Embed Size (px)
Citation preview

DOI: 10.1093/jxb/erg149
RESEARCH PAPER
Arbuscular mycorrhizal fungi enhance aluminiumresistance of broomsedge (Andropogon virginicus L.)
Jonathan R. Cumming1 and Jianchang Ning2
Department of Biology, West Virginia University, Morgantown, WV 26506 USA
Received 30 October 2002; Accepted 12 February 2003
Abstract
In the eastern United States, broomsedge (Andropogon
virginicus L.) is found growing on abandoned coal-
mined lands that have extremely acidic soils with high
residual aluminium (Al) concentrations. Broomsedge
may be inherently metal-resistant and nutrient-ef®cient
or may rely on the arbuscular mycorrhizal (AM) fungal
association to overcome limitations on such sites.
Broomsedge plants were grown with and without an
acidic ecotype AM fungal consortium and exposed to
controlled levels of Al in two experiments. The AM fun-
gal consortium conferred Al resistance to broomsedge.
Arbuscular mycorrhizal fungi reduced Al uptake and
translocation in host plants, potentially re¯ecting meas-
ured reductions in inorganic Al availability in the rhizo-
sphere of mycorrhizal plants. Mycorrhizal plants
exhibited lower shoot P concentrations, higher phos-
phorus use ef®ciency, and lower root acid phosphatase
rates than non-mycorrhizal plants. Aluminium signi®-
cantly reduced calcium (Ca) and magnesium (Mg) tissue
concentrations in both mycorrhizal and non-mycorrhizal
plants. However, plant response to any change in nutri-
ent acquisition was substantially less pronounced in
mycorrhizal plants. The exclusion of Al and greater sta-
bility of tissue biomass accretion±tissue nutrient rela-
tionships in mycorrhizal broomsedge plants exposed to
Al may be important mechanisms that allow broom-
sedge to grow on unfavourable acidic soils.
Key words: Acidic soils, calcium, magnesium, nutrient
homeostasis, phosphorus.
Introduction
Broomsedge (Andropogon virginicus L.) is an earlysuccessional species colonizing disturbed sites in theeastern United States (Campbell, 1983). In the coal®elds ofthe Appalachian Mountains, broomsedge dominates aban-doned surface strip-mined areas, even after signi®cantperiods of time have elapsed since mining (Nellessen andUngar, 1993). Depending upon the underlying geology,residual soils on such sites may be extremely acidic,nutrients may be limiting, and soil chemistry may bedominated by high concentrations of aluminium (Al) andiron (Fe) (Zeleznik and Skousen, 1996; Fernandez-Marcoset al., 1998). The early and persistent colonization of suchsites by broomsedge suggests that this species may beinherently nutrient-ef®cient and metal-resistant. In aprevious study, however, Ning and Cumming (2001)found that broomsedge was highly dependent on thesymbiotic association formed with arbuscular mycorrhizal(AM) fungi, and this association played a critical role inincreasing phosphorus use ef®ciency and nutrient home-ostasis of broomsedge plants under phosphorus (P)limitation.
Aluminium (Al) present in acidic soils is toxic to plants(Haug, 1984; Andersson, 1988; Kochian, 1995; Ritchie,1995). Aluminium reduces root growth, alters nutrientavailability in the rhizosphere, and impacts nutrient uptakeand translocation by plants (Roy et al., 1988; Taylor, 1988;Nichol et al., 1993; Matsumoto, 2000). For example, Alreduces inorganic phosphorus (Pi) availability by formingAl±Pi precipitates in the rhizosphere and restricts Ptranslocation within plants (Randall and Vose, 1963;Clarkson, 1966; Cumming et al., 1986; de Miranda andRowell, 1989; Macklon et al., 1994). In addition, Alinterferes with Ca and Mg uptake and translocation inplants (Haug and Caldwell, 1985; Keltjens and Tan, 1993;
1 To whom correspondence should be addressed. Fax: +1 304 293 6363. E-mail: [email protected] Present address: Delaware Biotechnology Institute, Newark, DE 19711, USA.
Journal of Experimental Botany, Vol. 54, No. 386, ã Society for Experimental Biology 2003; all rights reserved
Journal of Experimental Botany, Vol. 54, No. 386, pp. 1447±1459, May 2003
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021

Ryan and Kochian, 1993; Delhaize and Ryan, 1995;Rengel et al., 1995; Lux and Cumming, 2001). Theseeffects result in nutrient imbalances in plants, consequentlyreducing plant growth.
Some plant species and genotypes within species exhibitresistance to Al exposure. Aluminium resistance in plantsmay be associated with an alteration in rhizosphere pH,release of organic acids, or PO4
3± ef¯ux from the roots(Delhaize and Ryan, 1995; Kochian, 1995; Matsumoto,2000). Many of these responses reduce Al3+ availability inthe rhizosphere, consequently reducing Al accumulation intissues (Delhaize et al., 1993; Yang et al., 2000). Someplants also develop speci®c mechanisms to detoxify Alinternally by increasing the production of compounds thatchelate Al intracellularly (Ma et al., 1998).
Arbuscular mycorrhizal (AM) fungi may play a role inthe protection of roots from Al toxicity by mediatinginteractions between Al, Pi, and plant roots (Marschner,1995). Arbuscular mycorrhizal fungi are widely estab-lished in acidic soils (Clark, 1997), and colonization ofplant roots by AM fungi often improves seedling survivaland enhances plant growth on such soils (Danielson,1985). The role of AM fungi in mediating Al toxicity maybe especially pronounced where Al-induced P-de®ciencydominates plant response to Al (Lux and Cumming, 2001).The extensive external fungal hyphae exploit a largevolume of soil and mine scarce nutrient resources that areotherwise unavailable for uptake by roots (Smith andRead, 1997). This effect is most pronounced for Pi (Bolan,1991; Smith and Read, 1997; Ning and Cumming, 2001),although AM fungal colonization may enhance the uptakeof copper (Cu) and zinc (Zn) as well (Ross, 1971;Pacovsky, 1986; Smith and Read, 1997; Lux andCumming, 2001; Ning and Cumming, 2001). In addition,AM fungi may reduce the accumulation of other elements,such as Al, Fe, and manganese (Mn), which are problem-atic in acidic soils (Pacovsky, 1986; Kothari et al., 1991;Lux and Cumming, 2001).
Colonization of roots by AM fungi may confer Alresistance to broomsedge plants colonizing acidic minespoils by facilitating nutrient acquisition for the host plantsunder Al exposure and/or by altering the availability/speciation of Al in the rhizosphere. A limited number ofexperiments have demonstrated that AM fungal coloniza-tion ameliorates Al toxicity in plants (Koslowsky andBoerner, 1989; Medeiros et al., 1994; Mendoza and Borie,1998; Lux and Cumming, 2001), however, it is not clearhow this amelioration is achieved. The present studyinvestigated the response of broomsedge plants to Al andthe in¯uence of AM fungi on the growth and nutrition ofplants exposed to Al. A community of AM fungi, includingGlomus clarum (Nicolson & Schenck) and Gigasporagigantea ((Nicol. & Gerd.) Gerd. & Trappe), isolated froman abandoned strip mine was used as the inoculum source.Two experiments were undertaken. The ®rst focused on
the in¯uence of AM fungi and Al on Pi relations ofbroomsedge plants at low to moderate Al exposures. Thesecond experiment spanned a greater concentration rangeof Al and focused on mycorrhizal mediation of Al-inducednutrient perturbations in broomsedge. It was hypothesizedthat mycorrhizal and non-mycorrhizal plants would differin their responses to Al in the rhizosphere and that suchdifferences would result from reduced Al±Pi interactionsand the maintenance of overall nutrient acquisitionmediated by AM fungal colonization.
Materials and methods
Preparation of AM fungal inoculum
Arbuscular mycorrhizal fungal inoculum was generated frombroomsedge plants collected from an abandoned coal mine inField Crest, near Morgantown, West Virginia, USA. Glomus clarumwas the primary fungal species, with Gigaspora gigantea alsopresent. Although the site has been abandoned for approximately 50years, much of the surface soil remains devoid of vegetation due tothe harsh chemical conditions. The vegetated surface consists of abroomsedge sward surrounding stunted red maple (Acer rubrum L.)and big-toothed aspen (Populus grandidentata Michx.) trees. Soilfrom the site has a pH of 3.0±3.3 (soil±water paste) and containedMehlich-extractable Al of 363 mg kg±1 soil. The inorganicmonomeric Al concentration in surface water at the site wasc. 600 mM, as measured by the eriochrome cyanine method(Anonymous, 1985; Lux and Cumming, 2001).
To produce AM fungal inoculum, broomsedge plants with intactmycorrhizal roots and adhering soil from the mine site weretransplanted into 15 cm diameter pots containing a mixture ofautoclaved mine-soil and sand (1:3 v/v). After 1 month, the potcontents became the source of inoculum for the experiments. Thesepots are termed `nursery pots' hereafter. A set of axenicallygerminated broomsedge plants grown in the same soil±sand mixturewith a bacterial extract from roots of ®eld-collected plants served asa source of inoculum for non-mycorrhizal treatments.
Preparation of plants
Broomsedge seeds were sown around the perimeter of bothmycorrhizal and non-mycorrhizal nursery pots. After 4 weeksgrowth, roots of a small subset of seedlings (c. 10) were examined todetermine mycorrhizal status. Seedlings were well colonized(7267%). Tissue P status was analysed after wet digestion inconcentrated H2SO4/30% H2O2 (Parkinson and Allen, 1975) by themolybdate blue method (Olsen and Sommers, 1982). Tissuephosphorus concentrations and tissue dry weights did not differsigni®cantly between mycorrhizal and non-mycorrhizal plants (Ningand Cumming, 2001).
