Embed Size (px)
Citation preview

THE JOURNAL 0 1989 by The American Society for Biochemistry and Molecular Biology, Inc.
OF BIOLOGICAL CHEMISTRY Vol . 264, No. Issue of September 15, PP. 15177-15181,1989 Printed in U.S.A.
Anticoagulant Low Molecular Weight Heparin Does Not Enhance the Activation of Plasminogen by Tissue Plasminogen Activator*
(Received for publication, March 28, 1989)
Patricia Andrade-Gordon and Sidney Strickland From the Department of Molecular Pharmacology, State University of New York, Stony Brook, New York 11 794-8651
The activity of tissue plasminogen activator (t-PA) and urokinase-type plasminogen activator (u-PA) is stimulated by heparin. Heparin binds tightly to t-PA, u-PA, and plasminogen and decreases the usual stim- ulatory effect of fibrin on t-PA activity. In the present study we have found that low molecular weight hepa- rin (LMW-heparin) preparations obtained by nitrous acid depolymerization or heparinase treatment of standard heparin have different properties with re- spect to their interaction with the fibrinolytic system. LMW-heparin prepared by either method does not stimulate plasmin formation by t-PA. However, these preparations of heparin still efficiently accelerate the inhibition of thrombin by antithrombin 111. Binding data show that LMW-heparin does not bind t-PA and Glu-plasminogen and only binds very weakly to Lys- plasminogen. These results illustrate that it is possible to selectively destroy the fibrinolytic stimulating prop- erties of heparin while leaving the classical anticoag- ulant characteristics intact.
Considerable attention has recently been focused on the use of tissue plasminogen activator (t-PA)’ for the treatment of thrombotic disorders (Collen et al., 1984; TIM1 Study Group, 1985; Verstraete et al., 1985; Verstraete and Collen, 1986; Braunwald et al., 1987). One possible problem with thrombolytic therapy is rapid reocclusion of the affected ar- tery. Therefore, as a preventive measure, patients in recent trials were given heparin before, during, and after the treat- ment with t-PA. There have been previous reports indicating that heparin can enhance fibrinolysis in vivo and in vitro (Vinazzer et al., 1982), but the mechanism has remained obscure. It has now been found that heparin interacts with t- P A and plasminogen and promotes fibrin-independent plas- min generation (Andrade-Gordon and Strickland, 1986; Pi- ques et al., 1986; Soeda et al., 1987; Fears, 1988). Thus, the concomitant use of heparin and t-PA may account for a higher than anticipated level of systemic plasminogen activation.
* This work was supported by New York State Science and Tech- nology Foundation Grant CAT873, National Institutes of Health Grant HD17875, and American Cancer Society Grant BC525. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
’ The abbreviations used are: t-PA, tissue-type plasminogen acti- vator; AT-111, antithrombin 111; u-PA, urokinase-type plasminogen activator; HMW-heparin, high molecular weight heparin; LMW- heparin, low molecular weight heparin; LMW/NA-heparin, low mo- lecular weight heparin obtained by nitrous acid depolymerization; LMW/ase-heparin, low molecular weight heparin obtained after lim- ited heparinase digestion of HMW-heparin; SDS, sodium dodecyl sulfate; pNA, p-nitroanilide; HHT, hexahydrotyrosyl; 20 Ac, diacetate salt.
Commercial heparin is heterogeneous with respect to chem- ical composition, molecular size, anticoagulant activity, bind- ing affinity for antithrombin-I11 (AT-111), and affinities for other coagulation proteases (Hook et al., 1976; Nesheim et al., 1986). Therefore, it appeared possible that preparations of heparin could be obtained that are effective anticoagulants but that do not interact with plasminogen, t-PA, or u-PA.
There is evidence that preparation of low molecular weight heparin (LMW-heparin) cause less bleeding than standard heparin (Turpie et al., 1986; de Prost, 1986). Here, we dem- onstrate that a preparation of LMW-heparin does not bind t- PA and Glu-plasminogen, although it binds weakly to Lys- plasminogen and complexes with urokinase-type plasminogen activator (u-PA). LMW-heparin prepared by two methods does not stimulate the action of t-PA and affects only slightly the activity of u-PA. Importantly, these preparations of hep- arin still efficiently enhance the inhibition of thrombin by AT-111.
EXPERIMENTAL PROCEDURES
Materials-Unfractionated heparin (HMW-heparin) (calcium salt) was derived from porcine intestinal mucosa (148 units/mg, Sigma). LMW/NA-heparin (sodium salt, Sigma) prepared by nitrous acid depolymerization of HMW-heparin had an average size of 4,000- 6,000 daltons by a low angle laser light scattering method and 4,000- 11,000 by gel filtration (see below). Anti-Xa activity was greater than 150 units/mg; activated partial thromboplastin time was greater than 40 IU/mg. Sephadex G-75 (superfine grade) and Sepharose 4B beads were purchased from Pharmacia LKB Biotechnology Inc. The chro- mogenic plasmin substrate H-D-Val-Leu-Lys-pNA (S-2251) was pur- chased from Kabi, and the thrombin substrate H-r-HHT-Ala-Arg- pNA-2OAc was from American Diagnostica. Iodoheads and Iodogen were obtained from Pierce Chemical Co.
