Embed Size (px)
Citation preview

ANDROGENIC DOUBLED HAPLOIDS IN INDICA RICE : IN VITRO INDUCTION, PLANT REGENERATION
AND EVALUATION
A Thesis Submitted for the Degree of
DOCTOR OF PHILOSOPHY
By
Rabindra Nath Bhuyan
NORTH·EASTERN HILL UNIVERSITY
SHILLONG
1996

.L..u .... !.KAlt..t/CJ 31- :J_~v/ Ac: ~o .••...........•• .-.......
~ Ac•: Br... ···fl._q~g::..·-;;-D• ......... ... /~2._1.;. CL .. ~ by . . . .. ~~e'f.-. .. Sub.Hee1 y .•• ~ ........ -Ent( .................... -YT-an.q -. · ~ ~,v ........... .._...
b <::'-::,
~8 Lt I q '?:,, ()J-t-Q~~ I 6 I ~ l,) I

We certi~y that the thesis entitled II Androgenic
doubled haploids in rice : in vitro induction~ plant regeneration
and evaluation" submitted by Mr. Rabindra Nath Bhuyan ~or the
degree o~ Doctor o~ Philosophy in Botany o~ the North Eastet·n
Hill University, Shillong embodies the t·ecm·d 0~ original
investigation carried out by him under our supervision. He has been
duly registered and the thesis presented is worthy ~or being
considered ~or the award o~ Ph.D. degree. The work has not been
submitted ~or any degree o~ any other university.
(Prcunod Tandon)
Department o~ Botany
North-Eastern Hill University
Shillong 793 022
Date: June 5, 1996
Place: Shillong
, ' J \,1 \".{ 0 5 I \ q q -b (H.S.Gupta)
Division o~ Plant Breeding
ICAR Research Complex ~or
North Eastern Hill Region
Barapani - 793103

ACKNOWLEDGEMENT
I express my deep sense o-f gratitude to Dr. H. S. Gupta,
Principal Scientist and Head, Division o-f Plant Breeding, ICAR
Research Complex -for North Eastern Hi 11 Region, Barapani and
Dr. PrQtnod Tandon, Department o-f Botany,
North - Eastern Hill University~ Shillong -for their constant
guidance, valuable suggestion, const t-uct i ve criticism and
g~nerous attitude throughout the course o-f this investigation and
instrumentalization o-f this dissertation.
I am deeply thank-ful to Dr. B. K. Sarma, and
Mr. A. Pattanayak, o-f the Division o-f Plant Breeding ICAR
Research
ways.
Compex -for NEH Region -for helping me in all possible
I am highly indebted to the Di t-ectm- o-f ICAR Research
Complex -for North Eastern Hill Region ·Fm- pt-oviding -ficilities
during the course o-f this investigation.
I -further express thanks to Drs. D . K • Pandey,
M. Singh , U. K. Hazarika • S. Banerjee and Mr. A • Khound -for
their generous help in computer works.
I also sincerely thank Miss. B. Bhattacharjee, Mr. Ram Murti
and Mr. Hari Govind -for their valuable assistance in laboratory
works.

I am indebted to Miss. Verginia Kalita and Ms Suman Kumaria
o-f Department o-f Botany Nor-th-- E"'stet-n Hill University
~or their kind cooperation at the time o-f seminars.
Sincere thanks are also due to Mt- • A. Ahmed and
Mr. Nandalal Chetri -for helping the author in matters related to
-field trials.
I wish to thank Bishnu Sita Sharma and
Jotshna Bora -for their sel-fless service to the author in
routine works o-f the laboratory~
Sincere thanks are also due to the sta-f-f memebers o-f Centre
o-f In-formation Technology, New Kenche's Trace, Shillong -for
technical help in preparing the manuscript.
Last but not the least, I great-fully acknowledge the
constant inspiration and help received -from the members o-f my
-fami 1 y without which it would have been impossible to camp lete
the present investigation.
~~si~l=~k ( RABINDRA NATH BHUYAN )

Chapters
1.
2.
2.1.
2.2.
2.2.1.
2.2.2.
2.2.2.1.
2.2.2.2.
2.2.2.3.
2.2.3.
2.2.3.1.
2.2.3.2.
CONTENTS
Title
Introduction
Materials and methods
Materials
Methods
Time or collection or panicles
Preparation of staining solutions
and staining procedure.
Acetocarmine ~tain
Feulgen stain
Iron alum - haematoxylin stain.
Anther culture
Methodology
List or media and their composition
used for anther callusing
pp. No.
1 14
15 - 35
15
16
16
16
16
17
17
18
18
18
2.2.3.2.1. Callus prolire~ation and regeneration 22
2.2.3.2.2. Ploidy status 24
2.2.3.3 Production or promising double haploids 25,
2.2.3.3.1. Selection and hybridiza~ion 25
2.2.3.3.2. Culturing or a~thers from F1 plants,
regeneration a~d production or double
haploid lines 25

2.2.4.
2.2.4.1.
2.2.4.2.
2.2.4.3.
2.2.4.4.
2.2.4.5.
2.2.4.6.
2.2.4.7.
2.2.4.8.
3.
3.1.
3.1.1.
3 .. 1.1.1.
3.1.1.2.
Field evaluation
Data collectjon
Data analysis
Analysis o~ variance
Coe~~icient o~ variation
Her it ab i 1 it y
Genetic: advance
Correlation
Path coe~~ic~ent analysis
Results
Anther culture callus induction and
plant regeneration ~rom local c:old
tolerant cultivars
Standardization o~ conditions ~br
c:allusing'o~ mic:rospores
Time o~ collection or panicles
Environmental conditions at the time or
collection or panicles
36 -
27
27
28
28
30
31
31
32
33
60
36
37
37
37
3.1.1.3. Length or cold pretreatment 38
3.1.1.4. Staging o~ microspores 39
3.1.1.4.1. Staging with acetocarmine stain 39
3.1.1.4.2. Staging with ~eulgen stain 39
3.1.1.4.3. Staging with iron alum-haematoxylin stain 40

3.1.2.
3.1.3.
3.2.
3.2.1.
3.2.2.
3.2.3.
3.2.3.1.
3.2.3.2.
3.2.3.3.
3.3.
3.3.1.
3.2.
3.3.3.
3.3.4.
3.3.5.
E~~ect o~ di~~erent culture media on
callusing ~requency and callus
proli-Feration
Regeneration and con-Firmation o~ ploidy
status
Production o~ double haploids
Selection of= parents and hybridization
Raising o~ F 1 and their comparision
with parents
Culture o~ anthers derived ~rom F 1 plants
Callusing
Regeneration
Evaluation o~ double haploid ( DH )
lines
Field evaluation and per~ormance o~ double
haploid ( DH 2 ) lines
Means and ranges ~or di~~erent characters
Genetic variability
Analysis o~ variance
Simple correlation coe~~icience among some
major yield contributing characters.
Direct and indirect e~~ects o~ di~f=erent
characters on grain yield
40
45
51
~51
51
52
52
53
55
57
57
58
59
59
60

4.
4.1.
4.2.
4.3.
4.4.
4.:S.
5.
Discussion 61 - 79
Standardization o~ conditions ~or
callusing o~ microspores in local cultivar 62
Anther /microspore culture
Ploidy status o~ plants derived through
anther culture
Production o~ promising doubled haploid
lines and their evaluation
Field evaluation and per~ormance o~
doubled haploid ( DH ) lines
Summary
Bibiliography
80
84
64
69
70
74
83
99

INTRODUCTION
Rice, the major source o-f calories -for 407. o-f the
global population, is ranked as the world's most important -food
crop. Consequently, it is one o-f the most intensively
investigated crop plant. The accumulated knowledge-base has been
o-f considet-able value in enhancing t-ice productivity and, in
turn, in the betterment o-f millions o-f people who depend on it as
a source o-f -food and income.

As in the case o~ other crop plants~ rice breeders have
mainly pursued the exploitation o~ natural variation ~or
tailoring cultivars suitable ~or the demands o~ agriculture. The
process o~ hybridization and selection have been the main
vehicles o~ this delivery system. The achievements in this regard
have been impressive. Taichung Native! (TN 1) and IR series
starting ~rom IRS to IR74 lines/varieties released by the
International Rice Linstitute, Manila, the Philippines and L Institution o~ National Agriculture Research System are known ~or
their impact on enhancing productivity per unit area and time.
The increase in rice production has helped
alleviate widespread hunger- and -famine by keeping pace with
ever gt·owing human population. I:Jith the rice-consuming
population increasing at an alarming rate of= hundred
million per
would exceed
year, by 2020 A.D. the .. --~
760 million ~,s_~) The
rice requirement o-f Asia
situation in India, the
J.:n·gest rice gtowing countt-y, is also not stable. Although we
are sel~ su~~icient at pn~sent, ot.n- rice n,·qu i rement is
estimated to increase at a rate o~ 25 to 30 million tones every
decade. This means /~- "'
producing 100 million ~s
tonnes) by the year 2000 A.D.
we should achieve the target o-f
pr~r (present 1 y 75 mi 11 ion
This requirement would increase
2

to 160 million tonr1es by 2020 A.D. Accordingly, the production o~
rice has to be doubled in next 25 years. It has to be recognised
here that plant breeding
has severe biological
is a slow and painstaking process that d_e--q....~-Ad
1 imitations, and th<'\t such d[stimatea-,,
increase in yield and productivity r:annot be sustained
inde~initely. It is in this context that rapidly emerging
techniques o~ plant cell and molecular biology have attracted
much attention -for they pt-ovide a powet-~ul and novel means to
supplement and complement the traditional methods n~ plant
improvement.
Since 1930's, plant tissue culture has progressed mostly
on dicotyledonous plants, and studies on monocotyledonous plants
lagged behind. The in vitro culture o~ rice started with the
organ culture, when Fujiwara and Ojima (1955) cultured excised
roots. The culture conditions ~or vat-ious stages o~ immature
embryos were examined by Amemi ya et al. ( 1956) and obtained
seedlings ~t-om even -five day old cultLu-ed embryos. F1 hybrids
~rom otherwise unsuccess~ul rice crosses were· subsequently
obtained by Bourhamont (1961)~"-Iyer and Govila (1964). Cultut-e o~
nodes and young seedlings on Heller's medium containing vitamins
and auxins was reported by Furuhashi and Yatazawa in 1964. This
was -followed by plant t-egenet-ation ~rom seed derived callus
(Tamura,1968; Nishi et. al.~ 1968 and M~eda(~-;<-~.,1968). Plants "---··--"""·""' ! "·

were also regenerated -from root callus (Kawata and
Ishihara,1969)as well as pollen derived callus (Niizeki and
Oono,1968; Cho and Zapata, 1988).
The demonstt-at ion ~ Guha and 1'1aheshwar i ( 1964, 1966,
1967)/ that immature anthet-s of D,:;tura .i. n.nox.i a when cui tured
in an appropriate nutrient medium can give rise to plantlets with
a haploid numbet- o-f chromosomesy opened up a new approach to the
induction o-f haploidy in plants genet-ally known as androgenesis.
Soon it was -found that this technique can be e-f-fectively employed
in Orvza (Niizeki and Oono, 1968; Guha et. a1,.1970), .Nicotianet
(Bourgin and Nitsch, 1967; Nitsch et. al.,1969; ~kata and Tanaka
,1968; Nitsch and Nitsch,1969; Sunderland and Wricks, 1969;
Helchers and Labib, 11J70), B:·-A":;t';_ic,::~ O<ameya and Hinata, 1970) and
1-fordeum and Lol i um (Clapham, 1971). In Or·l;za sat i \-'a, varietal
g were-reported by Guha etj al.(1970).
The underlying principle o-f androgenesis is to direct the
...v=--~ . ./ development o-f the microspores whose -fate is~ to become~
gamete~ and to -force its development dit-ectly into a plant or
callus. This abnormal pathway is possible i-f the pollen cell is
provided with certain speci-fic conditions.
Utilization o-f anther culture technique in the breeding
programmes has been in practice, mostly in China, -for more than
4

20 years. Since the release for cultivation of Hua-Yu No. 1 and
Hua-Yu No. 2, the rice vat- iet i es developed through anther
culture breeding in 1976, Chinese t-esearchers ~e: • rwpur.t;ea M3
have developed over 100 varieties and lines with objectives like
earliness, yield, resistance to disease and grain quality
(Shen et al.~ 1983). Reseat-chet-s in China, have also successfully
used anther culture for several other purposes like, solving
sterility problems in indica-japonica hybrids. Outside China,
utilization of anther culture in rice breeding programme has been
comparatively of a lesser magnitude. In Korea, the technique has
been mainly used for developing blast resistan~ and cold
tolerant varieties (Gun~ 198~). In laiwan, application of anther
culture was mainly to produce inter-subspecific and inter
specific Fl hybt-ids (Woo and Chen,. 1982). International Rice
Research Institute, Manila's programme of anther culture has been
focussed qn production of varieties fOt- cold tolerance using Fl
hybrids of japonica-indica and indica-japonica cresses.
Researchet-s in Nepal (Agrawal et al., 1994) have also t-egenerated
plants from anthers of cold tolerant rice. The biggest programme
outside China is at CIAT (F'ulvet- and Jennings, 1986) as for
example, 20,000 diploid R2 lines are reported to have been raised
from approximately 100 triple crosses for obtaining Rl lines for
blast resistance, Fe-tolerance and white belly determination.
5

Factor-s a-f-fecting andr-ogenesis in rice have been
r-eviewed by Chen et al. ( 1991). Cold m- heat shock -for shor-t
period is known to enhance the yield o~ pollen embr-yos and gr-een
plants r-emad~ably. This bene~icial e~-fect o-f cold pr-etr-eatment
was -first repor-ted by Nitsch and Norreel ( 1973) in Datura
innoxia. It has also been shown to increase the r-atio o-f
responding anther-s in many species, such as N."ic:otiana tabac:um
(Nitsch, 1974), Datur.:.; :innoxic:t., llvo.c::.c.,·i.imus n.i.gc::-r. fkn-deum "'Lilgare
(Sunder-land and Roberts, 1977) and i}rvzc.· sBt:i-.,'a (Chale-f-f et.
al., 1975). In t-he_~ t·ir.e anthet- culture, this
tr-eatment has been an essential step and broadly adopted by many
r-esQat-cher-s. Callus induct ion -ft-equency 0~ Keng ( a~-vza StElt .i ,,,a
Subsp. japonica) or Hsien ((Jryz,::i .c::. •. :~tiva Subsp. indica) rice can
be r-aised -from two to more than tm1 times (Ying, 1986) via an
appropr-iate cold tr-eatment.
The time needed -fot- pt-etreatment depends on the
temper-atur-e used. In general, lower the temper-atur-e (as low
as 3°- 5°C) shorter- the time needed,
( as high as 10°- 15°C longer
and
the
higher the temper-atur-e
period o-f treatment
r-equir-ed. Through the experiment by Wang and his associates
( 1974) in indica rice, 10 day pretreatment at 3° - 5°C, 10 to
15 day at 6° - soc, or 15 to 20 day at 9° - 10°C was optimum -for
6

the enhancement o~ callus induction ~requency. ~hen the period o~
cold pretreatment is too long, the ~requency o~
di~~erentiated ~rom callus decreased dramatically
green plantlet (U-1'
while Lo~ albinoJ
increased greatly. The appropriate cold pretreatment bene~its the
di~~erentiation o~ green plantlets and raises the di~~erentiation
~requency and ~ormation o~ green 0
(!ti} 1983; Go and Chen , 1 983a) •
7?5f'
r ·~
plantlets (Zhou and(Chang? 1981; '-.. ---· ~~--- .. ~ ?
Physiological investigation has revealed that cold
pretreatment could reduce the respiration intensity o~ anther and
the consumption o~ materials, and could prolong the li~espan o~
anther. In addition, it was ~ound that the weight o~ anther also
increased during cold prett·eatment. All the!Se e~-fects would be
advantageous to the initiation and development o~ pollen grains
inoculated (~, 1983). Qu and Chen, ( 1983 1;:1) demonstrated that 'S the cold pretreatment supported the sut-vi val o~ most o~ the
grains and completion o-f the induction process -for androgenesis
at the same time, and thet·e-fore lef'd to a mLICh higher induction "<..
~requency o-f callus than that o-f the control.
Other treatments such as heat shock (Kellet- and
Armstrong 1978, 1979), placing the detached tillers in watet· at
room temperature (Wilson et al. 1978 ) and putting the excised
anthers in a watet· satLu·ated atmosphet·e (DLmwell, 1981 ) have
7

also been ~ound bene~icial to production o~ embryos and plants.
Formation o~ embryo or callus depends greatly on the
appropriate nutrients in the culture medium (Nitsch, 1969; Wang
et al., 1974). Clapham ( 1973) discovered that high concentration
o~ ammonium ions in LS medium (Linsmaier and Skoog, 1965) was
disadvantageous ~or callus ~ormation in barley microspores.
Subsequently, Chu et al.(1975) developed N6 medium which is
characterised by having a low concentration o~ (NH4 >2so4 and high
KN03 • This medium proved to be very e~~icient ~or rice anther
culture (Chu, 1978; Genovesi and Maqill, 1979; Chen et al. 1982;
Tsay et al., 1982). Specially i~1
·'iraminae, a high sucrose
concentration has been ~ound to be bene~icial -fot- plant
production through anther culture (Clapham,1973; Ouyang et al.,
1973; Dono and Lat-tet-, 1976; Miao et al., 1978). Chen (1978)
reported that in t- ic:e 6% sucrose in callusing medium and 3'Y.
sucrose in regenerating medium resulted in highest -frequency o-f
callus and green plant regeneration @~pability in rice. But some
investigators have n~pm-ted that 6% sucrose is too high and 5%
was recommended (Huang et al., 1978. Chu,1978; Chale~~ and 1-
Stolarz, 1981). It has been observed that high concentration o-f
sucrose is responsible -for maintaining osmotic pt-essure o-f the
nutrient medium (Wang et. al., 1974; Ch;de~-f and Stolarz,1981).
Plant growth regulatot-s like au:dns and cyf:okinins play a crucial
8

role in the process o~ androgenesis. In earlier days, 2,4-
dichlordphenoxyacetic ac:in (2.4--D) ,.,as thought to be the only ._.,.
hormone essential ~or androgenesis and it was always added to the
culture medium with or without cytokinin (usually kinetin)
and/or other auxins (Niizeki and Oono, 1968; Harn, 1969; Nishi
and t'1itsuoka, 1969; Guha- t1ukhfr- jee, 1973; Wang et al., 1974).
The inhibitory e~~ect o~ 2, 1l-D, and that it can be
replaced by <X - naphthylacetic acid (NAA), was ~it-st reported by
Niizeki and Oono,(1971). A~ter~anJs it was also -found out that
~or obtaining callus o~ high morphogenic potential cytokinin is
also needed (Chen, 1977; Chalef~ and Stolarz, 1981; Lee and Chen,
1982). However, this conclusion o~ hormonal requirement may not
be applicable to all cultivars as genotypic di-f~erences in
hormone requirement has been reported (Liang, 1978; Cornejo
-Martin and Primo- Millo, 1981).
Light and temper-atw-e play an impm-tant role in
androgenesis (Maheshwari et al., 1980). The light conditions
varied ~t-om complete dar-kness (Niizeki and Dono, 1968; Hat-n,
1969; Woo and Tung, 1972; Guha -- t-1ukhfr jee, 1973) to continuous
illumination (Nishi and Mitsuoka, 1969; Niizeki and Dono, 1971).
Darkness is ~avourable ~or initiation o~ callus (Cornejo - Martin
and Primo- Millo,1981). For plant regeneration high intensity o~
light is essential.
9

Other -factors that influence anther response are
composition o-f the gas mixture in the culture vessel (Dunwell,
1979; Johansson et al.,1982) and the density o-f anther inoculated
in the culture medium (Bajaj et al., 1977; Sunderland et al.,
1981).
The genotype o-f the donm- plantj has a strong e-f-fect on
pollen plant -formation. There at-e several reports on genotypic
variation in anther response o-f rice (Guha et al., 1970; Iyer and
Raina, 1972; Wang et al-~1974; Raina and Iyer,1974; Tsai and Lin,
1977; Chen and Lin,1976; Chu, 1982; Rush and Shao 1982; Ding
et al 1983; Zapata, 1985; Hu, 1985; Miah et al.,1985; Karim et
al., 1985; Boyajiev and Kuong, 1986; Guidet-doni et al., 1986;
Mikami and Kinoshita,1988). In the initial study o-f lyer and
Raina (1972), out o-f 15 cultivars only 5 showed callusing and o-f
these only one (IARI 5788) t-egenerated roots and shoots.
Genotypic variation is now seen mostly in the extent o-f response,
rather than in the existence o-f some responsive genotypes and
some unresponsive , genotypes (Raina, 1989). In general,
japonica rice varieties are known to respond better than indicas
(Chen and Lin, 1976; Hu, 1978; ~:w::h and Shao, 1982; Chu 19B2;Woof
and Chen,1982; Gun, 1982; Zapata 1985). Hu (1984. 1985) t-eported
10

that the induction ~requency o~ green plants was more than lOX in
japonica cultivars, whereas it was only 1% in indica rice.
The reason that this technique oF androgenesis has
attracted such wide attention is the possibility o~ obtaining
haploid cells, tissues and complete ~-~anism~, which has a single
dose o~ gametic in~ormation. Instant homozygous lines could then
be obtained by doubling the chromosome complement in these
haploids without having to resort to a lengthy and time consuming
programme o~ inbreeding, which is indispensable ~or obtaining
homozygosity by conventional methods. In addition, recessive
mutants can be readily discovered because o~ presence o~ only one
set o~ chromosomes, and hence only one copy o~ each gene.
The other attt·act ion o~ anthet· cui ture is the
possibility o~ early ~ixation o~ recombinants. Pollen plant
population derived ~rom anther culture o~ Fl hybrids will express
segregation patterns otherwise expressed in the F2 or F3
generation. Diploidization oF haploids will render them
homozygous, resulting in the t·apid establishment o~ pure lines
that otherwise would require several generations o~ sel~ing. The
saving o~ time and resources would be ~ar greater in long
duration compan~d to dut-at ion cultivars, in which
homozygosity can be achieved rathet- speedily through conventional
11

methods such as single seed descent, by growing two or three
crops per year. In rice, there~ore, anther culture breeding would
be more advantageous in situations involving temperate
environment, photosensitivity ot- environmental stresses because
in these cases only one generation can be raised in a year.
Increased selection e~~iciency is the other advantage
o~ anther culture breeding, especially when dominance variation
is signi~icant. In conventional breeding, early generation lines
(F2 - F4) show phenotypic di~rerences to which both additive and
dominance e~~ects contribute. In contrast, double haploid lines
derived ~rom F1 anther culture will show only additive variance
and, there~ore, high heritability due to elimination o~ dominance
e~~ects. Thus, compared to the F2 population, ~ewer double
haploid plants will be required -for the purposes or selection or
the desired recombinant(s). To elabm-ate this point, Baenziger
and Schi\e~-fer-,(1983) cited the hypothetical example o-f a cross
segregating ~or three recessive genes and ror three dominant
genes. In the case o-f three recessive genes, 1/8 o-f the double
haploids derived -from such a cross would be selected as compared
to 1/64 or the F2. In the case of three dominant genes, wherea~
1/8 o~ the double haploids, ~~~ of which would breed true, 27/64
o-f F2 plants would be selected, on the basis or phenotype, but
only 1/64 would be true bn.:>edino. Chinese scientists have
12

estimated that about 150 pollen plants det-ived -from Fl anthers
would be enouqh instead o~ 4 - 5.000 F2 plants. ~or the purpose
o-f selection o~ desirable genotypes (Shen et. al., 1983). Double
haploid breeding would be particulat-ly use-ful in situation in
which extensive recombination is not impm-tant (Chale-f-f, 1979;
Chu, 1982). In rice, e-f~orts to combine the desirable traits o~
indica and japonica cultivars have o-ften been ~rustrated by the
high degree o~ sterility o-f the hybrids and vigorous segregation
toward the parental types in F2 and subsequent generations.
Double haploids produced through anther culture o-f the Fl hybrid,
or o-f a promising segt-egant, ~·muld ensure stabi 1 ization and
thereby produce and arrest segregation in successive generations.
Use o-f anther culture ~or solving the pt-oblems arising -from
inter-subspeci~ic and intet--spec i ~ic hybt- idi zat ion can be
regarded as the most important area o-f its application in rice
improvement.
Another area o~ use o-f double haploid is the selection
o-f the races o-f pathogen o-f insect biotypes (Rush and Shao,
1982). Use o~ 'pure line' varieties or breeding lines may cause
considerable experimental error due to residual heterozygosity o-f
the genotypes investigated. Anther culture could be used to
obtain homozygous lines Fm- utilization in such situations, as
well as in sct-eeninq -rm- t-esistr.1nce to herbicides, physiological
13

studies, and in puri~ication o~ A~ B and R lines ~or hybrid rice
progt·ammes.
The present investigation is aimed at standardization
o~ conditions ~ot- anthet- cul tw-e-det· i ved callus induct ion and
plant regenet·ation -from cold tolet-ant t-ice vat·ieties qrown in
high altitude areas o-f Nm-th Easten1 Hill of= India. All these
varieties are red kerneled, possess cold tolerance and they are
poor yielders. Having standar-dized the conditions -for
androgenesis, we produced sevet-a I double haploid 1 ines tht-ouqh
anther culture o-f Fl derived hybrids between IR 70, an elite IRRI
bt·eeding line and Khonm-ullo. a local cold tolerant line. The
double~ haploid 1 ines wet-e evaluated .fot· agronomic charactet·s
undet· medium altitude ( 950 m. msl Q.
~~2_~-~!:nd condition .:md
promising ones with high yield and \l'lhite ket·nel colour wet·e
identi-fied -for -further evaluation.
14

MATERIALS AND METHODS
2.1 MATERIALS
The -following locally adapted varieties of= rice (Oryza
satival L). were used initially to assess the response to callus
induction during anther culture:
1. Khonorullo
2. RCPL 1 - 1C (Meqha Rice 2)
-~. RCPL 1 - 2C
4. RCPL 1 - 3C
I:' ..Jo DR 92 and
6. Pus a 33
15

They were qrown in the experimental ~ields o~ Plant
Breeding Division, Indian Council o~ Agricultural Research l~or
North Eastern Hills Region, Barapani~ Meghalaya, with normal
~ertilizer and crop management.
In addition, IR 65, IR 68, IR 70~ Khonorullo, Yamuk, Pusa
33 and DR 92 (Subhadra) were grown in pots, with normal
~ertilizers ~or crossing. One cross i.e., IR70 X Khonorullo was
made and attempted ~or daub le hap laid pt·oduct ion.
2.2 METHODS
2.2.1 Tim• of collection of panicle•
Panicles were collected while still inside the sheath,
mostly in the mornings (between 8.30 AM - 10.30 AM) between
middle or August to the end o~ September i.e, in the rlowerinq
period. They were wrapped in polythene bags with a little water
and stored at 8 - 10°C ~or 1 to 16 days.
2.2.2 Preparation o~ staining solutions and staining procedure
2.2.2.1 Acetocarmine stain:
2 gm o~ carmine was dissolved in 100 ml of 45X acetic
acid and was kept ovet·night. Ne:::t day the stain was ~iltered and
used ~or staging o~ microspores. A ~ew anthers were squashed in a
drop o~ stain and observed under a research microscope.
16

