Embed Size (px)
Citation preview

JOURNAL OF CLINICAL MICROBIOLOGY, Mar. 1993, p. 629-6350095-1137/93/030629-07$02.00/0Copyright © 1993, American Society for Microbiology
Vol. 31, No. 3
Analysis of the Humoral Response to the Flagellin Protein ofBorrelia burgdorferi: Cloning of Regions Capable of
Differentiating Lyme Disease from SyphilisJOHN M. ROBINSON,`* TAMI J. PILOT-MATIAS,2 STEVEN D. PRATT,2 CHANDU B. PATEL,1
TERRY S. BEVIRT,1 AND J. C. HUNT1Infectious Disease and Immunology' and Corporate Molecular Biology, 2
Abbott Laboratories, Abbott Park; Illinois 60064
Received 10 August 1992/Accepted 25 November 1992
Selected regions of the Borrelia burgdojferi flagellin gene (fla) that exhibit high or low homology with relatedgenes from other bacterial species were amplified by the polymerase chain reaction and expressed as fusionproteins in Escherichia coli. Purified fusion proteins were assayed for antibody reactivity in a microtiter plateenzyme-linked immunosorbent assay with sera from Lyme disease patients as well as syphilitic and normalsera. Immunoglobulin G antibody from Lyme disease patient sera reacted predominantly with the centralportion of the protein. The region of the flagellin protein encompassing amino acids 64 to 311 detected nearlyall of the immunoglobulin G-positive Lyme sera and only reacted with 1 of 26 syphilis patient serum samples.In contrast, 12 of 26 syphilis patient serum samples and 2 of 47 normal serum samples reacted with the aminoterminus of the flagellin protein, whereas 4 of 26 syphilis patient serum samples and 7 of 47 normal serumsamples reacted with the carboxyl terminus. The central region containing amino acids 64 to 311 may beemployed diagnostically to differentiate antibodies to B. burgdorferi from antibodies to Treponema pallidum. Inaddition, this region also was recognized by immunoglobulin M in the Lyme patient sera, indicating itspotential usefulness as a marker for early Lyme disease.
Lyme disease is a multisystem illness caused by thetick-transmitted spirochete Borrelia burgdorferi (5, 21).Lyme borreliosis is the most common arthropod-borne in-fection in the United States and has been reported in manycountries throughout Asia and Europe (2, 21). The earlyclinical feature of this disease is a local infection of the skin,erythema migrans, which may be followed by the develop-ment of systemic disease involving the nervous system,heart, or joints (2, 21).
Culture of the spirochete from human body fluids andantigen detection methods often are falsely negative, leadingto misdiagnosis of Lyme disease (2, 21). Currently, serolog-ical methods to detect antibodies to B. burgdorferi are themost appropriate means to diagnose infection or exposure.Commonly used assays for the diagnosis of Lyme diseaseinclude the indirect immunofluorescence assay, enzyme-linked immunosorbent assay (ELISA), and Western immu-noblot, which has been shown to be more sensitive than theimmunofluorescence assay or the ELISA for the detection ofearly disease (11). However, analysis of Western blot reac-tivity is subjective with regard to band intensity; clearcriteria for blot interpretation are difficult to establish (25).Most ELISAs for Lyme disease utilize whole or sonicated B.burgdorferi cells as the test antigen, although several inves-tigators demonstrated improved performance of these immu-noassays when subcellular fractions of the spirochete wereused (6, 8, 13, 15).The B. burgdorferi flagellar protein, flagellin, is highly
immunogenic and generally elicits the earliest detectableimmune response after infection (6, 11). Flagellin-enrichedfractions of B. burgdorferi have been shown to improve theperformance of Lyme antibody assays (12, 13). However,
* Corresponding author.
the specificity of these assays may be reduced because ofcross-reactivity of the B. burgdorferi flagellin with the fla-gella of other spirochetes, most notably with Treponemapallidum, the causative agent of syphilis (16). It also hasbeen demonstrated that antibodies to oral spirochetes asso-ciated with gingivitis or periodontitis can react in serologicalassays for Lyme disease (17). Cross-reactivity with serafrom these patients can be reduced by adsorption of thepatient sera with the Treponema Reiter strain (17, 24), butthis also can reduce the sensitivity of the Lyme antibodyassays.The nucleotide and amino acid sequences have been
determined for the flagellin proteins of several B. burgdorferiisolates (7, 9, 23). Comparisons of the B. burgdorferi flagellinprotein sequence with those of flagellar proteins from T.pallidum, Salmonella typhimurium, Escherichia coli, Serra-tia marcescens, and Bacillus subtilis show regions with highsequence homology at each end of these proteins but morevariability in the central region (9, 23). In this study, we usedthe polymerase chain reaction to isolate the gene encodingthe B. burgdorferi flagellin protein as well as selected frag-ments of the gene and expressed them as fusion proteins inE. coli. After they were purified, their reactivity with serafrom Lyme disease and syphilis patients was evaluated.
MATERIALS AND METHODS
B. burgdorferi. B. burgdorferi B31 (ATCC 35210) wasgrown in BSKII medium (1) for 5 days at 37°C. Cells wereharvested by centrifugation at 10,000 x g for 15 min andused for isolation of DNA or as an antigen for Western blots.
Construction of recombinant flagellin proteins. Oligonucle-otide primers for polymerase chain reaction amplification ofthe entire flagellin gene and selected fragments were de-signed on the basis of the published sequence of the flagellin
629
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
on A
ugust 14, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
on A
ugust 14, 2019 by guesthttp://jcm
.asm.org/
Dow
nloaded from

J. CLIN. MICROBIOL.630 ROBINSON ET AL.
GGGc ACCTTA10191TC
pl-137 AAATAGTCTCGACCCG1TTTCAATCTAATAe0
p137-262 AAATAGkTCCAGAG CA A 28
p262-336 AATAG&TC&GACCCGG7TC&AGLC&AWAITGG
GGGCGGTACCTTA1 TGATACATGTGC&TTTGGTT391
G=MGCT CTTIT7786 767
GGGGGTACCTTA1011 ATCTGCAATGACAAC991
190 .29 933C913p64-311 AIATAGATCCAG&CCCG 0AGATACT GGCTAiT GGGCACTTkATTA933AC TATTGAC
FIG. 1. Oligonucleotide primers used for polymerase chain reaction amplification of B. burgdorferi flagellin gene DNA. The primersencode a BglII restriction site in the sense primer and a HindIll or KpnI site in the antisense primer. Flagellin gene nucleotide sequencepositions are indicated by superscripts.
