from allthe cultivars of barley germinated for 12 h (B12) (Table 2). Thehighest increase was observed for DWR-28 and the lowest wasfor RD-2503. After germination for 12 h a significant change wasalso observed among the cultivars for bran (B12). The AOA rangedfrom 44.15% to 62.23% with the highest and the lowest being forRD-2508 and DWR-28, respectively. The TPC and AOA showed apositive correlation (R2 = 0.852) for bran (B12). Increase in durationof germination to 24 h, increased the AOA and the AOA rangedfrom 48.69% to 69.54% with the highest and the lowest being ob-served for RD-2508 and DWR-28, respectively. A significant posi-tive correlation (R2 = 0.903) was observed between TPC and AOAin the bran from barley germinated for 24 h (B24). The outermost
layer of barley cultivar showed the highest antioxidant activity(46–69%) as compared to the inner part of pearled barley byMadhujith et al. (2006).
The antioxidant activity varied significantly among cultivars forrefined flour (RF0) (Table 3). The highest and the lowest being ob-served for PL-172 and DWR-28 cultivar, respectively. The germina-tion up to 12 h significantly increased the AOA in all the cultivars.The highest percentage increase in AOA was showed by RD-2552while it was the lowest for RD-2035 (Table 3). A significant changewas observed in antioxidant activity among cultivars for refinedflour from barley germinated for 12 h (RF12). The antioxidant activ-ity ranged from 11.56% to 16.89% and the highest and the lowestAOA was exhibited by PL-172 and DWR-28, respectively.
When the duration of germination was increased up to 24 h theantioxidant activity significantly increased in all cultivars for re-fined flour from barley germinated for 24 h (RF24). The antioxidantactivity of germinated refined barley flour (RF24) showed a signifi-cant variation among cultivars and ranged from 15.56% to 22.40%.The highest and the lowest AOA of refined flour (RF24) was shownby PL-172 and RD-2052, respectively. Beta et al. (2005) reportedthat the inner part of wheat kernel had a low TPC and AOA activityas compared to the outer fraction. The observed DPPH-radical scav-enging activity ranged from 2.5% to 5.3% for pearled wheatcultivars.
In contrast to the total phenolic content the antioxidant activitycontinuously increased in whole flour, bran and refined flour of allthe cultivars as duration of their germination progressed. Thiscould be due to the synthesis of compounds like vitamin C and toc-opherols which are responsible for antioxidant activity (Dickoet al., 2005). During germination the hydrolytic enzymes modifythe endosperm and may liberate some of the bound componentswhich play a role in antioxidant activity (Doblado, Frias, &Valverde, 2007). The correlation between TPC and AOA for WF0,WF12 and WF24 (R2 = 0.82, 0.75 and 0.50, respectively) graduallydecreased as germination time increased. The germination signifi-cantly increased the ascorbic acid in sorghum (Malleshi & Klopfen-stein, 1998). On the other hand, correlation coefficient betweenTPC and AOA for B0, B12 and B24 increased (R2 = 0.80, 0.85 and0.90, respectively) with increase in the duration of germination.Bran is a rich source of free and bound phenolic compounds there-fore the bound phenolic compounds are released during germina-tion and increase the antioxidant activity. Maillard et al. (1996)also observed similar results for barley and malt.
