Embed Size (px)
Citation preview

Antifungal efficacy of a citrus fruit extract against
Candida albicans cells
by
Daniel Joel Kobric
A thesis submitted in conformity with the requirements for the degree of Masters of Science
Discipline of Periodontology Faculty of Dentistry
University of Toronto
© Copyright by Daniel Joel Kobric 2012

ii
Antifungal efficacy of citrus fruit extracts against
Candida albicans cells
Daniel Joel Kobric
Masters of Science
Discipline of Periodontology
Faculty of Dentistry
2012
Abstract
The number of superficial candidal infections has grown due to an increase in the elderly cohort
and ever-increasing immunocompromised population, and the increased prevalence of antifungal
drug resistance. The aim of this research was to investigate the antifungal efficacy of a citrus
fruit extract, Biosecur c320c (B320), against two strains of Candida albicans (nystatin sensitive
and resistant). The viability of C. albicans strain treated with 10% B320 was reduced by 90-99%
depending on biofilm age. A B320/nystatin combination demonstrated even greater efficacy at
killing biofilm cells in either strain. The nystatin sensitive strain did not develop resistance to
B320 while resistance was developed following long-term exposure to nystatin. Scanning
electron micrographs of C. albicans biofilms revealed cellular debris after treatment with
combined B320/nystatin. Citrus fruit extracts containing polyphenols might contribute to the
development of novel therapeutics for the treatment of oral candidiasis, particularly in those
refractory to nystatin therapy.

iii
Acknowledgments
First and foremost a big thank you to my supervisors and mentors, Dr. Céline Lévesque and
Dr. Howard Tenenbaum for their countless hours of scheduled and impromptu meetings, endless
email communication, thesis editing and research tangents, turning “lemon juice” into this thesis
and above all else, believing in me.
Thanks to my committee members, Dr. Michael Sigal Dr. Iona Leong, and the late Dr. David
Locker for their tough questions and support in the creative and technical aspects of the work
presented herein.
To the Levesque Lab: Steph, Delphine and Vince. Thank you for helping me with lab “stuff” and
tolerating my “bugs” in your space. To Adnan, thank you for helping me kill some yeast, your
help was very appreciated. Dr. Victor Charles, and Jian Wang, thanks for the stats and SEM.
To my mom, Marilyn, Thank you for pushing me to do more. I know I have made you proud.
To my late father, Marvin, (1947-2001), you taught me to persist, ask questions and make people
laugh (with good or bad jokes). I hope I have made you proud.
To Ilana, my wife, my whose love, support and tolerance of me going “back to
school” for another 3 years made this possible. I could not have done any of this without you.
No more school, well…maybe a part time PhD.
To my children, Mikayla Emily, Sydney Nicole and Hudson Meyer, follow your dreams and
your heart to your goals. Live your lives to the fullest and take every opportunity to get there. All
you need to do is try. Don’t give up; you have it within you to succeed.

iv
Table of Contents
Abstract …………………………………………………………………………………………. ii
Acknowledgments ........................................................................................................................ iii
Table of Contents ......................................................................................................................... iv
List of Abbreviations .................................................................................................................. vii
List of Tables .............................................................................................................................. viii
List of Figures............................................................................................................................... ix
1. Introduction ........................................................................................................................... 1
2. Review of the literature ......................................................................................................... 2
2.1 Candida albicans ............................................................................................................................2 2.2 A polymorphic species ..................................................................................................................2 2.3 Candida albicans plasma membrane and cell wall .....................................................................3
2.3.1 Plasma membrane ....................................................................................................................4 2.3.2 Cell wall structure ....................................................................................................................4 2.3.3 Components of cell wall...........................................................................................................5
2.3.3.1 Cell wall proteins .............................................................................................................................5 2.3.3.2 Specific CWPs..................................................................................................................................5
2.3.3.2.1 Adhesins ...................................................................................................................................5 2.3.3.2.2 Secreted aspartyl proteinase (SAPs).........................................................................................5 2.3.3.2.3 Phospholipases .........................................................................................................................6
2.3.3.3 Cell wall carbohydrates- Chitin........................................................................................................6 2.3.3.4 Cell wall carbohydrates- ß-glucans ..................................................................................................6 2.3.3.5 Cell wall carbohydrates- Mannans ...................................................................................................7
2.3.4 Immune response to the components of the cell wall ..............................................................7 2.4 Biofilms of Candida .......................................................................................................................7
2.4.1 Persister cells............................................................................................................................8 2.4.2 Quorum-sensing .......................................................................................................................9 2.4.3 Biofilm resistance...................................................................................................................10
2.5 Commensalism vs. virulence ......................................................................................................11 2.5.1 Host-fungal interactions .........................................................................................................12
2.5.1.1 Host ligands....................................................................................................................................12 2.5.2 Candidal infections- oral and esophageal ..............................................................................12

v
2.5.2.1 Candida in periodontal disease, caries, and endodontic infections................................................13 2.6 Treatment protocols....................................................................................................................14
2.6.1 Current treatments ..................................................................................................................14 2.6.2 Antimycotic drugs and mechanisms of action .......................................................................15
2.6.2.1 Polyenes .........................................................................................................................................15 2.6.2.1.1 Nystatin ..................................................................................................................................15
2.6.2.2 Azoles.............................................................................................................................................16 2.6.2.3 Pyrimidine analogues .....................................................................................................................16 2.6.2.4 Echinocandins ................................................................................................................................16 2.6.2.5 Allylamines ....................................................................................................................................17
2.6.3 C. albicans methods of drug resistance to antimycotics ........................................................17 2.6.4 Resistance to polyenes ...........................................................................................................18 2.6.5 Resistance to azoles................................................................................................................18 2.6.6 Resistance to pyrimidine analogues .......................................................................................19 2.6.7 Resistance to echinocandins...................................................................................................19 2.6.8 Resistance to allylamines .......................................................................................................19 2.6.9 Combination therapy ..............................................................................................................19
2.7 Plant extracts ...............................................................................................................................20 2.7.1 Essential oils...........................................................................................................................20 2.7.2 Polyphenols ............................................................................................................................20
2.7.2.1 Flavonoids ......................................................................................................................................21 2.8 Polyphenol activity ......................................................................................................................21
2.8.1 Antimicrobial properties ........................................................................................................21 2.8.2 Protective properties...............................................................................................................22
2.8.2.1 Antioxidant.....................................................................................................................................22 2.9 Biosecur c320c .............................................................................................................................23
3. Statement of the problem.................................................................................................... 24
3.1 Objective ......................................................................................................................................24 3.2 Aims ..............................................................................................................................................24 3.3 Rationale ......................................................................................................................................24 3.4 Hypothesis ....................................................................................................................................24
4. Materials and Methods........................................................................................................ 25
4.1 Fungal cultures and growth conditions.....................................................................................25 4.2 Test compounds ...........................................................................................................................25 4.3 Biofilm growth .............................................................................................................................25

vi
4.4 Determination of the minimum inhibitory concentration .......................................................26 4.5 Treatment of biofilm with test compounds ...............................................................................26 4.6 Scanning electron microscopy....................................................................................................26 4.7 Resistance assay...........................................................................................................................27 4.8 pH assay .......................................................................................................................................27 4.9 Drug synergy................................................................................................................................28 4.10 Statistical analysis .....................................................................................................................28
5. Results................................................................................................................................... 29
5.1 Determination of CFU from stationary phase cultures ...........................................................29 5.2 Antimicrobial susceptibility to B320 and nystatin ...................................................................29 5.3 Antimycotic activity of B320 ......................................................................................................29 5.4 Combined application of B320 and nystatin.............................................................................30 5.5 Determination of synergistic antimycotic activity....................................................................30 5.6 Resistance assay...........................................................................................................................31 5.7 SEM analysis................................................................................................................................31
6. Discussion ............................................................................................................................. 40
6.1 B320 as an antifungal..................................................................................................................40 6.2 Effects of pH on Activity of B320...............................................................................................41 6.3 MIC of B320.................................................................................................................................41 6.4 Effects of combinations of B320 and nystatin on C. albicans ..................................................42 6.5 Development of resistance to B320 and nystatin ......................................................................43 6.6 Cell morphology of C. albicans with and without treatment ..................................................44 6.7 Possible sources of error .............................................................................................................44 6.8 Summary ......................................................................................................................................46
7. Conclusions and future studies........................................................................................... 47
References .................................................................................................................................... 48

vii
List of Abbreviations
ATCC American Type Culture Collection
B320 Biosecur c320c ®
CFU Colony forming units
CWP Cell wall protein
FIC Fractional inhibitory concentration
FICI FIC index
GPI glycosyl-phosphatidyl-inositol
HCD High cell density
LCD Low cell density
NYS Nystatin
NYSR Nystatin resistant
SDA Sabouraud Dextrose Agar
SEM Scanning electron microscopy
WT Wild-type
YPG Yeast-Peptone-Glucose

viii
List of Tables
Table 1. Clinical forms of oral candidiasis, description and location ……….……………… 13
Table 2. Antifungal drugs and biological targets ………………………….………………… 14
Table 3. MIC of tested drugs against C. albicans isolates cultivated planktonically …........... 32
Table 4. Fractional inhibitory concentrations (FIC) and FIC indices (FICI)………….….….. 32
Table 5. Development of drug resistance……………………………………………..……… 33

ix
List of Figures
Figure 1. Morphological forms of C. albicans……………………………………………….. 2
Figure 2. Cell of C. albicans with its major components…………………………………….. 4
Figure 3. Backbone of a polyphenol molecule………………………………………………. 21
Figure 4. Percentage of cell survival of 24-h-old C. albicans WT biofilms following treatment
with B320 (0.5, 1, 10%), nystatin (100,000 IU/ml), or glycerin (control) for 60 s……….… 34
Figure 5. Percentage of cell survival of 24-h-old C. albicans WT biofilms following treatment
with 10% B320 (pH 2.8; non-pH-adjusted), 10% B320pH (pH 6.8; pH adjusted with NaOH), and
carrierpH solution (pH 2.8; adjusted with HCl) for 60 s………………………………………. 35
Figure 6. Percentage of cell survival of 6-h-, 24-h-, and 72-h-old C. albicans WT biofilms
following treatment with 10% B320, nystatin 100,000 IU/ml, or 10% B320 with 90,000 IU/ml
nystatin for 60 s……………………………………………………………………………….. 36
Figure 7. Percentage of cell survival of 6-h-, 24-h-, and 72-h-old C. albicans NYSR biofilms
following treatment with 10% B320, nystatin 100,000 IU/ml, or 10% B320 with 90,000 IU/ml
nystatin for 60 s……………………………………………………………………………….. 37
Figure 8. SEM of 24-h-old C. albicans WT biofilms exposed to B320 (10%), nystatin (100,000
IU/ml), or B320 (10%) combined with nystatin (90,000 IU/ml) for 60 s……………………. 38
Figure 9. SEM of 24-h-old C. albicans NYSR biofilms exposed to B320 (10%), nystatin
(100,000 IU/ml), or B320 (10%) combined with nystatin (90,000 IU/ml) for 60 s………….. 39

1
1. Introduction Antibiotics are one of the greatest discoveries of the early twentieth century medicine. Before
their accidental discovery, millions of people died from otherwise simple bacterial infections.
However, through overuse and misuse of these agents resistant microorganisms have been
created inadvertently. A recent study showed that tertiary care hospitals are using antimicrobial
and antifungals agents inappropriately (1). This growing trend is an ever-increasing threat to the
health of humans who are now at risk of dying from previously treatable infections.
Unfortunately, drug resistance (alteration of the drug target site, drug modification or restricted
drug penetration) develops at a pace faster than that of new drug development. This phenomenon
is also occurring in the pathogenic microorganism Candida albicans, an otherwise commensal
species found in and on the human body. Current antifungal agents are less effective against
C. albicans now than when they were first introduced because Candida species have developed
resistance to these agents. Additionally, research by the major pharmaceutical companies in the
development of antibiotic agents has been on the decline due to low return on investment. A
paper from 2005 indicated, “…Bristol-Myers Squibb, Lilly, and Wyeth discontinued all anti-
infective discovery in 2003, while GlaxoSmithKline, Abbott, and Aventis are in the process of
down-sizing their programs. Only a handful of companies…are continuing antimycotic drug
discovery” (2).
Innovation has led to the rediscovery of a group of naturally occurring agents with antimicrobial
properties. Plant derived extracts (i.e. polyphenols and essential oils) show promise as antifungal
agents in the ongoing battle between humans and microorganisms. The modern use of plant-
derived extracts as antimicrobials is in its infancy but these agents possess properties that make
them ideal in the fight against pathogenic microorganisms.

2
2. Review of the literature
2.1 Candida albicans
Candida albicans is one of 200 organisms of the genus Candida (3-5). Its genome was
sequenced in 2004 and a database is available for online searches (www.candidagenome.org) (6).
In general, this organism lives as a benign commensal entity in a variety of locations in the
human host, most notably the oral cavity, epidermis, genitalia and gastro-intestinal tract. This
species comprises 75% of the fungal species sampled from the oral cavity (7) but makes up only
a minor part of the total oral microflora. C. albicans can be isolated as a commensal microbial
organism from the oral cavity in 30% to 90% of healthy adults but in most cases, those colonized
do not show signs or symptoms of infection (8-13). The possible reasons for this range include:
detection method (swab, smear, denture impression) and population tested (immune status, age).
2.2 A polymorphic species
C. albicans grows (Fig. 1) in several ways (14, 15). It can form blastospores (daughter cells)
through budding. Pseudohyphae form when the budding cells grow by elongation and do not
detach. The hyphae (mycelia form) are the germ tubes that form as a result of filamentous
growth. Both cell forms appear elongated but hyphal cells have no septal constrictions (14).
Under suboptimal conditions C. albicans can also form chlamydospores, round spores with thick
walls (16).
Figure 1. Morphological forms of C. albicans.

3
In relation to the virulence of C. albicans, an important feature is its ability to switch between the
various morphological forms noted above (17, 18). It is also important to understand that the
yeast and hyphal forms are essential for infection (15). The cell wall proteins of the yeast form
covalently linked to the skeletal cell wall polysaccharides of C. albicans, play an important role
in the virulence of C. albicans. They contribute to cell wall integrity and to the formation of
biofilm, and also mediate adherence to host cells, commensal microbial flora, and the surfaces of
medical devices (e.g., prosthesis made from denture acrylic or indwelling catheters) (7, 19). The
hyphal cells express and exhibit a number of factors/receptors allowing C. albicans to adhere to
and penetrate the epithelium (17) of the host. The C. albicans hyphal form also secretes enzymes
including aspartyl proteases, phospholipases, and lipases (7, 19). Taken together, these factors
promote invasion of epithelial layers and offer protection against attacks by the innate immune
system further contributing to the virulence of C. albicans. Some of these factors, which are
present on the external surface of the cell, are described below.
2.3 Candida albicans plasma membrane and cell wall
The plasma membrane is selectively permeable to the molecules required by C. albicans for
survival (Fig. 2). It contains free fatty acids, glycolipids, phospholipids, sphingolipids, and
sterols. The fungal cell wall acts as an exo-cytoskeleton and is composed of 90% carbohydrate
and 10% protein (20, 21). The polysaccharides are present in three different but structurally
significant forms including mannans, ß-glucans and chitin. Based on dry weight analysis, the cell
wall consists of roughly: 40% manno-proteins, 40% ß-1, 3 glycans, 20% ß-1, 6 glycans and
1-2% chitin. The composition of the cell wall varies in polysaccharide content between the yeast
and hyphal forms (more chitin in the hyphal form), and is also affected by environmental factors
including presence of antifungal agents (21, 22).

