Embed Size (px)
Citation preview

Pestic. Sci. 1998, 53, 133È140
Antifeedant Effect, Biological Efficacy and HighAffinity Binding of Imidacloprid to AcetylcholineReceptors in persicae and nicotianaeMyzus Myzus
Ralf Nauen,* Heike Hungenberg, Bienvenu Tollo, Klaus Tietjen & Alfred Elbert
Bayer AG, Agrochemicals Division, Research Insecticides and Molecular Target Research,D-51368 Leverkusen, Germany
(Received 1 May 1997 ; revised version received 6 November 1997 ; accepted 26 January 1998)
Abstract : It is known from laboratory studies that tobacco-associated forms ofMyzus persicae (Sulzer) and the closely related tobacco aphid Myzus nicotianae(Blackman) are often somewhat less susceptible to imidacloprid than non-tobacco strains of M. persicae. Choice tests (Ñoating leaf technique) showed thattobacco aphids were also less susceptible to the antifeedant potential of imid-acloprid in contact bioassays. Synergists like piperonyl butoxide or DEF did notenhance the susceptibility of tobacco-associated morphs of Myzus ssp. to imid-acloprid, thus providing evidence that neither oxidative detoxication nor hydro-lytic metabolization took place. However, in an attempt to study the inÑuence ofendosymbiotic bacteria on the efficacy of imidacloprid, we allowed small popu-lations of tobacco aphids to feed on diets containing the antibiotic chlortetracy-cline prior to imidacloprid treatment. While the e†ectiveness of imidacloprid, i.e.lower values, could be improved in all strains, including the susceptibleLC50reference strain, there was no change in overall tolerance factors. In order toinvestigate any possible alteration of the target site, the affinity of imidaclopridand nicotine to nicotinic acetylcholine receptors in whole-aphid homogenateswas measured. All strains (and clones) showed the same high-affinity bindingsites and no detectable di†erence.
Studies using the FAO dip method revealed that the lower susceptibility of M.nicotianae is not restricted to chloronicotinyls like imidacloprid or acetamiprid,because other insecticides with di†erent modes of action such as pymetrozineand Ðpronil were also a†ected in laboratory studies. It is considered that theobserved tolerance to chloronicotinyls in certain strains of Myzus ssp. is anatural variation in response, probably not coupled with any known mechanismof resistance in this species complex. 1998 SCI(
Pestic. Sci., 53, 133È140 (1998)
Key words : imidacloprid ; Myzus ; acetylcholine receptor ; antifeedant ; cartap
1 INTRODUCTION
Since its Ðrst description as a separate taxon, thetobacco aphid Myzus nicotianae (Blackman), which isclosely related to the green peach aphid Myzus persicae
* To whom correspondence should be addressed.E-mail address : RALF.NAUEN.RN=bayer-ag.de
(Sulzer), has been the object of several papers dealingwith resistance of this species and the related tobacco-feeding morphs of M. persicae to di†erent types ofinsecticides, including carbamates, organophosphatesand pyrethroids.1h5 During 1986È1988, numerousaccounts of control failure against the tobacco aphidwere reported in most tobacco-growing regions of theUnited States.6 The predominant form of the normally
1331998 SCI. Pestic. Sci. 0031-613X/98/$17.50. Printed in Great Britain(

134 Ralf Nauen et al.
green-coloured tobacco aphids became the red-colouredmorph in most areas of the world. The red-colouredform has some advantages in surviving, developing andreproducing at temperatures above 25¡C, thus probablyexplaining its predominance in the Ðeld.7
Surprisingly, some Ðeld populations or even labor-atory clones of M. nicotianae showed reduced suscepti-bility towards the chloronicotinyl insecticideimidacloprid, although these aphids had never been incontact with imidacloprid before. The observed toler-ance factors were in general very low and not obviousin the Ðeld at recommended application rates, but sig-niÐcant in specialized laboratory bioassays.3,8 Further-more, a positive cross-tolerance to nicotine has beendescribed in these aphids, suggesting the possible selec-tion of a mechanism to tolerate certain levels of nicotinewhich may also cover the chloronicotinyl insecti-cides.3,5,8 Nicotine is one of the major alkaloids intobacco plants, the natural host of M. nicotianae, but itis translocated in the xylem, while Myzus ssp. feedmainly in phloem tissue.9 However, some authors sug-gested that one possible reason for the reduced suscepti-bility of M. nicotianae towards imidacloprid lies in themutation of the target site, the nicotinic acetylcholinereceptor.8 This was found not to be the case for atobacco-associated red-coloured morph of M. persicaefrom Japan which was closely related to M. nicotianae.5While it showed a 5È8 fold tolerance to imidacloprid,Nauen et al.5 could demonstrate high-affinity bindingsites for tritiated imidacloprid in homogenates of theJapanese strain as well as in a susceptible referencestrain. There was no statistical di†erence in correspond-ing receptor binding curves.
The objective of the present work was to investigatethe nature of imidacloprid tolerance in M. nicotianaeusing a range of biological, behavioural and biochemi-cal assays, including the measurement of imidaclopridbinding to nAChR in M. nicotianae.
