Embed Size (px)
Citation preview

Journal of Immunogenetics (1975) 2,231-243.
ANTIBODY SYNTHESIS I N VITRO X I I . A C T I V I T Y O F R E C O N S T I T U T E D A N D H Y B R I D
R E C O M B I N E D R I B O S O M E S I N C E L L F R E E S Y N T H E S I S O F A N T I B O D Y *
D. J A C H E R T Z , H. TRACHSEL A N D ADA HOIDA-WERNER
Institut fur Hygiene und Medizinische Mikrobiologie der Universitat Bern, Switzerland
(Received 10 May 1975)
SUMMARY
In a cell free system derived from peripheral human leucocytes an informative RNA (i-RNA) was synthesized after antigenic stimulation with benzoyl-penicilloyl- formyl-L-lysine (BPoFLys). This i-RNA was used to synthesize antibody in a second cell free system. The ribosomes used in the antibody synthesizing system were derived from peripheral human leucocytes, from human lymphoblasts CCRF and from HeLa cells. Furthermore, hybrid recombined ribosomes from 60s HeLa-subunits and 45 S CCRF-subunits as well as from 45s HeLa- and 60s CCRF-subunits were used to synthesize antibody. Only ribosomes from peripheral human leucocytes, from CCRF-cells and hybrid recombined ribosomes from 60s HeLa- and 45s CCRF-subunits were able to synthesize antibody in the cell free system. Antibody synthesized in the cell free system was detected by phage neutralization test and by equilibrium dialysis. By these methods new markers could be demonstrated which are essential for the differentiation to an antibody synthesizing cell.
I N T R O D U C T I O N
Most immunogenetical markers described until now concern recognition mechanisms which are thought to be expressed on T-cells. They are linked to the histocompatibility antigens (Bailey, 1971 ; Bailey & Hoste, 1971 ; Bluestein et al., 1972; Ellman et QZ., 1970; Gasser et al., 1969, 1970,1971 ; Green et al., 1970, 1971 ; Guenther et al., 1972; Lieberman & Humphrey, 1971; Martin et al., 1971; McDevitt et al., 1965, 1967, 1968, 1969; Merryman & Maurer,
* Paper no. XI of this series is D. Jachertz, Uta Opitz, and H. G. Opitz (1 972) ‘Gene amplification in cell-free systems’ Zeitschriyt f i r Immunitatsforschung, 144,260.
Correspondence: Professor Dr D. Jachertz, Bern, Friedbuhl-Str. 51, Switzerland. 23 1

232 D. Jachertz, H. Trachsel and A . Hoida- Werner 1971, 1972; Mozes et al., 1969; Rathbun & Hildeman, 1970; Snell et al., 1967; Vaz et al., 1970; Vladutiu & Rose, 1971). Other immunogenetical markers are well known as allotypes in the Gm and the Inv system as well. Furthermore, it was demonstrated that the ability to synthesize antibody against certain antigens is genetically linked to certain allotypes (Blomberg et al., 1972; Wells et al., 1971). These observations allow a differentiation of (a) markers concerning regulatory mechanisms during the recognition of the antigen. These markers are suitable to elucidate single steps of the recognition of a certain determinant; (b) structural markers which are linked to a certain allotype and which are phenotypically expressed as the deficiency to synthesize antibody against a certain antigen. These markers are thought to be suitable to elucidate single steps of the synthesis of antibody.
Both types of markers are related to the immunological specificity. Concerning the unknown sequence of steps finally leading to the differentiation of an antibody producing cell one rather should use markers which are not related to immunological specificity but to im- munological competence.
In a previous publication (Jacherts, 1969) it could be demonstrated that ribosomes pre- pared from HeLa cells are not able to synthesize antibody in a cell free system after addition of a preparation of informative RNA (i-RNA). The activity to code for an antibody of such an i-RNA preparation could be demonstrated in another cell free system using ribosomes from peripheral leucocytes. Therefore, a new class of immunogenetic markers seems to be available which correspond to the differentiation of certain cellular organelles necessary for the synthesis of immunoglobulins. In the present publication we describe a distinct functional ability of ribosomes derived from immunocompetent cells the phenotypical expression of which is a condition sine qua non for the synthesis of an immunoglobulin.
Since a human marker related to differentiation of cells cannot be studied in a reasonable way by classical genetic techniques, such as crossing experiments or statistical analysis, in order to elucidate genetic linkage groups we used a new technique of genetic analysis in cell free systems. The basic idea of the genetic experiment, i.e. the recombination of different phenotypes is modified to a recombination of pre-existent subunits of phenotypical different cells on a subcellular level. The rationale of our experiments was the following:
Step 1 A cell free system derived from human peripheral leucocytes was used to recognize a
certain antigenic determinant. The first detectable product of antigenic recognition in this cell free system is an i-RNA. The biological and physicochemical properties of this i-RNA have been described elsewhere (Jachertz, 1973).
Step 2 Ribosomes competent to synthesize immunoglobulin were derived from peripheral
leucocytes or from human lymphoblastic leukaemia cells CCRF-SB. Ribosomes incompetent for the synthesis of immunoglobulins were isolated from HeLa cells. So we came to three types of human ribosomes : (a) ribosomes derived from a mixed cell population which is able to recognize antigens and which is able to synthesize immunoglobulins; (b) ribosomes derived from a uniform cell population which is not able to recognize the antigen but the ribosomes of which are able to synthesize antibody; (c) ribosomes which are derived from cells which are not able to recognize the antigen and which are not able to synthesize imrnuno- globulins.

Antibody synthesis in vitro. XI1 233 Step 3
All types of ribosomes were dissociated in 45S- and 60s-subunits and reconstituted to monosomes. After reconstitution the biological activity of the monosomes to synthesize protein was tested.
