Embed Size (px)
Citation preview

Camp. Biochem. PhysioL Vol. 103B, No. 3, pp. 645-650, 1992 0305-0491/92 $5.00 + 0.00 Printed in Great Britain © 1992 Pergamon Press Ltd
ANATOMICAL DISTRIBUTIONS AND TEMPORAL VARIATIONS OF LIPID CLASSES IN SEA SCALLOPS
PLACOPECTEN MAGELLANICUS (GMELIN) FROM GEORGES BANK (NOVA SCOTIA)
GUILLERMO E. NAPOLITANO* and ROBERT G. ACKMAN t Department of Oceanography, Dalhousie University, Halifax, Nova Scotia, Canada B3H4J1; and ,Canadian Institute of Fisheries Technology; Technical University of Nova Scotia, Halifax, Canada
B3J 2X4 (Tel.: 902 420-7758; Fax: 902 420-0219)
(Received 2 April 1992; accepted 5 May 1992)
Abstract--l. Lipid contents, lipid class compositions and their temporal variations were studied in the sea scallop Placopecten magellanicus (Gmelin) from Georges Bank.
2. Analyses of the major organs (digestive gland, gonad, adductor muscle, mantle, and gills) presented different and characteristic lipid profiles.
3. The lipid contents of adductor muscle, gills, mantle and male gonad usually ranged from 0.6 to 1% of the wet weight. Lipids in these organs were mostly structural phospholipids and sterols, and therefore these tissues were not functioning as sites for lipid reserves.
4. The digestive gland and the female gonad exhibited dramatic changes in lipid content and composition associated with feeding and reproduction.
5. The relatively high lipid contents in digestive gland and mature female gonad (4 and 8% of wet weights, respectively) were consequences of the accumulation of both triacylgiycerols and phospholipids. Accumulations of phospholipids in somatic organs of marine invertebrates have not been previously documented.
INTRODUCTION
Lipids in marine organisms are associated with a variety of functions, reflecting special biochemical and environmental conditions of the marine milieu. Lipids are the major metabolic reserve in most marine animals, including mammals, fish and crustaceans (Sargent, 1976). It is now agreed that carbohydrates are a major energy reserve in most adult bivalves, while lipids in the form of triacylglycerols (TG) are accumulated by the larvae (Holland, 1978; Napolitano et al., 1988). Concurrent with this tem- poral segregation between energy storage as lipids in larvae and as carbohydrates in the postlarval stages of bivalves, there is also a spatial segregation of these components in the adult organisms. Many molluscs store complex carbohydrates (mostly glycogen) in the mantle and in the adductor muscle, while lipids are accumulated in the digestive gland.
The sea scallop Placopecten magellanicus is a bivalve mollusc (Bivalvia, Pectinidae) commonly occurring along the east coast continental shelf of North America (Posgay, 1957). The major offshore Canadian fishery is located in the northern portion of Georges Bank, which is one of the most biologically productive areas in temperate latitudes (Riley, 1982). Secondary fishing grounds are in the nearby Bay of
*Present address: Center for Environmental Biotechnology, University of Tennessee, 10515 Research Dr. Knoxville, TN 37932, U.S.A.
tAuthor to whom correspondence should be addressed.
Fundy, in the southern Gulf of St. Lawrence, and on the St. Pierre Bank and in Port au Port Bay, Newfoundland (Bourne, 1964).
Further study of the biochemical composition of sea scallops will enhance our understanding of basic aspects of their biology and relationship with environmental factors. This report presents a study of the anatomical distribution and temporal variations of the major lipid classes in sea scallops from Georges Bank.
MATERIALS AND METHODS
Lipid analysis: general procedures Sea scallops P. magellanicus were collected in the
Canadian sector of Georges Bank at approximately N 41°35'09"; W 61 ° 10'1 I". Specimens were captured during commercial fishing operations between August 1987 and October 1989 according to the schedule shown in Table 1.
Scallops used for lipid analyses were transported live to the laboratory. Adult animals (shell height > 10cm) were sexed and their major organs, i.e. digestive gland, gonad, adductor muscle, mantle and gills, were dissected out and individually weighed.