4
Figure 2. Cell of C. albicans with its major components.
2.3.1 Plasma membrane
The plasma membrane of C. albicans is comprised of a lipid bilayer similar to eukaryotic cells in
higher animals with the main difference being that instead of cholesterol being present, fungal
cell membranes contain primarily ergosterol. Ergosterol is found in both the yeast and hyphal
forms of C. albicans. As in higher animals, sterols modulate the fluidity of the membrane (23).
This sterol difference provides a useful target (i.e. ergosterol synthesis) in the development of
antifungal agents. However, not all ergosterol targeting anti-fungal agents retain their
antimycotic properties, but this will be discussed later. The lipid content of the plasma membrane
can be altered by changes in host substrate, biofilm age, and by changes in pH and in oxygen
tension, presence of antifungal agents and morphological form (23).
2.3.2 Cell wall structure
The cell wall has two distinct layers; the inner and outer cell wall (21, 22). The outer wall is
composed of mannan and cell wall proteins and is approximately 180 nm thick. The inner wall is
comprised of ß-1,6 glucans, ß-1,3 glucans and chitin, and is approximately 100 nm thick. The
organization of the cell wall from the cell membrane outward starts with the chitin layer
followed by a network of ß-1,3 glucan molecules held together by hydrogen bonds. ß-1,6 glucan
is present as a side chain linked to the ß-1,3 glucan layer. Interspersed within the ß-1,3 glucan
layer are cell wall proteins (CWPs) that are either bound to chitin or to the ß-1,3 glucan layer via
a glycosyl-phosphatidyl-inositol (GPI) anchors. The mannans are attached to the terminal ends of
the CWPs via covalent linkage.

5
2.3.3 Components of cell wall
2.3.3.1 Cell wall proteins
CWPs are highly glycosylated and have negatively charged phosphate groups in their
carbohydrate side chains (22). The predominant function of the CWPs is thought to be related to
limitation in permeability of the cell wall, protection against degrading enzymes (chitinases and
glucanases) and antifungal substances; determination of hydrophobicity and the antigenicity of
the cell wall. The CWPs are also thought to contribute to remodeling of the cell wall as well as to
cellular adhesion, which would also affect the organism’s virulence (21). There are three classes
of CWPs (24). This first class includes GPI proteins (e.g. adhesins) and are linked to ß-1,6
glucans. The second set of proteins are referred to as ‘proteins with internal repeats’ (PIR) which
are essential for maintaining the architecture of the cell wall (22, 24). The third class of proteins
are those secreted into the extracellular space including hydrolytic enzymes like phospholipases
and proteases (24).
2.3.3.2 Specific CWPs
2.3.3.2.1 Adhesins
The agglutinin-like sequence (ALS) glycoprotein of C. albicans is one protein responsible for
adherence. Eight ALS genes (ALS1-7, 9) have been sequenced in C. albicans and each encodes a
slightly different cell surface glycoproteins (24). ALS1-3 and 9 are almost always expressed in
the C. albicans strains of those infected with candidiasis. The ALS GPI-CWP of C. albicans
allows for adhesion to buccal and vaginal epithelial cells, fibronectin, laminin and to other yeast
cells. Deletion of ALS1 and ALS3 genes reduces binding of C. albicans mutants to epithelial
cells by 20-35% and 60% respectively. Additionally, ALS1/ALS3 mutants lose the ability to
form mature biofilms (24). Hwp1p and Ywp1p are other adhesion proteins expressed by hyphae
in biofilms. The difference is that Hwp1p promotes biofilm retention while Ywp1p promotes
biofilm dissolution (24).
2.3.3.2.2 Secreted aspartyl proteinase (SAPs)
Ten SAP genes have been identified in C. albicans. These genes encode proteases that are
secreted into the extracellular space. These factors are essential for virulence as their functions
include degradation of the host’s extracellular matrix, keratin, mucin, stratum corneum and

6
proteinase inhibitors. Additionally, they also degrade immune response factors including
immunoglobulin G (IgG) and can activate the pro-inflammatory cytokine interleukin-1ß (IL-1ß)
(24). The expression of specific SAP genes is regulated by temperature (SAP8) and early biofilm
development (SAP2). Specific SAPs (SAP 4/6) are expressed primarily when the yeast grows in
hyphal form (24).
2.3.3.2.3 Phospholipases
Phospholipase enzymes can also be anchored to GPI or secreted into the extracellular space.
There are five phospholipase genes that encode for enzymes that may be present in the yeast or
hyphal forms. These enzymes localize to the hyphal tips or growing end of the yeast form. The
expression of phospholipase genes is site dependent, however phospholipase B1 (PLB1) is found
more frequently in those with candidiasis (24).
2.3.3.3 Cell wall carbohydrates – Chitin
Chitin is a homopolymer of ß-1,4-N-acetyglucosamine (GlcNAc) and is synthesized by one of
four chitin synthases. Although it constitutes only a minor component of the inner cell wall,
chitin is essential for the formation of hyphae, pseudohyphae, septum and hyphal growth. Null
mutants for the gene encoding chitin synthase 1 form long chains without septa and eventually
lyse (25). This polysaccharide is located close to the cell membrane and this likely makes it a
poor target for recognition by the host’s innate immune system. However, it is often exposed on
the cell surface at the location of a yeast bud scar. If exposed to the human host, chitin can
induce macrophages to produce inflammatory mediators (26). As such it may play some role in
pathogenicity (27).
2.3.3.4 Cell wall carbohydrates – ß-glucans
Glucans are a class of glucose polymer of D-glucose that are linked by ß-glycosidic bonds (i.e.
atoms are in the same plane). Chains of this polymer can be short or long and can be branched or
unbranched. The glucan polymers present in C. albicans include the ß-(1,3) D-glucopyranosyl
polymers with varying amounts of ß-(1,6) D-glucopyranosyl side chains (28). This ß-glycan
linkage makes the cell wall insoluble as there are no ß-glucan hydrolases in humans (29).

7
2.3.3.5 Cell wall carbohydrates – Mannans
On the outer surface of the cell wall, mannans are present in the form of glycoproteins. The
mannan chain consists of 1,6-linked mannose residues comprised of highly substituted and short
oligomannose side chains with 1,2 and 1,3 linkages (30). Using this glycoprotein, taxonomists
divide C. albicans into Serotype A (longer chains) and Serotype B (shorter chains) strains.
Mannoproteins act as virulence factors with immunosuppressive activity (31). Mannan can
activate or depress cell-mediated NK-cells and phagocytic cells and humoral immune functions
(16); oligosaccharide fragments of mannan appear to be potent inhibitors of cell-mediated
immunity. For a more detailed review please refer to reference (31).
2.3.4 Immune response to the components of the cell wall
The immunological response to any pathogen is a multifaceted event, however, recent findings
have shown that there are several specific elements associated with the direct human response to
the cell wall of C. albicans. One such example comprises the ß-glucan receptor of Dectin.
Dectin 1 and Dectin 2 are type II transmembrane proteins of the C-type lectin family with a
single carbohydrate recognition domain in their extracellular region (32). Dectin-1 recognizes
ß-glucans and can be found on the surface of dendritic cells/macrophages. It has a signaling
motif which interacts with the immunoreceptor tyrosine-based activation motif (ITAM). Dectin-
2 recognizes α-mannans and can be found on the surface of Langerhans cells (32, 33). This
receptor lacks a signaling motif but links with Fc receptor g (FcRg) which has an ITAM motif.
Upon Dectin-ligand binding, spleen thyrosine kinase (syk) is recruited to ITAM. This in turn up
regulates production of cytokines and reactive oxygen species (ROS) which in turn mediate the
production of IL-12, IL-23 and IL-1ß. These cytokines mediate the activation of netrophils and
macrophages through the activation of T helper cells (32). Other receptor pathways, like Toll-
like receptor 2 and Toll-like receptor 4 have also been implicated in cascade reactions to ß-
glucans and mannans, respectively (27).
2.4 Biofilms of Candida
It is estimated that 65% of all human infections are related to the formation of biofilms by the
causative or pathogenic microbes (34) and importantly, up to 40% of the adult population are
carriers of C. albicans in the oral cavity (35). With respect to C. albicans, formation of a biofilm

8
begins with adhesion of the microbe to a suitable surface (biotic or abiotic) (36). Adhesion
occurs via non-specific factors (cell surface hydrophobicity/electrostatic forces) and specific
factors (cell surface receptors that recognize serum proteins/fibrinogen). These specific factors
promote retention of the organism in the oral cavity or elsewhere in or on the body
(esophagus/vagina). Additionally, C. albicans can adhere to other microbial biofilms that are
already present (e.g. dental plaque) (37). In the oral cavity, C. albicans has been found to
coaggregate with other oral pathogens, such as Actinomyces, Streptococcus, and Fusobacterium
(35). The formation of hyphae usually occurs 3-6-hours following the yeast cells initial
colonization (38). Adhesion is followed by colonization, proliferation, and maturation of the
biofilm structure. The adherent yeast cells form a basal layer that attaches firmly to the biofilm
substrate (i.e. palatal epithelial cells, denture acrylic, etc.) while subsequent germination
generates the bulk of the newly developing biofilm. After 48 hours, a mature biofilm typically
contains cells with hyphae and pseudohyphae (38). The mature biofilm is a well-organized and
spatially structured complex surrounded by an extracellular polysaccharide matrix (34, 37, 39).
The extracellular matrix is composed of proteins, hexosamine, as well as phosphorus and uronic
acid (39). The last step, detachment of cells, is followed by colonization of distant sites and can
be triggered by a process known as quorum-sensing (34, 37, 39). Quorum-sensing or cell-cell
signaling benefits the biofilm by preventing over population and by controlling competition (e.g.
when faced with limited nutrients). Two molecules used for quorum sensing by C. albicans
include farnesol and tyrosol, which are described below (37).
2.4.1 Persister cells
Persister cells are a subpopulation of regular cells and form sporadically in microbial biofilms.
C. albicans persister cells are tolerant to several antifungal drugs, whether they are used alone or
in combination (40). Even strains of yeast that are generally considered to be highly susceptible
to antimycotic agents are known to produce drug tolerant persister cells (41). Persister cells make
up 0.2% to 9% of the biofilm in individuals who are infected chronically with C. albicans, but
are undetectable in those individuals who have become infected only transiently (40).
Interestingly the carriage rate of C. albicans may be related to the presence and/or prevalence of
persister cells. It has been demonstrated in vivo with concurrent treatment, those with low levels
of persister cells (0.9%), C. albicans cells can be isolated for an average of 2.8 weeks. However,
those with higher levels of persister cells (3.7%), C. albicans cells can be isolated for 15.4 weeks

9
(41). This means that current 2-week treatment protocols for oral candidiasis may not be
sufficient, particularly considering the presence of persister cells. However, there is no evidence
that persister cells lead to re-infection (41).
2.4.2 Quorum-sensing
Quorum-sensing is a mechanism of cell-cell communication by which microbes detect their
population density through diffusible signal molecules. Microbes use quorum-sensing to regulate
their gene expression in response to fluctuations in the cell population density (42, 43).
C. albicans produces two primary alcohol quorum-sensing molecules, farnesol and tyrosol.
Farnesol is a sesquiterpine molecule, a small organic hydrocarbon molecule; and can be cyclic or
acyclic, saturated or unsaturated. Farnesol is commonly found in plants and in citrus fruits.
Tyrosol is derived from the aromatic amino acid tyrosine. The quorum-sensing molecules
influence the morphological transition between the yeast and filamentous forms in response to
environmental cues (44). Tyrosol acts to increase the rate of biofilm development and enhances
the formation of hyphae within the biofilm (44). One of the earliest findings regarding the
function of farnesol was that it prevents the development of candidal hyphae but did not affect
existing hyphal growth. Additionally, this molecule has been observed to prevent the initial
attachment of yeast cells to a target surface thereby regulating the initial formation of biofilms.
In a mature biofilm, low levels of farnesol has been observed to promote cell dispersal (42).
Farnesol has also been shown to interact with the ATPase binding cassette (ABC) transporter
family, specifically Candida drug resistance pumps (43). The proposed mechanism is related to
the level of farnesol. At high levels of farnesol, a cascade is observed such that ROS are
accumulated within the mitochondria, leading to an increased caspase activity and ultimately
leads to apoptosis in C. albicans. This cascade leads to a reduction of yeast cells within the
biofilm. At low levels of farnesol, no interaction is observed between the quorum sensing
molecule and the ATPase pumps. Interestingly, when low levels of farnesol and low levels of an
antifungal agent (fluconazole, amphotericin B or ketoconazole) are combined, apoptosis is
observed (43). Curcumin is a polyphenol found in tumeric root and competes for drug binding
sites on the ABC transporter. The observed effect of curcumin on Candida is similar to that of
farnesol in that on exposure to high levels, apoptosis or programmed cell death occurs (45)
suggesting a connection to the ABC transporter system. And, just like farnesol, when curcumin

10
is combined with an azole or polyene antifungal, synergy is observed and apoptosis occurs via
generation of ROS (43, 45). The importance of this finding will be discussed later. (See Section
2.6.9 for combination therapy).
2.4.3 Biofilm resistance
There is no single factor that leads to the development of resistance of candidal biofilms to
various antimycotic agents. It has been postulated that one of the mechanisms related to the
increased antimicrobial resistance demonstrated in biofilms might be related to physical or ionic
sequestration of drugs by these structures (37). In this regard it has been suggested that the
matrix can act as a physical barrier to the drugs by preventing penetration of the charged
antimycotic agents leading to their inactivation/sequestration. This latter concept has not stood
up to experimental testing but nonetheless biofilms do appear to reduce the rate of diffusion of
drugs towards susceptible cells (46). A decrease in the basal growth rate (i.e. cellular metabolic
activities) occurring during biofilm development may represent an innate mechanism of
resistance (47). C. albicans has several antimicrobial efflux pumps that take part in
antimicrobial resistance. However, when these specific efflux pump genes are deleted from
certain antifungal resistant strains of C. albicans, their resistance pattern is not altered (48).
Alterations in the fluidity of the plasma membrane is another resistance mechanism. As the
biofilm matures, the ergosterol content of the C. albicans cells is reduced compared to planktonic
cells. Since sterol metabolism is a target of many antifungal agents, a decrease in this molecule
could lead to the observed drug resistance (48). Candida cells within biofilms also up regulate
several classes of genes associated with drug resistance. The application of amphotericin B,
ketoconazole, or caspofungin on C. albicans biofilms in vitro up regulated genes for CWP
(ALS3 and HWP1), for hyphal growth (ALS1) and down regulated other non-cell wall specific
genes (49). In planktonic Candida cells, these same genes are either down regulated or non-
responsive to the same antifungal agents (49, 50).
Another mechanism of host resistance involves a change from yeast form to filamentous form.
The development of hyphae impairs phagocytosis. The hyphae produce more SAPs which
impairs IgA function, and hyphae have more complement binding factors leading to antigen
masking (39). The hyphal form can also invade the host via penetration through non-intact
epithelial surface and the production of phospholipase/SAPs damage host cells (39). Thus the

11
mechanisms contributing to drug resistance include: high cell density, biofilm matrix effects,
decreased growth rate, limitation in the diffusion of antimicrobial molecules as well as
expression of antimicrobial resistance genes. A multitude of factors are responsible for the
observed candidal resistance making this a complex phenomenon. In addition to characteristics
associated with the pathogen that affect virulence and resistance, several host factors also
increase the virulence of C. albicans. These include the use of medications (antibiotics, steroid
inhalers, xerostomia causing agents), extremes of age, endocrine disorders, immunosuppression
(pathological or drug-induced). Research on the above mechanisms are based on in vitro studies,
but current in vivo models of biofilms have confirmed some of the in vitro findings (37).
2.5 Commensalism vs. virulence
In recent decades, considerable interest has developed in relation to opportunistic candidal
infections due to an increase in the size of the population of individuals who are
immunocompromised (e.g. those with active HIV infections or fully developed AIDS, patients
undergoing immunosuppressive treatments or patients with cancer and undergoing chemotherapy
and/or radiotherapy). Various investigations have shown that 30 to 90% of the general
population are C. albicans carriers and yet, are often asymptomatic. Institutionalized patients
have a higher infection rate (51) which may be related to some form of removable denture (52).
As well, many of these patients also take several medications that cause xerostomia (53).
Systemic conditions (immune or metabolic) with associated inflammation can also predispose
these individuals to develop candidiasis (7, 12, 51, 54). Successful treatment of oral candidiasis
is difficult because C. albicans infection is primarily the consequence of various host factors.
Thus, following standard treatment, reinfection can develop due to the persistent host conditions
(8, 11, 55, 56).
Diagnosis involves performing an inspection of the oral cavity (corner of the lips, inner cheek).
If a clinician is unsure of the diagnosis, there are a few ways to obtain a sample of the oral
microflora for assessment. Saliva can be collected, or a smear sample taken from the epithelial
surfaces/abiotic surfaces (denture) for subsequent microscopic assessment following staining of
the sample with a fungus specific stain (Periodic Acid Schiff) (57, 58). However the detection of
candidal spores or hyphae cannot differentiate those that are carriers vs. those that are truly
infected without a clinical examination.

