Embed Size (px)
Citation preview

and the purified inducers should reveal whetherGABA and these algal inducers bind to the samelarval receptors . Elucidation of the structure ofthese inducers can lead to experiments in mammali-an systems to determine whether these algal mole-cules also mimic GABA in mammalian tissues, andwhether similar GABA-mimetic substances occurnaturally in mammals. Such investigations couldprove useful in unraveling the processes controllingdevelopment and neuronal function . Lastly, suchstudies may provide insight into the origins andeffective treatment of diseases associated withGABA functions .
Acknowledgements
This research was supported by the US NavyOffice of Naval Research Contract #N00014-80-C-0310, and by the US Dept . of Commerce (NOAA)-University of California Sea Grant College Pro-gram Grant #R/ A-43. The assistance of N . Hookerin providing Haliotis larvae, C . Froyd for excellenttechnical assistance and the Marine Science Insti-tute of the University of California, Santa Barbara,is gratefully acknowledged .
References
Bradford, M . A ., 1976 . A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding . Analyt . Biochem . 72:248-254 .
Morse, A . N . C . & D . E . Morse, 1984 . Recruitment and meta-morphosis of Haliotis Linnaeus larvae are induced by mole-cules uniquely available at the surfaces of crustose red algae .J . exp . mar . Biol . Ecol . 75 : 191-215 .
Morse, D . E ., H . Duncan, N. Hooker& A . Morse, 1977 . Hydro-gen peroxide induces spawning in mollusks, with activationof protaglandin endoperoxide synthetase . Science 196 : 298-300 .
Morse, D . E ., N . Hooker & A. Morse, 1978 . Chemical control ofreproduction in bivalve and gastropod molluscs, 3 : An ínex-pensive technique for mariculture of many species . Proc.World Maricult . Soc. 9: 543-547 .
Morse, D . E ., N . Hooker, H . Duncan & L . Jensen, 1979a.y-Amínobutyric acid, a neurotransmitter, induces planktonicabalone larvae to settle and begin metamorphosis. Science204:407-410 .
Morse, D . E ., N . Hooker, L . Jensen & H . Duncan, 1979b.Induction of larval abalone settling and metamorphosis byy-aminobutyríc acid and its congeners from crustose redalgae : 2 . Applications to cultivation, seed production andbioassays; principle causes of mortality and interference .Proc. World Maricult. Soc . 10 : 81-91 .
Morse, D . E., N . Hooker & H . Duncan, 1980 . GABA inducesmetamorphosis in Halloos, 5 : Stereochemical specificity.Brain Res . Bull . 5 : 381-387 .
Troxler, R . F . & R . Lester, 1967 . Biosynthesis of phycocyanobil-in. Biochemistry 6 : 3840-3846 .
Antimicrobial activity from marine algae : Results of a large-scale screening programme
John L. Reicheltl & Michael A. Borowitzka 2,3
I Sir George Fisher Centre of Tropical Marine Studies, James Cook University, Townsville, Qld . 4811,
Australia2 School of Environmental and Life Sciences, Murdoch University, Murdoch, WA 6150, Australia3 To whom correspondence should be addressed
Keywords: seaweed, algae, antibiotics, biologically active compounds
Introduction
Between 1974 and 1981, scientists at the Roche Re-search Institute of Marine Pharmacology(RRIMP)in Dee Why (Sydney, Australia) carried out anintensive study of biologically active chemical sub-stances from marine flora and fauna with the aim offinding new compounds which were suitable fortherapeutic medical and veterinary applications .
© 1984, Hydrobiologia 116/117 .
One aspect of this work was the screening of extractsfrom marine plants for potential antibiotic com-pounds .
In the literature there are many reports of antibio-tics isolated from marine algae (e.g., Hornsey &Hide 1979 ; Burkholder, et al 1960; Ohta 1977; Feni-cal, et al 1977 ; Rao & Parekh 1981) . Although thesepapers and many others clearly show that marinealgae produce a wide range of compounds with

antibiotic action in vitro, these compounds weregenerally tested against only a limited number ofmicroorganisms, and were rarely tested in in vivosystems .
In this paper we summarize the results of theRRIMP screening programme and discuss the im-plications of the results .
Materials and methods
Algae were collected either by diving, in the inter-tidal zone, or by collecting in the drift around Aus-tralia and New Zealand . Approximately 20 kg wetweight of each species were collected and frozen assoonn as possible after collection . Reference speci-mens of each collection were kept and are nowlodged in the National Herbarium, Botanical Gar-dens, Sydney . The frozen material was returned tothe laboratories at Dee Why where it was ground to apowder under liquid nitrogen for extractions.
