Embed Size (px)
Citation preview

Anatoly B. Kolomeisky
Department of Chemistry
Growth Dynamics of Cytoskeleton Proteins: Multi-Scale Theoretical
Analysis

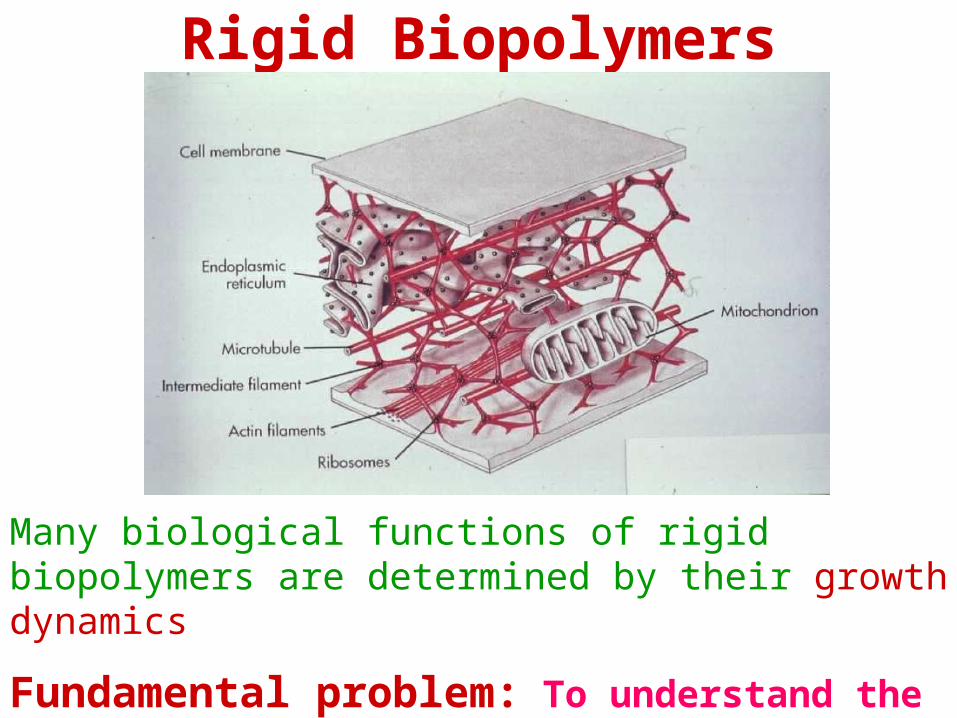
RIGID BIOPOLYMERS
actin filaments
microtubules
intermediate filaments
Rigid Biopolymers
Many biological functions of rigid biopolymers are determined by their growth dynamics
Fundamental problem: To understand the mechanisms of growth and coupling to biological processes
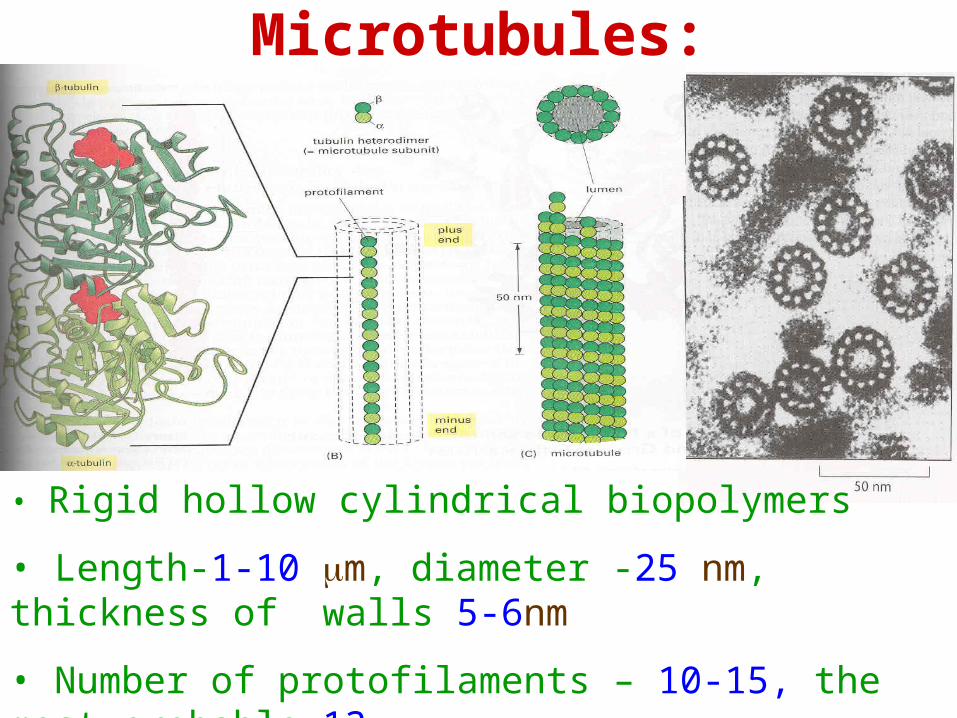
Microtubules:
• Rigid hollow cylindrical biopolymers
• Length-1-10 m, diameter -25 nm, thickness of walls 5-6nm
• Number of protofilaments – 10-15, the most probable 13