12
2.5.1 Host-fungal interactions
2.5.1.1 Host ligands
Several mechanisms play a role in the adherence of C. albicans yeast or hyphae to surfaces in the
oral cavity. These include glycoprotein components of the host’s extracellular matrix (ECM)
including laminin, fibronectin, collagen, entactin, vitronectin, cadherin, and tenascin. C. albicans
uses surface proteins to adhere to immune response factors present in the oral cavity. The
complement regulator acquiring surface proteins in the cell wall allows adherence of C. albicans
to various elements of the alternative and classic complement pathway, including c3b (24, 59,
60). This process leads to their inactivation. Additionally, the cell wall can also bind
plasminogen, the precursor to plasmin, an enzyme that cleaves fibrinogen and degrades ECM
proteins. This host derived enzyme can then be used by yeast cells to invade the ECM of the host
(59-61).
2.5.2 Candida infections – oral and esophageal
Oral candidiasis develops when certain (often multi-factorial) conditions arise including:
pregnancy, drug administration (antibiotic/steroid usage), uncontrolled diabetes and/or
compromised immunity (3, 4). The first line of defense against candidiasis is the presence of an
intact mucosal epithelial surface. Once overgrowth of C. albicans occurs, it becomes a local
irritant that can produce an acute inflammatory reaction causing local edema and the formation
of micro-abscesses (3, 4), which can put the host at risk for the loss of integrity of the underlying
epithelial surface. In fact, the critical event for the development of candidiasis occurs when the
microorganism actually penetrates the mucosal surface (3, 4). This is often observed in long
standing infections and in these instances it is the hyphal form of C. albicans that is observed on
cytological smears. Ill-fitting or poorly maintained maxillary dentures provide a good
environment for infection, particularly in areas of traumatically injured palatal surface in the
oropharynx (62). Once the epithelial surface has been traumatized, C. albicans can penetrate the
mucosal barrier easily and bypass the natural defense mechanism, leading to the initiation of
candidiasis (3, 4). Patients with oral candidiasis report sensations of oral burning, difficulty
swallowing, bad breath, pain at the corners of the lips and generalized or localized mucosal
soreness (with or without a maxillary denture). Due to variation in clinical presentation, oral
candidiasis has been classified as follows (3, 8-10, 63, 64): 1) Acute forms: pseudomembranous
13
(thrush) and erythematous; 2) Chronic forms: hyperplastic (nodular/plaque-like), erythematous,
and pseudomembranous; and 3) Candida-associated lesions: denture stomatitis, angular cheilitis,
median rhomboid glossitis, and linear gingival erythema. Erythematous candidiasis is generally
thought to describe denture stomatitis (chronic or acute), as there appears to be a correlation
between traumatization of the palatal surface by the prosthesis surface and subsequent
development of candidal infection (10). Angular cheilitis occurs at the corners of the lips and is
often associated with an over-closed mouth. This finding is often related to a loss of vertical
facial height leading to accentuation of the commissures of the lips, thereby providing the ideal
niche for C. albicans. Please refer to Table 1.
Table 1. Clinical forms of oral candidiasis, description and location (adapted from (65)).
Clinical type Pseudo-
Membranous
(thrush)
Erythematous Median
rhomboid
glossitis
Angular
cheilitis
Denture
stomatitis
Hyperplastic
Appearance Creamy white
removable
plaques,
Red areas Red mucosa Red
fissured
lesions
Red on denture
bearing area
White, non-
removable
plaques
Symptoms Burning
sensation and
bad taste
Burning
sensation
Asymptomatic Sore Asymptomatic Asymptomatic
Common
Sites
Buccal
mucosa,
tongue, palate
Hard palate,
buccal mucosa,
tongue
Midline of
tongue
Corners
of lips
Opposing the
denture surface
Buccal mucosa
2.5.2.1 Candida in periodontal disease, caries, and endodontic infections
In addition to adhesins that mediate the binding to buccal epithelial cells, C. albicans can bind
oral and non-oral bacteria. These interactions appear to be effected through multiple
mechanisms. Aggregation or agglutination of C. albicans has been observed with strains of
Streptococcus sanguinis, Streptococcus salivarius, Streptococcus mutans, Streptococcus mitis,
Fusobacterium nucleatum, Actinomyces viscosus, Lactobacillus amylovorus, and
Porphyromonas gingivalis (16). Reports demonstrate both association of C. albicans with caries,

14
periodontitis and endodontic infections while others do not (caries (66, 67); periodontitis (68, 69)
endodontics (70, 71)). The evidence provides controversial results and is beyond the scope of
this thesis and will not be discussed further.
2.6 Treatment protocols
2.6.1 Current treatments
Several antimycotic agents are used for the treatment of oral candidiasis (72). Specific
C. albicans targets have been taken advantage of in the development of these agents and
generally include the sterols, the carbohydrates in the cell wall, and the nucleic acids (Table 2).
Table 2. Antifungal drugs and biological targets.
Pharmacological agent Target
Azoles, Allylamines, Polyenes Sterols
Echinocandins Cell wall
5- Flucytosine DNA
Compliance (11, 51, 73), cost (13) and in particular, side effects (7, 9, 12, 55, 74) have been
shown to be important obstacles when focusing on treatment or prevention of oral candidiasis in
the population at risk. For instance, it is known that the polyene amphotericin B can cause
nephrotoxicity, especially at higher doses (75, 76). Azoles target yeast cytochrome P450 and the
co-administration of other cytochrome P450 medications (e.g. warfarin and acetaminophen),
could elevate or decrease the blood levels of either drug thus making them either toxic or
insufficient to mediate a clinical effect (77). The echinocandins can only be administered
intravenously and have minimal side effects (78).

15
2.6.2 Antimycotic drugs and mechanisms of action
The classes of antimycotic drugs include those that are either fungicidal or fungistatic. The
fungistatic agents include: 1) fluorinated pyrimidine analogues (e.g. 5-fluorocytosine) that inhibit
DNA synthesis; 2) azoles (e.g. ketoconazole) which alter sterol synthesis. The fungicidal agents
include: 1) polyenes (e.g. nystatin/amphotericin B) that create leaks in the lipid membrane; 2)
echinocandins (e.g. caspofungin), which alter cell wall synthesis; 3) allylamines (e.g.
terbinafine), which alter sterol synthesis. The mechanism of action of these agents is described
below.
2.6.2.1 Polyenes
Polyene antifungal agents were the standard of therapy for systemic fungal infections prior to the
development of azoles. Polyenes interact with fungal membrane sterols resulting in the
production of aqueous pores in the membrane. All organisms that contain ergosterol in their cell
membranes are susceptible to polyenes (e.g. yeast, algae, and protozoa) (79). The development
of these aqueous pores in the cell membranes leads to alterations in membrane permeability,
which causes several problems including leakage of ions, a property that leads to cell death. Of
two well-known polyenes, amphotericin B is more effective at killing fungal organisms than
nystatin. However, the former has a narrow therapeutic window, which limits its usefulness in
the clinic. (79).
2.6.2.1.1 Nystatin
Nystatin (originally called fungistatin) is a polyene. It was discovered in New York State (and
renamed to identify the place of discovery “NYSTATin”) by a biochemist (Brown) and
microbiologist (Hazen) in 1949. The company E.R. Squibb (currently Bristol-Myers Squibb)
developed the trade name Mycostatin in 1954 (80). This agent was isolated from the organism
Streptomyces noursei. Nystatin is the prototypical polyene (a class of natural compounds with an
amphipathic nature) and one of the first antimycotic agents to be discovered (72). Nystatin
targets ergosterol within the fungal membrane (as noted above, ergosterols confer fluidity to the
membrane) and creates pores that allow potassium ions to diffuse across the membrane, causing
cell death (81).

16
Nystatin binds to the fungal membrane ergosterol with up to 10 times more affinity than to the
human membrane sterols (i.e. cholesterol) providing selective binding and thus antifungal
activity (74) with minimal effects if any on host cells. Therefore, nystatin can be used at a dose
that is low enough to be bound to candidal cell membranes but not to mammalian cells, making
this agent non-toxic to human while still retaining its antifungal activity. In suspension, nystatin
(especially the form for oral use) is not absorbed in the gut and therefore has no systemic effects
or side effects.
2.6.2.2 Azoles
The target of azole based antimycotic drugs (e.g. ketoconazole and fluconazole) is the fungal
enzyme P450 cytochrome lanosterol 14α-demethylase (79). The function of this enzyme is the
demethylation of the ergosterol precursor at the C4 position. Inhibition of this enzyme leads to
depletion of ergosterol in the cell membrane and accumulation of sterol precursors, resulting in
the formation of a plasma membrane with altered structure and function (79).
2.6.2.3 Pyrimidine analogues
Flucytosin or 5-Flurocytosine (5-FC) is a synthetic antimycotic compound and is administered
intravenously for systemic candidal infections. 5-FC has no intrinsic antifungal capacity, but
after uptake and transformation by susceptible fungal cells, its metabolites become incorporated
into DNA and RNA molecules (82). 5-FC enters fungal cells and is converted to 5-fluorouracil
(5-FU), and then into 5-fluorouridylic acid (FUMP). This pyrimidine analog is then incorporated
into RNA, resulting in disruption of protein synthesis. 5-FU is also converted to
5-fluorodeoxyuridine monophosphate and interferes with DNA synthesis (79). The main adverse
effects of 5-FU, hepatotoxicity and myelotoxicity, have been ascribed to the occurrence of
significant plasma levels of 5-FU during treatment with 5-FC (76). An increased hematotoxicity
is expected when 5-FC is administered together with cytostatic and immunosuppressive drugs
(82).
2.6.2.4 Echinocandins
Echinocandins (e.g. caspofungin and micafungin) are semi-synthetic derivatives of fungal
fermentation products (76). They are cyclic lipopeptides that specifically inhibit the fungal
ß-glucan synthase. They are administered intravenously for systemic candidal infections (83).

17
ß-glucan inhibitors act as specific noncompetitive inhibitors of ß-(1,3)-glucan synthase.
Treatment of fungi with these compounds inhibits the synthesis of the structural glucan
components. Ultrastructural changes in fungi characterized by thickened cell walls and buds
failing to separate from mother cells have been observed. Cells also become osmotically
sensitive and lysis is observed in actively growing hyphae (79). There are few side effects but
several drug interactions can be expected. Concomitment administration of caspofungin and
immunosuppressant (i.e. cyclosporin A) increases the serum levels of the echinocandin. However
there is reduced serum availability when it is administered with highly active anti-retroviral
therapy (HAART) drugs (efavirenz, nevirapine, rifampicin or tacrolimus), anticonvulsants
(phenytoin or carbamazepine) and steroids (dexamesasone) (76). This is especially important to
recognize when treating individuals on HAART.
2.6.2.5 Allylamines
Allylamines (e.g. terbinafine and naftifine) act by inhibiting early steps of ergosterol
biosynthesis. This inhibition coincides with accumulation of the sterol precursor squalene and
the absence of any other sterol intermediate, suggesting that allylamines inhibition of sterol
synthesis occurs at the point of squalene epoxidation, a reaction catalyzed by squalene
epoxidase. Cell death is related primarily to the accumulation of squalene leading to increase
membrane permeability (79). These agents are typically applied topically to dermal surfaces
(skin/fingernails) and can also be used to treat oral candidiasis in conjunction with azoles (84).
There are minimal side effects but there are several drug-drug interactions with some
psychopharmacologic agents (e.g., paroxetine and codeine) and cardiovascular drugs (e.g.
metoprolol) (85). However, unlike azoles, it does not interact with HAART drugs or warfarin
(85).
2.6.3 C. albicans methods of drug resistance to antimycotics
Resistance to various classes of antifungal agents was reviewed recently (86). Antifungal
resistance has been well documented and the mechanisms have been elucidated through in vitro
studies. In addition, biofilms can increase antimicrobial resistance due to alterations in microbial
metabolic pathways, as well as the presence of extracellular materials that prevent penetration of
the agent, or variations in gene expression as previously mentioned (86). There is no specific
gene that imparts resistance to the variety of antifungal agents; rather it can be a combination of

18
altered drug targets, drug modification, and restricting drug penetration (79). This combination
can include: ABC drug efflux pumps (CDR1-4, MDR4), alteration of ergosterol synthesis (Erg1,
3, 11, and 25), synthesis of cell wall ß-glucans (Kre1, Skn1, Fks1), and modification of biofilm
matrix proteins (Zap1, Gcal1, Gcal2, Adh5, Csh1). Listed here are more specific examples of the
three basic drug resistance mechanisms observed in C. albicans (79): 1) Up-regulation of target
enzymes to overwhelm the antifungal agents ability to cause the desired effect. 2) The drug
binding site is altered in such a manner that the drug becomes non-functional. 3) The cells’ drug
efflux pumps remove the antifungal agent. 4) The cell is able to bypass the target enzyme and
compensates by the development of an altered physiological pathway that does not use the
previously targeted enzyme. 5) The antifungal is blocked by the cell wall, whether by enzyme or
diffusion. 6) Metabolic inactivation (intracellular or extracellular) of the antifungal agent through
sequestration in the ECM or enzymatic breakdown.
2.6.4 Resistance to polyenes
Resistance to polyene antibiotics, such as amphotericin B and nystatin, is rare with a suggestion
that development of resistance occurs by selection of naturally occurring resistant cells, present
within the population (79). These naturally resistant cells produce modified sterols that bind
nystatin with lower affinity. Detection of the naturally resistant cells can only be detected in the
presence of nystatin (79). If resistance to polyenes is attained through concurrent alteration in
membrane composition, it can also be lost after serial passage on culture media devoid of
nystatin. The mechanisms underlying shifts in sterol content are not well known. Mechanisms of
resistance to polyenes are determined by studying mutants generated by (i) growing cells in the
presence of increasing concentrations of antifungal agents (multistep mutants), (ii) exposing the
cells to a gradient concentration (4), or (iii) creating mutants by one-step mutation with
mutagenic agents. The most accepted mechanism of resistance involves the alteration of the
sterol synthesis pathway, thereby limiting the amount of sterols present in the cell membrane,
leading to resistance to polyene types of antifungals (79).
2.6.5 Resistance to azoles
Reports of resistance include alterations to the 14α-demethylase gene, alterations in the sterol
synthesis pathway, reduction or over expression of the target enzyme and increased
activity/number of drug efflux pumps (79).

