Embed Size (px)
Citation preview

[CANCER RESEARCH 46, 2588-2595, May 1986]
Anti-Neurofilament Antibodies in the Sera of Patients with Small Cell Carcinomaof the Lung and with Visual Paraneoplastic Syndrome1
Steven E. Kornguth,2 Tom Kalinke, Gerald B. Grunwald,3 Henry Schutta, and Doris Dahl
Departments of Neurology ¡S.E. K., T. K., H. S.J and Physiological Chemistry fS. E. K.J, University of Wisconsin, Madison, Wisconsin; Laboratory of BiochemicalGenetics, National Heart, Lung, and Blood Institute, NIH, Bethesda, Maryland [G. B. G.J;and Veterans Administration Hospital, West Roxbury, Massachusetts [D. D.]
ABSTRACT
The sera of patients with small cell carcinoma of the lung (SCCL) andan associated visual paraneoplastic syndrome (VPNS) have high titerimmunoglobulins that react with retinal ganglion cells and with clonedlines of the SCCL. The immunoglobulins in the sera of two patients withSCCL and VPNS reacted with at least one common antigen shared byneural cells and cloned lines of the SCCL. The molecular weights of thepredominant neural and tumor antigens were 205,000, 145,000, 65,000,and 20,000-24,000 as determined by Western blots. Three of the antigensfrom neural tissue copurify and comigrate electrophoretically with neu-rofilament proteins. Polyclonal antibodies prepared against authenticneurofilament proteins react with antigens having molecular weightsidentical to those of proteins that react with immunoglobulins from theSCCL-VPNS patients. Polyclonal antibodies that were prepared againstisolated retinal ganglion cells and that were shown previously to causethe immunoablation of the ganglion cells in vivo reacted most intenselywith the 17, 205,000 antigen and weakly with the M, 145,000 and M,70,000 antigens. Treatment of the Western blots with alkaline phospha-tase from Escherìchiacoli did not affect the immunoreactivity betweenthe immunoglobulins and the purified neurofilament proteins. It is proposed that the immunoglobulins in the sera of patients with SCCL-VPNSmay be involved etiologically in the development of the VPNS.
Previous studies in our laboratory revealed that the intra-vitreous injection of polyclonal immunoglobulins, produced inrabbits against large retinal ganglion cells, could cause theimmunoablation of these retinal cells in cats (9, 10). The lossof the large ganglion cells in retina and of their projections tothe lateral geniculate nucleus was determined by histológica!analysis of retinal wholemounts (10) and by electrophysiologi-cal studies of the lateral geniculate nucleus (11).
The immunoablation of the ganglion cells of the cat retinas,resulting from intravitreal injection of the polyclonal immunoglobulins, is similar in several respects to the visual PNS inthe patients with SCCL. The rabbit immunoglobulins that reactwith the ganglion cells in cat retina also react with bovine brainproteins having molecular weights of 205,000 and 145,000. Theretinas obtained from one patient with SCCL and visual PNSexhibited a marked loss of large ganglion cells,5 as did the
retinas from the cats. This report describes the characterizationof the neurofilament antigens that react with the immunoglobulins from the patients with SCCL-VPNS. It also presents thehypothesis that the immunoglobulins in these patients may beinvolved etiologically in the development of the visual PNS.
INTRODUCTION
A subset of patients with carcinomas develops neurologicaldysfunction even when the nervous system is not invaded withthe primary tumor or métastases.These dysfunctions are identified as PNS,4 and the syndromes were reviewed in 1982 by
Minna and Bunn ( 1). It has been shown immunohistochemicallythat patients with ovarian carcinoma and cerebellar PNS havehigh titer immunoglobulins reactive with Purkinje cells (2, 3).We have previously shown immunohistochemically that hightiter immunoglobulins, reactive with retinal ganglion cells (4,5), were present in the sera of those patients with SCCL whohad VPNS. These SCCL-VPNS immunoglobulins recognize asmall number of protein antigens in extracts of both retina andsmall cell cancer tumors. The protein antigens have molecularweights of 205,000, 145,000, 65,000 and 20,000-24,000 (5).The similarity of the molecular weights of the proteins in theneurofilament triplet (6, 7) to three of the antigens reportedhere (205,000, 145,000 and 65,000) suggested that the neuralantigens may be neurofilaments (5). It has been demonstratedhistochemically that neurofilaments are present in oat cell lungcancers (8).
Received 9/9/85; revised 1/27/86; accepted 1/31/86.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by a Grant from the Graduate School and from the Departmentsof Neurology and Radiology, University of Wisconsin, Madison, Wl.
2To whom requests for reprints should be addressed, at Department ofNeurology and Physiological Chemistry, Waisman Center, Rm. 659, Universityof Wisconsin, Madison, Wl 53705.
3 Present address: Anatomy Department, Thomas Jefferson University, Phil
adelphia, PA.' The abbreviations used are: PNS, paraneoplastic syndromes, SCCL, small
cell carcinoma of the lung; VPNS, visual paraneoplastic syndrome: SDS, sodiumdodecyl sulfate; PBS, phosphate buffered saline (0.01 Mphosphate, pH 7.4); HA,hydroxylapatite.
MATERIALS AND METHODS
Human Serum Samples
The sera from seven patients with SCCL and VPNS were obtainedover a 5-year period. Two of these seven sera were described earlier (4,5); the clinical history of patient 1 and the ¡mmunohistological reactionof his serum was reported in (4), and the clinical history from both ofthese patients was described briefly in (5). In addition to these sevenpatients, the present study examined the sera of five normal controlsubjects, of patients with SCCL and no VPNS, and of patients withdifferent forms of lung cancer.
Polyclonal Antibodies Produced against Authentic Neurofilament Proteins
The polyclonal antibodies were produced in rabbits against purifiedneurofilaments by Dr. Doris Dahl and were a gift from her laboratory.
Preparation of the Neural Protein Extracts for Gel Electrophoresis
The neural tissues were placed in four volumes of sample buffercontaining 2% SDS, 12% glycerol, and 63 HIM Tris, pH 6.8. Thesamples were homogenized with a Potter-Elvehjem tissue grinder, andthe homogenates were boiled for 2 min. Then 2-mercaptoethanol wasadded to a final concentration of 5%. These samples were used for gelelectrophoresis and Western blot studies.