gene of strain B31 (23) (Fig. 1). DNA was isolated from a200-ml culture of B. burgdorferi by standard methods (10).Briefly, cells were suspended in 8.5 ml of 25 mM Tris-10mMEDTA (pH 8.0) containing 50 mM glucose and 2 mg oflysozyme per ml. After 15 min at room temperature, 1.25 mlof 20% Sarkosyl-0.25 M EDTA (4:1, vol/vol) was added; thepreparation was mixed gently, and 9.3 g of cesium chlorideand 0.5 ml of a 5-mg/ml solution of ethidium bromide wereadded. The mixture was centrifuged at 150,000 x g for 40 h.The DNA band was isolated, extracted with NaCl-saturatedisopropanol, precipitated with ethanol, and resuspended in10 mM Tris-1 mM EDTA (pH 8.0).Each polymerase chain reaction consisted of a mixture of
20 mM (NH4)2SO4, 80 mM Tris (pH 9.0), 10 mM MgCl2, 85ng of B. burgdorfeni DNA, 60 pmol of each primer, 0.4 mM(each) dATP, dCTP, dGTP, and dTTP, and 2.5 U of Taqpolymerase. The DNA was amplified by using the followingcycles: 1 cycle of 97°C for 2 min; 4 cycles of 95°C for 0.5 min,40°C for 0.5 min, and 72°C for 1 min; and then 25 cycles of95°C for 0.5 min and 65°C for 1.5 min.The polymerase chain reaction products were digested
with BglII and HindIII and cloned into pTPM210 or digestedwith BglII and KpnI and cloned into pTB210 by standardtechniques. Plasmids pTPM210 and pTB210 are expressionvectors containing the kdsB gene, which codes for CMP-2-keto-3-deoxyoctulosonic acid synthetase (CKS) (4), which isused as a fusion partner. The resultant plasmid was trans-formed into competent E. coli K-12 strain XL-1 Blue (Strat-agene Cloning Systems, La Jolla, Calif.).Cloned regions of the flagellin protein gene were se-
quenced by the dideoxynucleotide chain-termination method(20) to verify fidelity of amplification.
Expression and purification of recombinant proteins. Bac-terial clones expressing flagellin proteins were grown over-night at 37°C in a rich medium (32 g of tryptone per liter, 20g of yeast extract per liter, 5 g of NaCl per liter, 3 mMglucose) containing ampicillin (50 ,ug/ml) and tetracycline (50,ug/ml). The overnight cultures were diluted 1:10 into thesame medium and incubated at 37°C with aeration until theculture reached an optical density at 600 nm of 1.0 and theninduced by the addition of isopropyl-,-D-thiogalactopyrano-side (IPTG; final concentration, 1 mM; Sigma). After incu-bation for 4 h, the cells were harvested by centrifugation at12,000 x g (4°C) for 15 min. Samples of preinduced andinduced cells were analyzed for recombinant protein produc-
tion by sodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE) as described below.To purify the recombinant proteins, the cells were lysed
by suspension and sonication in TE buffer (50 mM Tris, 10mM EDTA [pH 8.5]) containing 1 mg of lysozyme per ml,100 ,ug of DNase per ml, and 0.5% Triton X-100. The celllysate was centrifuged at 25,000 x g for 30 min, and therecombinant proteins were recovered in the pellet. SolubleE. coli proteins were removed from the pellet by washing inTE buffer containing 5% Triton X-100, TE buffer containing1% sodium deoxycholate, and then TE buffer containing 0.5M NaCl. After a water wash, the insoluble pellet containingthe recombinant fusion protein was solubilized in TE buffercontaining 8 M urea and 1 mM dithiothreitol and stored at40C.The expression levels and purities of the recombinant
proteins were evaluated by SDS-PAGE as described byLaemmli (14) with 12% acrylamide running gels and 5%acrylamide stacking gels. Samples were suspended in load-ing buffer (final concentrations: 62.5 mM Tris, 2% SDS, 10%glycerol, 5% 2-mercaptoethanol, and 0.1 mg of bromophenolblue per ml) and boiled for 2 min. Gels were stained in asolution of 0.125% Coomassie blue dye in 50% methanol and10% acetic acid for 1 h and then destained in a solution of 7%acetic acid and 5% methanol until a clear background wasobtained.
Sera and Western blot evaluation. The Lyme disease seraused in this study were kindly provided by T. Quan, Centerfor Disease Control, Fort Collins, Colo.; R. Ryan, Univer-sity of Connecticut Health Center, Farmington; WisconsinState Laboratory of Hygiene, Madison; J. Nelson, RushMedical Center, Chicago, Ill.; and D. Bussiere, BostonBiomedica, West Bridgewater, Mass. These sera were col-lected by physicians from individuals residing in endemicareas for Lyme disease including Wisconsin, Connecticut,New York, and Massachusetts, and diagnosed with Lymedisease on the basis of dermatological, neurological, cardiac,and/or arthritic manifestations. Sera from patients withprimary, secondary, or late-stage syphilis (syphilis-positivesera) were kindly provided by the Center for Disease Con-trol, Atlanta, Ga. Antibody titers of these specimens in theVenereal Diseases Research Laboratory test ranged from 1:2to 1:128, and the specimens were R3+ or R4+ reactive byfluorescent treponemal antibody absorption.Western blots were employed to evaluate the sera from
Region
pl-336
Antisense priter
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

LYME DISEASE-SPECIFIC B. BURGDORFERI FLAGELLIN REGIONS 631
38%
I52%o
1 13713%
336
137
30%
26254%
262 336
64 311FIG. 2. Regions of the B. burgdorfen flagellin gene chosen for cloning as CKS fusion proteins in E. coli. Numbers below the lines indicate
amino acid positions. The percentages of identical amino acid residues within each region compared with the T. pallidum endoflagellar subunitFlaB2 protein are indicated above the line.