Sand culture and growth conditions
Mycorrhizal and non-mycorrhizal broomsedge seedlings weretransplanted into and grown in D16 Deepots (5 cm diameter 3 18 cmheight) (Stuewe and Sons, Inc., Corvallis, Oregon, USA) pre®lledwith 220 cm3 of a 3:1 (v/v) mixture of coarse:®ne acid-washed sand.Deepots were placed into a growth chamber with 14 h of light at 28°C, 60% RH, and 10 h of darkness at 21 °C, 50% RH. Average lightintensity at pot height in the chamber was 260 mmol m±2 s±1 frommixed ¯uorescent and incandescent sources. Plants received abaseline nutrient solution containing NO3 (1.5 mM), NH4 (0.5 mM),
1448 Cumming and Ning
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021

K (0.79 mM), Ca (0.675 mM), Mg (0.25 mM), SO4 (0.25 mM),H2PO4 (40 mM), Fe (25 mM), B (23.14 mM), Mn (4.57 mM), Zn (0.38mM), Cu (0.16 mM), and Mo (0.06 mM). The pH of this formulationwas c. 6.5 prior to adjustment to 4.0. For Al exposures, this solutionwas modi®ed as indicated by the addition of Al as AlCl3. Allsolutions were adjusted to pH 4.0 after Al was added and beforeapplication. Aluminium concentrations in the delivered solutionswere within 10% of target concentrations as measured by theeriochrome cyanine method for inorganic monomeric Al. Solutions(approximately 15 ml) were automatically delivered to the plantsthree times each day for 8 weeks.
Experiment 1: impacts of Al on Pi relationships ofmycorrhizal broomsedge
Aluminium was delivered to broomsedge plants at concentrations of0, 10, 50, 100, and 200 mM Al. Chemical speciation analysis of thesesolutions by the program GEOCHEM (Parker et al., 1995) indicatedthat Al3+ concentrations were 0, 5.72, 29.91, 62.75, and 133.90 mMin the treatment solutions, respectively. For data analysis hereafter,delivered Al concentrations are presented.
Root zone pH and Al concentration were measured at weeks 4, 6,and 8 by collecting leachate ¯owing from the deepots followingdelivery of nutrient solutions. Solution Al was determined using theeriochrome cyanine method. Plants were harvested following 8weeks exposure to Al. At harvest, plants were gently removed fromdeepots and roots were excised from shoots. Shoots were rinsed indeionized H2O and roots were rinsed free of sand with deionizedH2O. To determine acid phosphatase (APase) activity, ®ve 8 cmroots were removed from the root system, washed in deionizedwater, cut into 1 cm pieces, and transferred into centrifuge tubescontaining 4.5 ml of the appropriate nutrient solution containing Al.Then, 0.5 ml of 1 mM p-nitrophenylphosphate (NPP) was added toeach tube and tubes were incubated for 1 h at room temperature.After the incubation period, 1 ml of 0.5 N NaOH was added to eachtube. Acid phosphatase activity (nitrophenol produced) was spectro-photometrically determined (Tabatabai and Bremner, 1969).
Percentage mycorrhizal colonization of roots was assessed on a5% fresh mass sample from the youngest portion of the root system.This region was sampled in order to assess the potential impacts ofAl on the colonization of newly produced roots. These root sampleswere cleared in boiling 10% KOH for 5 min, neutralized by 2% HClfor 2 min, and stained with 0.05% trypan blue for 5 min. Roots werestored in cold H2O while they were assessed for mycorrhizalcolonization by the gridline-intersect method (Giovannetti andMosse, 1980).
Shoots and remaining roots were dried at 60 °C, weighed, andground in a Wiley mill. Ground roots and shoots were digested and Pand Al concentrations of the digests were determined spectro-photometrically using the molybdate blue and eriochrome cyaninemethods, respectively.
Experiment 2: high Al concentration dose response
Aluminium was delivered to broomsedge plants at concentrations of0, 200, 400, and 1000 mM Al. Chemical speciation analysis of thesesolutions by the program GEOCHEM indicated that Al3+ concen-trations were 0, 133.9, 379.4, and 945.8 mM in the solutions,respectively. At the termination of the experiment following 8 weeksof treatment, plants were separated into roots and shoots. The tillernumber of each plant was counted and the cumulative length of allthese tillers was recorded. Shoots were rinsed in deionized water.Sand was removed from the roots under running deionized water.Mycorrhizal colonization of roots was assessed on a 5% fresh massroot subsample as noted above. Shoots and the remaining roots weredried at 60 °C, weighed, ground, and digested as noted above. Theconcentration of P in the digests was determined as noted above.
Other mineral elements (Al, K, Ca, Mg, Fe, Mn, Zn, Cu) in thedigests were analysed by inductively coupled plasma (ICP) emissionspectrophotometry by the National Research Center for Coal andEnergy Analytical Laboratory at West Virginia University.
Data calculation and analysis
Phosphorus use ef®ciency (PUE) (Boan et al., 1993) was calculatedas:
(plant dry weight)/(plant P content)
The Al±Pi experiment was a 235 (mycorrhizal-by-Al) factorialdesign with ®ve replicate plants for each treatment combination.Plant mass data were not normally distributed according to theShapiro±Wilk W Test and were natural-log transformed prior toanalysis of variance (ANOVA). Other measures (leachate pH and Alconcentration, APase, PUE) were analysed by ANOVA. The high Alconcentration dose response experiment was a blocked 234(mycorrhizal-by-Al) factorial design with 10 replicate plants foreach treatment combination. Blocks accounted for environmentalvariation within the growth chamber environment. Plant growth dataover this wider Al concentration range were normally distributed, aswere tissue nutrient data. Treatment means within mycorrhizaltreatments were compared using the Tukey±Kramer HSD procedure.Analysis of covariance (ANCOVA) was used to investigate therelationships between growth and tissue element concentrations.Data were analysed using the statistical package JMP (SAS Institute,Cary, North Carolina, USA).
Results
Impacts of Al on Pi relationships of mycorrhizalbroomsedge
The leachate pH of solutions from mycorrhizal and non-mycorrhizal plants was higher under low Al exposures, andpH from mycorrhizal plants was higher than non-mycorrhizal plants at higher Al concentrations (P=0.006for the mycorrhiza-by-Al interaction) (Fig. 1A). Theleachate pH of most treatments was below 4.5 and, atthis pH, Al would exist primarily as Al3+ in the root zonesof both mycorrhizal and non-mycorrhizal broomsedgeplants (Snoeyink and Jenkins, 1980). However, the analy-sis of leachate Al concentrations indicated that inorganicmonomeric Al in the rhizosphere of mycorrhizal broom-sedge plants was consistently less than that measured fornon-mycorrhizal plants (P <0.001) (Fig. 1B).
The mean percentage colonization of inoculatedbroomsedge plants was 60% across all Al treatments andthe rate of colonization was unaffected by Al (P=0.711)(data not presented). None of the non-mycorrhizal plantswere colonized by mycorrhizal fungi.
Mycorrhizal and non-mycorrhizal plants diverged intheir growth responses to Al in solution (Fig. 2). Following8 weeks of Al exposure, shoot and root growth ofmycorrhizal plants were stimulated by low Al concentra-tions up to 50 mM and were unaffected by higherconcentrations of Al in solution. In non-mycorrhizal plantsshoot and root mass were signi®cantly reduced by Alabove 50 mM, with reductions of up to 47% and 57% at 200mM Al, respectively (P=0.031 and 0.033 for the mycor-
Mycorrhizae and Al resistance in Andropogon 1449
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021
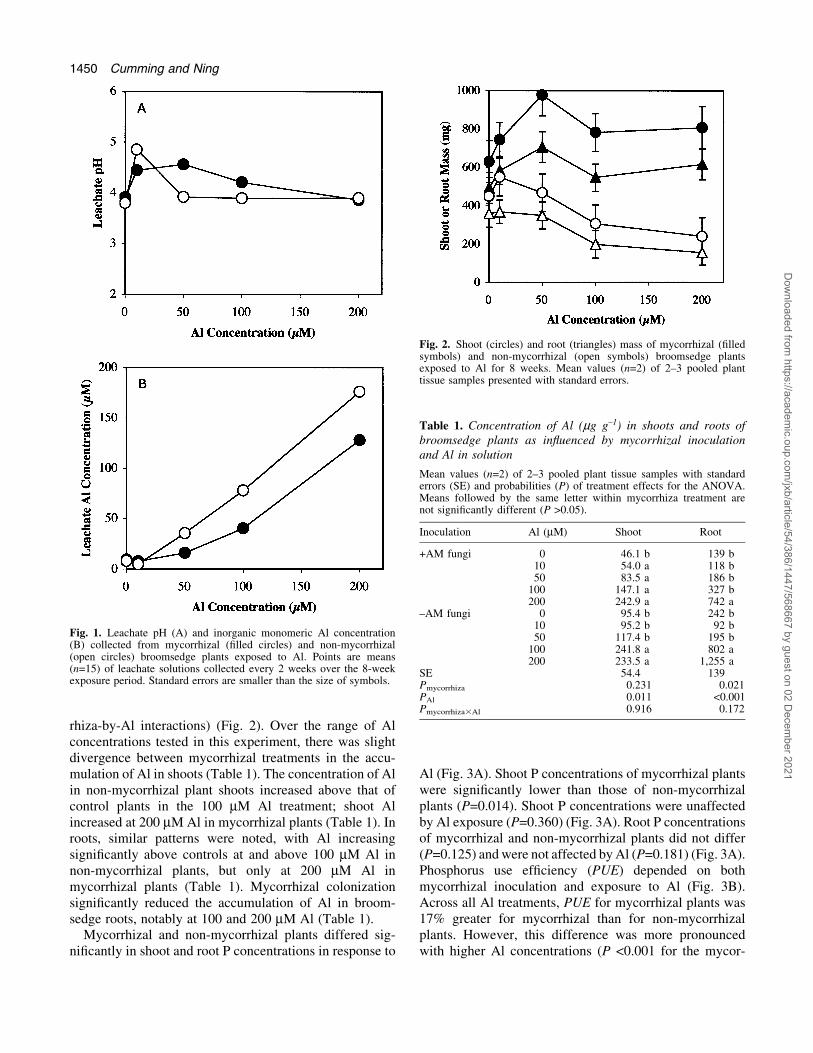
rhiza-by-Al interactions) (Fig. 2). Over the range of Alconcentrations tested in this experiment, there was slightdivergence between mycorrhizal treatments in the accu-mulation of Al in shoots (Table 1). The concentration of Alin non-mycorrhizal plant shoots increased above that ofcontrol plants in the 100 mM Al treatment; shoot Alincreased at 200 mM Al in mycorrhizal plants (Table 1). Inroots, similar patterns were noted, with Al increasingsigni®cantly above controls at and above 100 mM Al innon-mycorrhizal plants, but only at 200 mM Al inmycorrhizal plants (Table 1). Mycorrhizal colonizationsigni®cantly reduced the accumulation of Al in broom-sedge roots, notably at 100 and 200 mM Al (Table 1).