Step 4 The 45S- and 60S-subunits of competent and incompetent ribosomes were recombined
and the ability of the hybrid recombined monosomes to synthesize antibody was tested in a cell free system.
It could be demonstrated that the 60s-subunit of ribosomes may be derived from any cell, immunocompetent or not, but the 45S-subunit must be derived from an immunocompetent cell which is able to synthesize antibody. In other words, a cell which is synthesizing antibody must have gone through a distinct differentiation process to be able to express a certain functional ability on the 45s-subunit of ribosomes which is needed for the correct synthesis of antibody.
M A T E R I A L A N D M E T H O D S
Cells (a) HeLa cells, originally obtained from Microbiological Associates, Bethesda (USA),
were cultured at 37°C in Roux flasks using MEM-Eagle medium fortified with 10% calf serum. After an incubation of 3 days, cells were washed three times with Hanks’ solution. Using a solution of 0.2 % trypsin in 0.02 M EDTA, cells were removed from the glass and sedimented at 150 x g for 5 min. The pellet was resuspended in 2-0 ml RSB (0.01 M Tris-HC1, pH 7.4, 0.01 M NaCl, 0.0015 M MgCIJ. In order to label the ribosomal RNA, cells were cultured for 24 hr in a medium containing 2.5 pCi/ml 5-3H Uridine (specific activity 23 Ci/mmol-Radiochemical Centre, Amersham).
(b) CCRF-SB cells, obtained from the American Type Culture Collection, were cultured in Roux flasks using MEM-Eagle medium modified for suspension culture at 37°C. Harvest- ing, washing, resuspension and labelling were the same as for HeLa cells.
(c) Human peripheral leucocytes were prepared from buffy coat of freshly withdrawn and centrifuged blood conserves as described (Jacherts, 1969).
Cell free system (a) DNA was isolated from cells using a phenol natrium-dodecylsulphate method as
described (Jachertz et af., 1972). The DNA preparation should have a quotient of 260/280 O.D. reading between 1.95 and 2.0 and a 2601240 O.D. reading between 1.9 and 1.95. The average molecular size of the DNA preparation should be lo8 daltons, and the hyper- chromicity effect after heating to 100°C for 30 min should be at least 20 %.
(b) The pH,-fraction, which is another component of the antigen recognizing cell free system (step l), as well as a component of the antibody synthesizing cell free system (step 4), is prepared from a 100,OOO x g supernatant of an extract obtained by freezing and thawing, ten times and mechanically destroying the cells mentioned above by adjustment of the pH from 8.0 to 5.0. The details of the preparation of a pH5-fraction have been described (Jachertz et al., 1972).

234 D. Jachertz, H. Trachsel and A. Hoida- Werner (c) Ribosomes from HeLa cells, from CCRF-SB cells and from peripheral human leuco-
cytes were isolated by sedimentation through a sucrose gradient. At first 2 . lo7 cells in a volume of 2.0 ml RSB-buffer were incubated for 30 min at 0°C and then homogenized by thirty strokes in a Dounce homogenizer A at 0°C. Cellular debris was sedimented at 1000 x g for 10 rnin at 0°C. A volume of 0.4 ml of this supernatant was layered on 4.6 ml of a linear sucrose gradient ranging from 5 to 30% sucrose in 0.02 M Tris HC1, pH 7.8, 0.04 M KCl, 0.005 M MgCI, and 0.002 M mercaptoethanol, and sedimented at 100,000 x g for 210 min at 4°C. After this, the centrifuge tubes were punctured at the bottom and the effluent was collected in fractions of 0-2 ml. To each fraction 0.6 ml of Tris buffer were added and the optical density was recorded in a Zeiss Spectrophotometer PMQ I1 (Trachsel, 1973). In some cases ribosomes were labelled by incorporation of 3H-Uridine. The fractions containing ribosomes were detected by scintillation counting (Trachsel, 1973). Subunits of ribosomes were obtained in a single step by sedimentation of the cell homogenate through a gradient ranging from 5 to 30 % sucrose in Tris buffer containing 0.3 M KCl in an SW 50.1 rotor at 4°C at 100,000 x g for 210 min. In other cases subunits of ribosomes were sedimented in an SW 25.1 rotor at 4°C at 57,000 x g for 16.5 hr using a gradient ranging from 15 to 40 % sucrose in Tris buffer containing 0-3 M KCl.
Reconstitution of subunits and recombination of hybrid ribosomes was performed by mixing the separated subunits and the adjustment of the potassium chloride concentration to 0.04 M by dialysis against Tris buffer (Trachsel, 1973; Falvey & Staehelin, 1970). The reconstituted and recombined ribosomes were isolated by a final sedimentation through a sucrose gradient (Trachsel, 1973 ; Falvey & Staehelin, 1970).