2 MATERIALS AND METHODS
2.1 Insecticides and Chemicals
All insecticides used were of highest purity available.Imidacloprid was obtained in-house. Nicotine was pur-chased from Sigma (St. Louis, USA) and cartap wasobtained from Promochem GmbH (Wesel, Germany).Fipronil (250 g litre~1 SC 250SC), pymetro-(Regent'zine 250 g kg~1 WP 25WP) and acetamiprid(Chess'200 g kg~1 WP 20WP) were purchased(Mospilan'
from the manufacturers. Stock solutions of technicalinsecticides were prepared in acetone (10 g AI litre~1)and diluted in aqueous solutions of “TritonÏ X-100 (1 glitre~1). Chlortetracycline was kindly supplied by Dr P.Meisner (Entomology, in-house). [3H]Imidacloprid
(9É25 ] 1014 Bq Mol~1) was a gift from J. E. Casida(University of California, Berkeley). All other chemicalswere of analytical grade.
2.2 Aphids
The susceptible strain NS of the peach-potato aphid, M.persicae, had been reared in the laboratory since 1967under the conditions described below. Strain JR is ahighly resistant red-coloured M. persicae from Japan,closely related to M. nicotianae as described pre-viously.5 934E, a clone of a North Carolina (USA)population of M. nicotianae, was kindly provided by A.L. Devonshire (Rothamsted Experimental Station, Har-penden, UK). Another M. nicotianae strain (FR) col-lected from tobacco Ðelds in France was kindlyprovided by Yves Bouchery (INRA, Colmar, France).10Distinction between M. persicae and M. nicotianae wasbased, in general, on the electrophoretic separation ofglutamate oxalacetate transaminase allozymes.11 Allstrains were reared on chinese cabbage at 22È23¡C, 60%RH and 16 : 8 h L : D photoperiod.
2.3 Bioassays
The bioassay procedure used was a modiÐcation of theFAO-recommended aphid dip test.12 After dipping ofapterous adults in aqueous insecticidal solutions con-taining “TritonÏ X-100 (1 g litre~1), the aphids weretransferred onto freshly excised cabbage leaves placed inplastic containers. The petioles of the cabbage leaveswere inserted into small plastic tubes containing water.In this modiÐed FAO-procedure, the percentage mor-tality of dipped aphids was evaluated 24 h and 48 hafter dipping.4
The synergists piperonyl butoxide and DEF were dis-solved in “TritonÏ X-100 (1 g litre~1) at a concentrationof 10 mg litre~1. The synergist at the concentrationschosen showed no e†ects on aphids when assessed after48 h. The above-mentioned synergist solution was usedto dilute a stock solution of 10 mg imidacloprid in 1 mlacetone. The starting test concentration for an aphiddip was 100 mg imidacloprid litre~1 in all cases. Per-centage mortality was scored after 24 and 48 h.
The antibiotic chlortetracycline was administeredorally to groups of aphids feeding on artiÐcial doublemembranes (sachets) including an aqueous sucrose solu-tion (150 g litre~1).13 The concentration of chlortetracy-cline in aqueous sucrose was 100 mg litre~1 and aphidswere allowed to feed on sachets 24 h prior dipping withdi†erent concentrations of imidacloprid. Mortality wasassessed after 24 and 48 h. All experiments were repeat-ed at least three times with a minimum of Ðve di†erentconcentrations. At each insecticide concentrations 10 to30 aphids were dipped. Probit analysis was performed

E†ects of imidacloprid on Myzus spp. 135
using the POLO-computer program (LeOra Software,Berkeley USA).
2.4 Choice tests
The method used to elucidate the antifeedant action ofdi†erent insecticides by contact and oral ingestion wasmodiÐed according to Powell et al.14 Leaf discs (25 mmdiam.) were painted on their upper side with 40 kl ofinsecticidal solution in “TritonÏ X-100 (1 g litre~1). Theconcentrations chosen for painting were sublethal (O
as calculated from biotests performed withLC10)dipped leaf discs. Control leaf discs were treated with asolution of “TritonÏ X-100 (1 g litre~1) without insecti-cide. After drying, the painted leaf discs (insecticide-treated and control) were placed on the surface of waterstanding in a plastic Petri dish (85 mm diam.). Leaveswere connected with a small plastic bridge, allowing theaphids to walk from one leaf disc to the other. Theexperiment was started after placing Ðve adult seven-to-eight-day-old apterous aphids from synchronized popu-lations onto each leaf disc. The set-up was covered withthe lid of the Petri dish to catch the excreted honeydewdroplets above each leaf disc. The distribution of theaphids as well as the number of honeydew droplets andimmatures were evaluated after 24 h. The experimentitself was replicated at least 10 times for each insecticideand strain/clone of Myzus ssp. tested. Statistical analysiswas performed with the aid of the computer programINSTAT (GraphPad Software, Inc.). Data wereanalyzed using the following tests : ANOVA, Kruskal-Wallis nonparametric ANOVA test, unpaired t-test andMann-Whitney U-test.