Microtubules
• 3-start helical structure with a seam
• -tubulin-GTP subunit
• Size of the dimer subunit 8x4x4 nm
• Polar structure
• Plus ends grow faster than minus ends
• Polymerization produces forces 1-20pN
• Important biological functions:
cell division, cell motility and
cellular transport

Microtubules: Dynamic InstabilityMicrotubules exist in two dynamic phases: growing or shrinking
Dynamic instability – non-equilibrium phenomenon
Understanding of dynamic instability is not complete

Single Microtubules: Experiments
Force generation by single microtubules:
1) Video microscopy
2) Optical trap spectrometry
Buckling shapes forces
Dogterom et al. Appl. Phys. 75, 331 (2002)
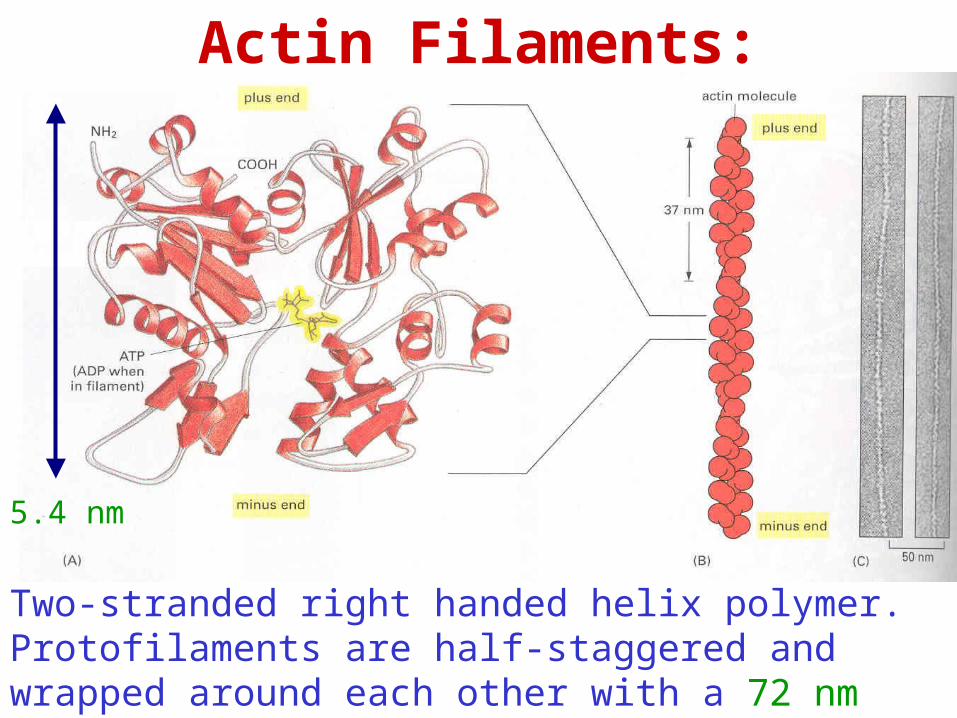
Actin Filaments:
5.4 nm
Two-stranded right handed helix polymer. Protofilaments are half-staggered and wrapped around each other with a 72 nm period.