2.2.2.2 Feulgen stain•
Feulgen reagent was prepared by adding 1 gm basic
fuschin, 2 gm potassium metabisulphite, 5 ml 1N HCl, 85 ml
water and 1 gm activated charcoal (Nitsch 1981). The solution was ,--~.
allowed to stand overnight in fudg~. , ___ /
The anthers were dissected out from the spikelets and
fixed in acetic alcohol (1 : 3) for a period ranqinq between
overnight to 24 hour. They were brought to water level through
alcohol grades and hydrolyzed with 1 N HCl at 60°C for 5 to 7
minutes. After the hydrolysis the vials were immediately put
into ice cold water to stop further action of HCl. After repeated
washing with water the anthers were squashed in a drop of the
staining solution and observed under the microscope after 5
minutes.
2.2.2.3 Iran Alum Haemataxylin staina
A stock solution of 4/. haematoxylin in 45/. acetic acid
was prepared and 1 gm of ir-on alum was added to it. The solution
was allowed to stand -for 24 houri.:_ To 10 ml of this stock ~ solution, 4 qm of chloral hydrate was added and after complete
dissolution)it was used (Chang et al.~ 1978)
Anthers were squashed in a drop of the staining
solution under a cover glass. After 2 to 5 minutes, the slides
17

were observed under a research microscope.
2.2.3 ANTHER CULTURE
2.2.3.1 Methodology
sterilized.
Panicles o~ the varieties mentioned above were sur~ace
Since individual panicles contain tfiicrospo~~~~---,at "-··~ . -.- . ..- - ~-
A stages o~ development at di~~et-ent places, the sheath di~-ferent
was opened at -five place~and ~~loret was taken out -from
opening. Anthers o-f these ~lorets were examined under
each
the
microscope
containing
-for ascertaining the nuclear stage.
microspores at mid-uninucleate stage were
Lfortions
taken out
and anthers were removed -from it. The anthers were then passed
through 70/. ethanol -for a -few seconds, and then "-sterilized in
O.l'l. mercuric chloride -for 7 to 8 minutes and rinsed 3 times 0
with sterile distilled "Jatet-. They wet-e then inoculated jn the
-following semisolid/liquid medium.
2.2.3.2 List OT media and their composition used TOr anther callusing
Medium No. 1 65 (Semisolid)
Medium No. 2 E24 (Semisolid)
Medium No. 3 Minimal (Semisolid)
Medium No. 4 LB (Semisolid)
18

Medium No. 5
Medium No. 6
Medium No. 7
Medium No. 8
MP2 (Semisolid)
E10G (Liquid)
Modi~ied Nb (Liquid)
E10 YK (Liquid)
Medium No. 1 G5 (Samisolid)a
Blaydes medium/.._ NaH2P04 (150 mg/1), (NH4 >2 so4 (134
mg/1), KN03 (2500 mq/1), MqS04 • 7H2 0 (250 mg/1), CaC1 2 • 2H2 0 (150
mg/1), KI (0.75 mg/1), Na2 Mo04 (0.25 mg/1), CuS04 .5H2o (0.025
mg/1), CoC1 2 (0.025 mg/1), [ Na2EDTA (745 mg), FeS04 .7H2 0 (557
mg) dissolved in 100 ml o~ water and 5 ml o~ this solution is
used ~or 1 litre], Sucrose (50 gm/1), Agar (8 gm/1), ~orti~ied
with 2,4-D (2 mg/1), Nicotinic acid (0.5 mg/1), Pyridoxin HCl
(0.5 mg/1), Thiamine HC1 (1.0 mg/1), Glycine (2.0 mg/1) and
Inositol (100 mg/1).
Medium No. 2 E24 (semisolid)•
G5 basic salts~orti~ied with 2,4-D (1 mg/1), BAP (0.5
mq/1), IAA (0.5 mg/1) and Inositol ( 160 mg/1).
Medium No. 3 Minimal (semisolid);
Med i urn No. 12 o+ Snnq ct . a I ( 1978) :
19

mg/lJ, KN03 ( 100 mg/1) !' KH2Po4 (300 mq/1), MqS04 • 7H2D (200 mg/1),
CaC1 2 • 2H20 (110 mg/1), MnS04 .4H2D (10 mq/1), [Na2EDTA (745 mg)~
FeS04 .7H20 (557 mg) dissolved in 100 ml o-F water and 5 m1 o-F
this solution was used -For 1 litre], Sucrose (30 qm/1) Agar
(0.8%) -Forti-Fied with 2,4-D (2 mg/1), Nicotinic acid (0.5 mg/1),
Pyridoxin HCl (0.1 mg/1), Thiamine HCl (0.1 mg/1) and Glycine (2
mg/1).
Medium No.4 LS C Semisolid ):
KN03 (3000 mg/1), l"lqSD4 .7H20 (185 mq/1), CaC1 2 .2H20
(150 mg/1), KH2Po4 (540 mq/1), [FeS04 .7H20 (557 mg), Na2EDTA (745
mq/1) dissolved in 100 m1 o-F water and 5 ml o-F this solution was
used -For 1 litre], Mnso4 .4H2 0 (22.30 mg/1), Znso4 .7H2 o (10 mq/1),
KI (1.0mg/l), H3 Bo3 (6.0 mq/1), Nicotinic acid (3.0 mg/1),
Thiamine HCl (2.5 mq/1), Pyridoxin HCl (5.0 mg/1), Inositol (100
mq/1), Glycine (2.0 mg/1) and Lactalbumin hydolysate or Ca~ein
hydrolysate (300 mg/1), Kinetin (1.0 mg/1), Sucrose (50 g/1) and
Agar (6 g/1).
- ~~---~-- --------.......
HRdium No. 5 ~( Liqu~-~ Potato extract 10% aqueous (-For 1 litre o-F medium, 100
gm o-F -Fresh potato tubers were weighed out and cleaned with
water, the sprouts on the tubers i-F any were.removed. The tubers
20

were cut into small pieces, boiled in a given volume o~ distilled
water -for 20-30 minutes and -fi 1 ter-ed tht-ouqh two layers o~ cheese.
cloth. The residual potato pieces were extracted once aqain in
th~ same manner. Finally, the two -filtered liquids were mixed ~or
immediate use), KN03 (100 mq/1), (NH4 >2so4 (100 mg/1), Ca(N03 >2
( 100 mg/1), MgS04 • 7H20 ( 125 mq/1), t<H2Po4 (200 mg/1), KCl (35
""'"'""A-~~~ mg/1), Sucrose (100 gm/1) and Agar (8 gm/1). ~- !~- Q
Medium No. 6 E10G ( Liquid ):
G5 medium without agar and -forti-fied with 2, 4-D (1
mg/1), BAP (0.5 mg/1), IAA (0.5 mg/1), Glucose (5 gm/1),
Nicotinic acid (0.5 mg/1), Pyridoxin HCl (0.5 mg/1, Thiamine HCl
( 1 mg/ 1), Glycine ( 2 mg/1), Inosi to1 ( 100 mg/ 10) and Glutamine
(47 mg/1).
Medium No. 7 Modi~iad N6 (Liquid)& J
CNH4)2S04 (463 mg/1), KN03 (2830 mg/1), KH2Po4 (400
mg/1), MgS04.?H20 (185 mg/1), CaC1 2 • 2H2 o (166 mg/1), Mnso4 .4H20
(4.4 mg/1), ZnS04 .7H2 0 (1.5 mg/1), H3 Bo3 (1.6 mq/1), KI (0.8
mg/1), [Na2EDTA (745 mg), Feso4 .7H2D (557 mg) -dissolved in 100
ml o-f water and 5 ml o-f this solution was taken ~er 1 litre or
medium], -fortiFied with 2,4-D (4 mg/1), Nicotinic acid (0.5
mg/1), Pyridoxin HCI (0.5 mg/1), Thiamine HCl (1.0 mg/1) and
21

Glycine (2 mg/1) Sucrose (60 g/1).
Medium No. 8 E10YK ( liquid )I
G5 salts without agar and ~orti~ied with 2,4-D (1.0
mg/1), BAP (0.5 mg/1), IAA (0.5 mg/1), Glucose (5 mg/1),
Nicotinic acid (0.5 mg/1), Pyridoxin HC1 (0.5mg/l), Thiamine HCl
(1.0 mg/1), Glycine (2.0 mg/1), Myo-inositol (100 mg/1), Yeast
extract ( 3g/l ) and Kinetin ( 1.0 mg/1 ).
The media pH was adjusted to 5.8 in all cases and they ':)
were autoclaved at 1.06 kg/cmL ~or 20 minuites.
2.2.3.2.1 Callus proliTeration and regeneration•
Small calli were trans~erred to proli~eration media
viz. N-19 and J-19 on a paper boat. The composition o~ J-19 and
N-19 media are given below.
J - 19 (Liquid ) 1
B5 basic sal tsri thout agat- and ~orti~ied with sucrose
(20 gm/1), Inositol (160 mg/1), Kinetin (1mg/l) and NAA (1 or 2
mg/1).
N - 19 (Liquid ) 1
MS (Murashige,--J:_t-97-4) basic salts without aqar and
~orti~ied with sucrose (30 gm/1), NAA (1 or 2 mg/1), Kinetin
22

(1 mg/1) and Inositol (100 mq/1).
Calli thus obtained were trans~erred to di~~erent
regenerating media. Ten media i.e., MSA, MSB, MSI, MSBT, MSC,
MSD, MSBS, RGA, RG2 and N6 without hormones were tried ~or
regeneration. The composition o~ the media are given in Table 1:
Table 11 Di~~arent regenerating media with their composition•
Media ----~ ~~tokinins,.)
------ ----- -·
MSA MS+
MSB MS+
MSI MS ..
MSBT MS+
MSC MS+
MSD MS+
MSBS MS+
RGA MS+
RG2 MS+
and N6 without
NAA (mg/1)
1.0
0.5
2.0
0.5
1.0
1.0
0.5
1.0
0.5
hormone.
Concentration or /
Vitamins and Sucrose.
BAP Kinetin (mg/1) (mq/1)
Thiamine HCL (mg/1)
1.0 0.5
3.0
1.0 2.0
3.0 10.0 10.0
0.5 0.6
0.1 3.0
3.0
'2.0 0.6
5.0
23
Auxins,
Sucrose (gm/1)
30
30
30
30
30
30
60
60
30

2.2.3.2.2 Ploidy Status
Excised roots o-f about 1 em long were pretreated with
2 mH (h d ~. L y roxyqu1J-1ne at room temperature -for 2-3 hours to
arrest/accumulate cells at metaphase. These were -fixed in
Carney's -fixative (6:3:1 absolute ethanol : chloro-form . . glacial acetic acid ; with two changes). The roots were stored at
4°C -for at least 2 days which can be extended upto 60 days.
The -fixed roots were washed in distilled water -for 2-3 times (5-6 minutes each) and then in ice cold O.OlH citrate
bu-f-fer (0.01M citric acid monohydrate in 0.01M sodium citrate, pH
4.6) 2-3 times (2-3 minutes each). Root tips (2-3 mm) were cut
and trans-ferred to an ependro-f-f tube containing an enzyme
solution [2-6'X Cellulase (Onozuka R-10) and 1-27. Pectolyase
(Yakult Y-23) in citrate bu-f-fer] held at 37°C in a water bath.
The rootf tips were digested -for 45-75 minutes and washed in
0.01H citrate bu-f-fer 2-3 times (10 minutes each) and then in
sterile distilled water twice (5-10 minutes each) to remove the
enzyme solution (Islam and Stich, 1989).
Root tips were trans-ferred singly to an acid cleaned
and ethanol washed slide. E>=:cess water was removed without
disturbing the slide. A drop o~ 3 : 1 absolute ethanol: glacial
acetic acid was placed onto the root tip and squashed immediately
with a needle. Another drop o~ 3 : 1 -fixative was added to and
24

the ~aterial was spread~ over the slide. Slides were air dried
~ and stored in a des~icator overniqht.
~ Slides were examined under phase contrast microscope.
For better contrast the chromosomes were stained by placing a
drop o~ tX acetocarmine or 1/. propionic carmine over the air
dried cells and covered with a cover slip and heated brie~ly over
a spirit lamp. Excess stain was removed be~ore applying liqht
pressure to the cover slip.
2.2.3.3 Production of promising double haploids• I
2.2.3.3.1 Selection and hybridization
A~ter standardizing the media and environmental
conditions ~or callus induction and plant regeneration, crosses
were made between IR70 and Khonorullo. The crossings were carried
out by ~allowing standard procedures o~ emasculation in the
evening and pollination next morninq.
2.2.3.3.2 Culturing of anthers from Fl, regeneration and
production o~ double haploid lines
In the next season, Fl derived seeds obtained from the
crosses were grown sep~-ately. The anthers obtained ~rom them ~
were cultured for callLisinq in di~~erent media as mentioned
earlier. The resultant calli were transFerred to regeneration
25

""' medium.
Regenerated plantlets were transferred singly to pots
where they matured. The data on agrobotanic characters of Fl's
were recorded and seeds were harvested from fertile plants.
During the ne}~t year, seeds of
--------
di-f-ferent double / I
haploid~~--1 ) 'lines were multiplied in upland, lowland and
pots and double haploid ~~--Y 1 ines thus obtained
in
were
evaluated for yield and yield contributing characters under
medium altitude ( 950 m. msl ) lowland condition.
26

2.2.4 Field evaluation
The experiment was conducted on the ~arm o~ the ICAR
Research Comple;, ~or NEH Region, Barapani in the wet season o~
1994. The test entries, 9 DH lines (line no. 1, 6, 7, 10, 13, 15,
16, 19 and 21) and 2 parents (IR70 and l<honot-u 11 o) were evaluated ~
design; in a 3m X 1m plot in }._randomized block with -five
rt=;::>lications. Each plot contained ~ive rows with 30 plants/row.
Single seelding was transplanted/hill at 20cm X 10cm spacing.
Recommended dose o~ NPK ~ertilizer was applied.
2.2.4.1 Data collection
Observations were recorded on plants in the three
middle rows (excluding border plants) ~or ~allowing characters:
Panicle length: Average length ~rom ~ive plants/plot measured in
em at maturity.
Panicle weight: Average weight ~rom ~ive plants /plot weighed as
gm at maturity.
Percent sound grain: Calculated as percentage o~ ~illed grains
~rom total number o~ spikelets/panicle o~ ~ive plants/plot.
Plant height: In em, ~rom the ground to the tip o~ the main
panicle at maturity.
E5'ar bearing t i llet-s: • Average number o~ ear bearing tillers -from ~
~ive plants/plot at matlu-ity.
27

Grain weight/panicle: Average weight o~ grain ~rom ~ive
panicles/plot at maturity.
Thousand grain weight: Recorded ~rom evenly oven dried seeds o~
~ive plants/plot.
Number o~ spikelet/panicle: Average number o~ spikelets ~rom
~ive panicles/plot.
Grain yield: Weight o~ clean and dry grains harvested ~rom the
whole plot (excluding border plants) computed at 14X moisture
content in t/ha.
2.2.4.2 Data analysis
In order to have the basic in~ormation on the extent o~ ~
genetic variability~ statistical analvsjs were carried out on
analysis o~ variance~ coe~~icient of variation and heritability.
2.2.4.3 Analysis of variance
The analysis o~ variance was carried out to test the
genetic di~~erence between the genotypes under investigation
~allowing the method suggested by Gomez and Gomez (1984).
The analysis o~ var1ance was based on the assumptions
underlying linear model. A simple additive model is qiven below:
28

r :; ( v t
where i = j = 1 •....• n
k = 1 •••••••••• r-
Y i jk= the phenotypic value o-f genotype i j gt-own in Kth
block.
u= the qeneral mean
qij= the generatio~ e-f-fect o-f the ijth genotype
bk= the kth e-f-fect
eijk= the environmental e-f-fect associated with each
observation.
The u,qij and bk are assumed to be -fixed unknown parameters. The
eijk's are assumed to be normally and independently distributed
with mean zero and variance~ The analysis o-f variance table is
qiven below:
Source o.f d.f Mean sum o-f E~<pected mean variation squ.:u-e squat-e
,.... ..., Block r-1 1 "'- x"-,, .
j g j t_g
'"") x2 Genotype g-1 1 .-"'-A
i (f2 ? + r- (f~ g
r- i f"g
Error (r-1) (g-1) By subtraction
29

~ J-, 0 /~': .J-.., ._2 ""i
Total rg-1 ;-:: I ' A ){...:. <_:
ij i rg
Various mean squares were tested against error variance by using
F test. The standard error o~ mean and di-f-ference o-f
means were calculated as ( EMS/t-) 11 2 and ( 2 EMS &/r) 1/2
respectively where EMS is the errot· mean square.
2.2.4.4 Coefficient of variation
For comparing the extent o-f variability between di-f-ferent
characters, coe-f-ficient o-f variation was calculated at phenotypic
and genotypic levels using the methodology o-f Burton ( 1952 ).
Phenotypic coe-f-ficient o-f vat·iation ( PCV ) :
PCV= X 1 (H)
X
Genotypic coe~-ficient o-f variation ( GCV )
~~ -l ' q.'
GCV= '..___./. X 1 00
30

Where p and are phenotypic and genotypic variances
~ obtained ~rom the expectation o~ mean squares in the analysis o~
variance tables andVs the qenet-al mean o~ the populations.
2.2.4.5 Heritability /:::::\
Heritability <tH )) in a broad sense was calculated by the L./
method described by Allard ( 1960 ) and was de~ined in percentage
where, -? ~~ h2 = Heritability is percentage
u 2 p = Phenotypic variance
u2g = Genotypic variance.
2.2.4.6 Genetic advance
It is calculated as
GA = vg :X ' K
~ vP
Where K is the selection di-f-ferential based on phenotypic
deviation ( K = 2.06 at 5/. selection intensity level )
Genetic advance as percentage o~ mean is calculated as
GA GAl. = X 100
mean
31

2.2.4.7 Correlation
Correlation coef~icient ( ,-) between various characters
studied were worked out according to the method suggested by
Robinson et al. ( 1951 ). The ~orm o~ analysis o~ covariance is
given below:
~ Analysis of covariance
Source d~ Mean sum 0~ E}~pectat ion o-f mean Products Sum of products
Replication r-1 r12 ee12 + ger12 J
) ~l Genotype q-1 t12 ee12 + reg12
Error ( r-1) ( g-1) e12 ee12
Also, di~ferent simple correlation coefficient among different
characters were worked out:
Phenotypic coe-fficient o~ correlation ( rp12 ):
-11 2 2
rp 12=u -l u-- >~ u--P12 Pt P2
Where,
rP12 = Correlation between phenotypic values o-f character 1 and 2.
2 u--- = Variation o~ phenotypic values o~ character 1
Pt
32

'") . ~ u--- = Variation o~ phenotypic values o~ character 2
P2
2 u--- = Covariance between 1 and 2
Pt2
The signi~icance o~ phenotypic coe~~icient o~ correlation was
tested against "r" values and error degrees o~ ~reedom at p=0.05
and p= 0.01 levels.
2.2.4.<6 Path coe~~icient analysis
Path coe~~icient is simply a standardised partial regression
coe~~icient and as such measut-es the direct in~luence o~ ~ one
variable upon another. Path coe~~icient analysis was done to
partition the total correlation into direct and indirect e~~ects
by usinq the concepts o~ Wriqht 1921 ). The phenotypic
correlation among di~~erent character combinations were utilized
~or this purpose. The path coe~ficient analysis was per~ormed in
accordance with the method given by Dewey and Lu ( 1959 ). The
basic theory for the calculation of path coe~ficient is as
~allows:
33

Where, ·
ryi is the total correlation between x 1 ·s (independent
variable) and Y ( dependent variable )
Pyi is the direct e~~ect o~ variance x 1 on Y
Pvn ezn is the indirect e~~ect a~ variable xi on Y through vat- iab le xn
Then the equation in matrix ~arm are as under:
=
Hence = }:: n .-, r yi
Where,
( R >,. x n = Correlation matrix
Pyi = Column vector o~ path coe~~icient
ryi = Column vector correlation coe~~icient o~ individual
trait with dependent variable
34

It ( R )-l r x n = ( C ) r R n 'the path coe~~icient are
calculated as under :
Direct e~~ects:
Ply = C11 rly + c12t-2y + c 1 -,t-"7 + . . . . . . . . . . ._:. ·-'Y
p2y = C21 rly + C22r-2y + c23r-3y + . . . . . . . . . . Pny = cnl t•l y + Cn--:.t-'1,. + Cn3t-3y + . . . . . . . . . .
.c. ..:..y
worked out as Piyrni·
P 1yr 12 is the indirect e~~ect o~ x1 on Y via .X2 :
P2yr23 is the indirect e~~ect o~ X2 on Y via X3 :
and similarly all the indirect e~~ects were calculated.
Residual e-f-fect:
The variation in Y may be due to some other variables
not studied, e~rect or pseudo variables which have no
correlation with any other independent variable etc. It is termed
as residual e-f~ect.
35

3.RESULTB
3.1 Anther culture- callus induction and plant reqeneration
from local cold tolerant cultivars
Microspores o~ rice during anther culture are known
to give riset:ingle or multiple calli which can be regenerated
into plants. In order to apply anther culture technique in rice
breeding ~or compressing the breeding cycle~ anther culturability
o~ the local varieties was ~irst tested. Apart ~rom in vitro
responsiveness~various physical parameters and suitable culture
conditions are also needed to be standardized. There~ore,
36

experiments were ~irst undertaken to work out suitable conditions
~or induction o~ callusing in the anthers.
3.1.1 Standardization OT conditions ~or callusino OT microspar•
3.1.1.1 Time DT collection DT panicles
The panicles collected in the morning hours gave better
callusing response compared to the panicles collected at
di~~erent times though the di~~erence was not signi~icant. The
experiments were done with panicles collected in the Torenoon
(between 8 AM to 10.30 AM ).
3.1.1.2 Environmental conditions at the time a~ collection a~
panicles
A signi~icant di~~erence in ~requency o~ callusinq was
observed in the panicles collected on di~~erent days. In order to
~ind out optimum conditions~ panicles were collected between 25th
August to 30th September and J~ept in cold at ___g_if.tlrrent
t~ TOr cold pretreatment. The range 0~ temperature,
humidity and sunshine hourL during the period when anthers were
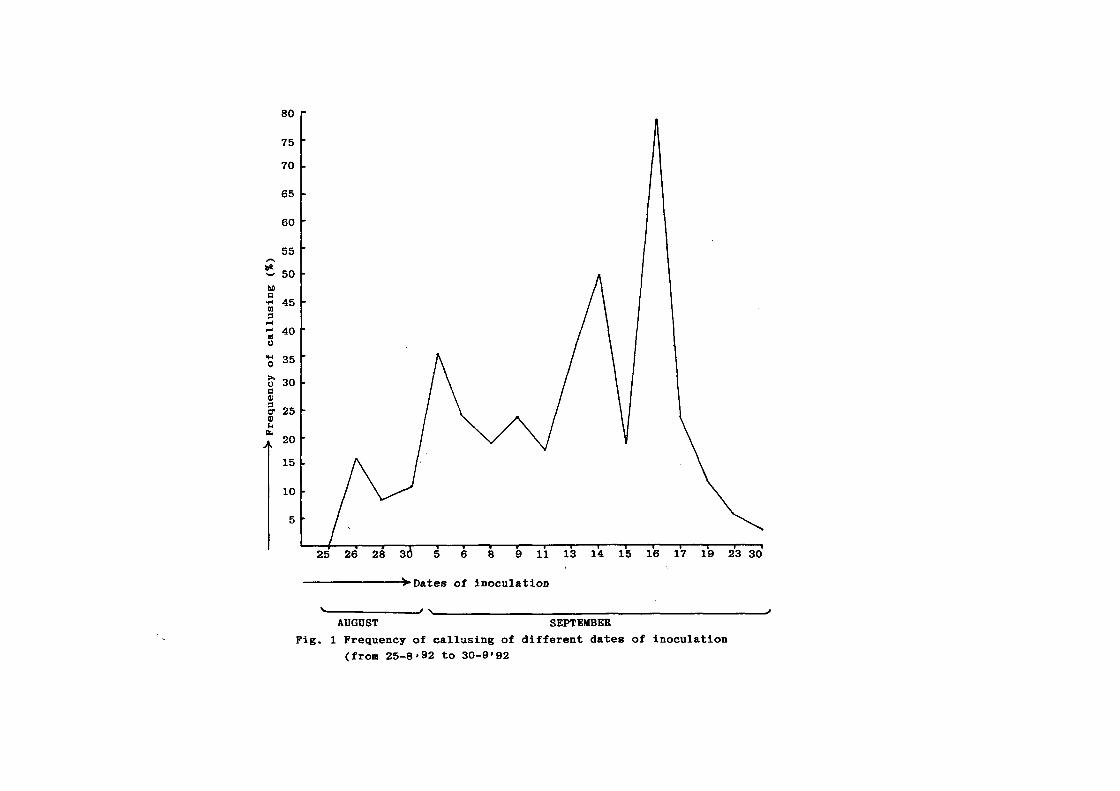
collected vis-a-vis ~requency o~ callusing is presented in Fig 1
and Table 1 •
Perusal o~ Fig 1 indicates that callusing ~requency
was maximum (78Z) in the anthers taken ~rom the panicles
collected Jn 16th ~ptembet- when the tempet~ature ranqed ~rom
37