The proteins used were prepared or obtained as follows: plasmin- ogen-free bovine fibrinogen (Mosesson, 1962; Strickland and Beers, 1976); soluble fibrinogen fragments, (Verheijen et al., 1982); human Glu-plasminogen (Deutsch and Mertz, 1970); human Lys-plasmino- gen, two-chain t-PA purified from human melanoma (Bowes) cells, fibrinolytic activity, 650,000-750,000 IU/mg, u-PA high molecular weight form, 63,000 units/mg (American Diagnostica); human recom- binant t-PA (Genentech); human thrombin, 3000 NIH units/mg (Sigma); AT-111, 5 IU/mg (Helena Laboratories).
Assay Procedures-The amidolytic plasmin activity of mixtures of Lys-plasminogen and t-PA in the absence or presence of LMW/NA- heparin was measured in a buffer containing 0.1 M Tris-HC1, pH 8.1, 37 “C as described for HMW-heparin (Andrade-Gordon and Strick- land, 1986) or in 0.15 M NaC1, 0.1 M Tris-HC1, pH 7.4 at 37 “ C ; similar results were obtained under both conditions. In addition, the results using melanoma or recombinant t-PA were the same; for all experiments shown, melanoma t-PA was used except for the binding studies (Fig. 6). Inhibition of amidolytic thrombin activity by AT-I11 and HMW- or LMW/NA-heparin preparations was analyzed using the p-nitroanilide substrate H-D-HHT-Ala-Arg-pNA-2OAc at a final concentration of 0.15 M NaCl (Teien et al., 1977; Andrade-Gordon and Strickland, 1986). The slopes of plots of absorbance uersus time (where slope = V,) were used as a measure of remaining thrombin activity. Zymographic analysis of plasminogen activator activity after electrophoresis through SDS-8% polyacrylamide gels was as described
15177
15178 Low Molecular Weight Heparin and Plasminogen Activation (Cranelli-Piperno and Reich, 1978; Vassalli et al., 1984).
Gel Filtration of Heparins-Gel filtration of heparins was per- formed on a Sephadex G-75 (superfine) column (1 X 60 cm) equili- brated in 0.1 M sodium phosphate buffer, pH 7.0, containing 1 mM EDTA and 0.001% Triton X-100. The column was calibrated with polystyrene sulfonates of known molecular weights kindly provided by Dr. Michael Silverberg, SUNY at Stony Brook. They were detected in the eluate by their absorbance at 240 nm. Heparin concentrations were determined by the modification of the procedure utilizing Azure- A (Jaques and Wollin, 1967); a 10-pl sample was added to 200 pl of Azure-A solution (20 pg/ml) in 0.02 M Tris-HCI, pH 8.0, and the absorbance at 490 nm was determined in a Bio-Tek model EL308 microplate reader. A standard curve was generated with known hep- arin concentrations.
Limited Heparin Degradation by Heparinme-HMW-heparin (10 mg, 50 mg/ml) in 0.025 M sodium acetate, 0.25 mM calcium acetate, pH 7.0 at 31 "C, was digested with purified heparinase (100 pl of 50 units/ml) obtained from Seihagaku, Kogyo Co., Japan. Aliquots (60 pl) containing 2 mg of partially degraded heparin were removed after 1 and 24 h of incubation. Aliquots were heated to 95 "C far 5 min, dialyzed against distilled H20, and lyophilized. The same procedure was followed using HMW-heparin without heparinase. Assays for heparinase activity were performed as described (Linhardt et al., 1982). Aliquots (10 p1) were removed over the 24-h digestion time, and the increase in absorbance at 232 nm was measured.
Preparation of LMW-heparin Sepharose-LMWINA-heparin was covalently attached to Sepharose by CNBr activation (Smith et al., 1980). LMW-heparin (250 mg) was dissolved in 60 ml of ice-cold deionized HzO adjusted to pH 7.8, and CNBr (1 g) was added. When the CNBr was completely dissolved, 15 ml of washed Sepharose 4B was added. The solution was kept at pH 11 for 25 min by the addition of 6 M NaOH and then stirred overnight at room temperature. The gel was filtered, washed with 400 ml of H20, and excess reactive groups were blocked with 1 M ethanolamine. Washings were per- formed with 150 ml of H20, 100 ml of 0.1 M NaOAc, pH 4.7, 100 ml of 0.5 M NaHC03, and 400 ml of HzO sequentially. The amount of LMW/NA-heparin coupled to Sepharose was determined colorimet- rically using the metachromatic dye toluidine blue (Smith et al., 1980).
Radioisotope Labeling-Human plasminogens, recombinant t-PA, u-PA, and AT-I11 were radiolabeled by the Iodogen method (Markwell and Fox, 1978) except that Sephadex G-50 (Pharmacia) spin columns were used to separate bound Iz5I from unreacted Iz5I . Specific activities were as follows: 0.3-0.5 mCi/mg for Glu- or Lys-plasminogen; 0.9 mCi/mg for t-PA and AT-111; and 0.8 mCi/mg for u-PA. The struc- tural integrity of all radiolabeled proteins was verified by SDS- polyacrylamide gel electrophoresis followed by autoradiography.
Binding Assays-Measurement of plasminogen, recombinant t-PA, u-PA, and AT-I11 binding to LMW/NA-heparin-Sepharose was per- formed as described for HMW-heparin-agarose (Andrade-Gordon and Strickland, 1986). Dissociation constants were obtained by Scatchard analysis. Nonspecific binding was determined by using unsubstituted Sepharose in place of LMW/NA-heparin-Sepharose. The proteins bound to heparin-Sepharose and the unbound proteins were examined by SDS-8% polyacrylamide gel electrophoresis.