Single Actin Filaments: Experiments
Fujiwara et al., Nature Cell Biology, 4, 666-673 (2002)
Direct observation of single actin polymerization/depolymerization processes from fluorescently labeled molecules
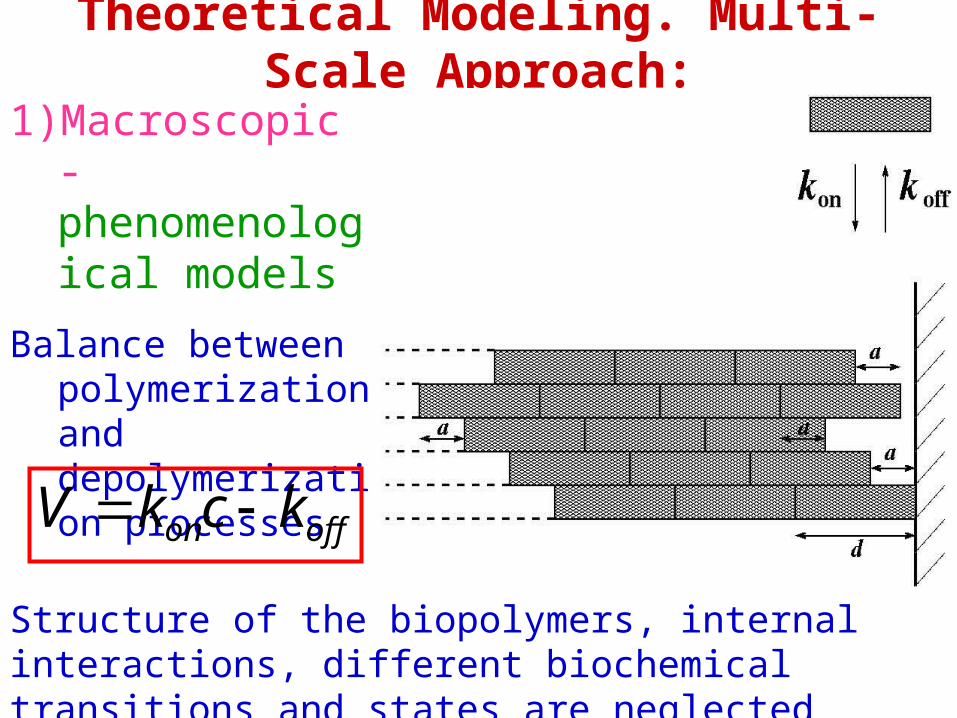
Theoretical Modeling. Multi-Scale Approach:
1) Macroscopic - phenomenological models
Balance between polymerization and depolymerization processes
offon kckV
Structure of the biopolymers, internal interactions, different biochemical transitions and states are neglected

1) Macroscopic Approach: Wrong!
)]/exp(
)/exp([)(
1
10
TkFdk
TkFdkdFV
Boff
Bon
V – mean velocity; F- load ; d0=d/n- mean increase in length; d=8.2 nm – dimer size; n=13 – number of protofilaments;
Fd1- the most probable work needed to add a single tubulin dimer against the load F
and load-distribution factors
For microtubules – phenomenological model

Load-Distribution Factors:Effect of an external load F:
TkFdoffoffoff
TkFdononon ekFkkekFkk B1B1 //
)0()(,)0()(
and load distribution factors 1
activation barrier aE
1Fd
1Fd
F=0F >0
j
TkEon
aek B/

Microtubules: Phenomenological Model
])/exp([)( 10 offBon kTkFdkdFV
Phenomenological (Thermodynamic) Theory: Dogterom and Yurke (Science,1997)
Assumption: d0=d1= d/13=0.63 nmkon= 1791 min-1
koff= -127 min-1Fit of experimental data
Unphysical! Chemical rates are always >0!
Phenomenological theory fails!
Stalling (V=0) is not an equilibrium