Scallop organ lipids were extracted with a mixture of chloroform-methanol (2:1) using a stainless steel Waring Blendor. In effect, the tissues were extracted following the classical method described by Bligh and Dyer (1959). The chloroform layer was washed with water, dried over anhydrous sodium sulfate and filtered. Small quantities of solvents were evaporated under a stream of nitrogen in glass centrifuge tubes placed in a water bath at 40-50°C. The evaporation of a large amount of solvent was conducted using a rotary evaporator under reduced pressure. Except for dissection steps, all procedures were carried out under a
645

646 GUILLERMO E. NAPOLITANO and ROBERT G. ACKMAN
Table 1. Schedule of sample collection of scallops P. magellanicus on Georges Bank
Date Boat
7 August 1987 Scotia Trawlers October 1987 Scotia Trawlers 7 June 1988 Scotia Trawlers 3 July 1988 Cape Race 15 October 1988 Scotia Trawlers 29 January 1989 Mersey 9 October 1989 Scotia Trawlers
nitrogen atmosphere. After the extraction, the lipid extracts were stored in chloroform in screw cap (Teflon-lined) glass vials at -35°C under a nitrogen atmosphere. Most tissue lipids were extracted immediately; a few samples were stored at -35°C for a few days for further analyses.
Total lipid contents were determined gravimetrically after exhaustive removal of the solvent from an aliquot of the lipid extract. Results are expressed in percentages of lipid on a wet weight basis. The different lipid classes present in these samples were first qualitatively studied by thin-layer chro- matography (TLC) and then quantitatively determined by Iatroscan TLC-FID (flame ionization detection) after prior separation on silica gel Chromarods-SII (Ackman, 198l).
Fatty acid and lipid standards were purchased from Serdary Research Laboratories (London, Ontario). The organic solvents were A.C.S. reagent grade (Anachemia), redistilled in glass before use. The acids and other reagents were A.C.S. grade from Fisher Scientific Company (Canada). Glassware was carefully cleaned before use by soaking overnight in a concentrated KOH-ethanol solution or by washing with detergent. It was then rinsed several times with tap water, distilled water and acetone before oven drying.
Lipid class separation by TLC TLC was performed on "Prekote" silica gel glass plates
(20 x 20 cm, 200/zm particle size, Applied Science Labora- tories, College Park, PA). Before use the plates were cleaned by developing in ethyl acetate and activated by heating at 110°C for 30 min. Lipids were applied as chloroform sol- utions using a plate streaker (Applied Science Laboratories). Plates were developed in solvent-saturated glass tanks. The common developing solvent was hexane~liethyl ether- acetic acid (80:20:1). Lipids were visualized by spraying with a 1% 2',7'-dichlorofluorescein solution in ethanol and observation under UV light. Standards and standard mix- tures were spotted alongside the samples to compare R r values.
Analyses of the lipid classes by TLC-FID An Iatroscan analyzer (Iatron Laboratories, Inc., Tokyo)
equipped with a FID was utilized for the analyses of the lipid classes in scallop tissues. The general procedure was that described by Sipos and Ackman (1978); for more details see Ackman (1981). Ten Chromarods-SII were activated
by passing through the FID scanner in a frame and then spotted with the sample or standard. Disposable glass micropipettes (1-5/zl; Drummond Sci. Co., Broomall, PA) were used for this purpose. The frames with rods were placed in appropriate solvent-saturated glass tanks and developed for 45 min. The common developing solvent was hexane-diethyl ether-formic acid (97:3 : 1.5). Developed rods in their frame were placed in an oven at I I0°C for I min to evaporate the solvents and then transferred to the scanning frame of the Iatroscan analyzer. The scanning operation was performed under the following conditions: flow rate of hydrogen, 160 ml/min; flow rate of air, 2 l/min; voltage of the detector, 8 mV full scale; scanning speed, 0.42 cm/s. The recording and integration of the peaks was provided by a Spectra Physics SP 4200 computing integra- tor. Before reuse the rods were soaked in 30% nitric acid overnight, washed and blank scanned on the FID of the Iatroscan analyzer. Iatroscan results are FID area per cent expressed as the mean of three separate samples, each applied to three different rods. Standard deviation of rod replicates never exceeded 10% of the mean.