RESULTS
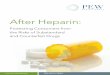
Effects of LMW-heparin on t-PA- or u-PA-mediated Acti- vation of Plasminogen-The influence of heparin on plasmin generation by t-PA or u-PA was analyzed using an amidolytic assay. As previously demonstrated (Andrade-Gordon and Strickland, 1986), HMW-heparin was able to enhance t-PA- mediated plasminogen activation 3-fold at therapeutic con- centrations (approximately 1 unit/ml or 10 pg/ml). On the other hand, t-PA activity was not stimulated when using nitrous acid-prepared LMW-heparin (LMW/NA-heparin) over a wide range of concentrations (0-150 pg/ml) (Fig. lA). The difference between HMW-heparin and LMW/NA-hep- arin was also observed when the assays were performed under conditions of physiological salt and pH (data not shown). It is important to note that although the reported mean mass of this LMW/NA-heparin (4000-6000 daltons) is 25% of that of HMW-heparin, the molar concentration of sugar monomer per given mass of heparin is identical.
, HMW-Heparin b LMW-Heparin
t-PA @
- so - 4 v,
2 0 gm- 88 0 25 50 75 100 125 150
I X
U-PA @
t z 2.0
7
175
101 0.0 1
0 25 50 75 100 125 150 175
HEPARIN (pg/ml)
FIG. 1. Effects of LMW/NA-heparin on the generation of amidolytic plasmin activity. Initial rates were obtained from the slopes of plots of change in absorbance at 405 nm as a function of time squared. A, amidolytic plasmin activity generated by t-PA. The assay was supplemented with either LMW/NA-heparin or HMW- heparin. E , amidolytic plasmin activity generated by u-PA in the presence of LMW/NA-heparin.
The amidolytic assay used to measure plasmin formation is indirect, and effects of LMW/NA-heparin could be obscured, e.g. an increase in plasminogen activation but a decrease in plasmin activity, resulting in no apparent effect. Therefore, plasminogen activation was tested directly by incubating plas- minogen and t-PA with and without LMW/NA-heparin in a solution containing 50% glycerol (glycerol preserves the struc- tural integrity of plasmin by inhibiting self-degradation). After 0, 12, 24, and 36 h, the samples were reduced with dithiothreitol in sample buffer and applied to an SDS-8% polyacrylamide gel. Plasmin generation was assessed by the appearance of bands corresponding to the heavy and light chains of plasmin. In this experiment, the rate of production of plasmin and consequently the consumption of plasminogen was exactly the same for samples with or without LMW/NA- heparin (data not shown). In contrast, the conversion of plasminogen to plasmin by t-PA is accelerated 5-fold in the presence of HMW-heparin (Andrade-Gordon and Strickland, 1986). Thus, LMW/NA-heparin does not enhance the plas- minogen-activating activity of t-PA.
Previously, it was observed that HMW-heparin can inter- fere with the fibrinogen fragment stimulation of t-PA (An- drade-Gordon and Strickland, 1986). LMW/NA-heparin was less effective in this regard; for example, at a concentration of 2.5 wg/ml mucopolysaccharide, HMW-heparin inhibited fibrinogen fragment stimulation 50%, whereas LMW/NA- heparin inhibited 25%.
The influence of LMW/NA-heparin on u-PA was also examined (Fig. 1B). In this case there was a slight dose- dependent stimulation of u-PA-generated plasmin activity by LMW/NA-heparin; this stimulation was about 1.3-fold at a dose of 25 pg/ml LMW/NA-heparin. At doses greater than 75 Fg/ml there was an inhibition of plasminogen activation. In contrast, at doses above 35 pg/ml HMW-heparin plasmin production was stimulated 3-fold (Andrade-Gordon and Strickland, 1986).