Microtubules: Theoretical Description
Fisher and Kolomeisky (Biophys. J. 2001)
)]/exp()/exp([)( 110 TkFdkTkFdkdFV BoffBon
No assumption of thermodynamic equilibrium at stalling (when V=0)
d1=d – complex parameter
Predictions:
off
onBS k
k
d
TkF ln
1
Stall force
kon= 1887 min-1
koff= 0.33 min-1
=0.22; unrealistic
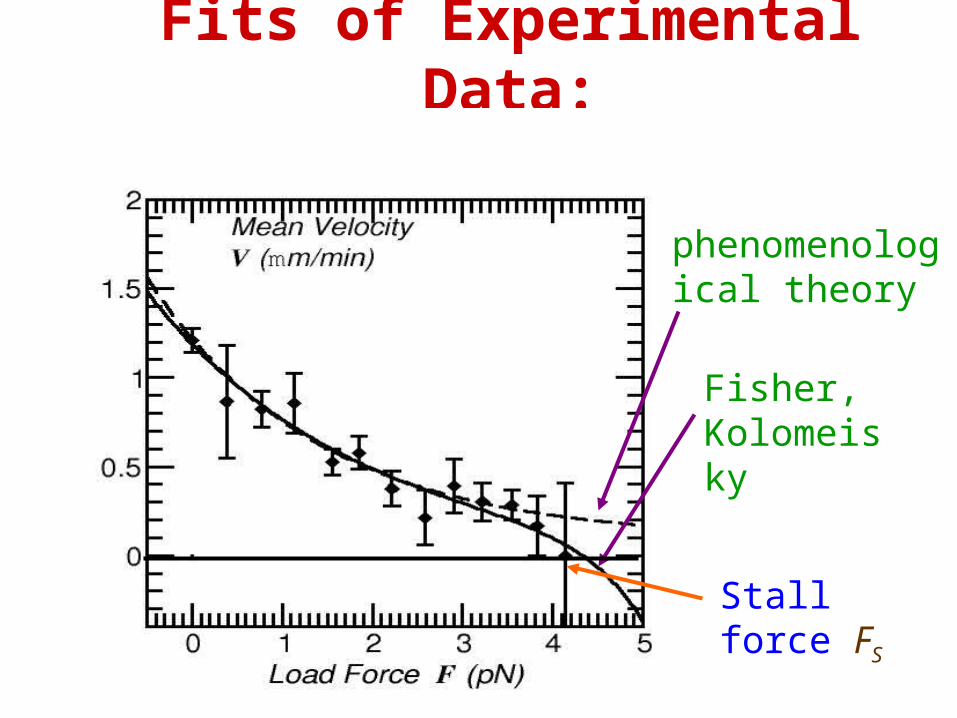
Fits of Experimental Data:
phenomenological theory
Fisher, Kolomeisky
Stall force FS

Theoretical Modeling. Multi-Scale Approach:
2) Microscopic approach – full atomistic simulations
Currently – do not exist!
Protein Data Bank:
- tubulin subunit
More than 10000 atoms!!!

Theoretical Modeling. Multi-Scale Approach:
3) Mesoscopic Approach:
Takes into account some structural and biochemical properties
Polymerization Ratchet Models:
Thermal fluctuations create gaps for inserting monomers
Mogilner and Oster, Biophys. J. 71, 3030 (1996)

Rigid Biopolymers: Theoretical Problems
Phenomenological models and polymer ratchet models cannot describe the growth dynamics, especially under external forces and concentration dependence
Main problem:
Geometrical structure of growing biopolymers, monomer-monomer interactions and biochemical transitions are neglected
Our approach: discrete stochastic models with lateral interactions, correct geometry of biopolymer’s tips and biochemical transitions

Rigid Biopolymers: Theoretical Description
Our Goal:To develop the simplest theoretical picture which will take into account the geometry and polymer lattice interactions
Problem:
Infinite number of polymer configurations!

Microtubules: Growth Mechanism
fast uj
slow ui
Inhomogeneity in growth rates
Different rates of association and dissociation for different protofilaments

Microtubules: Theoretical Description
Approximate theory:
One-Layer Model
Idea: only few configurations are relevant for growth dynamics

Microtubules: One-Layer ModelAssumption: Only configurations of microtubules with distances from the leading protofilament tip less than d allowed
There are N such configurations
N-number of protofilaments
Explicit expressions for mean growth velocity, V, and for the dispersion, D, for any N and any geometry in terms of {uj,wj}

Microtubules: One-Layer ModelHow good is the
approximation?
1) Comparison with the full dynamic solution for the specific value of N
2) Monte Carlo simulations
For N=2 the full dynamic solution exists;
Relevant for actin filaments

Microtubules: Theoretical Description
Compare with N = 2 model
Full dynamics : 4 different types of transitions
u
w
da
]2
exp[Tk
ggg
w
u
B
imvh
gv -free energy of creating head-to-tail bond
gh – free energy of creating lateral bond
1)
gim- free energy of monomer immobilizing
=a/d-fraction of created or broken lateral bond