A number of extraction methods were used eithersingly or sequentially . These included extraction incold water, dilute ammonia, cold methanol, colddichloromethane and cold petroleum ether . These`crude' extracts were tested for antimicrobial activityin vitro, and if such activity was found then this wasfollowed up by in vivo testing. Promising in vivoactivities were followed by fractionation of the`crude' extract and further in vitro and in vivo test-ing. Any pure compounds isolated during this frac-tionation or during fractionation to isolate pharmo-cologically active compounds were also tested in theantimicrobial screen .
Antimicrobial activity was tested against theGram-positive bacteria Staphylococcus aureus (Sa),Streptococcus pyogenes (Spy) and Streptococcuspneumoniae (Spn), the Gram-negative bacteriaEscherichia colí (Ec), Pseudomonas aeruginosa(Pa), Proteus mirabilis(Pm) and Salmonellaschott-muelleri ( Ss), the yeasts Candida albicans (Ca) andCryptococcus neoformans ( Cn), the fungi Tricho-phyton mentagrophytes (Tm) and Microsporumaudouini (Ma), and the protozoan Trichomonasfoetus (Tf) .
The bacteria were grown in a medium containingglucose(4 g l -'),NH4C1(2 g l- ~),NH4NO3 (0.3 g I- '),Na2SO4 (0.6g1 I), K2HPO4 (1 g 1 1 ), KH2PO4(0 .3 g l'), MgSO4 .7H2O (0.03 g 1 - '), ferric ammo-
1 59
nium citrate (0 .03 g 1-1 ) and Oxoid trypticase soybroth (6 g 1 - ) . The yeasts and fungi were grown in amedium containing glucose (20 g 1 -~), inositol(0.01 g l) and Difco Bacto-yeast nitrogen base(6.7 g 1 - ~) . Serial twofold dilutions of the test com-pounds were made using the appropriate growthmedium, which was then inoculated with exponen-tially growing cells of the test organism . In the case ofthe fungi, the inoculum consisted of a spore suspen-sion and the medium was then solidified by theaddition of an equal volume of4%(w:v) Difco-agar .The yeasts were incubated at 28 ° C ; the other organ-isms were incubated at 37 ° C . After 24 h for thebacteria and 48 h for the yeasts, the cultures wereexamined for turbidity to determine the minimuminhibitory concentration(MIC) . To avoid the possi-bility of misinterpretation due to the turbidity ofinsoluble compounds, these cultures were plated outon the homologous medium to determine the min-imum bacterial concentration (MBC) . In general theMIC and the MBC were very similar . In the case ofthe fungi, the cultures were observed microscopical-ly for colony formation after ten days to determinethe MIC .
In vivo evaluations were performed with 18-20-gFüllinsdorf mice, using ten mice per dose . Acutetoxicities were determined to allow the compoundsto be tested at their maximum tolerated doses . Toassess systemic activity, mice were given overwhelm-ing infections (100to 1 000 minimum lethal doses) andthen treated with the maximum tolerated doses ofthe compounds administered either orally or subcu-taneously (Grunberg, et al 1967) . Topical antimi-crobial activity was assessed by establishing in thesubcutaneous tissues of mice a localized infectionand infusing the test compound into the area ofinfection (Grunberg & Schnitzer 1959 ; Grunberg, etal 1967; Bach, et al 1978) .
Trichomonas foetus was maintained by passagebi-weekly in mice . For testing, 0 .2 ml of Trichomo-nas cells at a concentration of5-7 X I0 cells ml - ~ wasinjected on the ventral surface of the mice . Thecompound under test or a standard drug (Ornida-zole) was then injected into the loose skin behind thehead on the dorsal side of the neck . The mice werekilled after 24 h and dissected . The opalescent mate-rial below the skin at the site of infection was thenremoved and examined microscopically for motileTrichomonas .

1 6 0
Results
The RRIMP approach differed significantly fromthe approach followed by most other workers . In-itial screening was generally carried out on `crude'extracts (extracts made from frozen algae using arange of aqueous and nonaqueous solvents) ratherthan on pure compounds . If promising in vitroactivity was detected in these `crude' extracts, theywere then tested in the in vivo screens, and only ifactive in the in vivo screen was fractionation of the`crude' extract undertaken to isolate and purify theactive constituent . The fractionation of the `crude'extract was monitored continually using the biolog-ical activity . In most cases at least three different
Table 1 . Summary of all species tested and of their activities . (-) = inactive; (+) = active in vitro; (A) = active in vivo; ( . ) = not tested . Thenumber before the species name refers to the collection number . * = laboratory culture .