Biological activity of the reconstituted and recombined ribosomes tested in cell free systems
System I. Synthesis of i-RNA. In order to provide a biologically active messenger RNA for the synthesis of antibody with ribosomes, reconstituted ribosomes and recombined hybrid ribosomes in a first step an i-RNA was synthesized which codes for antibody against the benzoylpenicilloyl-determinant. Therefore, a cell free system was stimulated with BPoFLys* as antigen. This cell free system contained DNA, derived from HeLa cells, in a concentration of 2 . pg/ml, pH,-fraction in an optimal concentration of protein corre- sponding to 0.3 pg N,/ml and lo5 molecules/ml BPoFLys as antigen. Furthermore, 3H UTP lo-' M (sp. act. 40 Ci/mmol-Radiochemical Centre, Amersham) and UTP, CTP, GTP and ATP M each were added. After incubation of aliquots of 0.6 ml at 37"C, for times ranging from 3 min to 5 min 30 sec, synthesis was stopped and RNA extracted by addition of 0-5 ml Kirby I and 0-5 ml Kirby I1 solution. After mixing the aqueous phase was removed. The optimal amount of 3H UMP incorporation was reached after an in- cubation for 210 sec at 37°C. After this time an aliquot of 0.1 ml of the stimulated system contained 2882 cpm of TCA insoluble material in contrast to the unstimulated control in which 270 cpm were found. The i-RNA synthesized was isolated by sedimentation through a 5.0 ml sucrose gradient ranging from 5 to 25 % for 2 hr at 100,000 x g and 0°C. Fractions of 0.3 ml were collected and aliquots of 0.1 ml of each fraction were counted in a liquid scintillation counter. Fractions with increased radioactivity in comparison to the control were pooled and resedimented under conditions as described (Jachertz 1973; Jachertz et al., 1972). By this procedure 0.3 ml of a purified I-RNA preparation was obtained, which contained
* BPoFLys = benzoyl-penicilloyl-formyl-~-lysine which was a gift from Professor Dr A. de Weck, Berne.

Antibody synthesis in vitro. XII 235 12,938 cpm of 'H UMP incorporated, which corresponds to 5.10* molecules of i-RNA or pg i-RNA on the basis of an average molecular weight of 2.5. lo6 of the i-RNA and a specific activity of 'H UTP in the pool of 4 Ci/mmol. The corresponding preparation derived from a non-stimulated cell free system shows only 1.743 cpm of incorporated 3H UMP. This is a background level of unspecific stimulation and/or incorporation of 3H UMP,
System 2. Poly-U-directed synthesis of polyphenylalanine. A volume of 0.3 ml contained 0.02 M Tris-HC1 (pH 7 .Q 0.1 M KCl, 0.01 M MgCl, and 0.005 M 2-Mercaptoethanol, 50 pg polyuridylic acid (sodium salt, mol. wt 1.1. lo5, Calbiochem), 10-50 pg ribosomes or sub- units of ribosomes, 0.2 mM GTP (Serva, Heidelberg, Western Germany), 5 mM creatin- phosphate (sodium salt, Serva Heidelberg), 6 jig creatinkinase (Fluka, Buchs), 2-5 pCi ~-3-~H-phenylalanine (sp. act. 1 Ci/mmol-Radiochemical Centre, Amersham) ; and 250 pg protein of S-100 which is a supernatant of an extract derived from lymphoblast cells CCRF after centrifugation at 100,000 x g in an SW 50 L rotor at 0°C for 3 hr. The details of this system have been described (Trachsel, 1973). The amount of polyphenylalanine synthesized was estimated by TCA precipitation of polyphenylalanine and determination of 3H labelled phenylalanine incorporated in the product.
System 3. Synthesis of antibody. A volume of 1 ml of 0.1 M Tris-HCl buffer, pH 8.0, with 0.04 M KCI and 0.05 M MgCl, contains the following components: i-RNA in an optimal concentration, which is usually lo6 molecules of i-RNA. This i-RNA codes for antibody against benzoyl-penicilloyl (BPo) determinant; pH,-fraction in an optimal concentration which is equivalent to 0.6 pg N,/ml; 2. lo-'' mol 'H-~-leucine (sp. act. 50 Ci/mmol) (-2. lo7 dpm); 3. lod1' mol 3H-~-valine (sp. act. 29 Ci/mmol-Radiochemical Centre, Amersham) (-1. lo7 dpm); the reconstituted or hybrid recombined monosomes in a con- centration corresponding to 10-1 pg nitrogen/ml.
Addition of triphosphates, amino acids and an energy generating system does not result in a higher rate of antibody synthesis. The amount of antibody synthesized in the cell free system was detected by TCA precipitation of the protein synthesized, by determination of specific phage neutralization using penicilloyl-labelled T,-phage as described (Jachertz et al., 1972) and by equilibrium dialysis of the radioactive labelled antibody synthesized.
Equilibrium dialysis. For equilibrium dialysis cellulose acetate filters (Millipore Corporation) with pore diameter of 0.45 pm were used. The BPo determinant was fixed on human serum albumin (HSA)-sepharose, a gift from Dr W. Schneider, Berne, Institut fur klinische Immunologic.* In microchambers as described by MacDonald (1971) one side was filled with sepharose without determinant (= 0 side), the other side with BPo-HSA- sepharose (= 0 side). In both sides antibody labelled with 3H alanine and 3H leucine was added. After incubation for 8 hr at 37°C from each side of the microchamber an aliquot of 0-05 ml was withdrawn and the redistribution of radioactivity was estimated by scintillation counting.
Since the pores of the filter can be passed by antibody the following calculation is used: cpm at the 0 side corresponds to the amount of free antibody; cpm at the 0 side corresponds to the amount of free and bound antibody; cpm of 0 plus @ side corresponds to the total amount of antibody; cpm @ side minus 0 side corresponds to the amount of bound anti- body.
The relative amount of bound antibody is given as a percentage of the total amount of antibody present in the chamber.
* We thank Dr W. Schneider very much for the generous gift of BFo-HSA-sepharose,

236 D. Juchertz, H. Trachsel and A. Hoidu- Werner Buffers: RSB, 0.01 M Tris-HC1, pH 7-4, 0.01 M NaCI, 0.0015 M MgClz; Tris A, 0.02 M
Tris-HCl, pH 743,0.04 M KC1,0.005 M MgCl,; Tns B, 0-02 M Tris-HC1, pH 7-8,0.3 M KCl, 0-005 M MgCl,, 0-002 M mercaptoethanol; Kirby I, 50.0 g phenol (fresh distiIlate), 7-0 ml m-cresol (fresh distillate), 0.05 g 8-hydroxychinolin, 5.5 ml A.d.; Kirby 11, 1.0 g Na- dodecylsulphate, 5.88 g PAS-Na-salt, ad 100 ml A.d.