Microtubules: Theoretical Description
2) u1-
w
])1(2
exp[1
1
Tk
ggg
w
u
B
imvh
u1
w1
3)
]2
exp[1
1
Tk
ggg
w
u
B
imvh

Microtubules: Theoretical Description
4)
u0
w0
]exp[0
0
Tk
gg
w
u
B
imv Define
]exp[Tk
g
B
h
2
0
0
1
1
)1(2
0
0
1
1
2
0
0
w
u
w
u
w
u
w
u
w
u
w
u
Assumption: thermodynamics ~ kinetics
)1
11)((
2)(
/)2(
/0
0 dda
dawu
dexactV
Growth velocity
1,,1,0 where
, ,
i
wwuu iii

Microtubules: One-Layer Model
N=2 - only 2 configurations
u+w
u+w
)1
11)((
2)(
1)()-(
)12(0
0
)12(0
0
w
ud
exactV
wudlayeroneVCompare
growth rates:
)()( For layeroneVexactV
Realistic values: gh kBT,
]exp[Tk
g
B
h

Ratio of Growth Velocities
for different shifts
stronger lateral interactions
Ratio of exact and approximate velocities

Comparison with Monte-Carlo Simulations
]exp[Tk
g
B
h
gh-lateral interactions between the monomers in rigid biopolymers
~3-7 kBT
Son, Orkoulas and Kolomeisky, J. Chem. Phys. (2005) in press
N=13 protofilaments

Effect of External Forces
Effect of an external load F:TkadF
jjjTkadF
jjjjjjj ewFwweuFuu BB /)(/)( )0()(,)0()(
jj and load-distribution factors 1
jj
activation barrier aE
1Fd
1Fd
F=0F >0
j
TkEj
aeu B/

Comparison with Phenomenological Models
Concentration dependence – nonlinear!
critical concentration
one-layer model with N=13
Phenomenological model
offon kckV

Microtubule Growth: Experiments
Non-linear dependence!
Biochemistry, 26, 4428-4437 (1997)