Tabla 1 I Frequency of callusinQ in the anthers ccll•ct•d on
di-Fferent dates ( -from 25.8.92 to 30.9.92 )
Datr of No. of No. of I Tr•paraturl' Hu1idit y Total colll'ct ion anther cal luling callusing "ax. "in I 1 I Sunshint
inoculated antht!rs ( in 'c J hour
25.8.92 30 NIL NIL 29.8 21.4 83 8.52
26.8.92 400 63 15.7 27.2 20.5 88 2.23
28.8.92 1600 150 9.3 28.2 20.0 95 4.77
30.8.92 390 42 10.7 27.2 20.8 79 8.25
5.9.92 320 112 35.0 28.4 18.5 90 9.50
6.9.92 970 226 23.2 28.0 19.6 74 9.20
0.9.92 420 77 18.3 27.0 19.6 90 3.62
9.9.92 710 163 22.9 28.3 19.6 93 7.55
I 1. 9. 92 300 52 17.3 27.3 19.3 87 6.23
13.9.92 200 68 34.0 26.7 19.0 78 5.70
14.9.92 200 96 48.0 26.5 18.6 81 6.20
15.9.92 1600 290 18.1 24.8 18.0 93 4.75
16.9.92 205 160 78.0 26.3 17.2 83 3.10
17.9.92 1700 340 20.0 24.6 17.8 100 2.50
19.9.92 2500 270 10.8 26.6 17.2 77 8.70
23.9.92 so 4 5.0 20.8 19.6 89 5.00
30.9.92 600 20 3.3 24.4 18.0 DB 2.60
80
75
70
65
60
55 ""' ~ 50 bD c
..... 45 rn = .-< '; 40 C)
'E 35
t' 30 c Q)
& 25 Q)
"" "" 20
15
10
5
25 26 28 3 5 6 8 9 11 13 14 15 16 17 19 23 30
--------------~~Dates of inoculation
AUGUST SEPTEMBER Fig. 1 Frequency of callusing of different dates of inoculation
(from 25-8•92 to 30-9'92

l7.2-26.3°C. humidity 83%- 100% ~nd sunshine. 3.10 hour/day. In
addition, Fig.l also shows ~requency o~ callusinq ranginq ~rom
9.3 - 48% during Auqust 28th to September 14th.
3.1.1.3 Length o~ cold pretreatment
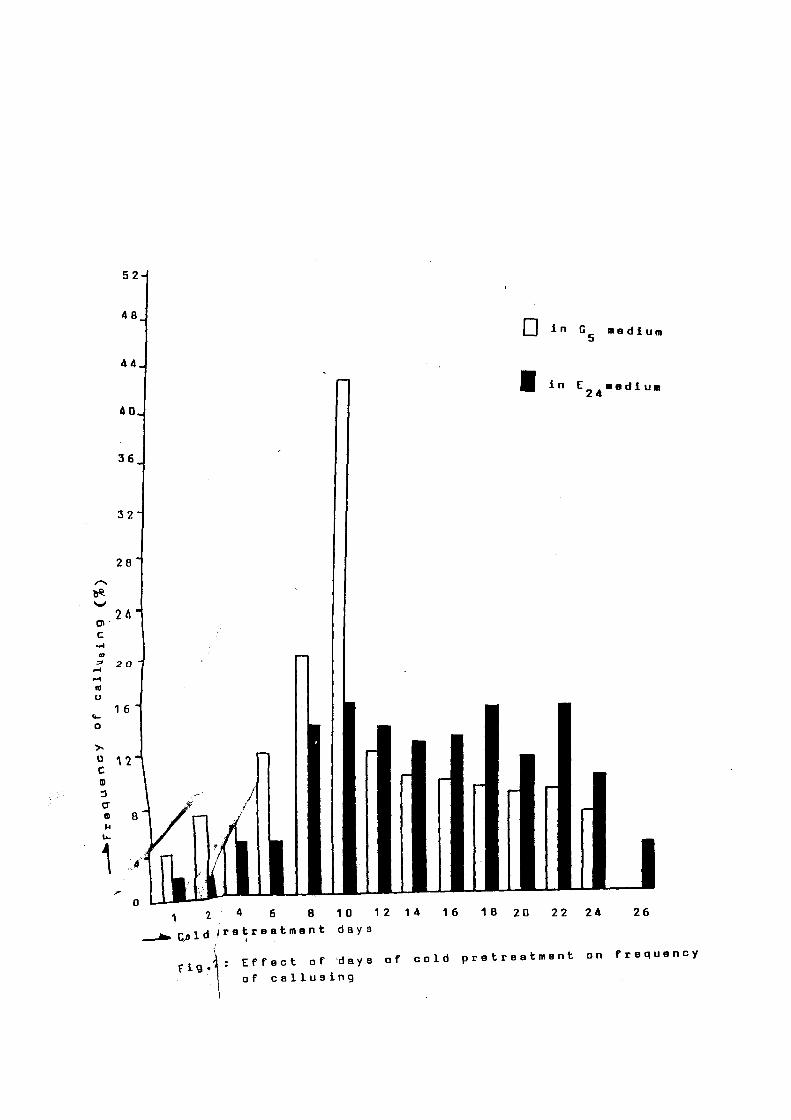
Cold pretreatment is known to exert pro~ound e~rect on
anther callusing. In the·present investigation it was observed
that ~requency
pretreatment (at
or callusing increased with the period
8-10° C). in both the media viz; G5
o-f cold
and
which were tested. Two to 4 % callusing was observed in the
anthers which were treated -for 1 day. Therea-fter, -frequency o-f
callusing increased with increase in ~ o-f cold pt-etreatment
and reached maximum (43.3% in G5 and 16% in E24 > a-fter 10 days o-f
pre- treatment. A-fter prolonged pre-treatment beyond 10 days, the
-frequency o-f callusing decreased abruptly -from 43.2% to 12.3% in
G5 medium a-fter 12 days. In case o-f E24 medium. it slowly
decreased with increase in days o-f cold pretreatment and reached
4% a-fter 26 days o-f cold pre treatment. None o-f the anther
callused in G5 medium a-fter 26 days o~ cold pre treatment
(Fig.2).
The
pr-ett-eatment
present study, thus. indicates that
10 days to the anthers
cold
be-fore
inoculation, is optimum -for obtaining maximum callusing in local
rice cultivf-s.
3R

3.1.1.4 Staging o~ micro5pores
Culture o~ anthers containing microspores at various
developmental stages ~ known to in~luence the ~requency o~
callusing. There~ore,
~evelopmental out a suitable stain
in order to inoculate microspores at
stage?, an attempt was -first made to -find J
to ascertain the exact developmental
staqe o~ the microspores. Anthers -from di-f-ferent parts o-f panicle
were stained with various stains and the results obtained are
pt-esented below:
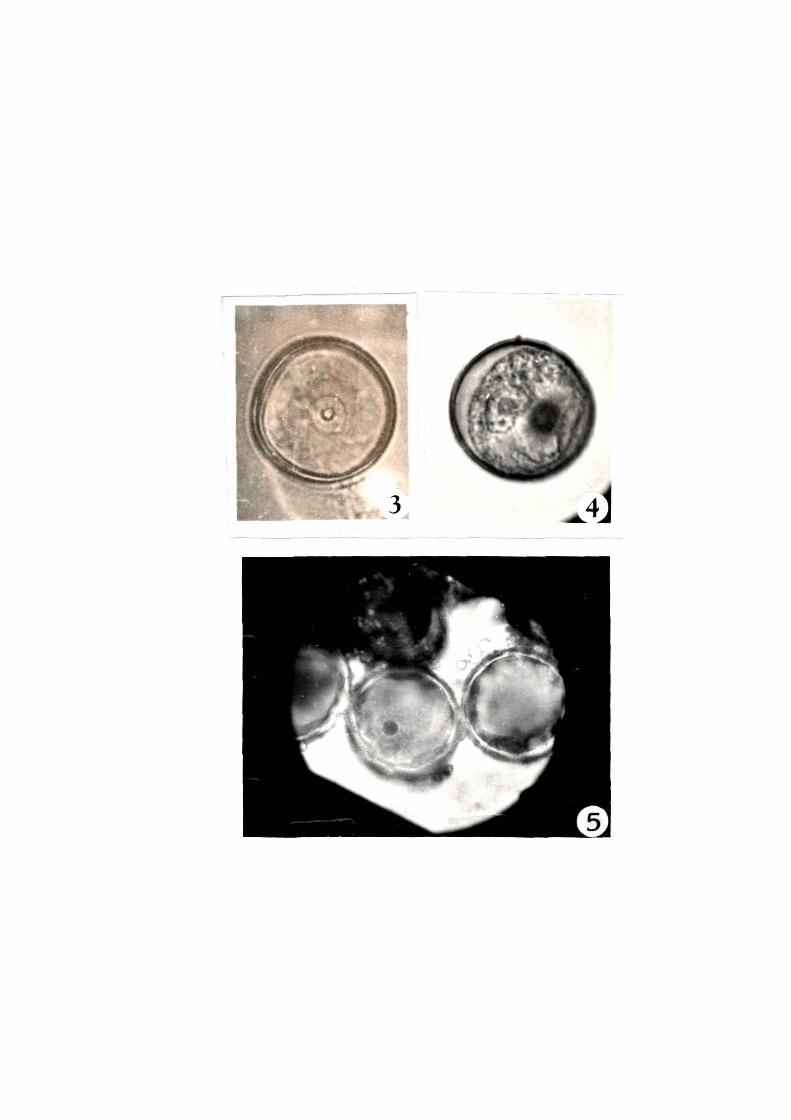
3.1.1.4.1 Staging•with acetocarmine stain
Mict-ospore nucleus could not be seen distinctly in_/
sqash preparations o-f 2% acetocarmine. Even the addition o~
mordants did not help in distinguishing nucleus ~rom the
cytoplasm (Fig. 3). There-fore~ tilis stain was not ~ound suitable
~or staging o~ rice mic~ospores.
3.1.1.4.2 Staging with Feulgen stain
The nuclei were seen distinctly using this stain
(Fig. 4). They appeared magenta coloured while cytoplasm remained
colourless. But this technique was time- consuminq and required
minimum 2 to 5 hours. There~ore. it had limited application,
especially when large number o~ anthers were to be inoculated
everyday.
39
52
48
44
40
36
32
28 .,.... ~ v
24 lJI• c ...! tiJ
:3 20 ...... ...... Ill u
.._ 0
'>-u c Gl ':l tr Gl ... t.-
\ ,-
'I 2 4 6 8 'I 0 1 2 1 4
__. C.D 1 d I r a ~ r a a t m a n t d a y s '
1 6
0 in G 5
I in
18 20 2 2 24
medium
26
f i 9
• ~ : E f f e c t o f 'd a y s o f c o l d p r e t r e a t m e n t o n f r e q u e n c y
\ of callusing

3.1.1.4.3 Staginq with iron alum - haematoxylin stain
Haematoxylin with iron alum stained the microspore
nuclei very distinctly. Nuclei were stained deep brown against
colourless cytoplasm (Fig. 5). Usinq this staining procedure,
lan1e numb~r oF spikelets could b~ staged within a short period
because microspores could be stained within 2-5 minutes and thus
this stain was used routinely.
3.1.2 Effect of different culture media on callusing frequency
and callus proliferation
Altoqether 12 semisolid and 10 liquid media
were tried ~ wR&cK the induction o~ callus~ in the local
cultivars. Preliminary screenina reve~led callus induction in
all the media but, there was wide variatio~ in the ~reauency. In -------------------'--·-~-----· ----- ---
order to narrow down the number o~ media and to Find out the
responsive one (s), only those media were tested which induced 5%
or more callusinq. Three semisolid media viz •• G5 , E24 ~minimal
and two liQuid media viz., N6 and E 10G were ~ound promising and
thus these were used For Further study. Anthers o~ Khonorullo,
RCPL 1-lC, RCPL 1-2C~ RCPL 1-3C, Pusa 33 and DR 92 were
inoculated in semisolid G5 , E24 (both containing B5 basic salts)
& minimal, and liquid E10G (containina B5 basic salts) L modiFied
N6 media.
40

Fig. 3 I Microspore stained with 2'Y. acetocarmine stain ( X 750 )
,">
tiU
Fig. 4 I Microspore stained with t=eulg~ stain ( X 750 )
i
Fig. ~ I Microspore stained with iron-alum haematoxylin stain ( X 750 )
4

It was observed that in case o~ semisolid medium
the anther started browning during ~irst three to ~our weeks o~
inoculation. Subsequently~ callus mass bursted out o~ the anthers
and the anthers turned deep brown a~ter 40-45 days o~ inoculation
(Fig.t, 6 L 7). The calli were cream coloured with organised
sections which appeared as embryoids.
In most o~ the cases microspores started callusing
a~ter ~our weeks o~ inoculation and continued upto ~our months.
The percent callusing was 13. 46. 33. 8 and 0.4% in 1st~ 2nd,
~ 3rd, 4th and 5th months respectivlv a~ter inoculation • Maximum
I ~ I callusing was, however observed between 40 to 50 days o~
J
inoculation.
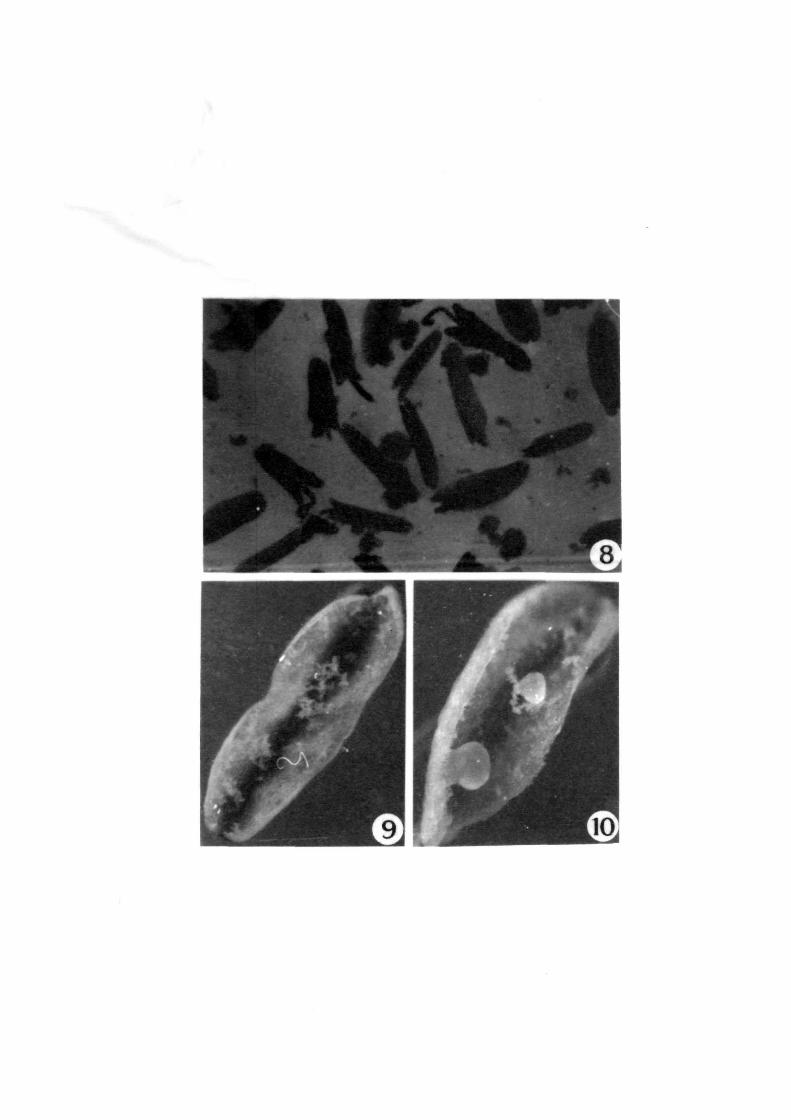
In case o~ liquid media (Fig. 8), anthers were ~ound
to split longitudinally thereby relefing microspores which were
seen as aggregates (Fig. 9). The microspores gave rise to calli
which remained attached to the anther wall in the initial stage
(Fig. 10) and were released a~terwards.
It was noticed that in most cases 200-300
microspores were released ~rom each anther a~ter splitting and
the maximum release was recorded between 4th to 6th day o~
culture. Three types o~ microspores viz •• highly elonqated- empty
; medium elongated- densely cytoplasmic and small unenlarged. in
the ratio o~ 5:4:1, respectively. were observed. Nuclear
41

~nJpaw
UOJ+e~aua6a~ o+ pa~~a~sue~+ aq o+ Apea~ Jtte3
tteM ~a4+ue a4+ ~o +no pa+s~nq Jtte~ 6UJM04S ~a4+u~
I
I
I
L
9
L-9
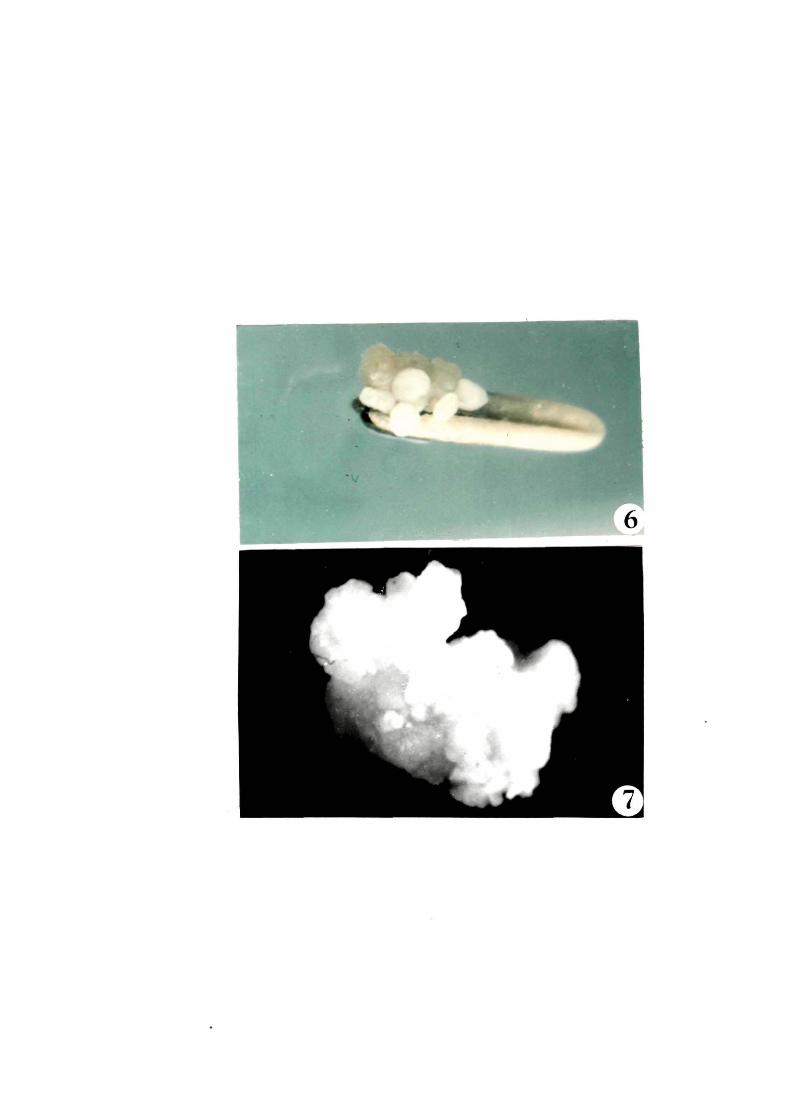

FiQB. B-10
B
.9
10
I
I
I
I
Callusing in liquid medium
Calli released in the medium ( single arrow) ( X 750 )
Longitudinal split anthers showing aggregates of microspores&( double arrow ) ( X 750 )
Anthers showing attached calli ( X 750 )

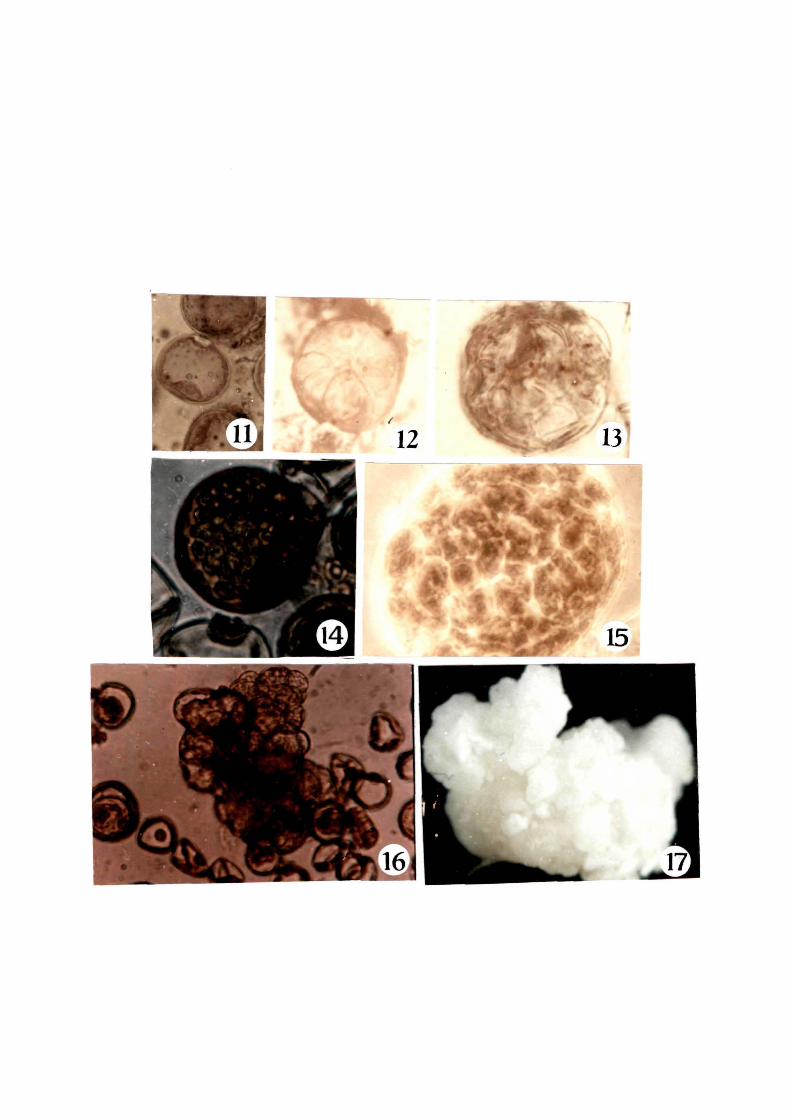
divisions occured mostly in the medium ~elongated and densely - . ~
cytoplasmic microspores a~ter 3rd day o~ culture and it continued
upto 16th day with wall ~ormation (Fig. 11-13). Further divisions
and wall ~ormation resulted in the ~ormation o~ enlarged ,--, /-----~---._""
multicellular r'~ generally composed o~ 50-60 cells -~-------
(Figs 14 & 15). The exine o~ the multicellular structur~
eventually bursted out and around 30th day. small calli were
released in the liquid mPdia. Globular and heart shaped calli
were observed a-fter 55 days o-f culture (Figs. (lb-~\~-< 17).
77"1. "J- In
qenerally
The overall -frequency o-f callusing ranqed -from 4.5 to
case o-f Khonorullo~ the ~requency
oood in almost all the media.
o-f callusing
However. liquid
I was\,
medium was the best where 57"1. callusinq was observed. In case o~
semisolid medium~ 50"1. callusinq was -found in G5 medium (Fiq. 18).
In some tubes callusing -frequency was as high as 98"1. in G5 medium
(Fig. 19).
In case o~ RCPL 1-lC, it was observed that liquid
medium~ E 10G was the best. where highest callusing (77''1.) was
observed. However, in semisolid medium, highest ~requency o-f
32.4% callusing was observed in E24 medium.
In RCPL 1-2C • the hiqhest callusino was recorded
in semisolid media where it was 45.5% in G5 • 3~).4% in E24 and
30.6% in minimal medium. In liquid N6 and E10G medium. the
42

Figs. 11-17 1 Various developmental stages o~ the microspores in liquid medium
" . '
11 1 Uninucleate microspore ( X 750 )
12 1 Microspore showing initial division ( X 750 )
13 1 Multicellular pollen ( X 750 )
14
17
1 Further divisions give rise to multicellular structure ( X 750 )
1 Individual multicellular pollen grains enclosed to show the densely staining cells ( X 750)
1 Microspore exine bursts out releasing microcalli ( X 750 )
1 Microcalli ready ~or trans~er to regeneration medium

~requency o~ callusing was 22.0 and 20.0% respectively.
In RCPL 1-3C. callusinq was equally good in both
semisolid and liquid medium. Fi~ty~our percent callusing was
observed in E24 semisolid (Fig. 20) and 58% in ElOG liquid
medium. However. only 5% callusinq was observed in G5 medium
which was generally good ~or other varieties.
In general, anthers o~ Pusa 33 did not respond
well in almost all the media and the ~requency o~ callusinq in
both semisolid and in liquid media ranqed ~rom 6.1% to 15% only.
DR92 anthers responded well in G5 semisolid (48%)
and N6 (31%) liquid medium. In minimal medium they did not
respond and the ~requency dropped to 4.5%. Data on the ~requency
o~ callusing in anthers o~ six varieties in semisolid and liquid
media at-e collated in~)ble 2.
Perusal o~ data on callusing o~ anthers in six
varieties (including parents and their progeny sister lines)
indicated a strong genotypic e~~ect. In qeneral, it was observed
that together with genotypic e~~ect , media containinq B5 salts
like G5, E24 and ElOG responded well to callusing o~ anthers in
local cult i vars except in Pusa 33 whet-e h ir--~hest ~t-equency o-f
callusing was observed in N6 medium. Khonorullo was in vitro
responsive variety whereas Pusa 33 was a recalcitrant one with
low ~requency o-f callus induction in almost all the media.
43

There~ore, apart ~rom presence /availability o~ appropriate
nutrients, genotype OT the donor plant exerts a
inTluence on callusing. Liquid media were relatively better
semisolid ones.
Proli~eration o~ calli
strong
than
Small pieces OT calli (weighing 10 to 15 mg) were
transTerred to N-19 and J-19 liquid medium TOr proliTeration. The
tiny callus grew into a big mass OT 2 to 3 q in 10 to 15 days
(Fiq.21}. Both the media were Tound to be equally good ~or
supporting proliTeration, but addition OT 2 mg/1 NAA in both the
media increased the ,rate OT callus growth in some cases.
Frequency OT regen~~ation was however,aTTected. ProliTeration 1n
dark or in low intensity light, was Tound better TOr
regeneration.
44

Fig•. 18-20 I Callus induction on semisolid media
18 I Callusing in Khonorullo on semisolid G5 medium
19 I Very high -frequency o-f callusing in Khonorullo on semisolid Gs medium
20 I Callusing in RCPL 1-3C on E24 medium
21 I Callus proli-feration in J-19 medium

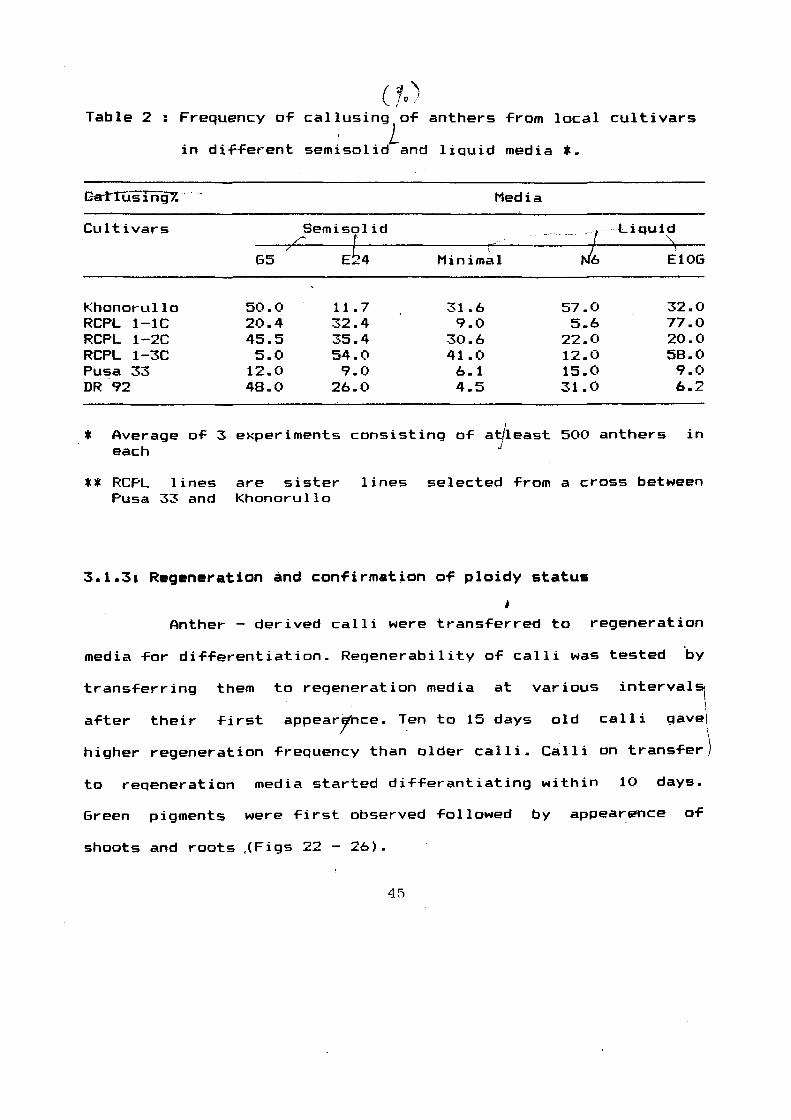
(}oJ Table 2 : Frequency o~ callusing o~ anthers ~rom local cultivars
. L in dif~erent semisolid and liquid media *·
c-rl1LiSTrlijY.·-·
Cultivars
Khonorullo RCPL 1-1C RCPL 1-2C RCPL 1-3C Pusa 33 DR 92
65
50.0 20.4 45.5 5.0
12.0 48.0
Semisolid 7 .::::::.
Eb4
11.7 32.4 35.4 54.0 9.0
26.0
Media
7 t J6 Minimal
31.6 57.0 9.0 5.6
30.6 22.0 41.0 12.0 6.1 15.0 4.5 31.0
· Li-quid ·,\.
I
E10G
32.0 77.0 20.0 58.0 9.0 6.2
* Average o~ 3 experiments consisting o~ at/least 500 anthers in each J
** RCPL lines Pusa 33 and
are sister Khonorullo
lines selected ~rom a cross between
3.1.31 Regeneration and ccn~irmation c~ ploidy status
I
Anther - derived calli were trans~erred to regeneration
media ~or di~~erentiation. Reqenerability o~ calli was tested by
trans~erring them to regeneration media at various
a~ter their ~irst appearfnce. ~en to 15 days old
higher regeneration ~requency than older calli. Calli
intervali
calli gavel'
on trans~er
to reqeneration media started di~~erantiating within 10 days.
Green pigments were ~irst observed ~allowed by appear~nce o~
shoots and roots ,(Figs 22 26).