Glassware: All glassware used has to be free of RNase and free of antigen.
RESULTS
Isolation of ribosomes.fiom peripheral leucocytes, LTCC cells and HeLa cells Washed cells were disintegrated by freezing and thawing or by treatment in a Dounce
homogenisator. Ribosomes were isolated by sedimentation in a sucrose gradient as described in Materials and Methods. The sedimentation profiles of such a preparation of ribosomes are demonstrated in Fig. 1. Ribosomes from peripheral leucocytes were prepared in a similar manner without labelling.
The ribosome content of each fraction was estimated by optimal density reading at 260 nm.
I 1
* !20b I P 10
5 Fraction no.
FIG. 1. Isolation of monosomes derived from HeLa cells and CCRF lymphoblasts. The cells were cultured for 24 hr in 3H-Uridine containing medium as described in Materials
and Methods. After 30 min incubation at 0°C in hypotonic RSB buffer cells were homogenized and nuclei sedimented. A volume of 0.3 ml of the clear homogenate was layered on a linear sucrose gradient (4-6 ml, ranging from 5 % to 30% sucrose in Tris buffer A, KCI O W M), sedimentation in rotor SW 50.1 was for 210 min at 4°C and 100,OOO x g. Fractions of 160 pl were made and the content of ribosomes was estimated by scintillation counting. Sedimentation from the right to the left. S-values calculations according to Strohmaier & Mussgay, 1959. (a) = HeLa cell ribosomes; (b) =ribosomes of CCRF lymphoblasts.
Isolation of subunits of HeLa- and CCRF-ribosomes Subunits of ribosomes derived from HeLa cells or CCRF lymphoblasts were obtained
by a single step procedure. The homogenate of the correspondent cells was layered on a sucrose gradient in Tris buffer containing 0-3 M KC1. Under these conditions the ribosomes dissociate during sedimentation. This is demonstrated in Fig. 2.
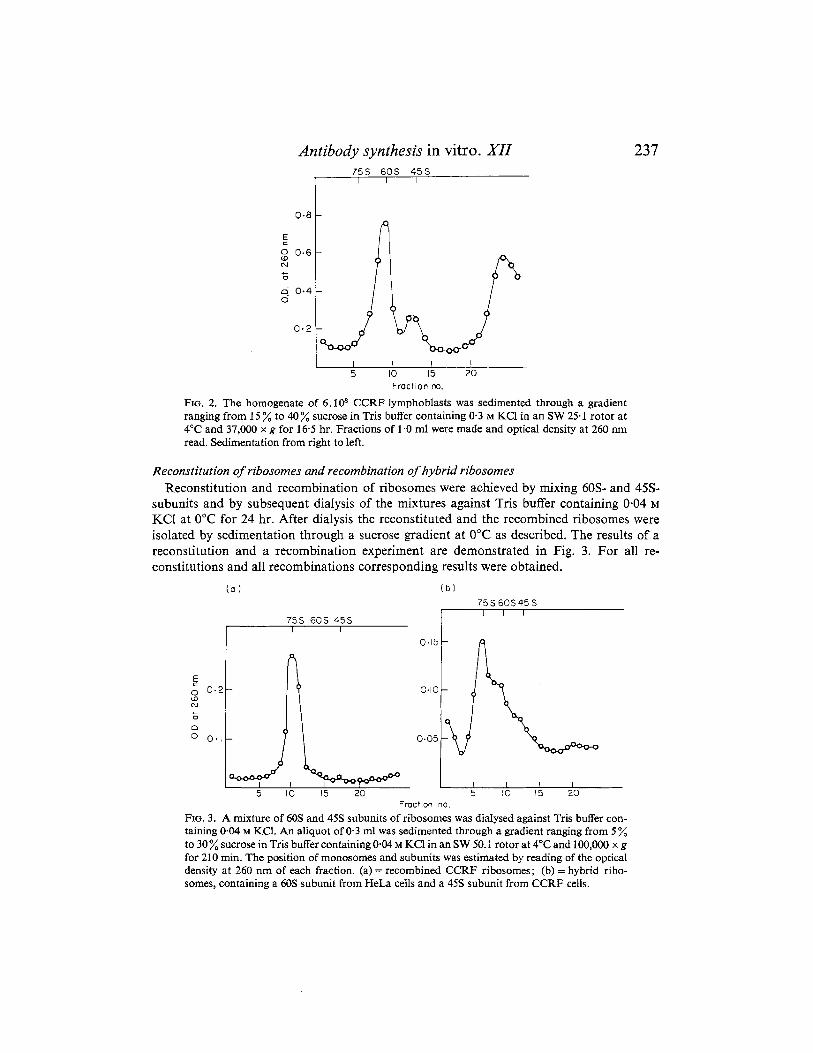
Antibody synthesis in vitro. XI1 7 5 s 60s 45s
1 I I I
237
E 0.6
N
o L
0: 0.4 0
0.2
L
L ! I I I 5 10 15 20
Fract ion no.
FIG. 2. The homogenate of 6.108 CCRF lymphoblasts was sedimented through a gradient ranging from 15 % to 40% sucrose in Tris buffer containing 0 3 M KCl in an SW 25.1 rotor at 4°C and 37,000 x g for 16-5 hr. Fractions of 1 .O ml were made and optical density at 260 nm read. Sedimentation from right to left.
