Embed Size (px)
Citation preview

1
Anaplastic Lymphoma Kinase (ALK) is Activated Through the Pleiotrophin (PTN)/ReceptorProtein Tyrosine Phosphatase (RPTP)b/z Signaling Pathway: An "Alternative Mechanism of
Receptor Tyrosine Kinase (RTK) Activation"P. Perez-Pinera1,¶, W. Zhang1, ¶, Y. Chang1, J.A. Vega† and T.F. Deuel1*
1 The Scripps Research Institute. La Jolla. California, † Universidad San Pablo-CEU, Madrid, Spain, ¶
These authors contributed equally to this workRunning title: Pleiotrophin activates ALK through RPTPb/z
* Address correspondence to: Thomas F. Deuel, M.D. The Scripps Research Institute. 10550 North TorreyPines Road. La Jolla, CA 92037. Tel. 858.784.7923. Fax. 858.784.7977. Email: [email protected]
Anaplastic Lymphoma Kinase (ALK) isa receptor tyrosine kinase (RTK) firstdiscovered as the constitutively activenucleophosmin (NPM)-ALK oncoprotein inanaplastic large cell lymphomas (ALCL). Fulllength ALK has a critical role in normaldevelopment and differentiation. Activated full-length ALK also is found in different malignantcancers. Nevertheless, the ligand to activateALK remained unknown, until recently, whenALK was proposed to be the physiologicalreceptor of the cytokine pleiotrophin (PTN,P t n ). However, earlier studies haddemonstrated the Receptor Protein TyrosinePhosphatase (RPTP)b/z is a physiological PTNreceptor. We now demonstrate thatphosphorylation of ALK in PTN-stimulatedcells is mediated through the PTN/RPTPb/zsignaling pathway. ALK is phosphorylatedindependently of a direct interaction of PTNwith ALK. The data thus support a uniquemodel of ALK activation. In cells notstimulated by PTN, RPTPb/z dephosphorylatesALK at the site(s) in ALK that is undergoingautophosphorylation through autoactivation. Incontrast, when RPTPb/z is inactivated in PTN-stimulated cells, the sites that areautophosphorylated in ALK no longer can bedephosphorylated by RPTPb /z and thus,autoactivation and tyrosine phosphorylation ofALK rapidly increase. The data indicate thePTN/RPTPb/z signaling pathway is a criticalregulator of the steady state levels of tyrosinephosphorylation and activation of ALK; thedata support the conclusion that ALKphosphorylation and activation in PTN-stimulated cells are increased through a unique"Alternative Mechanism of Receptor TyrosineKinase (RTK) Activation".
Anaplastic Lymphoma Kinase (ALK) is areceptor-type transmembrane tyrosine kinase(RTK) of the insulin receptor superfamily (1).ALK was first identified as the oncogenicchimeric nucleophosmin (NPM)-ALK fusionprotein that results from the (2;5)(p23;q35)chromosomal translocation in anaplastic large celllymphomas (ALCL) (2-4); NPM-ALK containsthe N-terminal domain of NPM fused with the C-terminal cytoplasmic (catalytic) domain of ALKand is constitutively active. Wild type ALK isrequired for normal embryonic development (5)and in the determination of cell survival and cellfate (6); however, it is also involved in thepathogenesis of different cancers. Nevertheless,the mechanisms through which ALK is activatedin these different contexts are not clear.
ALK is known to be activated throughautoactivation in vitro, as is the case with otherreceptor tyrosine kinases (RTKs) (7,8).Autoactivation is the consequence of ligandenforced dimerization of RTKs that results frominduced conformational changes in the active siteand leads to autophosphorylation which furtherenhances catalytic activity and introduces sites fordifferent adapter proteins to engage and activatedownstream signaling pathways. However, theligand to enforce ALK dimerization andautoactivation of ALK was unknown and thusALK has been regarded as an orphan receptor.
Recently, ALK was proposed to be thephysiological receptor of pleiotrophin (PTN, Ptn)(9-11). Pleiotrophin is a 136 amino acid cytokinethat has diverse functions in both normal cellgrowth and differentiation and in growth ofdifferent malignant cells (12,13). The proposal thatALK is the receptor for PTN was surprising, sinceearlier studies had demonstrated that the ReceptorProtein Tyrosine Phosphatase (RPTP)b/ z is afunctional receptor of PTN (14). In those studies,
http://www.jbc.org/cgi/doi/10.1074/jbc.M704505200The latest version is at JBC Papers in Press. Published on August 6, 2007 as Manuscript M704505200
Copyright 2007 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
PTN was shown to signal through enforceddimerization of RPTPb/z which, in turn, results ina loss of the RPTPb /z catalytic tyrosinephosphatase activity. The PTN-inducedinactivation of RPTPb/z in turn leads to increasedtyrosine phosphorylation of each of the substratesof RPTPb/z (14-19), since the sites that normallyare dephosphorylated by RPTPb/z continue to bephosphorylated by constitutive tyrosine kinaseswhen RPTPb/z is inactivated in PTN-stimulatedcells.
The proposal that ALK is thephysiological receptor for PTN also wassurprising, since, in Drosophila, Jelly Belly (Jeb)was identified as a ligand of ALK (20,21). Jeb hasno apparent sequence or structural relationshipwith PTN and, the human genome databases donot contain a human homolog of Jeb. Furthermore,neither Miple1, the Drosophila homolog of Ptn,nor Miple2, the Drosophila homolog of midkine(Mk), which also signals through RPTPb/z (22),were found to be spatially localized with DAlk nortemporally co-expressed with DAlk duringDrosophila development. Furthermore, in yetanother study, Moog-Lutz et al. (23) developedtwo anti-ALK monoclonal antibodies each ofwhich effectively activated ALK. However, underthe same conditions, PTN failed to activate ALK;thus, in this study also, the data did not supportthat ALK is a physiological receptor of PTN.
The conflicting views of ALK is thephysiological receptor of PTN have now led us totest the possibility that ALK is activated by PTNthrough an alternative mechanism. In the presentstudies, the data demonstrate that PTN stimulatestyrosine phosphorylation of ALK through thePTN/RPTPb/z signaling pathway. It stimulatesphosphorylation of ALK at the same site that isautophosphorylated when ALK is autoactivated invitro; this site is also recognized by RPTPb/z anddephosphorylated by RPTPb/z. The data suggestthe PTN/RPTPb/z signaling pathway functions toactivate ALK through an “An AlternativeMechanism of Receptor Tyrosine KinaseActivation” that is independent of a directinteraction of PTN with RPTPb/z.
EXPERIMENTAL PROCEDURESCell lines - HeLa, U87MG, U373, and
MCF-7 cells were obtained from American Tissue
Collection Center (ATCC) and grown in DMEMor MEM supplemented with 10% fetal bovineserum (FBS) and 1% penicillin/streptomycin at37°C in a 5% CO2 atmosphere. HeLa cells expressRPTPb/z and low levels of ALK, U87MG cellsexpress RPTPb/z and ALK, U373 cells expressRPTPb/z but not ALK, and, MCF-7 cells expressneither ALK nor RPTPb/z.
Reagents – Rabbit Anti-ALK antibodies(catalog number 51-3900) were obtained fromZymed (currently Invitrogen, Carlsbad, CA),mouse anti-phosphotyrosine antibodies (catalognumber 05-321) were obtained from Upstate(Charlottesville, VA), mouse anti-RPTPb /zantibodies (catalog number R20720) wereobtained from BD Biosciences (San Diego, CA),PTN was obtained from R&D Systems(Minneapolis, MN), and HRP-conjugated anti-mouse and anti-rabbit antibodies were obtainedfrom San Cruz Biotechnology (Santa Cruz, CA).
Plasmids, Transfections - The EpidermalGrowth Factor Receptor (EGFR)/RPTPb /zchimeric protein was constructed by fusing thesequences to encode the extracellular andtransmembrane domains of EGFR (amino acids 1to 671) with the cytoplasmic domain of RPTPb/z(amino acids 1662 to 2315), which then wereinserted into the vector pcDNA3.1.