It w~s also observed that proli~erated calli tend~ to
l~se their regeneration capability as compared to directly
trans~erred ones. In Khonorullo and RCPL 1-2C , small calli were
proli~erated in both J-19 and N-19 media and a~ter 10-15 days
they were trans~erred to MSB regeneration medium together with
10-15 days old primary calli. Out o~ 10 proli~eratin~ calli, only
one regenerated in MSB media,whereas ~rom the primary calli
directly trans~en-ed to MSB medium, around 501. showed l regeneration and produced green plantlets. There~ore~ in ~urther
experiments• only primary calli were used ~or regeneration.
Regeneration o~ calli obtained ~rom di~~erent varieties in
various regeneration media is presented in Bable 3.
Table 31 Regeneration percentage o-f anthar-darived calli on
dif-ferent regeneration media
Khonorullo
Media
MSI MSA MSB MSBT MSC MSD MSBS RGA RG2 N6-H
Total no. 0~ 12 16 16 8 12 6 3 5 5 9 tubes with calli
Number 0~ tubes '4 7 8 e 3 1 (- {! ·-./
1 2 green pigment a-tion
Plant 33 43 50 (- 26 16 / 20 20 regenera- t -t ' .. __...,/
tion percentage \ . ~/
46
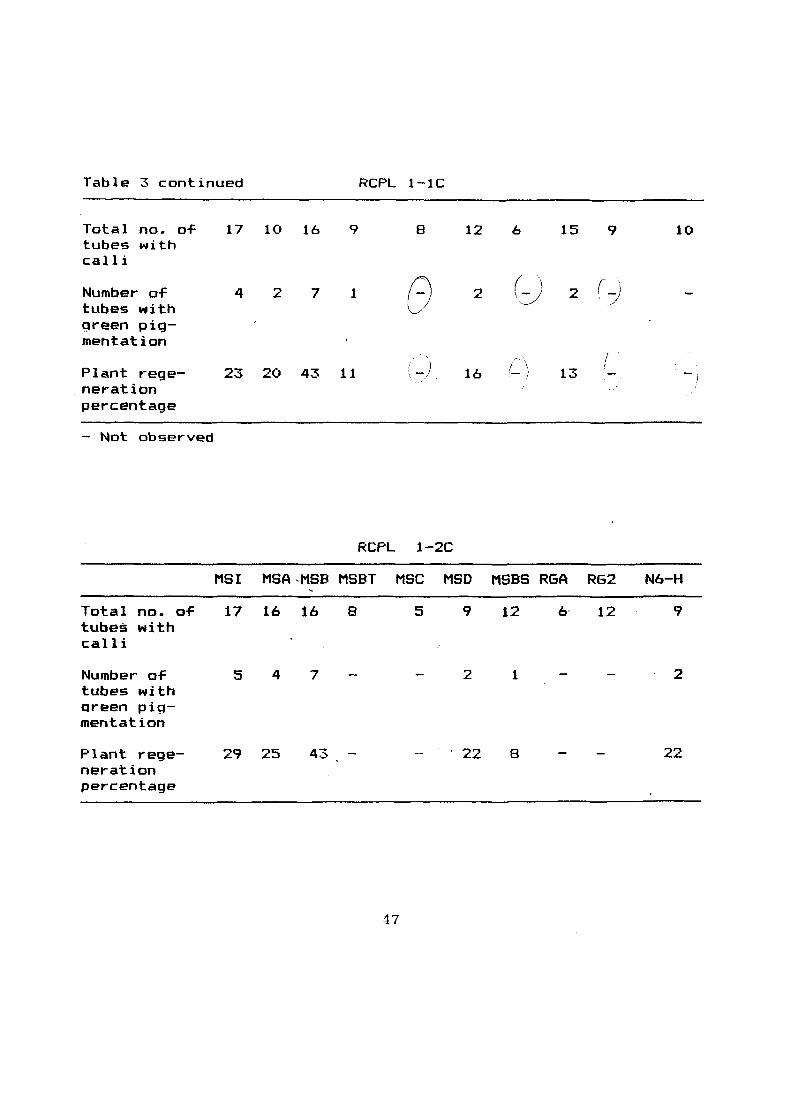
Table 3 continued RCPL 1-1C
Total no. o-F 17 10 16 9 8 12 6 15 9 10 tubes with calli
Number o-f 4 2 7 1 G 2 () 2 (j tubes with green pig-mentation
_) (_ \ I I
Plant rege- 23 20 43 11 16 13 -neration percentage
- Not observed
RCPL 1-2C
MSI MSA,MSB MSBT MSC MSD MSBS RGA RG2 N6-H
Total no. o-f 17 16 16 8 5 9 12 6- 12 9 tubes with calli
Number o-F 5 4 7 2 1 2 tubes with Qreen pig-mentation
Plant rege- 29 25 43 . 22 8 22 neration percentage
17

RCPL 1-3C
Total no. o-f 15 12 12 10 4 12 6 6 7 10 tubes with calli
Number oi= 2 2 4 2 1 1 1 1 tubes with qreen pig-mentation
Plant rege- 13 16 33 20 25 8 16 10 neration percentage
DR 92
Total no. o-f 6 5 9 4 5 6 6 6 tubes with calli
Number o-f 1 2 2 1 1 1 tubes with ~
qreen pig-mentation
Plant reg- 16 40 22 20 16 16 eneration pet·centage
- Not observed
48

Table 3 continued
Total no. tubes with calli
Number o-f tubes with qreen pigmentation
Plant regneration percentage
o-f
- Not observed
MSI
10
1
10
Pus a 33
MSA MSB MSBT MSC MSD
12 9 5 6 Is j C....-/
1 '2 1 1
8 22 20 16
't>'O \1 ~' ~~...,. ~\)r !+t..f lot el ') J Y.,..t-1- M f'{;...:J\... ;
MSBS RGA RG2 N6-H
r--: l ~~- .....
6 /- -
1
16
l<))
It was observed that light intensity o-f 2500 to
3000 lux is suitable -for plant regeneration. A continuous light
-for 5 days a-fter transfe~ to regeneration media followed by 16
hr/8 hr liqht/dark was -found to be the best -for regeneration.
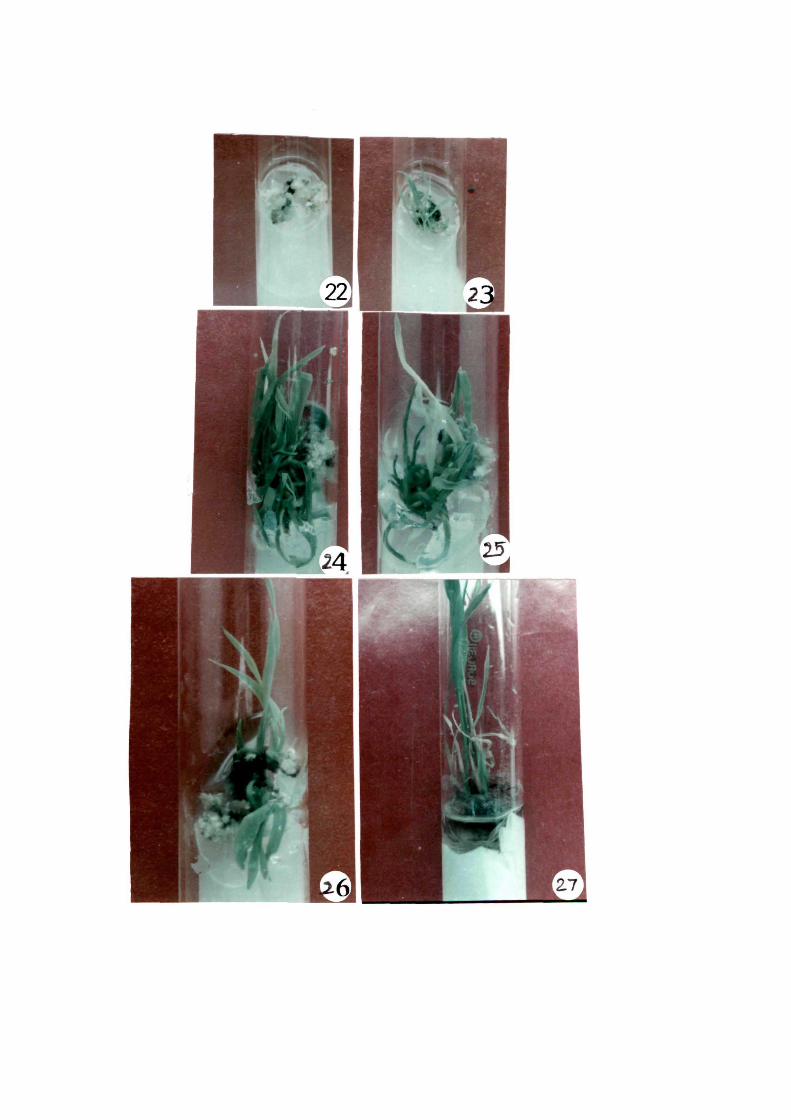
In Khonorullo, highest regeneration percentage
(50%) was observed in MSB medium (Figs 22 & 23) -followed by 43
and 33% in MSA and MSC. In regeneration media MSBT, MSBS and RGA
there was no regeneration response.
Almost all cultivars responded well in MSB
medium except DR 92 whet-e 40% regeneration was recorded in MSA
medium compared to 22% in MSB medium.
49

Forty three percent regeneration was recorded in RCPL
1-lC anthers in MSB regeneration medium ~allowed by 23, 20, 16,
13 and 11% in MSI, MSA, MSD, RGA and MSBT. There was no
reqeneration in MSC. RG2, MSBS and N6 media.
Forty three percent regeneration was observed in
RCPL 1-2C anthers on MSB medium followed by 29, 25, 22, 22 and B%
' in MSI, MSA, MSD, N6 and M~BS, respectively. MSBT, MSC, RGA and
RG2 media did not support regeneration. In cases o~ RCPL 1-3C
and Pusa 33 maximum regeneration o~ 33 and 22% respectivly was
observed in MSB.
It was observed that MSB , MSI and MSA supported
better regeneration than MSC, MSBS. MSBT, RGA, RG2 and N6, MSB
being the best ~or almost all local cultivars.
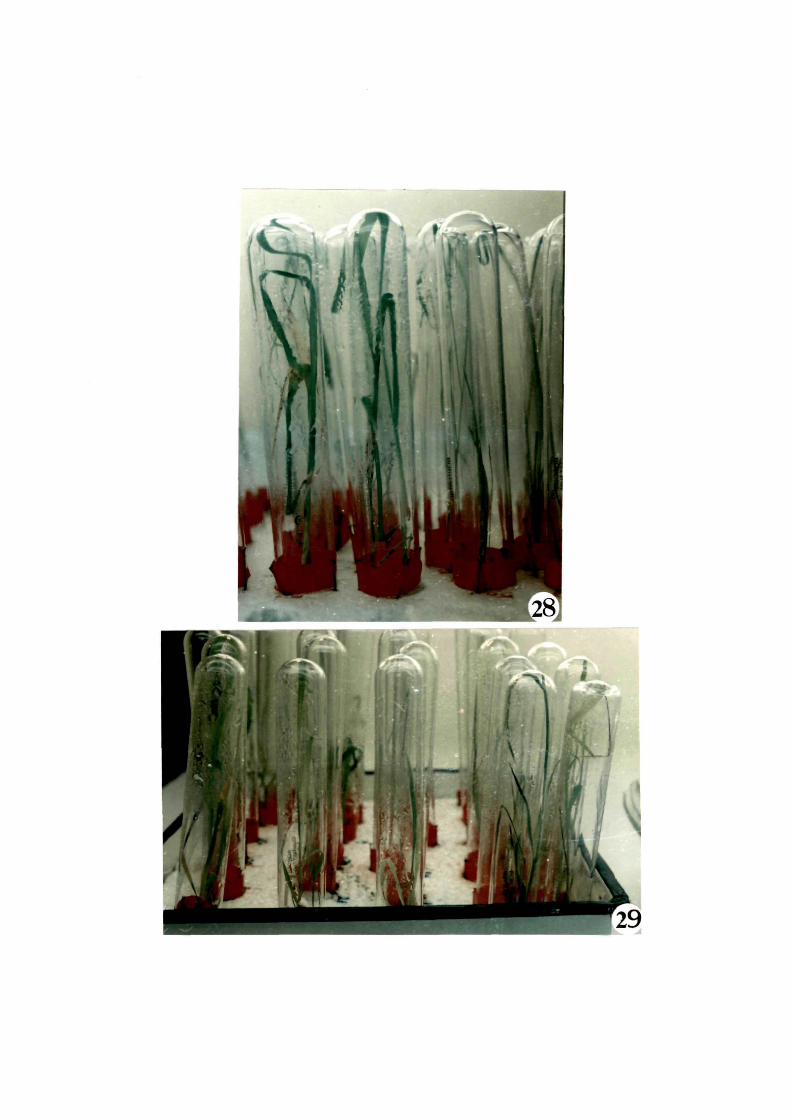
A~ter the plantlets attained 6 to 7 inches
height, they were trans~erred to Yoshida's liquid culture medium r----< and were covered with tubes to check evaporation (Figs 27-29).
A~ter about 15 to 20 days, they were trans-fer.red to pots for
~urther growth and maturity.
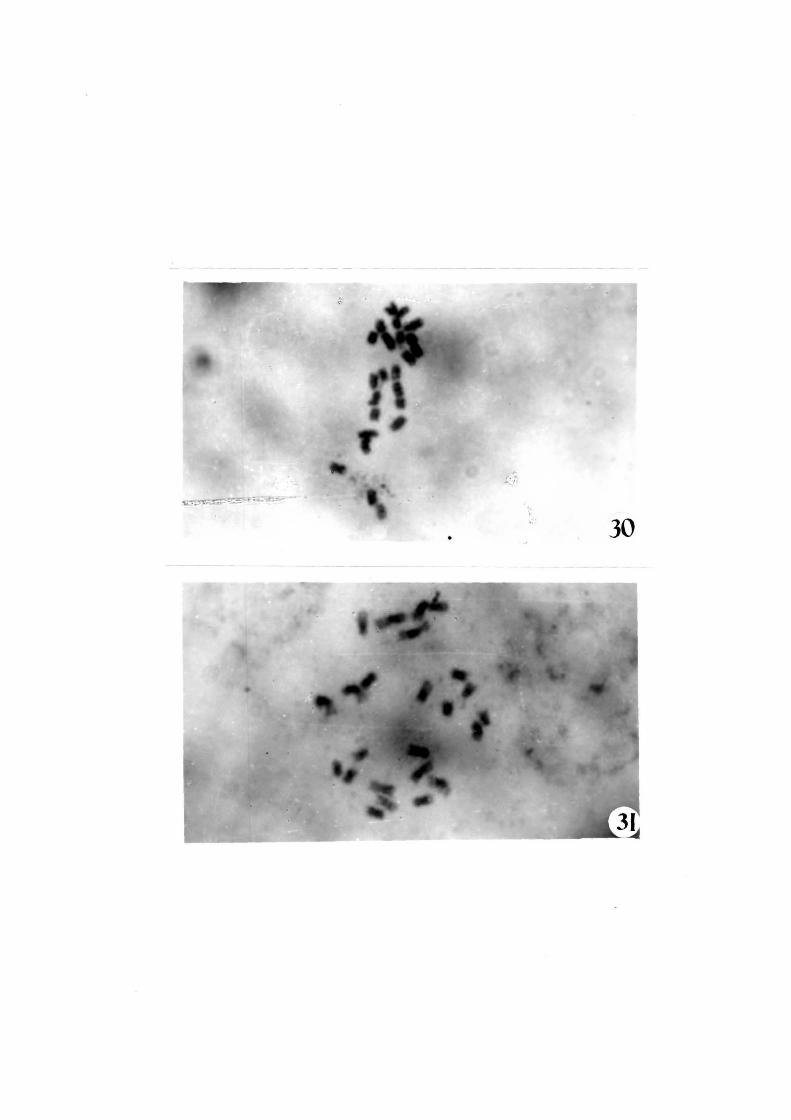
Ploidy status•
Root tip cytoloqy showed that most (70 to 75%)
o-f the regenerated plants showed diploid (Fiqs 30 and 31)
chromosome numbers, while 15 to 20Y. were haploids or mix~loids. f
50

Figs. 22-27 1 Various stages or plant regeneration rrom anther derived callus or local varieties
22 1 Initiation or greeen pigmentation in Khonorullo
23 1 Shoot rormation in Khonorullo
24 1 Leaf initiation in RCPL 1-1C
2~ 1 Plantlet or RCPL 1-1C
26 1 Plantlet in advanced stage in RCPL 1-1C and
27 1 Plantlet of Khonorullo on paperboat transrerred to Yosida's liquid medium

#
Figs. 28-29 1 Plantlets in Yoshida's liquid medium covered with tubes
28 1 Plantlets'o~ RCPL 1-1C
29 1 Plantlets o~ Khonorullo

Figs. 30-31 1 Squash preparation ~rom root tip o~ regenerated
plants showing diploid (2n=24) chromosomes
( X 3000)
.. ...... -~ ..
30

3.2 Production of double haploids
3.2.1 Selection of parents and hybridization
In order to trans~er cold tolerance to hiqh
yielding genetic background, IR 70, an elite breedinq line ~rom
IRRI1
and Khonorullo, a local cold tolerant cultiva~ ~ selected 1_'
for hybridization. IR 70 was used as ~emale whereas Khonorullo
was used as male. In all, 9 hybrid seeds were obtained.
3.2.2 Raising of F1 and their comparison with parents
The nine hybrid seeds were grown sep,rately in
the field. Data were recorded to compare the agrobotanic
characters o~ the Fl plants with their parents. Fl plants showed
hybrid vigour and performed better than both the parents in yield
and yield contributi~g characters (Table 4).
Days to 50% ~lowering (98 days) remained within
the range of both the parents which.was 109 and 91 days in IR 70
and Khonorullo, respectively. Lea~ length (29.0 em) was reduced
as compared to both the parents (34.0 and 36.5 in IR- 70 and
Khonorullo, respectively). Plant height remained within the range
o~ both the parents which was 87 em as compared to 60 and 131 em
in IR 70 and Khonorullo, respectively. Number of ear bearing
tiller increased remarkably to 7.1 as compared to 5.3 and 4.2 in
IR 70 and Khonorullo, respectively. Likewise, panicle length
51
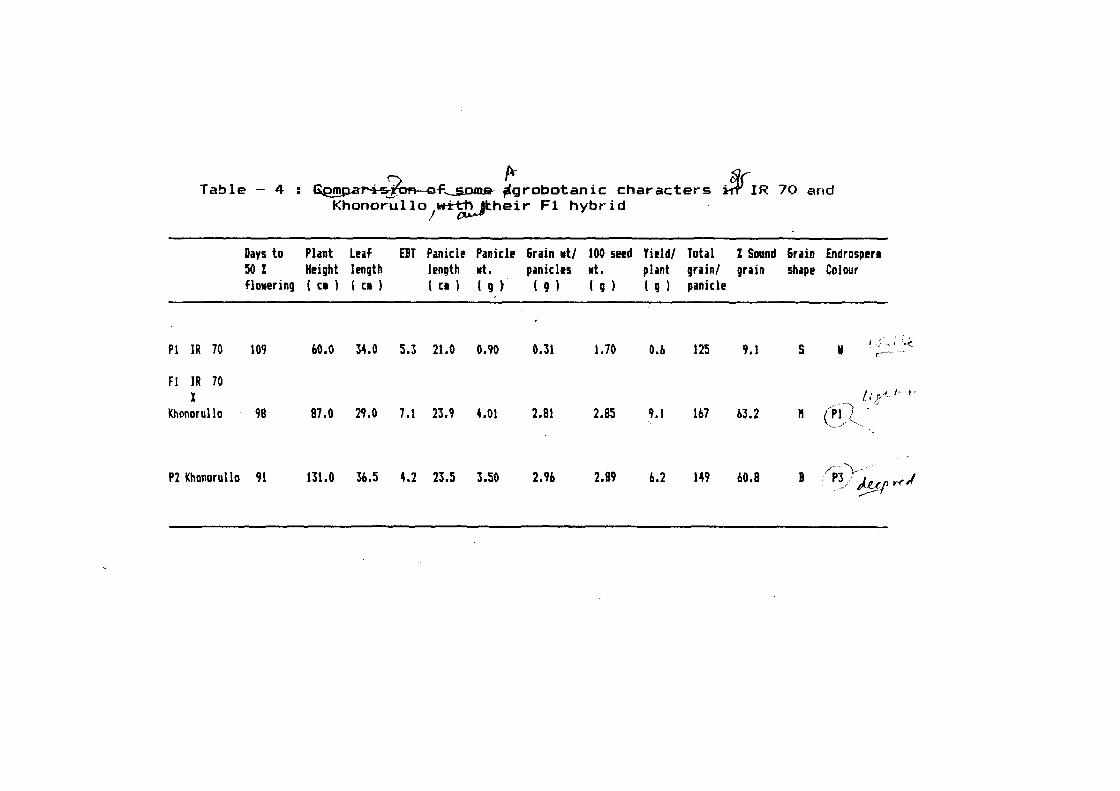
Table - 4 : ~.f.._..JiClfD.&-- ~robotanic characters $IR 70 and Khonorullo 1~~heir Fl hybrid
Days to Plant Luf EBT Panicl! Panicl! &rain •t/ 100 se!d Yi!ld/ Total 1 Sound &rain Endrosp!rt 50 % H!ight l!ngth l!ngth •t. panicl!s .t. plant grain/ grain shap! Colour flo•ering ( Cl ) ( [I ) ( [I ) ( g ~ ( g ) ( g ) ( g ) panicle
Pl IR 70 109 60.0 34.0 5.3 21.0 0.90 0.31 1.70 O.b 125 9.1 s II 1 .... ~ ' 1' ~·.<. r---
F! IR 70 X t ~· r-"'· r ~,c•
Khonorullo 98 87.0 241.0 7.1 23.9 4.01 2.81 2.85 9.1 1b7 b3.2 " .~P~--~ -(i
\,_./ ··,·-.
P2 khonorullo 91 131.0 3b.5 4.2 23.5 3.50 2.9b 2.89 6.2 149 bO.B B /~-\/
,' ~~/ ~-f rl' ,I