19
2.6.6 Resistance to pyrimidine analogues
The initial promise of pyrimidine analogues has been diminished by the high prevalence of
primary resistance in many fungal species. Monotherapy with the pyrimidine analogue 5-FC is
thus limited because of primary resistance. Two basic mechanisms of resistance can be
distinguished (82): (i) deficiency in the enzymes necessary for cellular transport and uptake of
5-FC or for its metabolism, (ii) increased synthesis of pyrimidines, which compete with 5-FC
during RNA synthesis and thus diminish its ability to affect RNA synthesis.
2.6.7 Resistance to echinocandins
Although few, there have been cases of echinocandin resistance. The mechanism involves
mutations in the genes that encode the targets of the echinocandins, the Fks1 and Fks2 alternative
β(1,3)-glucan synthase subunits (87). Additionally, there is a compensatory cell wall remodelling
that increase C. albicans cell wall chitin biosynthesis permitting some degree of resistance to
echinocandins. Yeast cells with elevated chitin levels are less susceptible to echinocandins.
Mutations to the Fks gene is heritable and subsequently leads to echinocandin resistance in
subsequent generations (83).
2.6.8 Resistance to allylamines
No clinical case reports of candidal resistance related to clinical use of terbinafine have been
published. However, Candida strains resistant to fluconazole express cross-resistance to
terbinafine (79).
2.6.9 Combination therapy
The usage of antifungal agents in combination is one method to circumvent the resistance
mechanism in C. albicans. Combination therapy with 5-FC and amphotericin B, 5-FC with
fluconazoles, amphotericin B and fluconazole, caspofungin and fluconazole have provided
mixed results (88). This means that the combined effects were either synergistic, indifferent or
antagonistic. Synergy occurs when the combination of two different agents reduced the effective
minimum inhibitory concentration of each agent individually. Antagonism occurs when the
combination of agents produces a less efficacious result that either agent alone. The term FIC or
fractional inhibitory concentration is the term given to the combination of therapies. The term
FICI or FIC index refers to the sum of the FIC values. The sum of these values determines

20
whether synergy (FICI <0.5), indifference (0.5<FICI<4) or antagonistic (FICI >4) occurs. A
combination of antifungal agents decreases the chance for resistance to develop and can even
make previously resistant strains susceptible (i.e. strain resistant to fluconazole made susceptible
by the addition of terbinafine (88)). Interestingly, synergy is observed in some studies where
plant derived extracts have been used in combination with pharmaceutical type antifungal agents.
The combination of essential oils from thyme and ketoconazole (89) and curcumin, a
polyphenol, with polyenes or azoles (90) also produce synergistic effects against C. albicans.
2.7 Plant extracts
Traditional medical practice relied on the use of plants and their components as new sources for
drugs. These include bronchodilators (theophylline from Chinese tea), pain inhibitors (morphine
from poppies), and antineoplastics (vincristine from periwinkle) (91). Many other plant derived
medicines also serve to modulate the human immune systems including essential oils and
polyphenols (91).
2.7.1 Essential oils
The components found in essential oils include mono- and sesquiterpines, alcohols, ethers,
aldehydes and ketones (92). These oils can be isolated from berries, bark, leaves and fruit, but
their antimicrobial activities differ depending on the primary source of the naturally occurring
agent. Citronellol has antioxidant activity (93) while linalool has antimicrobial properties and act
synergistically with fluconazole on C. albicans biofilms (94).
2.7.2 Polyphenols
As the name implies, polyphenols are compounds with more than one phenol group (Fig. 3).
They are found ubiquitously in nature. These plant-derived molecules function as to protect the
plants against pathogenic microorganisms including predatory insects (95). They are present in
the human diet in fruits and vegetables, however, precise dietary intake has not been defined
(96). Polyphenols are classified as tannins, lignans, stilbenes, and flavonoids and in fact the latter
comprise the most studied groups because of their multifunctional activities including
antioxidant, antimicrobial, and wound healing properties.

21
Figure 3. Backbone of a polyphenol molecule.
2.7.2.1 Flavonoids
The largest class of the polyphenol family are the flavonoids (97). They are the pigments
responsible for the colouration found in vegetables, flowers, tea, red wine, and citrus fruits (98).
The basic structure is two benzene rings linked through heterocyclicpyran or pyrone (Fig. 3).
Flavonoids are classified into several groups. These include: 1) flavones (e.g. luteolin- parsley),
2) flavonols (e.g. quercetin – broccoli), 3) flavanones (e.g. naringenin – citrus fruits), 4)
flavanols (e.g. epicatechin – apples), 5) anthocyanidins- (e.g. anthocyanin – grapes), 6)
isoflavones (e.g. genistein – soya beans) (98). The estimated daily intake of flavonoids by
humans is 1-2g per day (99).
2.8 Polyphenol activity
2.8.1 Antimicrobial properties
The use of polyphenols as antimicrobial agents was introduced in the late 1960’s (100).
However, the oldest known recorded use (>2000 years) is with propolis, a polyphenol derived
from honey (99). In vitro, polyphenols have been shown to interfere with C. albicans in several
ways but the in vivo concentration required to provide antifungal activity is unknown (99, 101).
Polyphenols act by sequestering metal ions required by fungal metalloproteases (Fe2+
specifically) and inhibit extracellular phospholipases (required for penetrating the epithelium).
Some polyphenols have also been shown to inhibit SAP and lipase activity in C. albicans by 10-
80% (102). Alteration of cell membrane is another mechanism whereby flavonoids can kill
C. albicans. Polyphenols can alter membrane fluidity as the molecules insert themselves into the
ordered lipid bi-layers (103). Once inserted, they disrupt the barrier function of the cell

22
membrane by altering its fluidity thereby causing leakage of ions. The change in transmembrane
ion potential alters the function of ATPases and other membrane-bound enzymes leading to cell
death (101). Polyphenols can also inhibit DNA and RNA synthesis. The B-ring of the polyphenol
(Fig. 2) in a proposed model can disrupt hydrogen bonding between the stacking molecules and
interfering with DNA topoisomerase activity once bound (104). This can lead to DNA cleavage
thereby inhibiting replication of Candida cells (101).
2.8.2 Protective properties
In vitro and in vivo studies have shown that polyphenols possess antioxidant and free radical
scavenging activities that impart, anti-allergic, anti-carcinogenic, anti-thrombotic, and anti-
inflammatory activities (98, 103). Polyphenols can complex with metal ions required by fungal
metalloproteases, thus inhibiting their growth. Polyphenols can also interact with the homeostatic
functions of white blood cells including neutrophils, mast cells, and basophils. They can deplete
host and microbial ROS (e.g. superoxide) thus inhibiting the initiation of inflammation via the
lipoxygenase, prostaglandin and cyclooxygenase pathways (97, 99). Additionally, lysozyme or
histamine release from these cells can be inhibited by polyphenols (105). In vitro studies on
specific polyphenols, like rutin, can block nitrous oxide production by LPS-activated
macrophages (106). In addition to protection, polyphenols can also promote wound healing,
though the exact mechanism is not known (107). For a recent review on the effects of flavonoids
on white blood cells please see (98).
2.8.2.1 Antioxidant
The immune response to candidal infections leads to the recruitment of cells capable of
phagocytosis ( i.e., monocytes, macrophges, neutrophils). These cells generate free radicals that
can kill invading microbial cells. Unfortunately this response can inadvertently target the host,
promoting inflammation leading to damage of host tissues. Some flavonoids can inhibit the
respiratory burst caused by the phagocytes. One mechanism involves inhibition of protein kinase
C and myeloperoxidase (98). This is very important since hydrogen peroxide can stimulate the
formation of hyphae, one of the principal features of pathogenic C. albicans (108).

23
2.9 Biosecur c320c
Biosecur c320c (B320) is a soluble product derived from citrus fruit which has antibacterial
properties (109). Several citrus fruits are used to manufacture this extract including lemons,
bergamots, limes, and oranges. Within the extract, identified components include ascorbic acid,
organic acids, and citrus bioflavonoid. The bioflavonoids include hesperindin, rutin, and
naringenin. The exact composition of B320 is proprietary information but we can determine the
flavonoids present based on other studies including the citrus fruit composition (110). Biosecur
Lab Inc. (www.biosecur.com) obtained a self-affirmed ‘Generally Regarded as Safe’ (GRAS)
designation in March 2011 in accordance with FDA regulations for B320, meaning it is safe for
human consumption as a food product or additive. This product currently has an EcoCert Canada
and USDA/NOP (National Organic Program) designation meaning that it is recognized in
Canada and USA as an organic product. In laboratory studies, B320 has been shown to possess
antibacterial (against other oral bacteria), antiparasitic, fungicidal (other Candida species),
bacteriostatic, and antiseptic properties (Yves Méthot, BioSecur Inc., personal communication).

24
3. Statement of the problem Antimicrobial resistance, in this case in relation to candidal biofilms, is a growing concern for
the human population. Through overuse and misuse, antimycotic drugs have become less
effective against oral pathogens. In particular, C. albicans, in the form of superficial candidal
infections, have become less susceptible to standard antifungal agents making treatment much
more difficult and less predictable. With the increasing size of the immunocompromised and
aging populations, the development of potent and safe antifungal agents with as few side effects
as possible is needed. It is postulated that a naturally produced agent, Biosecur c320c (B320),
could fill this need.
3.1 Objective
To investigate the antifungal efficacy of citrus fruit flavonoids against C. albicans cells.
3.2 Aims
1. To determine the antifungal efficacy of B320 against C. albicans cells.
2. To evaluate the effect of B320 combined with nystatin on C. albicans cells.
3. To determine if C. albicans cells develop resistance to B320.
3.3 Rationale
Previous in vitro studies demonstrated that 83% of C. albicans cells are eliminated after
immediate contact with B320 at 1%, while 99.9% are eliminated after seven days of
treatment with B320 at 1% (Yves Méthot, BioSecur Inc., Personal communication). The
rationale for suggesting that a combination of B320 with nystatin will act synergistically or
at least additively with regard to killing of candidal organisms is based on investigations
carried out with flavonoids combined with antifungals (111).
3.4 Hypothesis
A citrus fruit extract (B320) is an effective antimycotic agent against planktonic and
biofilm cells of C. albicans and; B230 combined with nystatin will have either a synergistic
or additive antimycotic effect against C. albicans cells.

25
4. Materials and Methods
4.1 Fungal cultures and growth conditions
Two laboratory strains were used in this study, C. albicans ATCC 10231 (nystatin-sensitive;
hereafter referred to as WT) and C. albicans ATCC 44831 (nystatin-resistant; hereafter referred
to as NYSR). C. albicans strains were routinely grown on solid Sabouraud Dextrose agar (SDA;
30 g per liter) or yeast peptone glucose (YPG; 1% (w/v) yeast extract, 2% (w/v) peptone, 2%
(w/v) glucose) agar at 37°C in air. For batch cultures, C. albicans cells were grown in YPG
liquid medium at 30°C in air with agitation. Microbial growth after 24-h was
spectrophotometrically measured at an absorbance of 530 nm (OD530).
4.2 Test compounds
Biosecur c320c (B320) was kindly provided by Biosecur Lab Inc. B320 is derived from citrus
fruits (lemon, bergamots, limes and oranges) and is composed of vegetable glycerin (60%), citrus
biomass (28.5%), ascorbic acid (5.5%), citrus polyphenols (3.5%), citrus bioflavonoids (1.3%),
and citric acid/lactic acid/citric pectin/sugar (1%). The bioflavonoids found in this product
include hesperindin, rutin, and naringenin. The exact nature of these components is proprietary
knowledge and thus not available to our laboratory. A carrier agent was used as control. This
carrier agent included the ingredients identified by Biosecur Lab Inc.: 60% glycerin, 5.5%
ascorbic acid, and 1% citric acid. A generic nystatin mouth rinse suspension (100,000 IU/ml)
was obtained from Ratio-Pharm (Mirabel, Quebec, Canada).
4.3 Biofilm growth
C. albicans biofilms were grown in 96-well and 12-well polystyrene microtiter plates. The
growth of the biofilm was initiated by inoculating ~1 x105 CFU/ml of an overnight culture into
300 µl of YPG medium in the individual wells of a 96-well microtiter plate or by inoculating
~1 x105 CFU/ml of an overnight culture into 3 ml of YPG medium in the individual wells of a
12-well microtiter plate. The microtiter plates were then incubated at 37°C in air without
agitation for ~6-h (early biofilm), ~24-h (intermediate biofilm), or ~72-h (mature biofilm). For
the development of mature biofilms, the planktonic phase was carefully removed after 24-h and
48 h and replaced with fresh YPG medium.

26
4.4 Determination of the minimum inhibitory concentration
The minimum inhibitory concentration (MIC) of B320 and nystatin was examined using
planktonic cells of C. albicans at low cell density (LCD; ~103 CFU) and high cell density (HCD;
~107 CFU). The experiments were performed in 96-well microtiter plates (in duplicate with three
technical replicates). Varying concentrations of B320 and nystatin were used (B320: 0.00005%,
0.0001%, 0.0005%, 0.001%, 0.005% and 0.01%, nystatin: 1, 3, 10, 30, 100, 300, 1000, 3000
IU/ml). Growth after 24-h was spectrophometrically measured by using an ELISA microtiter
plate reader (model IMark, BioRad Laboratories). The MIC was determined at the lowest test
concentration needed to ensure that culture did not grow over 10% of the relative cell density.
4.5 Treatment of biofilm with test compounds
Biofilms were grown as described previously (see section 4.3). After the incubation, the
planktonic phase was carefully removed and the biofilm cells left intact on the bottom of the
wells were washed once with sterile phosphate-buffered saline (PBS, pH 7.2) to remove loosely
bound cells. The biofilm cells were then treated with 200 µl of the test compound for 60 s at
room temperature. A short incubation time was used to mimic the extremely short exposure to
mouth rinses. Biofilm cells were then washed once with sterile PBS, collected by centrifugation,
and resuspended in PBS. Cells were serially diluted and spot plated on YPG agar and the
percentage of cell survival obtained after 60 s of treatment was determined from plate counts. All
assays were performed at least in duplicate or triplicate from three independent cultures. The test
components included: 1x PBS (control), 0.5% B320, 1% B320, 10% B320, 10% glycerin, 10%
carrier, nystatin at 100,000 IU/ml, and a combination of 10% B320 and nystatin at 90,000 IU/ml.
All test compounds were diluted in 1x PBS (pH 7.2).
4.6 Scanning electron microscopy
Scanning electron microscopy (SEM) was performed using biofilms of C. albicans (WT and
NYSR) formed on the surface of glass discs deposited in 12-well polystyrene microtiter plates.
Biofilms were treated with the test compounds for a total of 60 s as follows: i) sterile PBS; ii)
10% B320; iii) 100,000 IU/ml nystatin; iv) 10% B320 and 90,000 IU/ml nystatin. Cells were
then washed once with PBS and fixed in 10% formalin. Cells were dehydrated through ethanol
rinses (30%, 50%, 70%, 95% and 100%), critical point dried with liquid CO2, mounted and

27
sputter coated with gold. Samples were examined using a scanning electron microscope (Model
S-2500, Hitachi Instruments, San Jose, CA).
4.7 Resistance assay
Cells of C. albicans WT strain were cultured in 96-well polystyrene microtiter plates. Growth
was initiated by inoculating ~1x104 CFU of an overnight culture into fresh YPG medium in the
individual wells of the microtiter plate. For day 1 of the experiment, using B320 as the
antimycotic, 100 µl of C. albicans, 100 µl of YPG, and 100 µl of 0.0075% B320 solution
(0.005% final concentration corresponding to 0.5x MIC) was placed in a single well (total
volume for each well was 300 µl). For day 1 of the experiment using nystatin as the antimycotic,
100 µl of a 15 IU/ml solution of nystatin (5 IU/ml final concentration corresponding to 0.5x
MIC) was used in place of B320. A well with YPG only was used as blank control for each
independent experiment. The plates were incubated overnight at 37°C without agitation. On the
second day, 10 µl of the cell suspension was used to inoculate five individual wells. The first
well (growth control) contained cells and YPG. The final concentrations of B320 used were
0.001%, 0.0025%, 0.005% and 0.01%. This experiment was carried out for 14 days (repeated
transfers/passages). Microbial growth was spectrophotometrically measured at an absorbance of
490 nm on even days. To observe resistance to nystatin, a concurrent experiment was carried out
where the final concentrations of nystatin were 3 IU/ml, 5 IU/ml, 10 IU/ml and 30 IU/ml. Every
day the contents in the wells with 0.5x MIC were resuspended and 10 µl of the microbial
suspension was used to inoculate individual wells as described above. If growth was observed in
the well with the MIC of the test agent, this was considered to be resistant.
4.8 pH assay
The pH of the test compounds were measured using a standard pH meter. The pH meter was
calibrated as recommended by the manufacturer. The pH of 10% B320 solution was adjusted to
6.8 using 1 N NaOH. The pH of the carrier was adjusted to 2.8 using 1 N HCl. Biofilms of
C. albicans were tested with 10% B320 (non-pH adjusted; pH 2.8), 10% B320 (pH 6.8), 10%
carrier (non-pH adjusted; pH 4.56), 10% carrier (pH 2.86) vs. a saline control (pH 7.2). Cell
viability was assessed by counting CFU on replica agar plates as described previously.

