Embed Size (px)
Citation preview

Antimicrobial lipopeptides: in vivo and in vitro synthesis
A. Etchegaray1 and M. T. Machini2,*
1Faculty of Chemistry, Pontifical Catholic University of Campinas, Rod. D. Pedro I Km 136, Pq. das Universidades, Campinas, SP, 13086-900, Brazil
2Department of Biochemistry, Institute of Chemistry, University of São Paulo, Av. Prof. Lineu Prestes, 748, B8 Sup., sala 0854, São Paulo, SP, 05508-000, Brazil
*Also cited as Miranda, MTM.
The non-ribosomally assembled lipopeptides display a number of biological activities, including antibiotic, antitumor, immune-modulating and immune-suppressive activities. This review starts with general comments on antimicrobial peptides (AMPs), a large family of compounds that includes non-ribosomally assembled lipopeptides. Then, it brings together information on in vivo synthesis (biosynthesis) of such hybrid biomolecules using as models the surfactins, potent anionic biosurfactants produced by Bacillus subtilis that have potential environmental and medical applications, but limited productivity due to low yields and the high costs associated with microbial growth and surfactant recovery. It follows a brief statement on how important is to overcome those limitations as well as a discussion about in vitro synthesis (chemical synthesis) of this type of compounds. Hence, the methods currently used for making peptides are outlined and given examples of lipopeptides and analogues that have been chemically synthesized by the solid-phase method using the Fmoc-based chemistry. It becomes clear that most of these compounds have only been successfully produced in small scale and that a challenge is set up to upscale productivity of non-ribosomally assembled lipopeptides with low costs, high yields and, hopefully, using environmental friendly methods, techniques and conditions. In this context, modifications of the protocols used for non-ribosomal synthesis and microwave-assisted chemical synthesis seem to be promising alternatives.
Keywords antimicrobial peptides; non-ribosomal peptides; lipopeptides; solid-phase peptide synthesis
1. Antimicrobial peptides
The continually evolving resistant microbes are challenges for medical care to overcome with novel antimicrobials. Therefore, the research on discovery, properties, production and mechanisms of action of bactericidal and/or fungicidal compounds has become more urgent than ever. To better appraise such urgency, one should look throughout time how many lives have been saved since the discovery of penicillin [1]. Among the natural antibiotics known so far [2,3] is the family of antimicrobial peptides (AMPs), which is composed of several subfamilies [4-6]. Based on how they are biosynthesized, these immune-defense molecules can also be classified in ribosomally and non-ribosomally assembled peptides [3]. The first class has a large number of members. Hundreds of them have been extensively studied worldwide [7], including those termed bacteriocins [5]. However, probably mostly due to their high salt-sensitivity and susceptibility to proteolysis, only a few have been approved for use in therapeutics or in food industry and are now in the market [8]. As to the non-ribosomally assembled peptides, many of them have already been commercialized and used, such as vancomycins (glycopeptides [9]), tyrocidines (cyclic peptides; [10]), gramicidins (cationic cyclic peptides [11]) and polymyxins (cyclic lipopeptide [8]). The last decade has seen a growing interest in anionic or cationic AMPs covalently attached to a fatty acid moiety (lipopeptides), also members of this class, as is the case of polymyxins [12].
2. Lipopeptides
Lipopeptides are linear or cyclic peptides acylated by a lipid, usually a fatty acid side chain (examples are shown in Figure 1). It has been described that these compounds are produced only in bacteria and fungi of various habitats during cultivation on carbon and nitrogen sources [13,14]. The literature is rich in descriptions of lipopeptide biological actions, such as antibacterial [15], antifungal [16,17], improved bacterial motility [18], humoral and/or cell-mediated immunity [19,20], surfactant [21,22] and many others [18]. It has been proposed that their natural functions are in reality strictly related to the biology of the producing microorganism [23,18]. Table 1 lists some lipopeptides, their roles and producing microorganisms. Due to their antimicrobial, toxic and surfactant properties, lipopeptides are scientifically and economically valued molecules. A good example is daptomycin, which, even though it is somewhat toxic to human cells, has been approved by the Food and Drug Administration (FDA) for therapeutic application in systemic and life-threatening infections caused by Gram-positive bacteria [24]. Presently, caspofungin, micafungin, and anidulafungin are other examples of lipopeptides used as commercial antibiotics [25]. Needless to say that, as a consequence, these compounds must be produced in large amounts using in vivo and in vitro synthetic methods (biosynthesis and chemical synthesis, respectively).
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
951

Table 1 Some lipopeptides, their biological roles and their producing microorganisms
Lipopeptide Function Producing microorganism (reference)
Surfactin Surfactant activity Bacillus subtilis [21] Fengycins Antifungal acitivity Bacillus subtilis [16] Viscosin Increase of bacterial motility Pseudomonas fluorescens [18] Lipopeptide present in MCF-7 cells
Cytotoxicity, cell-cycle arrest and apoptosis in MCF-7 cell
Bacillus natto [26]
Iturins Antifungal activity Bacilus subtilis [29]
3. Biosynthesis of lipopeptides
Lipopeptides can be produced by the non-ribosomal biosynthetic pathway, an assembly line that parallels protein synthesis but it is ribosomal-independent and elaborated by a complex mechanism involving multifunctional synthetases. Figure 1 shows the structure of two lipopeptides elaborated by the non-ribosomal pathway, an antibiotic of the polymyxin family and a biosurfactant of the surfactin family [27,21]. The uniqueness of these the surfactins is the macrocyclization that is accomplished, in the case of surfactin, produced by Bacillus subtilis, due to the substrate beta-hydroxy fatty acid, which is linked to the N-terminal amino acid by an amide bond (involving the carboxyl moiety of the fatty acid) and to the C-terminal amino acid by an ester bond (involving the beta-hydroxyl side chain of the fatty acid). A database of non-ribosomal peptides and, specifically, non-ribosomally assembled lipopeptides can be consulted at the Norine database (http://bioinfo.lifl.fr/norine/ [28]).