RESULTS
Lipid content and lipid class composition
Lipids in the adduc to r muscle, gills and mant le accounted for a b o u t 0 . 6 - 1 % of the wet weight (Table 2), and showed very similar lipid profiles t h r o u g h o u t the year. Phosphol ip ids (PL) and sterols in these three " lean o rgans" accounted for abou t 80 and 18% of the total lipids, respectively, dur ing the winter (Table 2). The remaining lipids were minor amoun t s of T G and trace levels of sterol esters and hydrocarbon- l ike compounds . A similar pa t tern of lipid compos i t ion was found in the summer samples of these tissues, a l though the sterol concent ra t ion was decreased. No changes in the total lipid content , and no T G accumulat ion, were observed in adduc to r muscle, mant le or gills dur ing the summer (Table 2).
Besides the presence of s t ructural lipids, the scallop digestive gland conta ined large quant i t ies of T G (Table 2). The level of these neutral lipids in this o rgan changed seasonally, showing max imum values in summer (60% of total lipids) and min imum values dur ing the winter (40-50%) .
The p ropor t ion of free fatty acids was relatively low in all the organs (~<1.6% of the total lipids). An exception to this observa t ion was found in summer samples of digestive glands, where free fatty acids were 6.9% of the total.
The male gonads were small and yielded too little lipid for accuracy. However, the lipid profile was similar to those of the " lean organs" . Al though
Table 2. Anatomical distribution of lipid classes in the sea scallop (P. magellanicus) from Georges Bank reported as uncorrected TLC-FID area per cent
Winter Summer
DG GO MU GI MA DG GO MU GI MA
Polar lipidst 48.6 91.0 80.3 76.0 82.8 31.2 68.0 91.2 83.4 88.3 Sterols 9.1 3.1 19.7 18.7 17.7 0.64 4.8 9.3 9.9 9.8 Triacylglycerols 41.1 5.1 1.2 3.6 0.55 60.2 26.6 0.74 6.1 0.56 Free fatty acids 1.4 1.6 1.1 0.38 0.34 6.9 0.20 tr 0.50 0.4 Sterol ester 0.70 0.20 0.33 0.11 0.39 0.18 tr nd 0.31 tr Total lipids~ 2.4 1.2 1.1 0.64 0.61 4.4 1.3 0.78 0.75 0.66
*DG = digestive gland, GO = female gonad, MU = adductor muscle, GI = gills, MA = mantle, tr = trace amounts (<0.01%), nd= not detected.
tMainly phospholipids. :~Wet weight, %. For male gonads the winter value was 0.78%, the summer value 1.18%.
Placopecten lipids 647
this gonad suffers dramatic seasonal growth and differentiation, these changes did not affect its total lipid concentration. All samples of the male gonad analyzed during the different seasons exhibited an approximate total lipid content of 1% of the wet tissue weight (Table 2).
Temporal variations of lipids in the digestive gland and in the female gonad
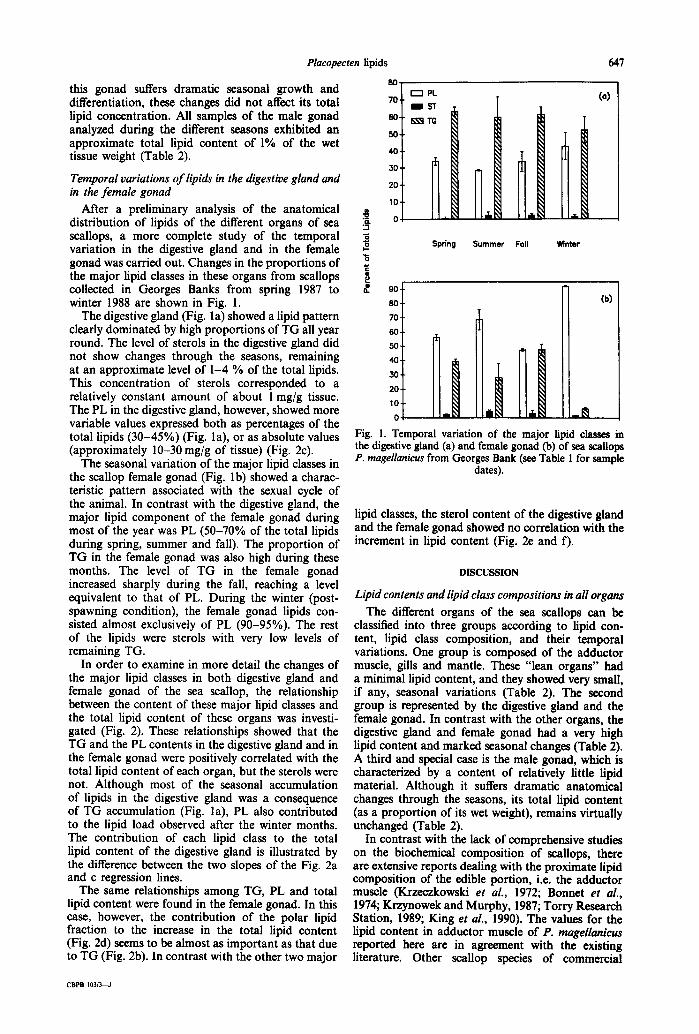
After a preliminary analysis of the anatomical distribution of lipids of the different organs of sea scallops, a more complete study of the temporal variation in the digestive gland and in the female gonad was carried out. Changes in the proportions of the major lipid classes in these organs from scallops collected in Georges Banks from spring 1987 to winter 1988 are shown in Fig. 1.
