Embed Size (px)
Citation preview

Vox Sang. 17: 393-405 (1969)
Antigenic Markers on Cultured Human Cells
III. The MN Antigens
ANNE DAWSON~
Department of Pathology, University of Cambridge, Cambridge
Summary. The cross-reactions of M and N antibodies and the MN antigens of human tissue cells in culture have been investigated by mixed agglutination. Cross-reactions have been found chiefly between the N antigen of tissue cells and anti-M and anti-N antibodies; and between anti-N antibodies and the M antigen of red cells.
Attempts were made to reduce cross-reactivity by testing the cells a t different temperatures and after treatment with enzymes. These conditions were also im- posed upon red cells in direct agglutination using the same antisera but there was little correlation between the results of the direct agglutination and mixed agglu- tination tests.
Introduction
MN antigens of human cells have been looked for using the mixed agglutination test by GURNER and COOMBS [8] and KELUS, GURNER and COOMBS [13] who reported that MN antigens were present on HeLa cells and leucocytes. ASHHURST, BEDFORD, COOMBS and ROUILLARD [l] were not able to detect MN antigens on freshly trypsinised skin cells, while EDWARDS, FERGUSON and COOMBS [6] showed that MN antigens were present on spermatozoa. It was found that there was cross-reactivity between anti-N serum and M indicator red cells on HeLa cells, and with human leucocytes there was a positive reaction when anti-M serum was used with group M indicator cells whatever the group of the leucocyte. The MN group of the person from whom HeLa cells were derived is not known. Similar cross-
Sir Halley Stewart Research Student.

394 DAWSON Antigenic Markers on Cultured Human Cells
reactions were found with spermatozoa. Mixed agglutination was only achieved with rabbit antisera, never with human antisera.
This paper describes the investigation of several human cell lines for the presence of the MN antigens and shows that there is wide cross-reaction between the M and N antigens and their antibodies. Cross-reaction can be described as the reaction of an antiserum, specific for one antigen with a closely related antigen. It must be distinguished in practice from contamination of the antiserum with other antibodies. Thus if the antiserum is absorbed with homologous antigen, the titre against the related antigen should also be lowered. If the antiserum is allowed to react with the homologous antigen, and the antibodies are then eluted off, these eluted antibodies should then be capable of reacting with the related antigen. The term cross- reaction has also been applied to the situation when two antigenic sites have determinants in common and the antibodies of a so-called specific antiserum are able to attach to either (e.g. Type XIV pneu- mococcal polysaccharide and blood group A substance). The extent t o which MN cross-reactivity fulfils either of these criteria is not known.
Since it is known that changes in temperature and treatment with enzymes affect the cross-reactivity of anti-M and anti-N antibodies [lo], the tissue cells were tested by the mixed agglutination tech- nique at different temperatures and after treatment of both test and indicator cells with enzymes. The cells are normally sub-cultured by the use of trypsin to remove them from the glass on which they grow, and since trypsin alters the MN antigens [15], the effect of trypsin was investigated to see if i t was this which was in- fluencing cross-reactivity. Neuraminidase splits off sialic acid; this enzyme was used to see whether it had any effect on the cross- reactivity or if the antigenic sites would be destroyed. As these various treatments made little difference to mixed agglutination, the same reagents were used 04 red cells to see whether direct agglutina- tion was affected.
It remained a possibility that the antigen being detected, especially after enzyme treatment, was the ‘T’ antigen (Hubener-Thomsen- Friedenreich phenomenon), as this antigen can be detected on most red blood cells which have been exposed to enzyme treatment or the activity of bacteria. This was investigated by using a specific anti-T lectin from Arachis hypogeae [2] and by absorption of antisera with enzyme treated red cells.

DAWSON Antigenic Markers on Cultured Human Cells 395
Materials and Methods
Tissue cells. Primary cultures were initiated from foetal lung, skin and kidney; and from adult thyroid tissues obtained from patients undergoing surgical thyroid- ectomy. Continuous cell lines were also maintained in the laboratory, including HeLa 229 (CCL 2.1), Detroit 98 (CCL 18) and AV3 (Amnion) (CCL 21). These were obtained initially from the American Type Culture Collection.