D-Phe L-Leu Dab Dab L-Thr Dab Dab L-Thr Dab fatty acid
Dab
(B)
(A)
L-Glu L-Leu D-Leu L-Val L-Asp D-Leu L-Leu
CO
CH2 CH O
(n = 8,9 or 10)CH2 (CH2)n CH3
Fig. 1 Primary structures of polymyxins, where Dba is 1-diaminobutyric acid, (A) and of surfactins, showing the beta-hydroxylated fatty acid that links the carboxyl and aminoterminals of the heptapeptide (B). In the biosynthesis of surfactins, in order to produce generate their unusual peptide structure, a combination of enzyme-catalyzed steps is carried out by three modular non-ribosomal peptide synthetases (NRPSs), the surfactin synthetases A, B and C [30,31] presented in Figure 2. The enzymatic steps involve aminoacyl-adenylation, thioesterification, condensation, epimerization and thioesterase activities, depending on the specific enzyme module.
Fig. 2 Enzymatic assembly line involved on the biosynthesis of surfactins (adapted from the work of Kraas et al. [31]). The NRPSs are involved in the steps leading to surfactin synthesis. On every enzyme module there are specific domains enrolled on adenylation of each amino acid as denoted for Glu, Leu, Val and Asp; a domain involved in condensation (C); a peptidyl carrier protein domain (PCP); an epimerization domain (Ep) and a thioesterase domain (Te).
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
952
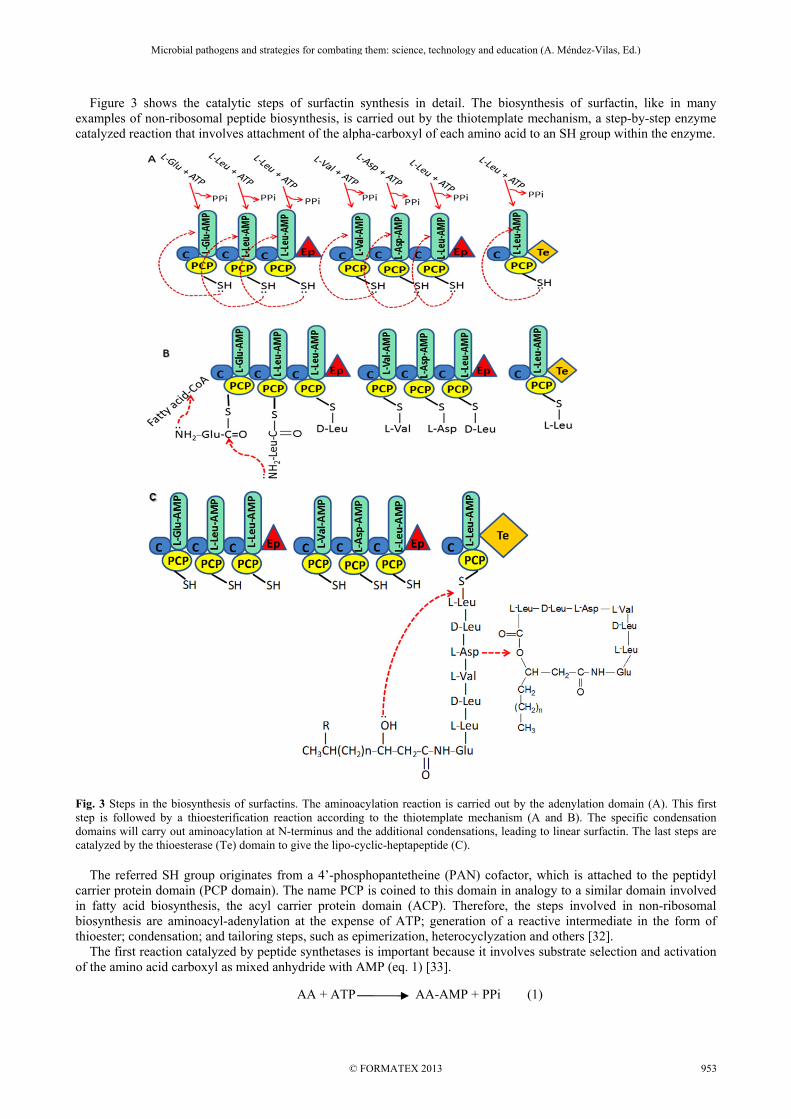
Figure 3 shows the catalytic steps of surfactin synthesis in detail. The biosynthesis of surfactin, like in many examples of non-ribosomal peptide biosynthesis, is carried out by the thiotemplate mechanism, a step-by-step enzyme catalyzed reaction that involves attachment of the alpha-carboxyl of each amino acid to an SH group within the enzyme.