The digestive gland (Fig. la) showed a lipid pattern clearly dominated by high proportions of TG all year round. The level of sterols in the digestive gland did not show changes through the seasons, remaining at an approximate level of 1-4 % of the total lipids. This concentration of sterols corresponded to a relatively constant amount of about 1 mg/g tissue. The PL in the digestive gland, however, showed more variable values expressed both as percentages of the total lipids (30-45%) (Fig. la), or as absolute values (approximately 10-30 mg/g of tissue) (Fig. 2c).
The seasonal variation of the major lipid classes in the scallop female gonad (Fig. lb) showed a charac- teristic pattern associated with the sexual cycle of the animal. In contrast with the digestive gland, the major lipid component of the female gonad during most of the year was PL (50-70% of the total lipids during spring, summer and fall). The proportion of TG in the female gonad was also high during these months. The level of TG in the female gonad increased sharply during the fall, reaching a level equivalent to that of PL. During the winter (post- spawning condition), the female gonad lipids con- sisted almost exclusively of PL (90-95%). The rest of the lipids were sterols with very low levels of remaining TG.
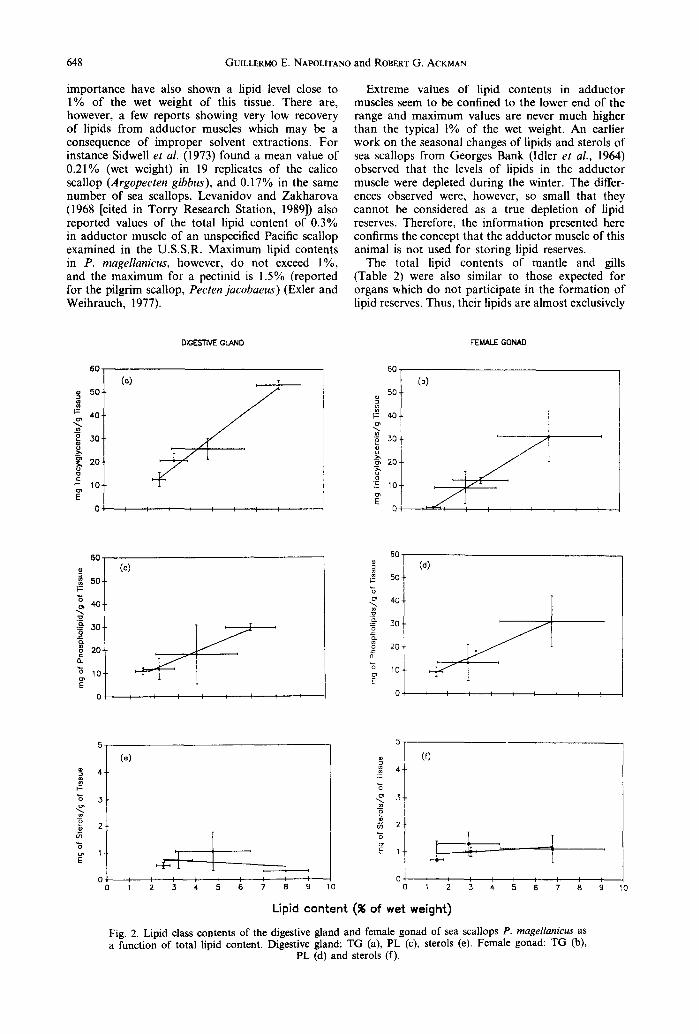
In order to examine in more detail the changes of the major lipid classes in both digestive gland and female gonad of the sea scallop, the relationship between the content of these major lipid classes and the total lipid content of these organs was investi- gated (Fig. 2). These relationships showed that the TG and the PL contents in the digestive gland and in the female gonad were positively correlated with the total lipid content of each organ, but the sterols were not. Although most of the seasonal accumulation of lipids in the digestive gland was a consequence of TG accumulation (Fig. la), PL also contributed to the lipid load observed after the winter months. The contribution of each lipid class to the total lipid content of the digestive gland is illustrated by the difference between the two slopes of the Fig. 2a and c regression lines.