patients with Lyme disease (Lyme disease sera) used in thisstudy. B. burgdorferi was denatured in SDS-PAGE loadingbuffer, and a volume containing 7.5 mg wet weight of cellswas electrophoresed into 12% acrylamide gels (13 by 13 cm).The separated proteins were electrophoretically transferredto nitrocellulose sheets (Bio-Rad Laboratories, Richmond,Calif.) at 700 mA for 45 min in buffer consisting of 25 mMTris (pH 8.3)-192 mM glycine-20% methanol (22), blockedovernight in 100 mM Tris-135 mM NaCl (TBS) containing3% gelatin, and cut into 3-mm-wide strips. Serum sampleswere diluted 1:50 in a diluent consisting of 100 mM Tris (pH7.5), 135 mM NaCl, 10 mM EDTA, 0.2% Tween 20, and 4%fetal calf serum (GIBCO, Grand Island, N.Y.) and allowedto react with the nitrocellulose strips for 2 h. After a washwith TBS, the antibody-antigen reactions were detected byincubation with peroxidase-labelled goat anti-human immu-noglobulin G (IgG; b-chain specific) or anti-human IgM(,u-chain specific) conjugates (Kirkegaard and Perry Labora-tories, Gaithersburg, Md.) diluted to 0.5 ,ug/ml in a diluentconsisting of 100 mM Tris (pH 7.5), 135 mM NaCl, and 10%fetal calf serum. The nitrocellulose was washed with TBScontaining 0.05% Tween 20, and bound conjugate wasdetected by incubation in TBS containing 2 mg of 4-chloro-1-napthol per ml, 0.02% hydrogen peroxide, and 17% meth-anol. Immunoblots were considered to be positive for IgG ifat least five proteins were reactive and three or moreproteins were from the group consisting of the 18-, 31-, 34-,41-, 60-, 66-, or 83-kDa proteins. An IgM immunoblot wasconsidered positive if at least three proteins were reactiveand two or more of the reactive proteins were from the groupof proteins listed above.ELISA. Purified recombinant antigens were diluted to 1 to
5 ,ug/ml in 50 mM carbonate buffer (pH 9.6), and 100 p1l wasadded to microtiter plate wells (MaxiSorp; Nunc, Inc.,Naperville, Ill.). The plates were incubated in the antigensolution for 1 h at 370C and overcoated in a solutionconsisting of 10% fetal calf serum and 3% gelatin in phos-phate-buffered saline for 30 min. Serum samples were di-luted 1:200 in the same antibody diluent used for the Westernblots, except 2% E. coli pTB210 lysate was added. Portions(100 pl) of each diluted serum sample were added to themicrotiter wells and incubated for 1 h at 37°C; then the plateswere washed three times with phosphate-buffered salinecontaining 0.05% Tween 20. Bound Ig was detected byincubation with peroxidase-labelled goat anti-human IgG oranti-human IgM conjugates as described above. After 1 h ofincubation (370C) with 100 pl of the desired conjugate per
well, the plate was washed as described above. The perox-idase substrate (0.02% hydrogen peroxide containing 2.5 mgof o-phenylenediamine dihydrochloride per ml) was allowedto react for 5 min at room temperature, and then the reactionwas stopped by the addition of 1 N H2SO4. The A490 wasread. The cutoff point for a positive result for each antigenwas determined to be the mean of results from a normalpopulation of 20 individuals with no known history ofspirochetal infection plus 0.13 A490 unit.The pTB210 lysate was included in the serum diluent to
block any possible serum reactivity with the CKS portion ofthe fusion protein or with any E. coli proteins remainingassociated with the purified recombinant proteins. Thislysate was prepared by sonicating E. coli cells containing thepTB210 vector plasmid in lysis buffer as described above andretaining the supernatant after centrifugation at 25,000 x gfor 30 min.
RESULTS
Expression of the flagellin protein and fragments. Theamino acid sequence of the B. burgdorferi B31 flagellinprotein (23) was aligned with the amino acid sequence of theT. pallidum endoflagellar subunit FlaB2 protein (19) by usingthe PALIGN program (PC-Gene; Intelligenetics, Inc.,Mountain View, Calif.). There are 38% identical residuesbetween the two sequences. When conservative changes areincluded, the two sequences are 51% homologous. Thehomology is greatest at the amino and carboxyl termini ofthe proteins, with the central region displaying the mostvariability.We cloned the B. burgdorferi full-length fla gene and four
gene fragments chosen on the basis of sequence homologywith the T. pallidum endoflagellar protein (Fig. 2; regionscloned are designated according to their amino acid residueendpoints). The regions were expressed as CKS fusionproteins under the control of a modified lac promoter asdescribed by Bolling and Mandecki (4). SDS-PAGE analysisof expression of each of the cloned fusion proteins beforeand after induction with IPTG (Fig. 3) indicated that fusionproteins of the approximate expected molecular masses (70,46, 45, 38, and 59 kDa for fusion proteins pl-336, pl-137,p137-262, p262-336, and p64-311, respectively) were pro-duced. Each of these proteins was expressed at 15 to 20% oftotal E. coli protein. Their identities were confirmed byimmunoblot analysis with anti-CKS monoclonal antibodiesand sera from patients with Lyme disease (data not shown).
VOL. 31, 1993
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from
632 ROBINSON ET AL.
MW 1 2 3 4 5 6 7 8 9 10
97--.
45- "Mu~'
21-1
14_
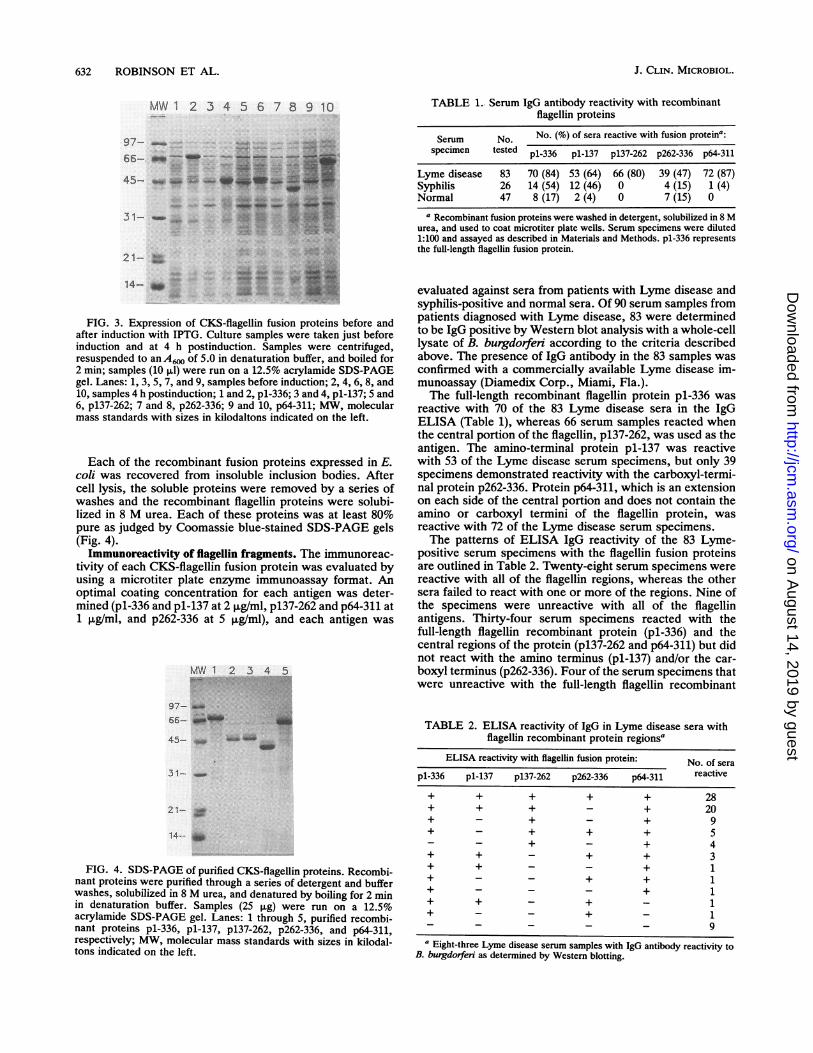
FIG. 3. Expression of CKS-fiagellin fusion proteins before and
after induction with IPTG. Culture samples were taken just before
induction and at 4 h postinduction. Samples were centrifuged,resuspended to an A6. of 5.0 in denaturation buffer, and boiled for
2 min; samples (10 pAl) were run on a 12.5% acrylamide SDS-PAGE
gel. Lanes: 1, 3, 5, 7, and 9, samples before induction; 2, 4, 6, 8, and
10, samples 4 h postinduction; 1 and 2, p1-336; 3 and 4, pl-137; 5 and
6, p137-262; 7 and 8, p262-336; 9 and 10, p64-311; MW, molecular
mass standards with sizes in kilodaltons indicated on the left.