Reconstitution of ribosomes and recombination of hybrid ribosomes Reconstitution and recombination of ribosomes were achieved by mixing 60s- and 45s-
subunits and by subsequent dialysis of the mixtures against Tris buffer containing 0.04 M KCI at 0°C for 24 hr. After dialysis the reconstituted and the recombined ribosomes were isolated by sedimentation through a sucrose gradient at 0°C as described. The results of a reconstitution and a recombination experiment are demonstrated in Fig. 3. For all re- constitutions and all recombinations corresponding results were obtained.
( a I ( b l
755 60s 455 I l l
0.1:
0.lC
0.0:
5 10 15 20
7 5 s 60s 45 S I l l
I ! I I 5 IC 15 20
Fraction no
FIG. 3. A mixture of 60s and 45s subunits of ribosomes was dialysed against Tris buffer con- taining 0.04 M KCl. An aliquot of 0.3 ml was sedimented through a gradient ranging from 5 % to 30% sucrose in Tris buffer containing 0.04 M KC1 in an SW 50.1 rotor at 4°C and 100,OOO x g for 210 min. The position of monosomes and subunits was estimated by reading of the optical density at 260 nm of each fraction. (a) = recombined CCRF ribosomes; (b) = hybrid ribo- somes, containing a 605 subunit from HeLa cells and a 45s subunit from CCRF cells.

238 D. Jachmtz, H. Trachsel and A . Hoida- Werner Activity of reconstitufed and recombined ribosomes in a poly( U) system
The polyphenylalanine synthesizing system contained poly(U), ribosomes (reconstituted or recombined ribosomes), HS-100-system and 3H phenylalanine in concentrations as descri- bed in Materials and Methods. After incubation for 30 min at 37"C, synthesis was stopped by addition of TCA and the amount of radioactivity incorporated in polyphenylalanine was estimated by scintillation counting. The results of such an experiment are summarized in Table 1. This shows that all reconstituted and all recombined ribosomes are able to synthesize polyphenylalanine. The most active ribosome is a recombinant between the HeLa 45s subunit and the CCRF 60s subunit. The background level of incorporation of 3H phenyl- alanine in phenylalanine with subunits of ribosomes is also mentioned in Table 1.
TABLE 1. Synthesis of polyphenylalanine in a cell free system using different types of reconsti- tuted and hybrid recombined ribosomes
Ribosomes Activity
Incorporation in %
CCRF ribosomes 60s + 45s (reconstituted monosomes) 2100 100
HeLa ribosomes 60s + 45s (reconstituted ribosomes) 11 67,926,782 53,44,37 recombined monosomes 60s (HeLa) + 45s (CCRF) 1372,951,615 65,45,29 recombined monosomes 60s (CCRF) + 45s (HeLa) 1590,1310,1140 75, 62, 52 CCRF-60s subunit 265 12,5
0 CCRF-45S subunit - HeLa-60s subunit 236 11
0 HeLa-45S subunit -
Samples of a volume of 0.15 ml Tris buffer containing 250pg S-lo0 protein, 15 pg monosomes or 10-1 1 pg 60s and 4-5 pg 455 subunits, 50 pg polyuridylic acid 0.2 lll~ GTP, 5 mM weatin- phosphate, 6 pg creatin-kinase and 2 5 pCi 3H-~-3-phenylalanine (sp. act. 1 Ci/mmol) were incubated for 30 min. at 37'C. Polyphenylalanine was precipitated and incorporation of 3H-phenylalanine estimated as described in Materials and Methods.
Biological activity of reconstituted and recombined ribosomes in an antibody synthesizing cell free system
In order to test the biological activity of the different reconstituted and the different recombined ribosomes a cell free system containing i-RNA, pH,-fraction and the respective monosomes was used as described in Materials and Methods. The antibody synthesized by the different reconstituted and recombined ribosomes was estimated by scintillation counting, by phage neutralization test and by equilibrium dialysis. This antibody is predominantly of the 19s-type (Jachertz, 1973). The results of such an experiment are summarized in Table 2. This shows a significant phage neutralization only after synthesis using reconstituted CCRF monosomes or recombined monosomes from the CCRF 45s subunit and the HeLa 60s subunit. The reconstituted HeLa ribosomes and the recombined monosomes from 45s HeLa subunit and 605 CCRF subunit are able to synthesize protein as in the poly(U) system but they are not able to synthesize phage neutralizing antibody. The corresponding observation is made in the equilibrium dialysis experiment.

TA
BL
E 2. C
ell f
ree
synt
hesi
s of
ant
ibod
y w
ith d
iffe
rent
reco
nstit
uted
and
hyb
rid
reco
mbi
ned
ribo
som
es. N
eutr
aliz
atio
n of
pen
icill
oyl-l
abel
led
T4-
phag
e pa
rtic
les a
nd fi
xatio
n of
ant
ibod
y on
benzoyl-penililloyl-HSA-sepharose in e
quili
briu
m d
ialy
sis
Equ
ilibr
ium
dia
lysi
s cp
m in
corp
orat
ed,
Rec
ipro
cal d
ilutio
n ca
lcul
ated
for
with
>3c
~ sign
ifica
nt
Tot
al
BPo
-bou
nd
% an
tibod
y 0.
6 rn
l ass
ay v
olum
e ph
age
neut
raliz
atio
n'
@ s
ide
0 sid
e an
tibod
y an
tibod
y bo
und
Leuc
ocyt
e-rib
osom
es
(mon
osom
es)
CC
RF-
ribo
som
es
(rec
onst
itute
d m
onos
omes
)
2132
2910
1000
16
70
874
2544
79
6 31
11
86
520
1705
66
5 39
10
00
1132
72
3 18
55
409
22
1108
72
3 18
31
385
21
410
206
61 6
204
33
HeL
a-ri
boso
mes
43
92
-
252
241
499
5 1
(rec
onst
itute
d m
onos
omes
) 36
7 32
6 69
3 41
5.