(23.9 em) and panicle weight (4.01 g) also increased ~rom the
range o~ both the parents. Grain weight per panicle (2.81 g) and
100 seed weight (2.85 g) remained within the range o~ both the
parents. Yield per plant (9.1 g) increased remarkably and
exceeded both the parents (0.6 q in case o~ IR 70 and 6.2 g in
case o~ Khonorullo). Total qrains per plant (167) also increased
and was higher than the parents. Per cent sound grain (63.27.)
also increased as compared to both the parents which was 9.17. in
IR 70 and 60.87. in Khonorullo. Medium grains were observed in
case o~ F1 as compared to slender grains in IR 70 and bold in
Khonorullo. Light red coloured endosperm was observed which was a
mixed character o~ both the parents; IR 70 was white where as
Khonorullo, possessed deep red endosperm colour.
3.2~3 Culturing of anthers derived from Fl
3.2.3.1 CallusinQ
Anthers ~rom F1 plants o~ the cross IR 70 X
Khonorullo were cultured as described previously (time o~
collection o~ panicle and preparation o~ staining solutions and
procedure) in 65, E24, L8 and MP2 medium. Percentage o~ callusing
ranged ~rom 11.7 (in G~ semi solid ·medium) to 33.67. (in E24
semisolid medium). Highest percentage o~ callusing was observed
in one tube containinq L8 medium (81.6%). "In MP2 medium. the
~requeny o~ callusing was generally good (31.1X). Frequency o~
callus induction in di~~erent media is presented in Table 5.
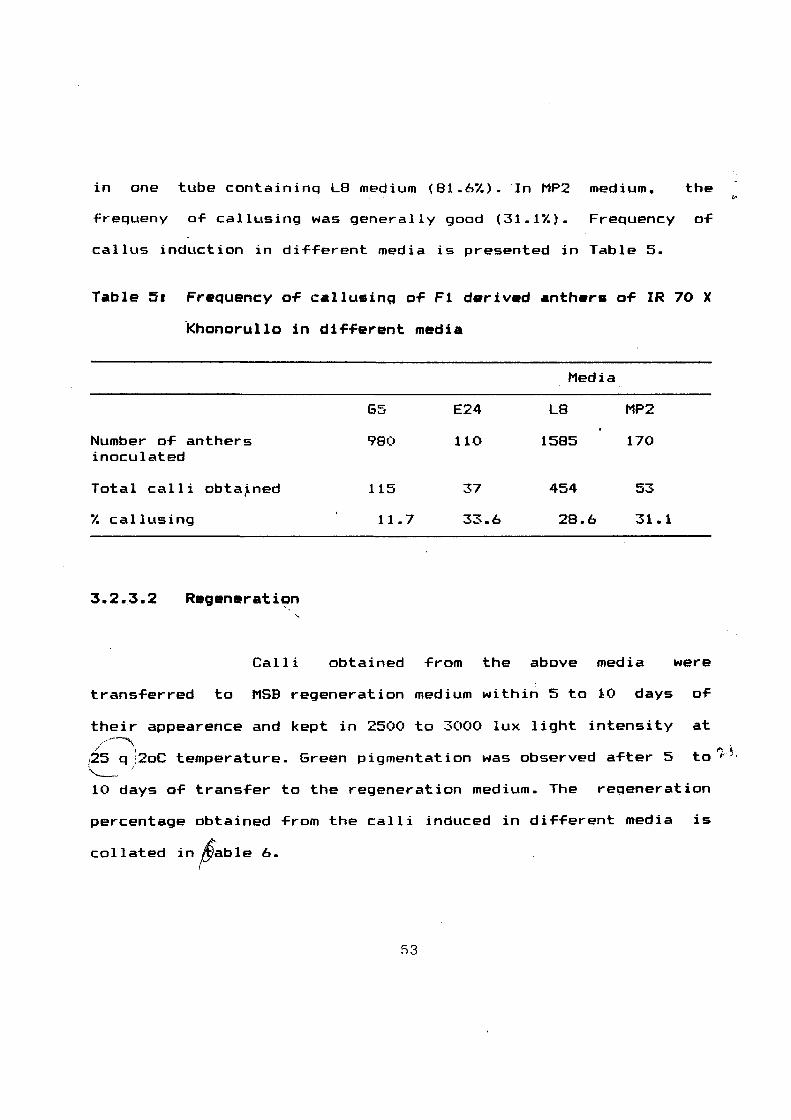
Table 51 Frequency o~ CAllusing o~ F1 d•riv•d •nth•r• o~ IR 70 X
Khonorullo in di~~erent media
Media
65 E24 L8 MP2
Number o~ anthers 980 110 1585 170 inoculated
Total calli obta~ned 115 37 454 53
'l. callusing 11.7 33.6 28.6 31.1
3.2.3.2 Regeneratic;m
Calli obtained ~rom the above media were
trans~erred to MSB regeneration medium within 5 to !0 days o~
their appearence and kept in 2500 to 3000 lux light intensity at ./---"'\
t25 q j2oC temperature. Green pigmentation was observed a~ter 5 to ··H. '"----· .' 10 days o~ trans~er to the regeneration medium. The reqeneration
percentage obtained ~rom the calli induced in di~~erent media is
collated in !able 6.
53
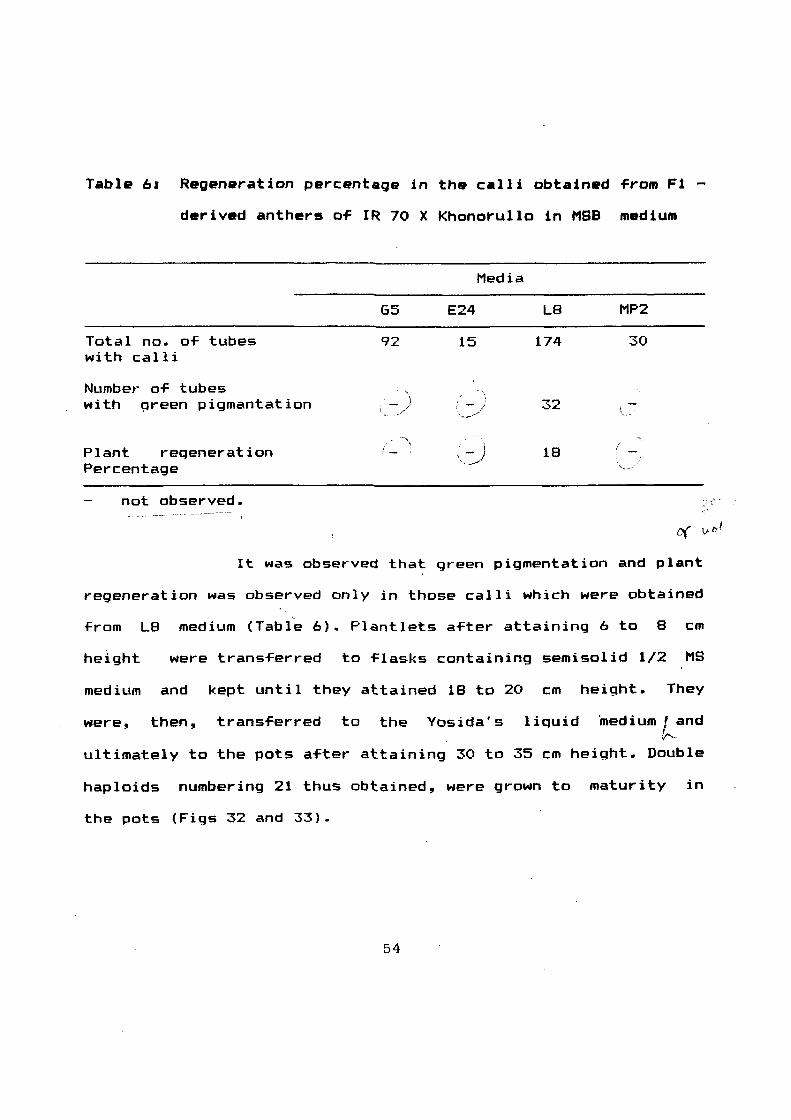
Table 61 Regeneration percentage in the calli obtained ~rom F1
derived anthers o~ IR 70 X Khonorullo in MSB medium
Total no. or tubes with calli
Number or tubes with qreen pigmantation
Plant reqeneration Percentage
not observed.
G5
92
--.\ /_
Media
E24 L8 MP2
15 174 30
32
18 (
It was observed that green pigmentation and plant
regeneration was observed only in those calli which were obtained
rrom L8 medium (Table 6). Plantlets arter attaining 6 to 8 em
height were transrerred to ~lasks containing semisolid 1/2 MS
medium and kept until they attained 18 to 20 em height. They
were, then, transrerred to the Yosida · s 1 iquid ·medium ! and ~
ultimately to the pots arter attaining 30 to 35 em height. Double
haploids numbering 21 thus obtained, were grown to maturity in
the pots (Figs 32 and 33).
54

Figs. 32-33 1 Double haploid plants growing in pots


3.2.3.3 Evaluation o~ double haploid (DH) lines
Out o~ 21 plants regenerated, nine showed promising
characters and they were selected. Seeds ~rom these lines were
collected and ~ound to be di~~erent ~rom each other. Agrobotanic
characters o~ the parents and the double haploid lines are
presented in ~~iguers 34 to 67.
Panicle type oF IR 70 was compact (Fiq. 34)
whereas that o~ Khonorullo was open (Fig. 36). Among double
haploid lines, panicle type varied ~rom compact as in line nos.
1,6.15,19 and 21 (Figs. ,38.40,45,47 and 48) to open as in line
no. 7 13 and 16 (Figs. 42,44 and 48). Line no 10 (Fig. 43) t._
exhibited intermediate type o~ panicle. Secondary branching was
observed in line nos. 1,6,15,19 and 21 (Figs. 38,40,45,47 and
48) whereas it was absent in the parents. Straight panicle was
observed in line nos. 6,13,15,16 and ·21 (Figs. 40,44,45,46 and I . .
48) whereas line nos. 1,7,10 and 19 showed droopy pariicle (Figs.
38,41,42 and 47).
Shape and size o~ grains o~ each line varied
greatly. It ranged From bold ( Khonorullo) to slender, ( IR 70 ).
Line no. 1 and 21 showed slender grains with thin and short hair
on upper portion o~ lemma and palea. (Figs. 51 & 67). Line no. 6
showed bolder grains with qlabt·ous lemma and~~-~~-~--==:~d) (Fig. 53). Line no. 7 and 10 possessed slender grains with
55

glabrous lemma and P~·._.~ubescen~J(Figs. 55 & 57 ). In ------
contrast~ I ine with qlabrous
lemma and pale pubescencF (Figs. 59 & 65). Line no 15 and 21
showed slender grains with thin and short hair on upper portion
o~ lemma and palea (Fig. 61 and 67). In line no. 16, qrains were
round with short hair (Fiq. 63). A typical character~ where lemma
and palea remained ope~at the tip was observed in most o~ the
grains o~ line no. 13 (Fig. 59). This character was-not ~ound in
any o~ the parents.
The endosperm colour, ranged ~rom dark red
CKhonorullo colour) to white (IR 70 colour) in the parents,
whereas~ it was intermediate in the progenies (Fig 70). Line no.
10 and 21 showed white k••~•l coloured grains. In most o~ the
lines, (line nos. 1~6~7~13 and 15) light red coloured kernel was
observed, whereas in 1 ine no 19 the kernel colour was •fiUiiiFI!!-"
darker. In line no. 16, the kernel colour was dark red like
Khonorullo.
Individual anther derived double haploid lines o~
. ~ the cross IR 70 X Khonorullo showed uni~ormity~ many agronomic
characters. The range o~ variability among the plants o~ anther
culture derived lines were within the limits o~ variability o~
their respective parents.
56
l

Fig. 34
Fig. 35
Fig. 36
Fig. 37
I
I
I
I
Panicle o~ Parent 1~ IR 70
Husked and dehusked grains o~ IR 70
Panicle o~ Parent 2, Khonorullo
Husked and dehusked grains o~ Khonorullo


Fig. 38: Panicle o~ line no. 1 with husked and dehusked grains
Fig. 39: Closeup o~ husked and dehusked grains o~ line no. 1


Fig. 40: Panicle o~ line no. 6 with husked and dehusked grains
Fig. 41: Panicle o~ line no. 7 with husked and dehusked grains


Fig. 42: Panicle o~ line no. 10 with husked and dehusked grains
FiQ. 43: Closeup o~ husked and dehusked grains o~ line no. 10


Fig. 44: Panicle or line no. 13 with husked and dehusked grains
FlQ. 43: Panicle or line no. 15 with husked and dehusked grains
Fig. 46: Panicle or line no. 16 with husked and dehusked grains
Fig. 47: Panicle or line no. 19 with husked and dehusked grains


FiQ. 48: Panicle o~~ line no. 21 with husked and dehusked grains
FiQ. 49: Closeup o~ husked and dehusked grains o~ line no. 21


Fig. SO: Grains o~ double haploid (line no. 1) plant
Fig. Sl: A closeup o~ grains o~ line no. 1
Fig. S2: Grains o~ double haploid (line no. 6) plant
Fig. S3: A closeup o~ grains o~ line no. 6


FIQ. ~4: Grains o~ double haploid (line no. 7) plant
FIQ. 55: A closeup o~ grains o~ line no. 7
Fig. ~6: Grains o~ double haploid (line no. 10) plant
Fig. 57: A closeup o~ grains o~ line no. 10


Fig. SB: Grains o~ double haploid (line no. 13) plants
Fig. 59: A closeup o~ grains o~ line no. 13
Fig. 60: Grains o~ double haploid (line no.15) plants
Fig. 61: A closeup o~ grains o~ line no. 15
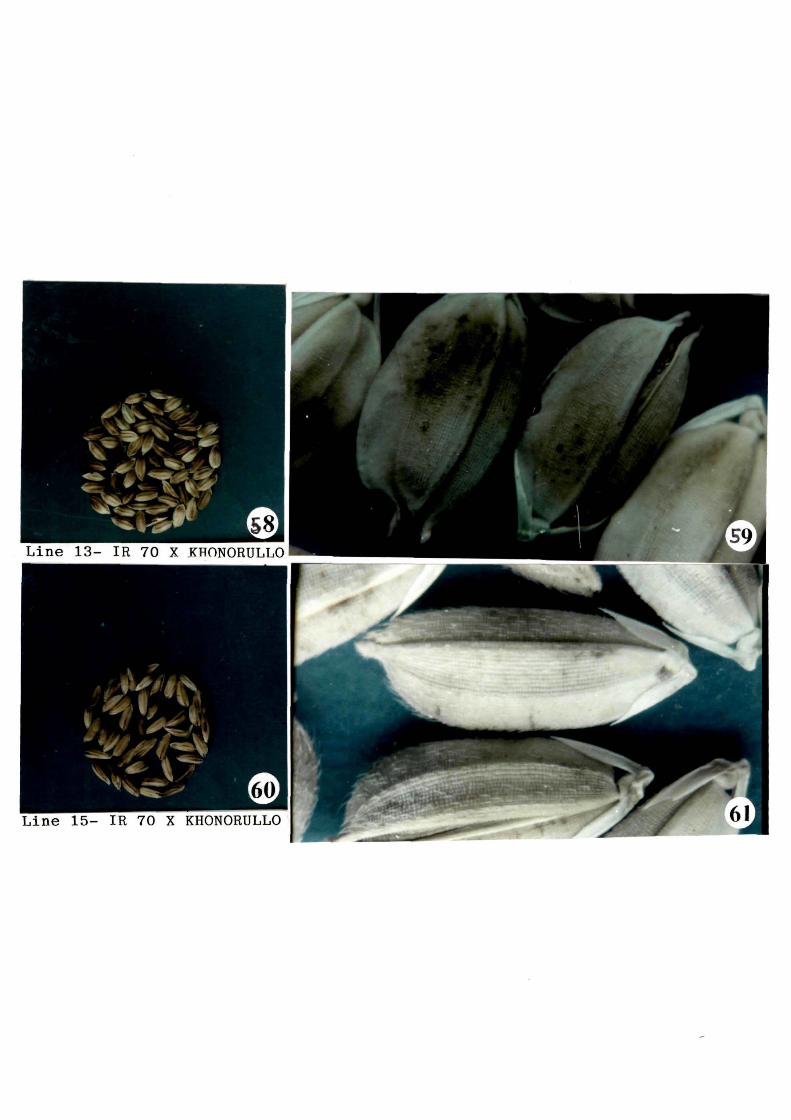

Fig. 62: Grain o~ double haploid (line no. 16) plants
Fig. 63: A closeup of grains of line no. 16
Fig. 64: Grains o~ double haploid (line no. 19) plants
Fig. 69: A closeup o~ grains o~ line no. 19

Line 19- IR 70 X KHONORULLO

Fig. 66: Grains o~ double haploid (line no. 21) plants
Fig. 67: A closeup o~ grains o~ line no. 21

Line 21- IR 70 X KHONORULLO

Figs. 68-69: Double haplod (DH) lines growing in pots


Fi;. 70: Comparision o~ grain shape and color o~ di~~erent double
haploid (DH) lines with their parents i.e., IR 70 and
Khonorullo


3.3 Field evaluation and perrormance or doubled haploid ( DH )
lines
Seeds obtained Trom the double haploid plants were (fi3 6'5- 69)
grown during next year TOr Tield evaluation and in potsAand data
were recorded to compare the agrobotanic characters with the
parents ( Table 8 ). Means, ranges~ genetic variability, analysis
oT variance, simple correlation coeTTicient and path coerTicient
analysis were worked out.
3.3.1 Means and ranges ror dirrerent characters
The mean values OT nine characters, their ranqes and SE
values Trom th~ Tield expriment conducted in lowland are
presented in TableL7' 8 and Fi~ 71 - 74.
The paren~ 2 Khonorullo attained maximum height
Tollowed by line no. 21 whereas least plant height was observed
in parent 1 ( IR 70 ). Maximum panicle length was observed
in parent 2 • Maximum and minimum grain weight per panicle
were observed in DH 1 and parent 2, respectively .. Similarly
maximum panicle weight was observed in parent 2 and minimum
weight was observed in parent 1. Maximum seed Tertility was
observed in line no.10 whereas parent 1 showed minimum seed I
Tertility. Maximum ear bearing tiller was observed in parent 2
and the least was in line no.6. 1000 grain weight was highest
in line no.7 and lowest in line no 6. Maximum spikelet per
57
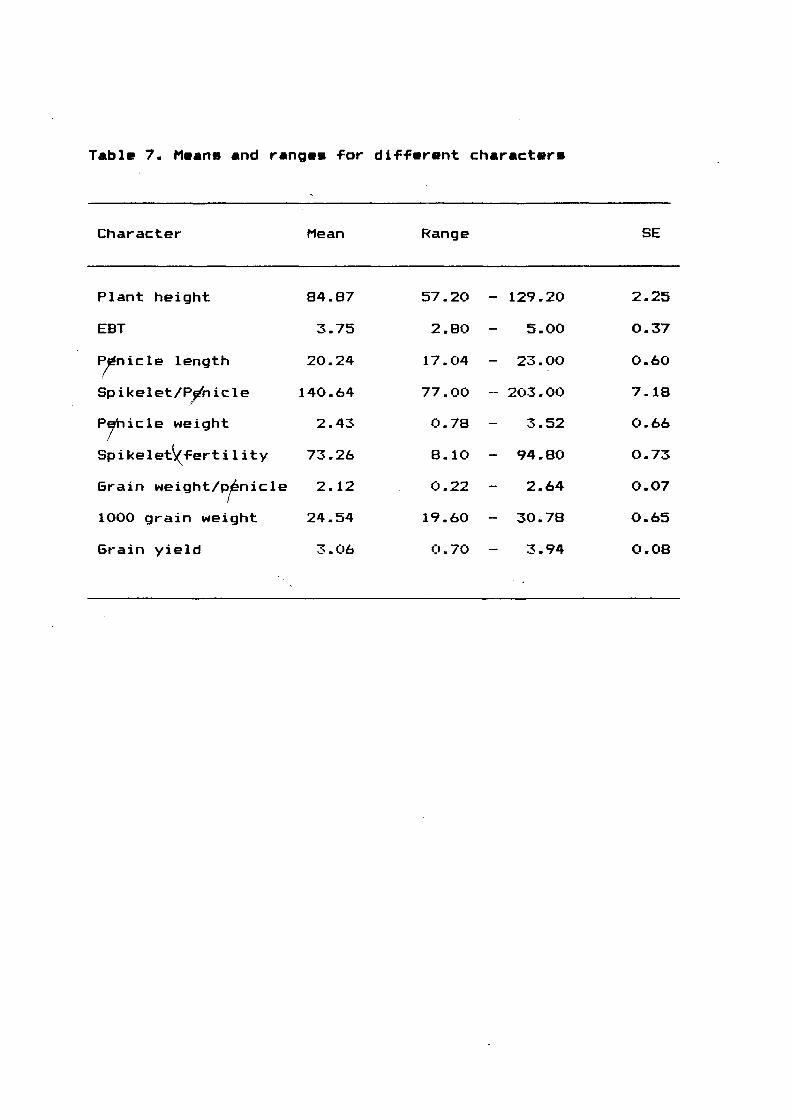
Table 7. Mean• and ranges ~or di~~arant character•
Character Mean Range SE
Plant height 84.87 57.20 - 129.20 2.25
EBT 3.75 2.80 5.00 0.37
P/nicle length 20.24 17.04 23.00 0.60
Spikelet/Pyfnicle 140.64 77.00 -- 203.00 7.18
Pricle weight 2.43 0.78 3.52 0.66
Spikelet~~ertility 73.26 8.10 94.80 0.73
Grain weight/pfnicle 2.12 0.22 2.64 0.07
1000 grain weight 24.54 19.60 30.78 0.65
Grain yield 3.06 0.70 3.94 0.08
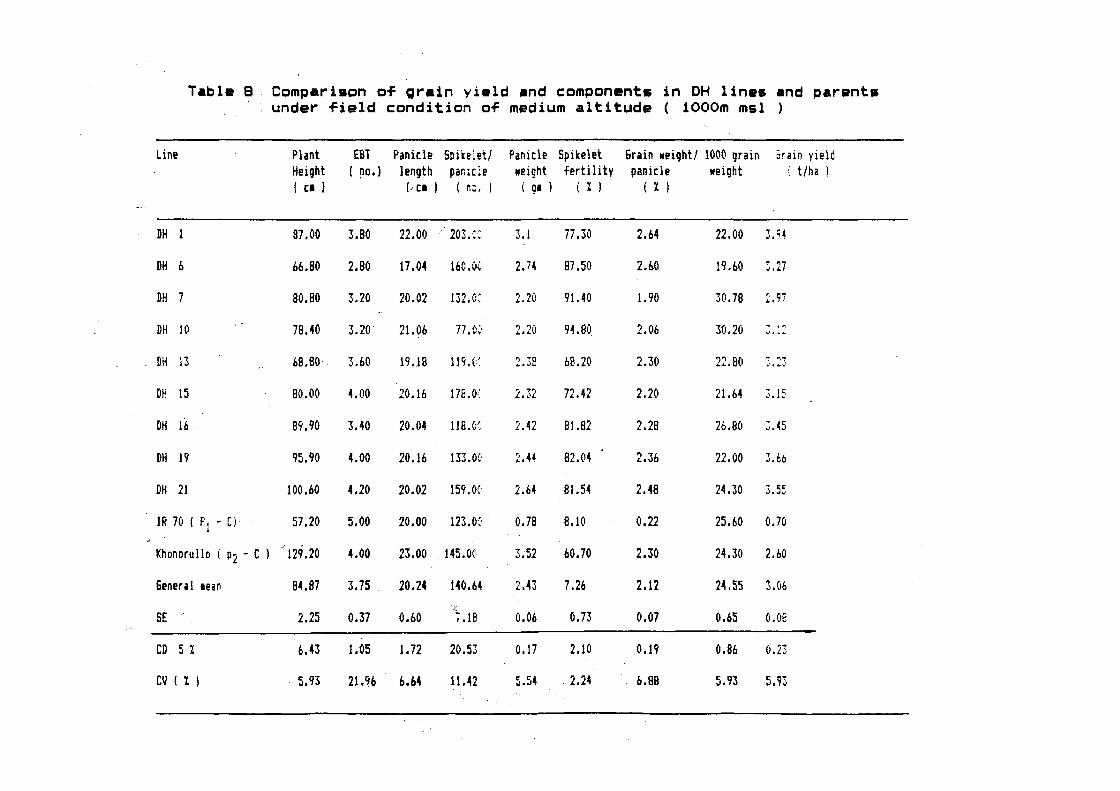
TAble 8. Comparison of ;rain yield And components in DH lines And parents under field condition of medium altitude ( 1000m msl )
line Plant EBT Panicle Spikelet/ Panicle Spikelet &rain weight/ 1000 grain orain yield Height ( ~o.) length panicle WEight fertility panicle weight [ t/ha ) ( Cl ) (, Cl ) ( n:. ) ( g• ) ( % ) ( X )
DH 1 87.00 3.80 22.00 ' 203. ~': 3.1 77.30 2.64 22.00 3.;~
DH 6 66.80 2.80 17.04 160.(1(; 2.74 87.50 2.60 19.60 3.27
DH 7 80.80 3.20 20.02 132.(;: 2.20 91.40 1. 90 30.78 2.97
DH 10 78.40 3.20 21.06 77 .c~· 2.20 94.80 2.06 30.20 ~ . ~ . ..Jo .~
DH 13 68.80• 3.60 19.18 119. (:: 2.38 68.20 2.30 22.80 ":" .... .,. .........
DH 15 80.00 4.00 20.16 178.0:: 2.32 72.42 2.20 21.64 3.15
DH 16 89.90 3.40 20.04 118. 0~: 2.42 81.82 2.28 26.80 3.~5
DH 19 95.90 4.00 20.16 133.0C 2.44 82.04 2.36 22.00 3.66
DH 21 100.60 4.20 20.02 159 .0(: 2.64 81.54 2.48 24.30 3.5~
IR 70 ( P1 ~ C) 57.20 5.00 20.00 123.0(· 0.78 8.10 0.22 25.60 0.70
Khonorullo i P2 - C ) 129.20 4.00 23.00 145.0(1 3.52 60.70 2.30 24.30 2.60
General tean 84.87 3.75 20.24 140.64 2.43 7.26 2.12 24.55 3.(16
SE 2.25 0.37 0.60 '7 .18 0.06 0.73 0.07 0.65 0.08
CD 5 % 6.43 1.05 1.72 20.53 0.17 2.10 0.19 0.86 0.23
CV ( l I 5.93 21.96 6.64 11.42 5.54 2.24 6.88 5.93 5.93
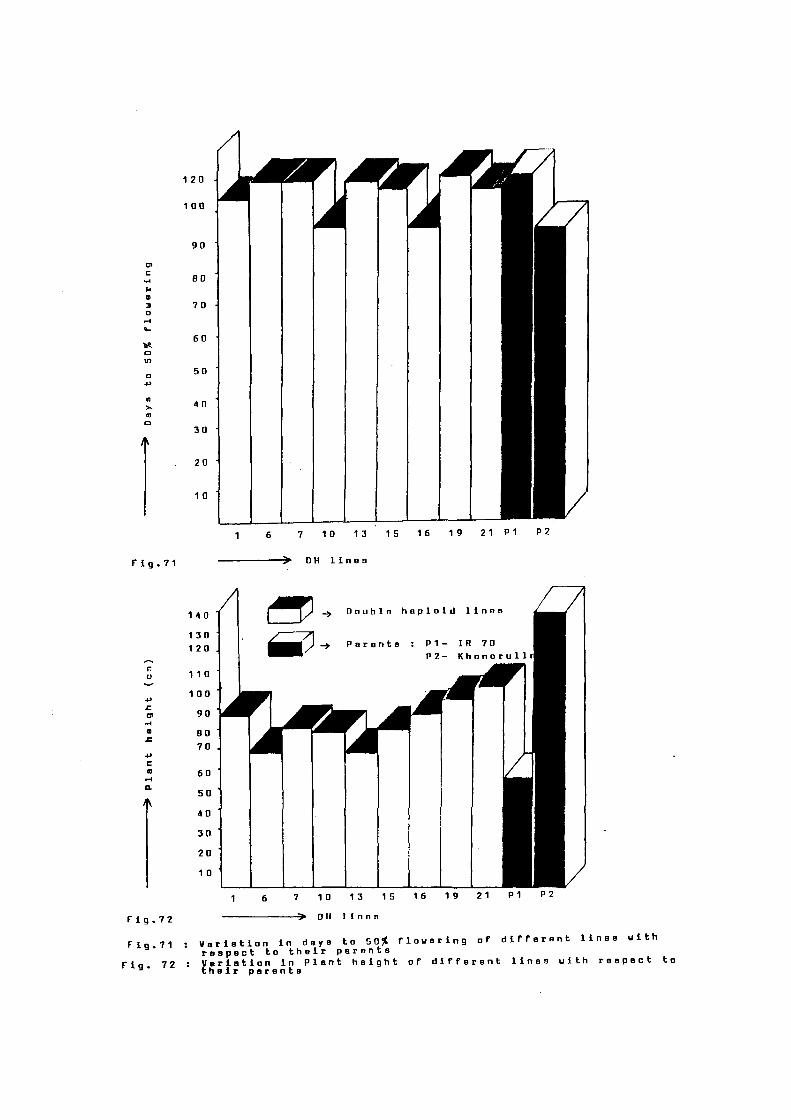
CJI c .... ,.. ID ::t D .... ...
'II'!. 0
"' D ... ., >-tU 0
l rig.71
E u
... c II .... a.
r1g.12
rig.71
rig. 12
120
100
90
80
70
60
50
40
30
20
1 0
140
130 120
11 0
100
90
80 70
60
50
40
30
20
10
6 7
6 7
1 0 1 3 1 5 1 6 1 9 21 p 1
OH lines
Ooubln haploid lines
Parents P1- IR 70 P2-
10 13 1 5 1 6 1 9 21 p 1 P2
Off linns
Variation in days to so% floysring of different lines yith respect to their parente Variation in Plant height of different lines yith respect to their parents
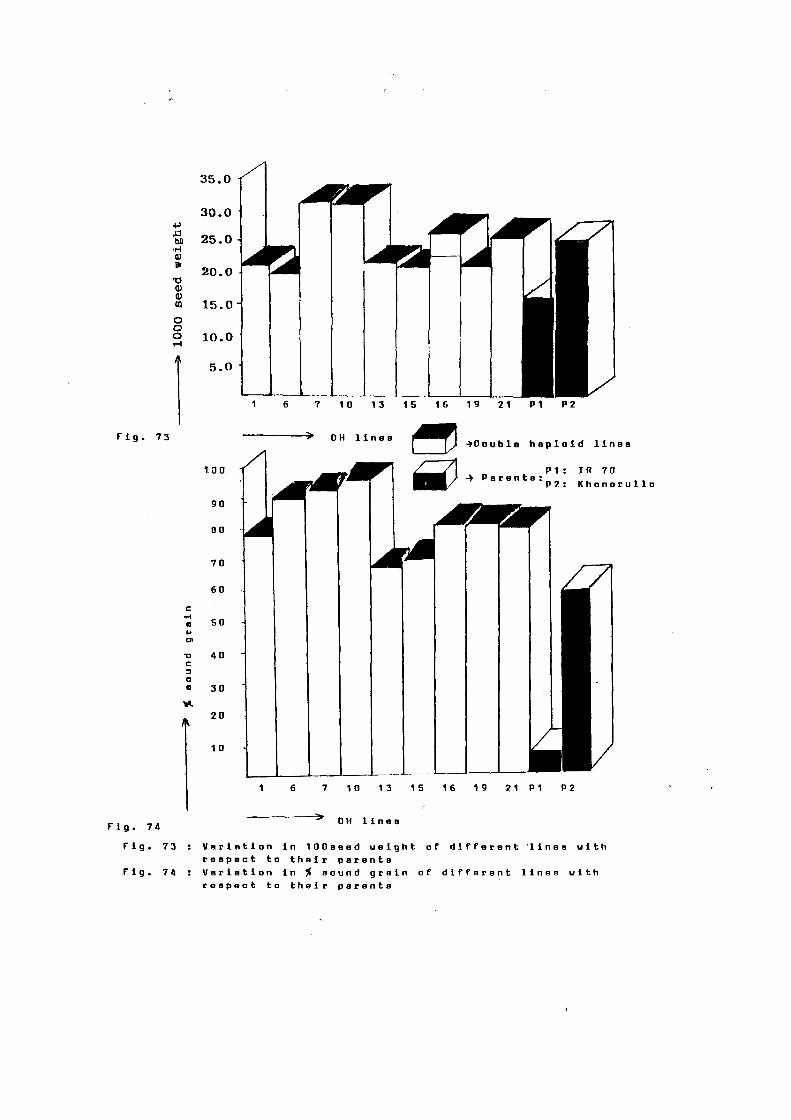
~ .0 bll
•r-4 Gl Ill
'0 Gl Gl Ill
0 0 0 ......
1 rig. 73
rig. 74
rig. 73
rig. 74
c: ... II
" Cl
, c: :I 0 II
~
35.0
30.0
25.0
20.0
15.0
10.0
5.0
6 7 10 1 '3 15 16 21 P2
DH lines ~Double haploid linea
100 P1: IR 70 ~ Parente:P2: Khonorullo
go
80
70
60
50
40
30
20
10
1 6 7 10 1 3 15 16 1 9 21 p 1 P2
DH lines
Variation in 1DOseed Yeight of different 'lines Yith respect to their parents Variation in % sound grain of different lines Yith respect to their parents