ChlorophytaCaulerpales
1496 Caulerpa brownii1441
Caulerpa fıliformis1686
Caulerpa flexilis3516 Caulerpa longifolia1494/1698172235013522I6873520191235451519
Sa
Spy Spn Ec
Caulerpa papi11osa - +Caulerpa racemosa +Caulerpa scalpe11íformisCaulerpa simp1iciuscu1a - +Caulerpa lrifaria + +Caulerpa vesiculifera - +Chlorodesmis fastigiata - +Halimeda cylindracea - +Halimeda opuntia
CladophoralesI553 Microdictyon umbílícatum3514 Rhizoclonium riparium
Codia1es1808 Codium adhaerens
+
+1 540 Codium fragile1809 Codium muelleri
+
+1003 Codium spongiosum
+
+Siphonocladales
1002 Cladophoropsis vaucheriaeformis
+Utvales
1534
Enteromorpha intestinalis1537
Ulva rígida
-
+Volvocales
2022
Dunalie11a tertiolecta*
solvent systems were used to prepare `crude' extractsof the alga for screening . The results obtained aresummarized in Table 1 . This Table shows that someactivity was found in most species examined . Of the159 species of marine plants screened, 118 (74 .2%)showed in vitro activity against one or more of themicrobes used in the screen . A total of 584 extractswas screened and 211 (36 .1%) of these showed invitro activity. This high rate of activity is comparableto the results obtained in other studies . However, ofthe 185 of these `crude' extracts with in vitro activitywhich were subsequently tested in the in vivoscreens, only 9 (4 .9%) showed in vivo activity .
The highest proportion of activities found wereagainst the Gram-positive bacteria and the least
Pm Ss Pa Ca Cn Tm Ma Tf

Table I . Continued .
1 6 1
Sa Spy Spn Ee Pm Ss
Pa Ca Co Tm Ma Tf
PhaeophytaEctocarpales
Notheiaceae1510 Notheia anomala + +
+Chordariaceae1806
Scytothamnus australis + +
-Chnoosporaceae1848 Chnoospora implexa + -
-
+Sphac Iariales13%
Cladostephus verticillatus -
-1876
Halopteris sp . +1978
Halopteris pseudospicata + + +Dictyotales
I361
Dictyota dichotoma + - +
-
-
+
+1892
Dictyota furce/lata + + +
+
+
+
+
+3509
Dictyota prolifera + + A
+I359
DicJ,vota prolificans + -
-
+I 100
Dictyopteris sp . + + +
-
-I041/164`
Dictyopteris aerostichoides + + +
-
-353
Dictyopteris muelleri - + +
-1360/3510
Dilophus marginatus A + +
-
-
+
+1362
Gymnosorus nigrescens - -
-1977
Lobospira bicuspidata + +
-3536 Spaıoglossum sp . - + +I51 :
Zonaria spiralis + + +
-
+Laminariales
I387
Ecklonia radiata - +
-I497
Macrocystis pyrífera + +
-Fucales
Cysroseiraceae1447 Acrocarpia paniculala + +
-
-
-1832
Acrocarpia paniculala + + +
+
+
+
-
-
-151 1
Cau1ocysıis cephalorniıhos + + +
-
+1821
Cystophora congesta A + +
+
+
+
-
+ -1825
CyJtophora expansa A A +
+
+
+
-
+16* Ćvstophora monilífera + + +
-
+1893
Cystophora monilífera A A +
-
+
+1853
Cystophora platylobium + + +
-
+
+
-
-1491
Ćvstophora polycysiidea + + +
-1827
Cystophora po1ycysiidea + A +
+
+
+
+1826
Cystophora retorta + + +
+
+
+
-
+1872
Cystophora scalaris A + +
+
+
+
-
+
+1513
Cýstophora ıorulosa + A +
+
-
+ +1997 Cystophyllum onustum + +
-
-
-1889
Scaberia agardhii - + +
-Durvilleaceae
- - +
-1878 Durvillea anıarcJicaI509 Durvillea potatorum + -
+1879 Durvillea willana - + -Fucaceae
- + +1805 Xiphophora chondrophylla1829 Xiphophora chondrophylla + + +
+
+
+
+

162
Table I . Continued .
Sa Spy Spn Ec Pm Ss
Pa Ca Cn Tm Ma Tf
Hormosiraceae1538 Hormosira banksii
+
+Sargassaceae1444 Sargassum sp .
-
-
+
-1445 Sargassum sp .1543 Sargassum sp .
-
+1828 Sargassum sp .
-
+
+ +1847 Sargassum sp .
-
+
+ +1975 Sargassum sp .
-
+
+
-1999 Sargassum sp .