Description of Experiments on Microtubules
force-velocity curve
Stall force Fs=5.6pN
Bond energies can be estimated
Phenomenol. theory:
2 parameters
Our theory:
3 parameters-
u0, w0,
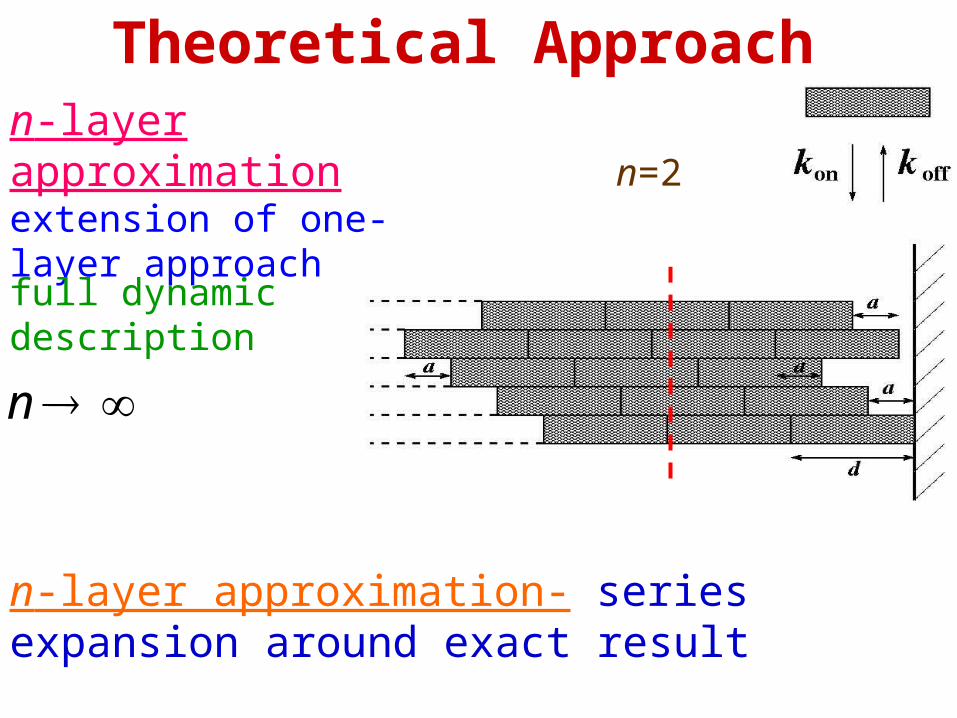
Theoretical Approachn-layer approximation extension of one-layer approach
n=2
n
full dynamic description
n-layer approximation- series expansion around exact result
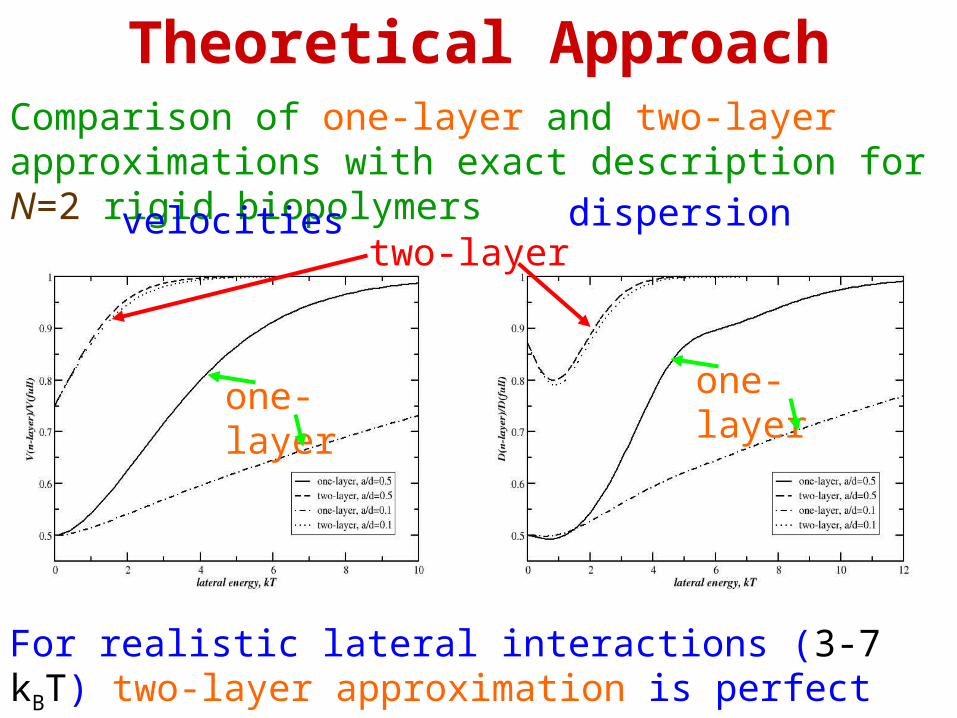
Theoretical ApproachComparison of one-layer and two-layer approximations with exact description for N=2 rigid biopolymers
velocities dispersion
For realistic lateral interactions (3-7 kBT) two-layer approximation is perfect
one-layer one-layer
two-layer

Theoretical ApproachKinetic explanations for n-layer approximations
Full kinetic scheme for N=2 rigid biopolymers
(k,m) – polymer configuration with k monomers in the 1-st protofilament, and m monomers in the 2-nd

Theoretical Approach
one-layer model
two-layer model
Kinetic justifications for n-layer approximations