2.5 Nicotinic acetylcholine receptor binding studies with[3H ] imidacloprid
Binding of imidacloprid to aphid nicotinic acetylcholinereceptors was measured according to Liu and Casida15with some minor modiÐcations. Whole aphids(1000 mg), stored frozen in liquid nitrogen before use,were homogenized in a blender in sucrose solution(320 mM ; 20 ml). After centrifugation for 10 min at1200g, the supernatant was Ðltered through Ðve layersof cheesecloth and used directly for binding assays. Theassay (total volume 1 ml) consisted of potassium phos-phate bu†er (0É1 M ; 0É7 ml, pH 7É4), containing 1 glitre~1 bovine serum albumin (binding bu†er ; 1 glitre~1), homogenate (0É25 ml) and [3H]imidacloprid(0É05 ml ; 333 Bq \ 0É36 pmol) in water containingmethanol (16 g litre~1). Unlabeled imidacloprid wasadded in binding bu†er containing up to 0É02 kldimethylsulfoxide. After incubation for 60 min at 22¡C,ice-cold binding bu†er (3 ml) was added, followedimmediately by Ðltration through pre-wetted WhatmanGF/C glass Ðber Ðlters and raising with ice-cold binding
bu†er (2] 3 ml). Bound radioactivity was determinedby scintillation counting of the Ðlters. All values weremeasured in duplicates. values ([lg M of the con-pI50centration of cold ligand displacing 50% of bound[3H]-imidacloprid) were calculated using a four-parameter logistic curve-Ðtting program (GraFit, Eri-thacus Software Ltd). Protein was determined accordingto Bradford16 and standard calibration curves to calcu-late protein content in aphid homogenates were estab-lished using bovine serum albumin.
3 RESULTS AND DISCUSSION
3.1 FAO dip test
Most data Ðtted well using the probit analysis regres-sion lines. In general the tests were aborted after 48 h,even though compounds like pymetrozine showed asomewhat slower action and lower values afterLC5072 h. Even after incubation periods of 72 h, however,tolerance factors were in most cases within the samerange. M. persicae NS was the most susceptible strainwith all compounds tested. Tables 1a and b summarizethe results obtained using the aphid dip technique. Allinsecticides tested were highly e†ective, with LC50values in the same range. Strain JR and clone 934Eshowed tolerance factors between 6 and 10 againstimidacloprid. Tolerance factors to the open-chain ana-logue, acetamiprid, were more or less in the same range.Strain JR was somewhat more tolerant to acetamipridthan clone 934E. However, the computed 95% con-Ðdence intervals were overlapping, indicating that thedi†erence is not signiÐcant (Table 1). The French Ðeldstrain FR of M. nicotianae was nearly as susceptible asthe reference strain NS. That strain losts its hardinessafter rearing for several generations in the laboratory.10Apart from the chloronicotinyls, other insecticides withdi†erent modes of action such as pymetrozine and Ðp-ronil also showed repeatedly a slightly lower efficacyagainst strain JR and clone 934E (but di†erencesbetween reference strain and the other strains were inmost cases non-signiÐcant).
3.2 Efficacy of imidacloprid against Myzus spp. inconjunction with synergists
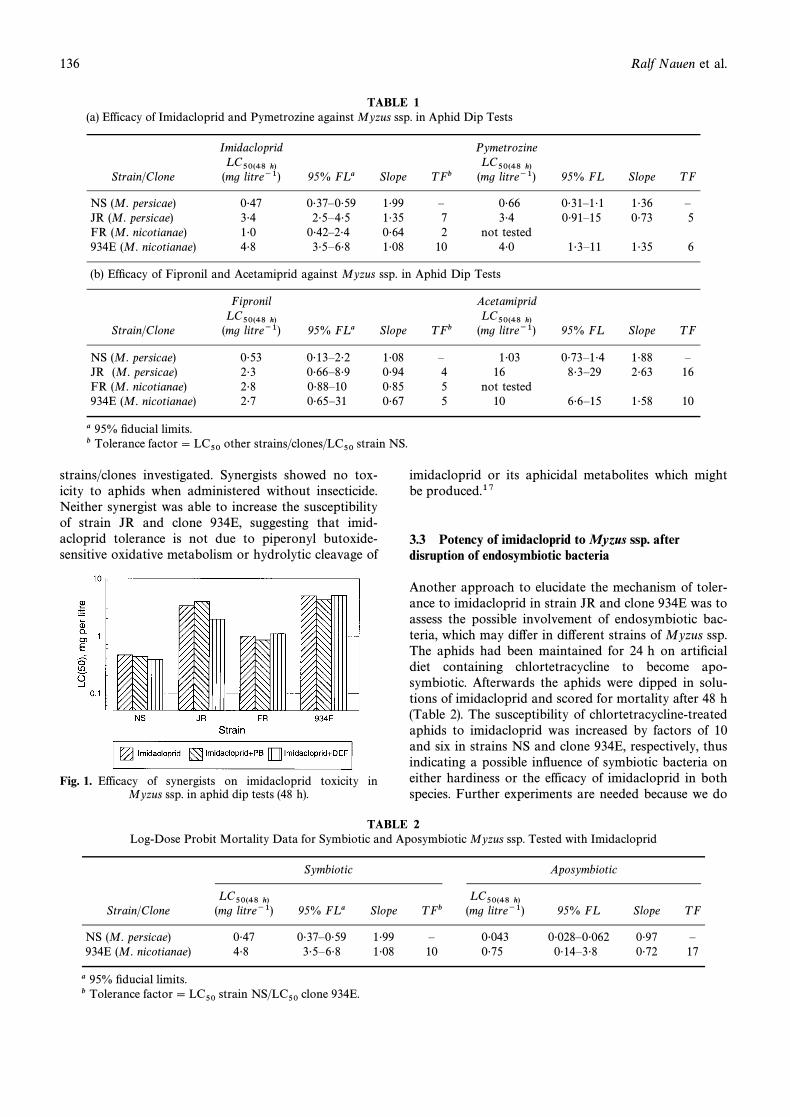
In order to investigate the possible involvement of oxi-dative degradation of imidacloprid, especially in strainJR and clone 934E, we tested the inÑuence of synergistssuch as piperonyl butoxide on values of imid-LC50acloprid. Another synergist included was DEF, which isprimarily active on hydrolytic enzymes, though hydro-lytic cleavage of imidacloprid, due to its structure, is notlikely. Figure 1 clearly indicates no e†ect of standardsynergists on the efficacy of imidacloprid in any of the