Human full-length ALK cDNA (GenBankAccession Number NM_004304), human full-length RPTPb/z cDNA (GenBank AccessionNumber, NM_002851), and a truncated ALKcDNA to encode the 8 membrane proximalextracellular amino acids and the intacttransmembrane and intracellular domains of ALK(amino acids 1027 to 1620) with the IgGk signalpeptide sequences were inserted in the pcDNA3.1vector. Transfections were performed using theFugene 6 transfection reagent (Roche Diagnostics,Alameda, CA) following the manufacturer'sinstructions.
The vectors containing shRNA targetingRPTPb/z and a control empty vector wereobtained from Open Biosystems (Huntsville, AL)[catalog numbers RHS1764-9687266 (shRNARPTPb/z 1) and RHS1764-9218363 (shRNARPTPb/z 2)] and delivered into cells using Arrest-In transfection reagent following themanufacturer's recommendations. Ninety-six hoursafter transfection, the levels of expression of
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
RPTPb/z were analyzed using Western blots andreal time RT-PCR as described before (24).
The vector pC4-Fv1E to enforcedimerization of RPTPb/z through AP20187 was akind gift from ARIAD Pharmaceuticals, Inc.(Cambridge, MA). The cDNA sequence encodingthe intracellular domain of RPTPb/z was insertedin frame with the Fv fragment of FKBP12.
RPTPb/z D1, D1 (C1932S), D1 (D1900A)GST "Capture" – Proteins recognized by the activesite (D1) domain of RPTPb/z (residues 1663-2034) in lysates of HeLa cells transfected withfull-length ALK were captured by GST-RPTPb/zD1, GST-RPTPb/z inactivated D1 (C1932S), andGST-RPTPb/z "substrate trap" D1 (D1900A) asdescribed previously (18) and analyzed in Westernblots probed with anti-ALK antibodies and anti-GST antibodies.
Dephosphorylation by RPTPb/z of ALKphosphorylated in tyrosine - MCF-7-EGFR/RPTPb/z cells which express very limitedlevels of EGFR into which EGFR/RPTPb/z wasintroduced were stimulated with EGF 150 ng/ml(R&D Systems, Minneapolis, MN) for 1 minute orwith sodium pervanadate 20 mg/ml for 20 minutes.Cell lysates were prepared, immunoprecipitatedwith ant i -ALK ant ibodies , and theimmunoprecipitates were incubated with GST-RPTPb/z D1 or GST-RPTPb /z (C1932S) andanalyzed by scanning densitometry of Westernblots probed with anti-phosphotyrosine antibodiesas described before (18).
In vitro kinase assays – ALK (Millipore,Billerica, MA, Catalog number 14-555) wasincubated with 200 mM ATP in a buffer containing25 mM Tris-HCl (pH 7.5), 5 mM b -glycerophosphate, 2 mM dithiothreitol (DTT), 0.1mM Na3VO4, and 10 mM MgCl2 for 20 minutes at37˚C. The reaction was stopped with boilingloading buffer and the samples were analyzed inWestern blots probed with anti-phosphotyrosineantibodies. As negative control, samples withoutATP were analyzed.
RESULTS
Pleiotrophin requires RPTPb/z to stimulatetyrosine phosphorylation of ALK.
As cited above, pleiotrophin signalsthrough enforced dimerization and inactivation of
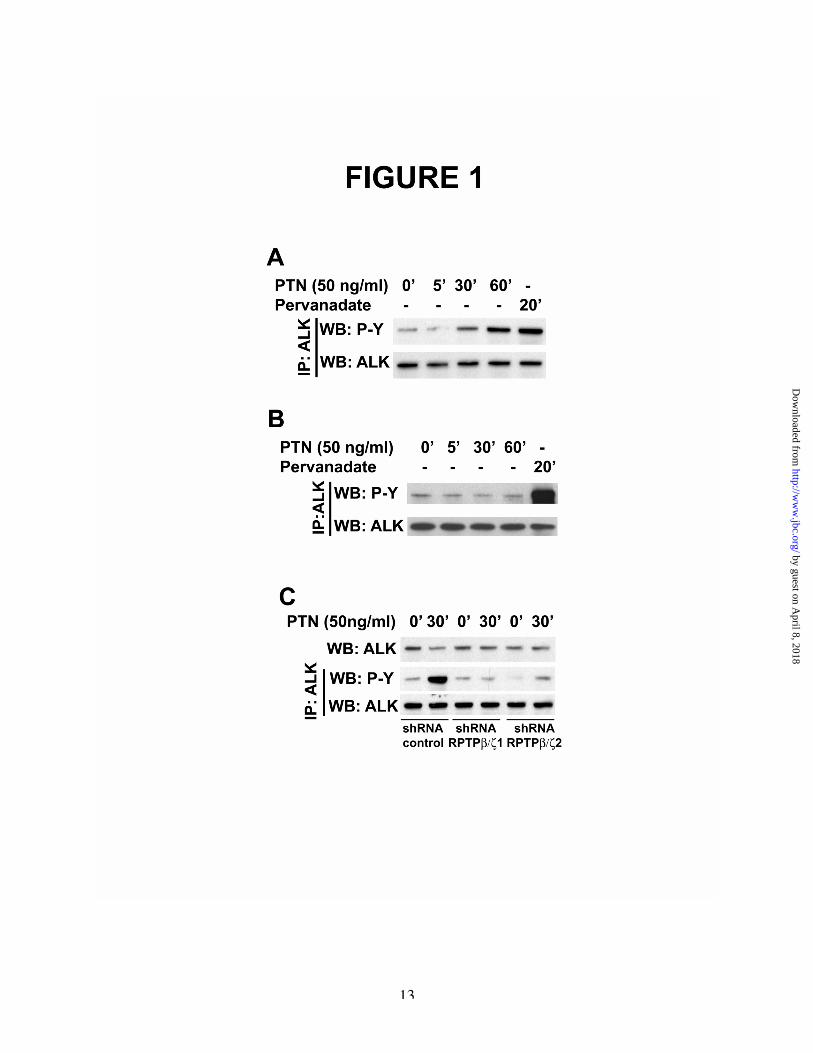
the cytoplasmic domain of RPTPb /z (14,25),leading to increased tyrosine phosphorylation ofeach of its substrates. To first determine if ALK isphosphorylated in PTN-stimulated cells throughthe PTN/RPTPb/z signaling pathway, MCF-7 cellsthat do not express ALK or RPTPb /z weretransfected with cDNAs to encode full-lengthALK and RPTPb/z. The cells were stimulatedwith PTN for 5, 30, and 60 minutes. Lysates fromstimulated and non-stimulated (control) cells werethen prepared and immunoprecipitated with anti-ALK antibodies. The levels of tyrosinephospho ry l a t i on o f ALK in t heimmunoprecipitates were measured by scanningdensitometry of Western blots probed with anti-phosphotyrosine antibodies. It was found thatbaseline levels of tyrosine phosphorylation ofALK in non-stimulated cells were low aspreviously shown (26-28) . Tyrosinephosphorylation of ALK was minimally increasedat 5 minutes and increased ~2.5-fold at 30 minutesand ~5.2-fold at 60 minutes after stimulation withPTN. MCF-7 cells were then transfected withALK alone but not with RPTPb/z. The cells werethen stimulated with PTN, and analyzed as above;it was found that PTN failed to increase tyrosinephosphorylation of ALK in MCF-7 cells that donot express RPTPb/z (compare Figure 1A, 1B).
The data demonstrate that PTN effectivelystimulates tyrosine phosphorylation of ALK inMCF-7 cells but, only when MCF-7 cells expressRPTPb/z; it is concluded that RPTPb/z is requiredfor PTN to stimulate tyrosine phosphorylation ofALK in PTN-stimulated cells. The data alsosupport the important conclusion that the level ofcatalytic activity of RPTPb/z regulates the steadystate levels of tyrosine phosphorylation of ALKwhen RPTPb /z is not inactivated in PTN-stimulated cells.
MCF-7 cells that express both ALK andRPTPb/z were then stimulated with the generalinhibitor of tyrosine phosphatases sodiumpervanadate for 20 minutes. Stimulation of theMCF-7 cells expressing both ALK and RPTPb/zwith sodium pervanadate increased levels oftyrosine phosphorylation of ALK by ~7-fold(Figure 1, A). MCF-7 cells that do not expressRPTPb/z were then stimulated with sodiumpervanadate; incubation of these cells with sodiumpervanadate also sharply increased tyrosine
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
phosphorylation of ALK (Compare Figure 1A,1B).