Mycorrhizal and non-mycorrhizal plants differed sig-ni®cantly in shoot and root P concentrations in response to
Al (Fig. 3A). Shoot P concentrations of mycorrhizal plantswere signi®cantly lower than those of non-mycorrhizalplants (P=0.014). Shoot P concentrations were unaffectedby Al exposure (P=0.360) (Fig. 3A). Root P concentrationsof mycorrhizal and non-mycorrhizal plants did not differ(P=0.125) and were not affected by Al (P=0.181) (Fig. 3A).Phosphorus use ef®ciency (PUE) depended on bothmycorrhizal inoculation and exposure to Al (Fig. 3B).Across all Al treatments, PUE for mycorrhizal plants was17% greater for mycorrhizal than for non-mycorrhizalplants. However, this difference was more pronouncedwith higher Al concentrations (P <0.001 for the mycor-
Fig. 1. Leachate pH (A) and inorganic monomeric Al concentration(B) collected from mycorrhizal (®lled circles) and non-mycorrhizal(open circles) broomsedge plants exposed to Al. Points are means(n=15) of leachate solutions collected every 2 weeks over the 8-weekexposure period. Standard errors are smaller than the size of symbols.
Fig. 2. Shoot (circles) and root (triangles) mass of mycorrhizal (®lledsymbols) and non-mycorrhizal (open symbols) broomsedge plantsexposed to Al for 8 weeks. Mean values (n=2) of 2±3 pooled planttissue samples presented with standard errors.
Table 1. Concentration of Al (mg g±1) in shoots and roots ofbroomsedge plants as in¯uenced by mycorrhizal inoculationand Al in solution
Mean values (n=2) of 2±3 pooled plant tissue samples with standarderrors (SE) and probabilities (P) of treatment effects for the ANOVA.Means followed by the same letter within mycorrhiza treatment arenot signi®cantly different (P >0.05).
Inoculation Al (mM) Shoot Root
+AM fungi 0 46.1 b 139 b10 54.0 a 118 b50 83.5 a 186 b
100 147.1 a 327 b200 242.9 a 742 a
±AM fungi 0 95.4 b 242 b10 95.2 b 92 b50 117.4 b 195 b
100 241.8 a 802 a200 233.5 a 1,255 a
SE 54.4 139Pmycorrhiza 0.231 0.021PAl 0.011 <0.001Pmycorrhiza3Al 0.916 0.172
1450 Cumming and Ning
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021
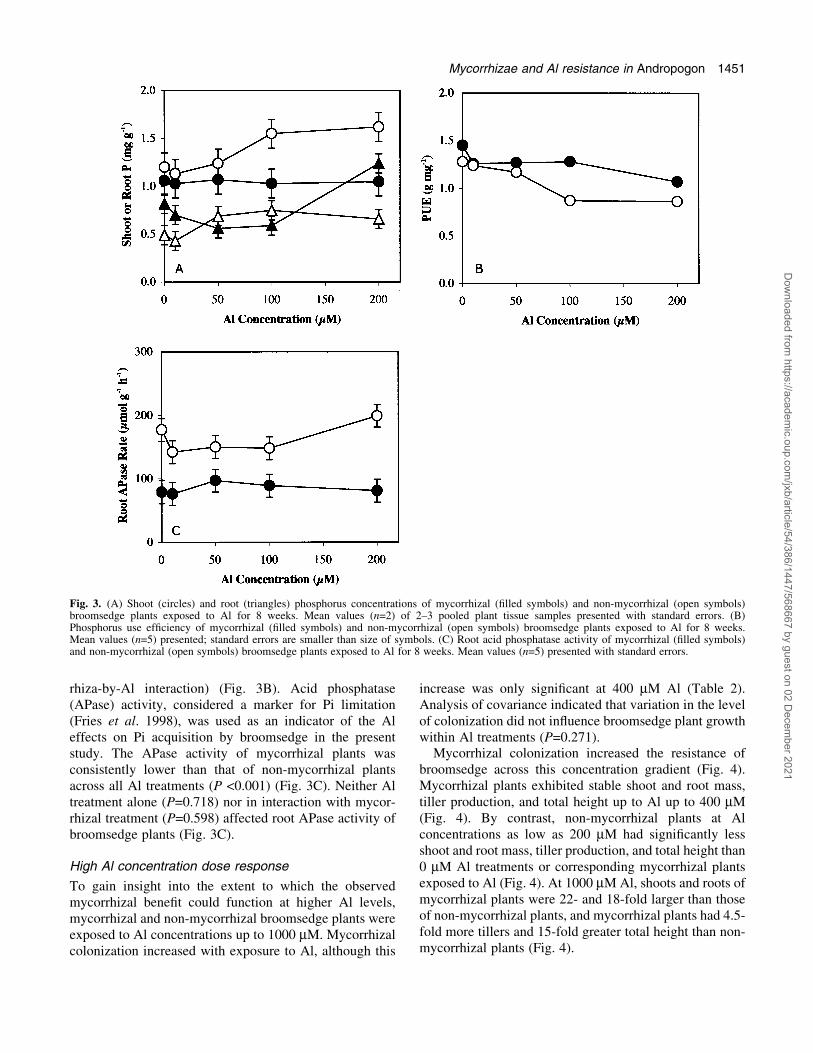
rhiza-by-Al interaction) (Fig. 3B). Acid phosphatase(APase) activity, considered a marker for Pi limitation(Fries et al. 1998), was used as an indicator of the Aleffects on Pi acquisition by broomsedge in the presentstudy. The APase activity of mycorrhizal plants wasconsistently lower than that of non-mycorrhizal plantsacross all Al treatments (P <0.001) (Fig. 3C). Neither Altreatment alone (P=0.718) nor in interaction with mycor-rhizal treatment (P=0.598) affected root APase activity ofbroomsedge plants (Fig. 3C).
High Al concentration dose response
To gain insight into the extent to which the observedmycorrhizal bene®t could function at higher Al levels,mycorrhizal and non-mycorrhizal broomsedge plants wereexposed to Al concentrations up to 1000 mM. Mycorrhizalcolonization increased with exposure to Al, although this
increase was only signi®cant at 400 mM Al (Table 2).Analysis of covariance indicated that variation in the levelof colonization did not in¯uence broomsedge plant growthwithin Al treatments (P=0.271).
Mycorrhizal colonization increased the resistance ofbroomsedge across this concentration gradient (Fig. 4).Mycorrhizal plants exhibited stable shoot and root mass,tiller production, and total height up to Al up to 400 mM(Fig. 4). By contrast, non-mycorrhizal plants at Alconcentrations as low as 200 mM had signi®cantly lessshoot and root mass, tiller production, and total height than0 mM Al treatments or corresponding mycorrhizal plantsexposed to Al (Fig. 4). At 1000 mM Al, shoots and roots ofmycorrhizal plants were 22- and 18-fold larger than thoseof non-mycorrhizal plants, and mycorrhizal plants had 4.5-fold more tillers and 15-fold greater total height than non-mycorrhizal plants (Fig. 4).
Fig. 3. (A) Shoot (circles) and root (triangles) phosphorus concentrations of mycorrhizal (®lled symbols) and non-mycorrhizal (open symbols)broomsedge plants exposed to Al for 8 weeks. Mean values (n=2) of 2±3 pooled plant tissue samples presented with standard errors. (B)Phosphorus use ef®ciency of mycorrhizal (®lled symbols) and non-mycorrhizal (open symbols) broomsedge plants exposed to Al for 8 weeks.Mean values (n=5) presented; standard errors are smaller than size of symbols. (C) Root acid phosphatase activity of mycorrhizal (®lled symbols)and non-mycorrhizal (open symbols) broomsedge plants exposed to Al for 8 weeks. Mean values (n=5) presented with standard errors.