136 Ralf Nauen et al.
TABLE 1(a) Efficacy of Imidacloprid and Pymetrozine against Myzus ssp. in Aphid Dip Tests
Imidacloprid PymetrozineL C50(48 h) L C50(48 h)
Strain/Clone (mg litre~1) 95% FL a Slope T Fb (mg litre~1) 95% FL Slope T F
NS (M. persicae) 0É47 0É37È0É59 1É99 È 0É66 0É31È1É1 1É36 ÈJR (M. persicae) 3É4 2É5È4É5 1É35 7 3É4 0É91È15 0É73 5FR (M. nicotianae) 1É0 0É42È2É4 0É64 2 not tested934E (M. nicotianae) 4É8 3É5È6É8 1É08 10 4É0 1É3È11 1É35 6
(b) EfÐcacy of Fipronil and Acetamiprid against Myzus ssp. in Aphid Dip Tests
Fipronil AcetamipridL C50(48 h) L C50(48 h)
Strain/Clone (mg litre~1) 95% FL a Slope T Fb (mg litre~1) 95% FL Slope T F
NS (M. persicae) 0É53 0É13È2É2 1É08 È 1É03 0É73È1É4 1É88 ÈJR (M. persicae) 2É3 0É66È8É9 0É94 4 16 8É3È29 2É63 16FR (M. nicotianae) 2É8 0É88È10 0É85 5 not tested934E (M. nicotianae) 2É7 0É65È31 0É67 5 10 6É6È15 1É58 10
a 95% Ðducial limits.b Tolerance other strain NS.factor \LC50 strains/clones/LC50
strains/clones investigated. Synergists showed no tox-icity to aphids when administered without insecticide.Neither synergist was able to increase the susceptibilityof strain JR and clone 934E, suggesting that imid-acloprid tolerance is not due to piperonyl butoxide-sensitive oxidative metabolism or hydrolytic cleavage of
Fig. 1. Efficacy of synergists on imidacloprid toxicity inMyzus ssp. in aphid dip tests (48 h).
imidacloprid or its aphicidal metabolites which mightbe produced.17
3.3 Potency of imidacloprid to Myzus ssp. afterdisruption of endosymbiotic bacteria
Another approach to elucidate the mechanism of toler-ance to imidacloprid in strain JR and clone 934E was toassess the possible involvement of endosymbiotic bac-teria, which may di†er in di†erent strains of Myzus ssp.The aphids had been maintained for 24 h on artiÐcialdiet containing chlortetracycline to become apo-symbiotic. Afterwards the aphids were dipped in solu-tions of imidacloprid and scored for mortality after 48 h(Table 2). The susceptibility of chlortetracycline-treatedaphids to imidacloprid was increased by factors of 10and six in strains NS and clone 934E, respectively, thusindicating a possible inÑuence of symbiotic bacteria oneither hardiness or the efficacy of imidacloprid in bothspecies. Further experiments are needed because we do
TABLE 2Log-Dose Probit Mortality Data for Symbiotic and Aposymbiotic Myzus ssp. Tested with Imidacloprid
Symbiotic Aposymbiotic
L C50(48 h) L C50(48 h)Strain/Clone (mg litre~1) 95% FL a Slope T Fb (mg litre~1) 95% FL Slope T F
NS (M. persicae) 0É47 0É37È0É59 1É99 È 0É043 0É028È0É062 0É97 È934E (M. nicotianae) 4É8 3É5È6É8 1É08 10 0É75 0É14È3É8 0É72 17
a 95% Ðducial limits.b Tolerance strain clone 934E.factor \LC50 NS/LC50
E†ects of imidacloprid on Myzus spp. 137
not know if all symbiotic bacteria were a†ected aftertreatment with chlortetracycline for 24 h.