The same relationships among TG, PL and total lipid content were found in the female gonad. In this case, however, the contribution of the polar lipid fraction to the increase in the total lipid content (Fig. 2d) seems to be almost as important as that due to TG (Fig. 2b). In contrast with the other two major
80-
"5. "1
"6 i
I~1 PL 70-
BI IS 3_ 60- K~ T
i 50-
40.
30.
20.
10.
0 1
Spdng
R.L
Summer Foil W~nter
(o)
90. 8o. (b) 70. 60.
Fig. 1. Temporal variation of the major lipid classes m the digestive gland (a) and female gonad (b) of sea scallops P. magellanicus from Georges Bank (see Table 1 for sample
dates).
lipid classes, the sterol content of the digestive gland and the female gonad showed no correlation with the increment in lipid content (Fig. 2e and f).
DISCUSSION
Lipid contents and lipid class compositions in all organs
The different organs of the sea scallops can be classified into three groups according to lipid con- tent, lipid class composition, and their temporal variations. One group is composed of the adductor muscle, gills and mantle. These "lean organs" had a minimal lipid content, and they showed very small, if any, seasonal variations (Table 2). The second group is represented by the digestive gland and the female gonad. In contrast with the other organs, the digestive gland and female gonad had a very high lipid content and marked seasonal changes (Table 2). A third and special case is the male gonad, which is characterized by a content of relatively little lipid material. Although it suffers dramatic anatomical changes through the seasons, its total lipid content (as a proportion of its wet weight), remains virtually unchanged (Table 2).
In contrast with the lack of comprehensive studies on the biochemical composition of scallops, there are extensive reports dealing with the proximate lipid composition of the edible portion, i.e. the adductor muscle (Krzeczkowski et al., 1972; Bonnet et al., 1974; Krzynowek and Murphy, 1987; Torry Research Station, 1989; King et al., 1990). The values for the lipid content in adductor muscle of P. magellanicus reported here are in agreement with the existing literature. Other scallop species of commercial
CBPB 103/3~J
648 GUILLERMO E. NAPOLITANO and ROBERT G. ACKMAN
importance have also shown a lipid level close to 1% of the wet weight of this tissue. There are, however, a few reports showing very low recovery of lipids from adductor muscles which may be a consequence of improper solvent extractions. For instance Sidwell et aL (1973) found a mean value of 0.21% (wet weight) in 19 replicates of the calico scallop (Argopecten gibbus), and 0.17% in the same number of sea scallops. Levanidov and Zakharova 0968 [cited in Torry Research Station, 1989]) also reported values of the total lipid content of 0.3% in adductor muscle of an unspecified Pacific scallop examined in the U.S.S.R. Maximum lipid contents in P. magellan&us, however, do not exceed 1%, and the maximum for a pectinid is 1.5% (reported for the pilgrim scallop, Pecten jacobaeus) (Exler and Weihrauch, 1977).
Extreme values of lipid contents in adductor muscles seem to be confined to the lower end of the range and maximum values are never much higher than the typical 1% of the wet weight. An earlier work on the seasonal changes of lipids and sterols of sea scallops from Georges Bank (Idler et al., 1964) observed that the levels of lipids in the adductor muscle were depleted during the winter. The differ- ences observed were, however, so small that they cannot be considered as a true depletion of lipid reserves. Therefore, the information presented here confirms the concept that the adductor muscle of this animal is not used for storing lipid reserves.
The total lipid contents of mantle and gills (Table 2) were also similar to those expected for organs which do not participate in the formation of lipid reserves. Thus, their lipids are almost exclusively
6 0 .
5 0 .
~" 4 0 .
~ 3 0 .