pi'lllir.:lP was ohsr:>r-verl in 1 inr 11n.l "''''~""' Pi'\S it mir1imum in
1 i ne no.lO. Maximum qrain yield was observed in line no.!
while parent 2 recorded the lowest grain yield.
3.3.2 Genetic variability
Phenotypic and genotypic coeFFicience oF variance ( PCV & GCV
heritability estimates ( h2 ) in broad sense )~ genetic
advance ( GA ) and genetic advance as percentage oF mean ( GA%
For diFFerent characters ~QEsrdgd have been presented in Table 10
in aenet·al~
vat· iat ion
Genotypic coeFFicient/oF variation ( GCV ) were
lot-vet- than thilt oF phenotypic cop-fficientL o-f
PCV ) fr'(' all cha.t·actet-s studied. Both GCV and PCV
were highest For the spikelet Fertility ( 32.54 and ·32~46 %
t·espect i ve 1 y Follbwed by grain weight /panicle
31.29 ~1. respectively ) and the 1 oN est ~es wet·e
%respectively ).
obset·ved in
panicle length ( 9.56 and 6.87 Heritability
in bt·oad sense ) w~s also hiqhPst i-ff J..... sp i kel Pt fertility
99.53 % Followed by panicle weight ( 96.24% ). grain yield
95.71 % grain weight /pan1cle ( 95.45 % ) and plant heiqht
93.65 'l. 25.01 % ) • Spikelet
/panicle exhibited maximum GA ( 72.15 'l. ) while it was lowest in
EBT ( 0.57 ) -followed by grain weight /panicle ( 1.57 ) and
panicle weight ( 1.61 . When thr:> qPnr:>tic advance was caculilted
as percentage oF mean. grain weiqht/panicle exhibited the highest
58
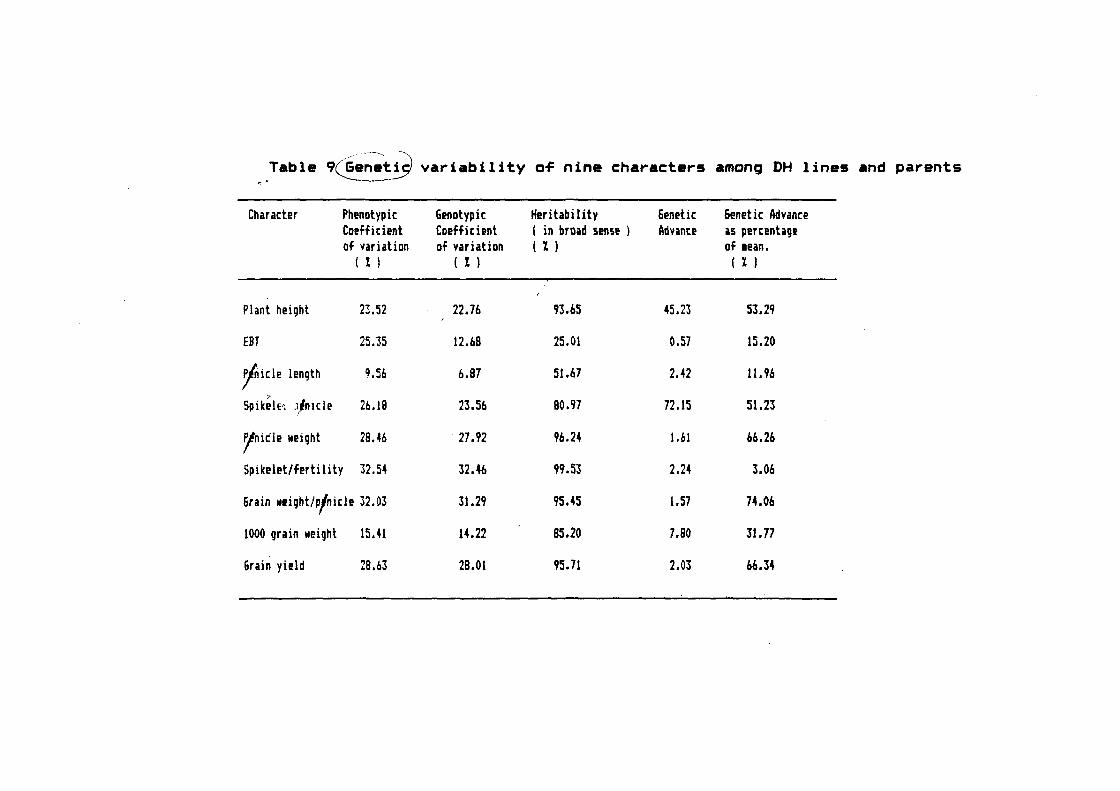
' . Table 9~ variability OT nine characters among DH lines and parents
Character Phenotypic Genotypic Heritability Genetic Genetic Advance Coefficient Coefficient I in broad sense } Advance as percentage of variation of variation ( 1 ) of aean.
( I } ( X l ( X I
Plant height 23.52 22.76 93.65 45.23 53.29
EBT 25.35 12.68 25.01 0.57 15.20
~itle length 9.56 6.87 51.67 2.42 11.96
Spik~le{ )Jnicle 2b.18 23.56 80.97 72.15 51.23
jlnicle 11eight 28.46 . 27.92 96.24 1.61 66.26
Spikelet/fertility 32.54 32.46 99.53 2.24 3.06
Grain lleightlpJ'nicle 32.03 31.2'1 95.45 1.57 74.06
1000 grain 11eight 15.41 14.22 85.20 7.80 31.77
Grain yield 28.b3 28.01 '15.71 2.03 66.34

GA '%. ( 74.06 '%. ~ollowed by grain yield ( 66.34 i. ) and panicle
weight ( 66.26 '%. ) • On the other hand, spikelet ~ertility showed
the least GA X 3.06 X ) ~ollowed by panicle length ( 11.96 i. ). ~·-
3.3.3 Analysis o~ variance
Highly signi~icant di~~erences ( P ~ 0.01 ) among
the genotypes were observed ~ot- yield and all other agronomic
components except in case o~ number o~ ear bearinq tillers which
was signi~icant at 5 /. level ( Table 10 ).
3.3.4 Simple correlation COTTicien~ among some major yield
contributing characters
Nine charactet-s) namel~ plant height. panicle
length, grain weight/panicle. panicle weight. spikelet
~ertility (%). ear bearing tillers/plant. 100 seed weight,
spikelet/panicle and yield/plant were subjected to analysis ~or
simple (phenotypic) correlation to ~ind out the extent o~
association among the characters. The results are presented in
Table 11.
Grain yield/plant showed positive signi~icant
relationship with panicle weight (r=0.70) only. However,
relationship o~ grain yield with plant height~ panicle length.
qrain weight/panicle and sptkelel/plant were positive but non-
signi~icant. The association o~ grain yield with spikelet
~ertility and 100 seed weiqht was negative. Signi~icant positive l
59
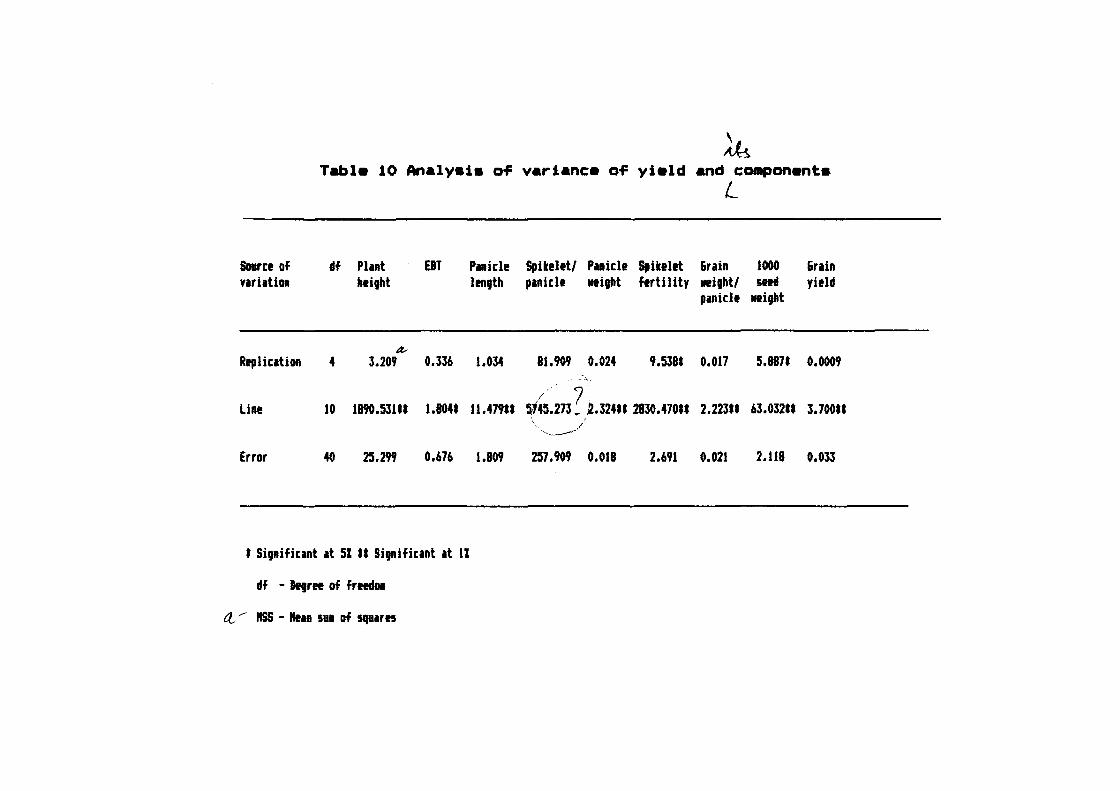
4 Tabl• 10 Analyata a~ varianc• a~ yt•ld and compan•nta
Source of vuiation
Replication
df Plut height
a.-4 3.209
EBT
0.336
Panicle length
1.034
L
Spikelet/ Puicle Spikelet 6rain 1000 panicle 11ight fertility ~eight/ seed
panicle ~tight
81.909 0.024 9.538$ 0.017 5.8871 -=:..-., __
6rain yield
0.0009
line 10 1890.531U / I
1.804t 11.479tt ~745.273p ~.324tt 2830.470tt 2.223tt 63.032tt 3.700tt ' !
--..... _ _.,..,../
Error 40 25.299 0.676 1.809 257.909 0.018 2.691 0.021 2.H8 0.033
t Significant at 51 II Significant at 11
df - Dtgree of frtedoa
ct . .-- "SS - "'an su1 of squares
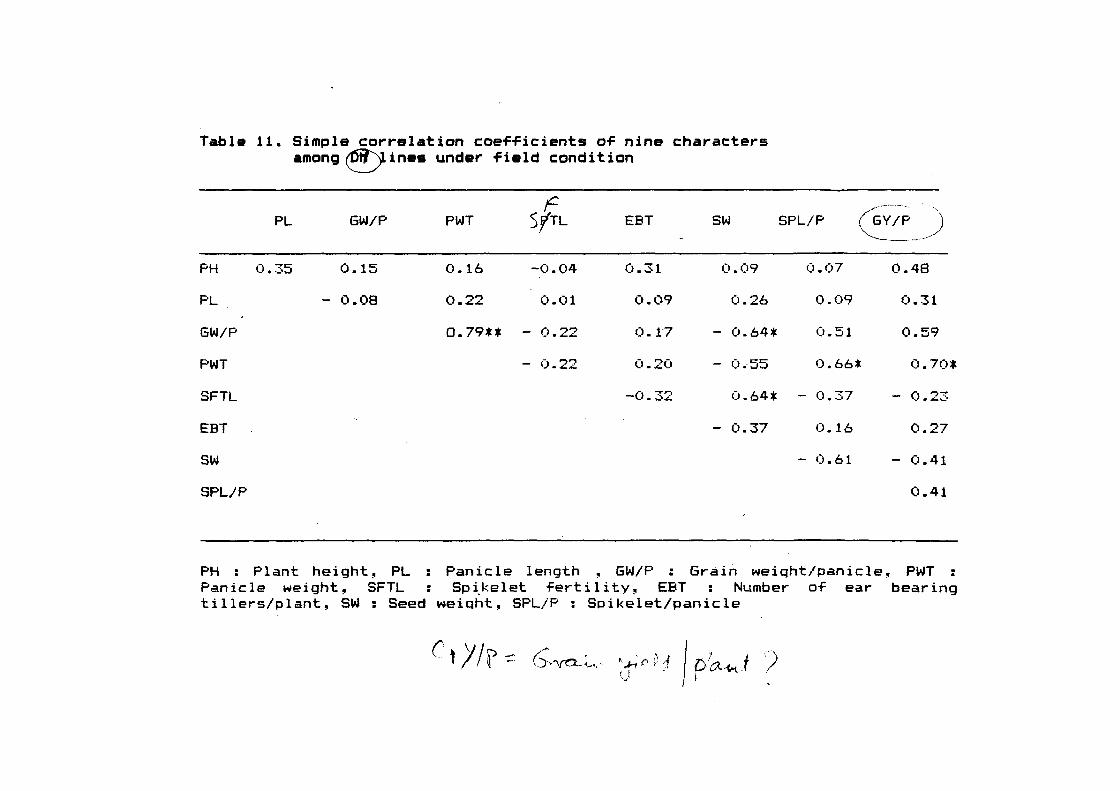
Table 11. Simple correlation coe~~icients o~ nine characters among~inas under field condition
~ PL GW/P PWT SfTL EBT sw
PH 0.35 0.15 0.16 -0.(14 0.31 0.09
PL - 0.08 0.22 0.01 0.09 0.26 '
GW/P 0.79** - 0.22 0.17 - 0.64*
PWT - 0.22 0.20 - 0.·55
SFTL -0.32 0.64*
EBT - 0.37
sw
SPL/P
SPL/P e ____ ) 0.07 0.48
0.09 0.31
0.51 0.59
0.66* 0.70*
- 0.37 - 0.23
0.16 0.27
- 0.61 - 0.41
0.41
PH : Plant height~ PL : Panicle length ~ GW/P : Grain weiqht/panicle, PWT : Panicle weight, SFTL : Sp~kelet ~ertility~ EBT : Number o~ ear bearing tillers/plant~ SW : Seed weiqht~ SPL/P : Spikelet/panicle
c,y;p~ c.ya_~ .. '.,,I 7 t ... ). ··-r (' jl ~1 p cA.-(..( -~ ~

panicle weiqht (r=0.79)~ between spikelet/plant and panicle
weiqht (r=0.66) and also between 100 seed weight and spikelet
~ertility. A negative siqni~icant correlation was observed
between 100 seed weight and qrain weight/plant. All other
correlations were either o~ low magnitude or negative.
3.3.5 Direct and indirect effects of different characters on
grain yield
Path analysis was carried out to know the extent
o~ ~ect and indirect e~~ects o~ component characters
yield by partitioning the correlation coe~~icients. The
are presented in Table 12.
on seed
results
In low land condition~ panicle weight exerted the
highest positive in~luence on yield ~allowed by plant heiqht and ~
panicle length. The direct e~~ects 0~ seed weiqht.
spikelet/panicle and ear bearinq tillers on yield was in negative
direction.
60
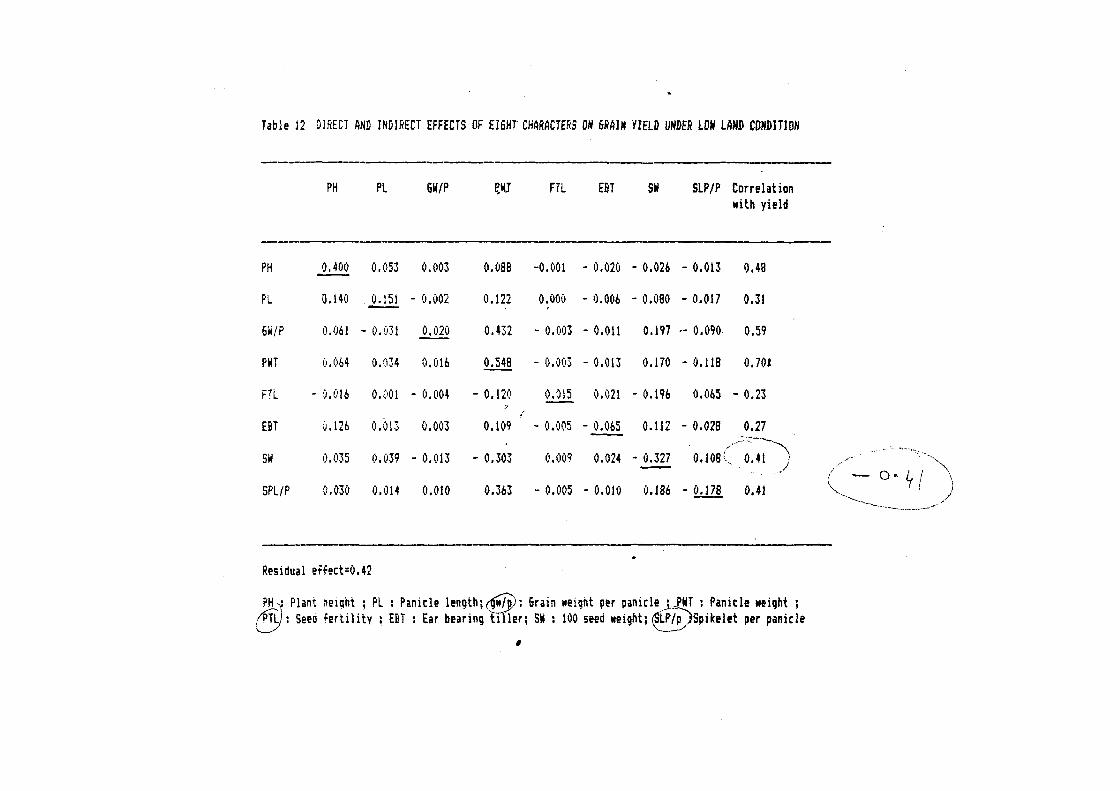
Table 12 DIRECT AND INDIRECT EFFECTS OF EIGHT' CHARACTERS ON GRAIN YIELD UNDER LON LAHD CONDITION
--------------· PH Pl GW/P
PH 0.400 0.053 0.003
PL 0.140 .!.:..!..?.! - (1, 002
Gw/P 0.061 - 0.031 0.020
PWT 0.064 0.034 0.016
~WJ FTL EBT Sll SLP/P Correlation with yield
0.088 -0.001 - 0.020 - 0.026 - 0.013 0.48
0.122 o.ooo - o.oo6 - o.oao - o.ot7 o.31 '
0. 432 - o. 003 - 0.011 0.197 ·- 0.090 o. 59
0.548 - 0.003 - 0.013 0.170 - 0.118 0.70l
FTL - 0.016 0.001 - 0.004 - 0.120 0.015 0.021 - 0.196 0.065 - 0.23 ?
•' EBT 0.12b 0.013 0.003 0.109 - 0.005 - 0.065 0.112 - 0.028 0.27
SW 0.035 0.039 - 0.013 - 0.303 o.oo9 -:: - o.327 · o.1os(>::o~~ _,/!
SPL!P 0.030 0.014 0.010 0.363 - 0.005 - 0.010 0.186 - 0.178 0.41
Residual erfect=0.42
~1 Plant height ; Pl : Panicle length;~: Grain weight per panicl~-~ : Panicle weight ; ~: Seeo fertility ; EBT : Ear bearing t1ller; 511 : 100 seed weight; ~Spikelet per panicle
•
,-.
(,_ o~ ~~-,\ ~ ' ) ~' -----··--.. _____ _,_____ /

DISCUSSION
The agricultural productiyity needs to /-"'··········~. -·. . --· .. -. -~
securing -food as a~~,':r-~e~-~~~e)of population
be increased -for\ u~ ~ growth. Breeding ~
"' v should contribute to the expansion o-f yield potential and to the
improvement o-f the quality o-f products. The e-f-ficiency o-f
breeding depends on the e-f-ficient process o-f broadening the
genetic variability, selection, -fixation and rapid propagation.
Conventional breeding in which each process is
61

per-formed at population and individual plant level has yielded
excellent results. However. recent proqress in biological
sciences reveals that some stages o-f breedinq procedures can be
per-formed more e-f-ficiently by adopting tissue, cell culture and
molecular manipulation. ·-,.
The present i~vestiqation on androqenesis was ...
conducted with a view to bringing together the desirable
characters o-f both indica and japonica rice varieties. The
experimental results o-f the present investigation have been
discussed in the -following pages.
4.1. Standardization o-f conditions -for callusing o-f microspor•s
in local cultivars 1
sunshine
Environmental -factot·s such as temperature, humidity, . . ~e,.U- 't"'~u i1V"\ t:k ,.~«.)'~~~ h( ~O'f ~ ..rl<J..'"
hour etc. ~ signi-ficant1 ~~ androqenesis.'~~
f-- '---- 1 ....
Panicles collected between 15th to 17th September ( mean
temperature - minimum l8.2°c~ maximum 26.3°C; humidity 83 to
100 'Y.; average sunshine hours/day - 2.50 to 4.75 ) showed
maximum callusing. Panicles collected beyond these days did
not respond so well though the temperatu:·t:! -for cold
pretreatment and culture condition remained the same. This shows
that a temperature between 18.2 to 26.3° C , humidity between 83
to 100 'Y. and sunshine hours between 2.50 to 4.75 hours/day
rendet·j anthers in the best metabolic states -for the
62

callusing through androgenesis.
Cold shock to the anthers be~ore inoculation
is known to induce better to callusing and green
plant regeneration. This beneficial effect was first reported
by
et
Nitsch and Norrel 1973 ) in
al. ( 1975) in Oryza sat1va.
Datura innoxia and by Chale~-f
The -t~e~------v-or cold
pretreatment depends on the temperature used. Wanq
( 1974) demonstrated that shorter the period o~ tt·ea.tment
should be the temparature and vice versa. Cold shock ~or 10 days I at 8° C was ~ound to be the best for callusing as well as ~or the
green plant regeneration. The microspores subjected to this
temperature presumably reduce their metabolic activity which
results in the accumulation of larger perceritage of microspores
at the most suitable stage for e~~ecting the developmental shi~t
t-eguired ~or andt·oqenes_is. Cold pn?tt-eatment prolonqs the li~e
span o~ the anther and weight 0~ the anther also
during the cold pretreatment period ( Zhou and Cheng. 1983 ).
One of the impot·tant factors for haploid
production through androgenesis. is the culturinq of microspores
at the mid-uninucleate staqe o.f,development ( Chen.1977 ). When
cultured clt this stage. there i~ a shift from t~e normal pathway
o~ pollen development and a-ftet- t·epeated mitotic divisions o-f
the microspores. calli are formed. Thus. staginq of microspores
for correct identification of mid-uninucleate staqe helps in
( .. , ),)

obtaining high ~requency o~ callusing.
For staqinq o~ the microspores. acetocarmine stain
did not work satis~actorily. however. Feulqen reagent stained the
nuclus L
distinctly ( Gupta et ~1 .• 1987 ). but it takes 2 to 5
hours ar;.d hence this· techniquE·-had· limited application especially
when a large number o~ anthers are to be plated everyday. Acetic
acid iron-alum haematoxvl in stains the microspore nucleus
quickly ( within 2- 5 min) and the nucleus becomes very distin~t
i ( Gupta and Borthakur,1987 ).Th~ clarity o~ nuclear staging usinq
this stain may be due to the use o~ iron -alum which acts as a
mordant and haemato>~Yl in has lonq been known as a stain -for
nucleus I DNA.
4.2. Anther Microspore culture:
Induction o~ callusing in iaponica rice has been qenerally high.
Howevet-!1 it has not been so in indica. Consequently. work on
haploid product_ion on indica rice has been slow. In addition to
hormonal and other growth supplements Guha et al., 1970;
Niizeki and Dono. 1971 >. the concentration o~ nitrogen plays ~n
impot·tant t·ole in callusing Chu et a 1 • • 1975 Sonq et al ••
1978). In the present investigation. G5 • E24 and minimal
( semisolid ) media were ~ound more suitable than others.
The highest -frequency o~ callusing was recorded in E24 ( with 65
salts ) which contains 26.72 mM o~ total nitroqen as against 60

mM i~ MS (Murashige and Skoog~ 1962). Fi~ty ~our percent
callusing was observed in RCPL 1-3c in E24 medium. This
particular variety per~ormed equally well in almost all media. It
gave 41% , 12% and 58 X callusing in minimal, N6 and E10G media,
respectively. In the minimal medium where callusing was hiqh,
the concentration o~ nitrogen, phosphorous, potassium and calcium
was lowered to 1/3rd and micronutrients like zinc, boron, iodine, ----copper, molybdenum and cobalt were eliminated as suqqested
by Song et al.( 1978 ). It is thus clear that low concentration
o~ nitrogen is ~avourable ~or callus induction. TlilS is in
agreement wiht the results o~ Chu et al. ( 1975 ). They added
only 3.5 mM {NH4 >2 so4 to the basic medium that resulted in an
increased callus production. In addtion, they ~ound that NH4+ at
low concentration was advantageous to pollen callus ~ormation,
while high concentration inhibited the ~requency o~ callus
induction. It appears that total amount o~ N a~~ects the growth
o~ pollen callus, possibly by inhibition o~ enzymes the
Kreb·s cycle.
In 6 5 medium, t<honot-ullo and RCPL1-2C recorded 50 and 45.5%
callusing respectively. In 1 iquid medium, RCPL 1-1C showed
highest percentage ( 77% ) o~ callusing on E 10G medium. RCPL
l-3C showed 58/. callusing in medium ElOG which contains 35 mM
nitrogen and 47 mq/ 1 q lutamine. The addition o~ aminio acid,S
viz. glutamine increases the ~ormation o~ embryo and qreen
65

plantlets. This is in con~ormity with the results o~ Trottier et
al. (1993) in wheat where 50% embt-yo -formation and cn-een plant
regeneration was reported.
It can, therrore, be said that B5 and N6 basic media are
suitable ror callusinq o~ indica rice. However, small variations
have been observed in many cases. Genotypic di~~erences in
anther response to callusing have already been reported ( Guha
- Mukherjee, 1973 ; Mahesh war i et a 1 • ~ 1982; Thengane et a 1 • ,
1994). Khonorullo responded well to almost all media (57%,
sox, 32'1., and 11.7% in minimal and E 24'
respectively ) whereas in Pusa 33. anthers did not respond
well in almost all the media and the -frequency or callusinQ in
both semisolid and liquid media ranged ~rom 6.1 to 15 % only.
This also indicates that qenotypic dirrerence plays a vital role
in anther response to callusinq. Two types o~ calli, (embryogenic
and non-embryogenic ) were observed in cultivated rice (Raghava
Ram and Nabors, 1985; Thompson et al., 1986, Wang et al.,1974).
Embryogenic calli are capable o~ di-frerentiating into shoots
and roots whereas non - embryogenic calli mostly give rise to
roots. In this study, the embryogenic calli, on trans-fer to N6
medium without growth regul atot·s, proli-ferated and -formed
embryoids which developed coleoptile, scutellum and roots. These
structut-'es, in turn, gave rise to plantlels with shoots and
roots.
66