3.4 Binding of imidacloprid to nicotinic acetylcholinereceptors in aphid homogenates
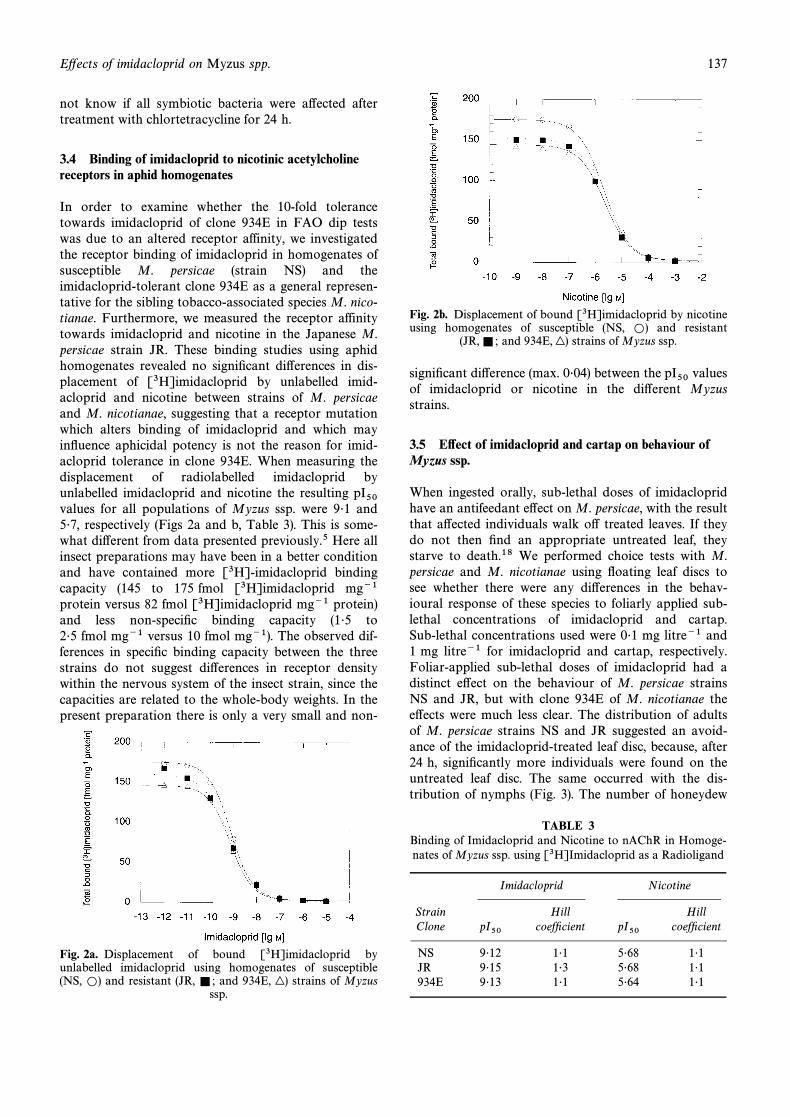
In order to examine whether the 10-fold tolerancetowards imidacloprid of clone 934E in FAO dip testswas due to an altered receptor affinity, we investigatedthe receptor binding of imidacloprid in homogenates ofsusceptible M. persicae (strain NS) and theimidacloprid-tolerant clone 934E as a general represen-tative for the sibling tobacco-associated species M. nico-tianae. Furthermore, we measured the receptor affinitytowards imidacloprid and nicotine in the Japanese M.persicae strain JR. These binding studies using aphidhomogenates revealed no signiÐcant di†erences in dis-placement of [3H]imidacloprid by unlabelled imid-acloprid and nicotine between strains of M. persicaeand M. nicotianae, suggesting that a receptor mutationwhich alters binding of imidacloprid and which mayinÑuence aphicidal potency is not the reason for imid-acloprid tolerance in clone 934E. When measuring thedisplacement of radiolabelled imidacloprid byunlabelled imidacloprid and nicotine the resulting pI50values for all populations of Myzus ssp. were 9É1 and5É7, respectively (Figs 2a and b, Table 3). This is some-what di†erent from data presented previously.5 Here allinsect preparations may have been in a better conditionand have contained more [3H]-imidacloprid bindingcapacity (145 to 175 fmol [3H]imidacloprid mg~1protein versus 82 fmol [3H]imidacloprid mg~1 protein)and less non-speciÐc binding capacity (1É5 to2É5 fmol mg~1 versus 10 fmol mg~1). The observed dif-ferences in speciÐc binding capacity between the threestrains do not suggest di†erences in receptor densitywithin the nervous system of the insect strain, since thecapacities are related to the whole-body weights. In thepresent preparation there is only a very small and non-
Fig. 2a. Displacement of bound [3H]imidacloprid byunlabelled imidacloprid using homogenates of susceptible(NS, and resistant (JR, and 934E, strains of MyzusL) =; |)
ssp.
Fig. 2b. Displacement of bound [3H]imidacloprid by nicotineusing homogenates of susceptible (NS, and resistantL)
(JR, and 934E, strains of Myzus ssp.=; |)
signiÐcant di†erence (max. 0É04) between the valuespI50of imidacloprid or nicotine in the di†erent Myzusstrains.