Low Molecular Weight Heparin and Plasminogen Activation 15179
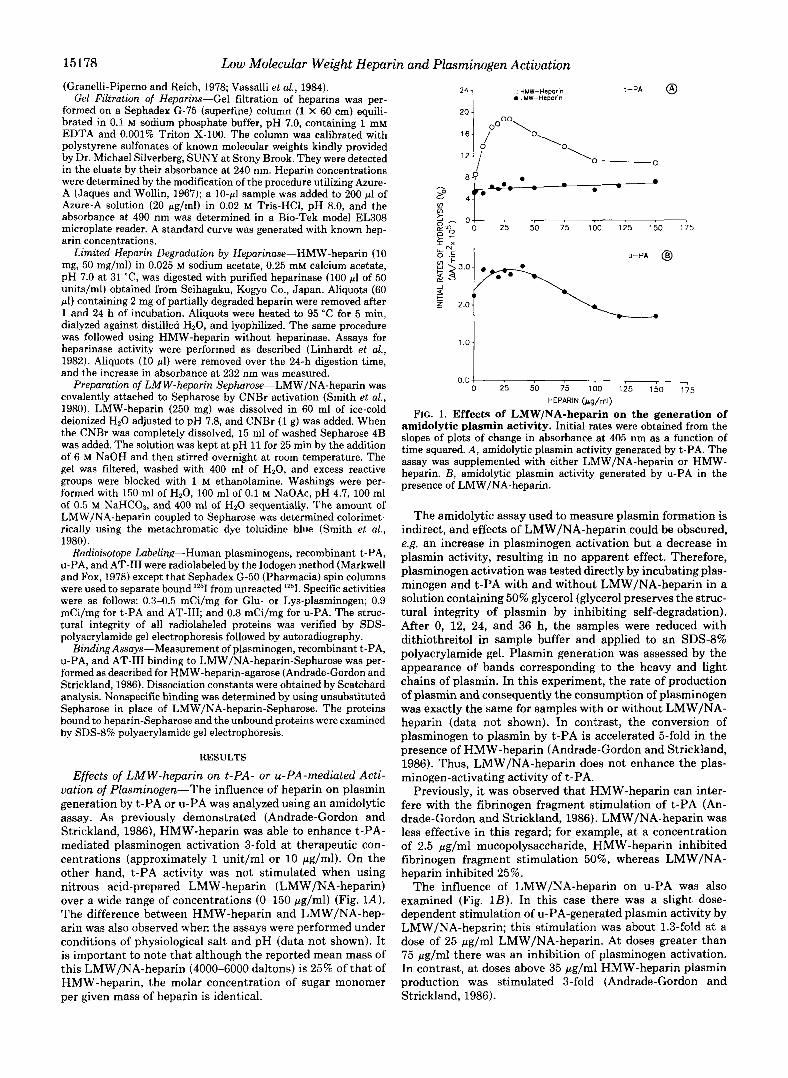
Preparation and Characterization of Low Molecular Weight Heparins-The fact that LMW/NA-heparin did not stimulate t-PA activity had two possible explanations: that the molec- ular size was too small to include this property or that chem- ical modification of sugar groups caused by the nitrous acid treatment had destroyed the activity. To define further the effect of molecular weight, we prepared another depolymer- ized heparin by partial digestion of HMW-heparin with hep- arinase (LMW/ase-heparin). Both preparations of LMW- heparin were then characterized by gel filtration (Fig. 2). Under our conditions, HMW-heparin preparation eluted from the column with a peak corresponding to an apparent molec- ular weight of 18,000. Nitrous acid-prepared LMW-heparin formed a broad peak with a molecular weight distribution of 4,000-11,000, with an average molecular weight of 6,500.
The heparinase-digested product was eluted a t an average M, of 5,700. Undigested sample (HMW-heparin treated in the same manner without addition of heparinase) and a 24-h digested product were also chromatographed. The undigested sample eluted as expected at the same position as HMW- heparin. The 24-h digested product showed several broad peaks of very low apparent molecular weights (data not shown).
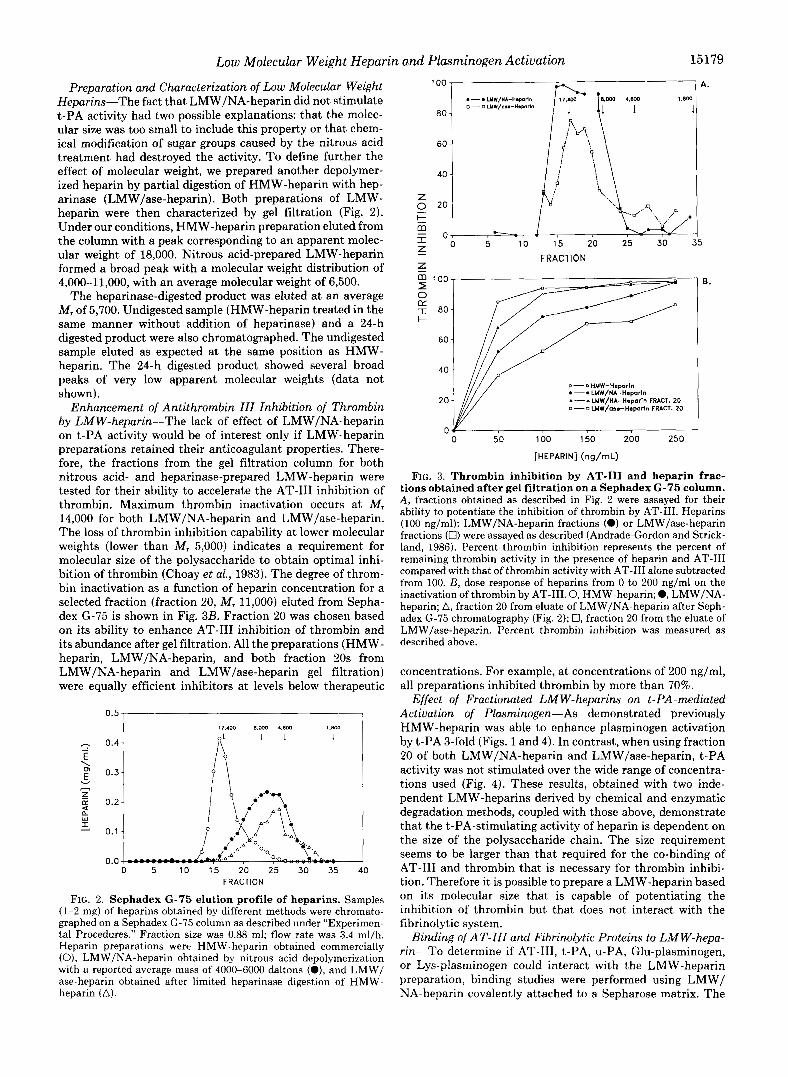
Enhancement of Antithrombin 111 Inhibition of Thrombin by LMW-heparin-The lack of effect of LMW/NA-heparin on t-PA activity would be of interest only if LMW-heparin preparations retained their anticoagulant properties. There- fore, the fractions from the gel filtration column for both nitrous acid- and heparinase-prepared LMW-heparin were tested for their ability to accelerate the AT-I11 inhibition of thrombin. Maximum thrombin inactivation occurs at M, 14,000 for both LMW/NA-heparin and LMW/ase-heparin. The loss of thrombin inhibition capability at lower molecular weights (lower than M, 5,000) indicates a requirement for molecular size of the polysaccharide to obtain optimal inhi- bition of thrombin (Choay et al., 1983). The degree of throm- bin inactivation as a function of heparin concentration for a selected fraction (fraction 20, M, 11,000) eluted from Sepha- dex G-75 is shown in Fig. 3B. Fraction 20 was chosen based on its ability to enhance AT-I11 inhibition of thrombin and its abundance after gel filtration. All the preparations (HMW- heparin, LMW/NA-heparin, and both fraction 20s from LMW/NA-heparin and LMW/ase-heparin gel filtration) were equally efficient inhibitors at levels below therapeutic
0.5 I I
I 17.400 8.000 4.600 ,-. 0.4- E 2 0.3-
_I
v
4 Q w I Y 0.1
0.0 0 5 10 15 20 25 30 35 40
FRACTION
FIG. 2. Sephadex G-75 elution profile of heparins. Samples (1-2 mg) of heparins obtained by different methods were chromato- graphed on a Sephadex G-75 column as described under “Experimen- tal Procedures.” Fraction size was 0.88 ml; flow rate was 3.4 ml/h. Heparin preparations were HMW-heparin obtained commercially (0), LMW/NA-heparin obtained by nitrous acid depolymerization with a reported average mass of 4000-6000 daltons (O), and LMW/ ase-heparin obtained after limited heparinase digestion of HMW- heparin (A).