Culture medium. Tissue culture medium 199 (Flow Laboratories) was used with added sodium pyruvate 100 mM (1 ml/l), penicillin (100 units/ml), and strepto- mycin (100 pg/ml). 10% foetal bovine serum (Flow Laboratories) was added which had been tested before use by immunoelectrophoresis to confirm that it was foetal serum.
Culture procedure. Unless otherwise stated, the tissue cells were allowed to grow to confluence in soda glass bottles and were then rinsed with 0 . 0 0 0 5 ~ EDTA, followed by treatment with 0.25% Difco Trysin (1:250) for 15 min or until the cells came off the glass. Medium was then added to inhibit the trypsin, the cells were spun down, the supernatant discarded, and the cells resuspended in fresh medium and distributed into fresh bottles.
The mixed agglutination test. This was carried out by the method of COOMBS, DANIEL, GURNER and KELUS [4] except that 0.02% crystalline bovine serum albumin (Armour) in buffered saline was used as the diluent instead of diluted normal rabbit serum. Indicator cells used were from group OM or ON donors taken into ACD anticoagulant from an earprick or venepuncture.
Antisera. These were rabbit anti-M and anti-N sera given by Dr. E. W. IKIN, MRC Blood Group Reference Laboratory; and human immune anti-M and anti-N sera from various sources.
Blood grouping of cell donors. The donors of cells for all primary cell lines were blood grouped for the ABO and MN blood groups. Heart blood was obtained from foetuses; red cells were obtained from thyroidectomy patients a t the time of operation.
Neurarninidase. Behringwerke (500 units/ml) neuraminidase was used diluted to give 20 units/ml in 0.45 M NaCl, 0.005 M CaC1, buffer, made pH 7.4 with 0.002 M NaHCO,. A 2% suspension of red cells was treated with diluted neuraminidase for 25 min a t 37°C. The cells were then washed in saline once before use.
Buccal cells were scraped from the buccal mucous membrane into saline just before use and washed once.
Absorption. Small volumes of serum, for example 0.2 ml, were absorbed with an equal volume of packed red cells, which had been treated with enzyme, for 15 min. The absorbed serum was tested for agglutination by a tile test and absorption repeated if necessary, until no agglutination was observed.
Direct agglutination. Doubling dilutions of antisera were made and one drop of diluted antiserum placed with one drop of a 2% suspension of red cells in an Rh tube and allowed to stand for 2 h. Agglutination was looked for with the aid of a microscope.
Preparation of Arachis Zectin. The seeds of Arachis hypogeae (‘monkey nuts’ or ‘peanuts’) were shelled, peeled, and ground. The lectin was then extracted in five times the dry weight of 10% saline, in the cold, for 24 h. The supernatant was dialysed against 0.9% saline for two to three days with frequent changes of saline.

396 DAWSON Antigenic Markers on Cultured Human Cells
Results
Mixed Agglutination Results
Mixed agglutination on various tissue cells. MN antigens were never demonstrated by the mixed agglutination test on freshly trypsinised adult (cadaver) and foetal skin cells, nor on buccal cells freshly scraped from the buccal mucous membrane. MN antigens were detected, however, on the continuous cell lines HeLa 229, Detroit 98, and AV3 (Amnion), and on primary cell lines of foetal cells and adult thyroid cells after a t least one subculture.
Mixed agglutination with human anti-M and anti-N sera. This was consistently negative.
