Embed Size (px)
Citation preview

Anchoring damages to benthic organisms in a subtropical scubadive hotspot
Vinicius J. Giglio1,2 & Maria L. F. Ternes3 & Thiago C. Mendes2,4 &
Cesar A. M. M. Cordeiro2 & Carlos E. L. Ferreira2
Received: 15 December 2016 /Revised: 26 February 2017 /Accepted: 17 March 2017# Springer Science+Business Media Dordrecht 2017
Abstract The physical damages to benthic organisms causedby boat anchorages were assessed in the Arraial do CaboMarine Extractive Reserve (ACMER), Brazil. It is one of themost visited scuba diving sites along the southwesternAtlantic. Through underwater visual observations, we ana-lyzed if benthic organisms were damaged by anchors and/oranchor cabling at two dive sites. A total of 112 anchorageswere sampled. Damages to benthic organisms were ob-served 139 times, mainly affecting epilithic algal matrix,the zoanthid Palythoa caribaeorum, and the fire coralMillepora alcicornis. Damages caused by anchor cableswere significantly higher than those caused by anchorsat one site. A significant difference between benthic orga-nisms damaged was observed only for P. caribaeorum, causedby the anchor’s cable. We present evidence that, at currentvisitation levels, anchors are a relevant stressor to benthicorganisms at dive sites in ACMER.
Keywords Subtropical reefs . Scuba divingmanagement .
Recreational diving . Tourismmanagement .
Marine protected area
Introduction
Impacts of anchoring on benthic organisms have beenwidely studied throughout tropical reefs. Mechanical dam-ages have been verified mainly on seagrasses (Creed andAmado Filho 1999; Milazzo et al. 2004) and coral reefs(Davis 1977; Jameson et al. 2007), where anchoring acti-vity can break, fragment and overturn several benthic or-ganisms. Not only the anchor itself can be detrimental tothese organisms, but also associated cables and chains canincrease the damaged area by dragging across the substra-tum or winding around reef organisms and structures(Dinsdale and Harriott 2004). The loss of benthic orga-nisms due to mechanical damage by anchors causes a con-siderable reduction in habitat complexity, especially in sys-tems characterized by tridimensionality, such as coral reefs(Francour et al. 1999). This disturbance may also have anindirect impact on associated faunal assemblages. For in-stance, scleractinian corals are important habitat-formingorganisms on coral reefs, providing critical refuge for fishagainst predation (Messmer et al. 2011; Coni et al. 2013),and suitable habitats for small predators to ambush theirprey (Holbrook et al. 2003).
With the increasing pressure of anthropogenic activities,the physical damage to benthic organisms caused by anchor-ing activity and divers has become of great concern amongcoral reef managers. Scuba diving has become one of the mostpopular aquatic recreational activities worldwide (Garrod andGossling 2012). By the mid-1980s, the major damage fromrecreational diving on corals was related to anchoring(Harriott et al. 1997; Jameson et al. 1999). To mitigate anchor-ing impacts, a simple system of mooring buoys has beenwidely used and proved to be a successful management ap-proach (Jameson et al. 1999; Tratalos and Austin 2001).Alongside these mooring buoys, implementation and
* Vinicius J. [email protected]
1 Programa de Pós-Graduação em Ecologia, Universidade Federal doRio de Janeiro, Rio de Janeiro, RJ, Brazil
2 Reef Systems Ecology and Conservation Lab, Universidade FederalFluminense, Niterói, RJ, Brazil
3 Programa de Pós-Graduação em Zoologia, Universidade Estadual deSanta Cruz, Ilhéus, BA, Brazil
4 Departamento de Ecologia, Universidade Federal do Rio de Janeiro,Rio de Janeiro, RJ, Brazil
J Coast ConservDOI 10.1007/s11852-017-0507-7

enforcement of rigorous carrying capacity programs (Zhanget al. 2016), and responsible diving practices (Roche et al.2016), represent management strategies that could be easilycarried out at dive sites but are still inexistent in many desti-nations, especially in developing countries.