9 31
0 27
7 58
7 33
5.
6 R
ecom
bine
d rib
osom
es
4032
H
60S-
HeL
a L
45S-
CC
RF
3000
85
0 16
8 10
18
682
67
819
210
1029
60
9 59
R
ecom
bine
d rib
osom
es
5178
-
Leu
cocy
te ri
boso
mes
in c
ell f
ree
756
-
H45
S-H
eLa
L60
S-C
CR
F
syst
em w
ithou
t i-R
NA
BPo
FLys
In a
vol
ume o
f 0.
6 m
l ca
lo6 m
olec
ules
of i
-RN
A B
PoFL
ys c
odin
g fo
r ant
ibod
y ag
ains
t the
benzoylpenicilloyl-determinant w
ere
incu
bate
d w
ith d
iffe
rent
ty
pes
of r
econ
stitu
ted
and
hybr
id r
ecom
bine
d ri
boso
mes
pg
nitr
ogen
lml)
, pH
,-fra
ctio
n de
rived
fro
m p
erip
hera
l leu
cocy
tes
(0.6
pg
nitr
ogen
lml)
, 3H
- va
line a
nd 3
H-le
ucin
e m
of e
ach
(sp.
act
. 20
Ci/m
mol
, Rad
ioch
emic
al C
entr
e, A
mer
sham
) for
45
min
at
37°C
. Aft
er th
is 0
.1 m
l of
eac
h sa
mpl
e w
as
prec
ipita
ted
with
TC
A a
nd p
rote
in s
ynth
esis
estim
ated
by
scin
tilla
tion c
ount
ing,
and
0.1
ml o
f eac
h sa
mpl
e was
dilu
ted
and
used
for p
hage
neu
tral
izat
ion
test
as
des
crib
ed. F
rom
sam
ples
indi
cate
d (1
-4)
a vo
lum
e of
0.3
rnl w
as p
ut o
n a
grad
ient
rang
ing
from
5 %
to 3
0% su
cros
e and
sed
imen
ted
in a
n S
W 5
0.1
roto
r (S
pinc
o) a
t 100
,OO
O x
g fo
r 16 h
r. A
fter
this
cen
trif
ugat
ion f
ract
ions
of 0
.4 m
l wer
e co
llect
ed. F
rom
eac
h fr
actio
n 0.
1 m
l wer
e us
ed f
or e
stim
atio
n of
inc
orpo
ra-
tion
of 3
H-l
abel
led a
min
o ac
ids
by s
cint
illat
ion
coun
ting.
The
frac
tions
of
the
19s r
egio
n w
ere
pool
ed a
nd u
sed
for
equi
libri
um d
ialy
sis a
gain
st B
PoFL
ys-
HSA
-sep
haro
se. T
he c
alcu
latio
n of
fre
e an
d bo
und
antib
ody
was
mad
e as
des
crib
ed in
Mat
eria
l and
Met
hods
.

240 D . Jacherts, H . Trachsel and A. Hoida- Werner
DISCUSSION
In the experiments described, we used different types of ribosomes to synthesize antibody: original monosomes, reconstituted monosomes and hybrid recombined monosomes. All of these exhibit comparable synthetic activity as far as the incorporation of tritium labelled amino acids is concerned. The two synthesizing systems used, i.e. the poly(U) system and the i-RNA dependent system, are suitable for synthesizing protein with different reconstituted or hybrid recombined ribosomes. However, these two systems are not comparable as far as the concentration of ribosomes is concerned. The poly-U-directed system needs a con- centration of 100-150 pg/ml of monosomes; in the i-RNA BPoFLys-directed system the optimal dose of the monosomes is in the area of 1 pg/ml. Other components, such as the pH,-fraction in the antibody synthesizing system, the correspondent S-100 protein fraction in the poly-U system, the concentration of the i-RNA in the antibody synthesizing system and the polyuridylic acid in the poly-U system, differ in the optimal concentration which is needed for synthesis of protein or in biological activity in several orders of magnitude. In this respect, the i-RNA dependent system is not only a system which is able to synthesize a biologically meaningful protein but also a system which exhibits synthetic activity which is much higher than the poly-U dependent system.
Besides the total incorporation of amino acids in protein, some other points should be discussed in connection with the antibody synthesizing system. For the detection of antibody activity we used a penicilloyl labelled T4 phage. Antibody activity against the penicilloyl determinant is able to neutralize the phage particle. However, this neutralization reaction is, as all antibody-antigen reactions, a reversible process, the equilibrium of which is dependent on concentration of antibody and antigen and furthermore, on the avidity of the antibody molecules. Since the cell free system synthesizes only antibodies with relatively low avidity, the competition between the neutralization reaction of antibody with the phage particle and the receptor dependent adsorption of the phage particle to the host bacterium is remark- able. This is the main reason why the maximum values of neutralization do not reach the high degree which is seen in hyperimmune sera prepared against phage particles. On the other hand, this phage neutralization test is a very sensitive one and minimal amounts of antibody may be detected, especially if statistical significance is assured by sufficient repetitions of single experimental points. Since the cell free system becomes exhausted as regards energy and information, the product of the i-RNA coded synthesis is not a homogeneous population of antibody molecules. Different stages of incompletely terminated translation processes give a population of many different molecules which may be precursors of the final product of antibody synthesis. Among these, molecules with specific binding to the determinant, and other molecules without the capability to fix the BPoFLys determinant, may be expected. In this respect, different aspects of antibody synthesis are tested, whether synthesis is estimated by incorporation of tritium labelled acid (TCA precipitation), whether the phage neutraliza- tion test is used or the binding of antibody to the BPo determinant is estimated by equilibrium dialysis. The results of the equilibrium dialysis show that the antibody molecules purified by gradient centrifugation represent a population not only of molecules with a homogeneous sedimentation velocity but also a population with a relatively high avidity of fixation to the BPo determinant. This means that the translation process exhibits a high fidelity and that the i-RNA preparation used must be a very homogeneous one in respect to information content.