-
+
+3565 Sargassum sp . - + - -1446 Sargassum globulariaefolium + - - -3575 Sargassum tristichum -Seirococcaceae1508 Phyllospora comosa
+
-1541 Phyllospora comosa
+
-
-
+
+
+
+
+1971 Scytothalia dorycarpa
+
+
-
-
+
-
-
-I831
Seirococcus axillaris
+
+
+
-
-
+
-
-
-Desmarestiales
1688 Perithalia caudata
+
+
+
-
-
-
-
+
+
+
+
RhodophytaFlorideophycideaeNemalionales
Bonnemaisoniaceae1671 Delisea elegans
+
+
+
+
-
-
-
+
+
+1368 Delisea fımbriata + + + + + - + + +1639 Ptilonia australasica + - - + + + + +Chaetangiaceae1410 Galaxauraarborea
-
+1520 Galaxaura rugosa
-
+
+1759 Galaxaura subfruticulosaGelidiaceae1456
Ce1idium sp.1810
Gelidium sp.1460 Gelidium glandulaefolium
CoraIlinales3561
Haliptilon sp .Cryptonemiales
1820 Pachymenia himantophora1868 Pachymenia lusoria
GigartinalesCaulacanthaceae1468
Rhabdonia verticillata
-
-
-Gigartinaceae1818
Gigartina aIveasa
+
-
+1869
Gigartina circumcincta
-
-1867 Gigartina cranwellae
+
+Gracilariaceae1970 Curdiea obesa
-
-
+
-
-
-
-1544 Gracilaria sp.
+
+
+ +
+189I
Gracilaria Iichenoides
-
+
+
-
-
-
-3556
Gracilaria sp.

Table 1 . Continued .
BangiophycideaePorphyridiales
I546
Porphyry denticulata
CyanophytaChroococcales
200!
Svnechococcussp .*Oscillatoriales35481670202635463542I907
Hormothamnion enteromorphoidesLyngbys sp .Lyngbys lutea*Oscillatoria bonnemaisoniiPhormidium hendersoniiRivularia firma
1 6 3
Sa
Spy Spn Ec Pm Ss
Pa Ca Cn Tm Ma Tf
Hypneaceae1461
Hvpnea sp .I979 Ĥvpnea episcopaIis1995
Hypnea valenıiaePhacelocarpaceae1834 Phacelocarpus complanatusI45I
Phacelocarpus Iabillardieri
+
+ +
+
+
+
+ +Plotamiaceae1506 Plocamium angusıum +
+
+I539 Plocamium cartilagineum1454 Plocamium cost alum +
+I492
Plocamium merlensiiRhizophyllidaceae1787
Chondrococcus sp .SoI eriaceae3569
Callophvcus harveyanus1457
Areschougia sp . +
+1493
Areschougia sp . +
+
+Ceramiales
Ceramiaceae1462
BaI1ia robertiana35I5
Centroceras clavulatum3525
Gri¡ftıhsia sp .
+ +Dayaceae3526 Dasya sp .
+ + +I895
Thuretia quercifolia
+ +RhodomelaceaeI607 Chondria dasyphylla1982
Dict'menia sonden
+ A352I
Laurencio sp.
+
+ +1381
Laurencia botrvoides
+ +
+
+
+1505
Launencia elataI981)
Laurencia fıliformis
+ +3538 Neurymenla fraxinѓfolia1415
Polysiphonia sp .3524 Polysiphonia sp .
+ +I819
Vida/ja co/ensi1983
Vidalfa spiralis
+ +

1 64
Table 1 . Continued.
ChrysophytaBacillariophyceae2014 Navicula sp .*
+
+
PyrrophytaGymnodiniales
2018 Amphidinium carterae*
+
Aquatic angiospermsHelobiae
1890 Amphiboles antarctica - +1953 Halophila ovalis - +1951
Posidonia australis (leaves)
-
+Dicotyledonae
1998
Avicennia marina (leaves)1923 Avicennia marina (pneumatoph .)
+
+
against the Gram-negative bacteria and yeasts (Ta-ble 2) . In vivo activities were observed only againstthe Gram-positive bacteria . Table 3 summarizes theresults for the four major algal phyla and shows thatthe highest proportion of in vitro activities wasobserved in the brown algae. In vivo activities werealmost totally restricted to the brown algae, with thegenus Cystophora accounting for most of the in vivoactivities .