Arraial do Cabo is one of the most visited dive sites on theBrazilian southeastern coast being known as the country’sBdiving capital^. The area represents a transitional zone betweenthe tropical and subtropical provinces along the Brazilian coastand is characterized by a relatively high biodiversity in south-eastern Brazil (Valentin 2001; Ferreira et al. 2001; Rogers et al.2014). In 1997, a sustainable use marine protected area namedArraial do Cabo Marine Extractive Reserve (ACMER) wascreated, whereby fishing is allowed exclusively to the localtraditional community, and recreational activities, such as divingand fishing, are permitted under specific management norms.Currently, there are 13 dive companies operating within theboundaries of the ACMER which perform approximately24,480 dives per year (Giglio, unp. data).
Dive sites of ACMER do not have mooring buoys so div-ing boats cast their anchors to remain stationary during divingoperations. Themain bow anchor of diving boats is placed in thesand bottom and the secondary anchor, located at the boat stern,is placed in shallow areas of the rocky reefs. To avoid majordamages to the environment, managers established that second-ary anchors should be placed manually in reefs by the dive staff,avoiding corals or other benthic organisms. However, there havebeen no assessments on the effectiveness of this approach toavoid anchoring damages to the reef biota. In this context, weassessed the damages caused by anchoring on benthic organ-isms along rocky reefs at two of the most visited dive siteswithin the ACMER. The effects of anchoring are discussed,
and the results of this study are extrapolated to current andpotential future levels of anchoring within the ACMER.
Material and methods
Benthic communities of Arraial do Cabo
The anchors of scuba diving boats were assessed in the divesites of Abobrinha and Anequim (Fig. 1). Both areas areamong the most visited by divers in the ACMER and repre-sent small inlets protected from the prevailing NE winds inCabo Frio island. The benthic sessile community ofAbobrinha is composed mainly of a conspicuous epilithic al-gal matrix (EAM) and the zoanthid Palythoa caribaeorumcovering up to 40% of the substratum (Rogers et al. 2014).Hard corals cover ~13% of the reef, represented by thebranching form Millepora alcicornis, and massive formsSiderastrea stellata and Mussismilia hispida (Rogers et al.2014; Lima and Coutinho 2015). At Abobrinha, EAM repre-sents 40 to 60% of benthic cover, withM. alcicornis occurringdown to 4 m deep, while sponges (mainly Aplysina) and theoctocoral Phyllogorgia dilatata are more abundant at lowerdepths (ca. 8 m) (Rogers et al. 2014). Likewise, EAM is thedominant benthic group at Anequim, while M. alcicornis isrestricted to shallow areas, and P. caribaeorum increases itscover towards deeper areas (ca. 9 m). As indicated by previousauthors cited above, both sites have similar benthic coverconcerning dominant groups. Our aim here in comparingthese two sites was to observe if there were differences indiving pressure between sites, and if that would lead to similareffects to benthic fauna.
Anequim
CardeirosIlha dos Porcos
Abobrinha
Saco do cherne
Ponta Sul
Prainha
Ponta Leste
Main dive siteSecondary dive site
*
*
* Surveyed site
Brazil
1500m0
23°S
42°WFig. 1 Map of the Arraial do Cabo indicating the main dive sites. The main dive sites are those which sustain 90% of total dive annually in the region(ca. 25,000). A map with all dive sites can be found at www.divepoint.com.br/english/divesites.html
V.J. Giglio et al.

Data collection and analysis
Data was collected between September 2014 and January 2016.Observers used scuba gear to sample the anchoring underwater.In order to evaluate possible damages caused by the anchors,the observer swam along the shallow reefs of the two dive sites,visually assessing all anchoring along the reef, whilst verifyingif the anchor and its cable were in physical contact with benthicorganisms. Since all physical contact between anchors and ben-thic organisms resulted in physical damage, dislodgement,breaks or scratches, we referred to them throughout the textas ‘damage’. We described whether the damage was causedby i) the anchor and ii) the cable that connects the anchor tothe boat (see Fig. 2a-d). Some anchors have a meter chainconnected between them and the cable. For analysis purpose,all were described only as ‘cable’. Damages were assignedto the following organisms: i) the fire coral Milleporaalcicornis; ii) the massive coral Siderastrea stellata; iii) thegorgonian Phyllogorgia dilatata; iv) the zoanthid Palythoacaribaeorum; v) sponges (genus Aplysina); vi) epilithic algalmatrix (EAM); and, vii) sea urchins. Additionally, it was re-corded if the anchoring caused damage to more than one or-ganism and if it was damaged in two different areas, for in-stance, through two parts of the cable.