Fig. 3 Steps in the biosynthesis of surfactins. The aminoacylation reaction is carried out by the adenylation domain (A). This first step is followed by a thioesterification reaction according to the thiotemplate mechanism (A and B). The specific condensation domains will carry out aminoacylation at N-terminus and the additional condensations, leading to linear surfactin. The last steps are catalyzed by the thioesterase (Te) domain to give the lipo-cyclic-heptapeptide (C). The referred SH group originates from a 4’-phosphopantetheine (PAN) cofactor, which is attached to the peptidyl carrier protein domain (PCP domain). The name PCP is coined to this domain in analogy to a similar domain involved in fatty acid biosynthesis, the acyl carrier protein domain (ACP). Therefore, the steps involved in non-ribosomal biosynthesis are aminoacyl-adenylation at the expense of ATP; generation of a reactive intermediate in the form of thioester; condensation; and tailoring steps, such as epimerization, heterocyclyzation and others [32]. The first reaction catalyzed by peptide synthetases is important because it involves substrate selection and activation of the amino acid carboxyl as mixed anhydride with AMP (eq. 1) [33].
AA + ATP AA-AMP + PPi (1)
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
953

The active form (Holo-form) of a peptide synthetase has a 4’-phosphopantheteine cofactor attached to the PCP domain. Owing to this functionalization, an SH group allows the formation of a reactive intermediate of the substrate amino acids, activated at the carboxyl end. The following equation (eq. 2) shows the formation of an enzyme-substrate intermediate, as thioester, catalyzed by the PCP domain.
AA-AMP + Enz-SH Enz-S-AA (2)
Before the first peptide bond is formed, as in the example of other biosynthetic clusters involved in the synthesis of lipopeptides, the N-terminal amino acid is acylated by a fatty acid. As stated above, in surfactins, the fatty acid incorporated into the N-terminal amino acid is a beta-hydroxyl-fatty acid, which can vary from 13 to 15 carbons in length [21]. The catalytic domain involved in the aminoacylation reaction is a special type of condensation domain, which is normally found in other synthetases that receive an aliphatic fatty acid or aryl compounds. This is also found at the N-terminus of several subunits of bacterial non-ribosomal peptide synthetases in order to transfer a growing peptide chain from one synthetase to the next collinear subunit. Such enzyme architecture can be seen in the synthetases presented in Figure 3. In our previous work, we showed that condensation domains located at N-terminus of peptide synthetases present higher sequence similarity. The alignment of C-domains also allowed us to group separately the condensation domains catalyzing acylation of aromatic derivatives and those that promote acylation of aliphatic fatty acids [34]. Thus, the formation of peptide bonds and N-terminal acylation reactions are catalyzed by condensation domains, where the catalytic mechanism involves conserved histidines, which will carry out the abstraction of protons from the reactive aminogroup [33]. Equation 3 shows the condensation reaction for two adjacent modules tethered with the respective amino acids as thioesters.
Enz-S-AA1 + Enz-S-AA2 Enz-S-AA2-AA1 (3)
The corresponding acylation of the N-terminal amino acid (glutamic acid) is similarly performed. However, the fatty acid is activated as thioester by a different enzyme that activates the fatty acid as coenzyme A intermediate, demonstrating that the condensation domain in fact acts on a preliminarily activated fatty acid thioester [35]. In surfactin biosynthesis, there are two tailoring conversions of L-leucine into D-Leucine. Both are catalyzed by an epimerase domain and take place at the C-terminal modules of the srfAA and SrfAB synthetases. According to the work of Luo et al. [36], epimerization occurs after amino acid esterification. These authors have studied the conversion of L to D-phenylalanine in the biosynthesis of gramicidin, another cyclic peptide produce by the Bacillus genus, having showed that the ratio of D-isomer in the thioester form over its adenylate counterpart is approximately 3,000 [36]. Therefore, in Figure 3 the aminoacyl-adenylates were presented as L-amino acids rather than in the D-form; conversely, the corresponding thioesters were shown as D-amino acids. The last biosynthetic step is the hydrolysis of the peptide from the last PCP domain, a reaction that is catalyzed by the thioesterase (Te) domain of srfAC (Figure 3). This step is a concerted reaction that also promotes cyclization of the peptide by a transesterification reaction in which the hydroxyl of the fatty acid makes an ester linkage with the carboxyl end of the C-terminal amino acid of surfactin, thus leading to a cyclic peptide as shown in Figure 1 and Figure 3C. There are many other lipopeptides, such as daptomycin, produced by a similar pathway. An interesting feature of the thiotemplate mechanism is the possibility of using the enzymatic machinery for combinatorial biosynthesis, given that substrate selection is not restricted to the primary substrates. The partially relaxed selection of substrates by the adenylation and thioesterification domains has been explored by some authors, allowing the biosynthesis of novel peptides using combinatorial biosynthesis, a strategy that can be used by feeding novel substrates or precursors in the growth medium [37]. Another approach that has been explored is based on the engineering of synthetases by swaping modules from one synthetase into another, a strategy that leads to a novel peptide structure and, therefore, an “unnatural product” that is yet produced as natural product [38]. However, the bottle neck in non-ribosomal production of AMPs, including lipopeptides, is the cost of production considering the low yields at the end of the growth of the producing organism. To overcome this problem, several studies have been carried out using cheaper or recycled substrate specifically for production of surfactins and other lipopeptide families [39]. Besides, chemical synthesis may provide higher yields and, undoubtedly, allow insertion of novel amino acids, cyclizations and modifications in the structures to create analogues with modified biological properties, which can be more appropriate for specific purposes such as specificity and capacity to avoid microbial resistance.