Mixed agglutination with rabbit anti-M and anti-N sera. Not all rabbit antisera obtained gave mixed agglutination, but most did : Of 8 rabbit anti-M sera, only 2 failed to give mixed agglutination and of 9 rabbit anti-N sera 4 failed to give mixed agglutination. All rabbit antisera which were used for the mixed agglutination tests were specific for their homologous red cells when tested for red cell agglutination, but all gave some degree of cross-reactivity in the mixed agglutination test. Some anti-M sera gave positive mixed agglutination on group MN tissue cells with both group M and N indicator cells; other anti-M sera were positive only with group M indicator red cells. Most anti-N sera only gave mixed agglutination with group N indicator cells on group MN tissue cells. With group M tissue cells there was no cross-reactivity. With group N tissue cells there was cross-reaction when group M indicator cells were used with both anti-N and anti-M antisera (table I).
Attempts to Prevent Cross-Reaction in the Mixed Agglutination Test
The effect of temperature. The entire mixed agglutination test was done a t three different temperatures: 4"C, 20°C and 37°C. Cells still gave mixed agglutination in the non-homologous systems (anti-M with N indicator cells; anti-N with M indicator cells) a t all tempera- tures.
The effect of trypsin. Cells were subcultured without the use of trypsin, that is, by scraping them from the glass with a piece of

DAWSON Antigenic Markers on Cultured Human Cells 397
Table I . Cross reactions in mixed agglutination tests
Tissue Indicator cell group Antiserum cell group Result
MN Anti-M M +++ MN Anti-M N - MN Anti-N M + t
MN Anti-N N +t+
M Anti-M M +ti
M Anti-M N - M Anti-N M - M Anti-N N -
N Anti-M M + .- N Anti-M N - N Anti-N M + + N Anti-N N +++
t = cross reaction.
silicone rubber. These were tested in parallel with the same cells which had been subcultured by using trypsin, but the test showed no significant difference in the number of positive cells cross-reacting, although the trypsinised cell showed an enhanced positive reaction. The ‘scraped’ cells suffer more damage than trypsinised cells, and there were more dead cells when the test was read when the cells had been removed by scraping.
The effect of neuraminidase. A monolayer of tissue cells, and tissue cells in suspension were treated with neuraminidase for 25 min a t 37°C. Both were tested in suspension by the mixed agglutination technique, but the positive reaction was not abolished, nor was there any difference in the number of cells giving mixed agglutination in non-homologous systems. Red cells were then treated similarly and used as indicator cells on both treated and normal tissue cells. No difference was observed in the amount of cross-reaction, nor was the reaction abolished, although i t appeared t o be somewhat weaker, that is, fewer tissue cells were positive, and the positive cells had fewer indicator cells adhering to them.
I n order t o maintain the conditions as much the same as possible these experiments were all done with group MN tissue cells using

398 DAWSON Antigenic Markers on Cultured Human Cells
anti-M and anti-N sera with M and N indicator cells. The antisera used were those which showed least cross-reaction in tests under normal conditions.
Investigation of the Effect of Temperature and Enzymes on Red Cells
The eflect of temperature. Rabbit and human anti-M and anti-N sera were titrated against M and N red cells a t 4"C, 20°C and 37°C with the following results: (table 11). Rabbit anti-M serum had a raised titre a t 37°C from 1:32 to 1:256. There was no cross-reaction with N cells a t any temperature. Rabbit anti-N serum showed no difference in titre a t any temperature, nor any cross-reaction with M cells. Human anti-M serum had a slightly increased titre at 37°C but there was no cross-reaction with N cells a t any temperature. With human anti-N serum, the reaction was abolished at 37°C and enhanced a t 4°C. There was no cross-reaction a t any temperature with M cells.