Mycorrhizae and Al resistance in Andropogon 1451
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021
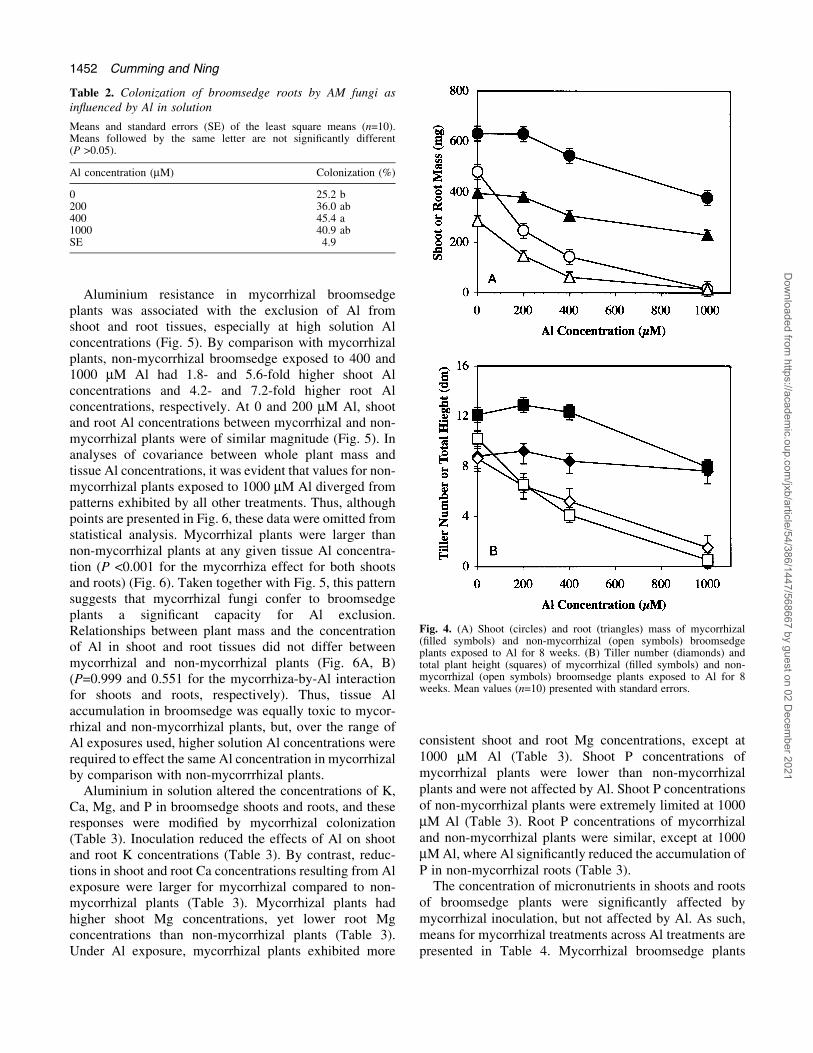
Aluminium resistance in mycorrhizal broomsedgeplants was associated with the exclusion of Al fromshoot and root tissues, especially at high solution Alconcentrations (Fig. 5). By comparison with mycorrhizalplants, non-mycorrhizal broomsedge exposed to 400 and1000 mM Al had 1.8- and 5.6-fold higher shoot Alconcentrations and 4.2- and 7.2-fold higher root Alconcentrations, respectively. At 0 and 200 mM Al, shootand root Al concentrations between mycorrhizal and non-mycorrhizal plants were of similar magnitude (Fig. 5). Inanalyses of covariance between whole plant mass andtissue Al concentrations, it was evident that values for non-mycorrhizal plants exposed to 1000 mM Al diverged frompatterns exhibited by all other treatments. Thus, althoughpoints are presented in Fig. 6, these data were omitted fromstatistical analysis. Mycorrhizal plants were larger thannon-mycorrhizal plants at any given tissue Al concentra-tion (P <0.001 for the mycorrhiza effect for both shootsand roots) (Fig. 6). Taken together with Fig. 5, this patternsuggests that mycorrhizal fungi confer to broomsedgeplants a signi®cant capacity for Al exclusion.Relationships between plant mass and the concentrationof Al in shoot and root tissues did not differ betweenmycorrhizal and non-mycorrhizal plants (Fig. 6A, B)(P=0.999 and 0.551 for the mycorrhiza-by-Al interactionfor shoots and roots, respectively). Thus, tissue Alaccumulation in broomsedge was equally toxic to mycor-rhizal and non-mycorrhizal plants, but, over the range ofAl exposures used, higher solution Al concentrations wererequired to effect the same Al concentration in mycorrhizalby comparison with non-mycorrrhizal plants.
Aluminium in solution altered the concentrations of K,Ca, Mg, and P in broomsedge shoots and roots, and theseresponses were modi®ed by mycorrhizal colonization(Table 3). Inoculation reduced the effects of Al on shootand root K concentrations (Table 3). By contrast, reduc-tions in shoot and root Ca concentrations resulting from Alexposure were larger for mycorrhizal compared to non-mycorrhizal plants (Table 3). Mycorrhizal plants hadhigher shoot Mg concentrations, yet lower root Mgconcentrations than non-mycorrhizal plants (Table 3).Under Al exposure, mycorrhizal plants exhibited more
consistent shoot and root Mg concentrations, except at1000 mM Al (Table 3). Shoot P concentrations ofmycorrhizal plants were lower than non-mycorrhizalplants and were not affected by Al. Shoot P concentrationsof non-mycorrhizal plants were extremely limited at 1000mM Al (Table 3). Root P concentrations of mycorrhizaland non-mycorrhizal plants were similar, except at 1000mM Al, where Al signi®cantly reduced the accumulation ofP in non-mycorrhizal roots (Table 3).
The concentration of micronutrients in shoots and rootsof broomsedge plants were signi®cantly affected bymycorrhizal inoculation, but not affected by Al. As such,means for mycorrhizal treatments across Al treatments arepresented in Table 4. Mycorrhizal broomsedge plants
Table 2. Colonization of broomsedge roots by AM fungi asin¯uenced by Al in solution
Means and standard errors (SE) of the least square means (n=10).Means followed by the same letter are not signi®cantly different(P >0.05).
Al concentration (mM) Colonization (%)
0 25.2 b200 36.0 ab400 45.4 a1000 40.9 abSE 4.9
Fig. 4. (A) Shoot (circles) and root (triangles) mass of mycorrhizal(®lled symbols) and non-mycorrhizal (open symbols) broomsedgeplants exposed to Al for 8 weeks. (B) Tiller number (diamonds) andtotal plant height (squares) of mycorrhizal (®lled symbols) and non-mycorrhizal (open symbols) broomsedge plants exposed to Al for 8weeks. Mean values (n=10) presented with standard errors.
1452 Cumming and Ning
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021
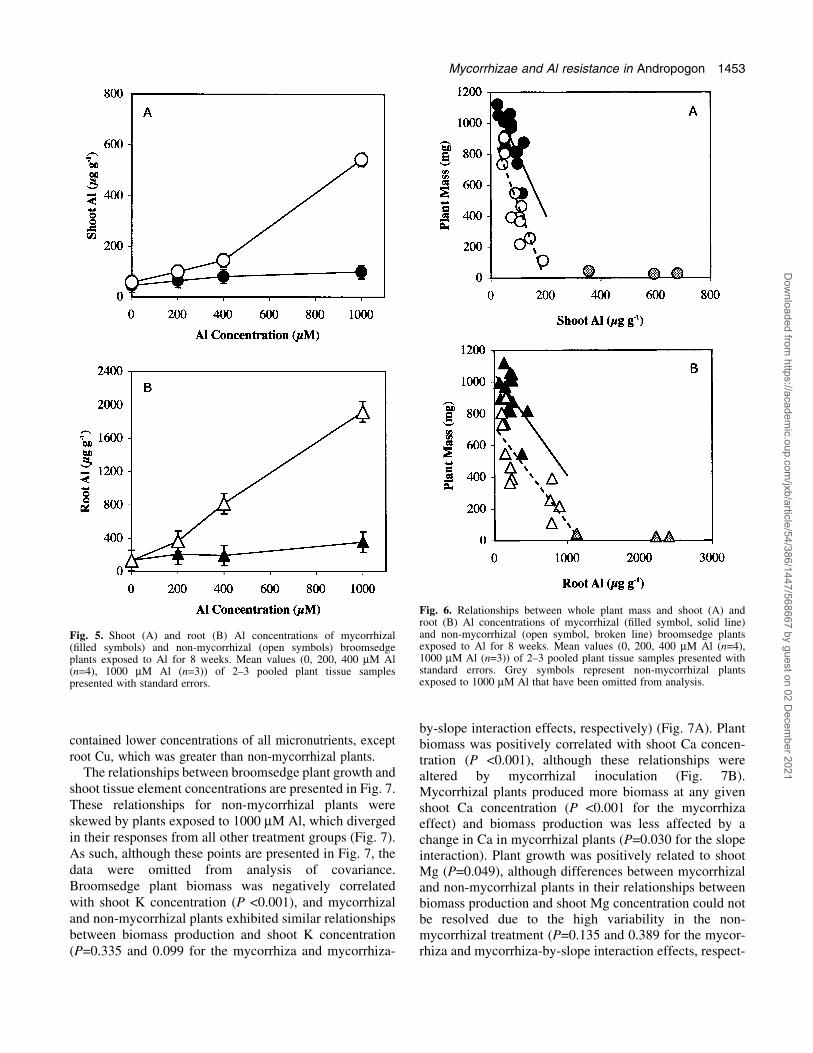
contained lower concentrations of all micronutrients, exceptroot Cu, which was greater than non-mycorrhizal plants.