Each of the recombinant fusion proteins expressed in E.
coli was recovered from insoluble inclusion bodies. After
cell lysis, the soluble proteins were removed by a series of
washes and the recombinant fiagellin proteins were solubi-
lized in 8 M urea. Each of these proteins was at least 80%
pure as judged by Coomassie blue-stained SDS-PAGE gels(Fig. 4).
Immunoreactivity of flagellin fragments. The immunoreac-
tivity of each CKS-fiagellin fusion protein was evaluated byusing a microtiter plate enzyme immunoassay format. An
optimal coating concentration for each antigen was deter-
mined (p1-336 and p1-137 at 2ing/ml, p137-262 and p64-311 at
1 plg/ml, and p262-336 at 5 10ml), and each antigen was
MW 1 2 3 4 5
97--
66-
45-
31-
21-
FIG. 4. SDS-PAGE of purified CKS-flagellin proteins. Recombi-nant proteins were purified through a series of detergent and bufferwashes, solubilized in 8 M urea, and denatured by boiling for 2 minin denaturation buffer. Samples (25 pg) were run on a 12.5%acrylamide SDS-PAGE gel. Lanes: 1 through 5, purified recombi-nant proteins pl-336, pl-137, p137-262, p262-336, and p64-311,respectively; MW, molecular mass standards with sizes in kilodal-tons indicated on the left.
TABLE 1.. Serum IgG antibody reactivity with recombinantflagellin proteins
Serum No. No. (%) of sera reactive with fusion proteina:specimen tested pl-336 pl-137 p137-262 p262-336 p64-311
Lyme disease 83 70 (84) 53 (64) 66 (80) 39 (47) 72 (87)Syphilis 26 14 (54) 12 (46) 0 4 (15) 1 (4)Normal 47 8 (17) 2 (4) 0 7 (15) 0
a Recombinant fusion proteins were washed in detergent, solubiized in 8 Murea, and used to coat microtiter plate wells. Serum specimens were diluted1:100 and assayed as described in Materials and Methods. pl-336 representsthe full-length flagellin fusion protein.
evaluated against sera from patients with Lyme disease andsyphilis-positive and normal sera. Of 90 serum samples frompatients diagnosed with Lyme disease, 83 were determinedto be IgG positive by Western blot analysis with a whole-celllysate of B. burgdorferi according to the criteria describedabove. The presence of IgG antibody in the 83 samples wasconfirmed with a commercially available Lyme disease im-munoassay (Diamedix Corp., Miami, Fla.).The full-length recombinant flagellin protein pl-336 was
reactive with 70 of the 83 Lyme disease sera in the IgGELISA (Table 1), whereas 66 serum samples reacted whenthe central portion of the flagellin, p137-262, was used as theantigen. The amino-terminal protein pl-137 was reactivewith 53 of the Lyme disease serum specimens, but only 39specimens demonstrated reactivity with the carboxyl-termi-nal protein p262-336. Protein p64-311, which is an extensionon each side of the central portion and does not contain theamino or carboxyl termini of the flagellin protein, wasreactive with 72 of the Lyme disease serum specimens.The patterns of ELISA IgG reactivity of the 83 Lyme-
positive serum specimens with the flagellin fusion proteinsare outlined in Table 2. Twenty-eight serum specimens werereactive with all of the flagellin regions, whereas the othersera failed to react with one or more of the regions. Nine ofthe specimens were unreactive with all of the flagellinantigens. Thirty-four serum specimens reacted with thefull-length flagellin recombinant protein (pl-336) and thecentral regions of the protein (pl37-262 and p64-311) but didnot react with the amino terminus (pl-137) and/or the car-boxyl terminus (p262-336). Four of the serum specimens thatwere unreactive with the full-length flagellin recombinant
TABLE 2. ELISA reactivity of IgG in Lyme disease sera withflagellin recombinant protein regions'
ELISA reactivity with flagellin fusion protein: No. of sera
pl-336 pl-137 p137-262 p262-336 p64-311 reactive
+ + + + + 28+ + + - + 20+ - + - + 9+ - + + + 5_ _ + - + 4+ + - + + 3+ + - - + 1+ - - + + 1+ - - - + 1+ + - + - 1+ - - + - 1_ _ _ - - 9
a Eight-three Lyme disease serum samples with IgG antibody reactivity toB. burgdorferi as determined by Western blotting.