Polyacrylamide Gel Electrophoresis
Polyacrylamide gels were prepared and run as described previously(12). The gels were prepared as either 5, 7.5,10, or 15% polyacrylamidegel slabs. The resolving gel was 0.375 MTris, pH 8.8, and the stackinggel was 0.125 MTris, pH 6.8. The protein extracts in the sample bufferswere applied to the gel (12 cm x 1.5 mm), and electrophoresis was done
*G. Grunwald, S. Kornguth, J. Towfighi, J. Sassani, M. Simmonds, C.Housman, and N. Papadopoulos. Autoimmune basis for visual paraneoplasticsyndrome in patients with small cell lung carcinoma, manuscript in preparation,1986.
2588
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTI-NEUROFILAMENT ANTIBODIES IN SMALL CELL CARCINOMA
at 30 mA until the tracking dye reached the bottom edge of the gel.The tracking dye was 0.01% bromphenol blue. The gel slabs that werestained for protein were placed in 0.25% Coomassie blue R-200 dissolved in an aqueous solution containing 9.2% acetic acid and 50%methanol. The gels were destained with an aqueous solution containing7.5% acetic acid and 12% isopropanol.
Western Blot Analysis
The gels that were transblotted onto nitrocellulose sheets (Bio-Rad)were not prestained with Coomassie blue. The proteins were transferredto the nitrocellulose membrane in a Bio-Rad apparatus by electropho-resis at 70 mA for 16 h in 20 mM Tris: 190 mM glycine buffer (pH 8.3)containing 20% methanol. The blots were incubated for 60 min witheither 10% normal goat serum or with 10% bovine serum albumin toblock the non-specific adsorption of immunoglobulins to the nitrocellulose matrix. The immune serum samples were diluted 1:100 to1:10.000 with PBS containing 5% normal goat serum and then placedon the blot and incubated for 16 h. Most of the serum dilutions were1:100. The blots were washed four times with PBS and then incubatedfor 2 h with appropriate second antibody (goat anti-human IgG, IgMcoupled to horseradish peroxidase; Kirkegaard and Perry). The secondantibody was diluted 1:3000 with PBS containing 3% normal goatserum. The blots were rinsed four times with PBS and then developedwith diamino benzidine and hydrogen peroxide.
Preparation of Neurofilaments
Urea Extraction. A modification of the methods of Tokutake et al.( 13) was used. Neural tissue samples were homogenized with a Polytrontissue grinder (Brinkman) in two volumes of buffer A. This buffercontained 10 mM sodium phosphate, pH 7.4, 1 mM EDTA, 0.1 Msodium chloride, 0.5 mM phenylmethyl sulfonyl fluoride, 0.85 M sucrose, and 1% Triton X-100. The homogenate was stirred for 16 h at4°Cand then centrifuged in a type 35 rotor at 100,000 x g for 30 min.
The recovered pellets were suspended in one-half the volume of bufferA (but without sucrose) used in the original extraction. The suspensionwas layered on buffer A which contained sucrose but not Triton andwas centrifuged for 30 min at 27,000 x g in an SW 27 rotor. Theresulting pellets were suspended in 10 mM sodium phosphate, pH 7.4,containing 1% of 2 mercaptoethanol and 8 M urea (half the volume ofbuffer A used in the original extraction). The suspension was stirredfor 30 min at 4°Cand then centrifuged at 100,000 x g for 30 min. This
last supernatant was used for gel electrophoresis.Thiocyanate Extraction. An alternate procedure was used to purify
the neurofilament proteins because urea carbamylates the proteins andbecause thiocyanate purification of the neurofilaments yielded increasedamounts of the M, 205,000 proteins compared with the urea. Neuraltissue samples were placed in two volumes of the buffer A describedabove. After homogenization with a Polytron tissue grinder and extraction as described above, the suspension was stirred for 90 min at 4°C
and then centrifuged at 20,000 x g for 30 min. This final pellet wassuspended in 10 volumes of 10 mM sodium phosphate, pH 7.4, containing 1% 2-mercaptoethanol and 1.5 M sodium thiocyanate. From230 g brain tissue, 34 g of pellet (wet weight) were obtained.
Batchwise Elution of the Neurofllament Protein from Hydroxylapatite
The HA (purchased from Sigma) was equilibrated with 10 mMsodium phosphate, pH 7.4, containing 8 M urea. The neurofilamentproteins were dissolved in 10 mM phosphate buffer, pH 7.4, containing1% 2-mercaptoethanol and 8 M urea. The solution was added to 15 gof the HA in 25 ml of the equilibration buffer. A total of 500 mgprotein were applied to the HA. The suspension was centrifuged at10,000 g for 10 min. The supernatant was removed and the HA wasthen eluted with 70 ml of 130 mM sodium phosphate, pH 7, containing8 M urea. The HA was finally eluted with 70 ml of 300 mM sodiumphosphate, pH 7, containing 8 M urea. The final eluate contained 60-80 mg protein as determined by absorbance at 280 nm and by theBradford method (14).
Chromatograph}' of Proteins on DEAE Cellulose
DE-32 (Whatman) was prepared for chromatography and then equil
ibrated with phosphate buffer (10 mM, pH 7) containing 1 mM EDTA,0.1% 2-mercaptoethanol, and 6 M urea. Thirty mg of neurofilamentproteins that were extracted with urea were dissolved in 25 ml of thesame phosphate:EDTA:urea:mercaptoethanol buffer and applied to theDEAE column (1.4 x 45 cm). The proteins were eluted at a flow rateof 18 ml/h with a 500-ml gradient of sodium phosphate; the gradientranged from 10 to 300 mM phosphate. All elution buffers contained 1mM EDTA, 0.1% mercaptoethanol, and 6 M urea.
The neurofilament proteins that were extracted with sodium thiocyanate were applied to the DE-32 in phosphate buffer (0.01 M, pH 7.4)and were eluted from the DE-32 with the same phosphate buffercontaining a gradient of sodium thiocyanate (0 to 0.5 Mthiocyanate).