The effect of trypsin. Rabbit and Human anti-M and anti-N sera were titrated against a 2% suspension of red cells which had been treated with 0.25% trypsin for 25 min a t 37°C: (table 111). Rabbit anti-M serum showed no difference in titre with treated or non- treated M red cells, but there was strong cross-reaction with trypsin- treated N cells. Rabbit anti-N serum had the same titre with trypsin
Table 11. Effect of temperature on agglutination of red cells
Red Cells Temperature Rabbit Human Anti-M Anti-N Anti-M Anti-N
- M 4°C + (32) - + (8) M 20°C + (64) - + (8) - M 37°C ++ (256) - ++(16) -
+ = agglutination ++ = enhanced agglutination ( ) = titre of antiserum 1 = undiluted serum

DAWSON Antigenic Markers on Cultured Human Cells 399
Table 111. Effect of trypsin treatment on agglutination of red cells
Red cells Rabbit Human Anti-M Anti-N Anti-M Anti-N
- - + (8) + (8) -
M rbc (normal) + (64) M (trypsin-treated) + (64) + (4)
+ (1) N rbc (normal) - N (trypsin-treated) + (16) + (4) + (8)
- + (4)
-
-c = agglutination - = no agglutination ( ) = titre of serum 1 = undiluted serum
treated N red cells as with normal cells and there was strong cross- reaction with trypsin treated M cells. The reaction between human anti-M serum and trypsin-treated M cells was abolished and there was no cross-reaction between human anti-M and trypsin-treated N cells. The reaction between human anti-N and trypsin-treated N cells was enhanced and cross-reaction occurred with trypsin-treated M cells.
A more highly purified trypsin preparation was obtained (Armour ‘Tryptar’) and used t o treat red cells at the same concentration (0.25%) as the Difco trypsin. Exactly the same results were obtained with both preparations.
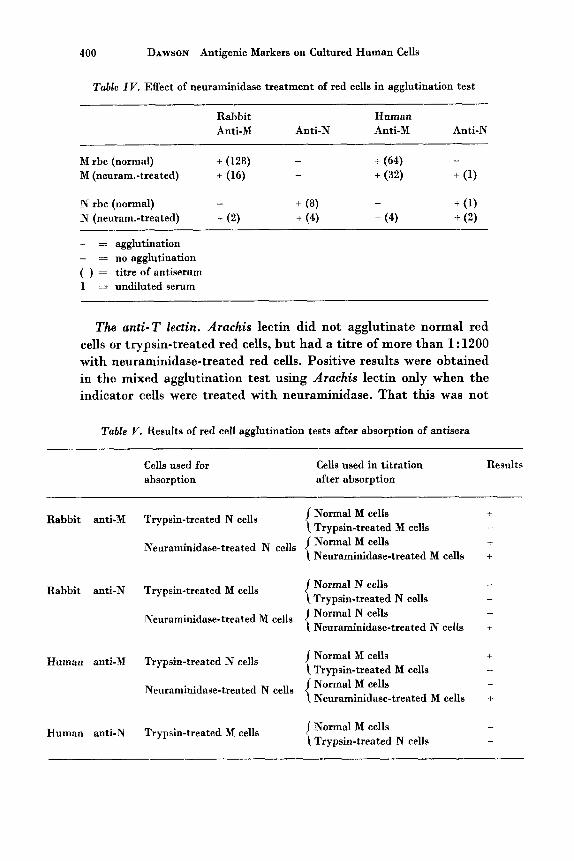
The eflect of neuraminidase. Two percent suspensions of red cells were treated with 20 units/ml neuraminidase for 25 min a t 37°C and used in the direct agglutination test against rabbit and human anti-M and anti-N antisera: (table IV). The titre of rabbit anti-M serum was lowered with neuraminidase-treated M cells, and there was some cross-reaction with treated N cells, but the titre of rabbit anti-N serum was reduced with neuraminidase-treated N cells, and there was no cross-reaction with treated M cells. Human anti-M serum had a lower titre with treated M cells and cross-reacted with treated N cells, while human anti-N serum had a higher titre with treated N cells and showed some cross-reaction with treated M cells. Neura- minidase-treated M and N cells were agglutinated by normal human AB serum and a rabbit serum which had no anti-M or anti-N activity.