The differences in the number of damages caused by theanchor and the cable was verified using Kruskal-Wallis test,
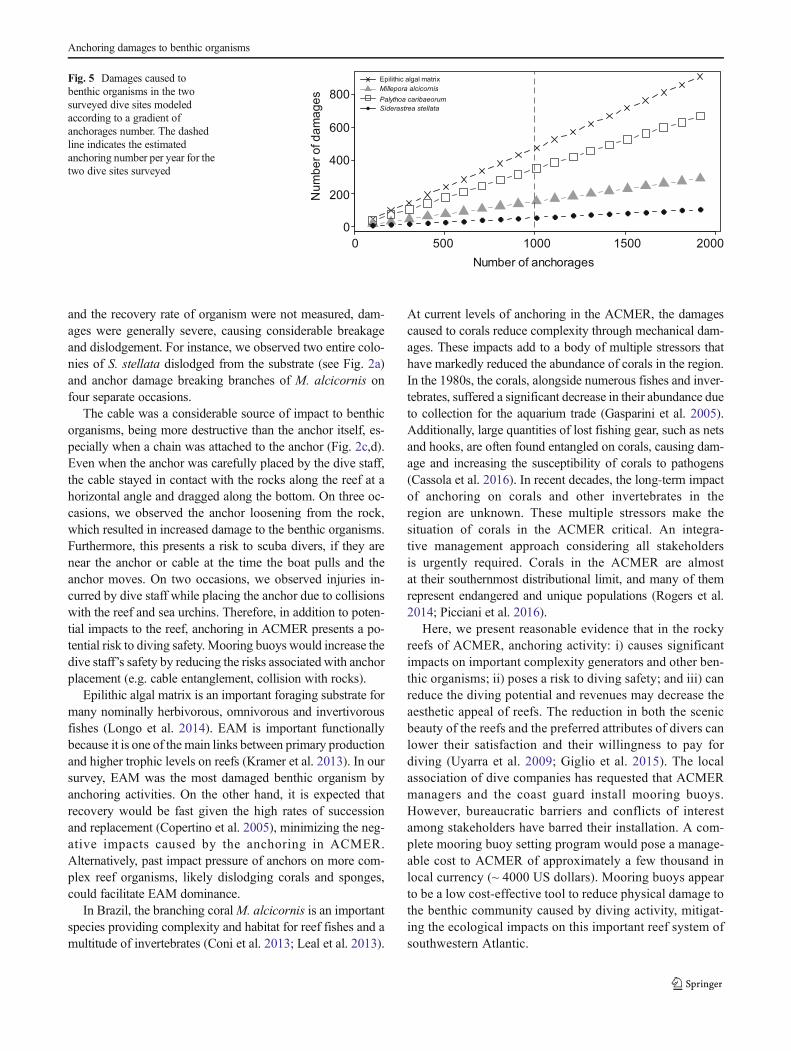
since data were nonparametric. The number of damagescaused by the anchor and cable according to benthic organismwere compared using the Wilcoxon test. A simple linear mod-el for estimating the number of damages caused by increasesin anchoring, was built to predict damages resulting from in-creasing diving activity. For instance, anchorages were sam-pled 112 times, and caused 39 damages on P. caribaeorum.This value was extrapolated to higher visitation levels by sim-ple multiplication of the results to 2,3,4 and so on.
Results
A total of 112 anchorages were observed, 57 in Abobrinha and55 in Anequim. Benthic organisms were damaged 139 times,in which the majority occurred in EAM (n = 53),P. caribaeorum (n = 39), M. alcicornis (n = 17), sea urchins(n = 15), sponges (n = 7), S. stellata (n = 6) and P. dilatata(n = 2). Forty-five percent (n = 52) of anchorages did notcontact any benthic organisms, only the bare rock.