- ~ 2 0 .
o
~ 1 0 . ? 0
(o)
DIGESTIVE GLAND
j 1 I I I I I I I I I
6 0
5O
~= 4O
3 0
~ , 2O-
~ 1 0 -
O l
(b)
FEMALE GONAD
I i I I
6 0 ¸
g 50
4 0
3o
~ 2o a.
0 .
(c) 60
5O
"6
.& N 3O
8 2o t "6 10
0
(d)
l
I I t I t I I
g
`6
ta
"5
(e) (f)
i
5 -
4 -
"6 3-
o 0~
2 -
`6
E' 1.
i b 0
9 I 0 a
1
f
1 2 3 4 5 I I / I I I ] 1 2 3 4 6 7 8
Lipid content (% of wet weight) Fig. 2. Lipid class contents of the digestive gland and female gonad of sea scallops P. magellanicus as a function of total lipid content. Digestive gland: TG (a), PL (c), sterols (e). Female gonad: TG (b),
PL (d) and sterols (f).
1 0

Placopecten lipids 649
made up of structural PL and sterols. There is very limited data on the lipid content and composition of mantle and gills in molluscs (Joseph, 1982; 1989). As in the case of the adductor muscle, however, there is no chemical or histological observation sup- porting the presence of lipid storage in these organs. In a study of the seasonal changes in the biochemical composition of the black scallop Chlamys varia (Shafee, 1981), large changes in lipid content of the animals occurred in the reproductive tissues, and much smaller variations were found in the "somatic tissues" (which included the lipid-rich digestive gland). The only information available for compari- son of the lipid content of mantle and gills in bivalves (Jarzebski et al., 1986, Tsuji and Nishida, 1988) confirms the low (< 1%) and steady lipid content reported here.
The male gonad presented a lipid profile similar to the one already described for adductor muscle, gills and mantle, (Table 2). Consequently, this gonad could be included in the group of "lean organs". Despite the marked seasonality of gamete prolifer- ation and maturation, no pronounced changes in the gross lipid composition of this organ could be observed. This may be explained if we assume that the proximate lipid composition of both reproductive and non-reproductive cells of the male gonad do not have to be different.
The female gonad and the digestive gland (in both male and female scallops) exhibited a much richer and more dynamic lipid profile. The main difference between the lipid composition of these two organs and the rest of the body is the presence of a large accumulation of reserves in the form of TG. Changes in the quantity and in the quality of the lipids in these organs are clearly related to the scallop feeding and reproductive cycles (Robinson et al., 1981).
The major changes observed in the digestive gland are those related to the availability of food. They are expressed as an increase in the total lipid content, and especially, but not exclusively, in the proportion of TG (Table 2 and Fig. l a). The digestive gland in scallops is the only organ truly engaged in storing lipid reserves (Giese, 1966), and high but variable proportions of TG were observed in this organ all year round. Although minimum levels of total lipids were measured in winter animals, neutral lipids, mostly TG, still accounted for at least one-half of the total. The persistence of a substantial amount of lipid material during the winter is consistent with the high primary production of Georges Bank (e.g. Riley, 1982). Georges Bank is characterized by high levels of photosynthetic primary production, not only during the typical spring and fall algal blooms, but also during the summer months. The primary production during the winter reaches a minimum, but its actual level is still above average values for other and comparable ecosystems (O'Reilly et al., 1987). Levels of primary production on Georges Bank varied sub- stantially from sub-area to sub-area, being particu- larly high in the shallowest sectors (O'Reilly et al., 1987). Estimated annual primary production ranges between 455 and 310g carbon/m 2 between the 60 and 100 m isobaths. These areas of high productivity coincide approximately with the location of the densest scallop populations (Bourne, 1964).
The gonad of the female scallop is also charac- terized by a high and variable lipid content. These changes are associated with the sexual maturation taking place during spring, summer and early fall, and with the final release of the mature eggs in late fall (Robinson et al., 1981). The lipid content of the female gonad remained high during spring and summer, reaching a maximum during the prespawn- ing period (late summer and early fall). The female gonad of postspawning animals exhibited a dramatic depletion of neutral lipids, and the total lipid content (mostly structural phospholipids) remained close to the baseline of 1% of the wet weight (Fig. lb).
Comparable changes in the lipid content and TG accumulation of the female gonad of a number of different scallop species have been documented (Ansell, 1974; Pollero et al., 1979; Taylor and Venn, 1979; Lubet et al., 1987; Tsuji and Nishida, 1988). Although the timing of the major fluctuations is geographic and species related, the association of these changes with the animal sexual cycle is maintained.