3.5 E†ect of imidacloprid and cartap on behaviour ofMyzus ssp.
When ingested orally, sub-lethal doses of imidaclopridhave an antifeedant e†ect on M. persicae, with the resultthat a†ected individuals walk o† treated leaves. If theydo not then Ðnd an appropriate untreated leaf, theystarve to death.18 We performed choice tests with M.persicae and M. nicotianae using Ñoating leaf discs tosee whether there were any di†erences in the behav-ioural response of these species to foliarly applied sub-lethal concentrations of imidacloprid and cartap.Sub-lethal concentrations used were 0É1 mg litre~1 and1 mg litre~1 for imidacloprid and cartap, respectively.Foliar-applied sub-lethal doses of imidacloprid had adistinct e†ect on the behaviour of M. persicae strainsNS and JR, but with clone 934E of M. nicotianae thee†ects were much less clear. The distribution of adultsof M. persicae strains NS and JR suggested an avoid-ance of the imidacloprid-treated leaf disc, because, after24 h, signiÐcantly more individuals were found on theuntreated leaf disc. The same occurred with the dis-tribution of nymphs (Fig. 3). The number of honeydew
TABLE 3Binding of Imidacloprid and Nicotine to nAChR in Homoge-nates of Myzus ssp. using [3H]Imidacloprid as a Radioligand
Imidacloprid Nicotine
Strain Hill HillClone pI50 coefÐcient pI50 coefÐcient
NS 9É12 1É1 5É68 1É1JR 9É15 1É3 5É68 1É1934E 9É13 1É1 5É64 1É1

138 Ralf Nauen et al.
Fig. 3. Sub-lethal e†ects of imidacloprid (0É1 mg litre~1) on Myzus ssp. feeding on foliar-treated and untreated leaf discs in choicetests (assessed after 24 h).
droplets excreted on the imidacloprid-treated leaf discover the whole testing period was signiÐcantly lowerthan on untreated ones for both strains (Table 4). In thecase of clone 934E, behavioural di†erences were less
clear and, based on honeydew excretion at 24 h, bothleaf discs were frequented equally. The distribution ofnymphs after 24 h revealed that smaller individuals ofclone 934E might be a†ected by sub-lethal doses of
TABLE 4Average Number of Honeydew Droplets Excreted by Myzus ssp. on Imidacloprid-Treated
and Untreated Control Leaf Discs
Imidacloprid 0É1 mg litre~1 Control SigniÐcanceStrain Species (n droplets) (n droplets) Imi. vs. control
NS M. persicae 27 (^13) 111 (^18) P\ 0É001JR M. persicae 46 (^10) 94 (^12) P\ 0É001934E M. nicotianae 75 (^33) 83 (^30) P\ 0É435
Fig. 4. Sub-lethal e†ects of cartap (1 mg litre~1) on Myzus ssp. in choice tests using treated and untreated leaves (assessed after24 h).

E†ects of imidacloprid on Myzus spp. 139
imidacloprid (Fig. 3). One-way analysis of variance(ANOVA) revealed that the average overall number oflarvae and honeydew droplets within a single test (atleast 10 replicates) was not signiÐcantly di†erent for allstrains of Myzus ssp. tested.
As shown with imidacloprid, foliar-applied sub-lethaldoses of cartap altered the behaviour of M. persicaestrain NS considerably. Cartap was much weaker inactivity against aphids than imidacloprid, hence a con-centration of 1 mg litre~1 was considered to be sub-lethal At this concentration no mortality was(LC10).observed throughout the testing period of 24 h. Thesame observation was made with imidacloprid at0É1 mg litre~1. A signiÐcant preference for the untreatedcontrol leaf disc by M. persicae was found whencounting the number of honeydew droplets excretedabove each leaf disc. However, the distribution of adultsand larvae of M. persicae also suggests an antifeedingpotential for cartap (Fig. 4). Judged from honeydew exc-retion levels, M. nicotianae showed exactly the oppositepreference, i.e. more honeydew droplets above thecartap-treated leaf disc, but statistically not quite signiÐ-cant (P\ 0É0545). However, when assessing the dis-tribution of adults between cartap-treated anduntreated leaf discs after 24 h, signiÐcantly more indi-viduals were feeding on the treated leaf.