In both sites, EAM and P. caribaeorum were damaged athigher frequency, followed by M. alcicornis and sea urchins(Fig. 3a-b). The number of damages caused by the anchor wasnot significantly different between sites (Fig. 4a) and the cablecaused significantly more damage to P. caribaeorum atAbobrinha (χ2 = 12.5, p < 0.001; Fig. 4b). Among the
Fig. 2 Damages caused byanchoring on benthic organismsin Arraial do Cabo. a Anchordamaging a colony of thefire coral M. alcicornis; b anchorand cable damagingM. alcicornisand the zoanthid P. caribaeorum;c cable (chain) damagingP. caribaeorum; and d suspendinga colony of the massive coralS. stellata
Anchoring damages to benthic organisms
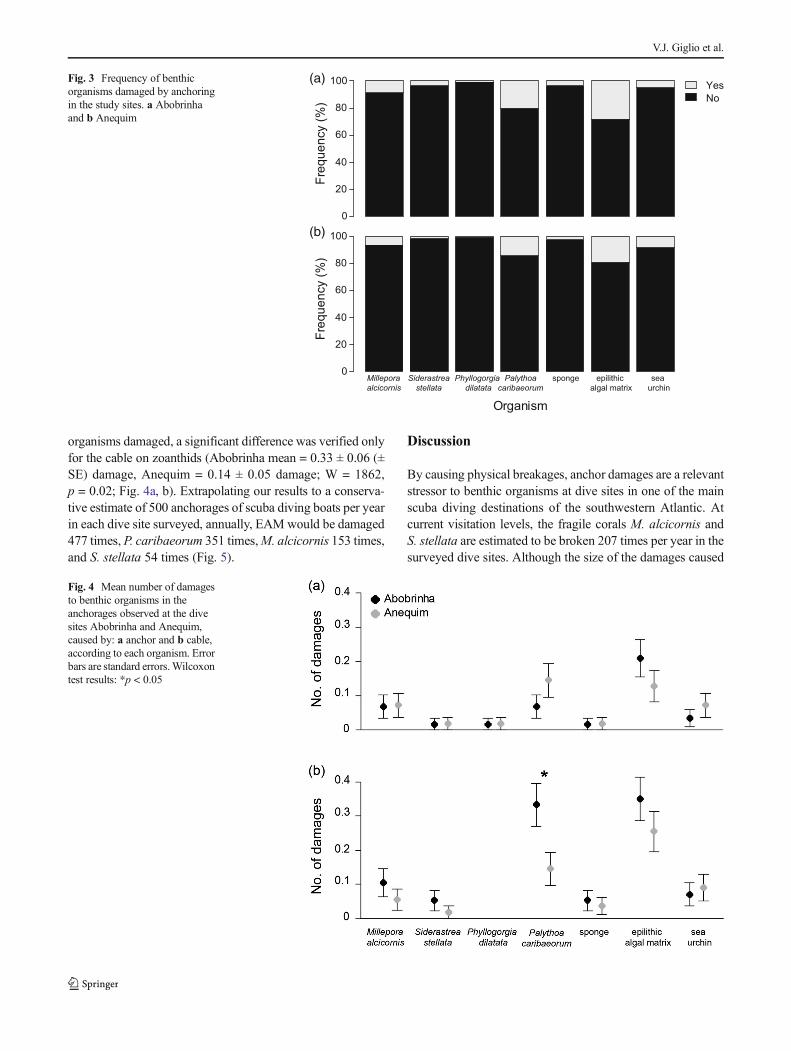
organisms damaged, a significant difference was verified onlyfor the cable on zoanthids (Abobrinha mean = 0.33 ± 0.06 (±SE) damage, Anequim = 0.14 ± 0.05 damage; W = 1862,p = 0.02; Fig. 4a, b). Extrapolating our results to a conserva-tive estimate of 500 anchorages of scuba diving boats per yearin each dive site surveyed, annually, EAMwould be damaged477 times, P. caribaeorum 351 times,M. alcicornis 153 times,and S. stellata 54 times (Fig. 5).