The female gonad, and both male and female digestive glands, exhibited interesting patterns of seasonal lipid accumulation. Figure 1 clearly indi- cates the importance of the neutral lipid fraction in enhancing the lipid accumulation of an animal. Similar correlations have been illustrated between total lipid and wax ester reserves in calanoid copepods (Lee and Barnes, 1975; Lee et al., 1971). Surprisingly, a positive and significant correlation has also been found between the PL and the total lipid contents of the digestive gland in scallops (Fig. lc). That a similar correlation was found in female gonad lipids (Fig. ld) is less surprising, since the eggs of many animals are known to store large amounts of PL as a preparation for the imminent process of cell division. However, accumulation of PL in somatic tissues, such as liver or digestive organs, is not well documented in invertebrates.
Therefore, the role of the digestive gland in scallops seems to be extended to accumulation not only of neutral lipids as energy storage, but also of structural PL, which may be further exported to the develop- ing gonads. In agreement with these observations, a radiotracer experiment using the scallop Chlamys hericia showed an initial incorporation of ~4C-lipids into the digestive gland, followed by an increase of activity in the gonads and, simultaneously, decreased activity in the digestive gland (Vassallo, 1973).
Sterols are necessary building blocks for the formation of new cell membranes during the process of gamete proliferation and growth. Nevertheless, no significant accumulation of this lipid class could be demonstrated in either digestive gland or female gonad of the sea scallops (Fig. 2e and 0. This is consistent with the fact that sterols are not used as energy reserves, and because their concentration in cellular membranes is, on the average, one order of magnitude smaller than that of PL.
Acknowledgements--We are grateful to Scotia Trawlers (Lunenburg, Nova Scotia) for providing the scallop, samples. This work was supported by grants from the Natural Sciences and Engineering Research Council of Canada to P. J. Wangersky and R. G. Ackman.

650 GUILLERMO E. NAPOLITANO and ROBERT G, ACKMAN
REFERENCES
Ackman R. G. (1981) Flame ionization detection applied to thin-layer chromatography on coated quartz rods. In Methods in Enzymology (Edited by Lowenstein J. M.), Vol. 74D, pp. 205-252. Academic Press, London.
Ansell A. D. (1974) Seasonal changes in biochemical com- position of the bivalve Chlamys septemradiata from the Clyde Sea area. Mar. Biol. 25, 85-99.
Bligh E. G. and Dyer D. J. (1959) A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911-917.
Bonnet J. C., Sidwell V. G. and Zook E. G. (1974) Chemical and nutritive values of several fresh and canned finfish, crustaceans, and mollusks. Part II. Fatty acid composition. Mar. Fish. Rev. 36, 8-14.
Bourne N. (1964) Scallops and the offshore fishery of the maritimes. Bull. Fish. Res. Bd Can. No. 145.
Exler J. and Weihrauch J. L. (1977) Comprehensive evaluation of fatty acids in food. J. Am. Diet. Assoc. 71, 518-531.
Giese A. C. (1966) Lipids in the economy of marine invertebrates. Physiol. Rev. 46, 244-298.
Holland D. L. (1978) Lipid reserves and energy metab- olism in the larvae of benthic marine invertebrates. In Biochemical and Biophysical Perspectives in Marine Biology (Edited by Malins D. C. and Sargent J. R.), pp. 85-123. Academic Press, New York.
Idler, D. R., Tamura, T. and Wainai, T. (1964) Seasonal variation in the sterol, fat and unsaponifiable components in scallop muscle. J. Fish. Res. Bd Can. 21, 1035-1042.
Jarzebski A., Wenne R. and Habermehl G. (1986) Anatomical distribution of lipids and sterols in Macoma baltica (L.). Comp. Biochem. Physiol. 85B, 135-137.
Joseph J. D. (1982) Lipid composition of marine and estuarine invertebrates. Part II. Mollusca. Prog. Lipid Res. 21, 109-153.
Joseph J. D. (1989) Marine invertebrates. In Marine Bio- genic Lipids, Fats and Oils (Edited by Ackman R. G.), pp. 49-143. CRC Press, Boca Raton.
King I,, Childs M. T., Dorsett C., Ostrander J. G. and Monsen E. R. (1990) Shellfish: Proximate composition, fatty acids, and sterols. J. Am. Diet. Assoc. 90, 677-685.