Antibody synthesis in vitro. XII 241 As previously published, the only cell population used in these experiments which is able
to recognize antigens is the population of peripheral leucocytes. Neither the CCRF cells nor the HeLa cells are able to recognize antigens. The ribosomes of peripheral leucocytes and CCRF cells are able to synthesize antibody in contrast to ribosomes of HeLacells which arenot able to do so. Consequently, the S-100 protein or the pH,-fraction derived from peripheral leucocytes is able to recognize antigens and to cooperate in antibody synthesis in cell free systems as well. The correspondent preparations derived from CCRF cells are only able to cooperate in the antibody synthesis and are not able to recognize antigens. Finally, the S-100 protein and the pH,-fraction derived from HeLa cells are neither able to recognize antigens nor able to synthesize antibody. However, all preparations are able to incorporate tritium labelled amino acid in TCA precipitable material (Jachertz, 1969). Since pH,-fraction derived from peripheral leucocytes cooperates with a hybrid recombined monosome CCRF45S- and HeLa 60s-subunit in antibody synthesis, a distinct biological activity should be postulated on the 45s subunit of the CCRF ribosome. On the other hand, it also may be postulated that the pH,-fractionderived from peripheral Ieucocytes contains a soluble factor (or factors) which is necessary for the expression of the above mentioned activity on the 45s subunit of the ribo- some or which cooperates with the ribosome or with the i-RNA in a still unknown reaction.
Since the genes for the variable part, and the genes for the constant part, of the light chain and the heavy chain of the antibody are thought to be situated on different loci of the genome, a recombination process must be postulated either on the level of the genome or the level of messenger RNA molecules or on the level of the translation process. A recombination process on the level of the genome needs a corollary of ad hoc hypotheses which cannot be tested experimentally until now. A recombination on the level of the messenger RNA mole- cule or on the level of the i-RNA has been described (Jacherts, 1968). This recombination process may be dependent on one or several factors present in the pH,-fraction derived from peripheral leucocytes or CCRF cells. The target size and the physico-chemical properties of the RNA indicate a rather high molecular i-RNA with an information content sufficient for variable and constant part of light and heavy chains, i.e. for the entire half molecule of antibody. The fact that i-RNA can be translated correctly only by ribosomes of peripheral leucocytes, CCRF cells and the HeLa 60s-CCRF45S hybrid ribosomes, is a challenge for further exploration of the translation process leading to the antibody molecule. In this context the possible recombination of informative subunits of the i-RNA and/or charac- teristic translational events for antibody synthesis should be mentioned.
R E F E R E N C E S
BAILEY, D.W. (1971) Allelic forms of a gene controlling the female immune response to the male antigen in mice. Transplantation, 11, 426.
BWY, D.W. & HOSTE, J. (1971) A gene governing the female immune response to the male antigen in mice. Transplantation, 11, 404.
BLOMFJERG, B., GEKELEV, W.R. & WEIGERT, W. (1972) Genetics of the antibody response to dextrans in mice. Science, 177,178.
BLUESTEIN, J.G., GREEN, I., MAURER, P.H. & BENACERRAF, B. (1972) Specific immune response genes of the guinea pig. V. Influence of the GA and GT immune response genes on the specificity of cellular and humoral immune response to a terpolymer of L-glutamic acid, L-alanine and r-tyrosine. Journal of Experimental Medicine, 135, 98.

242 D. Jachertz, H . Trachsel and A. Hoida- Werner ELLMAN, L., GREEN, I., MAR^, W.J. & BENACEBRAF, B. (1970) Linkage between the poly-L-Lysine gene
and the locus controlling the major hisrocompatibility antigens in strain 2 guinea pigs. Proceedings of the National Academy of Science, United States of America, 66,322.
FALVEY, A.K. & STAEHELIN, T. (1970) Structure and function of mammalian ribosomes. I. Isolation and characterization of active liver ribosomal subunits. Journal of Molecular Biology, 53,1.
GASSER, D.L. (1969) Genetic control of the immune response in mice. I. Segregation data and localization to the fifth linkage group of a gene affecting antibody production. Journal of Immunology, 103, 66.
GASSER, D.L. (1970) Genetic control of the immune response in mice. 11. A two-way selection experiment for specific immunologic competence. Journal of Immunology, 105,908.
GASSER, D.L. & SILVERS, W.L. (1971) Genetic control of the immune response in mice. nI. An association between H-2 type and reaction to H-Y. Journal of Immunology, 106, 875.
GREEN, I. & BENACERRAF, B. (1971) Genetic control of immune responsiveness to limiting doses of proteins and hapten protein conjugates in guinea pigs. Journal of Immunology, 107,374.
GREEN, I., INMAN, J.K. & BENACERRAF, B. (1970) Genetic control of the immune response of guinea pigs to limiting doses of bovine serum albumin: relationship to the poly-L-lysine gene. Proceedings of the National Academy of Sciences, United States of America, 66,1267.
GUENTHER, E., RUEDE, E. & STARK, 0. (1972) Antibody response in rats to the synthetic polypeptide (T, G)- A-L genetically linked to the major histocompatibility system. European Journalof Immunology, 2,151.
JACHERTZ, D. (1968) Antikorpersynthese in vitro. VIII. ober die Wirkung der informatorischen RNS- Molekiile auf die Milzzellkultur. Zeitschrvt f i r medizinische Mikrobiologie und Zmmunologie, 154, 1.