4. Chemical synthesis of lipopeptides
It is well known that chemical peptide synthesis is based on amide bond formation between amino acid or peptide derivatives as shown in the following reaction, in which R-COOH represents an amino acid or a peptide derivative, X represents a good leaving group and H2N-HR1 represents an amino acid or a peptide derivative:
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
954

RO
OHR
O
XNH2 R R
O
NH
R1
1+
Peptide chain assembly can be achieved by the classical or the solid-phase method using one of the two strategies specially developed for such purpose: tert-butyloxycarbonyl (Boc) and fluorenylmethyloxycarbonyl (Fmoc) (Figure 4). As shown, these methods and synthetic strategies share as the common features the needs for (i) orthogonal protection and for α-carboxyl-group activation during peptide chain building as well as (ii) full deprotection when the assembly is complete. Their main differences are the blocker of the α-aminogroup of the amino acids that act as the acyl donors in peptide bond formation (temporary: Boc or Fmoc) and protecting groups of their reactive side-chains (semi-permanent: benzyl or tert-butyl group derivative, respectively) [40,41]. In classical or traditional peptide synthesis, the starting materials (fully or partially protected amino acids), coupling reagents, bases, acids, byproducts and products are fully soluble in the reaction media (Figure 4A; [42,43]). In such anhydrous organic conditions, it is imperative to monitor formation of the products by a colorimetric or chromatographic method as well as to separate them from the reagents, solvent and byproducts using extraction, precipitation and/or liquid chromatography. As the whole synthetic process comprise many steps of α-aminogroup deprotection, α-carboxylgroup deprotection and amino acid coupling, classical peptide synthesis is time-consuming and not always an easy task. Full deprotection gives the crude desired peptide to be purified and chemically characterized prior to use. Solid-phase peptide synthesis (SPPS) has been conceived to make peptide chain assembly fast and simple. In fact, as the amino acids, partially or fully protected, are incorporated on a resin that is insoluble in the reaction media, the soluble reagents, bases, acids and byproducts are removed by simple filtration followed by alternate washings of the aminoacyl- and growing peptidyl-resin with solvents able to swell or shrink them (Figure 4B; [44,45]). As a result, SPPS has become the method of choice to synthesize trouble-free peptides manually [46] or automatically, separately or in parallel [47], conventionally (at room temperature as first proposed by Merrifield; [48]) or at elevated temperature, using conventional heating [49- 51], or with the assistance of microwaves [52-54]. Detachment from the resin and simultaneous full deprotection give the crude peptide, which is usually contaminated with byproducts generated from undesired reactions that also take place during the steps of amino acid deprotection and coupling. Despite the advances and general applicability of both methods, it is of interest to improve them even more because there are still peptides extremely difficult to be obtained by the classical or solid-phase methods. Moreover, many research groups around the world have attempted to chemically synthesize modified proteins [55], not found in natural sources and impractical to produce by the recombinant DNA methodology. For such purposes, peptide derivatives, rather than amino acid derivatives, are used as starting materials.
Fig. 4 Chemical synthesis of a tripeptide containing three different amino acid residues using the classical (A) or solid-phase method (B). Aa1, Aa2 and AA3 represent the amino acids, R represents the resin employed, R1, R2 and R3 are the amino acid side-chains; circles and ovals represent the protecting groups for the reactive amino acid side-chains and the triangle is the Boc or Fmoc group.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
955

The methods of classical chemical synthesis and, mostly, SPPS have been extremely useful to study biologically active peptides [56], including those with antibiotic activity [57]. Indeed, the last two decades have witnessed an overflow of reports on the properties, structure and mode of actions of antimicrobial peptides (AMPs) that would not exist without the possibility of using their synthetics forms (generally, isolation from natural sources provides little amounts of purified bioactive peptides). In addition, structure-activity relationship of natural AMPs has been widely explored simply because the chemical synthesis methodologies have made possible to prepare their truncated, modified and/or cyclic analogues [58-60]. Table 2 illustrates this point by listing some synthetic AMPs and analogues that have been obtained and studied, expanding the current knowledge on such remarkable biomolecules. There are several excellent reviews about lipopeptides focusing on their biosynthesis, properties, biological roles, structures and potential to be employed in the industry [37,61-64,18,65], but not on their chemical synthesis. In 2001 and in commemoration of 100 years of peptide synthesis, Pegoraro and Moroder [66] competently reviewed this subject. With an impressive collection of information, these authors showed that chemical synthesis has been successfully used for peptide assembly during the preparation of N-myristoyl peptides, S-prenylated peptides, S-palmitoylated peptides, doubly lipidated peptides, nonnatural mono- and multiple-lipidated peptides [66]. Besides giving chemical reactions and detailed protocols for the preparation of modified amino acids (building blocks), the authors reminded that modification of synthetic lipopeptides with groups that can be traced by proper analytical techniques is essential to make them suitable for biological studies [67]. Therefore such additional step(s) must be considered when