RESULTS
Interaction of Neural Proteins with Immunoglobulins fromSera of Patients with SCCL and Visual PNS. The sera of sevenpatients with SCCL and visual PNS contained immunoglobulins that reacted with a limited number of proteins in retinaand in cloned lines of SCCL. The molecular weights of theantigens were 205,000, 145,000,65,000, and 20,000-24,000 asdetermined by western blots. Fig. 1 illustrates the reaction ofthe retinal antigens with immunoglobulins in the sera of twosuch patients at high serum dilutions. The serum from one ofthese two (patient 1) reacted with retinal antigens of molecularweight 65,000 and 20,000-24,000, while that from patient 2reacted with the antigens of molecular weight 145,000 and205,000. These two patients were described earlier (4, 5) andhave been examined in detail in this report because (a) serumsamples were obtained from them at different times during theirclinical course and (b) retinal and neural tissue was obtainedfrom patient 2 after his death. The sera from the other fivepatients with SCCL and VPNS also reacted with subsets ofthese four retinal proteins but not with other proteins at thedilutions examined. The serum immunoglobulins from control
MW X ICT3
200-
92-
68-
45-
a b cdefghijklmnoFig. 1. Reaction of retinal antigens with antibodies in sera of patients with
SCCL-VPNS. Retinal proteins were solubilized, resolved by SDS-PAGE, andtransferred to nitrocellulose paper. For immunodetection, the nitrocellulose wasincubated with normal or SCCL-VPNS sera and then with goat-anti-human (IgG+ IgM + IgA) conjugated to horseradish peroxidase. The bands were visualizedwith diaminobenzidine and hydrogen peroxide. Lane a, protein molecular weightstandards (indicated at left): lane b, total protein detected with Amido Black; lanec. no primary or secondary antibodies; lane d, secondary antibody alone; lanes ethrough h, normal human serum at 1:8000, 4000, 1000, and 500-fold dilutions;lanes i through k, SCCL-VPNS patient 1 serum at 1:4000, 1000, and 500; lanesI through 0, SCCL-VPNS patient 2 serum at 1:8000, 4000. 1000, and 500. MW,molecular weight.
2589
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTI-NEUROFILAMENT ANTIBODIES IN SMALL CELL CARCINOMA
subjects did not react with these antigens at the dilutionsexamined.
The sera from two of the patients with SCCL and VPNS(patients 1 and 2) reacted with antigens solubili/ed from clonedlines of SCCL obtained from Dr. John Minna (National CancerInstitute). The immunoglobulins from patient 1 reacted with aA/r 65,000 protein shared by all four SCCL clone lines andhuman and chick retina (Fig. 2). The immunoglobulins frompatient 2 reacted with a M, 150,000 protein shared by all fourSCCL clone lines and human and chick retina. Immunoglobulins from patient 1 also reacted with a M, 90,000 protein inchick retina and a M, 20,000-24,000 protein in human retinathat are not shared by the SCCL lines. Immunoglobulins frompatient 2 reacted with a M, 205,000 protein in human and chickretina not shared by the SCCL lines; they also react with a M,40,000-44,000 protein in two of the four SCCL clones that isnot present in the other cell lines.
The immunoglobulins also reacted with frozen sections ofretina. Neither the immunohistochemical nor the Western blotstudies indicated species specific reaction of the immunoglobulins, since neural tissue sections and protein extracts fromchick, human, cat, and dog retinas reacted similarly. Sera frompatients with small cell carcinoma of lung but without visualPNS did not react immunohistochemically with the retinalganglion cells. The sera of one of these SCCL patients, whohad no visual or other PNS, contained immunoglobulins thatreacted with protein antigens having molecular weights of205,000 and 145,000 (Fig. 3); no reaction was detected withthe M, 65,000 protein. The serum of a patient who had both ahronchoalvcolar carcinoma of the lung and a squamous cellcarcinoma of the parotid gland and who had weakness anddysphagia attributed to PNS but with no visual PNS had hightiter immunoglobulins against a M, 65,000 neural protein (Fig.3). The serum from this patient reacted weakly with the A/r205,000 protein but not with the M, 145,000 antigen. Theserum from each of these two patients did not react immunohistochemically with retinal sections. The data from these twopatients demonstrate that immune reactivity with the M,205,000, 145,000, or 65,000 proteins in neural tissue does not
abode abcdef abcdef
MW X 10-3
Fig. 2. Similarity of human retinal antigens in molecular weight and immunoreactivity to SCCL cell line antigens and chicken retinal antigens. Sameconditions as in Fig. 1. except that all sera were diluted 1:5000. Panel I, normalhuman serum; panel II. serum from one patient with SCCL-VPNS; panel III,scrum from a second patient with SCCL-VPNS. Lanes a through d are SCCLcell lines 345. 187. 145. and 69 in each respective panel. Human retina is in lanec. panel I and lanef, panels II and III. Adult chicken retina is in lane e. panels IIand III. \ portion of this figure was published previously (5).
40-
a b e d eFig. 3. Reaction of neural antigens with antibodies from cancer patients who
had no VPNS and effects of alkaline phosphatase treatment. Lane a, serum fromSCCL patient without VPNS, dilution 1:100; lane b, serum from control, dilution1:100: lane c, serum from patient with squamous cell carcinoma of the parotidand bronchoalveolar carcinoma, dilution 1:100; lane d, electroblot treated withalkaline phosphatase from /,. coli and then incubated with serum from the patientwith SCCL but without VPNS (same as lane a); lane e, electroblot treated withalkaline phosphatase from bovine intestine and then incubated with serum frompatient with SCCL but without VPNS. The second antibody in all cases was goat-anti-human IgG, IgM coupled to horseradish peroxidase. A/H , molecular weight.
necessarily indicate reaction of the serum immunoglobulinswith retinal neurons.