400 DAWSON Antigenic Markers on Cultured Human Cells
Table IV. Effect of neuraminidase treatment of red cells in agglutination test
Rabbit Human Anti-M Anti-N Anti-M Anti-N
+ (64) M rbc (normal) + (128) - -
M (newam.-treated) + (16) + (32) + (1) -
+ (1) N rbc (normal) -
N (neuram.-treated) + (2) + (4) + (4) + (2) -
+ ( 8 )
+ = agglutination - = no agglutination ( ) = titre of antiserum 1 = undiluted serum
The anti- T lectin. Arachis lectin did not agglutinate normal red cells or trypsin-treated red cells, but had a titre of more than 1 : 1200 with neuraminidase-treated red cells. Positive results were obtained in the mixed agglutination test using Arachis lectin only when the indicator cells were treated with neuraminidase. That this was not
Table V. Results of red cell agglutination tests after absorption of antisera
Cells used for absorption
Cells used in titration Result3 after absorption
Normal M cells + Trypsin-treated M cells + Normal M cells Neuraminidase-treated M cells +
c Rabbit anti-M Trypsin-treated N cells
+ Neuraminidase-treated N cells
Normal N cells Trypsin-treated N cells
Rabbit anti-N Trypsin-treated M cells _ _
Normal N cells - Neuraminidase-treated N cells +
Neuraminidase-treated M cells
Normal M cells + Trypain-treated M cells - Normal M cells Neuraminidase-treated M cells +
c Human anti-M Trypsin-treated N cells
+ Neuraminidase-treated N cells
Normal M cells - c Trypsin-treated N cells Human anti-N Trypsin-treated M cells -

DAWSON Antigenic Markers on Cultured Human Cells 40 1
due to the enzyme treatment of the red cells was shown by the fact that the natural anti-T in sera which normally gives no mixed agglutination also failed to give it when the indicator cells were treated with trypsin or neuraminidase.
Absorption tests (table V). Rabbit anti-M serum was absorbed with both trypsin-treated and neuraminidase-treated N cells. After ab- sorption there was still some agglutinating activity with untreated M red cells. One anti-M serum which was absorbed with trypsin- treated M red cells, gave agglutination with untreated cells but failed t o agglutinate the untreated M cells of another donor. Rabbit anti-N serum which was absorbed with trypsin-treated and neuraminidase- treated M cells failed t o agglutinate normal N cells, but agglutinated neuraminidase-treated N cells. Human anti-M serum which was ab- sorbed with trypsin-treated and neuraminidase-treated N cells still agglutinated normal M cells, but human anti-N serum which was absorbed with trypsin-treated M cells failed to agglutinate normal N cells.
Discussion
The MN antigens cannot be demonstrated on freshly disaggregated tissue cells but after a few subcultures in tissue culture they become readily demonstrable. This may be contrasted with the behaviour of the AB antigens which are readily detected on freshly disaggregated tissue cells [12] but gradually disappear when these cells are grown in tissue culture [5, 111. Similarly, buccal cells scraped from the buccal mucous membrane have the A and B antigens but not the M and N antigens. Attempts have been made t o retain the A and B antigens in culture by growing them in precursor sugars, without success, so i t would not appear t o be the lack of these sugars in the medium which leads to their loss [7]. It may be that the cells cease to metabol- ise the antigens in order t o survive a t the least complex level in the simple environment of cell culture, but i t is difficult t o see why the M and N antigens are any more necessary to cell survival than A or B or that they are more readily synthesised. It could be due to differences of cell shape when they become adherent to glass, some antigenic sites being hidden and others exposed. Since both fibro- blasts and epithelioid cells show loss of A and B and retention of M and N antigens the morphology of the cell probably plays no part.
It is difficult to explain why human antisera should not give mixed agglutination. If i t is postulated tha t the trypsin treatment of the

402 DAWSON Antigenic Markers on Cultured Human Cells
tissue cells during culture has the same effect as trypsin treatment of red cells subsequently tested for direct agglutination, then, al- though the agglutinating activity of human anti-M serum is abolished, the anti-N activity should not be. It may be that human antisera having a generally lower titre than rabbit antisera, do not possess the ability of giving mixed agglutination. However, a human anti-M serum was available which had a high direct agglutination titre of 1 : 64 and which reacted more like a rabbit antiserum in that the agglutinating activity was not abolished when titrated with trypsin- treated M cells, although it was diminished. This antiserum did not give mixed agglutination in spite of its high antibody titre. Further differences are shown between rabbit and human antibodies against M and N antigens in their differing reaction against enzyme treated red cells. These observations suggest that it is possible that rabbit and human antisera t o the MN antigens may not have exactly the same specificity.