This data thus support the conclusion thattyrosine phosphatases other than RPTPb/z alsoregulate the steady state levels of tyrosinephosphorylation of ALK when their catalyticactivities are not inhibited by sodium pervanadate,presumably at tyrosine phosphorylation sites thatare different from the site(s) regulated through thePTN/RPTPb/z signaling pathway.
ShRNAs to “knock down” RPTPb/z preventPTN-stimulated tyrosine phosphorylation ofALK.
The requirement of RPTPb/z for PTN tostimulate tyrosine phosphorylation of ALK inPTN-stimulated cells was investigated further.Two shRNA’s to “knock-down” RPTPb/z wereexpressed in U87MG cells. U87MG cells expressboth endogenous RPTPb/z and endogenous ALK(29). Both the two shRNA’s were first shown toeffectively knock down expression of RPTPb/z; itwas found in these control experiments that thelevels of RPTPb/z mRNA were reduced ~66% incells expressing shRNA RPTPb/z-1 and ~73% incells expressing shRNA RPTPb/z-2, as measuredby qRT-PCR.
Pleiotrophin failed to increase tyrosinephosphorylation of ALK in the U87MG cells thatexpressed either of the two shRNA’s. However,PTN sharply increased tyrosine phosphorylation ofALK in U87MG cells into which a “scrambled”shRNA (control) was expressed (Figure 1C).
This data thus demonstrate that“knockdown” of RPTPb /z prevents PTN-stimulated tyrosine phosphorylation of ALK inPTN-stimulated U87MG cells; the data indicateendogenous RPTPb/z is required for PTN tostimulate tyrosine phosphorylation of ALK inU87MG cells that express both endogenous ALKand RPTPb/z.
Epidermal Growth Factor (EGF) stimulatesincreased tyrosine phosphorylation of ALK inMCF-7 cells through the EGF Receptor(EGFR)/RPTPb/z chimeric receptor.
To further pursue the requirement ofenforced dimerization of the cytoplasmic domainof RPTPb /z for PTN to stimulate tyrosinephosphorylation of ALK, a chimeric
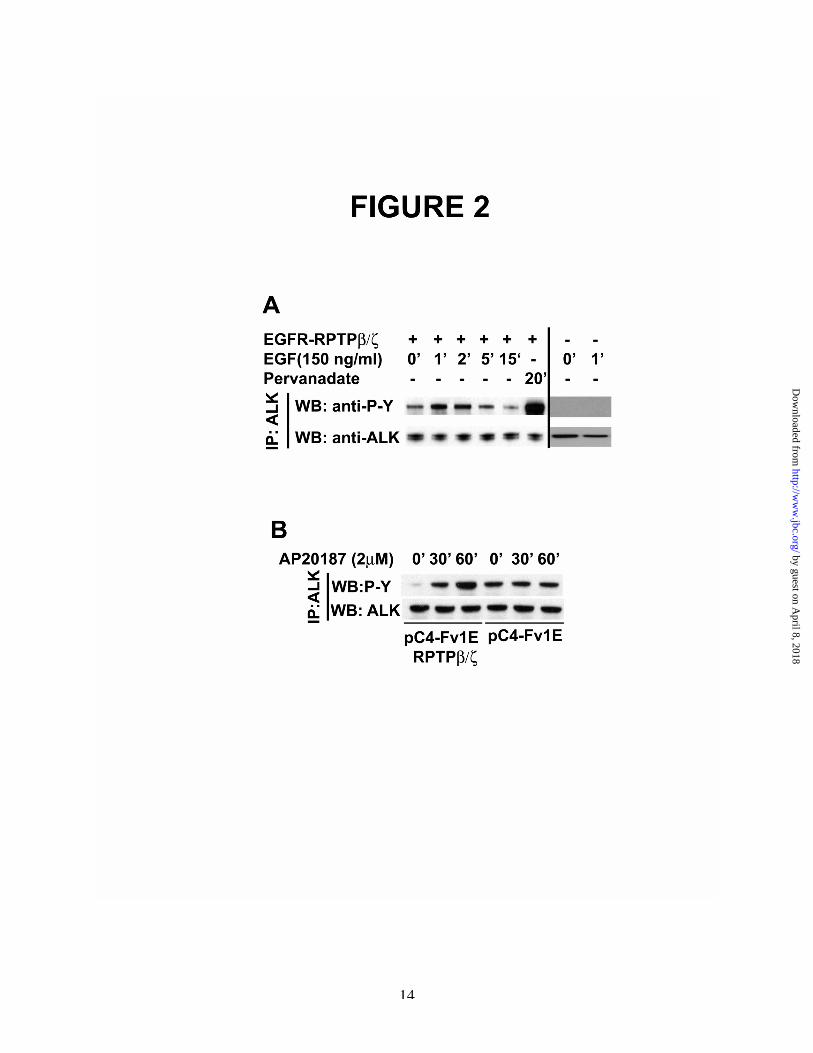
EGFR/RPTPb/z receptor was stably expressed inMCF-7 cells (MCF-7-EGFR/RPTPb/z cells). ALKwas introduced into MCF-7-EGFR/RPTPb/z cellsand, 48 hours later, the cells were stimulated withEGF. The levels of tyrosine phosphorylation ofALK in EGF-stimulated cells then were measuredas described above. EGF was found to stimulatetyrosine phosphorylation of ALK ~2.2-fold afterMCF-7-EGFR/RPTPb/z cells were stimulatedwith EGF for 1 and 5 minutes (Figure 2, A).
The data thus provide additional supportthat it is enforced dimerization of the intracellulardomain of RPTPb/z that has been induced by EGFthrough the EGFR extracellular domain thatstimulates tyrosine phosphorylation of ALK.
As anticipated, an ~4-fold increase inlevels of tyrosine phosphorylation of ALK wasfound when MCF-7-EGFR/RPTPb/z cells werestimulated with sodium pervanadate (Figure 2A),again supporting that multiple tyrosinephosphatases regulate levels of tyrosinephosphorylation of ALK, presumably at sites thatare different from the site(s) regulated through thePTN/RPTPb/z signaling pathway.
To exclude the possibility that EGFstimulates tyrosine phosphorylation of ALKthrough the endogenous EGFR, MCF-7 cells thatdo not express EGFR/RPTPb/z were transfectedwith ALK, stimulated with EGF, and analyzed asabove. EGF did not stimulate tyrosinephosphorylation of ALK (Figure 2, A).
Chemically Enforced Dimerization of RPTPb/zStimulates Tyrosine Phosphorylation of ALK.
To directly demonstrate that enforceddimerization of RPTPb/z alone is sufficient tostimulate tyrosine phosphorylation of ALK, MCF-7 cells were transfected with a vector pC4-Fv1Ethat contains the cDNA sequence to encode the Fvdomain of FKBP12 in frame with the intracellulardomain of RPTPb/z (see Methods). The FVdomain of FKBP12 is needed to enforcedimerization of RPTPb /z that is inducedchemically by AP20187 (25). The MCF-7 cellswere then transfected with a cDNA to encode full-length ALK and, 48 hours later, the cells werestimulated with AP20187 (2mM) for 30 and 60minutes. The levels of tyrosine phosphorylation ofALK were then analyzed in Western blots of cell
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
lysates probed with anti-phosphotyrosineantibodies.
Enforced dimerization induced byAP20187 was found to stimulate tyrosinephosphorylation of ALK ~12-fold and ~25-foldwhen analyzed 30 and 60 minutes after the cellswere stimulated with AP20187 (Figure 2B). In thecontrol cells that expressed ALK but contained an“empty” pC4-Fv1E vector, stimulation of the cellswith AP20187 failed to increase tyrosinephosphorylation of ALK.
These data thus directly demonstrateenforced dimerization of RPTPb/z alone in MCF-7 cells that express pC4-Fv1E in frame with theintracellular domain of RPTPb/z and ALK issufficient to initiate tyrosine phosphorylation ofALK. The data support further that the activitylevels of RPTPb/z in cells in which RPTPb/z hasnot been inactivated regulates the steady statelevels of tyrosine phosphorylation of ALK. Thevery high level of increase of tyrosinephosphorylation of ALK in cells stimulated withAP20187 suggests AP20187 is more effective ininducing dimerization of RPTPb/z expressed withthe Fv domain of FKBP12 than found in PTN-stimulated cells.