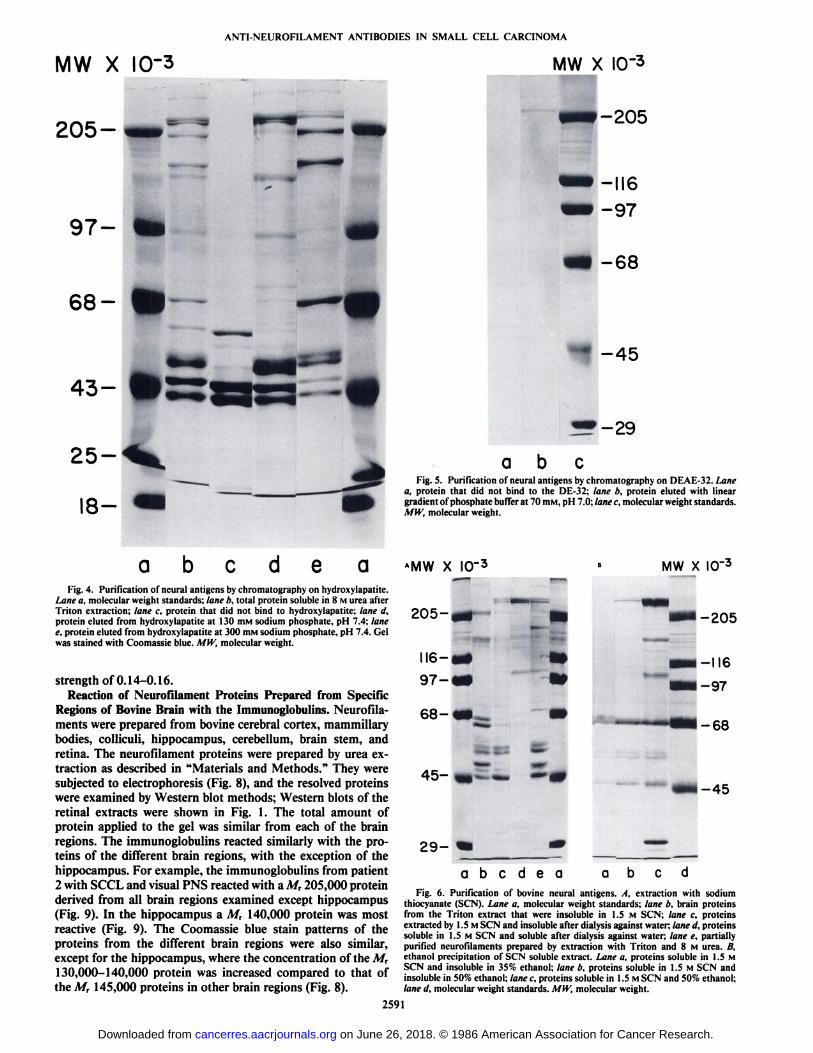
Purification of the Neurofilament Triplet Proteins. Neuro-filament proteins from bovine white matter were solubilizedeither using urea or sodium thiocyanate as described in "Materials and Methods." The urea solubilized proteins were puri
fied further by batchwise elution from HA (Fig. 4) and bychromatography on DE-32 cellulose (Fig. 5). The neurofila-ment proteins were eluted from the HA at 0.3 M sodiumphosphate and from the DE-32 at an ionic strength of 0.14-0.16, with a gradient of phosphate or sodium thiocyanate asthe eluting solvent.
The neurofilament proteins from bovine brain were solubilized with 1.5 M sodium thiocyanate (Fig. 6.Õ)and purifiedfurther by ethanol fractionation (Fig. 6Ä).Although a smallportion of the high molecular weight (M, 205,000) componentof the neurofilament proteins did precipitate in 50% ethanol,most of it remained in solution (Fig. (ìli).The ethanol causedthe removal of other proteins and nucleic acids by precipitation.The purified neurofilament proteins reacted with the immunoglobulins obtained from the sera of patients with SCCL andvisual PNS; Fig. 7 illustrates the reaction with immunoglobulins from patient 2. The neurofilaments extracted with thethiocyanate were purified further by chromatography on acolumn of DE-32 as described in "Materials and Methods."
The neurofilaments were eluted from the column at an ionic
2590
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
ANTI-NEUROFILAMENT ANTIBODIES IN SMALL CELL CARCINOMA
MW X 10-3 MW X 10-3
205- *•»- '-205
68-
43-
-68
-45
^-29
aFig. 5. Purification of neural antigens by chromatography on DEAE-32. Lane
a, protein that did not bind to the DE-32; lane b, protein eluted with lineargradient of phosphate buffer at 70 mM, pH 7.0; lane c, molecular weight standards.MW, molecular weight.
a b e a e aFig. 4. Purification of neural antigens by chromatography on hydroxylapatite.
Lane a, molecular weight standards; lane b, total protein soluble in 8 M urea afterTriton extraction; lane c, protein that did not bind to hydroxylapatite; lane d,protein eluted from hydroxylapatite at 130 mM sodium phosphate, pH 7.4; lanee, protein eluted from hydroxylapatite at 300 mM sodium phosphate, pH 7.4. Gelwas stained with Coomassie blue. MW, molecular weight.
strength of 0.14-0.16.Reaction of Neurofilament Proteins Prepared from Specific
Regions of Bovine Brain with the Immunoglobulins. Neurofila-ments were prepared from bovine cerebral cortex, mammillarybodies, collidili, hippocampus, cerebellum, brain stem, andretina. The neurofilament proteins were prepared by urea extraction as described in "Materials and Methods." They were
subjected to electrophoresis (Fig. 8), and the resolved proteinswere examined by Western blot methods; Western blots of theretinal extracts were shown in Fig. 1. The total amount ofprotein applied to the gel was similar from each of the brainregions. The immunoglobulins reacted similarly with the proteins of the different brain regions, with the exception of thehippocampus. For example, the immunoglobulins from patient2 with SCCL and visual PNS reacted with a M, 205,000 proteinderived from all brain regions examined except hippocampus(Fig. 9). In the hippocampus a M, 140,000 protein was mostreactive (Fig. 9). The Coomassie blue stain patterns of theproteins from the different brain regions were also similar,except for the hippocampus, where the concentration of the M,130,000-140,000 protein was increased compared to that ofthe Mr 145,000 proteins in other brain regions (Fig. 8).