CONCLUSIONS
The results of the FAO dip test showed low but signiÐ-cant tolerance in clone 934E, a typical representative ofM. nicotianae derived from North Carolina tobacco, tochloronicotinyl insecticides such as acetamiprid andimidacloprid. Compounds like pymetrozine and Ðpronilwere also less active against clone 934E, but di†erencesrelative to the susceptible reference strain were non-signiÐcant, at least for Ðpronil. For Ðpronil, conÐdenceintervals for all strains overlapped substantially and noconclusions can therefore be drawn regarding lower sus-ceptibility in the imidacloprid-tolerant strains.However, baseline susceptibility of clone 934E com-pared to the reference strain NS of M. persicae wasrepeatedly lower for all compounds tested, suggesting acommon natural hardiness rather than resistance basedon biochemical mechanisms evolved to survive underinsecticide pressure. Strain FR, a former Ðeld strainfrom tobacco in France, loses its tolerance to imid-acloprid when reared under laboratory conditions, asshown very recently.10 Imidacloprid tolerance in M.nicotianae or tobacco-associated M. persicae is wide-spread and correlates often with nicotine tolerance.3,5,8Clone 934E has never been treated with imidacloprid inthe Ðeld, but was derived from tobacco and someauthors have suggested that a possible reason for imid-acloprid tolerance could be reduced receptor affinity,
because of high levels of nicotine in tobacco plants.8However, nicotine is transported apoplastically,whereas the preferred feeding site of Myzus on tobaccois the phloem.9 Nevertheless it is possible that smallamounts of nicotine were imbibed on a regular basiswhen penetrating mesophyll cells in search of thephloem, as is suggested from analysis of electrical pen-etration graph recordings, where certain aphid species,including M. persicae also penetrate the xylem.13,19 Asshown for tobacco-associated morphs of M. persicae,M. nicotianae clone 934E showed no reduced affinity ofthe nicotinic acetylcholine receptor to imidacloprid andnicotine.5 However, in fact, it could be clearly demon-strated for the Ðrst time that homogenates of M. nico-tianae have the same high-affinity binding sites fortritiated imidacloprid as M. persicae. Hence target-siteinsensitivity can probably be excluded as a mechanismin low-level tolerance to imidacloprid of clone 934E andlikewise in other strains of M. nicotianae.
Imidacloprid has strong antifeeding potential againstM. persicae after oral ingestion in systemic biotestsusing excised leaves.8,10,18 It is interesting to note that,in contact bioassays, the antifeedant e†ect of imid-acloprid on M. nicotianae (clone 934E) is considerablylower than against M. persicae. However, it seems to bea common feature of M. nicotianae to tolerate sub-lethaldoses of imidacloprid as shown in studies performedrecently with a number of di†erent clones.8 The reasonfor this general tolerance remains unclear and it may beassociated with the adaption of M. nicotianae totobacco plants. In this connection, one of the most sur-prising results was the fact that, in choice tests, clone934E showed an obvious preference for leaf discstreated with sub-lethal doses of cartap. The reason forthis behaviour is not clear and it would be interesting toinvestigate further e†ectors of the nicotinic acetylcholinereceptor to see if certain structural parts in a moleculedetermine whether a compound acting on the receptoras a neurotoxicant has also a potential as an anti-feedant. It still remains unclear if this antifeedant pro-perty of imidacloprid is related to binding toacetylcholine receptors and therefore to a low-levelintoxication, or to another still unidentiÐed mechanismof action.
Piperonyl butoxide, an inhibitor of microsomalmixed function oxidases, did not enhance the e†ects ofimidacloprid on clone 934E, i.e. co-application of thissynergist with imidacloprid did not result in LC50values closer to the reference strains NS, thus suggestingthat there was no piperonyl-butoxide-suppressible oxi-dative potential in clone 934E to detoxify imidacloprid.The efficacy of piperonyl butoxide in M. persicae hasbeen demonstrated in earlier studies, where it was foundto synergize certain carbamates and theorganophosphate parathion.20,21 Another speculativehypothesis was the involvement of symbiotic bacteriawhich might be able to degrade imidacloprid to a

140 Ralf Nauen et al.
certain extent. In preliminary experiments we tested thishypothesis using the antibiotic chlortetracycline whichhas been successfully used to kill symbiotic bacteria inaphids.22 The so-called aposymbiotic aphids of clone934E became more susceptible to imidacloprid, but sus-ceptible M. persicae of strain NS treated in the sameway became even more susceptible, resulting in toler-ance factors of [10.
In earlier studies it has been shown that the over-expressed esterases E4 or FE4 do not confer resistanceto imidacloprid in Myzus ssp., hence we did not test theinÑuence of esterases in this study.3,5,8 Moreover, theobserved factors of tolerance towards the compoundsused throughout this study would never lead to a Ðeldfailure at recommended application rates for M. per-sicae and M. nicotianae, respectively (Dobri, L., 1996,pers. comm.).
M. nicotianae or tobacco-associated M. persicae ingeneral are aphid species, which often show a loweredsusceptibility to chloronicotinyls (or N-heterocycleinsecticides), probably due to their adaption to a plantrich in secondary plant metabolites such as alkaloidslike nicotine. More detailed investigations in the futuremight reveal the mechanisms that are involved in theobserved tolerance. Detoxication mechanisms like aputative alkaloid pump at its excretory organs/Ðltersystems (malpighian tubules are lacking in aphids) havebeen reported for another nicotine-tolerant pest, thetobacco hornworm Manduca sexta (Joh.) but have notbeen described in aphids so far.23
REFERENCES
1. Blackman, R. L., Morphological discrimination of atobacco-feeding form from Myzus persicae (Sulzer)(Hemiptera : Aphididae) and a key to New World Myzus(Nectarosiphon) species. Bull. Entomol. Res., 77 (1987)713È30.
2. Abdel-Aal, Y. A. I., Lampert, E. P., Roe, R. M. & Semtner,P. J., Diagnostic esterases and insecticide resistance in thetobacco aphid, Myzus nicotianae Blackman (Homoptera :Aphididae). Pestic. Biochem. Physiol., 43 (1992) 123È33.