It was found that the levels of tyrosinephosphorylation of ALK in non-stimulated cellsthat express pC4-Fv1E alone were very high. Aswill be demonstrated subsequently, ALK is asubstrate of RPTPb/z. Thus, it is predicted that the“empty” vector that lacks RPTPb/z cannoteffectively reduce tyrosine phosphorylation ofALK through the activity of RPTPb/z. It is noticedalso that the levels of tyrosine phosphorylation ofALK in non-stimulated cells that express pC4-Fv1E-RPTPb/z are very low, suggesting thepossibility that RPTPb/z with the Fv domain ofFKBP12 may function as a more effective tyrosinephosphatase than RPTPb/z “tethered” through itstransmembrane and extracellular domains.
ALK that lacks an extracellular domain(truncated ALK) is phosphorylated in PTN-stimulated cells; PTN requires RPTPb/z butnot the extracellular domain of ALK tostimulate tyrosine phosphorylation of ALK.
To exclude the possibility that PTNinteracts directly with ALK alone or with ALKand RPTPb / z to st imulate tyrosine
phosphorylation of ALK, MCF-7 cells were co-transfected with plasmids to encode RPTPb/z and"truncated ALK" that lacks the extracellulardomain; truncated ALK contains only the first 8amino acids of the extracellular domain but retainsthe intact transmembrane and intracellulardomains (amino acid 1027-1620). Thus, in cellsexpressing truncated ALK but not endogenousALK, PTN cannot directly interact with ALK tostimulate tyrosine phosphorylation of ALK.
MCF-7 cells transfected together withRPTPb/z and truncated ALK were thereforestimulated with PTN and the levels of tyrosinephosphorylation of ALK were measured as above.
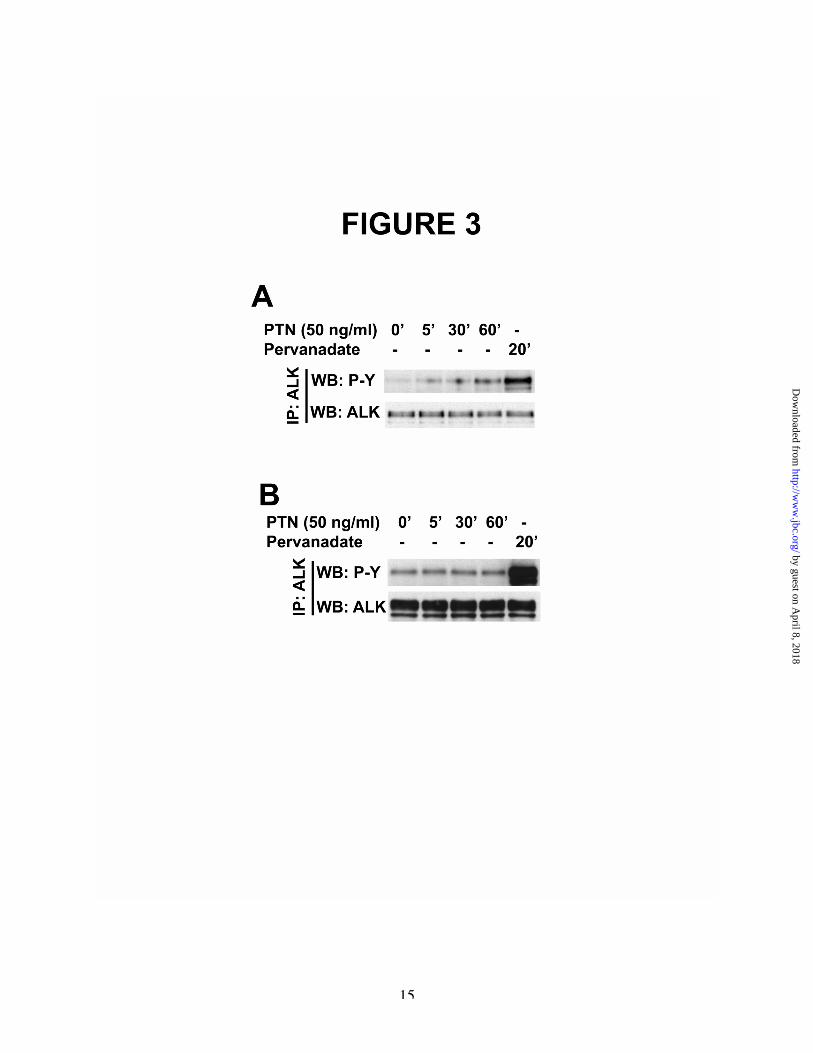
PTN was shown to stimulate tyrosinephosphorylation of truncated ALK ~5-, ~13-, and~26-fold after stimulation for 5, 30, and 60minutes (Figure 3A). The data thus demonstratedirectly that PTN stimulates an increase intyrosine phosphorylation of ALK independently ofan interaction of PTN with the extracellulardomain of ALK. The data furthermoredemonstrate that PTN requires RPTPb/z for PTNto stimulate tyrosine phosphorylation of ALKwithout an extracellular domain. As anticipated,PTN failed to stimulate tyrosine phosphorylationof the truncated ALK in MCF-7 cells thatexpressed truncated ALK but not RPTPb/z.
It was found that the baseline levels oftyrosine phosphorylation of ALK in non-stimulated cells were low. Again, as shownsubsequently, ALK is a substrate of RPTPb/z; thelow baseline level of ALK phosphorylationsuggest the possibility that the extracellulardomain of ALK may restrict the availability ofALK in some manner as a substrate of RPTPb/zthat is not the case in truncated ALK that lacks theextracellular domain.
The remarkably high levels of tyrosinephosphorylation of truncated ALK in PTN-stimulated MCF-7 cells transfected with bothRPTPb/z and truncated ALK suggests thepossibility that the levels of ALK phosphorylationare negatively regulated by the extracellulardomain of ALK; ALK phosphorylation may befacilitated when ALK dimerization is notencumbered by the extracellular domain of ALK.
As also anticipated, sodium pervanadatestimulates increased tyrosine phosphorylation of
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
truncated ALK to levels in excess of tyrosinephosphorylation stimulated by PTN (Figure 3B).
A L K c o - i m m u n o p r e c i p i t a t e s w i t hEGFR/RPTPb/z.