28
4.9 Drug synergy
Synergism between B320 and nystatin was assessed by the checkerboard assay method using a
96-well microtiter plate (112). From the first to eighth horizontal column, the following
concentrations of B320 were used: 0, 0.00001%, 0.00005%, 0.0001%, 0.0005%, 0.001%,
0.005%, and 0.01%. Along the first to eighth vertical row, the following concentrations of
nystatin (IU/ml) were used: 0, 0.3, 1, 3, 10, 30, 100, and 300 IU/ml. Each well received 100 µl of
test compound, 100 µl of cells and 100 µl of YPG broth. The plates were incubated at 37°C for
24-h without agitation. The MIC was defined as the lowest concentration of the test compound at
which visible growth was inhibited. The synergism between B320 and nystatin was determined
by calculating the fractional inhibitory concentration (FIC) index using the following formula:
FIC index (FICI) = FIC of B320 + FIC of nystatin; where FIC of B320 = MIC of B320 and
nystatin in combination /MIC of B320, and where FIC nystatin = MIC of B320 and nystatin in
combination /MIC of nystatin. The interaction is synergistic (FICI<0.5), indifferent
(0.5<FICI<4.0) or antagonistic (FICI >4.0)
4.10 Statistical analysis
Cell counts were compared across treatment and/or time using linear mixed models
(Multivariable analysis of ANOVA) for the data shown in Figures 4-7. Linear mixed models
allow the pooling of multiple experiments by adjusting for the intra-experiment correlation (i.e.,
clustering of outcomes within experiment). Linear mixed models were constructed assuming an
exchangeable correlation matrix. Post hoc or multiple comparisons were used to compare
individual treatments conditions with other treatment conditions in a pair wise manner. To adjust
for the inflation in type I error from use, the use of multiple comparisons Tukey-adjusted post
hoc T-tests were used with a level of significance set at <0.05.

29
5. Results
5.1 Determination of CFU from stationary phase cultures
For each C. albicans strain used in this study, serial dilutions of a stationary phase culture
cultivated in YPG-medium were plated on replica agar plates to determine the number of cells.
Our results showed that a stationary phase culture of C. albicans ATCC 10231 (WT) contained
approximately 2.2 x106 CFU/ml, while a stationary phase culture of C. albicans ATCC 44831
(NYSR) contained approximately 1.7 x106 CFU/ml. The WT strain grew as opaque white
colonies and the NYSR strains grew as opaque pink colonies on SDA and YPG-agar plates.
5.2 Antimicrobial susceptibility to B320 and nystatin
In order to determine the antimicrobial susceptibility of C. albicans WT and NYSR strains to
Biosecur c320c (B320) and nystatin, we examined the minimum inhibitory concentration (MIC)
of both test compounds using cultures of C. albicans at low cell density (LCD; ~103 cells) and at
high cell density (HCD; ~107 cells). The MIC of B320 at LCD was 0.005% and 0.001% for the
WT and NYSR strains, respectively. At HCD, the amount of B320 required increased 10 fold for
WT and 100 fold for NYSR. The MIC of nystatin at LCD was determined to be 10 IU/ml and
>100 IU/ml for WT and NYSR strains, respectively. At HCD, the MIC of nystatin increased as
well; approximately 10 fold for WT and 30 fold for NYSR (Table 3). These results demonstrated
that B320 was effective at killing nystatin-sensitive and nystatin-resistant C. albicans planktonic
cells.
5.3 Antimycotic activity of B320
We next assessed the in vitro susceptibility of C. albicans biofilm cells to B320. In order to
mimic the extremely short exposure to a mouth rinse, 24-h-old biofilms of WT strain were
exposed to the test compounds for 60 s. At a concentration of 10%, B320 is effective at reducing
the viability of C. albicans biofilms by ~90% (Figure 4). In contrast, a nystatin treatment of 60 s
failed to reduce biofilm cell viability.
To rule out the possibility that the acidic pH of B320 solution could be responsible for the
observed reduction of cell viability, we adjusted the pH of B320 solution to a more neutral pH
(pH 6.8) and adjusted the pH of the carrier to an acidic pH (pH 2.8). When tested against

30
24-h-old biofilms, 60 s treatment with the carrier solution at pH 2.8 was unable to reduce the
biofilm cell viability (Figure 5). In contrast, B320 solution adjusted to a pH of 6.8 was still
effective at killing more than 90% of the biofilm cells. Altogether, our results clearly
demonstrated that the low pH of B320 was not responsible for the observed reduction of biofilm
cell survival.
5.4 Combined application of B320 and nystatin
There are reports suggesting that a combination of polyphenol and polyene might have additive
or synergistic effects on the killing of C. albicans (90). In order to evaluate the activity of B320
in combination with nystatin, we exposed 6-h, 24-h, and 72-h-old biofilms of WT and NYSR
strains to a mixture of 10% B320 and 90,000 IU/ml nystatin for 60 s. B320 killed approximately
99% of 6-h-old WT biofilm cells and 99.9% were killed when B320 was combined with nystatin
(Figure 6). Less than 90% of the 24-h-old WT biofilm cells survived with B320 alone and over
99.9% were killed with B320 in combination with Nystatin. At 72-h, WT biofilm cell viability
was reduced by 90% with B320 alone or in combination with nystatin. However for the 72-h-old
biofilms, there was no significant difference when B320 was used alone or with nystatin. B320
alone or with nystatin yielded a significant difference compared to the control or nystatin only
treatments at all-time points.
Interestingly, no cell survival was observed when 6-h-old NYSR biofilm cells were treated for
60 s with B320 alone or in combination with nystatin (Figure 7). For 24-h-old NYSR biofilms,
B320 alone reduced cell survival by ~99% while less than 0.1% cell survival was observed when
B320 was used in combination with nystatin. For 72-h-old biofilms, B320 alone or in
combination with nystatin reduced cell viability by at least 90%. Our results demonstrated that
B320 possesses a fungicidal activity in all phases of C. albicans biofilm development. Moreover,
a combination of B320 and nystatin is more efficient at kiling C. albicans early and intermediate
biofilms than either drug alone suggesting that B320 and nystatin may act together.
5.5 Determination of synergistic antimycotic activity
Since the results (presented in section 5.4) were suggestive of B320 and nystatin acting together,
we designed a drug synergy experiment to evaluate the activity of B320 in combination with