Actin Filaments:
Fujiwara et al., Nature Cell Biology, 4, 666 (2002)
)(2
1 2offon kckaD a =2.7 nm
Experimental observations: large length fluctuations in actin filaments in stationary phase. D(exp)/D(calc) =35-40!!!
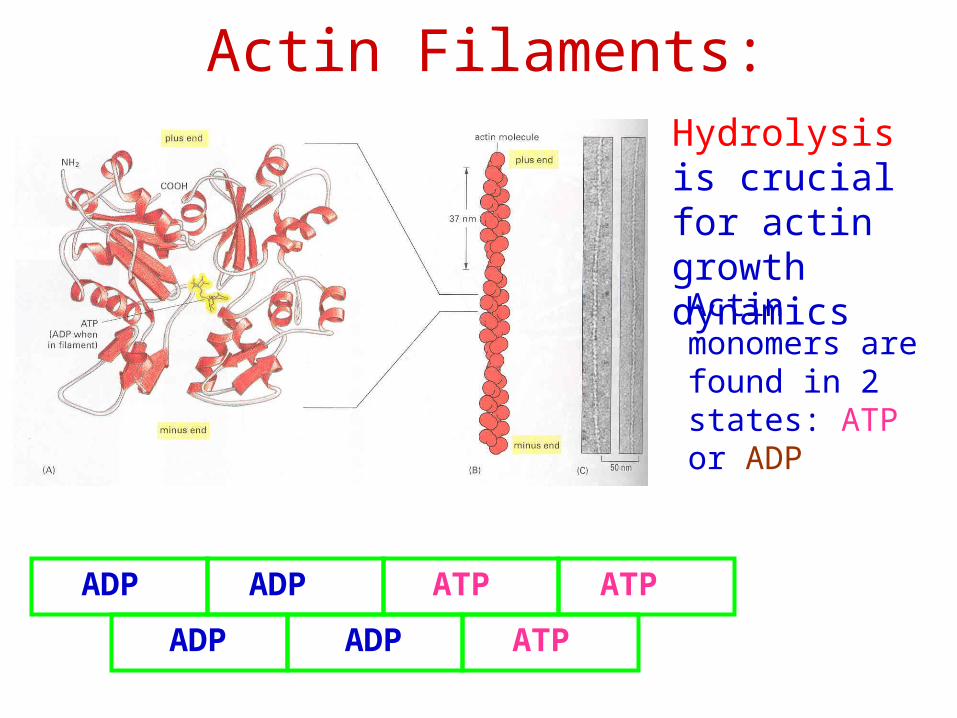
Actin Filaments:Hydrolysis is crucial for actin growth dynamics
ADP ADP
ADP ADP
ATP ATP
ATP
Actin monomers are found in 2 states: ATP or ADP

Actin Filaments. Hydrolysis1) Random mechanism
1) Sequential (vectorial)
ADP
ADP
ADP
ADP
ADP
ADP
ADP ADP
ATP ATP
ATP
ATP ATP
ATP
One interface between hydrolyzed and unhydrolyzed segments
Many interfaces between hydrolyzed and unhydrolyzed segments
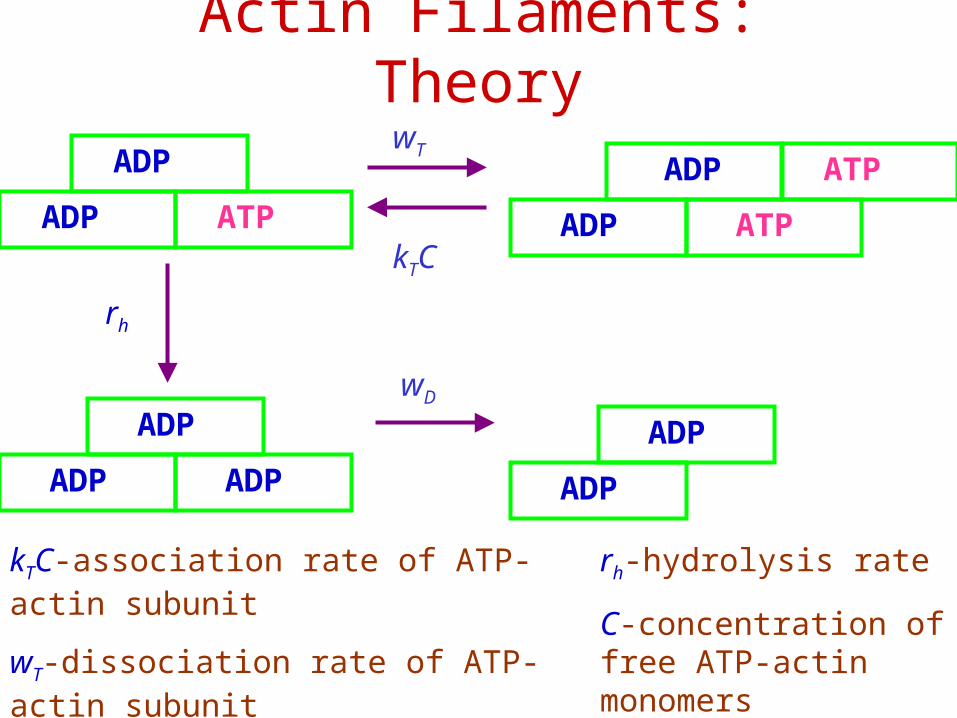
Actin Filaments: Theory
kTC
wT
ADP ATP
ADP
ADP
ADP ADP ADP
ADP
ADP
ADP
ATP
ATP
rh
wD
kTC-association rate of ATP-actin subunit
wT-dissociation rate of ATP-actin subunit
wD-dissociation rate of ADP-actin subunit
rh-hydrolysis rate
C-concentration of free ATP-actin monomers