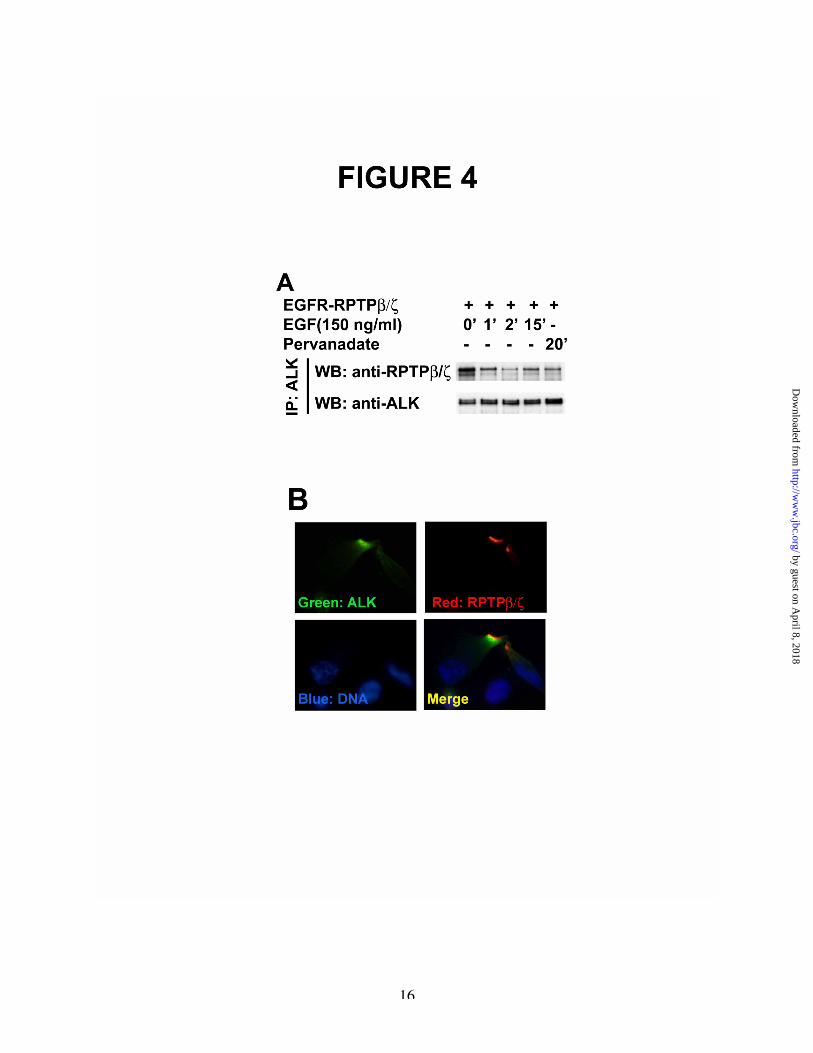
To address the mechanism through whichenforced inactivation of RPTPb /z increasestyrosine phosphorylation of ALK in PTN-stimulated cells, the possibility suggested abovethat ALK is a substrate of RPTPb/z was pursued.To first determine if RPTPb/z and ALK associatetogether, lysates of MCF-7-EGFR/RPTPb/z cellsthat t ransient ly express ALK wereimmunoprecipitated with anti-ALK antibodies.The immunoprecipitates were then analyzed inWestern blots probed with anti-RPTPb/zantibodies as above. It was found thatEGFR/RPTPb/z and ALK co-immunoprecipitatetogether (Figure 4A). However, an ~60% loss ofthe EGFR/RPTPb/z that co-immunoprecipitatedwith ALK from lysates of unstimulated cells wasfound in lysates from MCF-7-EGFR/RPTPb/zcells that were stimulated with EGF for 1 minute.An ~75% loss of association was found at 2minutes in lysates of stimulated cells. Thus,inactivation of RPTPb/z by enforced dimerizationinduced by EGF in MCF-7-EGFR/RPTPb/z cellssharply reduced the levels of association of ALKand EGFR/RPTPb/z.
The data suggest the possibility that ALKassociates with RPTPb/z through an interaction ofALK with the active site of RPTPb/z in cells thathave not been stimulated with EGF. When theactive site of RPTPb/z is no longer accessible toALK because of enforced dimerization access ofALK to the active site of RPTPb/z is denied. Thedata thus are consistent with the possibility thatALK is a substrate of RPTPb/z.
To further pursue whether ALKassociates with RPTPb/z in vivo, U373 cells whichexpress high levels of endogenous RPTPb/z (14)were then transfected with a cDNA to encodeALK and stained with FITC-tagged anti-ALKantibodies (Green), Texas-red-tagged anti-RPTPb/z antibodies (Red), and DAPI (Blue).Using confocal microscopy, ALK and RPTPb/zwere seen to co-localize (Figure 4B), thussupporting the likelihood that ALK and RPTPb/zassociate together in vivo.
ALK is a substrate of RPTPb/zTo more directly test whether ALK is a
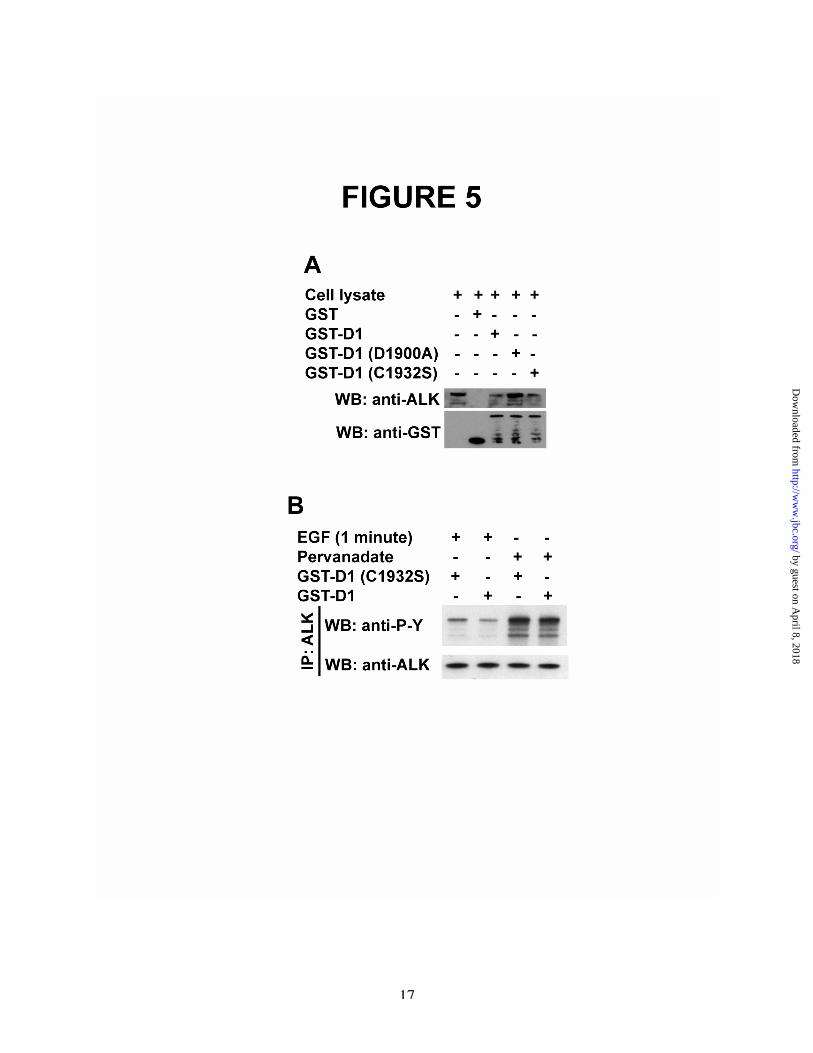
substrate of RPTPb/z , HeLa cells, which expressRPTPb/z , were transfected with a cDNA toencode ALK and stimulated with PTN. Lysateswere then prepared from the PTN-stimulated cellsand incubated with the GST-D1 domain ofRPTPb/z or with the GST-tagged RPTPb/z D1domain mutants, GST-RPTPb/z D1 (C1932S), orGST-RPTPb/z D1 (D1900A). Proteins thatassociated with GST-RPTPb/z D1, GST-RPTPb/zD1 (C1932S), and GST-RPTPb/z D1 (D1900A)were then captured with glutathione-Sepharose,eluted, and analyzed in Western blots probed withanti-ALK antibodies (18). The D1 domaincontains the active tyrosine phosphatase site ofRPTPb/z and itself is an active tyrosinephosphatase. RPTPb/z D1 (C1932S) is aninactivated active site D1 domain with thecatalytic cysteine mutated to serine. RPTPb/z D1(D1900A) is the “substrate trap” D1 mutant; thesubstrate trap mutant “captures” substrates ofRPTPb/z with very high affinity and specificity(30). It was found that the GST-tagged RPTPb/zD1 and each of the GST-tagged RPTPb/z D1domain mutants captured ALK (Figure 5A); thecapture of ALK by the D1 (D1900A) substratetrap mutant establishes with near certainty thatALK is a substrate of RPTPb/z.
To demonstrate directly that ALK isdephosphorylated by RPTPb/z and thus a substrateof RPTPb/z, ALK was then immunoprecipitatedfrom lysates of EGF-stimulated MCF-7-EGFR/RPTPb/z cells. The immunoprecipitateswere resuspended and incubated with the activesite D1 domain of RPTPb/z or with the inactiveRPTPb/z D1 (C1932S) mutant. After 2 hours at37°C, the proteins were then analyzed in Westernblots probed with anti-phosphotyrosine antibodiesand scanning densitometry. It was found thatlevels of tyrosine phosphorylation of ALK werereduced by ~43% in lysates derived from EGF-stimulated MCF-7-EGFR/RPTPb /z c e l l sincubated with the active GST-RPTPb /z D1domain (Figure 5B, lane 2) when compared withlevels of tyrosine phosphorylation of ALK thathad been incubated with the inactive GST-
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
RPTPb/z D1 (C1932S) mutant (Figure 5B, lane1).
The data thus strongly support RPTPb/zdephosphorylates ALK at the site in ALKphosphorylated when RPTPb/z was inactivated inEGF-stimulated MCF-7-EGFR/RPTPb/z cells.