J. CLIN. MICROBIOL.
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

LYME DISEASE-SPECIFIC B. BURGDORFERI FLAGELLIN REGIONS 633
TABLE 3. Serum IgM antibody reactivity with recombinantflagellin proteins
Serum No. No. (%) of sera reactive with fusion protein:specimen tested pl-336 pl-137 p137-262 p262-336 p64-311
Lyme disease 37 28 (73) 2 (5) 29 (78) 0 35 (95)Syphilis 26 0 0 0 0 0Normal 47 2 (4) 3 (6) 0 0 0
protein were reactive with regions p137-262 and p64-311,possibly because of serologically favorable conformations ofthese proteins compared with that of full-length flagellin. Thevariable central flagellin protein p137-262 was unreactivewith three specimens that reacted with all other constructs;however, each of these sera contained IgM antibodies thatwere reactive with this region. Three additional specimensthat were reactive with pl-336 and p64-311 were unreactivewith the more restricted region p137-262 but were not IgMreactive. Only two serum specimens were reactive with thefull-length protein but unreactive with p137-262 and p64-311in the IgG assay, yet both specimens contained IgM anti-body that was reactive with both proteins as well as thefull-length protein.The reactivity of each recombinant flagellin protein with
26 syphilis-positive serum samples and 47 normal serum
samples was also evaluated (Table 1). As predicted from thesequence homology with T. pallidum flagellin, many of thesyphilitic sera, all of which had fluorescent treponemalantibody absorption titers of R3+ to R4+, were reactivewith the amino and carboxyl termini of the flagellin as well aswith the full-length protein. Four of the syphilitic serumsamples were from persons diagnosed with primary syphilis;three of these specimens contained IgG antibodies that werereactive with the full-length flagellin. However, in contrastto Lyme sera reactivity, syphilitic sera were unreactive withthe central region defined by p137-262, indicating that thisregion of flagellin is specific for Lyme disease. Further, onlyone syphilis-positive serum sample, from a person withsecondary syphilis and a Venereal Diseases Research Lab-oratory titer of 1:32 and a fluorescent treponemal antibodyabsorption result of R4+, reacted with the p64-311 protein.This additional cross-reactive epitope(s) probably resides inthe region containing amino acid residues 64 through 137,since this syphilitic specimen cross-reacted with the amino-terminal protein pl-137. Central regions of the flagellin alsohave the advantage of being unreactive with normal sera
compared with the full-length protein. Reactivity of normalsera with the amino- and carboxyl-terminal proteins mayreflect the sequence homology in these regions of flagellinwith a wide variety of other flagellin proteins (9, 23).The reactivity of IgM antibodies in Lyme disease sera with
each recombinant flagellin protein also was evaluated. Thir-ty-seven of the 90 Lyme disease-positive serum samplesevaluated were found to be IgM positive by Western blot andcommercial (Diamedix) enzyme immunoassay. The reactiv-ities of these sera with recombinant antigens in the IgMassay indicated that the majority of the sera reacted with thecentral region of the flagellin (Table 3). The p64-311 proteinwas reactive with all but 2 IgM-positive serum samples, thep137-262 protein reacted with 29 specimens, and the full-length protein was reactive with 28 serum samples. Only twoserum samples contained IgM antibodies that were reactivewith the amino terminus of the flagellum, pl-137, and none ofthe sera was reactive with the carboxyl terminus, p262-336.
TABLE 4. ELISA reactivity of IgM in Lyme disease sera withflagellin recombinant protein regionsa
ELISA reactivity with flagellin fusion protein: No. of sera
pl-336 pl-137 p137-262 p262-336 p64-311 reactive
+ - + - + 21_ _ + - + 7_ _ _ - + 3
+ - - - + 2+ + + - + 1+ + - - + 1+ - - - - 1_
-_ - - 1
a Thirty-seven Lyme disease serum samples with IgM antibody reactivityto B. burgdorferi as determined by Western blotting.
These data indicate that the IgM response is directed prima-rily to the central region of the flagellin protein.Eleven of the IgM-positive serum samples were unreac-
tive with the full-length flagellin protein pl-336, yet 7 of thesesamples were reactive with the central region p137-262 and10 were reactive with p64-311 (Table 4). Six specimensdemonstrated IgM reactivity with p64-311 but not with thesmaller central region, p137-262. Conformational effects inthese proteins may explain this apparent lack of consistencyin serologic reactivity. The central flagellin regions werebetter serological targets than the full-length protein was forbinding of IgM. However, one serum sample was IgMreactive only with the full-length protein, and one IgM-positive serum sample was unreactive with all proteins.The central regions of flagellin offer the apparent advan-
tage of being unreactive with potential cross-reactive sera.None of the syphilitic sera was reactive with the recombi-nant flagellin proteins in the IgM assay, and only minorreactivity with proteins pl-336 and pl-137 occurred with thenormal sera.
DISCUSSION
In this paper we describe the cloning, expression, anddiagnostic utility of regions of the B. burgdorferi flagellinprotein, which may be employed to improve the specificityof detection of antibodies against B. burgdorferi. Flagellinelicits an early and strong immune response in infectedindividuals (6, 8). Hansen et al. (12) demonstrated that aflagellum-based ELISA was more sensitive and specific thanthe ELISA with a whole-cell sonic extract as the antigen inthe serodiagnosis of Lyme disease; this observation wasconfirmed by Karlsson et al. (13).The flagellin proteins from three strains of B. burgdorferi,
B31 (23), GeHo (9), and CA12 (7), have been cloned andsequenced. The amino acid sequences of the flagellin pro-teins from the European strain GeHo and the Americanstrain CA12 (California) are identical, whereas the Americanstrain B31 (New York) has alternate residues at positions 180(threonine for asparagine) and 279 (asparagine for asparticacid). Conservation of the flagellin sequence is one featurethat makes this protein an attractive target for use as anantigen in a serological assay. However, there also is con-siderable homology between the flagellin of B. burgdorferiand the flagellar proteins of other bacterial species, includingT. pallidum. Comparison of the amino acid sequence of theB. burgdorferi flagellin with that of the T. pallidum endo-flagellar FlaB2 protein reveals a high degree of homology,particularly at the amino and carboxyl termini. The amino
VOL. 31, 1993
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

634 ROBINSON ET AL.
terminus, pl-137, and the carboxyl terminus, p262-336,exhibit 52 and 54% homology, respectively, with those of theT. pallidum flagellar subunit; many of the identical aminoacids are in consecutive runs of five or six residues. Incontrast, the central region, p137-262, displays only 13%homology with few consecutive identical residues. Theregion designated p64-311 is a larger region displaying rela-tively low homology (30%) with the T. pallidum sequenceand minimal consecutive identical amino acid residues.The specificity of assays for the detection of antibodies to
B. burgdorfeni in patients with suspected Lyme disease maybe increased by using the central flagellin regions andomitting the cross-reactive amino- and carboxyl-terminalregions. Our IgG ELISA results indicate that the centralflagellin region (pl37-262) is unreactive with sera from pa-tients with syphilis. In contrast, 54 percent of the syphilis-positive specimens tested were reactive with the full-lengthflagellin protein, pl-336. As predicted from the amino acidhomologies, this reactivity was restricted to the amino-terminal region (pl-137) and the carboxyl-terminal region(p262-336) of the protein. Expansion of the central region(pl37-262) of flagellin to contain amino acids 64 to 311(p64-311) resulted in only 1 of the 26 syphilis specimensbecoming reactive in the IgG ELISA. Also, some normalsera cross-reacted with full-length (pl-336), amino-terminal(pl-137), and carboxyl-terminal (p262-336) regions of flagel-lin. One possible explanation is that some normal sera
contain antibodies to sequences shared among flagellar pro-teins from a variety of bacteria (9, 23) and that some of thesesera contained antibodies to oral treponemes associated withperiodontal infections (17). However, normal sera were
clearly differentiated from Lyme sera by using the p137-262and p64-311 proteins.