FRACTION FRACTION
00
80
60
40
0-OHMW-Heporln e--. LMW/NA-Haporln
0 - 0 LMW/ars-Heparln FRACT. 20 ~ - - b LMW/NA-Hsparln FRACT. 20 20
0
00
80
60
40
0-OHMW-Heporln e--. LMW/NA-Haporln
0 - 0 LMW/ars-Heparln FRACT. 20 20 ~ - - b LMW/NA-Hsparln FRACT. 20
0
E.
0 50 100 150 200 250
[HEPARIN] (ng/rnL)
FIG. 3. Thrombin inhibition by AT-I11 and heparin frac- tions obtained after gel filtration on a Sephadex G-75 column. A , fractions obtained as described in Fig. 2 were assayed for their ability to potentiate the inhibition of thrombin by AT-111. Heparins (100 ng/ml): LMW/NA-heparin fractions (0) or LMW/ase-heparin fractions (0) were assayed as described (Andrade-Gordon and Strick- land, 1986). Percent thrombin inhibition represents the percent of remaining thrombin activity in the presence of heparin and AT-I11 compared with that of thrombin activity with AT-I11 alone subtracted from 100. B, dose response of heparins from 0 to 200 ng/ml on the inactivation of thrombin by AT-111.0, HMW-heparin; 0, LMW/NA- heparin; A, fraction 20 from eluate of LMW/NA-heparin after Seph- adex G-75 chromatography (Fig. 2); 0, fraction 20 from the eluate of LMW/ase-heparin. Percent thrombin inhibition was measured as described above.
concentrations. For example, at concentrations of 200 ng/ml, all preparations inhibited thrombin by more than 70%.
Effect of Fractionated LMW-heparins on t-PA-mediated Actiuation of Plasminogen-As demonstrated previously HMW-heparin was able to enhance plasminogen activation by t-PA 3-fold (Figs. 1 and 4). In contrast, when using fraction 20 of both LMW/NA-heparin and LMW/ase-heparin, t-PA activity was not stimulated over the wide range of concentra- tions used (Fig. 4). These results, obtained with two inde- pendent LMW-heparins derived by chemical and enzymatic degradation methods, coupled with those above, demonstrate that the t-PA-stimulating activity of heparin is dependent on the size of the polysaccharide chain. The size requirement seems to be larger than that required for the co-binding of AT-111 and thrombin that is necessary for thrombin inhibi- tion. Therefore it is possible to prepare a LMW-heparin based on its molecular size that is capable of potentiating the inhibition of thrombin but that does not interact with the fibrinolytic system.