*MW X IO'3
205-,
116-
97-
29-
MW X IO"3
-205
I-II6
1-97
1-68
1-45
a b c d e aFig. 6. Purification of bovine neural antigens. A, extraction with sodium
thiocyanate (SCN). Lane a, molecular weight standards; lane b, brain proteinsfrom the Triton extract that were insoluble in 1.5 M SCN; lane c, proteinsextracted by 1.5 MSCN and insoluble after dialysis against water; lane d, proteinssoluble in 1.5 M SCN and soluble after dialysis against water; lane e, partiallypurified neurofilaments prepared by extraction with Triton and 8 M urea. B,ethanol precipitation of SCN soluble extract. Lane a, proteins soluble in 1.5 MSCN and insoluble in 35% ethanol; lane b, proteins soluble in 1.5 M SCN andinsoluble in 50% ethanol; lane c, proteins soluble in 1.5 MSCN and 50% ethanol;lane d, molecular weight standards. MW, molecular weight.
2591
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTI-NEUROFILAMENT ANTIBODIES IN SMALL CELL CARCINOMA
MW X 10-3 MW X 10-3
205--
145-
70-
a b c dFig. 7. Western blots of bovine neural proteins extracted with sodium thiocy-
anate (SCN) incubated with human serum and with rabbit anti-neurofilamentimmunoglobulins. Lane a, polyclonal rabbit serum prepared by Dr. D. Dahlagainst isolated M, 205.000 neurofilament protein, diluted 1:500: lane b, serumfrom a patient with SCCL-VPNS, diluted 1:100: lane c. control human serum,diluted 1:100: lane d. polyclonal rabbit immunoglobulins prepared against isolatedlarge ganglion cells from bovine retina, diluted 1:250. The second antibody wasgoat-anti-rabbit IgG. IgM coupled to horseradish peroxidase for those lanesincubated with rabbit immunoglobulins and goat- anti-human IgG. IgM, IgAcoupled to horseradish peroxidase for lanes incubated with human immunoglobulins. Mil', molecular weight.
Reaction of the Immunoglobulins with Neurofilament ProteinsPrepared from Bovine and Rat Brain. Neurofilaments wereprepared by urea extraction and by thiocyanate extraction frombovine and rat brains. The proteins were resolved on acrylamidegels and were then examined by the Western blot technique.The proteins recovered from each of the two species reactedsimilarly with the immunoglobulins from the sera of the patients with carcinomas. Fig. 10 illustrates the reaction of theserum immunoglobulins from a patient with SCCL and noVPNS with the M, 205,000 protein from rat and bovine neuraltissue. Several patients with SCCL and VPNS, including patient 2, also reacted with the Mr 205,000 protein.
Interaction of Anti-Retinal Ganglion Cell Immunoglobulinswith Neural Proteins. Polyclonal antibodies were prepared inrabbits against large ganglion cells isolated from ox retinas (9).The immunoglobulin fractions of the rabbit sera were separatedfrom the remaining serum proteins by chromatography on DE-32. The injection of the purified immunoglobulins into thevitreous of cats caused the immunoablation of the large ganglion cells in retina (10). The injections of the immunoglobulinsalso caused the marked inhibition of the electrophysiologicalactivity of nerve cells in the lateral geniculate nucleus thatreceived input from the large ganglion cells in the retina (11).
To obtain information about the molecular weights of theneural antigens, the purified rabbit immunoglobulins were in-
205-*«'
«116- «•97- «•
es- m—
'
45-
29-
abcde fgaFig. 8. Coomassie blue patterns of neurofilament fractions from various
regions of bovine brain. Lane a, molecular weight standards: lane b, proteins frombrainstem: lane c, from cerebellum; lane d. from colliculi: lane e, from cerebralcortex: lane f. from hippocampus: lane g, from mamillary bodies. Proteins weresolubili/cd with urea and stained with Coomassie blue. MW, molecular weight.
cubated with Western blots of the bovine neural tissue and thepartially purified neurofilaments (Fig. 7). It was found thatthere was an immunological reaction with the M, 205,000protein (Fig. 7, lane d) and weaker reactions with the M, 70,000and M, 145,000 proteins. The protein antigens that reactedwith the polyclonal antibodies against authentic neurofilamentprotein (Fig. 7, lane a) reacted both with rabbit immunoglobulins against the large retinal ganglion cells and with the humanSCCL-VPNS immunoglobulins, as determined from their elec-
trophoretic mobilities. The similarity of the molecular weightsof the antigens and the copurification with neurofilament proteins suggest that the rabbit immunoglobulins and the humanimmunoglobulins are directed against similar proteins.
Alkaline Phosphatase Treatment of Neurofilaments Does NotAffect Interaction with Immunoglobulins. The neurofilamentproteins were subjected to electrophoresis on polyacrylamidegels and then electroblotted onto nitrocellulose. The electro-blots were treated with alkaline phosphatase prepared fromeither bovine intestinal mucosa (Worthington, similar to SigmaType VII) or Escherichia coli (Sigma, Type III). Five units ofeach enzyme were used. The incubations were maintained inPBS (pH 7.4, 0.01 M phosphate) at 25°Cfor 2 h. The E. coli
phosphatase had no effect on the interaction of the immunoglobulins with the neurofilament proteins (Fig. 3). From ourstudies with Coomassie blue stained polyacrylamide gels, thebovine enzyme is contaminated with protease. This resulted inthe hydrolysis of the protein bands on the nitrocellulose. Elevated concentrations of phosphate (50 mivi) did not inhibit theloss of immunoreactivity of the M, 205,000 neurofilamentprotein treated with bovine phosphatase, even though phospha-
2592
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTI-NEUROFILAMENT ANTIBODIES IN SMALL CELL CARCINOMA
MW X 10-3
200-
140-
- 205-
abcdefghFig. 9. Western blots of neurofilament proteins from various regions of bovine
brain. Lanes a and b, cerebellar cortical antigens; lanes c and d, cerebral corticalantigens; lanes e and f, hippocampal antigens; and lanes g and h, bovine wholebrain antigens. Lanes a, c, e, and g were from a patient with SCCL-VPNS and b,d, f, and h were from normal human controls. Primary sera were diluted 1:100.Second antibody was goat anti-human IgG, IgM, IgA coupled to horseradishperoxidase. MW, molecular weight.
tase activity is inhibited at this concentration of phosphate.These observations indicate that (a) the antigen is a protein and(b) the reaction of the immunoglobulins from the patients withSCCL and visual PNS is not dependent upon the phosphoryl-ation of the neurofilament protein (15).