Importantly also, and as anticipated,tyrosine phosphorylation of ALK in sodiumpervanadate stimulated cells was reduced by only~16% by incubation with GST-RPTPb /z D1(Figure 5B, lane 4) compared with incubation withthe inactive RPTPb/z mutant (Figure 5B, lane 3),consistent with data cited above that multipletyrosine phosphatases dephosphorylate ALK insites that are not recognized by RPTPb/z.
ALK is autoactivated and autophosphorylatedin tyrosine in vitro: ALK autophosphorylatedand autoactivated is dephosphorylated byRPTPb/z.
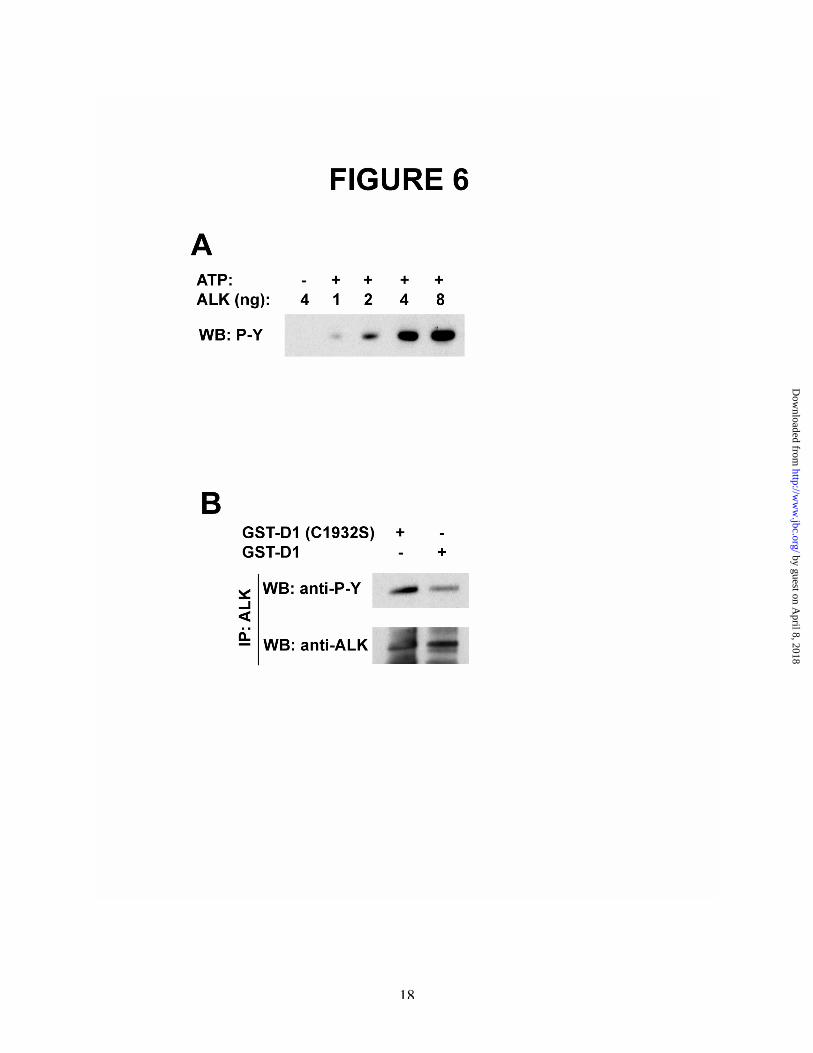
To pursue the mechanism through whichALK is phosphorylated in PTN-stimulated cells,ALK was incubated in an in vitro kinase assay.The levels of tyrosine phosphorylation of ALKwere measured in Western blots probed with anti-phosphotyrosine antibodies (Figure 6A); tyrosinephosphorylation of ALK was readily apparent oneminute after initiation of the in vitro kinasereaction and increased in levels for up to 20minutes. ALK was then immunoprecipitated withanti-ALK antibodies and the immunoprecipitateswere incubated with either GST-RPTPb/z D1 orwith GST-RPTPb/z D1 (C1932S) and analyzed inWestern blots probed with anti-phosphotyrosineantibodies as described above (Figure 6B). It wasfound that ALK autophosphorylated in tyrosine invitro was dephosphorylated by GST-RPTPb/z D1,and thus likely to be the same site in ALKphosphorylated in tyrosine in PTN-stimulatedcells, that also is dephosphorylated by RPTPb/z.
The data thus support the importantconclusion that ALK is phosphorylated at thesame site in PTN-stimulated cells that isphosphorylated when ALK is autoactivated invitro. In both cases, this site(s) is a substrate ofRPTPb/z.
DISCUSSIONReceptor protein tyrosine kinases (RTKs)
are activated through engagement of ligands (7,8)
which enforce dimerization and lead toautoactivation and autophosphorylation of thecognate receptors. In the present study, it isdemonstrated that PTN stimulates tyrosinephosphorylation and activation of ALK through amechanism that is independent of ligand enforceddimerization of ALK; it is suggested that ALK isactivated through a unique “AlternativeMechanism of RTK Activation”. The data supportthat a mechanism of activation of ALK isautoactivation and autophosphorylation of ALKthat continues to advance when theautophosphorylation sites in ALK no longer aredephosphorylated by RPTPb/z that has beeninactivated in PTN-stimulated cells; the steadystate activity of ALK is regulated through thelevels of activity of RPTPb / z thatdephosphorylates the crit ical tyrosinephosphorylation site(s) phosphorylated duringALK activation. Activation thus is not the result ofa direct interaction of PTN with ALK. The modeldoes not preclude unidentified tyrosine kinasesalso may phosphorylate the site in ALK that isautophosphorylated through autoactivation ofALK. The model thus underscores the steady-statelevels of tyrosine phosphorylation of ALK areregulated through the activity of thePTN/RPTPb/z signaling pathway.
The Alternative Mechanism of RTKActivation of ALK also raises the importantpossibility that through the PTN/RPTPb /zsignaling pathway, PTN may regulate multipleRTKs and thus effectively regulate differentimportant cellular functions; it potentially alsoeffectively expands the range of different systemsregulated through this pathway.
In studies of Mourali et. al. (6), ALK wasshown to have proapoptotic activity that isrelieved when engaged by agonists that activate itscatalytic activity. Our experiments thus implicatePTN as a potential regulator of the low levels oftyrosine phosphorylation of ALK that are criticalto cell survival. Since many transformed cells areknown to constitutively express Ptn, PTN may beanti-apoptotic through maintaining levels ofactivation of ALK sufficient to support survival ofmalignant cells.
This study identifies ALK as a new andimportant downstream of the PTN/RPTPb/zsignaling pathway whose levels of tyrosine
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
phosphorylation are regulated by PTN. Additionaltargets of the PTN/RPTPb/z signaling pathwaythus far identified include b -catenin (14),GIT1/Cat-1(15), Fyn (16), b-adducin (18), andP190RhoGAP (19). In each case, PTN stimulates asharp increase in the levels of tyrosinephosphorylation of the substrates of RPTPb/z inPTN-stimulated cells. Furthermore, in each casestudied (Fyn, b -catenin, b-adducin), RPTPb/zdephosphorylates each of the substrates ofRPTPb/z whose levels of tyrosine phosphorylationhave been increased in PTN-stimulated cells.Through this pathway, PTN thus coordinatelyregulates steady state levels of tyrosinephosphorylation of critical proteins important indiverse systems, such as cytoskeletal stability andfunction, intracellular and transmembrane tyrosinekinase activities, and chromatin remodeling. It issuggested this pathway profoundly influencescellular functions at different levels andcoordinately influences different cellular systemsto alter the cellular phenotype.
The view is supported in studies thatdemonstrate that PTN stimulates an epithelial tomesenchymal transition (EMT), in which loss ofcell-cell adhesion, profound cytoskeletonalternations, and a morphological transition to amore motile and invasive phenotype are inducedthrough the PTN/RPTPb/z signaling pathway (24).The EMT is a critical event in early developmentand a critical component of progression ofmalignant cells to a more aggressive phenotype(31,32).