designing the synthetic strategy to be employed. From 1994 to date, dozens of lipopeptides and analogues with variable biological functions have been chemically synthesized, being most of the synthesis performed on resin by the Fmoc strategy with the aim to demonstrate the efficiency of the strategy studied or to obtain the desired lipopeptide to be used in exploratory works. In fact, for instance, Casey and Buss reported optimized synthetic procedures specially developed to furnish simple lipoderivatives [68]. Okeley et al. proposed a procedure to prepare prenylated peptides, which is based on chemoselective transformations of peptide containing Se-phenylselenocystein [69]. DeOgny et al. described the rapid and convenient synthesis of tripalmitoylglyceryl analogs of the N-terminal sequences of several bacterial lipopeptides, which also present immunogenic activity [70]. Pagadoy et al. synthesized the racemic mixture and the R isoform of surfactin, lipopeptide still regarded as the most powerful biosurfactant [71]. Into et al. reported the synthesis of a dipalmitoylated lipopeptide derived from paralogous lipoproteins of Mycoplasma pneumonia with immunostimulatory activity [72]. JølcK et al. [73] developed a novel approach for the synthesis of PEGylated lipopeptides based on SPPS and the use of the Cu(I) catalyzed azide/alkene Huisgen 1,3-dipolar cycloaddition (CuAAC “Click” conjugation) previously reported by Meldal and coworkers [74] and Sharpless and coworkers [75]. Schieck et al. prepared the myristoylated-peptide Myr-HBVpreS/2-78, which is a Hepatitis B virus entry inhibitor, by using solid-phase synthesis at 50 °C [76]. Bionda et al. have just described the suitability of guanilation of resin-bound peptidyl amines for the preparation of fusaricidin/LI-F class of cyclic lipopeptides, which present broad antimicrobial activity [77]. Vilà et al. also have just reported different synthetic strategies for obtaining cyclic lipopeptides with potential to act against plant pathogens [78]. In these examples, peptide assembly was performed in small scale on different resins, such as MBHA (Boc strategy), Fmoc-RINK-MBHA, TentaGel R RAM, Rapp Polymere or NovaSyn TentaGel (Fmoc strategy). The lipid moiety was added to the assembled peptide-resin. The desired lipopeptides were obtained after acylation with the desired lipids with satisfactory yields, indicating that chemical synthesis is indeed suitable for in vitro production of such compounds and their analogues in miligrams. Unfortunately, the need for high quality and well characterized Nα-protected side chain-protected amino acids, functionalized resins, coupling reagents, carbocation scavangers, anhydrous solvents, acids and bases, makes large scale production of lipopeptides by chemical synthesis very expensive. On the other hand, this is the only option when a large amount of one particular chemically modified lipopeptide (for instance, with non-natural functionalities) is desired.
Table 2 A few examples of synthetic analogues of antimicrobial peptides (AMPs) with improved features
Original AMP Analogue (reference)
Bovine α-hemoglobin fragment 33-61 (Hb33-61)
amidated bovine α-hemoglobin fragment 40-61 (Hb40-61a) [58]
Tachyplesin (TP) [Gly6]ccTP* [79] Crotamine Nucleolar targeting peptides (NrTPs) [80]
Pyrrhocoricin (pyrr) Pip-pyrr-MeArg** dimer [81] Mellitin Cyclic mellitin [82]
* cc is cyclic cystine; **MeArg is N-methyl-arginine and Pip is 4-amino-piperidine-4-carboxylic acid
Acknowledgements The support by Fapesp, CNPq, RUSP and PROPESQ (PUC-Campinas) is gratefully acknowledged.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
956

References
[1] Bennett JW, Chung K-T. Alexander Fleming and the discovery of penicillin. Advances in Applied Microbiology. 2001;49: 163-184.
[2] Rutkowski J, Brzezinski B, Structures and properties of naturally occurring polyether antibiotics. BioMed Research International. 2013;2013:162513.
[3] Hancock REW, Chapple DS, Peptide Antibiotics. Antimicrobial Agents and Chemotherapy. 1999;43:1317-1323. [4] Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415: 389-395. [5] Hassan M, Kjos M, Nes IF, Diep, DB, Lotfipour F. Natural antimicrobial peptides from bacteria: characteristics and potential
applications to fight against antibiotic resistance. Journal of Applied Microbiology. 2012;113:723-736. [6] Piotto SP,, Sessa L, Concilio S, Iannelli P, YADAMP: yet another database of antimicrobial peptides. International Journal of
Antimicrobial Agents. 2012;39:346-351. [7] Papagianni M. Ribosomally synthesized peptides with antimicrobial properties: biosynthesis, structure, function, and
applications. Biotechnology Advances. 2003;6:465-499. [8] Hull R, Katete R, Ntwasa M. Therapeutic potential of antimicrobial peptides from insects. Biotechnology and Molecular
Biology Review. 2012;7:31-47. [9] Micek ST. Alternatives to Vancomycin for the Treatment of Methicillin-Resistant Staphylococcus aureus Infections. Clinical
Infectious Disease. 2007;45:S184-S190. [10] Marques MA, Citron DM, Wang CC. Development of Tyrocidine A Analogues with improved antibacterial activity.
Bioorganic & Medicinal Chemistry. 2007;15:6667-6677. [11] Marr AK, Gooderham WJ, Hancock REW. Antibacterial peptides for therapeutic use: obstacles and realistic outlook. Current
Opinion in Pharmacology. 2006;6:468–472. [12] Mogi T, Kita K. Gramicidin S and polymyxins: the revival of cationic cyclic peptide antibiotics. Cellular and Molecular Life
Sciences. 2009;23:3821-3826. [13] Ongena M, Jourdan E, Adam A, Paquot M, Brans A, Joris B, Arpigny J-L, Thonart P. Surfactin and fengycin lipopeptides of
Bacillus subtilis as elicitors of induced systemic resistance in plants. Environmental Microbiology. 2007;9:1084–1090. [14] Kirana GS, Thomas TA, Selvina J, Sabarathnama B, Liptonb AP. Optimization and characterization of a new lipopeptide
biosurfactant produced by marine Brevibacterium aureum MSA13 in solid state culture. Bioresearch Technology. 2010;101: 2389-2396.