31
nystatin. Our results clearly demonstrated that there is no synergy between B320 and nystatin in
the treatment of planktonic cells of C. albicans WT and NYSR strains (Table 4).
5.6 Resistance assay
In order to determine if resistance to B320 could develop, we designed an in vitro assay to detect
the change in sensitivity after successive passages of C. albicans WT cells. Our results,
presented in Table 5, showed that after 14 passages, C. albicans WT strain was still inhibited by
B320 at 0.01%. In contrast, a progressive increase in resistance to nystatin was observed.
5.7 SEM analysis
We used scanning electron microscopy to investigate whether noticeable difference could be
observed between treated and untreated C. albicans biofilms. There is formation of an
amorphous mass of cell debris following a 60 s treatment with a combination of B320 and
nystatin (Figure 8 and 9). Interestingly more cell debris was observed for C. albicans NYSR
strain in comparison with WT strain. Surprinsingly, the NYSR strain produced no hyphae under
the conditions tested. For these results several fields were examined and images of representative
fields were taken and show in the figures stated above.
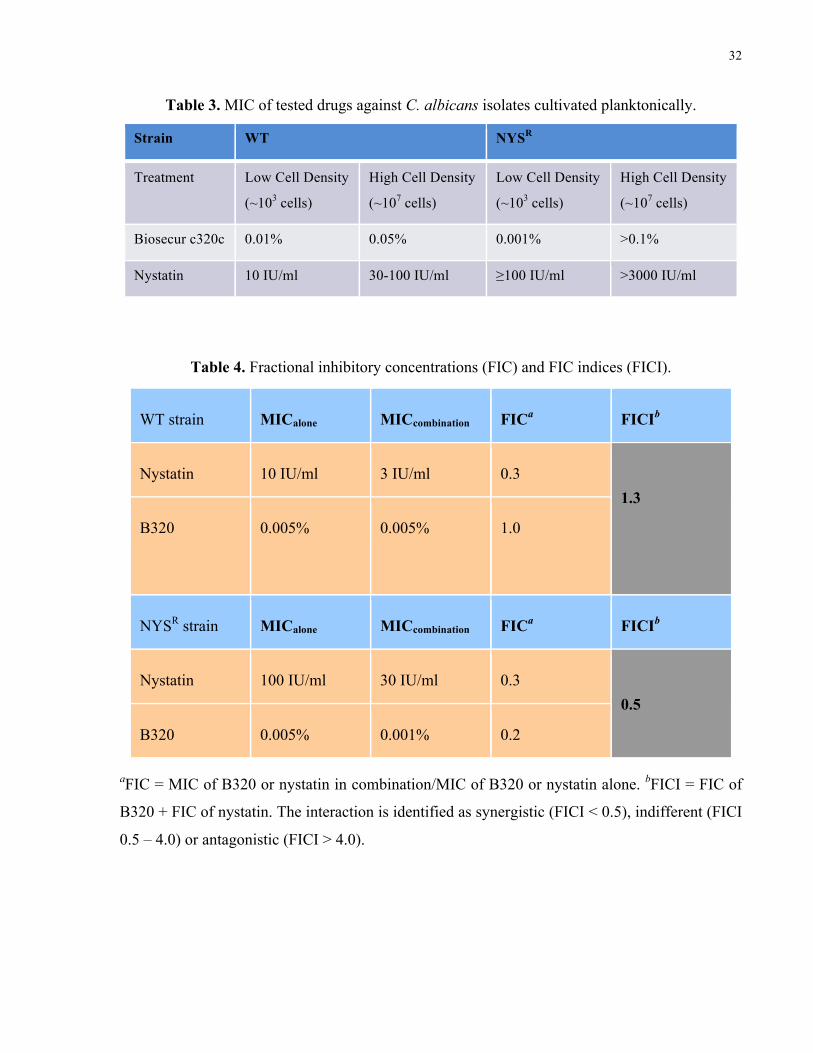
32
Table 3. MIC of tested drugs against C. albicans isolates cultivated planktonically.
Strain WT NYSR
Treatment Low Cell Density
(~103 cells)
High Cell Density
(~107 cells)
Low Cell Density
(~103 cells)
High Cell Density
(~107 cells)
Biosecur c320c 0.01% 0.05% 0.001% >0.1%
Nystatin 10 IU/ml 30-100 IU/ml ≥100 IU/ml >3000 IU/ml
Table 4. Fractional inhibitory concentrations (FIC) and FIC indices (FICI).
WT strain MICalone MICcombination FICa FICIb
Nystatin 10 IU/ml 3 IU/ml 0.3
B320
0.005% 0.005% 1.0
1.3
NYSR strain MICalone MICcombination FICa FICIb
Nystatin 100 IU/ml 30 IU/ml 0.3
B320 0.005% 0.001% 0.2
0.5
aFIC = MIC of B320 or nystatin in combination/MIC of B320 or nystatin alone. bFICI = FIC of
B320 + FIC of nystatin. The interaction is identified as synergistic (FICI < 0.5), indifferent (FICI
0.5 – 4.0) or antagonistic (FICI > 4.0).
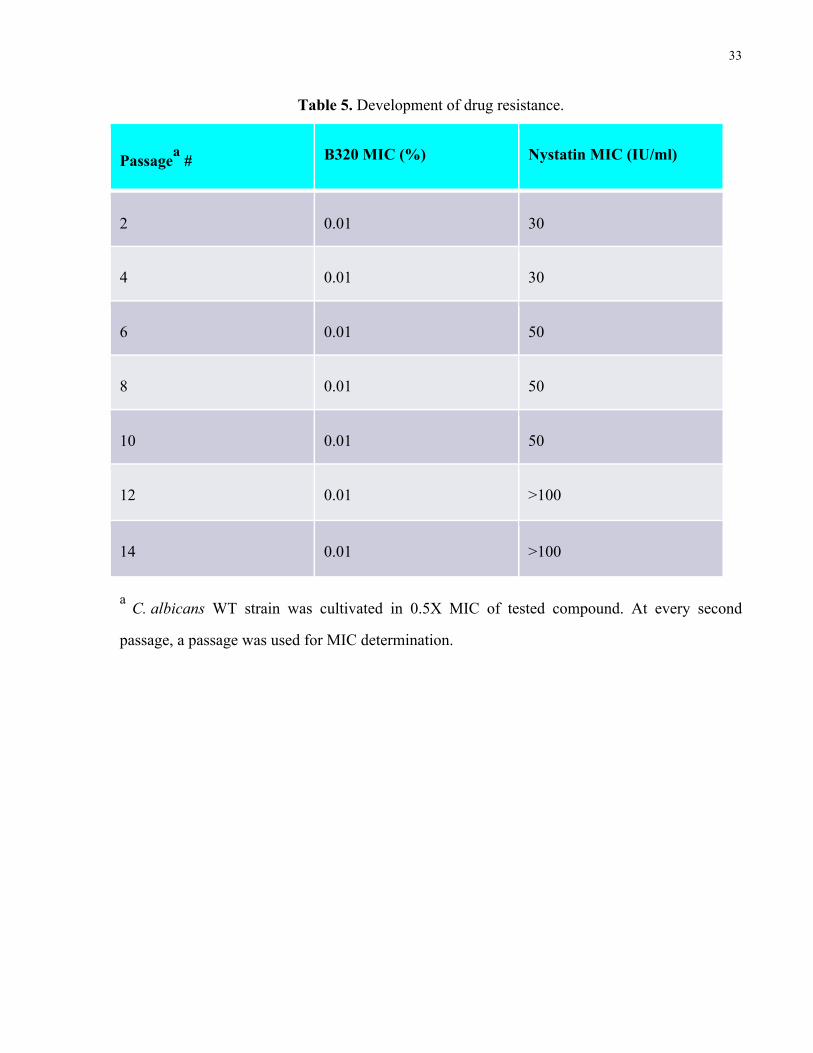
33
Table 5. Development of drug resistance.
Passagea # B320 MIC (%) Nystatin MIC (IU/ml)
2 0.01 30
4 0.01 30
6 0.01 50
8 0.01 50
10 0.01 50
12 0.01 >100
14 0.01 >100
a C. albicans WT strain was cultivated in 0.5X MIC of tested compound. At every second
passage, a passage was used for MIC determination.
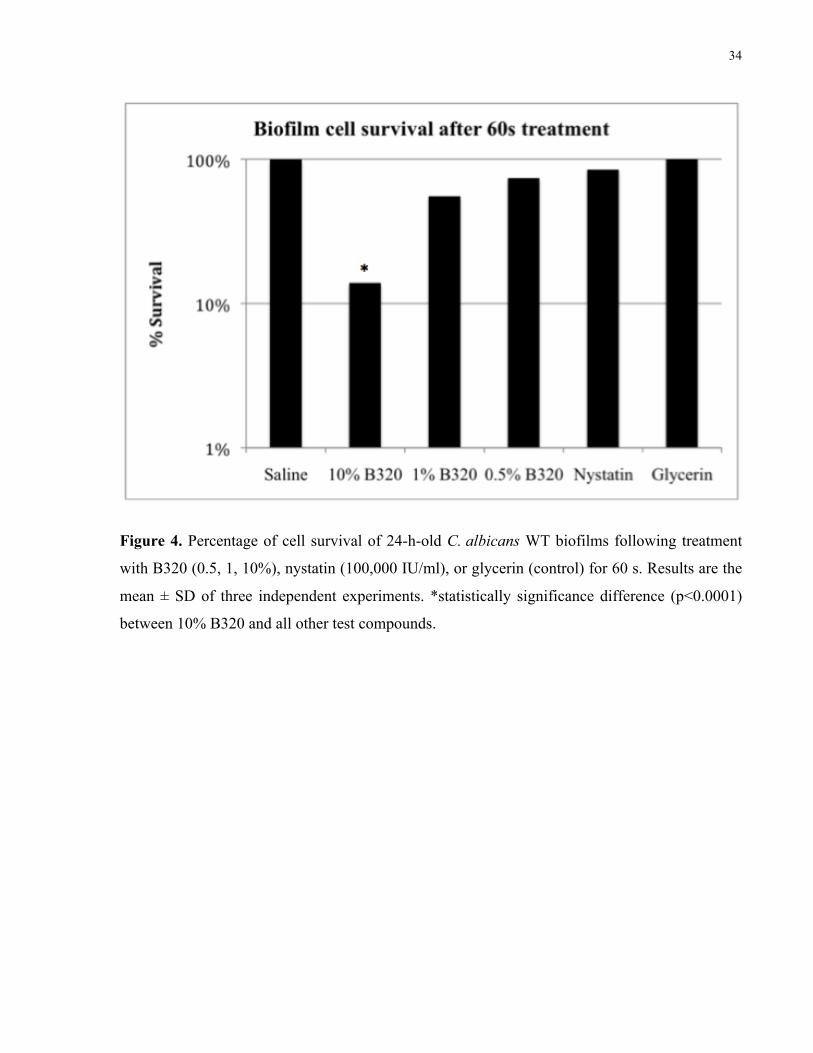
34
Figure 4. Percentage of cell survival of 24-h-old C. albicans WT biofilms following treatment
with B320 (0.5, 1, 10%), nystatin (100,000 IU/ml), or glycerin (control) for 60 s. Results are the
mean ± SD of three independent experiments. *statistically significance difference (p<0.0001)
between 10% B320 and all other test compounds.
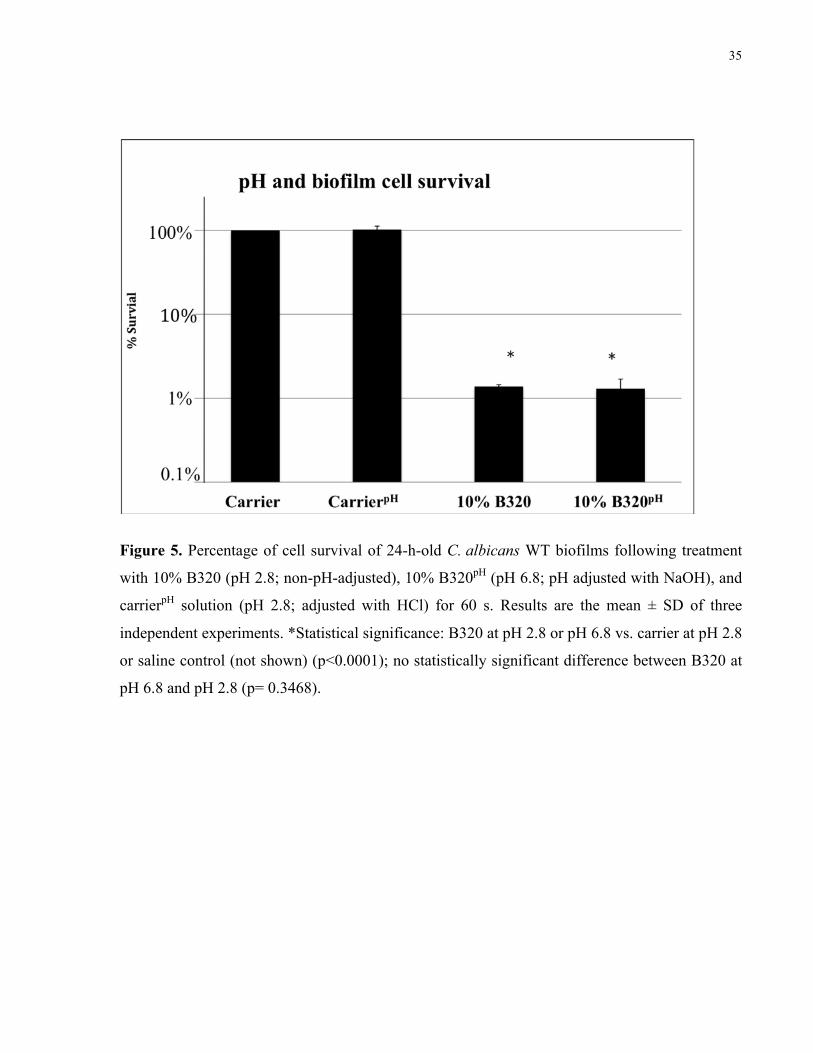
35
Figure 5. Percentage of cell survival of 24-h-old C. albicans WT biofilms following treatment
with 10% B320 (pH 2.8; non-pH-adjusted), 10% B320pH (pH 6.8; pH adjusted with NaOH), and
carrierpH solution (pH 2.8; adjusted with HCl) for 60 s. Results are the mean ± SD of three
independent experiments. *Statistical significance: B320 at pH 2.8 or pH 6.8 vs. carrier at pH 2.8
or saline control (not shown) (p<0.0001); no statistically significant difference between B320 at
pH 6.8 and pH 2.8 (p= 0.3468).
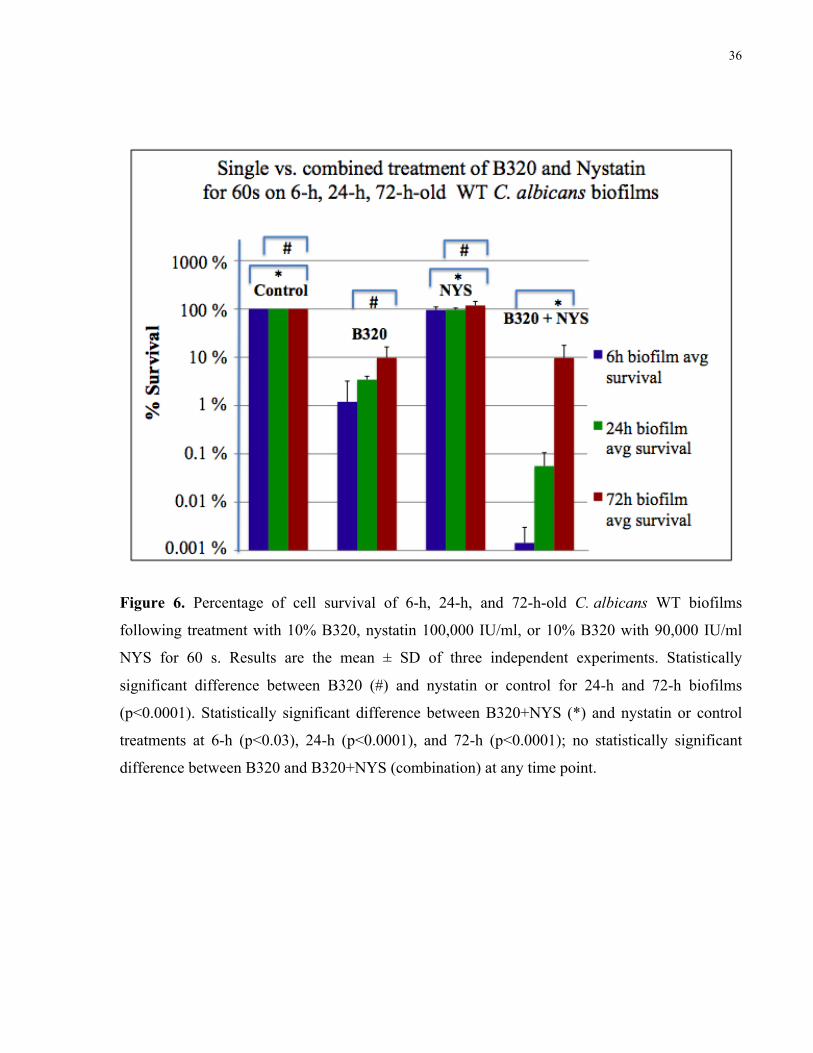
36
Figure 6. Percentage of cell survival of 6-h, 24-h, and 72-h-old C. albicans WT biofilms
following treatment with 10% B320, nystatin 100,000 IU/ml, or 10% B320 with 90,000 IU/ml
NYS for 60 s. Results are the mean ± SD of three independent experiments. Statistically
significant difference between B320 (#) and nystatin or control for 24-h and 72-h biofilms
(p<0.0001). Statistically significant difference between B320+NYS (*) and nystatin or control
treatments at 6-h (p<0.03), 24-h (p<0.0001), and 72-h (p<0.0001); no statistically significant
difference between B320 and B320+NYS (combination) at any time point.
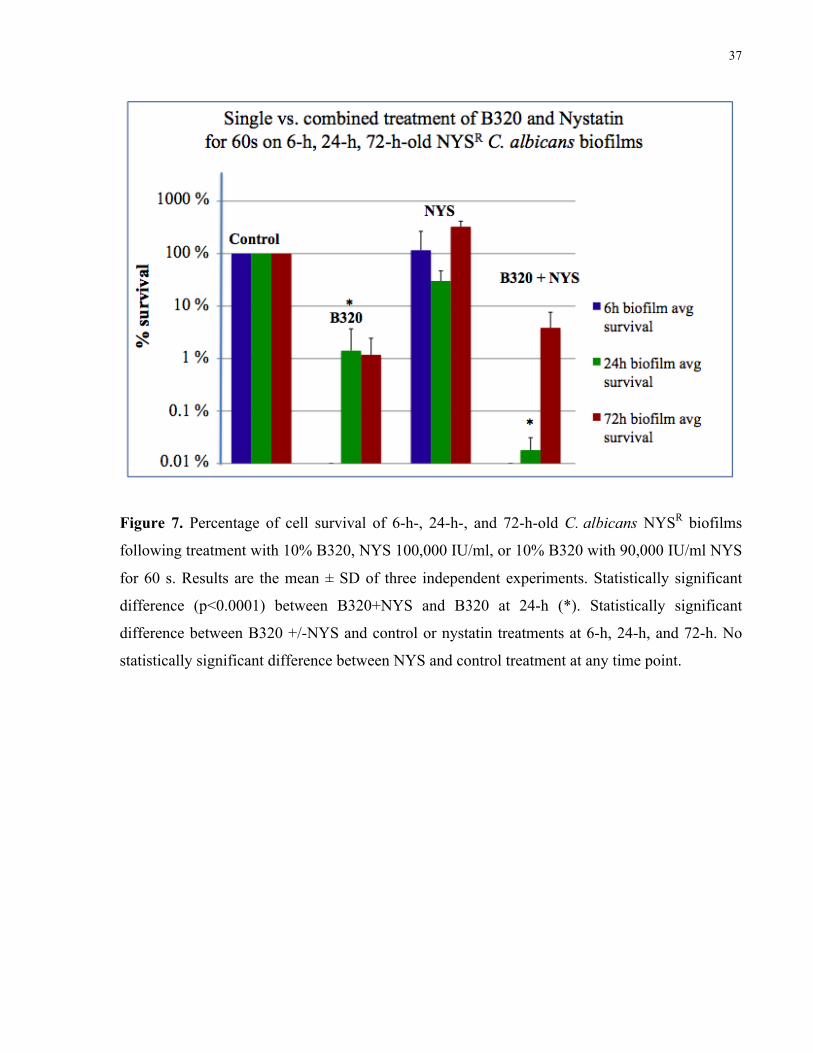
37
Figure 7. Percentage of cell survival of 6-h-, 24-h-, and 72-h-old C. albicans NYSR biofilms
following treatment with 10% B320, NYS 100,000 IU/ml, or 10% B320 with 90,000 IU/ml NYS
for 60 s. Results are the mean ± SD of three independent experiments. Statistically significant
difference (p<0.0001) between B320+NYS and B320 at 24-h (*). Statistically significant
difference between B320 +/-NYS and control or nystatin treatments at 6-h, 24-h, and 72-h. No
statistically significant difference between NYS and control treatment at any time point.

38
Figure 8. Scanning electron micrograph of 24-h-old C. albicans WT biofilms exposed to B320
(10%), nystatin (100,000 IU/ml), or B320 (10%) combined with nystatin (90,000 IU/ml) for 60 s.
There is formation of an amorphous mass of cell debris following exposure the combination
(arrow). Magnification of 1,000 X.

39
Figure 9. Scanning electron micrograph of 24-h-old C. albicans NYSR biofilms exposed to B320
(10%), nystatin (100,000 IU/ml), or B320 (10%) combined with nystatin (90,000 IU/ml) for 60 s.
Formation of an amorphous mass of cell debris following exposure to the combination (arrows).
Magnification of 1,000 X. (Note: no hyphae observed).

40
6. Discussion The components of B320 include extracts from lemon, bergamot, orange, and lime. These
ingredients make up the citrus extracts, which include flavonoids and essential oils. Determining
the flavonoids present in the fruits used to manufacture B320 is relatively easy (see review
(110)), however, the composition of the mixture of citrus biomass would require further analysis
as the composition of B320 is proprietary and therefore was unavailable to the investigator.
However, the antifungal properties of the flavonoids and essential oils have been previously
investigated. Citrus essential oils have been shown to possess antimicrobial properties against
C. albicans (113). The antifungal properties of the essential oils may be related to activation of
quorum-sensing pathways, which direct the cessation of Candida hyphal growth and induces
yeast cell dispersion. The polyphenol curcumin acts to kill Candida by disrupting cell membrane
sterols, making it an effective antifungal agent (114). Since we do not know the concentration of
the components of B320, it cannot be speculated as to what particular component(s) is
responsible for the biological activity of this antimycotic agent. However, the polyphenols
(hesperindin, rutin, and naringenin) are likely to be present in B320, we can hypothesize as to
how the agent may work and which components could be responsible for the antifungal
properties observed in this study. Moreover, the findings reported here are consistent with the
literature in that polyphenols from citrus fruits are effective as antimycotic agents (115). The
hypothesis is that the polyphenols present in B320 act to disrupt the plasma membrane. It is also
possible that the polyphenols trigger a cascade leading to cell death as >90% cell death is
observed after a 60 s application. However, the checkerboard assay for the combined application
of a B320 and nystatin with planktonic C. albicans biofilm cells demonstrated an indifferent
result unlike those in the literature showing synergistic effects between nystatin and a polyphenol
(curcumin) (45, 90).
6.1 B320 as an antifungal
In this investigation it has been demonstrated that concentrations of 0.5%, 1%, and 10% of B320
possess significantly greater (p<0.05) fungicidal properties relative to nystatin at 100,000 IU/ml
(Figure 4). A 10% solution of B320 applied to a 24-h old biofilm for 60 s causes ~95% of the
cells to die relative to a saline control (Figure 6), which is substantially greater than what would
be obtained by the use of nystatin. We can also glean from Figure 6 that when B320 is applied

41
for 60 s to a 6-h and 72-h old WT biofilms, 99% and 90% of the cells die, respectively. Taken
together, these data demonstrate that B320 is an efficient fungicidal agent, and only needs to be
applied for a period of 60 s, a finding that was reproduced across several of the different
experiments shown here. B320 mechanism of action is still to be determined but based on other
studies we can speculate on what they may include. It may act to disperse yeast cells in the
biofilm or act to induce cell death via a quorum-sensing regulating pathway. As well, B320
components may act in a manner that increases the fluidity of the membrane by itself or by
disrupting ergosterol synthesis. It may inhibit formation of hyphae by reducing the availability of
oxidizing agents required for host invasion thereby decreasing the virulence of C. albicans.
Recent work with curcumin has suggested that the antifungal properties of polyphenol may be
related to yeast cell dispersion or cell death via a quorum-sensing regulating pathway (114).
Others have also shown that the polyphenol curcumin (a polyphenol) acts synergistically with
antifungals known as azoles.
6.2 Effects of pH on Activity of B320
The pH of B320 solution is about 2.8. This was of some concern since it was possible that the
low pH, and not the so-called active ingredients in B320 might have been responsible for the
antimycotic activity of B320. Moreover, if this agent is to be used in the mouth, then a pH of 2.8
could theoretically cause irritation to the patients’ mucosa and could lead to dental
demineralization lesions. Therefore it was critically important to determine whether the acidic
pH of B320 was responsible for its antimycotic effects. As shown in Figure 5, it was not the
effect of low pH itself that killed C. albicans cells. Moreover, when the pH was adjusted
upwards to a more physiological level, the antimycotic activity of B320 was retained. This
basically suggests that if B320 were approved for use in humans, the pH of the solution could be
raised without losing the antimycotic activity. It is also important to point out that since altering
the pH of B320 can be a rather time-consuming effort, and since the activity of B320 is identical
at the different pH’s tested, it was decided that it would be appropriate to continue our
experimentation using the original formula for B320 without adjusting the pH.
6.3 MIC of B320
The MIC assay allows for speculation as to the putative mechanism that might explain how B320
is able to kill C. albicans cells. We postulate that the antimycotic effects of B320 are possibly