Wallich et al. (23) and Gassmann et al. (9) predictedthrough sequence analysis and reactivity with rabbit andmouse sera that the central regions of the flagellin proteins(amino acids 130 to 250 and amino acids 180 to 260, respec-
tively) were likely to be specific for the diagnosis of Lymedisease. Our results with human sera agree with thesepredictions and demonstrate that syphilitic sera and normalsera are unreactive with the region from amino acids 137 to262. Minor cross-reactivity with only one syphilitic serum
samples in the IgG ELISA when p64-311 was used as antigenindicates that the specific region of the B. burgdorferiflagellin may be larger than predicted. It is not likely thatthese regions would be able to distinguish Lyme diseasefrom relapsing fever caused by Borrellia hermsii, becausethe flagellin proteins from these organisms are 95% homol-ogous (9).
Sera from individuals with Lyme disease reacted broadlyagainst multiple flagellin regions in the IgG ELISA, incontrast to the amino- and carboxyl-terminal restricted re-
activity of syphilitic sera. However, the pattern of IgG
reactivity revealed that more of the Lyme disease sera
reacted with the central region (pl37-262) than with theamino- or carboxyl-terminal region. By extending the central
region of the flagellin to span amino acids 64 through 311, six
additional serum samples in addition to those that reacted
with the p137-262 protein were recognized as IgG positive.
Differences in the conformations of the fusion proteins or
variations in assay performance may explain some inconsis-
tencies in the patterns of serum IgG reactivity, such as the
four sera that were reactive with p137-262 and p64-311 but
unreactive with the full-length flagellin protein.Our data regarding IgG reactivity with regions of the entire
flagellin protein contrast with those of Collins and Peltz (7),
who found that only the amino-terminal portion of theflagellin defined by the first 90 amino acids of the proteinreacted with sera from Lyme disease patients. On the basisof these findings, they concluded that a specific assay forLyme disease based on the flagellin protein might not bepossible. The patient population used in their study was
restricted to those displaying arthritis, whereas the sera
evaluated in this study included various stages of the dis-ease. It is possible that the concentration of IgG directedagainst certain flagellin epitopes varies throughout thecourse of infection. These results need to be confirmed witha well-defined population of Lyme disease patients at vari-ous stages of disease.
Berland et al. (3) recently demonstrated through immuno-blot analysis of overlapping flagellin fragments that an IgGbinding immunodominant domain could be mapped to aminoacids 197 to 241 but that additional regions were immunore-active with Lyme disease-positive sera. They also showedthat there was no reactivity of normal sera in this immun-odominant region, but they did not evaluate syphilitic sera.Our results agree with their conclusions and indicate that wemay be able to increase Lyme serological assay specificitywithout compromising sensitivity by using the central flagel-lin protein p137-262 or p64-311.The central region of flagellin apparently elicits a strong
IgM response. Of the 37 Lyme disease IgM-positive serumsamples, 35 reacted with the p64-311 protein and 29 reactedwith p137-262. Conformational effects may explain thegreater IgM reactivities of the smaller central regions com-pared with that of the full-length protein. These data indicatethat the central region of the flagellin protein may be asignificant marker for the detection of early Lyme disease.This region also provides specificity, since none of thesyphilis positive sera or normal sera contained reactive IgM.The IgM titers to T. pallidum in these sera were notdetermined, so additional sera from patients with primarysyphilis and known IgM titers will be evaluated.The observation that the central domain of the flagellin
protein is the predominant epitope for the detection of IgMagrees with observations on the secondary structure of theflagellum filaments (18). X-ray fiber diffraction analysis ofthe secondary flagellin structure indicated that the terminalportions of the protein influence the structure of the filamentand are located toward the center of the filament, whereasthe central region of the protein is not involved in confor-mation and is located on the outside of the filament struc-ture. Wallich et al. (23) postulated that this exposed middledomain could function to elicit the immune response to B.burgdorferi.The utility of the central region of the B. burgdorferi
flagellin protein as an antigen in serologic assays for Lymedisease was evaluated. The benefit derived by the use of thisregion in the IgG ELISA assay was in increased specificity;there was little or no cross-reactivity with syphilitic ornormal sera. In the IgM ELISA, the central flagellin regiondisplayed greater reactivity with Lyme sera than the full-length protein did, leading to a more sensitive assay for thepossible detection of early disease. Additional Lyme dis-ease-specific proteins are currently being cloned and evalu-ated to complement the specific flagellin regions to detectLyme disease sera that are not reactive with the flagellinprotein.
REFERENCES1. Barbour, A. G. 1984. Isolation and cultivation of Lyme disease
spirochetes. Yale J. Biol. Med. 57:521-525.
J. CLIN. MICROBIOL.
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

LYME DISEASE-SPECIFIC B. BURGDORFERI FLAGELLIN REGIONS 635
2. Barbour, A. G. 1988. Laboratory aspects of Lyme borreliosis.Clin. Microbiol. Rev. 1:399-414.
3. Berland, R., E. Fikrig, D. Rahn, J. Hardin, and R. A. Flavell.1991. Molecular characterization of the humoral response to the41-kilodalton flagellar antigen of Borrelia burgdorfen, the Lymedisease agent. Infect. Immun. 59:3531-3535.
4. Boiling, T. J., and W. Mandecki. 1990. An Escherichia colivector for high-level production of heterologous proteins infusion with CMP-KDO synthetase. Biotechniques 8:488-490.
5. Burgdorfer, W., A. G. Barbour, S. F. Hayes, J. L. Benach, E.Grunwaldt, and J. P. Davis. 1982. Lyme disease-a tick-bornespirochetosis? Science 216:1317-1319.
6. Coleman, J. L., and J. L. Benach. 1987. Isolation of antigeniccomponents from the Lyme disease spirochete: their role inearly diagnosis. J. Infect. Dis. 155:756-765.