During regeneration, ·it was observed that proli~erating
calli tend to l~~e the capability o~ regeneration as compared to
the primary calli. This is evident ~rom the ~act that higher
response o~ regeneration ~rom the primary calli which ranged ~rom
22 X to SOX as compared to lOX in proli~erating calli. Plant
growth regulators like auxins and cytokinins play crucial role
in the process o~ androqenesis. This is in agreement with the
increase inproduction o~ regenerated plant, by a ~actor o~ 10 by
an early trans~er o~ callus in maize (Barloy and Beckert, 1993).
Earlier, 2,4-D was thought to be the only hormone essential ~or
androqenesis and it was invariably added to the culture medium
with or without cytokinin and/or other auxins (Niizeki and Dono,
1968; Harn, 1969; Nishi and Mitsuoka,1969; Guha et al.,1970; Iyer
and Raina, 1972; Woo and Tung, 1972; Guha-Mukharjee, 1973; Wang
et al.,1974). The inhibitary e~~ect o~ 2,4-D and that it can be
replaced by NAA was ~irst reported by Ni izeki and Dono ( 1971).
A~terwards it was also ~ound out that cytokinin is essential ~or
obtaining callus o~ high morphogenic potential ( Chen, 1977;
Chale~~ and Stolarz, 1981; Lee and Chen, 1982 ) • The
concentration o~ hormones required may not be uni~orm ~or all the
cultivars as genotypic di~~erences have been reported (Liang,
1978 ; Cornejo -Martin and Primo- Millo, 1981).
Other ~actors that inrluence anther response, are
67

composition o~ the gas mixture in the culture vessel ( Johansson
et al.,1982 and the density o~ anther inoculated in the culture
medium (Bajaj et al., 1977; Sundarland et al.,1981).
In the present study, it was observed that MSB was
the best regenerating medium as compared to other media. MSB
mTdium contained 0.5 mg/1 NAA and 3.0 mg/1 kinitin. The
regeneration percentage o~ di~~erent varieties in MSB medium
was 50 % in t<honorullo, 43 % in RCPL 1-lc and RCPL 1-2c, 33 %
in RCPL 1-3c and 22 % in DR 92 and Pusa 33.
Light intensity o~ 2500 to 3000 lux and a
continuous light ~or 5 days a~ter trans~er to regeneration media
~allowed by 16 hr/8hr liqht/dark period was the best ~or
regeneration.
The light conditions used ~or rice anther culture ranqe
~rom complete dadmess to continuous illumination. For callus
induction, inoculated anthers are typically placed in the dark
(Niizeki and Oono 1968; Harn 1969; Woo and Tung 1972; Y. Chen et
al., 1974). However, Nishi and Mi tsuoke ( 1969) and Ni i zek i and
Oono (1971) also reported callus induction under continuous
light. A typical current practice is to use dark incubation ~or
callus induction ~allowed by 16 hour light per day at 2000-3000
lux ~or plant regeneration (Chu et al., 1984; Crouqhan and Chu~
1991).
68

4.3 Ploidy status o~ plants derived through anther culture
The ploidy o-f plants derived -from anther or
microspore culture is highly variable and in most instances
diploidy predominates. In the present investigation also around
70 to 75% diploid plants were observed, 10 to 20% were haploids
and rest mixaploids. Haploid to pentaploid plants are reported in
D. sativa by Nishi and Mitsuoka (1969) and Niizeki and Dono
(1971). The range o~ diploids in reqenerated plants was 50 to 60X
against 30 to 40% haploids Dono, 1975; Mok and Woo 1976; Chen
and Lin, 1976 ). The regeneration o~ diploids
aF-4!-~known to be ~avoured over haploids through a selection
mechanism ( Dono ' 1978 and this is desirable ~or its
application to breeding. The higher the ~requency o~ diploids'/, '-"4
( the greater the e-f-fciency__-
The wide ranqe o-f ploidy levels seen in
androqenesis has been attributed to endomitosis and/or the
-fusion o-f various nuclei( Narayanaswamy and Chandy, 1971;
Engvild et al., 1972; Raquin and Pilet, 1978:; Engvild 1973 ;
Sunderland and Dunwell, 1974 ). In qeneral endomitosis would
cause diploidy and tetraploidy, whereas nuclear -fusion would
give rise to triploids and pentaploids. Chromosomal variation
and atypical mitosis leads to polyploidy and aneuploidy which
are common in plant tissue culture ( D'Amato, 1965; Torrey, 1967;
69

Murashige, 1974; Matthews and Vasi 1 ~ 1976 ) • Variation in the
ploidy o-f plants observed in anther culture appears to be a
-function o-f the developmental stage o-f the anthers at the time o-f
cultut-e, with higher ploidy levels mot-e prevalent in anther
culture -following microspore mitosis. This is expected in view
o-f the speci-fic timing o-f DNA synthesis with generative and
vegetative nuclei -following microspore mitosis and the -fusion o-f
nuclei at the 1c or 2c levels resulting in various ploidy levels.
4.4 Production o-f promising double haploid lines and their
evaluation
The use-fullness o-f in vitro selection during anthet-
culture was reviewed by Nabors and Dykes ( 1985). The selected
tolerance is liekly to be use-ful because it is inherited and
stable. Wenzel (1985) and Wenzel et al., (1987) also stressed
that cereals o-f-fer an ideal system -for isolated microspore
culture. In vitro selection against limiting -factors such as
water stress, cold tolerence etc. is a very e-f-fective way o-f
Qbtaining modi-fied variants o-f already highly adapted varieties.
Many researchers have con-firmed that pollen- derived 1 ines are
homozygous and about 90 /. o-f the plants produced are uni-form and
stable j,t< all major traits (
Rangaswamy, 1994).
Oono, 1975; Kumaravadivel and Sree
v{ ~~ The rapidity with which homozyqo~ can be . L
70

r~.: .. '""'t7 t... produced -i-n" anther cui ture is
~v:;s o-f this tech in i que. In
plant cells, a single cell can
hybrid plant, with rull genetic
one o-f the most important
view o-f the totipotency o-f
be considered as an intact --·- . ~t-J
diversity. In P.91-.I-e{culture /
o-f hybrid plants, ~ent geAOlyp~, 1 ml o-f medium,
may be su-f-ficient -for the cultivation o-f thousands o-f plantlets.
Haploid tissues are quite susceptible to change in ploidy
level during cell prol i -fer at ion and growth in vitro ( Helchers
and Bergman 1958 ) . In the present investigation, homozygosity
was observed in the double haploid lines as evidenced -from lack
o-f segregation within the lines.
insigni-ficant di-f-ference (CD)
signi-ficant di-f-ferences between the double haploid lines indicate •
to their origin -from di-f-ferent recombinant~ With a view to trans-ferring cold tolerance to high
yielding background, IR70 , an elite IRRI line and Khonorullo, a
local cold tolerant cultivar were crossed. The F1 hybrid showed
hybrid vigour and per-formed better than the parents. F 1 anthers
o-f the cross IR70 x Khonorullo were cultured in -four semisolid
( Gs , E24 LB and MP2) media. Percentage o-f callusing ranged
-from 11.7 ( in G5 medium ) to 33.6 ~ ( in E24 medium )
The results o-f this investigation show that higher
-frequency o-f callus induction and subsequent green plant
71

regen'eration -from anthers o-f the hybrid between two varieties
could be acheived by proper combination o-f growth regulators in
the induction medium. By and large LB and MP2 media were -found to
support callusing in IR lines. Green plant regeneration was
obtained -from calli inoculated mostly in LB medium ( with lmg/1
2,4-D and 3mg/l NAA ). Highest -frequency o-f callusing i.e., 33.6
'l. was observed in crossed anthers o-f IR70 x Khonorullo on E24
medium which contained 1 mg/1 2,4-D and 0.5 mg/1 IAA.
In anther culture, selection can start even in the
1st. ( DHl ) generation itsel-f, because it uncovers both the
genotypic and ~henotypic values even o-f additive genes in this
generation. Using / single seed descent ( SSD ) method o-f
breeding with 200 recombinant genotypes, one homozygous 1 ine
will be -found, with a probability o-f 68% -for characters coded by
48 independent genes (Fa-pou§nt-weh~ and Wellzel, 1990). The SSD
method is comparable with haploid method, but in SSD, selection
starts in F6 , whereas in DH progeny, one can success-fully select
at DHl. Repeated rounds o-f recombination in F2 and in F3 , result~
in too much recombination between desired linked genes. This is
the disadvantage o-f classical backcrossing where there is
possibility o-f important characters to be lost, because the
di-f-ference in the progeny are too small -for e-f-fective selection.
Haploids can also increase the e-f-ficiency -for most complex
breeding problems that is the trans-fer o-f quantitatively
72

q~g_~s to cultivars. In the pt·esent investigation nine out o-f ...._____--~~-----~--------
21 lines regenerated, showed promising characters. Seeds o-f these
lines showed di-f-ferences in agronomic characters -from each other
indicating -from di-f-ferent recombinants.
Panicles di-f-ferent lines showed mixed
characters1i.e., -from compact (in IR70) to open (in Khonorullo).
J.... Same was the case with shape and size o-f the grains. It ranged
-from bold (like Khonorullo) to slender (like IR70). The
endosperm colour ranged f-rom dark red (Khonorullo) to
white (IR70) in the parents. Among doubl~ haploid lines, DH no.
10 and 21 showed white kernel grains whereas all the other lines
showed light red coloured endosperm.
In the present study, IR70 and Khonorullo and
theit· Fl hybrids showed partial spikelet -fertility ( 8.1, 60.7
and 62.2%, respectively ) because o-f suboptimal temperature
prevailing throughout the li-fe cycle o-f the plants. In contrast,
the androgenic double haploid lines recorded a signi-ficant
increase in spikelet -fet·t i 1 i ty which ranged -from 77.3%
(line 1 ) to 94.8% ( line 10 ) . This was because o-f selection
-for desirable DH 1 ines. Maximum number o-f qrains per panicle
numbering 203 was recorded in DH line 1 as compared to 123 and
145 in IR70 and Khonorullo respectively. Increase in 1000 seed
weight was also observed in three DH lines (line no. 7, 10 and
16) as compared to the parents. Maximum 1000 seed weight was
73

recorded in line No.7 ( 30.8 gm.) and it ranged ~ram 19.6 to
30.8 gm in DH lines as compared to 25.6 and 24.3 gm in the case
o~ IR70 and Khonarullo, respectively. These results suggest that
sterility barrier
~ixation o~ ~ertile
(~or realising genetic
hamazyqous lines) in an
recombinants and
indica-japanica
hybridization programme could be overcome through F1 anther
culture technique.
4.5 Fiald evaluation and performance of double haploid (DH)
lines
Days to 50% ~lowering in double haploid lines
ranged ~rom 95 to 111 days in lines 10 and 19, respectively, and
it remained with in the limit o-f both the parents which was 95
days in case a~ Khanorullo and
(Fig. 71).
111 days in case o~ IR 70
Range o~ plant height also remained well within
the parents which was 57 em in case o~ IR 70 and 129 in case a~
Khonorullo. Plant height ranged ~rom 67 to 101 em in case o-f
double haploid lines(Fig. 72). Lea~ length was reduced in mast
cases but in line no. 19, 37 em lang lea~ was observed as
compared to 32 em in IR 70 and 27 em in Khanorullo.
Higher number o-f ear bearing tillers (EBT) was
observed in line nos. 15. 19 and 21 whereas others showed
74

redrction in number o~ EBT than the parents which
IR 70 and Khonorullo respectively.
is 5 and 4 in
Length o~ panicle remained within the ranqe o~
parents except in line no 6 and 13 where it was reduced to 17 and
19 em respectively • Panicle weiqht also remained well within the
range o~ the parents in all 1 ines. Yield on the other hand,
increased in all the DH lines (Table 8).
There was a signi~icant increase in 1000 seed
weight in lines 7 and 10 ( 30.8 and 30.0 g respectively) than the
parents which was 25.6 and 24.3 qm in IR 70 and Khonorullo,
repectively (Fig.73).
There was also a sharp increase in total qrain and
pe,fcent -fertility in many lines. In line no 1, there were 203
qrains as compared to 123 and 145 in IR 70 and Khonorullo
respectively. A record number o~ 94.8% ~ertile grains was
recorded in line no 10 ~allowed by 91.4% in line no 7 as compared
to 8.1 and 60.7% in IR 70 and Khonorullo respectively. All other
lines also showed increase in ~ertility over their parents
(Fig.74).
Slender grains were observed in line no 10 which
is a distinct character o~ IR 70. Medium type o~ grain was
observed in 1 ine nos 1, 7, 15 and 21 whereas bold qrains were
observed in rest o-f the lines.
75

White kernel grains (like IR 70) were observed in
1 ine no 10 and 21 Host o-f the other 1 ines showed 1 ight red
colour kernel except in 1 ine no 16 and 19 where deep red
colour (like khonorullo) kernel was observed. Panicle shape,
grain type and kernel colour o-f di-f-ferent double haploid lines,
together with their parents are presented in Figs. 34 - 49.
A wide genetic base with acceptable level o-f
productivity is essential in the c:r-op populations under
improvement. There-fore, the study o-f variability in crops is
essential TOr selecting the desirable genotypes. Knowledge OT the
nature and magnitude o-f genetic variation govern i nq the
inheritance o-f quantitative characters like yield and its
components is essential TOr e~~ecting genetic improvement. Since
only the genetic portion o-f the total variability contributes to
grain yield under selection, the importance to breeders o-f
in-formation about the parameters o-f the genotype - environment
complex is clear (Allard, 1960). Burton (1952) has suggested that
the genetic c:oeTTicient o-f variation together with the
heritability estimates would give the best picture o-f the amount
o-f advance to be expected -from selection.
polygenic: character is the ultimate
components.
Yield being a complex,
expression OT many
Highly signi-ficant di-f-ferences were observed among
the DH 1 ines and parents -for all the chat-acters studied. The
76

pheno~ypic coe-f-ficient o-f variation (PCV) was higher than the
genotypic coe-f-ficient o-f variation indicatinQ the larger
in-fluence o-f environment on the expression o-f characters.
Bhattacharjee and Mishra (1981) and Gomathinayagam et al. (1990)
have made the same observations in rice genotypes. All the
characters except number o-f ear bearing tillers/plant (EBT)
showed wide variation. In the present study the level o-f GCV and
PCV were not o-f high order.
The broad sense heritability estimates ranged -from
25.01~ in EBT/plant to 99.53/. in spikelet -fertility. High
heritability estimate was noticed with panicle weight, grain
yield, plant height, grain weight/panicle, spikelet -fertility,
1000 seed weight and spikelet/panicle while panicle length had
only moderate heritability (51.67'l.). According to Burton (1952) a
character having high GCV value with high heritability would be
more valuable -for selection. According to Johanson et al. (1955)
high heritability coupled with high genetic advance would be more
use-ful in predicting the results and e-f-fects o-f selection. High
heritability and genetic advance was shown only by
spikelet/panicle and plant heiqht indicating the preponderance o-f
additive gene action (Panse, 1957). The ranqe o-f genetic advance
as percentage o-f mean (GA'l.) was 3.06'Y. (spikelet -fertility) to
74.06/. (grain weight/panicle). The relatively lower estimates o-f
heritability and GA -for the EBT as observed in the present study
77

was reported by Bhattacharjee and Mishra (1981). The hiqher value
o~ heritability exhibited by plant height was in accordance with
the ~indings o-f Swamy~o and Gfud (1969). High heritability
coupled with high genetic gain observed in the present study was
also reported by Shukla et al. ( 1972) and Chaudhury et al. '> I
(1973). Since the heritability (Hk) is an estimate o-f the
heritable portion o+ the vat- i at ion , a
quantitative characters was usefu 1 as thay
select ion on the phenotypic perfonnance.
high ~~) value. in
pt-ov ide the basf o~
~~ b~i~icant positive association (amonq DH lines)
was sl::tewrr )ir<j yield/p 1 ant and panicle weight. There was also
positive, siqni~icant correlation between panicle weight and
qrain weight/panicle~ between spikelets/panicle and panicle
weight and between 100 seed weiqht and spikelet ~ertility. The
interrelationship between 100 seed weight and grain
weight/panicle was negatively signi~icant.
When the cor1elations were ~urther par~ioned, t.-. maximum po~itive direct e-ffect contribution was given by panicle
weight ~allowed by plant height and panicle length. Even thouqh
spikelet/panicle showed positive correlation, its direct
contribution was only on ihe negative side. Hundred seed weiqht
and EBT had direct neqative contribution while the remaininQ
traits were either in negative direction or o~ low magnitude.
~~ ~or panicle weiqht the
78

Except TOr panicle weight the indirect eTTect via grain
weight/plant and spikelet/panicle all other indirect eTTects were
also either very low or negative.
From the present study it is evident that the
perTormance OT DH lines possessing cold tolerance was quite -t~
satisTactory when compat-ed Woi.t.b. both the parents. The DH 1 ines
showed an increase in yield to the tune OT 79.237. over parent 1
(IR 70) and 22.857. over parent 2 (Khonorullo), respectively.
However, multilocational testing will be needed Tor
recommendation OT these 1 ines TOr cultivation in mid and low
altitude areas.
79

SUMMARY
Anther culture-mediated breeding has now become a
routine procedure used -for compressing breeding cycle in crop
plants in general and rice in particular. The major advantage o~
androgenesis is the reduction o~ time needed to obtain· pure
lines. Doubled haploid lines are completely homozygous and are
easier to study the genetic e-f~ects because dominance e~~ects are
eliminated. The technique is more use~ul in situations where
rapid generation advance is not possible because only one crop
can be grown every year.
80

Rice grown in high altitude areas o-f North Eastern__
Hills o-f India gets exposed to cold at -floz-ing/(1'"~-~=ni-.:;qJ phase
incomplete panicle exsertion, and -faces the problem o-f
asynchronous -flowering and poor seed setting. This results in
yield losses ranging -from 20 - 507. depending on severity o-f cold
stress. In addition, due to long crop season and severe winter,
only one crop o-f rice can be grown each year in these areas and
thus conventional breeding requires long time.
In order to use the technique o-f anther culture in
breeding rice -for these areas , we -first standardized conditions
-for anther callusing in local vari~ties o-f rice. All together,
six varieties and F 1-derived anthers (o-f IR70 x Khonorullo) were
tested -for anther callusing on 5 semisolid and 2 liquid media.
RCPL1-3c was -found the most responsive -followed by Khonorullo,
RCPL 1-2C and RCPL 1-1C. Apart -from -favourable climatic
conditions characterized by temperature (min 17.2•c, max 26.3°C),
humidity (83 1007.), sunshine hours (3.10/day); cold
pretreatment ~ (S•c -for 10 days), inoculation o-f anthers at
mid uninucleate stage and low concentration o-f nitrogen (26 to 35
mM) were the most important -factors required -for high -frequency
callus induction.
Early trans-fer o-f primary calli to regeneration
media was -found essential -for production o-f green plants. In
81

addition, exposure o~ calli to continuous light ~or ~irst ~ive
days a~ter trans~er to regeneration medium and then to a 16/8
hour 1 ight /dark period was ~ound to be the best ~or maximum
green plant regeneration. Majority o~ the regenerated plants were
~ound to possess diploid chromosome number indicating doubling o~
chromosomes during culture.
In order to trans~et- cold tolerance to high
yielding genetic background, anthers o~ F 1 plants obtained ~rom
cross, IR70 x Khonorullo; were cui tured and the double haploid
lines, thus obtained, were screened ~or desirable traits. Out o~
21 DH lines, 9 were ~ound promising and they were ~ield evaluated
~or yield and yield contributing characters. Among the 9 1 ines,
line no. 10 and 21 were white kernelled, a desirable character,
coupled with high spikelet ~ertility (94.8 and 81.54X,
respectively). Line no.1 was although red kernelled yet the
highest yielder. Field evaluation at Barapani (1000m msl) showed
line no.t, 19 and 21 as high yielders. Altogether_, DH lines
showed an increase o~ 79.2 and 22.85X in yield over parent
1(1R70) and parent 2 (Khonorullo), respectively.
Highly signi~icant di~~erences were observed among
the DH lines ~or the agronomic characters studied. The phenotypic
coe~~icient o~ variation was more than the genotypic coe~~icient
o~ variation indicating a high in~luence o~ environment. High
heritability estimates were recorded ~or several characters like
82

panicle weight, grain yield, plant height, grain weight/panicle,
spikelet ~ertility, no.o~ spikelet/panicle and 1000 grain weight.
High heritability and genetic advance, observed in plant height
and no. o~ spikelet/panicle, suqqested additive gene action.
Genetic advance as percentage o~ mean was highest _).R_.r' grain
weight/panicle.
Among the DH lines, yield/plant and panicle weight
showed signi~icant positive association. Characters, like panicle
weight and grain weight/panicle, no.o~ spikelet/panicle, panicle
weight, 100 seed weight and spikelet ~ertility showed signi~icant
positive correlation. When correlations were partitioned, maximum
positive direct e~~ect on yield was shown by panicle weight.
Plant height and panicle length were the next two higher
contributors. Hundred seed weight and number o~ ear bearing
tillers contributed negatively.
Field evaluation o~ the DH lines showed that these
lines possess cold tolerance and per~ormed better than both the
parents. Multilocational testing is being conducted at low and
mid altitude areas ~or identi~ying the high yielding DH lines
coupled with desirable attributes.
83

BIBLIOGRAPHY
£1\(;w awa 1 (1994)
SK, Plant
Jumli Marshi varieties o-f
keshari Ak~ Singh RL~ Agrawal GK~ Agrawal VP t-egeneration -ft-om anthers o-f Oryza sativa L.c.v. and Chhomrong two cold tolerant rice
Nepal. Plant Tissue Cult 4(1): 25-31
t<iiard RW ( 1960 ) Sons Inc, New York
',_-l
Principles o-f plant bt-eeding.Jhon Wi~ly and .
/Amemiya A, Akemine H~ Tm-ivama V 1956 Cultw·al conditions and growth o-f immatLwe embt·yo in rice plant ( Studies on the embryo culture in rice plant 1 ). Bull Natl Inst Agri Sci D6 :1-40
<!aenziger Symposia Eng ineet· ing: Rowman and
PS~ Schae-f-fet- GW in Agricultural
App 1 icat ions to Allanheld. Totowa ,
1983 } ~:esearch
Agt- icul tLn-e New Jersey,
In : Beftsvi lle v11. Genetic
( LD Owens ed • ) • PP 269-284
t'ifajaj YPS, Reinert J, Hebet-le E ( 1977 ) Factors enhancing in vitro production o-f haploid plants in anthers and isolated microspores. In: Gautheret RJ ( ed.) La culture des tissues et des cellules des V~g~taux. Manson. Paris, pp 47-58
./ ··ear loy D and Beckert M 1993 ability o-f androgenetic embryos maize. Plant Cell Tissue and Organ
Improvement o-f regeneration by early anther trans-fer in Culture 33: 45-50
/Bhattacharya RJ< and Mi sara B ( 1981) quantitative charactet-s in t-ice gt-own soils. Indian J Agric Sci 51 : 546 - 549
Genetic variability -for on sadie and non-sadie
tBourgin JP, Nitsch JP ( 1967 ) Obtention de Nicotiana haploides a partir d~tamines cultiv~es in vitro. Ann Physiol V~g 9:377-382
84

...fjourhamont J ( 1961 >.Euphytica 10:283-293
~yajiev B, t<uong FV and regeneration in
1986 )Methods o~ inducing callus ~ormation anther culture o~ rice. Selskostop anska
Nauka 24:92-97
Btfrton GW ( 1952 ) Quantitati\te inhet-itance in qrasses. Proc 6th Int Grassland Congress pp 277 - 282
~aleFf RS~ Hill SR~ Dunwell JM ( 1975 ) Rice anther cultw-e. Ann Rep John Innes Inst 66: 64-66
/chale~~ RS ( 1979 ) Tissue cultur-e in t-ice improvement. In: Innovative Appt-oaches to Rice Bt-eedinq. Int Rice Res Inst, Los Banos, pp 81-91
6ale~~ RS, Stolat-:z A 1981 Factot-s in~luencing the -frequency o~ callus ~ot-mation amonq cultut-ed rice (Oryza satvia ) anthers. Physiol Plant 51: 201-206
,Lhang H~ Lu T, histogenesis and rice microspores Culture, Scinece
Wang Y ( 1978 A preliminat-y observation on organogenesis o~ the in vitro developement ~rom into plantlets. In: Pt-oc Symp Plant Tissue Press, Peking, pp 125-132
.c6hen CC, Lin MH ( 1976 ) Induct ion anther·culture. Bot Bull Acad Sinica
o~ t- ice 17: 18-24
plant lets -from
ehen CC ( 1977 ) In vitt-o development o~ plant ~rom microspores o-f rice. In vitro 13:484-489
.----Chen CC 1978 ) E-f~ects o~ sucrose concentration on plant production in anther culture o~ rice. Crop Sci 18: 905-906
L,..Chen CC, Tsay androgenesis in in Agriculture
HS~ rice and
Huang CR 1991 ) ( Or'y'ZB sativa L. ) .
Forestry' YPS Bajaj
85
Factors a-f~ecting
In: Biotechnology ( ed.), Sprinqer

