8/2/2019 Anatomy of the Basal Ganglia
1/2
Anatomy of the Basal Ganglia
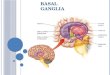
The striatum (caudaute/putamen) and subthalamic nucleus (STN)
receive excitatory input from
the cerebral cortex. Dopamine-releasing neurons in the
substantia nigra pars compacta (SNpc) connect
to neurons in the striatum and modulate the inputs from the
cortex. There is an inhibitory connection
from the striatum to the globus pallidus internal segment (GPi)
and the substantia nigra pars reticulara(SNpr), as well as an
excitatory projection from STN to GPi that is divergent (one STN
neuron contacts
many Gpi neurons). This pathway is called the direct pathway.
Another, indirect pathway of
inhibitory connections extends from the striatum to the globus
pallidus external segment (Gpe) to the
STN to the GPi. The GPi and SNpr send inhibitory output via
collaterals to the thalamus and brain stem.
The SNpr is involved in eye movements.
Types of Neurons in the Striatum
Medium spiny neuronsmake up 95% of the total. Use GABA as a
transmitter. Are the output
neurons of the striatum.
Large aspiny neuronsinterneurons that use ACh as a
transmitter.
Medium aspiny cellsinterneurons that use somatostatin as a
neurotransmitter.
Small aspiny cellsinterneurons that use GABA.
Neuron Firing in the Basal Ganglia
Neurons in the putamen and throughout the basal ganglia fire
after those in the motor cortex,
implying that the striatum is not involved primarily in the
initiation of movement.
The firing of SNpc neurons increases in response to behaviorally
significant events such as
reward or the presentation of instructional cues. These neurons
can also learn to respond to a cue
that predicts a reward.
Consequences of Damage to the Basal Ganglia
Inactivating the putamen leads to slowed movement of the
contralateral limb. Huntingtons
Disease, which causes involuntary movements, is linked to the
death of neurons that project from the
putamen to the GPe.
Damage to the STN causes large involuntary movements of the
limbs.
Lesions to the GPi cause slowness of movement, linked to a
tendency of the limbs to assume an
abnormally flexed posturethat is, an inability to turn off
muscle activity.
Damage to the SNpc causes symptoms of Parkinsons diseasetremor
and slowed movement.
8/2/2019 Anatomy of the Basal Ganglia
2/2
Model of Basal Ganglia Function
One hypothesis suggests that the basal ganglia automatically
execute learned movement
sequences. This theory is supported by the fact that patients
with PD have difficulty coordinating
complex movements of several body parts in sequence, such as
when writing.
Another suggests that the basal ganglia form two opposing motor
pathways, the direct and
indirect pathways described above. Increased activity in the
direct pathway causes excessive
movement, while activity in the indirect pathway inhibits
movement.
A third suggests that the basal ganglia act as a brake on motor
movement. The theory
suggests that STN neurons excite the GPi widely, inhibiting
motor output. At the same time, signals sent
from the cortex to the striatum to the GPi inhibit a small part
of the GPi, selecting a certain motor
pattern to be disinhibited and suppressing surrounding
patterns.
Anatomy of the Cerebellum
The cerebellar cortex is divided into three lobes: anterior,
posterior, and flocculonodular. Each
lobe consists of thin folds called folia. This sheet is laid
over four cerebellar nuclei (CN) on each side.
Three cerebellar peduncles on each side connect the cerebellum
to the brain stem.
The cortex consists of three layers. The granular cell layer, on
the bottom, contains an enormous
number of granule cells. The Purkinje cell layer contains cell
bodies of PCs, and the molecular layer
contains only dendrites and axons.
Incoming information from all over the brain sends information
via mossy fibers (MFs) to the
granule cells. These cells relay the information to the
molecular layer via parallel fibers (PFs), which
make contact with the dendrites of PCs. In addition, error
correction signals are sent from the inferiorolive to PCs via
climbing fibers (CFs). Each PC is only contacted by one CF.
The PCs send inhibitory signals to the CN. These nuclei are
linked reciprocally to populations of
neurons in other parts of the brain, forming attractor
networks.
Long-term Depression (LTD) Takes Place Between the Synapses of
PFs and PCs
Unlike LTP, LTD requires the presence of three factors in order
to take place:
1) Depolarization of the dendrite
2) Activation of glutamate receptors on a particular spine on
the dendrite
3) The firing of a CF (training signal) hundreds of milliseconds
later
This process marks synapses causing movements that result in
error and causes them to be less
effective. Plasticity in the cerebellum is thought to be
involved only in adjusting motor responses, not in
forming a link between a conditioned and unconditioned
stimulus.