7. Collins, C., and G. Peltz. 1991. Immunoreactive epitopes on anexpressed recombinant flagellar protein of Borrelia burgdorfen.Infect Immun. 59:514-520.
8. Craft, J. E., D. K. Fischer, G. T. Shimamoto, and A. C. Steere.1986. Antigens of Borrelia burgdorfen recognized during Lymedisease. J. Clin. Invest. 78:934-939.
9. Gassmann, G. S., E. Jacobs, R. Deutzmann, and U. B. Gobel.1991. Analysis of the Borrelia burgdorferi GeHo fla gene andantigenic characterization of its gene product. J. Bacteriol.173:1452-1459.
10. Glover, D. M. 1985. DNA cloning, vol. 2. IRL Press, Oxford.11. Grodzicki, R. L., and A. C. Steere. 1988. Comparison of
immunoblotting and indirect enzyme-linked immunosorbent as-say using different antigen preparations for diagnosing earlyLyme disease. J. Infect. Dis. 157:790-797.
12. Hansen, K., P. Hindersson, and N. S. Pedersen. 1988. Measure-ment of antibodies against the Borrelia burgdorferi flagellumimproves serodiagnosis in Lyme disease. J. Clin. Microbiol.26:338-346.
13. Karlsson, M., G. Stiernstedt, M. Granstrom, E. Asbrink, and B.Wretlind. 1990. Comparison of flagellum and sonicate antigensfor serological diagnosis of Lyme borreliosis. Eur. J. Clin.Microbiol. Infect. Dis. 9:169-177.
14. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
15. Magnarelli, L. A., J. F. Anderson, and A. G. Barbour. 1989.Enzyme-linked immunosorbent assay for Lyme disease: reac-tivity of subunits of Borrelia burgdorferi. J. Infect. Dis. 159:43-49.
16. Magnarelli, L. A., J. F. Anderson, and R. C. Johnson. 1987.Cross-reactivity in serological tests for Lyme disease and otherspirochetal infections. J. Infect. Dis. 156:183-188.
17. Magnarelli, L. A., J. N. Miller, J. F. Anderson, and G. R.Riviere. 1990. Cross-reactivity of nonspecific treponemal anti-body in serological tests for Lyme disease. J. Clin. Microbiol.28:1276-1279.
18. Namba, K., I. Yamashita, and F. Vonderviszt. 1989. Structure ofthe core and central channel of bacterial flagella. Nature (Lon-don) 342:648-654.
19. Pallesen, L., and P. Hindersson. 1989. Cloning and sequencing ofa Treponema pallidum gene encoding a 31.3-kilodalton end-oflagellar subunit (FlaB2). Infect. Immun. 57:2166-2172.
20. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-termination inhibitors. Proc. Natl. Acad. Sci.USA 74:5436-5467.
21. Steere, A. C., R. L. Grodzicki, A. N. Kornblatt, J. E. Craft,A. G. Barbour, W. Burgdorfer, G. P. Schmid, E. Johnson, andS. E. Malawista. 1983. The spirochetal etiology of Lyme dis-ease. N. Engl. J. Med. 308:733-740.
22. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 76:4350-4354.
23. Wallich, R., S. E. Moter, M. M. Simon, K. Ebnet, A. Heiberger,and M. D. Kramer. 1990. The Borrelia burgdorfeni flagellum-associated 41-kilodalton antigen (flagellin): molecular cloning,expression, and amplification of the gene. Infect. Immun. 58:1711-1719.
24. Wilske, B., G. Schierz, V. Preac-Mursic, K. Weber, H. W.Pfister, and K. Einhiupl. 1984. Serological diagnosis of ery-thema migrans disease and related disorders. Infection 12:331-337.
25. Zdller, L., S. Burkard, and H. Schafer. 1991. Validity ofWestern immunoblot band patterns in the serodiagnosis ofLyme borreliosis. J. Clin. Microbiol. 29:174-182.
VOL. 31, 1993
on August 14, 2019 by guest
http://jcm.asm
.org/D
ownloaded from

JOURNAL OF CLINICAL MICROBIOLOGY, Sept. 1993, p. 2553-25550095-1137/93/092553-03$02.00/0
Letters to the EditorAnalysis of the Humoral Response to the Flagellin Protein of
Borrelia burgdorferi
We read with interest the article by Robinson et al.reporting cross-reacting and specific epitopes on the flagellarprotein of Borrelia burgdorferi (5). The authors have over-looked previous findings, including our own study, whichprecisely characterized the common and specific epitopes ofthe flagellar protein by using overlapping decapeptides (6)and which have been confirmed since by others usingrecombinant fragments (1, 3). This is especially unfortunatebecause the authors thus did not realize that they interruptedthe integrity of the N- and C-terminal cross-reacting epitopes(6) by their choice of fragments (5). In addition, by sera beingpreabsorbed with Escherichia coli cell sonicate (5), which isnot done in routine diagnostic studies, potential cross-reacting antibodies were excluded from their study, despiteknown sequence homologies between the flagellin of E. coliand that of B. burgdorfeni (2). Furthermore, the unspecificbinding of immunoglobulins to a region between amino acids(aa) 49 and 68 is already known (6) and can be blocked bypreincubation with ovalbumin (4, 6). The question posed byRobinson et al. regarding the regions recognized preferen-tially from sera from patients at different stages of Lymedisease has also been answered in our previous study: serafrom stage I and II patients recognize the epitope within aa151 to 166 but not the epitope within aa 205 to 226, which isthe preferential epitope recognized by sera of patients atstage III and the Borrelia-specific monoclonal antibodyH9724 (1, 6). We hope that Table 1, summarizing what isknown, will enable future investigators to extend the currentknowledge on B. burgdorferi epitopes. Nonetheless, weagree with Robinson et al. that an enzyme-linked immu-nosorbent assay based on the central region of the B.burgdorferi flagellin would substantially improve the serodi-agnosis of Lyme disease; however, we suggest excluding aa121 to 148 and 255 to 280, because the main cross-reactingepitopes of sera from healthy controls and patients withsyphilis are located in these regions.
TABLE 1. Characteristics of important epitopes of the flagellarprotein of B. burgdorderi
Characteristic(s) of aa Sequence ofepitopes B. burgdorferepltopes ~~~~~~~~flagellin
Causes unspecific binding of immunoglobulin ......... 49-68Cross-reacting, N terminal .................................. 121-148Recognized by stage I and II sera ........................ 151-166Borrelia specific, recognized by stage III sera ........ 205-226Recognized by mouse monoclonal antibody H9724.... 208-224Cross-reacting, C terminal .................................. 255-280
REFERENCES1. Fikrig, E., R. Berland, M. Chen, S. Williams, L. H. Sigal, and R.