3.3. Polyphenol oxidase activity
The polyphenol oxidase activity is associated with the discolor-ation of the products made from barley flour (Kruger, Hatcher, &DePauw, 1994). The polyphenol oxidase (PPO) activity of wholebarley flour (WF0) significantly (p < 0.05) varied among cultivars(Fig. 1a) and ranged from 0.147 to 0.382 D475/min g. The highestvalue of PPO activity was observed for RD-2503 while the lowestwas noted for RD-2552. A negative correlation (R2 = �0.41,p < 0.05) was observed between the PPO activity and lightness ofthe whole flour from control barley (L*, Hunter colour value, datanot reported). Quinde-Axtell and Baik (2006) also reported a similarnegative correlation between PPO activity and L* of dough sheetmade from barley flour. Increasing the duration of germination to12 h significantly lowered the PPO activity in all the cultivarstested. The highest decrease was observed in RD-2552 and the low-est in RD-2035 barley cultivar. The PPO activity of barley germi-nated for 12 h (WF12) ranged from 0.115 to 0.255 D475 /min gand significantly varied among cultivars with the highest and thelowest being for RD-2508 and DWR-28, respectively. There was anegative correlation (R2 = �0.55) between the PPO activity of wholeflour (WF12) and L* of barley grain germinated for 12 h. Increase in
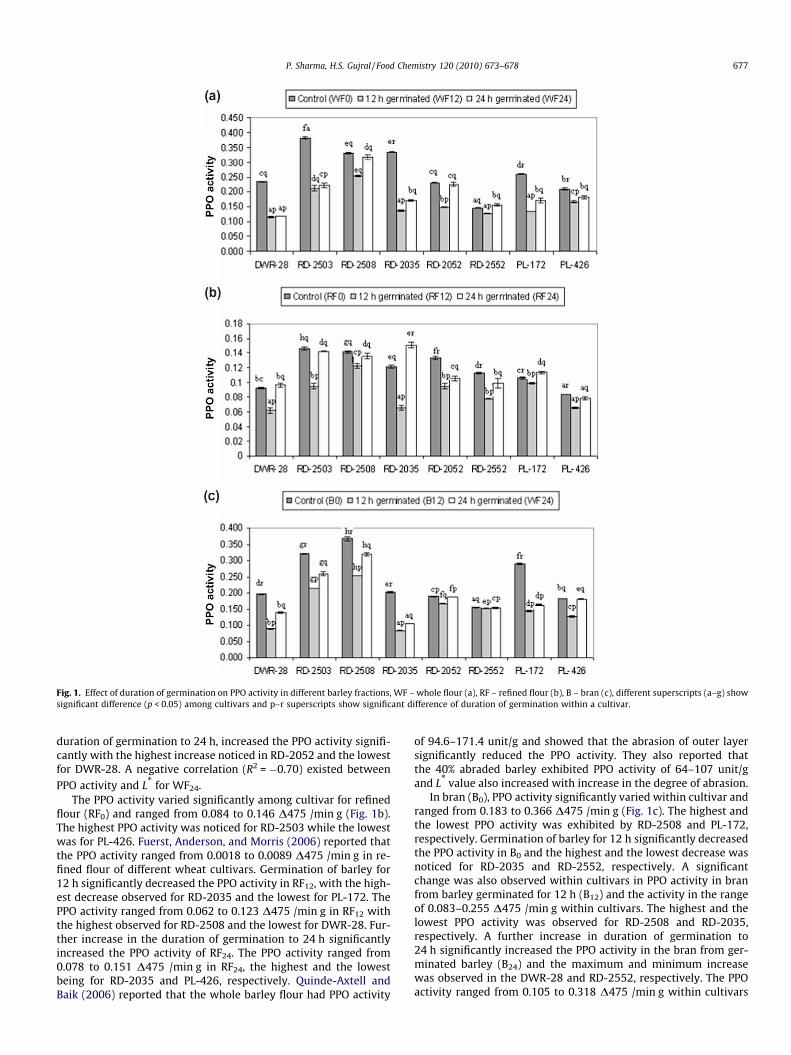
Fig. 1. Effect of duration of germination on PPO activity in different barley fractions, WF – whole flour (a), RF – refined flour (b), B – bran (c), different superscripts (a–g) showsignificant difference (p < 0.05) among cultivars and p–r superscripts show significant difference of duration of germination within a cultivar.
P. Sharma, H.S. Gujral / Food Chemistry 120 (2010) 673–678 677
duration of germination to 24 h, increased the PPO activity signifi-cantly with the highest increase noticed in RD-2052 and the lowestfor DWR-28. A negative correlation (R2 = �0.70) existed betweenPPO activity and L* for WF24.
The PPO activity varied significantly among cultivar for refinedflour (RF0) and ranged from 0.084 to 0.146 D475 /min g (Fig. 1b).The highest PPO activity was noticed for RD-2503 while the lowestwas for PL-426. Fuerst, Anderson, and Morris (2006) reported thatthe PPO activity ranged from 0.0018 to 0.0089 D475 /min g in re-fined flour of different wheat cultivars. Germination of barley for12 h significantly decreased the PPO activity in RF12, with the high-est decrease observed for RD-2035 and the lowest for PL-172. ThePPO activity ranged from 0.062 to 0.123 D475 /min g in RF12 withthe highest observed for RD-2508 and the lowest for DWR-28. Fur-ther increase in the duration of germination to 24 h significantlyincreased the PPO activity of RF24. The PPO activity ranged from0.078 to 0.151 D475 /min g in RF24, the highest and the lowestbeing for RD-2035 and PL-426, respectively. Quinde-Axtell andBaik (2006) reported that the whole barley flour had PPO activity
of 94.6–171.4 unit/g and showed that the abrasion of outer layersignificantly reduced the PPO activity. They also reported thatthe 40% abraded barley exhibited PPO activity of 64–107 unit/gand L* value also increased with increase in the degree of abrasion.