Binding of AT- 111 and Fibrinolytic Proteins to LM W-hepa- rin-To determine if AT-111, t-PA, u-PA, Glu-plasminogen, or Lys-plasminogen could interact with the LMW-heparin preparation, binding studies were performed using LMW/ NA-heparin covalently attached to a Sepharose matrix. The
15180 Low Molecular Weight Heparin and Plasminogen Activation
h
v 9 vr A-A WW/NA-H.pmrln FWICT. 20
0-0 HYW-Haporin
- LYW/NA-H.PO~~~
- 4 t z 07
0 10 20 30 40 50 [HEPARIN] (pg/mL)
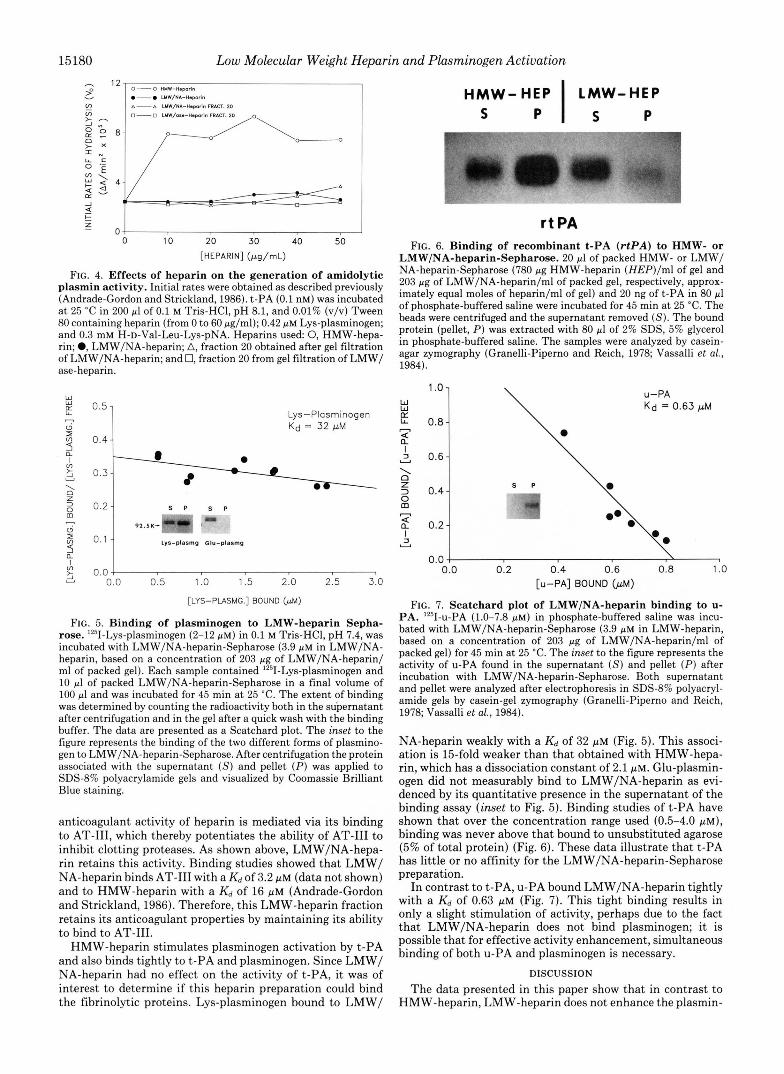
FIG. 4. Effects of heparin on the generation of amidolytic plasmin activity. Initial rates were obtained as described previously (Andrade-Gordon and Strickland, 1986). t-PA (0.1 nM) was incubated a t 25 "C in 200 pl of 0.1 M Tris-HC1, pH 8.1, and 0.01% (v/v) Tween 80 containing heparin (from 0 to 60 pg/ml); 0.42 pM Lys-plasminogen; and 0.3 mM H-D-Val-Leu-Lys-pNA. Heparins used 0, HMW-hepa- rin; 0, LMW/NA-heparin; A, fraction 20 obtained after gel filtration of LMW/NA-heparin; and 0, fraction 20 from gel filtration of LMW/ ase-heparin.
W
Lys-Plasminogen Kd = 32 pM
6 2 0.04 !
u 0.0 0.5 1.0 1.5 2.0 2.5 3.0
[LYS-PLASMG.] BOUND (pM)
FIG. 5. Binding of plasminogen to LMW-heparin Sepha- rose. '251-Lys-plasminogen (2-12 pM) in 0.1 M Tris-HCI, pH 7.4, was incubated with LMW/NA-heparin-Sepharose (3.9 p~ in LMW/NA- heparin, based on a concentration of 203 pg of LMW/NA-heparin/ ml of packed gel). Each sample contained 1251-Lys-plasminogen and 10 pi of packed LMW/NA-heparin-Sepharose in a final volume of 100 pi and was incubated for 45 min at 25 "C. The extent of binding was determined by counting the radioactivity both in the supernatant after centrifugation and in the gel after a quick wash with the binding buffer. The data are presented as a Scatchard plot. The inset to the figure represents the binding of the two different forms of plasmino- gen to LMW/NA-heparin-Sepharose. After centrifugation the protein associated with the supernatant ( S ) and pellet (P) was applied to SDSd% polyacrylamide gels and visualized by Coomassie Brilliant Blue staining.
anticoagulant activity of heparin is mediated via its binding to AT-111, which thereby potentiates the ability of AT-I11 to inhibit clotting proteases. As shown above, LMW/NA-hepa- rin retains this activity. Binding studies showed that LMW/ NA-heparin binds AT-I11 with a K d of 3.2 p~ (data not shown) and to HMW-heparin with a K d of 16 pM (Andrade-Gordon and Strickland, 1986). Therefore, this LMW-heparin fraction retains its anticoagulant properties by maintaining its ability to bind to AT-111.
HMW-heparin stimulates plasminogen activation by t-PA and also binds tightly to t-PA and plasminogen. Since LMW/ NA-heparin had no effect on the activity of t-PA, it was of interest to determine if this heparin preparation could bind the fibrinolytic proteins. Lys-plasminogen bound to LMW/
HMW- HEP LMW-HEP S P I s P
r t PA FIG. 6. Binding of recombinant t-PA (rtPA) to HMW- or
LMW/NA-heparin-Sepharose. 20 pl of packed HMW- or LMW/ NA-heparin-Sepharose (780 pg HMW-heparin (HEP)/ml of gel and 203 pg of LMW/NA-heparin/ml of packed gel, respectively, approx- imately equal moles of heparin/ml of gel) and 20 ng of t-PA in 80 pl of phosphate-buffered saline were incubated for 45 min a t 25 "C. The beads were centrifuged and the supernatant removed (S). The bound protein (pellet, P ) was extracted with 80 pl of 2% SDS, 5% glycerol in phosphate-buffered saline. The samples were analyzed by casein- agar zymography (Granelli-Piperno and Reich, 1978; Vassalli et al., 1984).