Discussion
By causing physical breakages, anchor damages are a relevantstressor to benthic organisms at dive sites in one of the mainscuba diving destinations of the southwestern Atlantic. Atcurrent visitation levels, the fragile corals M. alcicornis andS. stellata are estimated to be broken 207 times per year in thesurveyed dive sites. Although the size of the damages caused
Fig. 4 Mean number of damagesto benthic organisms in theanchorages observed at the divesites Abobrinha and Anequim,caused by: a anchor and b cable,according to each organism. Errorbars are standard errors.Wilcoxontest results: *p < 0.05
Freq
uenc
y (%
)
0
20
40
60
80
100
Freq
uenc
y (%
)
0
20
40
60
80
100
(a)
(b)
Milleporaalcicornis
Siderastreastellata
Phyllogorgia dilatata
Palythoacaribaeorum
sponge epilithic algal matrix
sea urchin
Organism
YesNo
Fig. 3 Frequency of benthicorganisms damaged by anchoringin the study sites. a Abobrinhaand b Anequim
V.J. Giglio et al.
and the recovery rate of organism were not measured, dam-ages were generally severe, causing considerable breakageand dislodgement. For instance, we observed two entire colo-nies of S. stellata dislodged from the substrate (see Fig. 2a)and anchor damage breaking branches of M. alcicornis onfour separate occasions.
The cable was a considerable source of impact to benthicorganisms, being more destructive than the anchor itself, es-pecially when a chain was attached to the anchor (Fig. 2c,d).Even when the anchor was carefully placed by the dive staff,the cable stayed in contact with the rocks along the reef at ahorizontal angle and dragged along the bottom. On three oc-casions, we observed the anchor loosening from the rock,which resulted in increased damage to the benthic organisms.Furthermore, this presents a risk to scuba divers, if they arenear the anchor or cable at the time the boat pulls and theanchor moves. On two occasions, we observed injuries in-curred by dive staff while placing the anchor due to collisionswith the reef and sea urchins. Therefore, in addition to poten-tial impacts to the reef, anchoring in ACMER presents a po-tential risk to diving safety. Mooring buoys would increase thedive staff’s safety by reducing the risks associated with anchorplacement (e.g. cable entanglement, collision with rocks).
Epilithic algal matrix is an important foraging substrate formany nominally herbivorous, omnivorous and invertivorousfishes (Longo et al. 2014). EAM is important functionallybecause it is one of the main links between primary productionand higher trophic levels on reefs (Kramer et al. 2013). In oursurvey, EAM was the most damaged benthic organism byanchoring activities. On the other hand, it is expected thatrecovery would be fast given the high rates of successionand replacement (Copertino et al. 2005), minimizing the neg-ative impacts caused by the anchoring in ACMER.Alternatively, past impact pressure of anchors on more com-plex reef organisms, likely dislodging corals and sponges,could facilitate EAM dominance.
In Brazil, the branching coralM. alcicornis is an importantspecies providing complexity and habitat for reef fishes and amultitude of invertebrates (Coni et al. 2013; Leal et al. 2013).
At current levels of anchoring in the ACMER, the damagescaused to corals reduce complexity through mechanical dam-ages. These impacts add to a body of multiple stressors thathave markedly reduced the abundance of corals in the region.In the 1980s, the corals, alongside numerous fishes and inver-tebrates, suffered a significant decrease in their abundance dueto collection for the aquarium trade (Gasparini et al. 2005).Additionally, large quantities of lost fishing gear, such as netsand hooks, are often found entangled on corals, causing dam-age and increasing the susceptibility of corals to pathogens(Cassola et al. 2016). In recent decades, the long-term impactof anchoring on corals and other invertebrates in theregion are unknown. These multiple stressors make thesituation of corals in the ACMER critical. An integra-tive management approach considering all stakeholdersis urgently required. Corals in the ACMER are almostat their southernmost distributional limit, and many of themrepresent endangered and unique populations (Rogers et al.2014; Picciani et al. 2016).