[15] Kanasaki R, Sakamoto K, Hashimoto M, Takase S, Tsurumi Y, Fujie A, Hino M, Hashimoto S, Hori Y. FR209602 and related compounds, novel antifungal lipopeptides from Coleophoma crateriformis No. 738 I. taxonomy, fermentation, isolation and physico-chemical properties. Journal of Antibiotics. 2006;59:137–144.
[16] Chen L, Wang N, Wang X, Hu J, Wang S. Characterization of two anti-fungal lipopeptides produced by Bacillus amyloliquefaciens SH-B10. Bioresearch Technology. 2010;101:8822-8827.
[17] Etchegaray A, Bueno CC, de Melo IS, Tsai SM, Fiore MF, Silva-Stenico ME, de Moraes LA, Teschke O. Effect of a highly concentrated lipopeptide extract of Bacillus subtilis on fungal and bacterial cells. Archives of Microbiology. 2008;190:611-622.
[18] Raaijmakers JM, de Bruijn I, Nybroe O, Ongena M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiology Review. 2010;34:1037–1062.
[19] Zeng W, Horrocks KJ, Robevska G, Wong CY, Azzopardi K, Tauschek M, Robins-Browne RM, Jackson DC. A modular approach to assembly of totally synthetic self-adjuvanting lipopeptide-based vaccines allows conformational epitope building. Journal of Biological Chemistry. 2011;286:12944–12951.
[20] Zeng W, Ghosh S, Lau YF, Brown LE, Jackson DC. Highly immunogenic and totally synthetic lipopeptides as self-adjuvanting immunocontraceptive vaccines. Journal of Immunology. 2002;169:4905-4912.
[21] Jacques P. Surfactin and other lipopeptides from Bacillus spp. In: Soberon-Chavez G, ed. Biosurfactants: from genes to applications: Microbiology Monographs 20. Berlin-Heidelberg, Germany: Springer-Verlag; 2011:57-91.
[22] Nazari M, Kurdi M, Heerklotz H. Classifying surfactants with respect to their effect on lipid membrane order. Biophysical Journal. 2012;102:498-506.
[23] Koch B, Nielsen TH, Sørensen D, Andersen JB, Christophersen C, Molin S,Givskov M, Sørensen J, Nybroe O. Lipopeptide Production in Pseudomonas sp. Strain DSS73 Is Regulated by Components of Sugar Beet Seed Exudate via the Gac Two-Component Regulatory System. Applied and Environmental Microbiology. 2002;68:4509–4516.
[24] Fowler VG, Boucher HW, Corey GR. Daptomycin versus standard therapy for bacteremia and endocarditis caused by Staphylococcus aureus. New England Journal of Medicine. 2006;355:653–65.
[25] Arendrup MC, Garcia-Effron G, Lass-Flör C, Lopez AG, Rodriguez-Tudela J-L, Cuenca-Estrella M, Perlin DS. Echinocandin susceptibility testing of candida species: comparison of EUCAST EDef 7.1, CLSI M27-A3, e-test, disk diffusion, and agar dilution methods with RPMI and iso sensitest media. Antimicrobial Agents and Chemotherapy. 2010;54:426–439.
[26] Xiao-Hong Cao X-H, Liao Z-Y, Wang C-L, Yang W-Y, Lu M-F. Evaluation of a lipopeptide biosurfactant from Bacillus Natto TK-1 as a potential source of anti-adhesive, antimicrobial and antitumor activities. Brazilian Journal of Microbiology. 2009;40:373-379.
[27] Chen LF, Kaye D. Current use for old antibacterial agents: polymyxins, rifamycins, and aminoglycosides. Medical Clinics of North America. 2011;95:819-842.
[28] Caboche S, Leclère V, Pupin M, Kucherov G, Jacques P. Diversity of monomers in nonribosomal peptides: towards the prediction of origin and biological activity. Journal of Bacteriology. 2010;192: 5143–5150.
[29] Maget-Dana R, Peypoux F. Iturins, a special class of pore-forming lipopeptides: biological and physicochemical properties. Toxicology. 1994;87:151-174.
[30] Menkhaus M, Ullrich C, Kluge B, Vater J, Vollenbroich D, Kamp RM. Structural and functional organization of the surfactin synthetase multienzyme system. Journal of Biological Chemistry. 1993;268:7678-7684.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
957

[31] Kraas FI, Helmetag V, Wittmann M, Strieker M, Marahiel MA. Functional dissection of surfactin synthetase initiation module reveals insights into the mechanism of lipoinitiation. Chemistry and Biology. 2010;17:872-880.
[32] Walsh CT, Chen H, Keating TA, Hubbard BK, Losey HC, Luo L, Marshall CG, Miller DA, Patel HM. Tailoring enzymes that modify nonribosomal peptides during and after chain elongation on NRPS assembly lines. Current Opinion in Chemical Biology. 2001;5:525-534.
[33] Marahiel MA, Essen LO. Nonribosomal peptide synthetases mechanistic and structural aspects of essential domains. Methods in Enzymology. 2009;458:337-351.