DISCUSSION
We have shown that the immunoglobulins from sera ofpatients with small cell carcinoma of lung and an associatedvisual paraneoplastic syndrome (SCCL-VPNS) react immuno-histochemically with the retina. From Western blot analyses itis concluded that these immunoglobulins react with proteins ofthe neurofilament triplet and with a M, 20,000-24,000 protein.Three of the four primary antigenic species are neurofilamentproteins because they copurify and comigrate electrophoreti-cally with the neurofilaments. They also react with polyclonalantibodies prepared against the neurofilaments. At the high(>!()()) serum dilutions used in this study the neurofilamentsand a protein of molecular weight 20,000-24,000 appear to bethe primary antigens with which the immunoglobulins react asdetermined by Western blot analysis. The possibility must beconsidered that other retinal antigens react histochemically withthe immunoglobulins but were not detected by the Westernblot. For example, the antigenic determinants may have beenaltered by the SDS-polyacrylamide gel electrophoresis procedure, or the antigen may be of low molecular weight (less thanM, 10,000) and therefore not retained on the gel. The experimental evidence to date indicates that the antigens that are
-abcdefgh
Fig. 10. Western blots of neurofilament proteins from rat and bovine brain.Lanes a and b, rat brain antigens insoluble in 1.5 M SCN; Lanes c and d, rat brainantigens soluble in 1.5 M SCN; lanes e andf, rat brain antigens extracted in 1.5MSCN and soluble in water; and lanes g and h, solubilized antigens from bovinebrain. Lanes a, c, e, and g were from SCCL without VPNS, and lanes b, d, f, andh were from a normal control subject. Primary serum samples were diluted 1:100.Second antibody was goat anti-human IgG, IgM, IgA coupled to horseradishperoxidase.
detected histochemically are the same ones as revealed on theWestern blots.
We propose that the anti-retinal immunoglobulins in thepatients with SCCL-VPNS may be etiologically connected with
the occurrence of the visual PNS. The criteria required for thishypothesis to be valid are as follows: (a) immunoglobulins thatreact specifically with neurons of the visual pathway must bepresent in the patients at sufficient titer; (b) these immunoglobulins must have access to the neurons in the central nervoussystem; (c) the immunoglobulins must recognize either theantigen against which they were induced or a cross-reactingmolecule on the surface of the neurons in the visual system; (d)those neurons in the visual pathway that react specifically withthe immunoglobulins should be reduced in numbers in thepatients with the visual PNS; and (e) immunoglobulins againstsimilar antigenic determinants should cause the immunoabla-tion of the neurons in the visual pathway.
Most of these conditions have been satisfied in the cases ofSCCL-VPNS, as will be discussed below. Patients with othertumors, malignant melanoma for example, were shown to havehigh titer serum immunoglobulins that reacted with the tumor(16, 17).
Immunoglobulin Specificity. We have demonstrated that theimmunoglobulins of patients with SCCL-VPNS react immu-nohistochemically with ganglion cells of retinas from humans,macaques, dogs, cats, rats, mice, and chicks (4, 5). The immunoglobulins were detectable at high dilutions, and their reactionwith the retinal ganglion cells was specific. The extent of serum
2593
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTI-NEUROFILAMENT ANTIBODIES IN SMALL CELL CARCINOMA
dilution in immunological assays is an important factor in theinterpretation of results. For example, Stefansson et al. (18)reported that over 90% of healthy individuals had immuno-globulins in their sera that reacted with neurofilament proteinson Western blots at serum dilutions of 1:50; they state that alltheir studies were done at a serum dilution of 1:50. In ourstudies, the sera from SCCL-VPNS patients and controls werediluted extensively, from 1:100 to 1:8000 (5). The titer againstthe M, 205,000 protein, detected in the sera of SCCL-VPNSpatients at these dilutions, was markedly elevated. Because ofthe difference in serum dilutions, our results are not contradictory to those of Stefansson et al. Although Stefansson et al.( 18) concluded that none of the disease categories was associated with high titers of antibodies to the M, 200,000 protein, asingle dilution of serum cannot reveal titer level, even thoughit may reflect the incidence of a specific immunoglobulin in apatient population.
The reaction between immunoglobulins from patients withSCCL-VPNS and retinal ganglion cells was another exampleof the specificity of the response. Other retinal neurons reactedweakly or not at all, and other central nervous system neurons(e.g., Purkinje cells) did not react with these immunoglobulins,as determined immunohistochemically. Of particular interestwas the lack of reaction of cerebellar Purkinje cells with serumimmunoglobulins obtained from the patients with SCCL-VPNS. It has been reported that patients with cerebellar PNSassociated with ovarian carcinoma had high titer antibodiesthat reacted immunohistochemically with the Purkinje cells (2)but not with retinal ganglion cells (3). Therefore the immunoglobulins from the patients with PNS react immunohistochemically with neurons of the system affected by the PNS (e.g.,retinal ganglion cells in the visual PNS and cerebellar Purkinjecells in the cerebellar PNS) but not with other neurons. Yet inthis study it has been shown that the neurofilament fractionfrom different brain regions reacts on Western blots with theimmunoglobulins from patients with the SCCL-VPNS. Theseobservations suggest that the antigenic determinants againstwhich the SCCL-VPNS immunoglobulins react are maskedduring the immunohistochemical reaction but become availableduring the SDS-polyacrylamide gel electrophoresis procedure.There is evidence for the existence of neurofilament proteinswith neurotypic regions that reflect either the masking of different antigenic determinants in different neuronal populationsor the existence of variable domains in the neurofilaments.Goldstein et al. ( 19) observed that some monoclonal antibodiesreacted with neurofilaments in one population of neurons, whiledifferent monoclonal antibodies reacted with the neurofila-ments in a second neuronal population. This molecular heterogeneity or "neurotypy" was observed with monoclonal im
munoglobulins prepared against the M, 200,000 and M,155,000 neurofilament antigens. Drager et al. (20) also observedthat a monoclonal antibody, reactive with the M, 205,000neurofilament protein, interacted preferentially with particularretinal neurons and not with other neurons.