The relationships between broomsedge plant growth andshoot tissue element concentrations are presented in Fig. 7.These relationships for non-mycorrhizal plants wereskewed by plants exposed to 1000 mM Al, which divergedin their responses from all other treatment groups (Fig. 7).As such, although these points are presented in Fig. 7, thedata were omitted from analysis of covariance.Broomsedge plant biomass was negatively correlatedwith shoot K concentration (P <0.001), and mycorrhizaland non-mycorrhizal plants exhibited similar relationshipsbetween biomass production and shoot K concentration(P=0.335 and 0.099 for the mycorrhiza and mycorrhiza-
by-slope interaction effects, respectively) (Fig. 7A). Plantbiomass was positively correlated with shoot Ca concen-tration (P <0.001), although these relationships werealtered by mycorrhizal inoculation (Fig. 7B).Mycorrhizal plants produced more biomass at any givenshoot Ca concentration (P <0.001 for the mycorrhizaeffect) and biomass production was less affected by achange in Ca in mycorrhizal plants (P=0.030 for the slopeinteraction). Plant growth was positively related to shootMg (P=0.049), although differences between mycorrhizaland non-mycorrhizal plants in their relationships betweenbiomass production and shoot Mg concentration could notbe resolved due to the high variability in the non-mycorrhizal treatment (P=0.135 and 0.389 for the mycor-rhiza and mycorrhiza-by-slope interaction effects, respect-
Fig. 5. Shoot (A) and root (B) Al concentrations of mycorrhizal(®lled symbols) and non-mycorrhizal (open symbols) broomsedgeplants exposed to Al for 8 weeks. Mean values (0, 200, 400 mM Al(n=4), 1000 mM Al (n=3)) of 2±3 pooled plant tissue samplespresented with standard errors.
Fig. 6. Relationships between whole plant mass and shoot (A) androot (B) Al concentrations of mycorrhizal (®lled symbol, solid line)and non-mycorrhizal (open symbol, broken line) broomsedge plantsexposed to Al for 8 weeks. Mean values (0, 200, 400 mM Al (n=4),1000 mM Al (n=3)) of 2±3 pooled plant tissue samples presented withstandard errors. Grey symbols represent non-mycorrhizal plantsexposed to 1000 mM Al that have been omitted from analysis.
Mycorrhizae and Al resistance in Andropogon 1453
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021
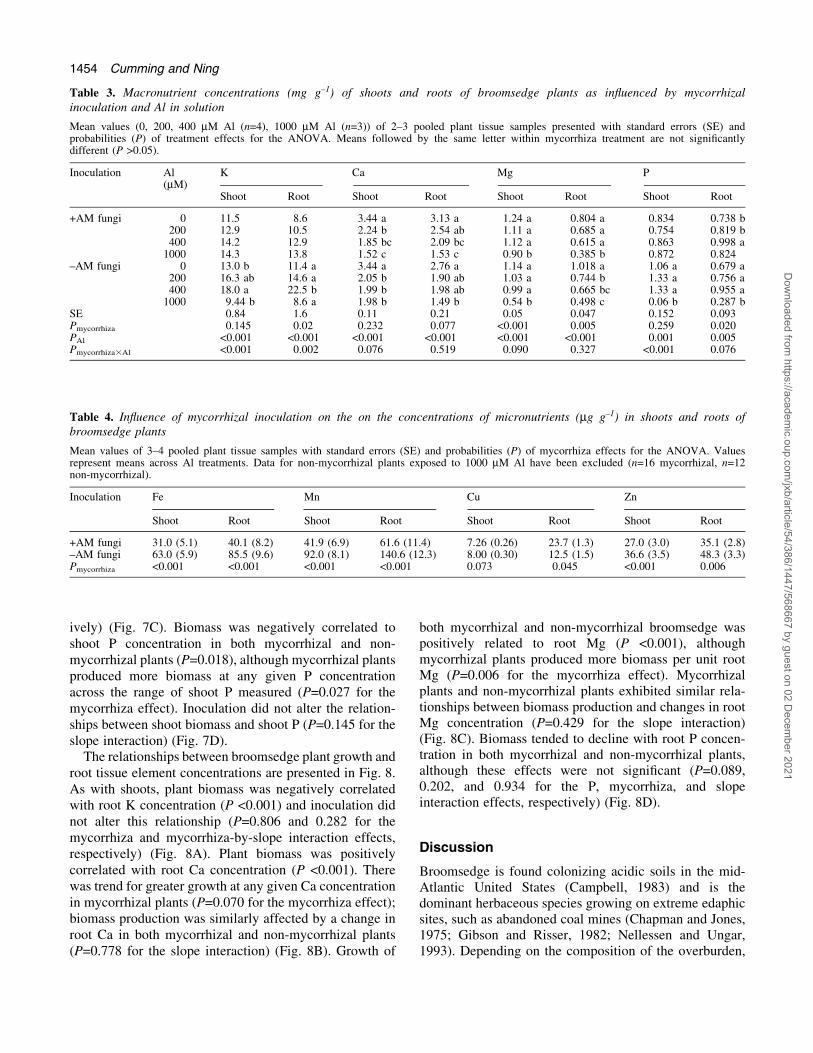
ively) (Fig. 7C). Biomass was negatively correlated toshoot P concentration in both mycorrhizal and non-mycorrhizal plants (P=0.018), although mycorrhizal plantsproduced more biomass at any given P concentrationacross the range of shoot P measured (P=0.027 for themycorrhiza effect). Inoculation did not alter the relation-ships between shoot biomass and shoot P (P=0.145 for theslope interaction) (Fig. 7D).
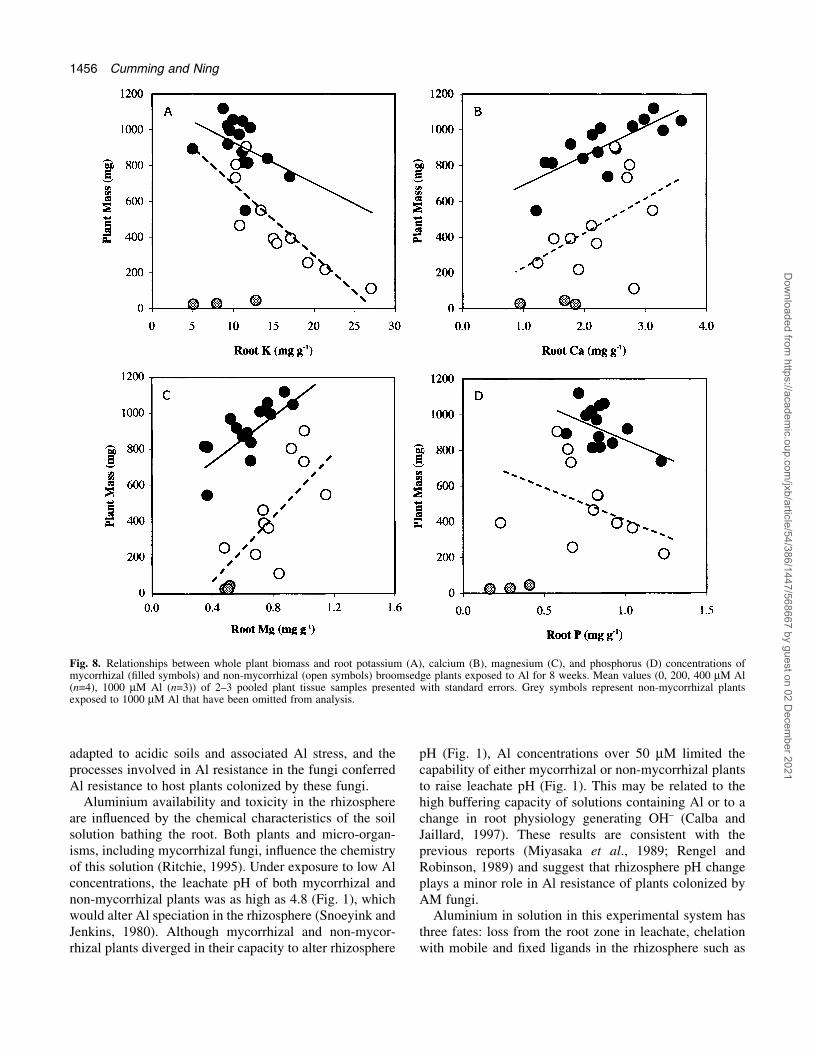
The relationships between broomsedge plant growth androot tissue element concentrations are presented in Fig. 8.As with shoots, plant biomass was negatively correlatedwith root K concentration (P <0.001) and inoculation didnot alter this relationship (P=0.806 and 0.282 for themycorrhiza and mycorrhiza-by-slope interaction effects,respectively) (Fig. 8A). Plant biomass was positivelycorrelated with root Ca concentration (P <0.001). Therewas trend for greater growth at any given Ca concentrationin mycorrhizal plants (P=0.070 for the mycorrhiza effect);biomass production was similarly affected by a change inroot Ca in both mycorrhizal and non-mycorrhizal plants(P=0.778 for the slope interaction) (Fig. 8B). Growth of
both mycorrhizal and non-mycorrhizal broomsedge waspositively related to root Mg (P <0.001), althoughmycorrhizal plants produced more biomass per unit rootMg (P=0.006 for the mycorrhiza effect). Mycorrhizalplants and non-mycorrhizal plants exhibited similar rela-tionships between biomass production and changes in rootMg concentration (P=0.429 for the slope interaction)(Fig. 8C). Biomass tended to decline with root P concen-tration in both mycorrhizal and non-mycorrhizal plants,although these effects were not signi®cant (P=0.089,0.202, and 0.934 for the P, mycorrhiza, and slopeinteraction effects, respectively) (Fig. 8D).