Here, we present reasonable evidence that in the rockyreefs of ACMER, anchoring activity: i) causes significantimpacts on important complexity generators and other ben-thic organisms; ii) poses a risk to diving safety; and iii) canreduce the diving potential and revenues may decrease theaesthetic appeal of reefs. The reduction in both the scenicbeauty of the reefs and the preferred attributes of divers canlower their satisfaction and their willingness to pay fordiving (Uyarra et al. 2009; Giglio et al. 2015). The localassociation of dive companies has requested that ACMERmanagers and the coast guard install mooring buoys.However, bureaucratic barriers and conflicts of interestamong stakeholders have barred their installation. A com-plete mooring buoy setting program would pose a manage-able cost to ACMER of approximately a few thousand inlocal currency (~ 4000 US dollars). Mooring buoys appearto be a low cost-effective tool to reduce physical damage tothe benthic community caused by diving activity, mitigat-ing the ecological impacts on this important reef system ofsouthwestern Atlantic.
0 500 1000 1500 20000
200
400
600
800
Number of anchorages
Num
ber o
f dam
ages
Epilithic algal matrixMillepora alcicornisPalythoa caribaeorumSiderastrea stellata
Fig. 5 Damages caused tobenthic organisms in the twosurveyed dive sites modeledaccording to a gradient ofanchorages number. The dashedline indicates the estimatedanchoring number per year for thetwo dive sites surveyed
Anchoring damages to benthic organisms

Acknowledgements We thank the diving operators association -Associação das Operadoras de Mergulho de Arraial do Cabo and theArraial do Cabo marine extractive reserve - ICMBio (through VivianeLasmar and Rafaela Farias) for research permits and support. C. Garlandfor English editing. The first author was supported by the BrazilianMinistry of Science and Technology (CNPq). TCM was supported byFAPERJ (postdoc grant # E-26/202.858/2016). CELF was supported bygrants from CNPq and FAPERJ.
References
Cassola GE, Pacheco MSC, Barbosa MC, Hansen DM, Ferreira CEL(2016) Decline in abundance and health state of an Atlantic subtrop-ical gorgonian population. Mar Pollut Bull 104:329–334
Coni EOC, Ferreira CM,Moura RL,Meirelles PM,Kaufman L, Francini-Filho RB (2013) An evaluation of the use of branching fire-corals(Millepora spp.) as refuge by reef fish in the Abrolhos Bank, easternBrazil. Environ Biol Fish 96:45–55
Copertino MS, Connell SD, Chesire A (2005) The prevalence and pro-ductivity of turf-forming algae on a temperate subtidal coast.Phycologia 44:241–248
Creed JC, Amado Filho GM (1999) Disturbance and recovery of themacroflora of a seagrass (Halodule wrightii, Ascherson) meadowin the Abrolhos marine National Park, Brazil: an experimental eval-uation of anchor damage. J Exp Mar Biol Ecol 235:285–306
Davis GE (1977) Anchor damage to a coral reef on the coast of Florida.Biol Conserv 11:29–34
Dinsdale EA, Harriott VJ (2004) Assessing anchor damage on coral reefs:a case study in selection of environmental indicators. EnvironManag 33:126–139
Ferreira CEL, Gonçalves JEA, Coutinho R (2001) Community structureof fishes and habitat complexity on a tropical rocky shore. EnvironBiol Fish 61:353–369
Francour P, Ganteaume A, Poulain M (1999) Effects of boat anchoring inPosidonia oceanica seagrass beds in the port-Cros National Park(north-western Mediterranean Sea). Aquat Conserv 9:391–400
Garrod B, Gossling S (2012) New Frontiers in marine tourism. Elsevier,Amsterdam