The cross reactivity of rabbit antisera in the mixed agglutination test shows somewhat surprising results. Even those antisera which were ‘specific’ showed a regular pattern of cross-reactivity. This appears to be mainly concerned with the N antigen of the tissue cell, as group M tissue cells show little cross reactivity. Both anti-M and anti-N antibodies seem to recognise the N antigen of the tissue cell and go onto the site during sensitization; when the red cells are added to these sensitized tissue cells, apparently the M red cell can go onto both the anti-M and anti-N antibodies, but the N cell attaches only to the anti-N antibody. This cross-reaction of anti-M with N antigen has not so far been observed between anti-M and untreated N red cells. Without knowing the exact composition, size and shape of the M and N antigenic determinants it is difficult to say why this cross- reaction occurs. It is known that bacterial antigens show cross- reaction when their determinants are very similar [9]. It would not seem that this cross-reaction between M and N is due to a purely cross-reacting antibody, as judged by the criteria of absorption and elution: (that the titre of the homologous antigen is lowered when the antiserum is absorbed by heterologous red cells; that if the anti- body is eluted from homologous red cells it will then agglutinate the heterologous red cells). Although anti-N satisfies these criteria t o some extent it appears from the mixed agglutination results that it is the anti-M antibody which fails to discriminate between the M and N tissue cell antigenic sites, but the anti-N antibody which

DAWSON Antigenic Markers on Cultured Human Cells 403
fails t o distinguish between the M and N antigenic sites of the red cell.
Variations in the temperature and use of trypsin had no effect on the cross reactivity between M and N antigens as detected by mixed agglutination. Since there was no difference a t any temperature in the direct agglutination test using these same sera this was not too unexpected. It is more surprising, however, that omitting the use of trypsin on tissue cells had no effect in the mixed agglutination test, as in the direct agglutination test there is a great deal of cross- reaction when trypsin-treated red cells are used. It seems reasonable that trypsin-treated tissue cells give an enhanced positive reaction in the mixed agglutination test as (a) the cells are in a better con- dition and (b) trypsin also increases the red cell agglutination reaction using rabbit antisera.
Neuraminidase is known to release sialic acid from both red cells and tissue culture cells, for example HeLa [3], but a t the strength of enzyme used in these experiments i t was obvious that the M and N antigenic sites were not destroyed when either tissue cells or red cells were treated. However, the number of red cells adhering to positive tissue cells was reduced when both tissue cells and indicator cells were treated in the same test, indicating that there may be some breakdown of MN sites. There may also be exposure of T antigenic sites, since cross-reactivity was increased in the direct agglutination test with treated red cells (with the exception of rabbit anti-N serum which did not cross-react with neuraminidase-treated M cells). Neuraminidase must break down the M and N determinants t o expose a different antigenic site. Longer treatment a t higher concentrations was not tried but this may have eventually removed most of the sialic acid and inactivated the antigenic sites, as indicated by the work of SPRINGER and ANSELL [14].
The tests with Arachis lectin confirm that neuraminidase exposes an antigenic determinant on red cells which is different from the site exposed by trypsin. It would also appear that this determinant occurs naturally on cultured tissue cells since mixed agglutination is obtained with tissue cells which have not been treated with neuraminidase but sensitized with Arachis lectin and using neuraminidase-treated in- dicator cells.
It seems unlikely that the cross-reaction obtained in these experi- ments with anti-M and anti-N can be due solely to the T antigen, as there is such a difference in the reactions between neuraminidase-

404 DAWSON Antigenic Markers on Cultured Human Cells
treated and trypsin-treated red cells with human and rabbit antisera. Also, since anti-M sera can be absorbed with enzyme treated N cells and still retain its anti-M activity, this shows that the serum contains a specific anti-M antibody as well as an anti-T antibody.