Lu et al. (29) isolated two forms of PTN,one, an 18 kDa form (PTN18) and second, a 15kDa form (PTN15) that result fromposttranslational modifications. The studies thatwe report use the 136 amino acid 18 kDa PTN
initially reported (12,13). Pleiotrophin 15 (residues1-124) lacks the 12 C-terminal amino acids(KKEGKKQEKMLD) of PTN18, and, thus, itlacks the 5 lysine residues that establish the strongnet positive charge of the C-terminal domain ofPTN. In their studies, immobilized PTN18 wasshown to stimulate migration of glioblastoma cellsin an RPTPb /z dependent manner whereasimmobilized PTN15 promoted proliferation ofglioblastoma cells in an ALK-dependent fashion.The data of Lu et al. raise the possibility thatPTN18 and PTN15 signal through differentreceptors. However, the PTN15-stimulatedproliferation response that was reported to occurthrough ALK was determined by demonstratingphosphorylation of Akt; thus it is possible thatALK is activated either through an interactiondirectly with PTN15 or, alternatively, through adifferent signaling pathway, thus, these data do notdistinguish the mechanism of activation of ALKand are not inconsistent with our data. However,the studies of Lu et al. (29), are consistent withprevious studies in this laboratory in which the N-and C-terminal domains of PTN wereindependently expressed with the endogenousPTN signal peptide; in those studies, the differenttruncated PTNs were found to stimulatetransformation and angiogenesis, respectively(33).
These data thus clearly demonstrate thatALK is activated through "An AlternativeMechanism of RTK Activation". To the best ofour knowledge, this study is the first todemonstrate that a cytokine-dependent inactivationof a transmembrane tyrosine phosphatase is themechanism to activate a transmembrane receptortyrosine kinase.
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
REFERENCES1. Morris, S. W., Naeve, C., Mathew, P., James, P. L., Kirstein, M. N., Cui, X., and Witte, D. P.
(1997) Oncogene 14(18), 2175-21882. Le Beau, M. M., Bitter, M. A., Larson, R. A., Doane, L. A., Ellis, E. D., Franklin, W. A., Rubin,
C. M., Kadin, M. E., and Vardiman, J. W. (1989) Leukemia 3(12), 866-8703. Mason, D. Y., Bastard, C., Rimokh, R., Dastugue, N., Huret, J. L., Kristoffersson, U., Magaud, J.
P., Nezelof, C., Tilly, H., Vannier, J. P., and et al. (1990) Br J Haematol 74(2), 161-1684. Morris, S. W., Kirstein, M. N., Valentine, M. B., Dittmer, K. G., Shapiro, D. N., Saltman, D. L.,
and Look, A. T. (1994) Science 263(5151), 1281-12845. Iwahara, T., Fujimoto, J., Wen, D., Cupples, R., Bucay, N., Arakawa, T., Mori, S., Ratzkin, B.,
and Yamamoto, T. (1997) Oncogene 14(4), 439-4496. Mourali, J., Benard, A., Lourenco, F. C., Monnet, C., Greenland, C., Moog-Lutz, C., Racaud-
Sultan, C., Gonzalez-Dunia, D., Vigny, M., Mehlen, P., Delsol, G., and Allouche, M. (2006) MolCell Biol 26(16), 6209-6222
7. Schlessinger, J. (2000) Cell 103(2), 211-2258. Schlessinger, J. (1993) Harvey Lect 89, 105-1239. Bowden, E. T., Stoica, G. E., and Wellstein, A. (2002) J Biol Chem 277(39), 35862-3586810. Powers, C., Aigner, A., Stoica, G. E., McDonnell, K., and Wellstein, A. (2002) J Biol Chem
277(16), 14153-1415811. Stoica, G. E., Kuo, A., Aigner, A., Sunitha, I., Souttou, B., Malerczyk, C., Caughey, D. J., Wen,
D., Karavanov, A., Riegel, A. T., and Wellstein, A. (2001) J Biol Chem 276(20), 16772-1677912. Li, Y. S., Milner, P. G., Chauhan, A. K., Watson, M. A., Hoffman, R. M., Kodner, C. M.,
Milbrandt, J., and Deuel, T. F. (1990) Science 250(4988), 1690-169413. Milner, P. G., Li, Y. S., Hoffman, R. M., Kodner, C. M., Siegel, N. R., and Deuel, T. F. (1989)
Biochem Biophys Res Commun 165(3), 1096-110314. Meng, K., Rodriguez-Pena, A., Dimitrov, T., Chen, W., Yamin, M., Noda, M., and Deuel, T. F.
(2000) Proc Natl Acad Sci U S A 97(6), 2603-260815. Kawachi, H., Fujikawa, A., Maeda, N., and Noda, M. (2001) Proc Natl Acad Sci U S A 98(12),
6593-659816. Pariser, H., Ezquerra, L., Herradon, G., Perez-Pinera, P., and Deuel, T. F. (2005) Biochem
Biophys Res Commun 332(3), 664-66917. Pariser, H., Herradon, G., Ezquerra, L., Perez-Pinera, P., and Deuel, T. F. (2005) Proc Natl Acad
Sci U S A 102(35), 12407-1241218. Pariser, H., Perez-Pinera, P., Ezquerra, L., Herradon, G., and Deuel, T. F. (2005) Biochem
Biophys Res Commun 335(1), 232-23919. Tamura, H., Fukada, M., Fujikawa, A., and Noda, M. (2006) Neurosci Lett20. Englund, C., Loren, C. E., Grabbe, C., Varshney, G. K., Deleuil, F., Hallberg, B., and Palmer, R.
H. (2003) Nature 425(6957), 512-51621. Lee, H. H., Norris, A., Weiss, J. B., and Frasch, M. (2003) Nature 425(6957), 507-51222. Maeda, N., Ichihara-Tanaka, K., Kimura, T., Kadomatsu, K., Muramatsu, T., and Noda, M.
(1999) J Biol Chem 274(18), 12474-1247923. Moog-Lutz, C., Degoutin, J., Gouzi, J. Y., Frobert, Y., Brunet-de Carvalho, N., Bureau, J.,
Creminon, C., and Vigny, M. (2005) J Biol Chem 280(28), 26039-2604824. Perez-Pinera, P., Alcantara, S., Dimitrov, T., Vega, J. A., and Deuel, T. F. (2006) Proc Natl Acad
Sci U S A 103(47), 17795-1780025. Fukada, M., Fujikawa, A., Chow, J. P., Ikematsu, S., Sakuma, S., and Noda, M. (2006) FEBS Lett26. Lamant, L., Pulford, K., Bischof, D., Morris, S. W., Mason, D. Y., Delsol, G., and Mariame, B.
(2000) Am J Pathol 156(5), 1711-172127. Dirks, W. G., Fahnrich, S., Lis, Y., Becker, E., MacLeod, R. A., and Drexler, H. G. (2002) Int J
Cancer 100(1), 49-56
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
28. Motegi, A., Fujimoto, J., Kotani, M., Sakuraba, H., and Yamamoto, T. (2004) J Cell Sci 117(Pt15), 3319-3329
29. Lu, K. V., Jong, K. A., Kim, G. Y., Singh, J., Dia, E. Q., Yoshimoto, K., Wang, M. Y.,Cloughesy, T. F., Nelson, S. F., and Mischel, P. S. (2005) J Biol Chem 280(29), 26953-26964
30. Dewang, P. M., Hsu, N. M., Peng, S. Z., and Li, W. R. (2005) Curr Med Chem 12(1), 1-2231. Thiery, J. P. (2002) Nat Rev Cancer 2(6), 442-45432. Thiery, J. P. (2003) Curr Opin Cell Biol 15(6), 740-74633. Zhang, N., Zhong, R., Perez-Pinera, P., Herradon, G., Ezquerra, L., Wang, Z. Y., and Deuel, T. F.
(2006) Biochem Biophys Res Commun 343(2), 653-658
AcknowledgementsThis is manuscript number 18518 from the Scripps Research Institute. This work was supported by grantCA84400 from The National Institutes of Health. The MEM core laboratory is supported by Sam andRose Stein Endowment Fund. The vector pC4-Fv1E was generous gift from ARIAD Pharmaceuticals, Inc.Pablo Perez-Pinera was supported by grant 2 T32 DK007022-26 from the National Institutes of Health.Wei Zhang and Yunchao Chang were supported by Skaggs training grant.