In bran (B0), PPO activity significantly varied within cultivar andranged from 0.183 to 0.366 D475 /min g (Fig. 1c). The highest andthe lowest PPO activity was exhibited by RD-2508 and PL-172,respectively. Germination of barley for 12 h significantly decreasedthe PPO activity in B0 and the highest and the lowest decrease wasnoticed for RD-2035 and RD-2552, respectively. A significantchange was also observed within cultivars in PPO activity in branfrom barley germinated for 12 h (B12) and the activity in the rangeof 0.083–0.255 D475 /min g within cultivars. The highest and thelowest PPO activity was observed for RD-2508 and RD-2035,respectively. A further increase in duration of germination to24 h significantly increased the PPO activity in the bran from ger-minated barley (B24) and the maximum and minimum increasewas observed in the DWR-28 and RD-2552, respectively. The PPOactivity ranged from 0.105 to 0.318 D475 /min g within cultivars

678 P. Sharma, H.S. Gujral / Food Chemistry 120 (2010) 673–678
for B24. The highest PPO activity in B24 was observed for RD-2508and the lowest for RD-2035.
The PPO is thoroughly distributed in the barley kernel althoughthe concentration of PPO is high in the outer layers (Bendelow &Laberge, 1979). The decrease in PPO activity after 12 h of germina-tion could be attributed to leaching of PPO in soaking water andformation of complex with protein and further increase in germi-nation activated the PPO enzyme, a similar result was also reportedfor germinated wheat (Demeke, Chang, & Morris, 2001).
4. Conclusions
The germination of barley up to12 h significantly decreased thetotal phenolic content in all the barley cultivars tested and theirmilling fractions. However, increasing the duration of germinationfrom 12 to 24 h significantly increased the TPC in all the cultivarsand their fractions. On other hand the antioxidant activity of barleysignificantly increased with duration of germination up to 24 h. Astrong positive correlation existed between total phenolic contentand antioxidant activity. The polyphenol oxidase activity signifi-cantly decreased in the all cultivars and their milling fractions after12 h germination, while germination for 24 h significantly in-creased the PPO activity in all cultivars and their fractions.
Reference
American Association of Cereal Chemists (erican Association of Cereal Chemists).Approved methods of the AACC, Method 22–85 (10 ed.). St. Paul, MN, USA:American Association of Cereal Chemists.
Baik, B. K., & Ullrich, S. E. (2008). Barley for food: Characteristics, improvement, andrenewed interest. Critical review. Journal of Cereal Science, 30, 1–10.
Bendelow, V. M., & LaBerge, D. E. (1979). Relationship among barley, malt and beerphenolics. Journal of American Society of Brewing Chemists, 37, 89–90.
Beta, T., Nam, S., Dexter, J. E., & Sapirstein, H. D. (2005). Phenolic content andantioxidant activity of pearled wheat fractions. Cereal Chemistry, 82(4),390–393.
Beta, T., Rooney, L. W., Marovatsanga, L. T., & Taylor, J. R. N. (1999). Phenoliccompounds and kernel characteristics of Zimbabwean sorghum. Journal of theScience of Food and Agriculture, 79, 1003–1010.
Bhatty, R. S. (1993). Physicochemical properties of roller milled b flour and bran.Cereal Chemistry, 70, 397–402.
Brand-Williams, W., Cuvelier, M. E., & Berset, C. (1995). Use of a free radical methodto evaluate antioxidant activity. Lebensmittel Wissenschaft and Technologie, 28,245–251.
Demeke, T., Chang, H. G., & Morris, C. F. (2001). Effect of germination, seed abrasionand seed size on polyphenol oxidase assay activity in wheat. Plant Breeding, 120,369–373.
Dicko, M. H., Gruppen, H., Traore, A. S., Van-Berkel, W. J. H., & Voragen, A. G. J.(2005). Evaluation of the effect of germination on phenolic compounds andantioxidant activity in sorghum varieties. Journal of Agriculture and FoodChemistry, 53, 2581–2588.
Doblado, R., Frias, J., & Valverde, C. V. (2007). Changes in vitamin C content andantioxidant capacity of raw and germinated (Vigna sinensis var. carilla) seedsinduced by high pressure treatment. Food Chemistry, 101, 918–923.
Fuerst, E. P., Anderson, J. V., & Morris, C. F. (2006). Polyphenol oxidase in wheatgrain: Whole kernel and bran assay for total and soluble activity. CerealChemistry, 83, 10–16.
Gao, L., Wang, S., Oomah, B. D., & Mazza, G. (2002). Wheat quality: Antioxidantactivity of wheat millstreams. In P. Ng & C. W. Wrigley (Eds.), Wheat qualityelucidation (pp. 219–233). St Paul, MN: AACC International.
Goupy, P., Hugues, M., Boivin, P., & Amiot, M. J. (1999). Antioxidant composition andactivity of barley (Hordeum vulgare) and malt extracts and of isolated phenoliccompounds. Journal of Science of Food and Agriculture, 79, 1625–1634.