Immunoglobulin Access to the Central Nervous System. Frozen sections of the retinas from one of our patients with SCCL-VPNS revealed deposits of IgG or IgM in the retinal ganglioncell layer.5 This observation indicates that the immunoglobulins
in this patient did cross the blood retinal barrier as required bythe second condition. This patient's retina also had reduced
numbers of retinal ganglion cells, indicating that these cellswere lost in v/vo.5 In this case there was an association of the
loss of retinal ganglion cells with the localization of humanimmunoglobulins in the retina.
Anti-Neurofilament Antibodies Interact with Neuronal CellSurfaces. The third requirement of the hypothesis is that antibodies must have access to the cells //; situ and recognize oneor more markers on the surface of the neural cells if they areto initiate a neuropathological process. Fields and Yen (21)demonstrated the interaction of the anti-Afr 200,000 neurofilament antibodies with dissociated but apparently intact neurons from rat dorsal root ganglia. In our earlier studies on theimmunoglobulins generated against the large ganglion cellsisolated from retina, it was shown that the fluorescein labeledimmunoglobulins reacted with intact but dispersed large ganglion cells (9). As is shown in this report, the polyclonalimmunoglobulins from the rabbits immunized against the isolated retinal ganglion cells reacted with the proteins of theneurofilament triplet, as did the immunoglobulins from thepatients with SCCL-VPNS. The antibodies may also haveinteracted with other antigens immunohistochemically thatwere not detectable on Western blots, as indicated at the beginning of the "Discussion."
The surface antigens with which the antibodies react may bethe Mr 205,000 antigen or a cross-reacting molecule. For example Drager et al. (22) have shown that antibodies preparedagainst a-melanocyte stimulating hormone react with the Mr150,000 neurofilament protein. These observations indicatethat the patients' immunoglobulins may have interacted either
with the high molecular weight neurofilament protein (M,205,000) or with a protein that contains a similar antigenicdeterminant.
Immunoablation of Neurons with Anti-Ganglion Cell Antibodies. The fourth condition of the hypothesis is that the interaction of the immunoglobulins with the neuronal surface leads toimmunoablation of the neuron. Our previous studies have demonstrated that the rabbit immunoglobulins prepared against thelarge retinal ganglion cells can cause the immunoablation ofthe large ganglion cells when injected into the vitreous (10, 11).The loss was demonstrated by anatomical analysis of the retinaand by electrophysiological recordings from the lateral genicu-late nucleus. As discussed above, retinal flatmounts from apatient with SCCL-VPNS, who also had high titer anti-retinalganglion cell antibodies, revealed a loss of the large ganglioncells.5 These observations are related to the fourth and fifth
conditions of the hypothesis and are consistent with the hypothesis presented.
Anti-Neurofilament Antibodies and Neurological Dysfunction.Immunoglobulins in the sera of patients with several neurological disorders and in some control subjects have been reportedto react with the M, 205,000 protein of the neurofilamenttriplet. Increased levels of immunoglobulins against the neurofilament triplet proteins were observed in the sera of patientswith kuru, Creutzfeldt-Jakob disease, Alzheimer's disease, sub-acute sclerosing panencephalitis, and Parkinson's disease (23-
25). Toh et al. (25) observed antibodies in approximately 30%of normal control subjects and in 60-70% of the patients withthe neurological disorders. The reaction of immunoglobulins,prepared against the M, 200,000 neurofilament protein, withthe neurofibrillary tangles present in Alzheimer's disease (26)
has resulted in the hypothesis that these antibodies may beinvolved etiologically in the neuropathological process (27). Inthe present study it was shown that the sera from two patientswith carcinomas, but with no visual dysfunction, containedimmunoglobulins against the Mr 205,000 protein, even thoughthe immunoglobulins did not react immunohistochemicallywith the retinal ganglion cells.
It is necessary to consider the apparent paradox that certain2594
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

ANTl-NEUROFILAMENT ANTIBODIES IN SMALL CELL CARCINOMA
control subjects with no evidence of neurological dysfunctionalso contain high titer immunoglobulins in their sera. Theseobservations indicate one of four possibilities: (a) the anti-neurofilament and anti-A/r 20,000-24,000 protein antibodiesin the subjects without neurological dysfunction exist in theserum and do not cross the blood-brain or blood-retinal barrierin sufficient concentration to cause dysfunction; (¿>)the antibodies in the patients with SCCL-VPNS are directed againstdeterminants that are accessible on the affected cells of thepatient but not on controls; (c) the antibodies in the patientswith SCCL-VPNS are directed against determinants that aredifferent from those that bind the antibodies present in patientswith other PNS or in control subjects; or (d) the immunoglobulins to these proteins are not involved in the PNS. To distinguish among these, it will be necessary to characterize thesequence of the amino acids involved in those antigenic determinants that react with SCCL-VPNS immunoglobulins.
One proposed mechanism by which the anti-ganglion cellantibodies may cause the VPNS is presented here: (a) the rapidgrowth of the SCCL is accompanied by central necrosis of thetumor; (b) tumor necrosis results in the release of the antigensfrom the tumor; (c) the antigens stimulate the production ofimmunoglobulins that react with the antigens or antigenicallysimilar proteins on neuronal cell surfaces; (d) the permeabilityof the blood-brain or blood-vitreous barriers increases in asubset of patients with the tumor. This results in the access ofthe immunoglobulins to the retina; and (<•)the immunoglobulins
react with the antigens on the surface of the retinal ganglioncells causing the immunoablation of the ganglion cells andvisual dysfunction.
REFERENCES
1. Minna, J., and Bunn, P. A., Jr. Paraneoplastic Syndromes. In: V. T. DeVita,S. Hollinan, and S. Rosenberg (eds.). Principles and Practice of Oncology,pp. 1476-1517. Philadelphia: J.B. Lippincott, 1983.
2. Greenlee, J., and Brashear, H. R. Antibodies to Purkinje cells in patientswith paraneoplastic cerebellar degeneration and ovarian carcinomas. Ann.Neurol., 7*609-613, 1983.