Gasparini JL, Floeter SR, Ferreira CEL, Sazima I (2005) Marine orna-mental trade in Brazil. Biodivers Conserv 14:2883–2899
Giglio VJ, Luiz OJ, Schiavetti A (2015) Marine life preferences andperceptions among recreational divers in Brazilian coral reefs.Tour Manag 51:49–57
Harriott VJ, Davis D, Banks SA (1997) Recreational diving and its im-pact in marine protected areas in eastern Australia. Ambio 26:173–179
Holbrook SJ, Brooks AJ, Schmitt RJ (2003) Variation in structural attri-butes of patch-forming corals and in patterns of abundance of asso-ciated fishes. Mar Freshw Res 53:1045–1053
Jameson SC, Ammar M, Saadalla E, Mostafa H, Riegl B (1999) A coraldamage index and its application to diving sites in the Egyptian RedSea. Coral Reefs 18:333–339
Jameson SC, Ammar MSA, Saadalla E, Mostafa HM, Riegl B (2007) Aquantitative ecological assessment of diving sites in the EgyptianRed Sea during a period of severe anchor damage: a baseline forrestoration and sustainable tourism management. J Sustain Tour 15:309–323
Kramer MJ, Bellwood O, Bellwood DR (2013) The trophic importanceof algal turfs for coral reef fishes: the crustacean link. Coral Reefs 32(2):575-583
Leal ICS, Pereira PHC, Araujo ME (2013) Coral reef fish association andbehaviour on the fire coral Millepora spp. in north-east Brazil. J MarBiol Assoc UK 93:1703–1711
Lima LFO, Coutinho R (2015) The reef coral Siderastrea stellata thrivingat its range limit: population structure in Arraial do Cabo, southeast-ern Brazil. B Mar Sci 92:107–121
Longo GO, Ferreira CEL, Floeter SR (2014) Herbivory drives large-scalespatial variation in reef fish trophic interactions. Ecol Evol 4:4553–4566
Messmer V, Jones GP, Munday PL, Holbrook SJ, Schmitt RJ, Brooks AJ(2011) Habitat biodiversity as a determinant of fish communitystructure on coral reefs. Ecology 92:2285–2298
Milazzo M, Badalamentib F, Ceccherelli G, Chemello R (2004) Boatanchoring on Posidonia oceanica beds in a marine protected area(Italy, western Mediterranean): effect of anchor types in differentanchoring stages. J Exp Mar Biol Ecol 299:51–62
Picciani N, Seiblitz GL, Paiva PC, Castro CB, Zilberberg C (2016)Geographic patterns of Symbiodinium diversity associated withthe coralMussismilia hispida (Cnidaria, Scleractinia) correlate withmajor reef regions in the southwestern Atlantic Ocean. Mar Biol163:236
Roche RC, Harvey CV, Harvey JJ, Kavanagh AP, McDonald M, Stein-Rostaing V, Turner JR (2016) Recreational diving impacts on coralreefs and the adoption of environmentally responsible practiceswithin the SCUBA diving industry. Environ Manag 58:107–116
Rogers R, Correal GO, Oliveira TC, Carvalho LL, Mazurek P, BarbosaJEF, Chequer L, Domingos TFS, Jandre KA, Leão LSD, MouraLM, Occhioni GE, Oliveira VM, Silva ES, Cardoso AM, CostaAC, Ferreira CEL (2014) Coral health rapid assessment in marginalreef sites. Mar Biol Res 10:612–624
Tratalos JA, Austin TJ (2001) Impacts of recreational SCUBA diving oncoral communities of the Caribbean island of grand Cayman. BiolConserv 102:67–75
Uyarra MC, Watkinson AR, Côté IM (2009) Managing dive tourism forthe sustainable use of coral reefs: validating diver perceptions ofattractive site features. Environ Manag 43:1–16
Valentin J (2001) The Cabo Frio upwelling system, Brazil. In: Seeliger U,Kjerfve B (eds) Coastal marine ecosystems of Latin America.Springer, New York, pp 97–105
Zhang L-Y, Chung S-S, Qiu J-W (2016) Ecological carrying capacityassessment of diving site: A case study of Mabul Island, Malaysia.J Environ Manage 183:253–259
V.J. Giglio et al.





