W W K L- 0.8 '.OI \ - 2 3 0.6 I
0.0 \ 0.0 0.2 0.4 0.6 0.8 1 .o
[u-PA] BOUND (pM)
FIG. 7. Scatchard plot of LMW/NA-heparin binding to u- PA. 1Z51-~-PA (1.0-7.8 p ~ ) in phosphate-buffered saline was incu- bated with LMW/NA-heparin-Sepharose (3.9 p~ in LMW-heparin, based on a concentration of 203 pg of LMW/NA-heparin/ml of packed gel) for 45 min a t 25 "C. The inset to the figure represents the activity of u-PA found in the supernatant ( S ) and pellet (P) after incubation with LMW/NA-heparin-Sepharose. Both supernatant and pellet were analyzed after electrophoresis in SDS-8% polyacryl- amide gels by casein-gel zymography (Granelli-Piperno and Reich, 1978; Vassalli et al., 1984).
NA-heparin weakly with a K d of 32 pM (Fig. 5). This associ- ation is 15-fold weaker than that obtained with HMW-hepa- rin, which has a dissociation constant of 2.1 PM. Glu-plasmin- ogen did not measurably bind to LMW/NA-heparin as evi- denced by its quantitative presence in the supernatant of the binding assay (inset to Fig. 5). Binding studies of t-PA have shown that over the concentration range used (0.5-4.0 p ~ ) , binding was never above that bound to unsubstituted agarose (5% of total protein) (Fig. 6). These data illustrate that t-PA has little or no affinity for the LMW/NA-heparin-Sepharose preparation.
In contrast to t-PA, u-PA bound LMW/NA-heparin tightly with a K d of 0.63 p~ (Fig. 7). This tight binding results in only a slight stimulation of activity, perhaps due to the fact that LMW/NA-heparin does not bind plasminogen; it is possible that for effective activity enhancement, simultaneous binding of both u-PA and plasminogen is necessary.
DISCUSSION
The data presented in this paper show that in contrast to HMW-heparin, LMW-heparin does not enhance the plasmin-

Low Molecular Weight Heparin and Plasminogen Activation 15181
generating activity of t-PA (Figs. L4 and 4). Moreover, unlike the HMW-heparin, the LMW/NA-heparin preparation does not bind t-PA or Glu-plasminogen and binds only weakly to Lys-plasminogen. Although it binds u-PA, the effect on its activity is not as great as with HMW-heparin (Fig. 1B). It should be noted that using a rabbit jugular vein thrombosis model, low molecular weight heparin fragments have been shown to enhance thrombolysis after injection of human t- PA (Stassen et al., 1987). The apparent discrepancy between these results and those reported here might be explained by differences: 1) between the heparin preparations used; 2 ) in the interactions of the mucopolysaccharide with rabbit plas- minogen; 3) related to endogenous rabbit t-PA.
Commercial heparin is complex and interacts with a wide variety of plasma proteins including clotting factors (Fujikawa et al., 1973), growth factors (Maciag et al., 1984; Shing et al., 1984), apolipoproteins (Shelburne and Quarfordt, 1977), and fibronectin (Stathakis and Mosesson, 1977). Some of these interactions are ionic in nature and involve a cationic region of the protein associating with the negatively charged groups on heparin. For others, including t-PA, the binding is less well understood. In addition to the different chemical moieties on heparin that could mediate its action, the molecular size can play a role. For example, a pentasaccharide is sufficient to catalyze factor Xa inhibition by AT-111, whereas this oligo- saccharide does not mediate thrombin inhibition by AT-I11 (Choay et al., 1983). This results has been interpreted as suggesting that a ternary complex (heparin-AT-111-thrombin) is necessary for acceleration of thrombin inhibition and that the pentasaccharide cannot accommodate both proteins. Our analysis of two heparin preparations also suggests that the size of the polysaccharide may be crucial; first, LMW-heparin prepared by two independent methods does not stimulate the activity of t-PA. The fact that the heparinase-depolymerized product behaves similarly to that obtained by nitrous acid treatment suggests that the formation of anhydrosugars (Shively and Conrad, 1976) but the latter procedure is not the cause of the loss of activity. Second, a size of 11,000 can catalyze inactivation of thrombin by AT-I11 but is inadequate for t-PA stimulation. If binding to both t-PA and plasminogen is required for heparin stimulation, a complex with only one of the components may not be sufficient.
Although other plasminogen activators such as streptoki- nase and u-PA can be used for thrombolytic therapy, t-PA has been reported to be more effective. The promise of t-PA lies primarily in one aspect of its enzymatic properties: under certain conditions, its ability to activate plasminogen is strongly enhanced by the presence of fibrin (Hoylaerts et al., 1982), the major protein component of blood clots.
In the presence of 1 unit/ml heparin, t-PA activates plas- minogen from 5- to 30-fold faster (depending on the assay conditions) than in the absence of the anticoagulant. Fur- thermore, heparin interferes with fibrin's ability to enhance the activation of plasminogen by t-PA. In fact, systemic proteolysis has been reported with the use of t-PA, leading in some cases to serious depletion of circulating clotting factors and hemorrhaging (Collen et al., 1984; TIMI Study Group, 1985; Verstraete et al., 1985; Verstraete and Collen, 1986; Braunwald et al., 1987). These harmful effects might be ex- plained by the influence of heparin on the activity of t-PA.
The concomitant presence of heparin and t-PA in thrombo- lytic therapy might have two deleterious consequences: 1) heparin could stimulate t-PA in the general circulation, caus- ing increased fibrin-independent destruction of blood com- ponents; and 2 ) heparin could inhibit binding of t-PA to fibrin, preventing the localization of the enzyme at the site of the clot.