Because in vitro activities were of little interest perse, only limited data are available as to the com-pounds which had the in vitro antimicrobial activity .However, some data on pure compounds are availa-ble, because fractionation of `crude' extracts wascarried out not only to establish the active com-pounds in the antimicrobial screens, but also to
Total numberof extracts
435
352
353
438
438
17Total in vitro
86
124
138
18
14
0active
(19.8%)** (35 .2%) (39.1%) (4.1%) (32%)Total numbertested in vivo 44 43 76 10 8 0Total in vivo 5 4 2 0 0 0active
(11 .4%)**
(93%) (2.6%)
* See Materials and methods for abbreviations used .** Figures in parentheses are % active of total tested .
Sa Spy Spn Ec Pm Ss
Pa Ca Cn Tm Ma Tf
Table 2 . Summary of results obtained with the various microbes used in the screens .
(4 .3%) (0 .7%)
purify compounds which showed activity in thepharmacological screens .
Compounds active in vitro isolated from red algaegenerally were halogenated and include the lipid-soluble pentabromopyrone (Fig. la) from Ptiloniaaustralasica, a tribromo compound isolated fromDelisea hypnoides (Fig . lb), and the fimbrolidesisolated from Delisea fìmbriata (Fig . lc, d, e) .
Much of the in vitro activity observed in the brownalgae could be attributed to various phenols whichoccur commonly in these plants. Of most interestwere those compounds with in vivo activity, andthese include an alkylated resorcinol isolated fromCystophora torulosa (Fig. 2a), a phloroglucinolfrom Cystophora scalaris (Fig. 2b) and a 8-toco-trienol from Cystophora expansa (Fig. 2c) .
Sa
Spy
Spn
Ec
Pm
Ss
Pa
Ca
Cn
Tm
Ma
TI*
439
434
150
435
76
22619
3
12
36
14
21(8.8%) (8 .3%) (18 .9%)
(9.3%)
16
0
0
4
0
100
0
0
0
0
0

Table 3 . Summary of activities against the various microbestested arranged according to algal division.
HO
o
Br Br
b R=H, X=Y=BrC R=H, X=Br, Y=Hd R=OAc, X-Br, Y=He R=OH, X=Br, Y=H
Fig. I . Examples of antimicrobially active compounds from theRhodophyta . a - from Ptilonia australasica ; b - from Deliseshypnoides ; c, d, e - from Delises fimbriata.
A typical spectrum of activity and toxicity of redalgal compounds is shown by the fimbrolides ofDeliseafımbriata . Their in vitro antifungal activitieswere comparable to the standard antibiotics (5-fluo-rocytosine, nystativ) used as controls . The antiyeastand antibacterial activities also approached those ofstandard antibiotics (5-fluorocytosine, ampicillin,carbenicillin), particularly in their activity againstEscherichia colí and Staphylococcus aureul, butwith much less activity on Proteus mirabilis andPseudomonas aeruginosa . Both compounds weremoderately toxic to mice . A single dose of com-pound 1 e had a 50%lethal dose (LD 50) of 12 mg kg- 'subcutaneously and 100 mg kg' orally ; a single doseof compound id had an LD50 of 24 mg kg' subcu-taneously and 360 mg kgl orally. When tested at
OH
0
C
165
Fig. 2. Examples of antimicrobially active compounds from thePhaeophyta. a - from Cystophora torulosa; b - from Cystophorascalaris ; c - from Cystophora expansa .
Totalnumber ofextracts
% in vitroactive
Total testedin vivo
% in vivoactive
Chlorophvta (Total number of algae tested = 26)Sa 71 7 .0 2 0Spy 55 32 .7 lI 0Spn 55 36 .4 10 0Ec 70 4 .3 0 0Pm 71 0 0 0Ss 7 0 0 0Pa 71 0 0 0Ca 68 0 0 0Cn 28 3 .6 0 0Tm 71 0 0 0Ma 29 13 .8 0 0Tf 15 13 .3 2 0
Total 702 1 I_3
Phaeophyta (Total number of algae tested = 61)Sa 175 32 .0 36 13 .9Spy 150 48 .0 18 22 .2Spn 151 5 1 .7 42 0Ec 176 6 .8 8 0Pm 175 6 .9 8 0Ss 8 0 0 0Pa 176 10 .8 16 0Ca 176 0 .6 0 0Cn 41 2 .4 0 0Tm 173 12 .1 4 0Ma 23 26 .1 0 0Tf 112 11 .6 6 0
Total 1792 22 .5
Rhodophyta (Total number of algae tested = 58)Sa 155 142 5 0Spy 113 19 .5 8 0Spn 113 25 .7 17 11.8Ec 158 1 .9 2 0Pm 158 1 .3 0 0Ss 2 0 0 0Pa 158 0 0 0Ca 156 1 .3 0 0Cn 72 13 .9 0 0Tm 157 8 .9 0 0Ma 21 19 .1 0 0Tf 70 8 .6 2 0
Total 1523 I1 .5
Cyanophyta (Total number of algae tested = 7)0Sa 11 9 .1 1
Spy II 36 .6 2 0Spn Il 36 .6 2 0Ec 11 0 0 0Pm 11 0 0 0Pa 11 0 0 0Ca 11 0 0 0Cn 9 0 0 0Tm I1 9 .1 0 0Tf 9 0 0 0
Total 106 9 .0

166