[34] Etchegaray A, Silva-Stenico ME, Moon DH, Tsai SM. In silico analysis of nonribosomal peptide synthetases of Xanthomonas axonopodis pv. citri: identification of putative siderophore and lipopeptide biosynthetic genes. Microbiological Research. 2004;159:425-437.
[35] Steller S, Sokoll A, Wilde C, Bernhard F, Franke P, Vater J. Initiation of surfactin biosynthesis and the role of the SrfD-thioesterase protein. Biochemistry. 2004;43:11331-11343.
[36] Luo L, Kohli RM, Onishi M, Linne U, Marahiel MA, Walsh CT. Timing of epimerization and condensation reactions in nonribosomal peptide assembly lines: kinetic analysis of phenylalanine activating elongation modules of tyrocidine synthetase B. Biochemistry. 2002;4:9184-9196.
[37] Bonmatin JM, Laprévote O, Peypoux F. Diversity among microbial cyclic lipopeptides: iturins and surfactins. Activity-structure relationships to design new bioactive agents. Combinatorial Chemistry High Throughput Screening. 2003; 6:541-556.
[38] Walsh CT. Polyketide and nonribosomal peptide antibiotics: Modularity and versatility. Science. 2004;303:1805-1810. [39] Al-Ajlani MM, Sheikh MA, Ahmad Z, Hasnain S. Production of surfactin from Bacillus subtilis MZ-7 grown on pharmamedia
commercial medium. Microbial Cell Factories. 2007;6:17-24. [40] Stewart JM, Young JD. Solid Phase Peptide Synthesis. Pierce Chemical Company, Rockford, USA. 1984:1-49. [41] Atherton E, Sheppard RC. Solid Phase Peptide Synthesis: a practical approach, Oxford: IRL press. 1989. [42] Bodansky M. Racemization. In: Principles of Peptide Synthesis, vol. 16, Bodanszky M. ed., Springer-Verlag, Berlin, 1984:158-
173. [43] Machado A, Liria CW, Proti PB, Remuzgo C, Miranda MTM. Sínteses químicas e enzimáticas de peptídeos: princípios básicos
e aplicações. Quimica Nova. 2004;27:781-789. [44] Fields GB, Noble RL. Solid phase peptide synthesis utilizing 9-fluorenylmethoxycarbonyl amino acids. International Journal of
Peptide and Protein Research. 1990;35:161-214. [45] Miranda MTM, Liria, CW, Remuzgo C. Difficult peptides, In: Hughes AB. ed. Amino Acids, Peptides and Proteins in Organic
Chemistry. Wiley-VCH Publishers, Weinheim, Germany. 2011:549-569. [46] Merrifield RB. Automated synthesis of peptides. Science. 1965;150:178-185. [47] Winkler DFH. Chemistry of SPOT synthesis for the preparation of peptide macroarrays on cellulose membranes. Mini-Review
in Organic Chemistry. 2011;8:114-120. [48] Merrifield RB. Solid phase peptide synthesis I synthesis of a tetrapeptide. Journal of the American Chemical Society. 1963; 85:
2149-2154. [49] Rivier JE, Miranda, MTM. Solid-Phase peptide synthesis at elevated temperature. In: Goodman M, Felix A, Moroder L,
Toniolo C eds. Synthesis of peptides and peptidomimetics: commemorating 100 years of peptide research. 2001/2004:806-813. [50] Souza MP, Tavares MFM, Miranda MTM. Racemization in stepwise solid-phase peptide synthesis at elevated temperatures.
Tetrahedron. 2004; 60:4671-4681. [51] Remuzgo C, Andrade GFS, Temperini MLA, Miranda MTM. Acanthoscurrin fragment 101-132: Total synthesis at 60 °C of a
novel difficult sequence. Journal of Peptide Science. 2009; 92:65-75. [52] Bacsa B, Desai B, Dibó G, Oliver-Kappe C. Rapid solid-phase peptide synthesis using thermal and controlled microwave
irradiation. Journal of Peptide Science., 2006;12:633-638. [53] Rizzolo F, Sabatino G, Chelli M, Rovero P, Papini AM. A convenient microwave-enhanced solid-phase synthesis of difficult
peptide sequences: case study of gramicidin A and CSF114(glc). International Journal of Peptide Research Therapeutics. 2007;13:203-208.
[54] Loffredo C, Assunção NA, Gerhardt J, Miranda, MTM. Microwave-assisted solid-phase peptide synthesis at 60ºc: alternative
conditions with low enantiomerization. Journal of Peptide Science. 2009;15:808-815. [55] Kent S, Sohma Y, Liu S, Bang D, Pentelute B, Mandal K. Through the looking glass–a new world of proteins enabled by
chemical synthesis. Journal of Peptide Science. 2012;18: 428–436 [56] Bryan A, Joseph L, Bennett JA, Jacobson HI, Andersen TT. Design and synthesis of biologically active peptides: a 'tail' of
amino acids can modulate activity of synthetic cyclic peptides. Peptides. 2011;32:2504-2510. [57] Jenssen H, Hamill P, Hancock RE. Peptide antimicrobial agents. Clinical Microbiology Review. 2006;19:491-511. [58] Machado A, Sforça M L, Miranda A, Daffre S, Spisni A, Pertinhez TA, Miranda MTM. Truncation of amidated fragment 33-
61 of bovine α-hemoglobin: effect on the structure and anticandidal activity. Biopolymers (Peptide Science). 2007;88:413-426. [59] Kim J, Park S, Yoon M, Hahm K, Park Y. C-terminal amidation of PMAP-23: translocation to the inner membrane of Gram-
negative bacteria. Amino Acids. 2011;40:183-195. [60] Machado A, Fázio MA, Miranda A, Daffres S, Machini MT. Synthesis and properties of cyclic gomesin and analogues. Journal
of Peptide Science. 2012;18:588-598. [61] Kosmidis C, Levine DP. Daptomycin: pharmacology and clinical use. Expert Opinion on Pharmacotherapy. 2010;11:615-625. [62] Moyle PM, Toth I. Self-adjuvanting lipopeptide vaccines. Current Meicinal Chemistry. 2008;15:506-516. [63] Ongena M, Jacques P. Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends in Microbiology.