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
FIGURE LEGENDSFigure 1A. Pleiotrophin stimulates tyrosine phosphorylation of ALK in MCF-7 cells that express both ALK andRPTPb/z. Complementary DNAs to encode ALK and RPTPb/z were transiently expressed in MCF-7cells. After 48 hours, cells were stimulated with 50ng/ml PTN for 5, 30, and 60 minutes or, as control,with sodium pervanadate for 20 minutes. Cell lysates were immunoprecipitated with anti-ALK antibodiesand the immunoprecipitates were analyzed using scanning densitometry of Western blots probed withanti-phosphotyrosine antibodies and with anti-ALK antibodies.B. Pleiotrophin fails to stimulate tyrosine phosphorylation of ALK in MCF-7 cells that do not expressRPTPb/z. The cDNA to encode ALK was transiently expressed in MCF-7 cells. After 48 hours, cellswere stimulated with PTN for 5, 30, and 60 minutes or, as control, with sodium pervanadate. Cell lysateswere immunoprecipitated with anti-ALK antibodies and analyzed by scanning densitometry of Westernblots probed with anti-phosphotyrosine antibodies and separately with anti-ALK antibodies.C. ShRNAs to “knock down” RPTPb/z block PTN-stimulated tyrosine phosphorylation of ALK. Aretroviral vector (pSM2) encoding two shRNAs to "knock down" RPTPb/z or pSM2 containing a“scrambled” shRNA were introduced into U87MG cells that express endogenous RPTPb/z and ALK.After 96 hours, the levels of expression of RPTPb/z in cells incubated with either of the two shRNA’s toknock down RPTPb/z or the “scrambled” (control) shRNA were measured by quantitative RT-PCR. Itwas found that expression levels of RPTPb/z were reduced ~66% and ~73% with each shRNA to knockdown RPTPb/z respectively relative to the control. The cells were then stimulated with 50 ng PTN/ml for30 minutes. Cell lysates were prepared and used to immunoprecipitate ALK. The immunoprecipitateswere analyzed by scanning densitometry in Western blots probed with anti-phosphotyrosine antibodiesand with anti-ALK antibodies.
Figure 2A. EGF stimulates tyrosine phosphorylation of ALK in EGF-stimulated MCF-7-EGFR/RPTPb/z cells. Achimeric protein that contains the extracellular and transmembrane domains of EGFR was fused with theintracellular domains of RPTPb/z and stably expressed in MCF-7 cells (MCF-7-EGFR/RPTPb/z cells)that transiently express ALK. Lysates from stimulated and control non-stimulated cells wereimmunoprecipitated with anti-ALK antibodies and analyzed by scanning densitometry in Western blotsprobed with anti-phosphotyrosine antibodies (left side). Control MCF-7 cells that do not expressEGFR/RPTPb/z (right side).B. Enforced dimerization of RPTPb/z stimulates tyrosine phosphorylation of ALK. ComplementaryDNAs to encode ALK and a vector pC4-Fv1E containing the cDNA sequence encoding the intracellulardomain of RPTPb/z or an empty pC4-Fv1E vector were transiently expressed in MCF-7cells. After 48hours, the cells were stimulated with 2mM AP20187 for 30 and 60 minutes. Cell lysates wereimmunoprecipitated with anti-ALK antibodies and analyzed with scanning densitometry of Western blotsprobed with anti-phosphotyrosine and with anti-ALK antibodies.
Figure 3A. Pleiotrophin-stimulated tyrosine phosphorylation of ALK does not require an interaction of PTN withthe extracellular domain of ALK. MCF-7 cells transfected with plasmids to encode full-length RPTPb/zand to encode a truncated ALK that contains the 8 membrane proximal extracellular amino acids and thetransmembrane and intracellular domains (amino acids 1028-1620) of ALK were stimulated with 50 ngPTN/ml for 5, 30 and 60 minutes or with sodium pervanadate for 20 minutes. Cell lysates wereimmunoprecipitated with anti-ALK antibodies and the immunoprecipitates were analyzed using scanningdensitometry of Western blots probed with anti-phosphotyrosine and anti-ALK antibodies.B. Pleiotrophin fails to stimulate tyrosine phosphorylation of truncated ALK in MCF-7 cells that expressALK but not RPTPb/z. A cDNA to encode truncated ALK transiently expressed in MCF-7 cells was
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
stimulated with PTN (50 ng/ml) for 5, 30 and 60 minutes or with sodium pervanadate for 20 minutes. Celllysates were immunoprecipitated with anti-ALK antibodies and analyzed by scanning densitometry ofWestern blots probed with anti-phosphotyrosine and anti-ALK antibodies.
Figure 4A. ALK associates with EGFR/RPTPb/z in lysates of MCF-7-EGFR/RPTPb/z cells; loss of association ofALK and EGFR/RPTPb/z is demonstrated in cells stimulated with EGF. MCF-7 cells that stably expressthe chimeric receptor protein EGFR/RPTPb/z were co-transfected with ALK and either not stimulated orstimulated with EGF for 1 minute (lane 2), 2 minutes (lane 3), 5 minutes (lane 4), or 15 minutes (lane 5),or with pervanadate for 30 minutes (lane 6). Cell lysates were used to immunoprecipitate ALK and theimmunoprecipitates were analyzed in Western blots probed with anti-phosphotyrosine antibodies (upperpanel) and anti-ALK antibodies (lower panel).B. Co-localization of ALK and RPTPb/z in vivo. U373 cells were transfected with an ALK cDNA andstained with FITC-tagged anti-ALK antibodies (Green) (upper left), Texas-red-tagged anti-RPTPb/zantibodies (Red) (upper right), and DAPI (Blue) (lower left). ALK and RPTPb/z were found to colocalizetogether in specific regions of the cell membrane (yellow, Merged) (lower right).
Figure 5A. ALK binds to and is a substrate of the D1 domain of RPTPb/z. GST-RPTPb/z D1, GST-RPTPb/z D1(A1900S), and GST-RPTPb/z D1 (C1932S) “capture” ALK in lysates of HeLa cells stimulated with PTN.Lysates of HeLa cells transfected with ALK stimulated with 50 ng PTN/ml were incubated with GSTalone (lane 2), GST-RPTPb/z D1 (lane 3), GST-RPTPb/z D1 (D1900A) (lane 4), GST-RPTPb/z D1(C1932S) (lane 5). Proteins captured were analyzed in Western blots probed with anti-ALK and anti-GSTantibodies.B. ALK is a substrate of RPTPb/z. MCF-7-EGFR/RPTPb/z cells were transfected with ALK andstimulated with EGF for 1 minute (lanes 1, 2) or pervanadate (lane 3, 4). Lysates wereimmunoprecipitated with anti-ALK antibodies and the immunoprecipitates were incubated with eitherGST-RPTPb/z D1 (lanes 2, 4) or GST-RPTPb/z D1 (C1932S) (lanes 1, 3). The samples were analyzed byscanning densitometry in Western blots probed with anti-phosphotyrosine antibodies (upper panel) andreprobed with anti-ALK antibodies (lower panel).
Figure 6A. ALK autophosphorylates itself in vitro. ALK (1 nM, 2 nM, 4 nM, and 8 nM) was incubated with ATP,or as control without ATP, in an in vitro kinase reaction at 37˚C for 10 minutes. The products of thereaction were analyzed in Western blots probed with anti-phosphotyrosine antibodies.B. ALK autophosphorylated in vitro is dephosphorylated in RPTPb/z. Autophosphorylated ALK (10 nM)from the reaction above was immunoprecipitated using anti-ALK antibodies and the immunoprecipitateswere incubated with GST-RPTPb/z D1 or GST-RPTPb/z D1 (C1932S) for 120 min at 37˚C and analyzedin Western blots.
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Pablo Perez-Pinera, Wei Zhang, Yunchao Chang, Jose Antonio Vega and Thomas F. Deuelof receptor tyrosine kinase (RTK) activation"
mechanismALIGN="BASELINE" ALT="zeta "> signaling pathway: An "alternative /<IMG SRC="/math/zeta.gif"β(PTN)/receptor protein tyrosine phosphatase (RPTP)
Anaplastic lymphoma kinase (ALK) is activated through the pleiotrophin
published online August 6, 2007J. Biol. Chem.
10.1074/jbc.M704505200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from