Actin Filaments: Theory
kTC
wT
ADP ATP
ADP
ADP
ADP ADP ADP
ADP
ADP
ADP
ATP
ATP
rh
wD
IDEA: large fluctuations of length at low concentrations due to dissociation of exposed ADP-actin monomers
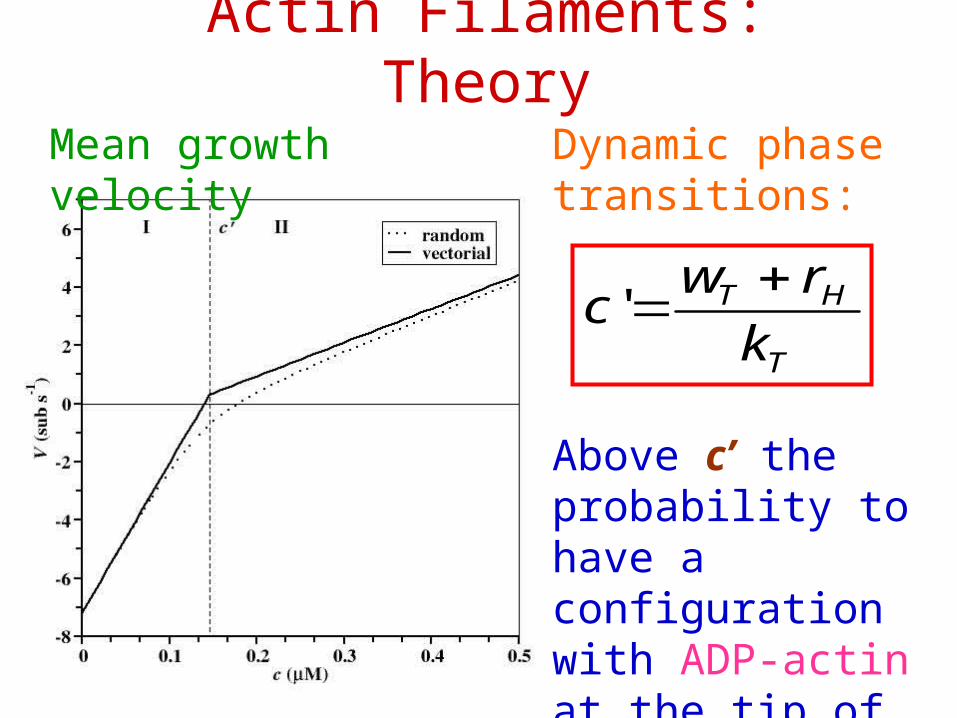
Actin Filaments: Theory
Mean growth velocity Dynamic phase transitions:
T
HT
k
rwc
'
Above c’ the probability to have a configuration with ADP-actin at the tip of the filament is zero

Actin Filaments: TheoryMean dispersion
c’
Large length fluctuations at c’ because of ATP-actin dissociation/association and ADP-actin dissociation
D(exp)=25-31 sub2s-1
D(theory)=31.6 sub2s-1

CONCLUSIONS
• Multi-scale analysis of the growth of rigid biopolymers is presented
• Mesoscopic models that accounts for geometry, lattice interactions and biochemical transitions are developed
• All dynamic properties can be calculated explicitly
• n-layer approximations of growth dynamics are presented
• Hydrolysis stimulates large length fluctuations in actin filaments at low concentrations

Acknowledgements• Dr. E. Stukalin (Rice University) and Prof. M.E.
Fisher (U of Maryland)• Financial support: NSF, Welch Foundation,
Dreyfus Foundation, Sloan Foundation• Publications:1) Kolomeisky and Fisher, Biophys. J., 80, 149
(2001)2) Stukalin and Kolomeisky, J. Chem. Phys., 121,
1097 (2004).3) Stukalin and Kolomeisky, J. Chem. Phys., 122,
104903 (2005).