Discussion
Broomsedge is found colonizing acidic soils in the mid-Atlantic United States (Campbell, 1983) and is thedominant herbaceous species growing on extreme edaphicsites, such as abandoned coal mines (Chapman and Jones,1975; Gibson and Risser, 1982; Nellessen and Ungar,1993). Depending on the composition of the overburden,
Table 3. Macronutrient concentrations (mg g±1) of shoots and roots of broomsedge plants as in¯uenced by mycorrhizalinoculation and Al in solution
Mean values (0, 200, 400 mM Al (n=4), 1000 mM Al (n=3)) of 2±3 pooled plant tissue samples presented with standard errors (SE) andprobabilities (P) of treatment effects for the ANOVA. Means followed by the same letter within mycorrhiza treatment are not signi®cantlydifferent (P >0.05).
Inoculation Al K Ca Mg P(mM)
Shoot Root Shoot Root Shoot Root Shoot Root
+AM fungi 0 11.5 8.6 3.44 a 3.13 a 1.24 a 0.804 a 0.834 0.738 b200 12.9 10.5 2.24 b 2.54 ab 1.11 a 0.685 a 0.754 0.819 b400 14.2 12.9 1.85 bc 2.09 bc 1.12 a 0.615 a 0.863 0.998 a
1000 14.3 13.8 1.52 c 1.53 c 0.90 b 0.385 b 0.872 0.824±AM fungi 0 13.0 b 11.4 a 3.44 a 2.76 a 1.14 a 1.018 a 1.06 a 0.679 a
200 16.3 ab 14.6 a 2.05 b 1.90 ab 1.03 a 0.744 b 1.33 a 0.756 a400 18.0 a 22.5 b 1.99 b 1.98 ab 0.99 a 0.665 bc 1.33 a 0.955 a
1000 9.44 b 8.6 a 1.98 b 1.49 b 0.54 b 0.498 c 0.06 b 0.287 bSE 0.84 1.6 0.11 0.21 0.05 0.047 0.152 0.093Pmycorrhiza 0.145 0.02 0.232 0.077 <0.001 0.005 0.259 0.020PAl <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 0.001 0.005Pmycorrhiza3Al <0.001 0.002 0.076 0.519 0.090 0.327 <0.001 0.076
Table 4. In¯uence of mycorrhizal inoculation on the on the concentrations of micronutrients (mg g±1) in shoots and roots ofbroomsedge plants
Mean values of 3±4 pooled plant tissue samples with standard errors (SE) and probabilities (P) of mycorrhiza effects for the ANOVA. Valuesrepresent means across Al treatments. Data for non-mycorrhizal plants exposed to 1000 mM Al have been excluded (n=16 mycorrhizal, n=12non-mycorrhizal).
Inoculation Fe Mn Cu Zn
Shoot Root Shoot Root Shoot Root Shoot Root
+AM fungi 31.0 (5.1) 40.1 (8.2) 41.9 (6.9) 61.6 (11.4) 7.26 (0.26) 23.7 (1.3) 27.0 (3.0) 35.1 (2.8)±AM fungi 63.0 (5.9) 85.5 (9.6) 92.0 (8.1) 140.6 (12.3) 8.00 (0.30) 12.5 (1.5) 36.6 (3.5) 48.3 (3.3)Pmycorrhiza <0.001 <0.001 <0.001 <0.001 0.073 0.045 <0.001 0.006
1454 Cumming and Ning
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021
the chemistry of such soils may be dominated byphytotoxic Al. Broomsedge may thus be inherentlyresistant to Al or may rely on symbiotic AM fungi toovercome edaphic limitations on such sites.
The Al resistance of broomsedge and the in¯uence ofAM fungi on the response of broomsedge to Al in therhizosphere was investigated. Inoculum for this study wascollected from an acidic (pH=3.0) coal mine, where Aldominated both soil (360 mg kg±1 soil) and soil water (600mM) chemistry. Although broomsedge is the dominantvegetation on the site and many others like it in the region,the present study indicates that broomsedge is not tolerantof Al at concentrations noted in these soils, and exhibited
signi®cant reductions in growth above 50 mM Al (Figs 2,4). However, the colonization of roots by AM fungiconferred Al resistance to broomsedge plants (Figs 2, 4), ashas been demonstrated previously for several other plantspecies (Medeiros et al., 1994; Mendoza and Borie, 1998;Lux and Cumming, 2001). Mycorrhizal broomsedge plantsdid not exhibit any Al toxicity symptoms up to 200 mM Al,and showed stimulated growth at low Al concentrations(Fig. 2). When solution Al was 400 mM and greater,mycorrhizal broomsedge plants exhibited only slightgrowth reductions, whereas the growth of non-mycorrhizalplants was severely limited (Fig. 4). This indicates that thenative AM fungal consortium used in the present study is
Fig. 7. Relationships between whole plant biomass and shoot potassium (A), calcium (B), magnesium (C), and phosphorus (D) concentrations ofmycorrhizal (®lled symbols) and non-mycorrhizal (open symbols) broomsedge plants exposed to Al for 8 weeks. Mean values (0, 200, 400 mM Al(n=4), 1000 mM Al (n=3)) of 2±3 pooled plant tissue samples presented with standard errors. Grey symbols represent non-mycorrhizal plantsexposed to 1000 mM Al that have been omitted from analysis.
Mycorrhizae and Al resistance in Andropogon 1455
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021
adapted to acidic soils and associated Al stress, and theprocesses involved in Al resistance in the fungi conferredAl resistance to host plants colonized by these fungi.
Aluminium availability and toxicity in the rhizosphereare in¯uenced by the chemical characteristics of the soilsolution bathing the root. Both plants and micro-organ-isms, including mycorrhizal fungi, in¯uence the chemistryof this solution (Ritchie, 1995). Under exposure to low Alconcentrations, the leachate pH of both mycorrhizal andnon-mycorrhizal plants was as high as 4.8 (Fig. 1), whichwould alter Al speciation in the rhizosphere (Snoeyink andJenkins, 1980). Although mycorrhizal and non-mycor-rhizal plants diverged in their capacity to alter rhizosphere
pH (Fig. 1), Al concentrations over 50 mM limited thecapability of either mycorrhizal or non-mycorrhizal plantsto raise leachate pH (Fig. 1). This may be related to thehigh buffering capacity of solutions containing Al or to achange in root physiology generating OH± (Calba andJaillard, 1997). These results are consistent with theprevious reports (Miyasaka et al., 1989; Rengel andRobinson, 1989) and suggest that rhizosphere pH changeplays a minor role in Al resistance of plants colonized byAM fungi.
Aluminium in solution in this experimental system hasthree fates: loss from the root zone in leachate, chelationwith mobile and ®xed ligands in the rhizosphere such as
Fig. 8. Relationships between whole plant biomass and root potassium (A), calcium (B), magnesium (C), and phosphorus (D) concentrations ofmycorrhizal (®lled symbols) and non-mycorrhizal (open symbols) broomsedge plants exposed to Al for 8 weeks. Mean values (0, 200, 400 mM Al(n=4), 1000 mM Al (n=3)) of 2±3 pooled plant tissue samples presented with standard errors. Grey symbols represent non-mycorrhizal plantsexposed to 1000 mM Al that have been omitted from analysis.
1456 Cumming and Ning
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021

root exudates and cell walls, and uptake by the plant. Overthe course of the ®rst experiment, less inorganic mono-meric Al was detected in leachate collected frommycorrhizal than non-mycorrhizal plants (Fig. 1).Although the fate of this Al is not known for certain, lessAl was detected in mycorrhizal plant tissues and Al mayhave been chelated in rhizosphere, for example, by organicacids (Kochian, 1995), or bound to fungal cell walls in thesand matrix surrounding mycorrhizal plant roots. Areduction in available Al in the rhizosphere of mycorrhizalplants would contribute to limited Al accumulation in bothshoots and roots of mycorrhizal plants observed in bothexperiments (Table 1; Fig. 5). This suppression of Aluptake and translocation to shoots of mycorrhizal plants isconsistent with most previous reports (Koslowsky andBoerner, 1989; Medeiros et al., 1994; Mendoza and Borie,1998).
Aluminium-induced organic acid exudation has beenassociated with Al resistance in higher plants (Kochian,1995; Delhaize and Ryan, 1995; Ma, 2000; Matsumoto,2000). Aluminium enhances the exudation of malate inwheat (Delhaize et al., 1993), oxalate in buckwheat (Zhanget al., 1998), and citrate in corn (Pellet et al., 1995),snapbean (Miyasaka et al., 1991), and soybean (Silva et al.,2001). Organic acid release into the rhizosphere appears tobe a functional Al resistance mechanism that chelates Alextracellularly, reducing Al uptake and subsequent im-pacts on root physiological processes. In the present study,the substantially lower accumulation of Al in mycorrhizalbroomsedge plants (Table 1; Fig. 5) similarly suggests achange in Al speciation and availability in the rhizosphereas a consequence of AM fungal colonization, although themechanism of such change remains to be elucidated.