3. Kornguth, S., Kalinke, T., Grunwald, G., Brooks, B., and Zimmerman, E.Antineurofilament antibodies associated with paraneoplastic syndrome. Fed.Proc., 44:966, 1985.
4. Kornguth, S., Klein, R., Appen, R., and Choate, J. Occurrence of anti-retinalganglion cell antibodies in patients with small cell carcinoma of the lung.Cancer (Phila.), SO: 1289-1293, 1982.
5. Grunwald, G., Klein. R., Simmonds. M.. and Kornguth, S. Autoimmunebasis for visual paraneoplastic syndrome in patients with small-cell lungcarcinoma. Lancet, /: 658-661, 1985.
6. Hoffman, P., and Lasek, R. The slow component of axonal transport:identification of major structural polypeptides of the axon and their generalityamong mammalian neurons. J. Cell Biol., 66:351-366, 1975.
7. Liem, R.. Yen, S. II.. Salomon, G., and Shelanski, M. Intermediate filaments
in nervous tissues. J. Cell Biol., 79:637-645, 1978.8. Lehto, V-P., Stenman, S., Miettinen, M., Dahl, D., and Virtanen, E. Expres
sion of a neural type of intermediate filament as a distinguishing featurebetween oat cell carcinoma and other lung cancers. Am. J. Pathol., 110:113-118, 1983.
9. Kornguth, S., Langer, E., and Scott, G. Antigenic properties of large ganglioncells isolated from ox retina. Exp. Eye Res., 33: 413-432, 1981.
10. Kornguth, S., Spear, P., and Langer, E. Reduction in numbers of largeganglion cells in cat retina following the intravitreous injection of antibodies.Brain Res., 245:35-45, 1982.
11. Spear, P., Jones, K., Zetlan, S., Geisart, E., and Kornguth, S. Effects ofantibodies to large ganglion cells on cat's retinogeniculate pathway. J. Neu -rophysiol., 47: 1174-1195, 1982.
12. I lames. B., and Rickwood, D. Gel Electrophoresis of Proteins: A PracticalApproach. Washington, D.C.: 1RL Press, 1983.
13. Tokutake, S., Hutchison, S., Pachter, J., and Liem, R. A batchwise purification procedure of neurofilament proteins. Anal. Biochem., 135: 102-105,1983.
14. Bradford, M. A rapid sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein dye-binding. Anal.Biochem., 72: 248-254, 1976.
15. Julien, J. P., and Mushynski, W. E. The distribution of phosphorylation sitesamong identified proteolytic fragments of mammalian neurofilaments. J.Biol. Chem., 258:4019-4025, 1983.
16. Lewis, M. G., Avis, P. J. G., Phillips, T. M., and Sheikh, K. M. A. Tumor-associated antigens in human malignant melanoma. Yale J. Biol. Mod.. 46:661-668, 1973.
17. Thomas, P. J., Kaur, J. S., Aitcheson, C. T., Roninson, W. A., and Tan, E.M. Antinuclear, antinucleolar, and anticytoplasmic antibodies in patientswith malignant melanoma. Cancer Res., 43: 1372-1380, 1983.
18. Stefansson, K., Marton, L., Dieperink, M., Molnar, G., Schlaepfer, W., andHelgason, C. Circulating autoantibodies to the 200,000 dalton protein ofneurofilaments in the serum of healthy individuals. Science (Wash. DC), 228:1117-1119,1985.
19. Goldstein, M., Sternberger, L., and Sternberger, N. Microheterogeneity(neurotypy) of neurofilament proteins. Proc. Nati. Acad. Sci. USA, 80:3101-3105, 1983.
20. Drager, U., Edwards, D., and Barnstable, C. Antibodies against filamentouscomponents in discrete cell types of the mouse retina. J. Neurosci., 2025-2042, 1984.
21. Fields, K., and Yen, S-H. Conventional antibodies to intermediate filamentsubunit proteins show cell-type specificity of 10-nm filaments. In: R. McKay,M. Raff, and L. Reichardt (eds.). Monoclonal Antibodies to Neural Antigens,Cold Spring Harbor, NY: Cold Spring Harbor Laboratory, 1981.
22. Drager, I !.. Edwards, P., and Kleinschmidt, J. Neurofilaments contain a-melanocyte-stimulating hormone (a-MSH)-like reactivity. Proc. Nati. Acad.Sci. USA, 80:6408-6412, 1983.
23. Bahmanyar, S.. Liem, R. K., Griffin, J., and Gajdusek, D. C. Characterizationof antineurofilament autoantibodies in Creutzfeldt-Jakob disease. J. Neuro-pathol. Exp. Neurol., 43: 369-375. 1984.
24. Elizan, T., Casals, J., and Yahr, M. J. Antineurofilament antibodies inpostencephalitic and idiopathic Parkinson's disease. Neurosci., 59:341-347,
1983.25. Toh. B., Gibbs, C., Gajdusek, D. C., Goudsmit, J., and Dahl, D. The 200
and 150 kDa neurofilament proteins react with IgG autoantibodies frompatients with Kuru, Creutzfeldt-Jakob disease, and other neurological diseases. Proc. Nati. Acad. Sci. USA, 82: 3485-3489, 1985.
26. Anderton, B., Breinburg, D., Downes, M., Green, P., Tomlinson, B.. Ulrich,J., Wood, J., and Kahn, J. J. Monoclonal antibodies show that neurofibrillarytangles and neurofilaments share antigenic determinants. Nature (Lond.),29«:84-86, 1982.
27. Gajdusek, D. C. Unconventional viruses. In: A. Notkins and M. B. Oldstone(eds.), Concepts in Viral Pathogenesis, pp. 350-357. New York: SpringerVerlag, 1984.
2595
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1986;46:2588-2595. Cancer Res Steven E. Kornguth, Tom Kalinke, Gerald B. Grunwald, et al. SyndromeCell Carcinoma of the Lung and with Visual Paraneoplastic Anti-Neurofilament Antibodies in the Sera of Patients with Small
Updated version
http://cancerres.aacrjournals.org/content/46/5/2588
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/46/5/2588To request permission to re-use all or part of this article, use this link
on June 26, 2018. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