42
related to the ergosterol content of the cell membrane. In this regard, we observed that the MIC
for NYSR strain, required 100x more B320 at HCD compared to LCD (Table 3). This is in
contrast to what was observed with WT planktonic cells where those at HCD required only 10x
greater concentration of B320 relative to the LCD cells. The decreased ergosterol content of the
NYSR cells may yield fewer targets for B320 to affect the membrane. However, the NYSR cells
require twice the amount of B320 at HCD vs. WT cells. The difference observed between the
WT and NYSR strains of this species in the MIC assay may be related to the ergosterol content of
the cells. However, further studies would be required to verify this hypothesis.
Another research group recently tested two bioflavonoid containing product, derived from citrus
fruits (Citrox®) on planktonic and biofilm cells (115). Their results are similar to those reported
here however their agent was applied for a 24-h duration to C. albicans biofilms and planktonic
cells, whereas our results were obtained after a 60 s application. Other notable issues concerning
the apparent differences between the findings reported here and those reported with Citrox®
were that the other investigators used the ‘McFarland standard’, which uses E. coli as the culture
from which the OD is determined to make a standard curve. In our study, standard curves were
generated using the test species of interest, C. albicans. It would seem to be more scientifically
valid to generate standard curves for calculation of cell counts by using the microbial species of
interest as opposed to another microbe. However, it also has to be recognized that the others’
use of E. coli for generation of their standard curves might not be completely invalid. In this
regard, while this approach might not actually produce true and accurate CFU counts for
C. albicans, but would constitute a systematic error in all tests meaning at least that intra-
experimental comparisons would be somewhat reliable. In addition it was unclear as to whether
the findings reported by others (115) were obtained using biofilms or planktonic presentations of
C. albicans. At any rate, our findings confirmed that the MIC of B320 against planktonic cells
of C. albicans at low and high cell density was 0.005% and 0.05% B320, respectively.
6.4 Effects of combinations of B320 and nystatin on C. albicans
This combination approach of antifungal and citrus extracts is not new. Terpenoids, including
essential oils, are known to impair yeast cell membrane integrity. Together with an azole, the
synergistic effect exhibited by these two agents affects the yeast plasma membrane and cell
signaling (112). Using biofilms of C. albicans (WT and NYSR) it was possible to observe

43
synergistic-like effects with regard to the killing of C. albicans by combining B320 with nystatin
(Figures 6 and 7). Based on the data it is probable that that biofilms at different stages of
maturity might respond differently to these antimycotic agents. As shown in this study, it appears
that early and intermediate biofilms are most susceptible to the synergistic-like effect of B320
and nystatin. However, no statistically significant difference between B320 alone or in
combination with nystatin was found after a multivariate analysis.
When it comes to considering more mature biofilms, the effects of B320 alone or in combination
with nystatin do not have synergistic effects while the cell survival rates are similar (~10%).
Using a checkerboard assay with planktonic cells of the WT and NYSR strains at LCD, synergy
was not observed using the FICI calculation. However, the NYSR cells with a FICI value of 0.5 is
quite close to the being indicative of synergy between B320 and nystatin (FICI < 0.5). As
reported above, the effects of B320 and nystatin were also tested against a nystatin-resistant
strain of C. albicans. This strain, as reported via the ATCC database, does not express ERG2, a
gene responsible for the conversion of fecosterol to episterol along the ergosterol pathway
altering membrane fluidity. It is hypothesized that the polyphenol acts to further disrupt the
already altered membrane lacking ergosterol, thus imparting antifungal effects to this nystatin-
resistant strains of C. albicans. Additionally, it can also be hypothesized that the polyphenol
somehow makes the nystatin-resistant strain susceptible to nystatin.
There is likely an alteration in the mature biofilm (cells or ECM) that prevents full synergy
between B320 and nystatin, as this is not observed in younger biofilms. Although synergy
between B320 and nystatin could not be shown in the planktonic yeast, statistical analysis
showed a statistically significant difference between B320 and nystatin with the intermediate
biofilms formed by the NYSR strain. Furthermore, even if pure synergy could not always be
demonstrated, there is at the very least an important additive effect when both compounds are
used.
6.5 Development of resistance to B320 and nystatin
Given the problems relating to the continual development of microbial resistance to
anti-infective medications, it was decided that it was necessary to demonstrate whether or not,
C. albicans could develop resistance to B320. Planktonic cells were incubated with levels of the
antimycotic agents at doses below the previously determined MIC. It was found that when cells

44
were incubated with sub-MIC levels of the antimycotic agents, they were able to form biofilms.
If the cells developed a biofilm at or above the MIC then it was concluded that the organism had
developed resistance to the agent used. Resistance to B320 was never observed in WT cells over
a two-week period of treatment. When WT cells were exposed to nystatin they were able to
overcome the MIC and develop biofilms at the MIC levels. A 10-fold increase in the MIC level
was observed as the MIC of nystatin increased from 10 to 100 IU/ml. These findings are
consistent with the development of resistance to nystatin for WT strain. This has been reported
by others, where 5-fold increases in resistance were noted by using this approach over a period
of 20 days (116). However, we can only conclude that resistance does not occur after 14 days
and further testing would be required to determine if resistance develops after more time.
6.6 Cell morphology of C. albicans with and without treatment
The SEM results of both C. albicans strains examined (WT and NYSR) demonstrated visible
changes following treatment with the combination of B320 and nystatin. These results are
congruent with a study by Sohn et al. on C. albicans (ATCC 10231) (117). In their investigation
it was surmised that the flavonoid alters the cell wall/membrane resulting in clefting, surface
collapse and wrinkled appearance, like that of a ‘shredded fabric’, of candidal cells, which is
observed in Figure 8 and Figure 9. Of interest is that their result is after incubation of C. albicans
with the flavonoid for 2 h at 30°C. Our results were based on a 60 s exposure at room
temperature. However, we cannot conclude that the morphological change is demonstrative of
cell death.
6.7 Possible sources of error
Any errors that may have occurred during the carefully planned experiments are likely due to the
following.
1) Difficulty in preparing the required concentrations of reagents is related to the high
viscosity of B320. This may affect the accuracy of the prepared solution, specifically the
10% B320 which was used in all the experiments. Additionally, exceptionally small
volumes were used in the microtiter plates; this may have led to some errors as well.
However, completing experiments with three technical replicates and in duplicate or
triplicate minimized this possibility.

45
2) Washing of the biofilms to remove the test agent (B320, nystatin). This action likely
disrupted some of the biofilm. The test agent may have also been left on the biofilm (not
washed off completely) resulting in treatment times greater than 60 s. As experiments
were completed multiple times and the results across different experiments had
reproducible results, it is probable that the potential sources of error discussed here had a
minimal impact or at most constituted a systematic error that would have affected all
cultures treated with one or the other agent. Furthermore, although intraoral use of an
antimycotic rinse involves expectoration after about a 30 s rinse, it is highly likely that
the antimycotic agent is still present (substantivity). In fact, if that was not the case, the
effectiveness of oral rinses might be quite poor. At any rate, given the potential for a
systematic error, intra-experimental comparison of data was deemed reliable while inter-
experimental comparison of data (apart from relative changes) could not be done.
3) There were some concerns associated with the use of microtiter plates and the
measurement of turbidity (OD) to determine viability. The reagents: B320, YPG broth
and nystatin are yellow in colour and can absorb incident light in a spectrophotometer at
various wavelengths. As a result of this, other investigators have used XTT (a tetrazolium
dye) to define cell viability. Candida metabolizes XTT to an orange colour.
Spectrophotometry is then used to compare the XTT reaction at specific wavelengths.
The protocol used for this investigation provided a step to wash away the yellow colour
with saline. The wavelengths used (490 nm or 530 nm) were kept constant within each
experimental protocol to minimize error further.
4) We observed what appeared to be synergy-like between B320 and nystatin with biofilm
but not planktonic C. albicans cells. Several studies have examined the effects of
combining natural products with antifungal agents. In some cases there was synergy
(118) and in others the effect was additive or indifferent (119). Given the inconclusive
results obtained for synergy, another method to test for synergy may be required. The
rationale for stating this is that the biofilm data suggest that there might well be
synergistic antimycotic effects when B320 and nystatin are combined, at least in early
biofilms. However, it must be recognized that both types of tests (biofilm and planktonic
cultures) are important, and could very well yield different types of interactions between

46
B320 and nystatin, since C. albicans exists in the mouth and elsewhere in both the
planktonic and biofilm configurations.
6.8 Summary
Overall this study demonstrated the antimycotic potency of a citrus fruit extract solution, B320
on C. albicans. A kill rate of over 90% after a 60 second application on a biofilm was observed
and pH played no role in the antimycotic activity of B320. No synergy was demonstrated
between B320 and nystatin (a polyene) on a planktonic population. C. albicans could not be
shown to develop resistance to B320 unlike what was observed for cells treated with nystatin.

47
7. Conclusions and future studies The citrus derived agent known as B320 is effective as an antifungal agent alone or in
combination with the polyene antifungal nystatin. It mediates killing of C. albicans in biofilm
and planktonic forms. The mechanism of action of B320 may be multifaceted. Further study of
the mechanism of action of this agent and the possible synergistic effects of B320, a citrus fruit
extract, with various antimycotic pharmaceuticals is required. Future studies using the flavonoids
present in lemons, bergamot, limes, and oranges are available from Sigma-Aldrich and can be
tested alone or in combination based on their relative concentration in the specified citrus fruits
(110). The determination of the minimum biofilm eradication concentration (MBEC) may be
more useful than the MIC. The MIC value pertains to planktonic cells whereas the MBEC relates
to biofilms. The techniques described in (120) can be used in future experiments. Further, animal
models should be used to test the efficacy in vivo before proceeding to human studies. This is
required to determine the most appropriate concentration of agent that is required to act
effectively, and whether it should be used alone or in combination with other antimycotic agents.
We could also further investigate the resistance to B320 by testing over a longer period of time
(>14 days). And since B320 is not specific to C. albicans, the effect of B320 on the normal oral
flora would also need to be evaluated.
This agent could represent an exciting and novel treatment alternative for superficial candidiasis
and possibly other biofilm related human illnesses given that it also has antibacterial effects. It is
as if there has been a re-awakening of ‘traditional medicine’ in the western world, which could
lead to safer and more effective treatments for various disorders (121).

48
References
(1) Nivoix Y, Launoy A, Lutun P, et al. Adherence to recommendations for the use of antifungal agents in a tertiary care hospital. J Antimicrob Chemother 2012.
(2) Cleary JD, Walker LA, Hawke RL. Antimycotic drug discovery in the age of genomics. Am J Pharmacogenomics 2005; 5: 365-386.
(3) Vazquez JA, Sobel, J.D. Candidiasis, In: Dismukes WE, Pappas, P.G., ed. Clinical Mycology. New York, USA: Oxford University press, 2003: 143-187.
(4) Segal E, Baum GL. Pathogenic Yeasts and Yeast Infections London, UK.: CRC Press, 1994.
(5) Sans-Blas G, Calderone RA. Pathogenic Fungi: host fungi interactions and emerging strategies for control Wymondham, UK: Caister Academic press, 2004.
(6) Jones T, Federspiel NA, Chibana H, et al. The diploid genome sequence of Candida albicans. Proc Natl Acad Sci U S A 2004; 101: 7329-7334.
(7) Cannon RD, Holmes AR, Mason AB, Monk BC. Oral Candida: clearance, colonization, or candidiasis? J Dent Res 1995; 74: 1152-1161.
(8) McCullough MJ, Savage NW. Oral candidosis and the therapeutic use of antifungal agents in dentistry. Aust Dent J 2005; 50: S36-39.
(9) Fotos PG, Vincent SD, Hellstein JW. Oral candidosis. Clinical, historical, and therapeutic features of 100 cases. Oral Surg Oral Med Oral Pathol 1992; 74: 41-49.
(10) Jenkins WM, Thomas HC, Mason DK. Oral infections with Candida albicans. Scott Med J 1973; 18: 192-200.
(11) Fotos PG, Lilly JP. Clinical management of oral and perioral candidosis. Dermatol Clin 1996; 14: 273-280.
(12) Zegarelli DJ. Fungal infections of the oral cavity. Otolaryngol Clin North Am 1993; 26: 1069-1089.
(13) Allen CM. Diagnosing and managing oral candidiasis. J Am Dent Assoc 1992; 123: 77-78, 81-72.
(14) Bastidas RJ, Heitman, J. Trimorphic stepping-stones pave the way to fungal virulence. Proceedings of the National Academy of Sciences 2009; 106: 351-352.
(15) Baillie GS, Douglas LJ. Role of dimorphism in the development of Candida albicans biofilms. J Med Microbiol 1999; 48: 671-679.

49
(16) Chaffin WL, Lopez-Ribot JL, Casanova M, Gozalbo D, Martinez JP. Cell wall and secreted proteins of Candida albicans: identification, function, and expression. Microbiol Mol Biol Rev 1998; 62: 130-180.
(17) Cannon RD, Chaffin WL. Oral colonization by Candida albicans. Crit Rev Oral Biol Med 1999; 10: 359-383.
(18) Lo HJ, JR K, B D, D L, A C, GR F. Nonfilamentous C. albicans mutants are avirulent. Cell 1997; 90: 939-949.
(19) Pendrak M, Klotz S. Adherence of Candida albicans to host cells. FEMS Microbiol Lett 1995; 129: 103-113.
(20) Gow N, Hube B. Importance of the Candida albicans cell wall during comensalism and infection. Current Opinion in Microbiology 2012; 15: 1-7.
(21) Klis FM, de Groot P, Hellingwerf K. Molecular organization of the cell wall of Candida albicans. Med Mycol 2001; 39 Suppl 1: 1-8.
(22) Tronchin G, Pihet M, Lopes-Bezerra LM, Bouchara JP. Adherence mechanisms in human pathogenic fungi. Med Mycol 2008; 46: 749-772.
(23) Mishra P, Prasad R. An overview of lipids of Candida albicans. Prog Lipid Res 1990; 29: 65-85.
(24) Chaffin WL. Candida albicans cell wall proteins. Microbiol Mol Biol Rev 2008; 72: 495-544.
(25) Ruiz-Herrera J, Elorza MV, Valentin E, Sentandreu R. Molecular organization of the cell wall of Candida albicans and its relation to pathogenicity. FEMS Yeast Res 2006; 6: 14-29.
(26) Lee CG. Chitin, chitinases and chitinase-like proteins in allergic inflammation and tissue remodeling. Yonsei Med J 2009; 50: 22-30.
(27) Mora-Montes HM, Netea MG, Ferwerda G, et al. Recognition and blocking of innate immunity cells by Candida albicans chitin. Infect Immun 2011; 79: 1961-1970.
(28) Goodridge HS, Wolf AJ, Underhill DM. Beta-glucan recognition by the innate immune system. Immunol Rev 2009; 230: 38-50.
(29) Miura NN, Miura T, Ohno N, et al. Gradual solubilization of Candida cell wall beta-glucan by oxidative degradation in mice. FEMS Immunol Med Microbiol 1998; 21: 123-129.
(30) Domer JE. Candida cell wall mannan: a polysaccharide with diverse immunologic properties. Crit Rev Microbiol 1989; 17: 33-51.

50
(31) Nelson RD, Shibata N, Podzorski RP, Herron MJ. Candida mannan: chemistry, suppression of cell-mediated immunity, and possible mechanisms of action. Clin Microbiol Rev 1991; 4: 1-19.
(32) Saijo S, Iwakura Y. Dectin-1 and Dectin-2 in innate immunity against fungi. Int Immunol 2011; 23: 467-472.
(33) Filler SG. Insights from human studies into the host defense against candidiasis. Cytokine 2012; 58: 129-132.
(34) Thein ZM, Seneviratne CJ, Samaranayake YH, Samaranayake LP. Community lifestyle of Candida in mixed biofilms: a mini review. Mycoses 2009; 52: 467-475.
(35) Jenkinson HF, Douglas LJ. Interactions between Candida species and bacteria in mixed infections, In: Brogden KA, Guthmiller JM, eds. Polymicrobial Diseases. Washington, D.C.: ASM Press, 2002: 357-374.
(36) Lynch AS, Robertson GT. Bacterial and fungal biofilm infections. Annu Rev Med 2008; 59: 415-428.
(37) Ramage G, Saville SP, Thomas DP, Lopez-Ribot JL. Candida biofilms: an update. Eukaryot Cell 2005; 4: 633-638.
(38) Nickerson KW, Atkin AL, Hornby JM. Quorum sensing in dimorphic fungi: farnesol and beyond. Appl Environ Microbiol 2006; 72: 3805-3813.
(39) Williams DW, Kuriyama T, Silva S, Malic S, Lewis MA. Candida biofilms and oral candidosis: treatment and prevention. Periodontol 2000 2011; 55: 250-265.
(40) Ramage G, Rajendran R, Sherry L, Williams C. Fungal biofilm resistance. Int J Microbiol 2012; 2012: 528521.
(41) Lafleur MD, Qi Q, Lewis K. Patients with long-term oral carriage harbor high-persister mutants of Candida albicans. Antimicrob Agents Chemother 2010; 54: 39-44.
(42) Albuquerque P, Casadevall A. Quorum sensing in fungi--a review. Med Mycol 2012; 50: 337-345.
(43) Sharma M, Prasad R. The quorum-sensing molecule farnesol is a modulator of drug efflux mediated by ABC multidrug transporters and synergizes with drugs in Candida albicans. Antimicrob Agents Chemother 2011; 55: 4834-4843.
(44) Alem MA, Oteef MD, Flowers TH, Douglas LJ. Production of tyrosol by Candida albicans biofilms and its role in quorum sensing and biofilm development. Eukaryot Cell 2006; 5: 1770-1779.
(45) Sharma M, Manoharlal R, Shukla S, et al. Curcumin modulates efflux mediated by yeast ABC multidrug transporters and is synergistic with antifungals. Antimicrob Agents Chemother 2009; 53: 3256-3265.