3. Elbert, A., Nauen, R., Cahill, M., Devonshire, A. L., Scarr,A. W., Sone, S. & Ste†ens, R., Resistance managementwith chloronicotinyl insecticides using imidacloprid as anexample. PÑanzenschutz Nachr. Bayer, 49 (1996) 5È55.
4. Moores, G. D., Devine, G. J. & Devonshire, A. L.,Insecticide-insensitive acetylcholinesterase can enhanceesterase-based resistance in Myzus persicae and Myzusnicotianae. Pestic. Biochem. Physiol., 49 (1994) 114È20.
5. Nauen, R., Strobel, J., Tietjen, K., Otsu, Y., Erdelen, C. &Elbert, A., Aphicidal activity of imidacloprid against atobacco feeding strain of Myzus persicae (Homoptera :Aphididae) from Japan closely related to Myzus nicotianaeand highly resistant to carbamates and organophosphates.Bull. Entomol. Res., 86 (1996) 165È71.
6. Harlow, C. D., Southern, P. S. & Lampert, E. P., Geo-graphic distribution, carboxylesterase activity and
chromosome conÐguration of two color form of thetobacco aphid (Homoptera : Aphididae) in North Carol-ina. J. Econ. Entomol., 84 (1991) 1175È9.
7. Reed, D. T. & Semtner, P. J., InÑuence of temperature onpopulation development of two color morphs of thetobacco aphid (Homoptera : Alphididae) on Ñue-curedtobacco. J. Entomol. Sci., 26 (1990) 33È40.
8. Devine, G. J., Harling, Z. K., Scarr, A. W. & Devonshire,A. L., Resistance to lethal and sub-lethal e†ects of imid-acloprid in nicotine-tolerant Myzus nicotianae and Myzuspersicae. Pestic. Sci., 48 (1996) 57È62.
9. Guthrie, F. E., Campbell, W. V. & Baron, R. L., Feedingsites of the green peach aphid with respect to its adaptionto tobacco. Ann. Entomol. Soc. Am., 55 (1962) 42È6.
10. Nauen, R. & Elbert, A., Apparent tolerance of a Ðeld-collected strain of Myzus nicotianae to imidacloprid dueto strong antifeeding response. Pestic. Sci., 49 (1997)252È8.
11. Blackman, R. L. & Spence, J. M., Electrophoretic distinc-tion between the peach-potato aphid, Myzus persicae andthe tobacco aphid, Myzus nicotianae (Homoptera :Aphididae). Bull. Entomol. Res., 82 (1992) 161È5.
12. FAO, Recommended methods for the detection and mea-surement of resistance of agricultural pests to pesticides :Method for adult aphidsÈFAO method No. 17. FAOPlant Prot. Bull., 18 (1979) 6.
13. Nauen, R. & Elbert, A., E†ect of imidacloprid on aphidsafter seed treatment of cotton in laboratory and green-house experiments. PÑanzenschutz Nachr. Bayer, 47 (1994)181È216.
14. Powell, G., Hardie, J. & Pickett, J. A., E†ects of the anti-feedant polygodial on plant penetration by aphids, assess-ed by video and electrical recording. Entomol. Exp. Appl.,68 (1993) 193È8.
15. Liu, M.-Y. & Casida, J. E., High affinity binding of[3H]imidacloprid in the insect acetylcholine receptor.Pestic. Biochem. Physiol., 46 (1993) 40È6.
16. Bradford, M. M., A rapid and sensitive method for thequantitation of microgram quantities of protein utilizingthe principle of proteinÈdye binding. Anal. Biochem., 72(1976) 248È53.
17. Nauen, R., Tietjen, K., Wagner, K. & Elbert, A., Efficacyof plant metabolites of imidacloprid against aphids.Pestic. Sci., 52 (1998) 53È7.
18. Nauen, R., Behaviour-modifying e†ects of low systemicconcentrations of imidacloprid on Myzus persicae withspecial reference to an antifeeding response. Pestic. Sci., 44(1995) 145È53.
19. Tjalingii, W. F., Electrical nature of recorded signalsduring stylet penetration by aphids. Entomol. Exp. Appl.,38 (1985) 177È86.
20. Attia, F. I., Hamilton, J. T. & Franzmann, B. A., Car-bamate resistance in a Ðeld strain of Myzus persicae(Sulzer) (Hemiptera : Aphididae). Gen. Appl. Entomol., 11(1979) 24È6.
21. Kirknel, E. & Reitzel, J., Insecticide resistance in the greenpeach aphid (Myzus persicae Sulz.). T idsskrift for Plan-teavl., 77 (1973) 191È9.
22. Wilkinson, T. L. & Douglas, A. E., Aphid feeding, as inÑu-enced by disruption of the symbiotic bacteria : an analysisof the pea aphid (Acyrtosiphon pisum). J. Insect Physiol.,41 (1995) 635È40.
23. Murray, C. L., Quaglia, M., Arnason, J. T. & Morris, C.E., A putative nicotine pump at the metabolic bloodÈbrain barrier of the tobacco hornworm. J. Neurobiol, 25(1994) 23È34.





