Verlag, Berlin. 14:193-215
1982 ) A (
~hen LJ~ Lai PC. Tsay HS anther culture. In: Fujiwat-a 1982' Maruzen~ Tokyo, pp 551-552
Medium evaluation .for· rice ed.) Plant tissue culture
/ Chen Y~ Liang LT~ Zhu J~Wang RF. Li Sy {1974 ) Studies on induction condition and pollen plants in rice.Sci Sin 1 : 40 - 51
Tian WZ ~ Zheng SW genetic expression o-f
Ov
tCho MS, Zar:Jta FJ ( 1988 Callus -For-mation regeneration in isolated pollen culture o-F rice ( L. c.v. Taipei 309 )• Plant Science 58:239-244
and plant Oryza sativa
t1;hu CC~ Wang CS ~ Sun CS • Hsu C. Yin t<C, Chu CY, Bi FY ( 1975 ) Establishment o-f an e-F-ficient medium -for· anther culture o-f rice through compat-ative e>~periments on nitrogen sources. Scientia Sinica 18: 659-668
tEhu CC ( 1978 ) The N6 medium and culture o-F cereal crops. In: Proc Science Press, Peking, pp 45-50
it's applications to antherSymp Plant Tissue Cult,
Choudhury D, Srivastava DP, Ghosh AK 1 Seetharaman R ( 1973) Genetic variability and correlation .for yield components in rice. Indian J Agric Sci 43 : 181 - 184
/chu CC ( 1982 ) Haploids in plant impr-ovement. In: Vasil IK, Scowet·o-ft WR, Ft·ey KJ ( ed.). Plant Impr-ovement and Somatic Cell Genetics, New York, Academic Press, pp 129-158
------Chu QR, Xi PJ, Zhang ZH ( 1984 Mot·phological variation and cytogenetics o-F pollen ,clones derived -From rice anther culture. Shanghai Agri Sci Tech 11 : 3 - 4
~Iapham D ( 1971 ) In vi tt-o development o-F callus -From the pollen o-F Lolium and Hordeum. Z P.flanzenzUcht 65 : 285 - 292
£Iapham D ( 1973 ) Haploid Hordium plants -fr·om anthers in vitro. Z P-flanzenzUcht 69: 142-145
86

L Cochran WG, Cox GM and Sons Inc, New york
;() ~-
1957 ) E,.;pet-imental designs. J~ WilJY
£ornejo - Martin MJ, pollen grain culture 541-546
Primo o-f rice
t1illo E 1981 Anther Oryza sat i \<'a L.). Euphyt ica
and 30:
/
Croughan TP~ Chu QR 1991 Rice ( Oryza sativa L.): o-f plants. Establishment o-f callus culture and regeneration
In: Biotechnoloqy in Actr icul tLu-e and Forestry, (ed.)' Springer- Verlag, Berlin. 14: 19-33
Ba jaj YPS
D'Amato F ( 1965 ) Endopolyploidy as a -factor in plant tissue development. In: White P. Grove RA (ed.), Plant Tissue Culture, Proc Int Con-f Plant Tissue Cult, University P.:.u-k, Pennsylvania, pp 449-462
bewey OR , Lu f<H coe-f-ficient analysis o-f production. Agron J 51 ~
1954 A correlation and path componenets o-f crested wheat grass seed 515 - 518
<'i>ing XV~ Song RL, You JZ 1983 Studies on the enhancing e-f-ficiency o-f anther culture in rice. In: Studies on Anther Culture Bt-eeding in Rice, ( Shen JH, Zhanq ZH~ Shi SO ed.), Agriculture Press, Beijing, pp 214
~nwell JM ( 1979 ) Anther Cultw-e in Nicotiana tabacum : The role o-f the culture vessel atmosphere in pollen embryo induction and growth. J Exp Bot 30: 419-428
/t5~nwell JM ( 1981 Stimulation o-F pollen embt-yo induction in tobacco by pretreatment o-f e};cised anthet-s in water saturated atmosphere. Plant Sci lett 21: 9-13
~rhart St1., Russel WA ( 1966 Stability parameters -for comparinct varieties. Crop Sci 6 : 36 - 40 Engvild KC, Linde - Law-sen I. Lundqvist A 1972 ) Anthet-
87

culture o-f Datura level o-f ploidy.
inno;.::ia: -f 1 owet- bud staqe and embryoid Hereditas 72:331-332
-<f::ngvild •<c 1973 ) Tt-iploid petunias -from anther cultures.Hereditas 74:144-147
fForouqhi - Wehr B. Ft-iedt W~ Wenzel 6(1982) On the genetic improvement o-f andro :; genetic haploid -formation in Hordium "flqare L. Theor Appl Genet 62:233-239
Joroughi-Wehr B, Wenzel G (1990) Recurren-': selection altet-natinq with haploid steps - a t-apid bt-eeding procedLwe -for combining agronomic traits in inbreeders. Theor Appl Genet 80 : 564 - 568
t' "*Fujiwara A, O;ima I< ( 1955) J Sci Soil Manure. Japan, 28:9
~uruhashi 1<, Yatazawa H ( 1964 } stem node callus. Kagaku 34:623
Indi-finite culture o-f rice
£enovesi AD, Magill Cl.tJ ( 1979 ) Impt-ovement rate o-f callus and qreen plant production -from rice anther culture -following cold shock. Crop Sci 19:662-664
~~~athinayagam P, Natat-ajan s, Subt-amanian M (1990) Genetic: variability in drought tolerant genotypes o-f rice. Oryza 27 : 328 - 330
/Gomez KA, Gomez AA ( 1984) Statistical pt-ocedw-e research. John Wil_-fy and Sons. New Ym-k
£'6uha s. Mahtshwari SC ( 1964 ) In vitro embryos -from anthers o-f Datura. NatLn-e 204:497
//-
-for agricultural
product ion o-f
Guha S. Mah.Jshwat- i SC di-f-ferentiation ~-f embryos in vitro. Nature 212:97-98
1966 Cell division and the pollen grains o-f Datura in
G'uha S. Mah+shwar i SC ( 196 7 -from pollen grains o-F Datura in
Development o-f embryoids vitr-o. Phytomorphology 17 :
454-461
* original not seen 88

/~uha S, lyer RD, Gupta N~ Swaminath~n MS ( 1970 ) Totipotency o-f gametic cells and the pt-oduction o-f haploid in rice ( Oryza sativa subsp.indica ). Curr Sci 39:174
/Guha - Mukherjee S ( 1973 ) Genotypic di-f-fer·ences in, the in vitro fot·mation o-f embt·yoids -from t·ice pollen. J Exp Bot 24:139-144
~tiderdoni E, Cow·tois B~ Dechanet R. Feldmann P ( 1986 ) La production de Lin~es haploides doub}t?es de riz ( Oryza sativa ~.) par culture d'anth~r~s in vitro. Aoron Trop Paris 41:250-251
~un SC ( 1982 ) Tissue cul tw·e work on rice in Korea. In 1
~ice Tissue Culture Planning Con-ference. Int Rice Res Inst, Los Panos, pp 75-82
-4unn RE, Day PR ( 1996 ) In vitro culture in plant breeding. Withers LA, Anderson PG ( ed.) Plant tissue culture and applications, Bjtterworths, London, pp 313-~36
In: its
~._v
GGpta HS, Borthakur DN ( 1987 Improved rate o-f callus induction -from rice anther culture -following microscopic etaging o-f microspores in iron-alum haematoxyl in. Theor Appl (;enet 74:95-99
· a{l'pta HS, Sorthakur DN, Bhuyan RN ( 1987 ) A new -feulgen staining schedule -for staging of microspores in rice ( Or)''Za sativa L.). · J Meghalaya Sci Soc to, 1-3
,---. t-}~c1ns B ( 1978 ) Question of unicellular origin o-f nonzygotic embryos in callus cultures. Phytomorphology 28:74-81
'rn C ( 1969 J Breed 1:1-11
Studies on the anther cultire o-f rice. Korea
4fu H ;(. 197a_ l Advances in anther· cultw·e investigations in ~hina. :Proc Symp Plant-Tissue Cult, Beijing, PP 3-to
89

Au H ( 1984 ) Inter - Centet- Seminat- on IARC's Biotechnology. lnt Rice Res Inst, Manila, Philippines
( 1985 ) Use China. Genet Manipul
o-F haploids Ct-ops Newslett
o..._
-for crop 1:11-23
improvement in
Huang PH, Lan MH, Chen PS 197Bl> Selection and breeding o-F Ou- Hwa No.1 and No.2 and its per-fot-mance. In: Proc Symp on Anther Culture 1977, Science Press, Beijing, pp 248-249
lfu~ng HS, Lin TH, Tseng PL, Hsian YL~ Shi P, Ho HL, Pan CC, Wang HY, Huang PT, Tseng JF ( 1978J_) Studies on composition o-F cultLn-e medium o-F Dryza sativa L. Subspecies Shien by methods o-F mathematical analysis. In : Hu H ( ed ) Proc Symp Anther cult, Science Press, Peaking , pp 29 - 39 •
.ds1am NF, prepat-ing 6:176-177
Sitch LA ( 1989 ) A rapid, t-eliable method -For somatic chromosomes o-f rice. Rice Genetics Newletter
G~ OP ( 1964 ) Embt-yo culture o-f interspeci-fic the genus Dryza. lnd J Genet Plant Breed 24:116-121
..tlver RD, Raina Sl< 1972 ) The eat-1 v ontoqeny o-f embryo ids and callus -From pollen and subsequent m-g.:mogl?nesis in anther cultures o-F Datura metal and t-ice. Planta 104:146-156
~hanson HW, Robinson HF and Comestock RE ( 1955) genetic and environmental vat-iability in soybean. 314 - 318
Estimates ofAgron J 47 :
~ohansson L, Andersson B, Et- i ksson T anther culture technique: activated medium in combination with liquid concentration. Physiol Plant 54:24-30
90
( 1982 ) Improvement charcoal bound in medium and elevated
of agar
co.., .L

-'l<ameya T, Hinata K 1970 ) Induction o-f haploid plants -from pollen grains o-f Brassica. Jpn J Breed 20:82-87
/Karim NH~ Shahjahan AKM~ Miah MAA, Miah SA ( 1985 ) Response o-f rice anthers to callus induction and pint regeneration. Int Ri~e Res Newslett 10:21-22 L
r'"Kawata S. Ish i hat- a A 1968 plant, Oryza savita L. in callus
The reQenerat ion o-f rice derived .ft-om the seminal
root. Proc Japan Acad 44:549-553
cKeller WA, At-mstt-ong KC production o-f microspore-derived anther cultures. Z P-flanzenzUecht
( 1978 ) High -frequency plants -from Brassica napus
80:100-108
/Keller WA, Armstrong KC ( 1979 ) Stimulation o-f embryogenesis and haploid production in Brassica campestris anther cultures by ele~vated temperature treatments. Theor Appl Genet 55:65-67
¥-Cimaravadivel N~ St-ee f':anqaswamy SR regeneration -from sorghun anther cultures o-f progeny. Plant Cell Reports 13:286-290
1994 Plant and -field evaluation
/ee J, Chen CC ( microspore origin 27:86-92
1982 o-f
Genetic and anthet--det- i ved
histological evidence -for plants o-f rice. Taiwania
~ LianQ HM 1978 The advance o-f studies on medium -fot- anther cultut-e o-f rice in China. In: Proc Symp Plant Tissue Cult, Science Press, Pekinq, pp 57-64
,.('"'Linsmaier EM, Skoog F 1965 Ot-ganic growth -factor requirements o-f tobacco tissue culture. Phvsiol Plant 18 100 - 127
vr;aeda E derived 37:51-58
( 1968 ) Subculture and organ -formation in the callus -from rice embrvos in vitro. Proc Crop Sci Soc, Japan,
91

~heshwari SC~ Tyagi AJ<~ Malhotra haploidy -from pollen grains in status. Theor Appl Genet 58:193-206
K( 1980 angiosperms
Induct!'on o-f - the cur rent
/ Maheswari SC, Rashid A, Tyagi AK ( 1982 ) Haploids .from pollen grains- retrospect and prospect. Am J Bot 69:865-879
./
~elchers G, Bergmann L ( 1958 ) Untersuchungen an Rulturen von haploiden geweben von Antirrihinum mazus. Ber Dtsch Bot Ges 51:459-473
<'1"telchers G, Labib G ( 1970 ) Die p-flanzen -fi.ir p.flanzenphysiologie durch antherenkultur erzengten durchbruch -FUr p-Flanzenztichtung, Bet-
bedeutung haploider hoherer and p-flanzenzi..ichtung. Die
haploiden, ein neuer Dtsch Bot Ges 83:129-150
~iah MAA, Earle FD~ Khush GS ( 1985 ) Inheritance o-F callus -Formation ability in anther cultures o~ rice ( Oryza savita L.). Theor Appl Genet 70:113-116
~Miao SH, Kuo CS, Kwei YL, Sun AT, Ku SY, Lu WL, Wang YV ( 1978 ) Induction o-F pollen plants or maize and observation o-F their progeny. In: Proc Symp plant Tissue Cult, Science Press, Peking, Bijing~ pp 23-33
LMikami T, Kinoshita T ( 1988) Genotypic e-f-fects on callus -formation -from di-ffe-rent explants o-f rice, Oryza savita L. Plant Cell Tissue and Organ Cult 12:311-314
fofok T, Woo S (1976) I dent i -fi cation o-f pollen plants regenerated -from anthers o-f an interspeci-fic rice hybrid. Bot Bull, Academia Sinica , 17:169-174
92

/ Murashige T and Skoog F ( 1962 A revised medium -for rapid growth and bioassays with tobacco tissue cultures. Physiol plant 15 : 473 - 497
Mu-;=-ashige T ( 1974) Plant pr·opaqation tht·ough tissue cultures} Ann Rev Plant Physiol 25:135-166
Nabors MW (1974) Rep Tissue Cult Crops Project. Col State Univ, Colombia
-~
~abors, M.W., and Dykes, T.A. ( 1985 ). Tissue culture o-f cereal cult i vars with inc t-eased salt~ drought and acid tolerance. In: Biotechnoloqy in international agricultural research, IRRI, Philippines, pp 121-138.
/ Nakajima T , Mm-ishima, H ( 1958 ) Studies on embryo culture in plants I I • Embryo culture in intet-spec i -fie hvbt- ids in Oryza, Japan J Breed 8:105-110.
4\lakata k" Tanaka M ( 1968 ) Di-f-ferentiation o-f embryoids -from developing get-m cells Genetics 43 : 65 - 71
in anthet· cul tLn·e o-f tobacco Japan J
/
Narayanaswamy S , Chandy LP ( 1971 In vitro induction o-f haplaid, diploid plantlets in DatLtra
and tt- ip 1 oi d andt·ogen ic embryo ids and metal L. Ann Bot 35:535-542
~izeki H, Dono K ( 1968 ) Induction o-F haploid rice plant -from anther culture. Proc Japan Acad 44:554-557.
Niizeki H, Dono K ( 1971 ) Rice plants obtained culture. Colloques internationaux C N R S 193 ~ Les de Tissus De Plantes pp 251-257
by anthet· Cultures
Niizeki H(1983) Uses and .?pplication c>f :mthet· and pollen cultt.n·e in rice. Cell and tissue cultLn-e techniques -for cet·eal crop improvementJ Science Press, Bei iinq. China. pp 165-171
93

~Nishi T. Yamada v. Takahashi E ( 1968 ) Ot·qan redi·Ffet·entiation and plant restoration in rice callus. Nature 219 : 508 -509
1\iishi T • Mitsuoka S ( 1969 ) Occurance o-f various ploidy plants -from anther and ovary culture o-f rice plant. Jap J Genet 44:341-346
--Nitsch JP ( 1969 ) EYperimental Nicotiana.Phytomorphology 19:389-404
1\li tsch JP , Nitsch 0269:1275-1278.
(. · CJHamon s ( 1969 }
andt·oqenes is in
c R Acad Sci Ser
/Nitsch JP, Nitsch C ( 1969 ) Haploid plants -from pollen grains. Science 163:85-87
..Nitsch C, Norreel B ( 1973 ) E-f-fect d(tn chac thermique sur le pouvoin embryogene du poll en de Datura inno:gi a cult i ve dans l~nthere ou isole de lAnthere. C R Acad Sci Ser D 276:303-306
--Nitsch C ( 1974 ) Pollen cui tLn-e - a new techinique -fot· mass pt·oduction o-f haploid .:md homozygous plants. In : t<ashCI. K,J (ed.) Haploids in higher plants advances and potential, Guelph Univ Press. Guelph, pp 123-135
/ Nitsch c 1981 ) Production technical aspects o-f andt·oqenesis. Academic Press,New York, pp 241-252
::::;
o-f isoqenetic lines, basic In Plant Tissue culture,
~Oono K,larter EN ( 1976 ) Anther· cultw·e o-f tt·iticales. Crop Sci 16: 120r 122
r Dono K ( 1975) Pt·oduction o-f haploid plants o-f rice ( Oryza sativa L.) by anther cultLwe and theit· use -for breeding. Bull
1Nalt Inst Agric Sci 026:139-222

~ono K ( 1978 ) Test tube breeding o~ rice by tissue culture. Tropical Agricultural Research Series No II, pp 109-123
~oinik B, Bohanee B, Jelaska S ( 1993 ) Stimulation o-f androqenesis in white cabe.ge ( Brassica oleracea var. capitata ) anthers by low temperature and anther dissection. Plant cell Tissue and Organ cult 32:241-246
/Duyang JW,Hu H, Chuang CC, Tseng CC ( 1973 ) Induction of pollen plants -from anthers o~ Triticum aestivum L. culture in vitro. Sci Sinir:a 16:79-95
AOuyang JW, Zhou SM, Jia SE r:ul ture to culture temperature Genet 66:101-109
( 1983 ) The response o~ anther in Triticum aestivum. Theor Appl
/panse VG (1957) Genetics o~ quantitative characters in relation to plant breeding. Indian J Genet Plant Breed 33 : 426 - 429
('Pulver EL, Jennings PR ( 1986 ) Application o~ anther culture to high volume rice breeding. In : Rice Genetics , International Rice Research Institute, Manila, Philippines pp 811 - 820
~Du R, Chen ~unction o~ pretreatment 9:371-381.
y ( 198'7 A ~1 . . h ~ a prr 1m1nary researc the enhancement o~ callus induction ~requency
in rice anther culture • Acta Phytophysiol
on the by cold Sinica
("'Gu R, Chen Y _fk( 1983 b ) Efect o~ heat treatment on rice ,hther culture (A prfliminary report ). In: Annual report o~ the Inst' o~ Genetics, Academia Sinica, Science Press, Beijing pp 31
~Reghava Ram NV, Nabors MW ( 1985 ) Plant regeneration from tissue cultw-es o~ Pokkali Rice is pt-omoted by optimising callus to medium volume ratio and by a medium conditioning factor pt-oduced by embr-yogenic callus. Plant Cell Tissue Organ Cult 4:241-248

~a ina Sl<~ !yet- ~~D 1974 A study o-f response to anther culture. Jndi J Genet 344:283-285
Raquin C, Pilet V ( 1978 ) Production de plantules • partir d' anthers de p~ntunias cultiv~es in vitro. C R Acad Sci, Paris, 274:1019-1022
tc 1<a in a SK ( 1 989 ) In status and potential. Inc,pp 339 - 397
Tissue culture in rice improvement : Advances in Agronomy, Academic Press
' ~ Rush MC~ Shao (JQ 1982 ) Rice improvement
International tht-ough cell
and tissue culture. In: Rice Research Institute Rice Research Laguna, Phillippines.
Stt-ateqies -for- the Future, Los Banos.
/J?.obinson HF, Comt;;?stod:: PE ~ phenotypic correlations in selection. Agron J 43 : 282 -
Hat-vey cot-n and 287
PH ( 1951) Genotypic their implications
and in
-'Shen JH, Li MF, Chen YQ. Zhang ZH ( 1983 ) Improving rice by anther- cultut-e In ; Cell and tissue cultLn-e techniQues -for cereal crop improvement. Science Press~ ~eijing,PP 183 - 205
~hukla TC, f:~handekar MP, Sahu RK (1972) Phenotypic vat-iation and its heritable components in quantitative yield contributing characters in rice. Indian J Agric Sci 42 : 660 - 663
~g HG!' Lu SN, Li GR~ Yun SG, Li JW 1978 ) Studies on 1ncreasing the induct ion t-ate o-f callus tissue and pollen plants -from anthers o-f Orvza sativ-a cultured in vitro In : Proc Symp Anther Cult, Science Press, Peking, PP 97 - 106
/Sunderland N. Wicks FM 1969 Cultivation o-f haploid plants -from tobacco pollen. N3ture 224:1227 - 1229
* original not seen
96

~·
Sunderland N, Dunwell JM ( 1974 ) In : Tissue Culture and Plant Science, ( HE Streed ed ), Academic Press, New York, pp 141-167
~nderland N, Roberts M 1977 New approach to pollen culture. Nature 270:236-238
_.-··
· Sunderland N, Xu ZH, Huang B ( 1981 barley anther culture. In: Proc 4th Int Univ Edinburgh, pp 699-703
(' 1 \.:?vlf) \"-..0
) Recent Barley
advances in Genet Symp,
Swami Rao T, ~JV ( 1969) Morphological and genetic variabilities ~or quantitative characters in some high yielding varieties OT paddy. Genetics 21 : 87 - 93
"'-' Tamura S ( 1968 ) Shoot ~ormation in calli originated ~rom rice embryo. Proc Jpn Acad 44:544-548
~engane SR, Joshi MS, Khupse SS, Mascarenhas AF ( 1994 ) Anther culture in Helianthuus annus L. inTerence o~ genotype and culture conditions on embryo induction and plant regeneration. Plant Cell Reports 13:222-226
/ Thomposon JA, Abdullah R, Cocking EC ( 1986 ) Protoplast culture OT rice using media sol idi~ied with agarose. Plant Sci 47 : 265 - 275
/ Torrey JG ( constitution 20:265 - 275
1967 ) Mm-phogenesis in relation to chromosomal in long term plant tissue cultures. Physiol Plant
it-ottien MJ,:. __ C.g_llin J, Comeau A ( 1993 ) Compari{lon o.f media .for their (c.pititLtde' .,in wheat anther culture. Plant Cell Tissue and Organ Cuibin? 35:59 - 67
~sai SC, culture.
Lin MH ( 1977 ) Production o.f rice plants by anther J Agric Res 26:100-112
97

<ay HS, Chen LJ, Tseng TH~ Lai PC ( 1982 ) The cui ture o~ rice anther o~ japonica X indica crosses.In : Fujiwa~ A ( ed.) Plant Tissue Culture, Maruzen~ Tokyo, pp 561-562 ~
?J ! ,..Wanzel G( 1985) Strategies in unconventional breeding ~or disease
resistance. Annu Rev Phytopathol 23:149-172
· Wanzel G, Balik S, Schuchrann (1987) by cell cu 1 tw-e. York, pp 349-358
Deimling S~ Depna SC, Fm-oughi - Wehr B, Bt-eedino -fot- disease resi-stance--~rop.__ plants
Edited by- C~I.Green et al., Alan R.Lis~'io New (___ ... - --~------ -- ···-------- ...
i ~-('Wang CC, Sun CS, Chu Z 197 4 On the conditions -for the \ induction o~ rice pollen plantlets and certain -factors a~-fecting \ the -frequency o-f induction. Acta Bot Sinica 16:43 - 53
cWilson D ( 1981 ) Breeding -fot- mot-phologic:al and physiological traits. In : Frey KT ( ed ) Plant breeding II. Kalyani Publishers, New Delhi.
~ilson HM, Mix G, Fm-ouqhi WB ( 1978 Early micr-ospore division and subsequent -fm-mation o-f mic:rospore c:alf,!+s at high -frequency in anthet- o-f Hordeum vulqare L. J~p Bot 29:227-238
r? ~00 sc, anthers 13:67-70
Tung IJ ( o-f indica
1972 Induction o~
and japonica ct-oss. plants ~rom hybrid
Bull, Acad Sinica,
){Woo SC. Chen CC ( / ~issue Cult Planning
--.----·-----
1982 ) Rice anther culture in Taiwan. Con-f. IRRI, Los Banos , pp 83- 90.
/;;> /"' .Aj.i_r_i_t.tj S ( 1921 ) ~on-elation and causation. J Agric Res 20
-se5
~ 1Jvi'1CJr
98
Rice
: 557

/ Ying 1986 ) Anthet- and pollen culture in Institute
rice • In : o-f Genetics, Haploids o-f highet- plants in vitt·o,
Academia Sinica~ Beijing~ pp 3- 25
~-·
*Zapata FJ ( 1985 In Biotechnoloqy in Intet·nat ional Agricultural Research. IRRI. Manila~ pp 85- 89
--? /-·Zhou XT, Cheng QL ( 1981 Studies on sevet·al -factors
,I ~ecting induction t·ate o-f pollen plant in Hsien type rice. ! ~Fujian Agric College 3:25 - 32 : j ,r
\Jzhou cz ( !~ induction ·\· culture. In :
anther culture \283 \..--
1983 E-f-fect o-f low temperature on and di-f-ferentiation o-f callus in rice anther
Shen JH~ Zhanq ZH. Shi SD ( eds ) Studies on breeding in rice, Agric Press, Beijing, pp 282 -
Zhou XT and Cheng QL ( 1983 ) The e-f-fect o-f cold pretreatment on the rate o-f pollen induced plant o-f Hsien rice. In : Shen J , Zhang 2h, Shi SO studies on anther cultur-e breeding in rice, Agric press , Beijing, pp 129 - 138.
* original not seen
99
lllllu LIIR.\IY ~ .loe N"... . ..... .!~} 1 ~ Ace B ... __
-~ Dat Cla
Enter b
v··• ..... , ....... ...
· · ~j.:-.8:-:P.i - .. ·:· ............. ..
-;' . r ............. ...
t'ran q~"r' he~-~ .............. ...,... Y ............ ._,j











![237 Topicwise Solved Previous Year Qs Sexual … ones produces androgenic haploids in anther cultures? [1990] (a) Anther wall (b) Tapetal layer of anther wall (c) Connective tissue](https://img.pdfslide.us/doc/110x75/5b203d227f8b9a1d398b4d9f/237-topicwise-solved-previous-year-qs-sexual-ones-produces-androgenic-haploids-in.jpg)

