2008;16:115-125. [64] Pirri G, Giuliani A, Nicoletto SF, Pizzuto L, Rinaldi AC. Lipopeptides as anti-infectives: a practical perspective. Central
European Journal of Biology. 2009:4:258–273. [65] Mandal SM, Barbosa AE, Franco OL. Lipopeptides in microbial infection control: scope and reality for industry. Biotechnology
Advances. 2013;31:338-345.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
958

[66] Pegoraro S, Moroder L. Synthesis of lipopeptides. In: Goodman M, Felix A, Moroder L, Toniolo C eds. Synthesis of Peptides and Peptidomimetics: Commemorating 100 years of Peptide Research. 2001/2004:333-374.
[67] Hoogerhout P, Stittelaar KJ, Brugghe HF, Timmermans JAM, ten Hove GJ, Jiskoot W, Hoekman JHG, Roholl, PJM. Solid-phase synthesis and application of double-fluorescent-labeled lipopeptides, containing a CTL-epitope from the measles fusion protein. Journal of Peptide Research. 1999;54:436-443.
[68] Lipid modification in Proteins. Methods in Enzymology. Casey PJ, Buss JE. (eds.) 1995;250. [69] Okeley NM, Zhu Y, van Der Donk WA. Facile chemoselective synthesis of dehydroalanine-containing peptides. Journal of
Organic Letters. 2000;2:3603-3606. [70] De Ogny L, Pramanik BC, Arndt LL, Jones JD, Rush J, Slaughter CA, Radolf JD, Norgard MV. Solid-phase synthesis of
biologically active lipopeptides as analogs for spirochetal lipoproteins. Peptide Research. 1994;7:91-97. [71] Pagadoy M, Peypoux F, Wallach J. Solid-Phase Synthesis of Surfactin, a Powerful biosurfactant produced by Bacillus subtilis,
and of four analogues. International Journal of Peptide Research and Therapeutics. 2005:11:195-202. [72] Into T, Dohkan J-I, Inomata M, Nakashima M, Shibata K-I, Matsushita K. Synthesis and characterization of a dipalmitoylated
lipopeptide derived from paralogous lipoproteins of Mycoplasma pneumonia. Infection and Immunity. 2007;75:2253-2259. [73] Jølck RI, Berg RH, Andresen TL. Solid-phase synthesis of PEGylated lipopeptides using click chemistry. Bioconjugate
Chemistry. 2010;21:807-810. [74] Tornøe CW, Christensen C, Meldal M. Peptidotriazoles on solid Phase: [1,2,3]-triazoles by regiospecific copper(I)-catalyzed
1,3-dipolar cycloadditions of terminal alkynes to azides. Journal of Organic Chemistry. 2002;67:3057-3064. [75] Rostovtsev VV, Green LG, Fokin VV, Sharpless KB. Angewandte Chemie International Edition. 2002;41:2596-2599. [76] Schieck A, Müller T, Schulze A, Haberkorn U, Urban S, Mier W. Solid-phase synthesis of the lipopeptide Myr-HBVpreS/2-78,
a hepatitis B virus entry inhibitor. Molecules. 2010;15:4773-4783. [77] Bionda N, Pitteloud JP, Pitteloud JP, Cudic P. Solid-phase synthesis of fusaricidin/li-f class of cyclic lipopeptides:
Guanidinylation of resin-bound peptidyl amines. Biopolymers. 2013;100:160-166. [78] Vilà S, Badosa E, Montesinos E, Feliu L, Planas M. A convenient solid-phase strategy for the synthesis of antimicrobial
cyclic lipopeptides. Organic and Biomolecular Chemistry. 2013;11:3365-3374. [79] Tam JP, Lu YA, Yang JL. Marked increase in membranolytic selectivity of novel cyclic tachyplesins constrained with an
antiparallel two-beta strand cystine knot framework. Biochem Biophys Res Commun. 2000; 267:783-790. [80] Rádis-Baptista G, de la Torre BG, Andreu D. A novel cell-penetrating peptide sequence derived by structural minimization of a
snake toxin exhibits preferential nucleolar localization. Journal of Medicinal Chemistry. 2008; 51:7041-7044. [81] Otvos, L Jr, Wade JD, Feng JQ, Condie BA, Hanrieder J, Hoffmann R. Designer antibacterial peptides kill fluoroquinolone-
resistant clinical isolates. J. Med. Chem. 2005; 48:5349-5359. [82] Unger T, Oren Z, Shai Y. The effect of cyclization of magainin 2 and melittin analogues on structure, function, and model
membrane interactions: implication to their mode of action. Biochemistry. 2001; 40:6388-97.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
959





