In a previous study, inoculation of broomsedge plants bythis same AM fungal consortium increased phosphorus useef®ciency and improved nutrient homeostasis in plantsunder Pi limitation (Ning and Cumming, 2001). Since thedisruption of Pi availability and acquisition by Al may beone toxic mechanism in plants (Clarkson, 1966; Foy, 1983;Cumming et al., 1986; Roy et al., 1988; Tan and Keltjens,1990; Macklon and Sim, 1992; Lux and Cumming, 2001),P physiology was investigated in the present work.Exposure to Al did not consistently affect shoot or root Pconcentrations in either mycorrhizal or non-mycorrhizalbroomsedge plants at concentrations <400 mM Al,although tissue P concentrations dropped dramatically innon-mycorrhizal plants exposed to 1000 mM Al (Fig. 3;Table 3). These patterns suggest that Al±Pi interactions didnot in¯uence broomsedge growth under Al exposure.Indeed, the relationships between plant mass and tissue Pconcentrations indicate that P became concentrated inplant tissues as Al inhibited plant growth (Figs 7, 8). Othermeasures of P physiology similarly re¯ected the signi®cantin¯uence of AM fungi on P acquisition, with little impactof Al on these processes. Patterns of APase activity
(Fig. 3C) indicate that mycorrhizal plants were under less Pstress than non-mycorrhizal plants, and this may be theresult of the higher PUE of mycorrhizal plants (Fig. 3B).These bene®ts of AM fungi on P relations have been welldocumented (Hayman, 1983; Bolan, 1991; Smith andRead, 1997) and support earlier work highlightingincreased P use ef®ciency in mycorrhizal broomsedgeunder P limitation (Ning and Cumming, 2001). However,these patterns do not support the original hypothesis thatinteractions between Al and Pi would determine thebroomsedge plant response to Al.
Numerous experiments have demonstrated that Alinterferes with the acquisition of cations in plants(Rengel and Robinson, 1989; Rengel, 1990; Huang et al.,1992, 1996; Nichol et al., 1993; Tan et al., 1993; Wheelerand Dodd, 1995; Lindberg and Strid, 1997; Lux andCumming, 1999, 2001). In the present study, Al depressedconcentrations of Ca and Mg in both shoots and roots inboth mycorrhizal and non-mycorrhizal plants (Table 4).However, mycorrhizal and non-mycorrhizal plants di-verged in their mass-tissue nutrient concentration relation-ships for these elements (Figs 7, 8; Table 4). The growth ofmycorrhizal plants was less affected by Al-inducedchanges in shoot or root Ca or Mg concentrations,suggesting that the colonization of broomsedge plants byAM fungi alters fundamental nutrient homeostasis.Mycorrhizal plants exhibited more stable tissue nutrient±biomass accretion relationships than non-mycorrhizalplants and this stability may increase Al resistance andallow successful colonization of stressful soil environ-ments. In a previous study, a similar phenomenon inbroomsedge plants grown under Pi limitation was noted(Ning and Cumming, 2001) and it was proposed thatenhanced micronutrient acquisition by mycorrhizal plantsaltered the nutrient±mass relationships. In the presentstudy, mycorrhizal and non-mycorrhizal plants differed,but concentrations in mycorrhizal plants were typicallylower than concentrations in non-mycorrhizal plants(Table 4), which may re¯ect a growth dilution inmycorrhizal plants. Thus, alternative in¯uences of AMfungi on host physiology, such as altered ¯ux of carbon tothe rhizosphere, altered water relations (Allen et al., 1981;Auge et al., 1995; Miller et al., 1997), or phytohormonechanges (Allen et al., 1980, 1982), may be involved in thegreater growth of mycorrhizal plants at any given tissuenutrient concentration. This capacity would clearly repre-sent an important adaptation for growth on acidic, nutrient-limited edaphic environments.
In conclusion, broomsedge, a species found colonizingextremely acidic soil environments in the eastern UnitedStates, appears not to be inherently resistant to Al in therhizosphere. However, an acidic ecotype AM fungalconsortium conferred Al resistance to broomsedge plants,promoting substantial growth at Al concentrations as highas 1000 mM. The exclusion of Al from tissues, a change in
Mycorrhizae and Al resistance in Andropogon 1457
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021

P homeostasis, a reduction in the impacts of Al on cationacquisition, and higher biomass accretion±tissue nutrientrelationships in mycorrhizal plants exposed to Al may beimportant mechanisms involved in Al resistance. Thus, thesymbiosis between AM fungi and broomsedge plants playscritical roles in Al resistance of this plant species, whichallows broomsedge to colonize and grow in unfavourableacidic edaphic environments.
Acknowledgements
We thank Dr JB Morton and Bill Wheeler for advice with AM fungalpropagation. This work was supported by the National ScienceFoundation (IBN-9723393) and the Graduate School of WestVirginia University.
References
Allen MF, Moore Jr TS, Christensen M. 1980. Phytohormonechanges in Bouteloua gracilis infected by vesicular-arbuscularmycorrhizae (Glomus fasciculatus). I. Cytokinin increases in thehost plant. Canadian Journal of Botany 58, 371±374.
Allen MF, Moore Jr TS, Christensen M. 1982. Phytohormonechanges in Bouteloua gracilis infected by vesicular-arbuscularmycorrhizae. II. Altered levels of gibberellin-like substances andabscisic acid in the host plant. Canadian Journal of Botany 60,468±471.
Allen MF, Smith WK, Moore TS, Christensen M. 1981.Comparative water relations and photosynthesis of mycorrhizaland non-mycorrhizal Bouteloua gracilis. New Phytologist 88,683±693.
Andersson M. 1988. Toxicity and tolerance of aluminium invascular plants: a literature review. Water, Air, and Soil Pollution39, 439±462.
Anonymous. 1985. Aluminium, eriochrome cyanine R method. In:Standard methods for the analysis of water and wastewater.Washington, DC: American Public Health Association, 183±187.
Auge RM, Stodola AJW, Ebel RC, Duan X. 1995. Leafelongation and water relations of mycorrhizal sorghum inresponse to partial soil drying: two Glomus species at varyingphosphorus fertilization. Journal of Experimental Botany 46,297±307.
Boan JB, Smith SE, Alston AM. 1993. Mycorrhizal responses ofbarley cultivars differing in P ef®ciency. Plant and Soil 157, 97±105.
Bolan NS. 1991. A critical review on the role of mycorrhizal fungion the uptake of phosphorus by plants. Plant and Soil 134,189±207.
Calba H, Jaillard B. 1997. Effect of aluminium on ion uptake andH+ release by maize. New Phytologist 137, 607±616.
Campbell CS. 1983. Systematics of the Andropogon virginicuscomplex (Gramineae). Journal of the Arnold Arboretum 64,171±254.
Chapman RH, Jones Jr SB. 1975. Ecotypic differentiation inAndropogon virginicus (Gramineae). Bulletin of the TorreyBotanical Club 102, 166±171.
Clark RB. 1997. Arbuscular mycorrhizal adaptation, sporegermination, root colonization, and host plant growth andmineral acquisition at low pH. Plant and Soil 192, 15±22.
Clarkson DT. 1966. Effect of aluminium on the uptake andmetabolism of phosphorus by barley seedlings. Plant Physiology41, 165±172.
Cumming JR, Eckert RT, Evans LS. 1986. Effect of aluminium
on 32P uptake and translocation in red spruce seedlings. CanadianJournal of Forest Research 16, 864±867.
Danielson RM. 1985. Mycorrhizae and reclamation of stressedterrestrial environments. In: Tate III RL, Klein DA, eds. Soilreclamation processes: microbiological analysis andapplications. New York, NY: Marcel Dekker, 173±201.
Delhaize E, Ryan PR, Randall PJ. 1993. Aluminium tolerance inwheat (Triticum aestivum L.). II. Aluminium-stimulated excretionof malic acid from root apices. Plant Physiology 103, 695±702.
Delhaize E, Ryan PR. 1995. Aluminium toxicity and tolerance inplants. Plant Physiology 107, 315±321.
de Miranda LN, Rowell DL. 1989. Aluminium±phosphateinteractions in wheat. New Phytologist 113, 7±12.
Fernandez-Marcos ML, Alvarez E, Monterroso C. 1998.Aluminium and iron estimated by Mehlich-3 extractant in minesoils in Galicia, northwest Spain. Communications in Soil Scienceand Plant Analysis 29, 599±612.
Foy CD. 1983. The physiology of plant adaptation to mineral stress.Iowa State Journal of Research 57, 355±391.
Fries LL, Pacovsky RS, Sa®r GR, Kaminski J. 1998. Phosphoruseffect on phosphatase activity in endomycorrhizal maize.Physiologia Plantarum 103, 162±171.
Gibson DJ, Risser PS. 1982. Evidence for the absence of ecotypicdevelopment in Andropogon virginicus (L.) on metalliferousmine wastes. New Phytologist 92, 589±599.
Giovannetti M, Mosse B. 1980. An evaluation of techniques formeasuring vesicular arbuscular mycorrhizal infection in roots.New Phytologist 84, 489±500.
Haug A. 1984. Molecular aspects of aluminium toxicity. CRCCritical Reviews in Plant Sciences 1, 345±373.
Haug A, Caldwell CR. 1985. Aluminium toxicity in plants: the roleof the root plasma membrane and calmodulin. In: John JB, BerlinE, Jackson PC, eds. Frontiers of membrane research inagriculture. Totowa, NJ: Rowman & Allanheld, 359±381.
Hayman DS. 1983. The physiology of VA endomycorrhizalsymbiosis. Canadian Journal of Botany 61, 944±963.