their maximum tolerated doses both orally andsubcutaneously, neither was able to cure mice withoverwhelming infections of either S. aureus or E.coli. Subcutaneous ampicillin treatment cured bothinfection with a 50% effective dose (EDS0) of0.5 mg kg* Hence these compounds do not func-tion as systemic antibiotics for bacterial diseases .Neither were they effective as topically appliedagents to clear infected wounds . This was assessed byestablishing an infection in the subcutaneous tissuesof the mice and delivering the test compound to thesite of infection, thereby avoiding the absorptionand elimination problems encountered by systemicagents. The maximum tolerated doses of these com-pounds were unable to clear subcutaneous infec-tions in mice by S. aureus or T. mentagrophytes,whereas the topical control agents neomycin andpolymyxin B sulfate clear S. aureus and E. colíinfections with ED Sa s of 7 ~g m1 - ' . The main reasonsforthese compounds failing to be active in tissues areinactivation and binding by serum . In vitro tests oftheDafimbriata compounds in the presence of 50%human serum showed that S. aureus activity wasreduced 500- to 1000-fold by the serum. Doses highenough to overcome this effect could not be adminis-tered to mice because of the toxicity of the com-pounds .
By far the most promising antimicrobial com-pounds isolated came from the brown algal genusCystophora . Most of those with in vivo activityagainst Gram-positive bacteria compounds belongto a series of resorcinols and phloroglucinols . Forexample, the alkyl resorcinol (Fig . 2a) isolated fromCystophora torulosa had a local in vivo EDSO of2300 ~g m1-~ and had weak local activity againstlocal infections of mice by S. pyogenes but not S .aureus . Phenolics are generally considered to betoxic compounds ; however, the alkyl group isknown to render them less toxic. The phloroglucinol(Fig. 2b) isolated from Cystophora scalarls with alocal EDSO of 250 ~g ml- ' also had local in vivoactivity against S. aureus . Neither of these com-pounds was found to have systemic activity . Studieson the in vivo activities of the `crude' extracts of thevarious Cystophora species were not completed dueto the untimely closure of the Institute ; however,preliminary data indicated that the other activitieswere probably due to compounds related to the onesmentioned above. The active dichloromethane ex-tract of Cystophoraexpansa contained a mixture of
small amounts of the phloroglucinols found in C .scalarls as well as the 5-tocotríenol (Fig . 2c) . Themajor metabolite isolated from C. monilifera was ahydroxylated derivative of the phloroglucinol origi-nally isolated from C. scalarls (Fig. 2b), althoughthis compound was found to be unstable on storage .Fractionation of the `crude' extracts from the otherCystophora species was not completed, and testingof some of the compounds isolated from them(Gregson, et al 1977 ; Kazlauskas, et al 1981 ; Ravi, etal 1982) was also not completed .
Discussion
Compounds with in vitro antimicrobial activityhave often been isolated from marine algae (seereferences listed in the Introduction), and theRRIMP screening programme obtained similar re-sults . Most of the previous studies have, however,not proceeded further. There would seem to be tworationales for studying antibiotic production by al-gae and aquatic plants : investigating the ecologicalrole of these antibiotics in preventing disease in thealgae or preventing fouling by bacteria, other algaeor other organisms, or the search for new antibioticsfor therapeutic use in medical and veterinary areas .
The ecological role of algal antibiotics has onlyrarely been examined . In those few studies whereactivity against marine bacteria has been examined,this was often disappointing. For example Fenicaletal( 1977) tested the antibiotic activity of a number ofhalogenated compounds isolated from algae fromthe red algal family Bonnemaisoniaceae. Many ofthese were very inhibitory to the growth ofS. aureusand Candida albicans but had only slight activityagainst the marine bacterium Vibrin anguillarum .S ignificant activity against marine bacteria has onlybeen shown so far by 3,6,7-trihydroxycoumarin, asubstance isolated from the green dasycladaleanalga Dasycladus vermicularis (Menzel, etal 1983) . Itis, however, also to be expected that the variousphenolics excreted by many brown algae (Glombit-za 1979) would also inhibit marine bacterial growthas well as retard fouling of the alga in general (AI-Og-ily & Knight-Jones 1979) . Ecologically, the mainaction of many of the substances with in vitro activi-ty against nonmarine microorganisms may be todeter herbivore grazing, and many of them havebeen shown to have piscicidal activity or other graz-

ing-deterrent activity (e.g ., Fusetani, et al 1976 ;Ohta 1979) .