In addition to the possible relevance of the properties of LMW-heparin for t-PA therapy, our results may bear on some of the complications associated with other uses of heparin. For example, if heparin interacts with endogenous t-PA or u- PA, systemic proteolysis might be promoted, and hemorrhagic complications could result from heparin's effect on the fibrin- olytic system rather than its effect on coagulation. Prepara- tions of LMW-heparin have been shown to have several advantages over HMW-heparin. Of special interest here is the fact that these preparations seem to cause less bleeding (Turpie et al., 1986; de Prost, 1986). A partial explanation of this advantage could stem from the lack of interaction of LMW-heparins with the fibrinolytic system.
REFERENCES Andrade-Gordon, P., and Strickland, S. (1986) Biochemistry 25,4033-4040 Braunwald, E., Knatterud, G. L., Passamani, E., Robertson, T. L., and Solomon,
R. (1987) J. Am. CoU. Cardiol. 10,970 Choay, J., Petitou, M., Lormeau, J.-C., Sinay, P., Casu, B., and Gatti, G. (1983)
Biochem. Biophys. Res. Commun. 116,492-499 Collen, D., Topol, E. J., Tiefenbrunn, A. J., Gold, H. K., Weisfeldt, M. L.,
Sohel, B. E., Leinbach, R. C., Brinker, J. A., Khaw, B. A., and Grosshard, B. W. (1984) Circuhtion 70,1012-1017
de Prost, D. (1986) Trends Pharmucol. Sci. 7,496-500 Deutsch, D. G., and Mertz, E. T. (1970) Science 170,1095-1096 Fe?, R. (1988) Biochem. J. 249,77-81 Fu~~kawa, K., Thompson, A. R. Legaz, M. E., Meyer, R. G., and Davie, E. W.
(1973) Biochemistry 12,49364945 Granelli-Piperno, A., and Reich, E. (1978) J. Ex Med 148,223-234 Hiiiik, M., Bjork, I., Hopwood, J., and Lindahl, 5. (1976) FEBS Lett. 66,90-
Hoylaerts, M., Rijken, D. C., Lijnen, H. R., and Collen, D. (1982) J. Biol. Chem.
Jaques, L. B., and Wollin, A. (1967) Can. J. Physiol. Pharmucol. 4 5 , 785-794 Linhardt, R. J., Grant, A., Cooney, C. L., and Langer, R. (1982) J. Biol. Chem.
Maciag, T., Mehlman, T., Friesel, R., and Schreiber, A. B. (1984) Science 2 2 5 ,
Markwell, M. A. K., and Fox, C. F. (1978) Biochemistry 17,4807-4817 Mosesson, M. W. (1962) Biochim. Biophys. Acta 67,204-213 Nesheim, M., Blackburn, M. N., Lawler, C. M., and Mann, K. G. (1986) J. Biol.
Piques, E-P., Stohr, H-A., and Heimburger, N. (1986) Thromb. Res. 42 , 797-
93
257,2912-2919
257,7310-7313
932-934
Chem. 261,3214-3221
xn7 SGiburne, F. A., and Quarfordt, S. H. (1977) J. Gin. Inuest. 60,944-950 Shing, Y., Folkman, J., Sullivan, R., Butterfield, C., Murray J., and Klagsbrun,
Shively J. E., and Conrad, H. E. (1976) Biochemistry 15,3932-3942 Smith, k'. K., Mallia, A. K., and Hermanson, G. T. (1980) Anal. Biochem. 109 ,
M. (1984) Science 223,1296-1299
466-473 Soeda, -S., Kakiki, M., Shimeno, H., and Nagamatsu, A. (1987) Biochim.
Bioohvs. Acta 91 6.279-287 -r I- ~~~
Stassen, J. M., Juhan-Vague, I., Alessi, M. C., De Cock, F., and Collen, D.
Stathakis, N. E., and Mosesson, M. W. (1977) J. Clin. Inuest. 60 , 855-873
Teien, A. N., Abildgaard, V., Hiiiik, M., and Lindabl, V. (1977) Thromb. Res. Strickland, S., and Beers, W. H. (1976) J. Biol. Chem. 251,5694-5702
, - . . - - . (1987) Thromb. Haemostasis 58,947-950
11.1n7-117 TIMI Study Group (1985) N . Engl. J . Med. 312,932-936 Turpie, A. G. G., Levin, M. N., Hirsh, J., Carter, C. J., Jay, R. M., Powers, P.
J., Andrew, M., Hull, R. D., and Gent, M. (1986) N . Engl. J. Med. 315,925-
", _I. ".
99q
v ~ s ~ h i , J.-D., Dayer, J.-M., Wohlwend, A,, and Belin, D. (1984) J . Exp. Med.
Verheijen, J. H., Mullaart, E, Chang, G. T. G., Kluft, C., and Wijngaards, G. 159,1653-1668
(1982) Thromb. Haemostasls 4 8 . 266-269 Verstraete, M., and Collen, D. (1986) B&d 6 7 , 1529-1541 Verstraete, M., Bernard, R., Bory, M., Brower, R. W., Collen D., De Bono, D.
J., Michels, H. R., Rutsch, W., Schartl, M., Schmidt, M., Uebis, R., and vo; P., Erbel, R., Huhmann, W., Lennane, R. J., Lubsen, J., Mkhey D., Meyer
Vinazzer, H., Sternberger, A,, Haas, S., and Bliimel, G. (1982) Thromb. Res. Essen, R. (1985) Lancet 1 , 842-847
27,341-352