Huang JW, Pellet DM, Papernik LA, Kochian LV. 1996.Aluminium interactions with voltage-dependent calcium transportin plasma membrane vesicles isolated from roots of aluminium-sensitive and -resistant wheat cultivars. Plant Physiology 110,561±569.
Huang JW, Shaff JE, Grunes DL, Kochian LV. 1992.Aluminium effects on calcium ¯uxes at the root apex ofaluminium-tolerant and -sensitive wheat cultivars. PlantPhysiology 98, 230±237.
Keltjens WG, Tan K. 1993. Interactions between aluminium,magnesium and calcium with different monocotyledonous anddicotyledonous plant species. Plant and Soil 155/156, 485±488.
Kochian LV. 1995. Cellular mechanisms of aluminium toxicity andresistance in plants. Annual Review of Plant Physiology and PlantMolecular Biology 46, 237±260.
Koslowsky SD, Boerner REJ. 1989. Interactive effects ofaluminium, phosphorus and mycorrhizae on growth and nutrientuptake of Panicum virgatum L. (Poaceae). EnvironmentalPollution 6, 107±125.
Kothari SK, Marschner H, Romheld V. 1991. Contribution of theVA mycorrhizal hyphae in the acquisition of phosphorus and zincby maize grown in a calcareous soil. Plant and Soil 131, 177±185.
Lindberg S, Strid H. 1997. Aluminium induces rapid changes incytosolic pH and free calcium and potassium concentrations inroot protoplasts of wheat. Physiologia Plantarum 99, 405±414.
Lux HB, Cumming JR. 1999. Effect of aluminium on the growthand nutrition of tulip-poplar seedlings. Canadian Journal ofForest Research 29, 2003±2007.
Lux HB, Cumming JR. 2001. Mycorrhizae confer aluminium
1458 Cumming and Ning
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021

resistance to tulip-poplar seedlings. Canadian Journal of ForestResearch 31, 694±702.
Ma JF. 2000. Role of organic acids in detoxi®cation of aluminiumin higher plants. Plant and Cell Physiology 41, 383±390.
Ma JF, Hiradate S, Matsumoto H. 1998. High aluminiumtolerance in buckwheat. II. Oxalic acid detoxi®es aluminiuminternally. Plant Physiology 117, 753±759.
Macklon AES, Sim A. 1992. Modifying effects of not-toxic levelsof aluminium on the uptake and transport of phosphate inryegrass. Journal of Experimental Botany 43, 915±923.
Macklon AES, Lumsdon DG, Sim A. 1994. Phosphate uptake andtransport in Agrostis capillaris L.: effects of non-toxic levels ofaluminium and the signi®cance of P and Al speciation. Journal ofExperimental Botany 45, 887±894.
Marschner H. 1995. Mineral nutrition of higher plants. San Diego,CA: Academic Press.
Matsumoto H. 2000. Cell biology of aluminium toxicity andtolerance in higher plants. International Review of Cytology 200,1±46.
Medeiros CAB, Clark RB, Ellis JR. 1994. Effects of excessaluminium on mineral uptake in mycorrhizal sorghum. Journal ofPlant Nutrition 17, 1399±1416.
Mendoza J, Borie F. 1998. Effect of Glomus etunicatuminoculation on aluminium, phosphorus, calcium, andmagnesium uptake of two barley genotypes with differentaluminium tolerance. Communications in Soil Science andPlant Analysis 29, 681±695.
Miller RM, Hetrick BAD, Wilson GWT. 1997. Mycorrhizal fungiaffect root stele tissue in grasses. Canadian Journal of Botany 75,1778±1784.
Miyasaka SC, Kochian LV, Shaff JE, Foy CD. 1989.Mechanisms of aluminium tolerance in wheat: an investigationof genotypic differences in rhizosphere pH, K+, and H+ transport,and root-cell membrane potentials. Plant Physiology 91, 1188±1196.
Miyasaka SC, Buta JG, Howell RK, Foy CD. 1991. Mechanismof aluminium tolerance in snapbeans. Root exudation of acidicacid. Plant Physiology 96, 737±743.
Nellessen JE, Ungar IA. 1993. Physiological comparisons of old-®eld and coal-mine-spoil populations of Andropogon virginicusL. (Broomsedge). American Midland Naturalist 130, 90±105.
Nichol BE, Oliveira LA, Glass ADM, Siddiqi MY. 1993. Theeffects of aluminium on the in¯ux of calcium, potassium,ammonium, nitrate, and phosphate in an aluminium-sensitivecultivar of barley (Hordeum vulgare). Plant Physiology 101,1263±1266.
Ning J, Cumming JR. 2001. Arbuscular mycorrhizal fungi alterphosphorus relations of broomsedge (Andropogon virginicus L.)plants. Journal of Experimental Botany 52, 1883±1891.
Olsen SR, Sommers LE. 1982. Phosphorus. In: Page AL, MillerRH, Keeney DR, eds. Methods of soil analysis. Part 2. Chemicaland microbiological properties. Madison, WI: American Societyof Agronomy and Soil Science Society of America, 403±430.
Pacovsky RS. 1986. Micronutrient uptake and distribution inmycorrhizal or phosphorus-fertilized soybeans. Plant and Soil 95,379±388.
Parker DR, Norvell WA, Chaney RL. 1995. GEOCHEM-PC: achemical speciation program for IBM and compatible computers.In: Leoppert RH, Schwab AP, Goldberg S, eds. Chemicalequilibrium and reaction models. Madison, WI: Soil ScienceSociety of America, 163±199.
Parkinson JA, Allen SE. 1975. A wet oxidation procedure suitable
for the determination of nitrogen and mineral nutrients inbiological material. Communications in Soil Science and PlantAnalysis 6, 1±11.
Pellet DM, Grunes LD, Kochian LV. 1995. Organic acidexudation as an aluminium tolerance mechanism in maize (Zeamays L.). Planta 196, 788±795.
Randall PJ, Vose PB. 1963. Effect of aluminium on uptake andtranslocation of phosphorus by perennial ryegrass. PlantPhysiology 38, 403±409.
Rengel Z. 1990. Competitive Al3+ inhibition of Mg2+ uptake byintact Lolium multi¯orum roots. Plant Physiology 93, 1261±1267.
Rengel Z, Pineros M, Tester M. 1995. Transmembrane calcium¯uxes during Al stress. Plant and Soil 171, 125±130.
Rengel Z, Robinson DL. 1989. Aluminium effects on growth andmacronutrient uptake by annual ryegrass. Agronomy Journal 81,208±215.
Ritchie GSP. 1995. Soluble aluminium in acidic soils: principlesand practicalities. Plant and Soil 171, 17±27.
Ross JP. 1971. Effect of phosphate fertilization on yield ofmycorrhizal and non-mycorrhizal soybean. Phytopathology 61,1400±1403.
Roy AK, Sharma A, Talukader G. 1988. Some aspects ofaluminium toxicity in plants. The Botanical Review 54, 145±178.
Ryan PR, Kochian LV. 1993. Interaction between aluminiumtoxicity and calcium uptake at the root apex in near-isogenic linesof wheat (Triticum aestivum L.) differing in aluminium tolerance.Plant Physiology 102, 975±982.
Silva IR, Smyth TJ, Raper CD, Carter TE, Rufty TW. 2001.Differential aluminium tolerance in soybean: an evaluation of therole of organic acids. Physiologia Plantarum 112, 200±210.
Smith SE, Read DJ. 1997. Mycorrhizal symbiosis, 2nd edn. SanDiego, CA: Academic Press.
Snoeyink VL, Jenkins D. 1980. Water chemistry. New York, NY:John Wiley & Sons.
Tabatabai MA, Bremner JM. 1969. Use of p-nitrophenylphosphate for assay of soil phosphatase activity. Soil Biologyand Biochemistry 1, 301±307.
Tan K, Keltjens WG. 1990. Interaction between aluminium andphosphorus in sorghum plants. II. Studies with the aluminiumtolerant sorghum genotype SC0283. Plant and Soil 124, 25±32.
Tan K, Keltjens WG, Findenegg GR. 1993. Evaluating thecontribution of magnesium de®ciency in the aluminium toxicitysyndrome in twelve sorghum genotypes. Plant and Soil 149, 255±261.
Taylor GJ. 1988. The physiology of aluminium phytotoxicity. In:Sigel H, Sigel A, eds. Metal ions in biological systems. NewYork, NY: Marcel Dekker, 123±163.
Wheeler DM, Dodd MB. 1995. Effect of aluminium on yield andplant chemical concentrations of some temperate legumes. Plantand Soil 173, 133±145.
Yang ZM, Sivaguru M, Horst WJ, Matsumoto H. 2000.Aluminium tolerance is achieved by exudation of citric acidfrom roots of soybean (Glycine max). Physiologia Plantarum110, 72±77.
Zeleznik JD, Skousen JG. 1996. Survival of three tree species onold reclaimed surface mines in Ohio. Journal of EnvironmentalQuality 25, 1429±1435.
Zhang FS, Ma J, Matsumoto H. 1998. High aluminium resistancein buckwheat. I. Al-induced speci®c secretion of oxalic acid fromroot tips. Plant Physiology 117, 745±751.
Mycorrhizae and Al resistance in Andropogon 1459
Dow
nloaded from https://academ
ic.oup.com/jxb/article/54/386/1447/568667 by guest on 02 D
ecember 2021





