The studies at RRIMP were of course directed totherapeutically relevant compounds, and in generalone must conclude that algae are a rather disappoint-ing source of these . Only a very small number of theextracts active in vitro showed any activity in vivo .Many of the compounds active in vitro were eithertotally inactive in vivo, or proved to be extremelytoxic . Toxicity was especially common in the ex-tracts of red algae. Many compounds with an accep-table level of toxicity were either rapidly inactivatedby binding with serum proteins, were not absorbedand circulated to the site of infection, or were rapidlyeliminated either by excretion or metabolism toinactive compounds .
Antimicrobial compounds may, however, alsofind use as soaps, lotions, shampoos, creams etc.,applied to the unbroken skin as general body cleans-ing agents. An important clinical application of thisclass of topical agents is the need to control skinlevels of S . aureus on newborn babies in hospitalsand on surgeons during operations . Besides havinggood antimicrobial activity, such compounds mustbe suitable for more frequent and prolonged usethan a systemic antibiotic or a wound cleanser . Inthis case, the nature and degree of toxicity are impor-tant. (Toxicity is also important for systemic antibi-otics, but a novel systemically active compound withsome degree of toxicity may still find application foruse against life-threatening infection or as a wound-cleanser). All topically applied chemicals wí11 beabsorbed through the skin to a certain extent, givingrise to the possibility of occasional episodes of acutetoxicity from abnormally excessive use, or to adeveloping chronic toxicity if the compound ac-cumulates or has cumulative effects . In no area didwe perform tests of chronic toxicity, but in the caseof many of the compounds from the red algae, suchas those from the Delisea species, the degree of acutetoxicity from a single dose precludes an applicationas a topical skin cleanser . On the other hand theactive compounds from the brown algae, such asthose isolated from the Cystophora species, couldpossibly be used in such applications if they werefound to be more active and less toxic than presentlyused skin cleansers such as hexachlorophene . Fromour experience at RRIMP it seems clear that thebrown algae may be a potential source of suchcompounds .
167
Antimicrobial compounds are also used in a varie-ty of industrial applications including the cleansingof surfaces and utensils (antiseptics), the preventionof microbial deterioration (preservatives, inhibitingagents) and the inhibition of growth of microbes incooling water, etc . (toxins) . Such antimicrobialcompounds have as a major requirement that they bevery cheap to produce, especially compared to theantibiotics and topicals in human use . This may,however, be tempered somewhat by the ever-in-creasing awareness of the environmental impact ofsome of the substances presently in use . Naturalproducts such as those isolated from marine algaemay be more readily biodegradable and thereforemore acceptable, even though at a higher cost, in thefuture. Thus the novel antimicrobial compoundsisolated from algae so far, and those which will beisolated in the future, may provide leads for thedesign ofantiseptics and industrial inhibitory agentsfor use in industry.
At RRIMP a large number of marine algae (andinvertebrates) were screened for their antibiotic ac-tivity, but it must be remembered that our totalsample of organisms tested is still very small com-pared with the large numbers of species of algae andother organisms living ín our oceans . Marine organ-isms clearly contain a wide range of very novelchemicals (Baker& Murphy 1981 ; Ragan 1981), andit is not improbable that some of these may be verypotent and effective antibiotics .
Acknowledgements
The results reported here are the combined workof our colleagues in the Microbiology, Biology andChemistry Sections of the Roche Research Instituteof Marine Pharmacology .
References
Al-Ogíly, S . M . & E . W . Knight-Jones, 1977 . Anti-fouling role ofantibiotics produced by marine algae and bryozoans . Nature(Lood .) 265 : 728-729 .
Bach, B ., G . S . Holland, R . P . Gregson, R . J . Quinn & J . L .Reichelt, 1978 . L-Azetidine-2-carboxilic acid, the antider-matophyte constituent of two marine sponges . Experientia34 : 688 .
Baker, J . T. & V. Murphy, 1981 . Compounds from MarineOrganisms . CRC Press, Florida 2: 233 pp .

Burkholder, P . R ., L . M . Burkholder& L . R . Almodovar, 1960 .Antibiotic activity of some marine algae of Puerto Rico . Bot .mar . 2 : 149-156 .