51
(46) Adam B, Baillie GS, Douglas LJ. Mixed species biofilms of Candida albicans and Staphylococcus epidermidis. J Med Microbiol 2002; 51: 344-349.
(47) Chandra J, McCormick TS, Imamura Y, Mukherjee PK, Ghannoum MA. Interaction of Candida albicans with adherent human peripheral blood mononuclear cells increases C. albicans biofilm formation and results in differential expression of pro- and anti-inflammatory cytokines. Infect Immun 2007; 75: 2612-2620.
(48) Mukherjee PK, Chandra J, Kuhn DM, Ghannoum MA. Mechanism of fluconazole resistance in Candida albicans biofilms: phase-specific role of efflux pumps and membrane sterols. Infect Immun 2003; 71: 4333-4340.
(49) Vediyappan G, Rossignol T, d'Enfert C. Interaction of Candida albicans biofilms with antifungals: transcriptional response and binding of antifungals to beta-glucans. Antimicrob Agents Chemother 2010; 54: 2096-2111.
(50) Watamoto T, Samaranayake LP, Egusa H, Yatani H, Seneviratne CJ. Transcriptional regulation of drug-resistance genes in Candida albicans biofilms in response to antifungals. J Med Microbiol 2011; 60: 1241-1247.
(51) Budtz-Jorgensen E, Mojon P, Rentsch A, Deslauriers N. Effects of an oral health program on the occurrence of oral candidosis in a long-term care facility. Community Dent Oral Epidemiol 2000; 28: 141-149.
(52) Budtz-Jorgensen E. Oral mucosal lesions associated with the wearing of removable dentures. J Oral Pathol 1981; 10: 65-80.
(53) Torres SR, Peixoto CB, Caldas DM, et al. Relationship between salivary flow rates and Candida counts in subjects with xerostomia. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2002; 93: 149-154.
(54) Oksala E. Factors predisposing to oral yeast infections. Acta Odontol Scand 1990; 48: 71-74.
(55) Bissell V, Felix DH, Wray D. Comparative trial of fluconazole and amphotericin in the treatment of denture stomatitis. Oral Surg Oral Med Oral Pathol 1993; 76: 35-39.
(56) Nairn RI. Nystatin and amphotericin B in the treatment of denture-related candidiasis. Oral Surg Oral Med Oral Pathol 1975; 40: 68-75.
(57) Arendorf TM, Walker DM. The prevalence and intra-oral distribution of Candida albicans in man. Arch Oral Biol 1980; 25: 1-10.
(58) Olsen I, Stenderup A. Clinical-mycologic diagnosis of oral yeast infections. Acta Odontol Scand 1990; 48: 11-18.
(59) Zipfel PF, Wurzner R, Skerka C. Complement evasion of pathogens: common strategies are shared by diverse organisms. Mol Immunol 2007; 44: 3850-3857.

52
(60) Meri T, Hartmann A, Lenk D, et al. The yeast Candida albicans binds complement regulators factor H and FHL-1. Infect Immun 2002; 70: 5185-5192.
(61) Crowe JD, Sievwright IK, Auld GC, Moore NR, Gow NA, Booth NA. Candida albicans binds human plasminogen: identification of eight plasminogen-binding proteins. Mol Microbiol 2003; 47: 1637-1651.
(62) Budtz-Jorgensen E, Bertram U. Denture stomatitis. I. The etiology in relation to trauma and infection. Acta Odontol Scand 1970; 28: 71-92.
(63) Holmstrup P, Axell T. Classification and clinical manifestations of oral yeast infections. Acta Odontol Scand 1990; 48: 57-59.
(64) Samaranayake LP, Cheung LP, Samaranayake YH. Candidiasis and other fungal diseases of the mouth. Dermatologic Therapy 2002; 15: 251-269.
(65) Neville BW, Damn D, Allen CM, Bouquout JE. Oral and maxillofacial pathology, 2nd edn.: W.B. Saunders, 2002: 843.
(66) Peretz B, Mazor Y, Dagon N, Bar-Ness Greenstein R. Candida, mutans streptococci, oral hygiene and caries in children. J Clin Pediatr Dent 2011; 36: 185-188.
(67) Yang XQ, Zhang Q, Lu LY, Yang R, Liu Y, Zou J. Genotypic distribution of Candida albicans in dental biofilm of Chinese children associated with severe early childhood caries. Arch Oral Biol 2012; 57: 1048-1053.
(68) Reynaud AH, Nygaard-Ostby B, Boygard GK, Eribe ER, Olsen I, Gjermo P. Yeasts in periodontal pockets. J Clin Periodontol 2001; 28: 860-864.
(69) Jarvensivu A, Hietanen J, Rautemaa R, Sorsa T, Richardson M. Candida yeasts in chronic periodontitis tissues and subgingival microbial biofilms in vivo. Oral Dis 2004; 10: 106-112.
(70) Baumgartner JC, Watts CM, Xia T. Occurrence of Candida albicans in infections of endodontic origin. J Endod 2000; 26: 695-698.
(71) Sundqvist G, Figdor D, Persson S, Sjogren U. Microbiologic analysis of teeth with failed endodontic treatment and the outcome of conservative re-treatment. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 1998; 85: 86-93.
(72) Ellepola AN, Samaranayake LP. Oral candidal infections and antimycotics. Crit Rev Oral Biol Med 2000; 11: 172-198.
(73) Blomgren J, Berggren U, Jontell M. Fluconazole versus nystatin in the treatment of oral candidosis. Acta Odontol Scand 1998; 56: 202-205.
(74) Odds FC. Antifungal agents: their diversity and increasing sophistication. Mycologist 2003; 17: 51-55.

53
(75) Arif T, Bhosale JD, Kumar N, et al. Natural products- antifungal agnets derived from plants. Journal of Asian Natural Products Research 2009; 11: 621-638.
(76) Bellmann R. Clinical pharmacokinetics of systemically administered antimycotics. Curr Clin Pharmacol 2007; 2: 37-58.
(77) Dresser GK, Spence JD, Bailey DG. Pharmacokinetic-pharmacodynamic consequences and clinical relevance of cytochrome P450 3A4 inhibition. Clin Pharmacokinet 2000; 38: 41-57.
(78) Stone EA, Fung HB, Kirschenbaum HL. Caspofungin: an echinocandin antifungal agent. Clin Ther 2002; 24: 351-377; discussion 329.
(79) Ghannoum MA, Rice LB. Antifungal agents: mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin Microbiol Rev 1999; 12: 501-517.
(80) Hamill RJ. Part II: 4. Lysosomal Nystatin, In: Dismukes WE, Pappas PG, eds. Clinical Mycology. New York: Oxford University Pres, 2003: 49-58.
(81) Pugh D, Cawson RA. The effect of nystatin on the morphology of Candida albicans. Sabouraudia 1978; 16: 307-311.
(82) Vermes A, Guchelaar HJ, Dankert J. Flucytosine: a review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. J Antimicrob Chemother 2000; 46: 171-179.
(83) Denning DW. Echinocandin antifungal drugs. Lancet 2003; 362: 1142-1151.
(84) Ghannoum MA, Elewski B. Successful treatment of fluconazole-resistant oropharyngeal candidiasis by a combination of fluconazole and terbinafine. Clin Diagn Lab Immunol 1999; 6: 921-923.
(85) Darkes MJ, Scott LJ, Goa KL. Terbinafine: a review of its use in onychomycosis in adults. Am J Clin Dermatol 2003; 4: 39-65.
(86) Niimi M, Firth NA, Cannon RD. Antifungal drug resistance of oral fungi. Odontology 2010; 98: 15-25.
(87) Balashov SV, Park S, Perlin DS. Assessing resistance to the echinocandin antifungal drug caspofungin in Candida albicans by profiling mutations in FKS1. Antimicrob Agents Chemother 2006; 50: 2058-2063.
(88) Johnson MD, MacDougall C, Ostrosky-Zeichner L, Perfect JR, Rex JH. Combination antifungal therapy. Antimicrob Agents Chemother 2004; 48: 693-715.
(89) Hemaiswarya S, Kruthiventi AK, Doble M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008; 15: 639-652.

54
(90) Sharma M, Manoharlal R, Negi AS, Prasad R. Synergistic anticandidal activity of pure polyphenol curcumin I in combination with azoles and polyenes generates reactive oxygen species leading to apoptosis. FEMS Yeast Res 2010; 10: 570-578.
(91) Licciardi PV, Underwood JR. Plant-derived medicines: a novel class of immunological adjuvants. Int Immunopharmacol 2011; 11: 390-398.
(92) Kalemba D, Kunicka A. Antibacterial and antifungal properties of essential oils. Curr Med Chem 2003; 10: 813-829.
(93) Zhuang SR, Chen SL, Tsai JH, et al. Effect of citronellol and the Chinese medical herb complex on cellular immunity of cancer patients receiving chemotherapy/radiotherapy. Phytother Res 2009; 23: 785-790.
(94) Khan MS, Ahmad I. Antibiofilm activity of certain phytocompounds and their synergy with fluconazole against Candida albicans biofilms. J Antimicrob Chemother 2012; 67: 618-621.
(95) Petti S, Scully C. Polyphenols, oral health and disease: A review. J Dent 2009; 37: 413-423.
(96) Landete JM. Updated Knowledge about Polyphenols: Functions, Bioavailability, Metabolism, and Health. Crit Rev Food Sci Nutr 2012; 52: 936-948.
(97) Ross JA, Kasum CM. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annual Review of Nutrition 2002; 22: 19-34.
(98) Middleton JR E, Kandaswami C, Theoharides TC. The effect of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacological Reviews 2000; 52: 673-751.
(99) Havsteen BH. The Biochemistry and medical significance of the flavonoids. Pharmacology & Therapeutics 2002; 96: 67-2002.
(100) Dabbah R, Edwards VM, Moats WA. Antimicrobial action of some citrus fruit oils on selected food-borne bacteria. Appl Microbiol 1970; 19: 27-31.
(101) Cushnie TPT, Lamb AJ. Review: Antimicrobial activity of flavonoids. International Journal of Antimicrobial Agents 2005; 26: 343–356.
(102) Yordanov M, Dimitrova P, Patkar S, Saso L, Ivanovska N. Inhibition of Candida albicans extracellular enzyme activity by selected natural substances and theri application in Candida infection. Canadian Journal of Microbiology 2008; 54: 435-440.
(103) Saija A, Scalese M, Lanza M, Marzullo D, Bonina F, Castelli F. Flavonoids as antioxidant agents: importance of their interaction with biomembranes. Free Radic Biol Med 1995; 19: 481-486.

55
(104) Webb MR, Ebeler SE. Comparative analysis of topoisomerase IB inhibition and DNA intercalation by flavonoids and similar compounds: structural determinates of activity. Biochem J 2004; 384: 527-541.
(105) Middleton jr E, Kandaswami C. Effects of flavonoids on immune and inflammatory cell function. Biochemical Pharmacology 1992; 43: 1167-1179.
(106) Han Y. Rutin has therapeutic effect on septic arthritis caused by Candida albicans. Int Immunopharmacol 2009; 9: 207-211.
(107) San Miguel SM, Opperman LA, Allen EP, Zielinski J, Svoboda KK. Bioactive antioxidant mixtures promote proliferation and migration on human oral fibroblasts. Arch Oral Biol 2011; 56: 812-822.
(108) Srinivasa K, Kim J, Yee S, Kim W, Choi W. A MAP kinase pathway is implicated in the pseudohyphal induction by hydrogen peroxide in Candica albicans. Mol Cells 2012; 33: 183-193.
(109) Levesque C. In Vitro Study of the Antimicrobial Activity of BioSecur C320C. Toronto: Faculty of Dentistry, Dental Research Institute, 2008: 1-6.
(110) Nogata Y, Sakamoto K, Shiratsuchi H, Ishii T, Yano M, Ohta H. Flavonoid composition of fruit tissues of citrus species. Biosci Biotechnol Biochem 2006; 70: 178-192.
(111) Jackson C, Agboke A, Nwoke V. In vitro evaluation of antimicrobial activity of combinations of nystatin and Euphorbia hirta leaf extract against Candida albicans by the checkerboard method. Journal of Medicinal Plants Research 2009; 3: 666-669.
(112) Zore GB, Thakre AD, Jadhav S, Karuppayil SM. Terpenoids inhibit Candida albicans growth by affecting membrane integrity and arrest of cell cycle. Phytomedicine 2011; 18: 1181-1190.
(113) Hammer KA, Carson CF, Riley TV. Antimicrobial activity of essential oils and other plant extracts. J Appl Microbiol 1999; 86: 985-990.
(114) Sharma M, Dhamgaye S, Singh A, Prasad R. Lipidome analysis reveals antifungal polyphenol curcumin affects membrane lipid homeostasis. Front Biosci (Elite Ed) 2012; 4: 1195-1209.
(115) Hooper SJ, Lewis MAO, Wilson MJ, Williams DW. Antimicrobial activity of Citrox bioflavonoid preparations against oral microorganisms. British Dental Journal 2010; 210: 1-5.
(116) Athar MA, Winner HI. The development of resistance by candida species to polyene antibiotics in vitro. J Med Microbiol 1971; 4: 505-517.
(117) Sohn H-Y, Kwon C-S, Son K-H. Fungicidal effect of prenylated flavonol, Papyriflavonol A, isolated from Broussonetia papyrifera (L.) Vent. against Candida albicans. Journal of Microbiology and Biotechnology 2010; 20: 1397-1402.

56
(118) Saad A, Fadli M, Bouaziz M, Benharref A, Mezrioui N-E, Hassani L. Anticandidal activity of the essential oils of Thymus moroccanus and Thymus broussonetti and their synergism with Amphoteracin B and fluconazol. Phytomedicine 2010; 17: 1057-1060.
(119) Rosato A, Vitali C, Piarulli M, Mazzotta M, Argentieri MP, Mallamaci R. In vitro synergic efficacy of the combination of Nystatin with the essential oils of Origanum vulgare and Pelargonium graveolens against some Candida species. Phytomedicine 2009; 16: 972-975.
(120) Ceri H, Olson M, Morck D, et al. The MBEC Assay System: multiple equivalent biofilms for antibiotic and biocide susceptibility testing. Methods Enzymol 2001; 337: 377-385.
(121) Williams D, Lewis M. Pathogenesis and treatment of oral candidosis. J Oral Microbiol 2011; 3.
![Research Article Comparison of the Antifungal Efficacy of ...downloads.hindawi.com/archive/2014/575748.pdfroot canal irrigants and medications to disinfect dentin [ ]. Despite the](https://img.pdfslide.us/doc/110x75/5f18593247a7ab036b42e2d7/research-article-comparison-of-the-antifungal-efficacy-of-root-canal-irrigants.jpg)
![in vitro and in vivo evaluation of efficacy of antifungal ...microbialcell.com/wordpress/wp-content/uploads/... · tissue infections [8]. Of more significance, microbial bio-films](https://img.pdfslide.us/doc/110x75/5e6272ae6daf8008472afe54/in-vitro-and-in-vivo-evaluation-of-efficacy-of-antifungal-tissue-infections.jpg)