JACHERTZ, D. (1969) Antikorpersynthese in vitro. X. Information zur Antikorpersynthese in der DNS von nicht-kompetenten Zellen. Zeitschr$t fiir medizinische Mikrobiologie und Zmmunologie, 154, 300.
JACHERTZ, D. (1973) Flow of information and gene activation during antibody synthesis. Annals of the New York Academy of Sciences, 207, 122.
JACHERTZ, D., OPITZ, U. & OPITZ, H.-G. (1972) Gene amplification in cell-free systems. Zeitschrift frir Immunitatsforschung und experimentelle Therapie, 144,260.
LIEBERMAN, R. & HUMPHREY, W. JR. (1971) Association of H-2 types with genetic control of immune res- ponsiveness to IgA allotypes in the mouse. Proceedings of the National Academy of Science, United States of America, 68,2510.
MACDONALD, R. (1971) In: Methods in Immunology and Immunochemistry, Vol. 111 (Academic Press, New York & London).
MARTIN, T.E., ROLLESTON, F.S., Low, R.B. & WOOL, I.G. (1969) Dissociation and reassociation of skeletal muscle ribosomes. Journal of molecular 'Biology, 43,135.
MARTIN, T.E. & WOOL, I.G. (1969) Active hybrid 80 s particles formed from subunits of rat, rabbit and protozoan (Tetrahymenapyriformis) ribosomes. Journal of Molecular Biology, 43,151.
MARTIN, W.J., MAURER, P.H. & BENACERRAF, B. (1971) Genetic control of immune responsiveness to a glutamic acid, alanine, tyrosine copolymer in mice. I. Linkage of responsiveness to H-2 genotype. Journal of Immunology, 107,715.
MCDEW, H.O. (1968) Genetic control of the antibody response. III. Quantitative and qualitative charac- teristics of the antibody response to (T, G)-A-L in CBA and C57 mice. Journal of Immunology, 100,485.
MCDEVITT, H.O. & CHINITZ, A. (1969) Genetic control of antibody response: relationship between immune response and histocompatibility (H-2) type. Science, 163, 1208.
MCDEVIIT, H.O. & SELA, M. (1965) Genetic control of the antibody response. I. Demonstration of deter- minant-specific differences in response to synthetic polypeptide antigens in two strains of inbred mice. Journal of Experimental Medicine, 122,517.
M c D E v ~ , H.O. & SELA, M. (1967) Genetic control of the antibody response. 11. Further analysis of the specificity of determinant-specific control, and genetic analysis of the response to (H, G)-A-L in CBA and C57 mice. Journal of Experimental Medicine, 126,969.
MCDEVITT, H.O., SHREFFLER, D.C. & STIMPFLING, J.H. (1969) A single chromosome region in the mouse controlling the major histocompatibility antigens and the ability to produce antibody to synthetic polypeptides. Journal of Clinical Investigation, 48, 57a.
MCDEVITT, H.O. & TYAN, M.L. (1968) Genetic control of the antibody response in inbred mice. Transfer of response by spleen cells and linkage to the major histocompatibility (H-2) locus. Journal of Experimental Medicine, 128, 1 .
MERRYMAN, C.F. & MAURER, P.H. (1971) Genetic control of immune response to synthetic polypeptides in mice. Federation Proceedings, 30, 470.

Antibody synthesis in vitro. XI1 243 MERRYMAN, C.F. and MAURER, P.H. (1972) Genetic control of immune response to glutamic acid, alanine,
tyrosine copolymers in mice. I. Association of response. Journal of Immunology, 108, 135. Moz~s, E., M C D E V I ~ , H.O., JATON, J.C. & SELA, M. (1969) The genetic control of antibody specificity.
Journal of Experimental Medicine, 130,1263. UTHBUN, W.E. & HILDEMAN, W.H. (1970) Genetic control of the antibody response to simple haptens in
oncogenic strains of mice. Journal of Immunology, 105,98. SNELL, G.D., CUDKOWICZ, G . & BUNKER, H.P. (1967) Histocompatibility genes of mice. VII. H-13, a new
histocompatibility locus in the fifth linkage group. Transplantation, 5,492. STROHMAIER, K. & MUSSGAY, M. (1 959) Bestimmung der Sedimentationskonstante eines infektiosen F’rizips
mit Nucleinsaurecharakter aus dem Virus der Maul- und Klauenseuche mit Hilfe der Gradienten- Zentrifugation. Zeitschrgt fiir Naturforschung, 14., 171.
TRACHSEL, H. (1973) Bildung hybrider Ribosomen aus HeLo- und Lymphoblasten- Unfereinheiten und PrCfung ihrer biologischen Aktivitat in der Poly( lJ)-abhiingigen Poly-Phenylalanin-Synthese und im antikorper- synthetisierenden System. Thesis, Bern.
Vm, N.M. & LEVINE, B.B. (1 970) Immune responses of inbred mice to repeated low doses of antigens : relation- ship to histocompatibility (H-2) type. Science, 168, 852.
VAZ, N.M., VAZ, E.M. & LEVINE, B.B. (1970) Relationship between histocompatibility (H-2) genotype and immune responsiveness to low doses of ovalbumin in the mouse. Journal of Immunology, 104,1572.
VLADUTIU, A.O. & ROSE, N.R. (1971) Autoimmune murine thyroiditis relation to histocompatibility (H-2) type. Science, 174,1137.
WELLS, V.J., FUDENBERG, H.H. & MACKEY, J.R. (1971) Relation of the human antibody response to flagellin to Gm-genotype. Journal of Immunology, 107,1505.