Krzeczkowski R. A., Tenney R. D. and Hayes M. L. (1972) Fatty acid content and proximate analysis of bay, calico, sea and weathervane scallop adductor muscle. J. Food Sci. 37, 300-301.
Krzynowek, J. and Murphy, P. (1987) Proximate compo- sition, energy, fatty acid, sodium, and cholesterol content in finfish, shellfish and their products. NOAA Tech. Rep. No. 55. U.S. Dept Of Commerce.
Lee R. F. and Barnes A. T. (1975) Lipids in the mesopelagic copepod Gaussia princeps. Wax ester utilization during starvation. Comp. Biochem. Physiol. 52B, 265-268.
Lee R. F., Nevenzel J. C. and Paffenhofer G. A. (1971) Importance of wax esters and other lipids in the marine food chain: phytoplankton and copepods. Mar. Biol. 9, 99-108.
Levanidov I. P. and Zakharova V. P. (1968) The chemical composition of food molluscs and echinoderms of the Sakhalin region. Izvestiya TNIRO 65, 221-230.
Lubet P., Besnard J. Y., Feveris R. and Robbins I. (1987) Physiologie de la reproduction de la coquille Saint-Jacques (Pecten maximus L.). Oceanis 13, 265-290.
Napolitano G. E., Ratnayake W. M. N. and Ackman R. G. (1988) Fatty acid components of larval Ostrea edulis (L.): importance of triacylglycerols as a fatty acid reserve. Comp. Biochem. Physiol. 90B, 875-883.
O'Reilly J. E., Evans-Zetin C. and Busch D. A. (1987) Primary production. In Georges Bank (Edited by Backus R. H. and Bourne D. W.), Chap. 21. MIT Press, Cambridge.
Pollero R. J., Re M. E. and Brenner R. R. (1979) Seasonal changes in the lipids of the mollusc Chlamys tehuelcha. Comp. Biochem. Physiol. 64A, 257 263.
Posgay, J. A. (1957) The range of the sea scallop. Nautilus 71, 55-57.
Riley, G. A. (1982) Biological Processes in Georges Bank. In Georges Bank: Past, Present and Future of a Marine Environment (Edited by McLeod G. C. and Prescott J. H.), pp. 61-76. Westview Press, Boulder.
Robinson W. E., Wehling W. E., Morse P. and McLeod G. C. (1981) Seasonal changes in soft-body component indices and energy reserves in the Atlantic deep-sea scallop Placopecten magellanicus. Fish. Bull. 79, 449-458.
Sargent J. R. (1976) The structure, metabolism and func- tion of lipids in marine organisms. In Biochemical and Biophysical Perspectives in Marine Biology (Edited by Malins D. and Sargent J. R.), Vol. 3, pp. 149-212. Academic Press, New York.
Shafee M. S. (1981) Seasonal changes in the biochemical composition and calorific content of the black scallop Chlamys varia (L.) from Lanveoc, Bay of Brest. Oceanol. Acta 4, 331-341.
Sidwell V. D., Bonnet J. C. and Zook E. G. (1973) Chemical and nutritive values of several fresh and canned finfish, crustaceans, and molluscs. Part 1: Proximate composition, calcium, and phosphorus. Mar. Fish. Rev. 35, 16-19.
Sipos J. C. and Ackman R. G. (1978) Automatic and rapid quantitative analysis of lipids with Chromarods. J. chromatogr. Sci. 16, 443-447.
Taylor A. C. and Venn T. J. (1979) Seasonal variation in weight and biochemical composition of the tissues of the queen scallop, Chlamys opercularis, from the Clyde Sea area. J. Mar. Biol. Ass. U.K. 59, 605-621.
Torry Research Station. (1989) Yield and nutritional value of the commercially more important fish species. FAO Fish. Tech. Paper 309, 187 pp.
Tsuji K. and Nishida H. (1988) Seasonal Changes in biochemical composition of the scallop, Patinopecten yessonensis in Nemuro Bay, Hokkaido. Sci. Rep. Hokkaido Fish. Exp. Station 31, 27-54.
Vassallo M. T. (1973) Lipid storage and transfer in the scallop Chlamys hericia Gould. Comp. Biochem. Physiol. 44A, 1169-1175.





