Anti-N sera absorbed with enzyme treated M red cells failed t o show the continued presence of a specific anti-N antibody, but this is not surprising as the anti-N antibody is easily absorbed out by normal M cells.
These observations of the reactions of anti-M and anti-N antisera show that there is a greater complexity of the M and N antigenic determinants than had been formerly recognised when only the cross- reaction of anti-N sera with M cells a t lowered temperatures and with trypsin treated M cells caused concern. The human anti-N antibody cross reacts with neuraminidase-treated M red cells, whereas the rabbit anti-N antibody does not. Both the anti-M and anti-N antibodies are capable of cross-reacting between tissue antigenic determinants and red cell determinants.
References
1. ASHHURST, D. E.; BEDFORD, D.; COOMBS, R. R. A. and ROUILLARD, L. M.: Blood-group antigens on human epidermal cells. Nature, Lond. 178: 1170 (1956).
2. BIRD, G. W.: Anti-T in Peanuts. Vox sang. 9: 748 (1964). 3. COOK, G. M. W.: Glycoproteins in membranes. Biol. Rev. 43: 363 (1968). 4. COOMBS, R. R. A,; DANIEL, M. R.; GURNER, B. W. and KELUS, A,: Recognition
of the species of origin of cells in culture by mixed agglutination. 1. Use of antisera to red cells. Immunology 4: 5 5 (1961).
5. DAWSON, A.: Unpublished observations. 6. EDWARDS, R. G.; FERGUSON, L. C. and COOMBS, R. R. A.: Blood group antigens
on human spermatozoa. J. Reprod. Fertil. 7: 153 (1964). 7. FLORY, L. L.; FRANKS, D.; DAWSON, A.; LISKE, R. and DANIEL, M. R.: Anti-
genic markers on cultured human cells. 11. The H antigen. Exp. Cell Res. (in press).
8. GURNER, B. W. and COOMBS, R. R. A.: Examination of human leucocytes for the ABO, MN, Rh, Tja, Lutheran and Lewis systems of antigens by means of mixed erythrocyte - leucocyte agglutination. Vox sang. 3: 13 (1958).
9. HEIDELBERGER, M. : Scope and significance of antigenic cross-reactivity ; in TRENTIN: Cross reacting antigens and neoantigens, p. 1 (Williams and Wilkins, Baltimore 1967).
10. HIRSCA, W.; MOORES, P.; SANGER, R. and RACE, R. R.: Notes on some reac- tions of human anti-M and anti-N sera. Br. J. Haemat. 3: 134 (1957).
11. HOGMAN, C. F.: Blood group antigens on human cells in tissue culture. The effect of prolonged cultivation. Exp. Cell Res. 21: 137 (1960).

DAWSON Antigenic Markers on Cultured Human Cells 405
12. HOLBOROW, E. J.; BROWN, P. C.; GLYNN, L. E.; HAWES, M. D.; GRESHAM, G. A.; O’BRIEN, T. F. and COOMBS, R. R. A.: The distribution of the blood group A antigen in human tissues. Brit. J. exp. Path. 41: 430 (1960).
13. KELUS, A.; GURNER, 13. W. and COOMBS, R. R. A.: Blood group antigens on HeLa cells shown by mixed agglutination. Immunology 2: 262 (1959).
14. SPRINGER, G. F. and ANSELL, N. J. : Inactivation of human erythrocyte agglu- tinogens M and N by influenza viruses and receptor-destroying enzyme. Proc. nat. Acad. Sci., Wash. 44: 182 (1958).
15. UHLENBRUCK, G.: Action of a proteolytic enzyme on the human erythroycte surface. Nature, Lond, 190: 181 (1961).
Author’s address: Miss ANNE DAWSON, Department of Pathology, University of Cambridge, Cambridge (England).