Fenical, W ., O . J . McConnell & A . Stone, 1977 . Antibiotics andantiseptic compounds from the family Bonnemaisoniaceae(Florideophyceae) . Proc . int . Seaweed Symp . 9 : 387-400 .
Fusetani, N ., C . Ozawa & Y . Hashimoto,1976 . Studies on marinetoxins . LITI . Fatty acids as ichthyotoxic constituents of agreen alga Chaetomorpha minima. Bull . jap . Soc . scient .Fish . 42 : 941 .
Glombitza, K . W ., 1979 . Antibiotics from algae. In H . A . Hoppe,T. Levring & Y. Tanaka(eds.), Marine Algae in Pharmaceut-ical Science . Walter de Gruyter, Berlin.
Gregson, R . P ., R . Kazlauskas, P . J . Murphy & R . J . Wells,1977. New metabolites from the brown alga Cystophoratorulosa . Aust . J . Chem. 30: 2527-2532 .
Grunberg, E. & R. J . Schnitzer, 1959 . Experimental approach totopical antibacterial therapy . Ana N . Y. Acad . Sci . 82 :114-118 .
Grunberg, E., J . Berger, G . Beskid, R. Cleeland, N . N . Prince &E. Titsworth, 1967 . Studies on the in vitro and in vivochemo-
Studies on the biologically active compounds of the algae from the Yellow Sea
Lí Lian Niang & Xue HungInstitute of Materia Medica, Chinese Academy of Medical Sciences, Beijing, People's Republic of China
Keywords: seaweed, bromophenol, diterpenes, Dictyota indica, dictyotriol A, dictyotriol B
Introduction
Marine algae have been used in traditional andfolk medicine in China for more than two thousandyears . Several species of algae are listed in earlyChinese materia medica (Shen Nung Ben Cao Jing,200 BC and Ben Cao Gang Mu, 1518-1593) fortheir antifebrile, antiedema, diuretic and expector-ant functions . In the past few years, screening forantibacterial activity, effect on coronary bloodflow, and in the hypoxia test in mice of more than80 species of algae has been carried out in severalinstitutions in China (Zhang J .-d ., Tan Mu., WuP.-p ., Li L., Fu D.-m. & Hang J .-x ., unpubl.) .During our investigation on the biologically activecompounds of the algae from the Yellow Sea, thecrude extracts of Rhodomela confervoides, Sym-phyocladia latiuscula (Rhodomelaceae) and Dicty-ota indica (Dictyotaceae) showed antiinflammato-
© 1984, Hydrobiologia 116/117 .
therapeutic properties of the antibiotic myxin . Chemothera-pia 12 : 272-281 .
Hornsev, I . S . & D. Hide, 1974 . The production of antiroicrobialcompounds by British marine algae, I . Antibiotic-producingmarine algae . Br . phycol . J . 9 : 353-361 .
Kazlauskas, R ., L . King, P . T . Murphy, R . G . Warren & R . J .Wells, 1981 . New metabolites from the brown algal genusCystophora. Aust . J . Chem . 34 : 439-447 .
Menzel, D ., R . Kazlauskas & J . Reichelt, 1983 : Coumarins inthe Siphonalean green algal family Dasycladaceae Kützing(Chlorophyceae) . Bot . mar . 26 : 857-864 .
Ohta, K ., 1979 . Chemical studies on biologically active substan-ces in seaweeds . Proc . int. Seaweed Symp . 9 : 401-412 .
Ragan, M . A ., 1981 . Chemical constituents of seaweeds . In C . S .Lobban & M. J . Wynne (eds .), The Biology of Seaweeds .Blackwell Scientific Publ . Oxford : 589-626 .
Rao, P . S . & K . S . Parekh, 1981 . Antibiotic activity of Indianseaweed extracts . Bot . mar . 24 : 577-582 .
Ravi, B . N ., P . T. Murphy, R . O . Lidgard, R. G . Warren & R . J .Wells, 1982 . C18 terpenoid metabolites of the brown algaCystophora moniliformis . Aust . J . Chem . 35 : 171-182 .
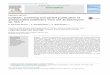
H
CH2ORtR2
Br 1 . Rt=R2=H~ . Ri CH3 ; R2=H
4 . Ri= CH3+ RřBr
5 . Rı = C2Hς, R2=B'
0
B
3
ry activities in mice (Gao Y.-g ., unpubl.) . Bromo-phenols 1, 2 and 3 were isolated from R. confer-voides, and 4 and 5 from S. latiuscula . Two newhydroazulene diterpenes, dictyotriols A (6) and B (7)were isolated from the anti-inflammatory-activeportion of D. indica. We now wish to report theisolation and structure elucidation of these two newditerpenes .
2