Kruger, J. E., Hatcher, D. W., & DePauw, R. (1994). A whole seed assay for polyphenoloxidase in Canadian prairie spring wheat and its usefulness as a measure ofnoodle darkening. Cereal Chemistry, 71, 324–326.
Lagasse, S. L., Hatcher, D. W., Dexter, J. E., Rossnagel, B. G., & Izydorczyk, M. S. (2006).Quality characteristics of fresh and dried white salted noodles enriched withflour from hull-less barley genotype of diverse amylose content. CerealChemistry, 83, 202–210.
Liu, Q., & Yao, H. (2007). Antioxidant activity of barley seeds extracts. FoodChemistry, 102, 732–737.
Lu, J., Zhao, H., Chen, J., Fan, W., Dong, J., Kong, W., et al. (2007). Evolution ofphenolic compounds and antioxidant activity during malting. Journal ofAgriculture and Food Chemistry, 55, 10994–11001.
Madhujith, T., Izydorczyk, M., & Shahidi, F. (2006). Antioxidant activity of pearledbarley fraction. Journal of Agriculture and Food Chemistry, 54, 3283–3289.
Madhujith, T., & Shahidi, F. (2006). Antioxidant properties of barley pearledfractions. Journal of Agriculture and Food Chemistry, 54, 3283–3289.
Madhujith, T., & Shahidi, F. (2007). Antioxidative and antiproliferative properties ofselected barley (Hordeum vulgare L.) cultivars and their potential for inhibitionof low density lipoprotein (LDL) cholesterol oxidation. Journal of Agriculture andFood Chemistry, 55, 5018–5024.
Madhujith, T., & Shahidi, F. (2008). Antioxidative and antiproliferative potential ofpearled barley (Hordeum vulgare L.). Pharmaceutical Biology, 46, 88–95.
Maillard, M. N., Soum, M. H., Boivin, P., & Berset, C. (1996). Antioxidant activity ofbarley and malt: Relationship with phenolic content. Lebensmittel Wissenschaftand Technologie, 29, 238–244.
Maillared, M. E., & Berset, E. (1995). Evolution of the antioxidant activity duringkilning: The role of insoluble and bound phenolic acids of barley and malt.Journal of Agriculture and Food Chemistry, 43, 1789–1793.
Malleshi, N. G., & Klopfenstein, C. F. (1998). Nutrient composition, amino acid andvitamin content of malted sorghum, pearl millet, finger millet, and theirrootlets. International Journal Food Science Nutrition, 49, 415–422.
Quinde-Axtell, Z., & Baik, B. K. (2006). Phenolic compounds of barley grain and theirimplication in food products discoloration. Journal of Agricultural and FoodChemistry, 54, 9978–9984.
Saper, G. M. (1993). Browning of foods: Control by sulfites, antioxidants, and othermeans. Food Technology, 47, 75–84.
Shahidi, F., & Pathirana, C. L. (2008). Antioxidant properties of wheat grain and itsfraction. In L. L. Yu (Ed.), Wheat antioxidants (pp. 7–17). New York: Wiley.
Shibuya, N. (1984). Phenolic acids and their carbohydrate esters in rice endospermcell wall. Journal of Phytochemistry, 23, 2233–2237.
Singh, N., Singh, H., Kaur, K., & Bakshi, M. S. (2000). Relationship between the degreeof milling, ash distribution pattern and conductivity in brown rice. FoodChemistry, 69, 147–151.
Tian, S., Nakamura, K., & Kayahara, H. (2004). Analysis of phenolic compounds inwhite rice, brown rice, and germinated brown rice. Journal of Agricultural andFood Chemistry, 52, 4808–4813.




![Phenolic Profiles and Polyphenol Oxidase (PPO) Gene ... · p-ethylphenol, a metabolite of biochanin A, have been linked to decreased bovine reproduction . However, [15] biochanin](https://img.pdfslide.us/doc/110x75/607469627bc5c160f7329b6c/phenolic-profiles-and-polyphenol-oxidase-ppo-gene-p-ethylphenol-a-metabolite.jpg)












![Novel Roles for the Polyphenol Oxidase Enzyme in ......Novel Roles for the Polyphenol Oxidase Enzyme in Secondary Metabolism and the Regulation of CellDeathinWalnut1[W][OPEN] Soha](https://img.pdfslide.us/doc/110x75/5f1a449c92fe23609900d20e/novel-roles-for-the-polyphenol-oxidase-enzyme-in-novel-roles-for-the-polyphenol.jpg)