Flavell. 1993. Serologic response to the Borrelia burgdorferiflagellin demonstrates an epitope common to a neuroblastomacell line. Proc. Natl. Acad. Sci. USA 90:183-187.
2. Gassmann, G. S., E. Jacobs, R. Deutzmann, and U. B. Gobel.1991. Analysis of the Borrelia burgdorferi GeHo fla gene andantigenic characterization of its gene product. J. Bacteriol.173:1452-1459.
3. Jiang, W., B. J. Luft, W. Schubach, R. J. Dattwyler, and P. D.Gorevic. 1992. Mapping the major antigenic domains of the nativeflagellar antigen of Borrelia burgdorferi. J. Clin. Microbiol.30:1535-1540.
4. Lange, R., H. Bocklage, T. Schneider, H. W. Kolmel, J. Heese-mann, and H. Karch. 1992. Ovalbumin blocking improves sensi-tivity and specificity of immunoglobulin M immunoblotting forserodiagnosis of patients with erythema migrans. J. Clin. Micro-biol. 30:229-232.
5. Robinson, J. M., T. J. Pilot-Matias, S. D. Pratt, C. B. Patel, T. S.Bevirt, and J. C. Hunt. 1993. Analysis of the humoral response tothe flagellin protein of Borrelia burgdorfen: cloning of regionscapable of differentiating Lyme disease from syphilis. J. Clin.Microbiol. 31:629-635.
6. Schneider, T., R. Lange, W. Ronspeck, W. Weigelt, and H. W.Kolmel. 1992. Prognostic B-cell epitopes on the flagellar proteinof Borrelia burgdorferi. Infect. Immun. 60:316-319.
Thomas SchneiderMedizinische KlinikDepartment of GastroenterologyKlinikum SteglitzFree University of Berlin1000 Berlin 45Germany
Robert LangeEmanuel KrankenhausDepartment ofRheumatology1000 BerlinGermany
Wolfgang WeigeltRobert Koch-Institut des Bundesgesundheitsamtes1000 Berlin 65Germany
Author's ReplyWe greatly appreciate the letter of Dr. Schneider et al.
regarding the specificity of epitopes located on the flagellinprotein of Borrelia burgdorferi, and we regret our failure toinclude their key study (6) in the discussion of our data.Their observation that major B. burgdorferi-specific epitopesreside within amino acids 151 to 166 and 205 to 226 of theflagellin protein correlates well with our observation that theflagellin polypeptide encompassing amino acids 137 to 262(pl37-262) is reactive with most of the Lyme disease serathat are flagellin reactive. Neither of the immunologicalcross-reactive regions identified in their study, those con-taining amino acids 121 to 148 and amino acids 255 to 280, isfully contained within the B. burgdorferi-specific flagellinfragnent, p137-262. We selected and cloned the p137-262region because of its lack of overall (13%) sequence homol-ogy with the Treponema pallidum endoflagellar subunitFlaB2 and because the homology is scattered throughout thesequence. On either side of the p137-262 polypeptide, the
2553
Vol. 31, No. 9

2554 LETT7ERS TO THE EDITOR
homology increases, as does the likelihood of immunologicalcross-reactivity.Our serological evaluation of the cloned full-length B.
burgdorferi flagellin and selected regions indicated thatsyphilitic and normal sera reactive with the full-length pro-tein also were reactive with either the amino-terminal (pl-137) or the carboxyl-terminal (p262-336) region but not withthe central region. We believe that these data agree with thedata of Schneider et al. The shared overlap between peptideamino acids 121 to 148 and fragment pl-137 and that betweenpeptide amino acids 255 to 280 and fragment p262-336 areregions of high homology with the T. pallidum flagellin andmay explain the immunological cross-reactivity of syphiliticand normal sera with these sequences.The inclusion of the Escherichia coli sonicate in the serum
dilution buffer was not included to preabsorb antibodiesdirected to E. coli flagellar proteins which might cross-reactwith B. burgdorfen flagellin. Rather, the sonicate was in-cluded as a precaution to absorb antibodies reactive with theCMP-2-keto-3-deoxyoctulosonic acid synthetase portion ofthe fusion protein or contaminating E. coli proteins orlipopolysaccharide remaining after the purification of therecombinant proteins. Experiments performed with cross-reactive sera demonstrated that the added sonicate did notplay a major role in improving assay specificity. Also, notethat the B. burgdorferi flagellin region p137-262 displays onlylimited homology with E. coli flagella, similar to that of T.pallidum (4).We previously demonstrated that the flagellin-specific
monoclonal antibody H9724 reacted with a synthetic peptiderepresenting amino acids 196 to 225 (5a). This peptide wasfound to react with many of the Lyme disease sera frompatients with late-stage disease, in agreement with Schneideret al. Berland et al. (1) also found that the immunodominantdomain comprising amino acids 197 to 241 was bound byserum samples from 17 to 18 patients exhibiting manifesta-tions of late-stage disease. In a subsequent study (3), theydetermined that the majority of sera from Lyme diseasepatients with arthritis and neuroborreliosis contained anti-bodies that bound to the region encompassing amino acids208 to 225. However, we also observed that patients witharthritic manifestations had antibodies reactive with the
region defined by amino acids 1 to 137, which agrees with theresults obtained by Collins and Peltz (2). Jiang et al. (5) alsodemonstrated that in addition to the dominant epitopes in thecentral portion of flagellin, additional Lyme serum-reactiveepitopes were identified in the amino-terminal region.The specificity of serodiagnosis of Lyme disease will
certainly be improved through the use of specific regions ofthe flagellin. The question remains regarding the ability ofsuch a target to detect all cases of Lyme disease. Additionalspecific B. burgdorferi proteins such as OspA, OspC, the39-kDa protein, and others will need to be- evaluated as totheir role in the diagnosis of Lyme disease.
REFERENCES
1. Berland, R., E. Fikrig, D. Rahn, J. Hardin, and R. A. Flavell.1991. Molecular characterization of the humoral response to the41-kilodalton flagellar antigen of Borrelia burgdorferi, the Lymedisease agent. Infect. Immun. 59:3531-3535.
2. Collins, C., and G. Peltz. 1991. Immunoreactive epitopes on anexpressed recombinant flagellar protein of Borrelia burgdorferiInfect. Immun. 59:514-520.
3. Fikrig, E., R. Berland, M. Chen, S. Williams, L. H. Sigal, andR. A. Flavell. 1993. Serologic response to the Borrelia burgdor-feri flagellin demonstrates an epitope common to a neuroblas-toma cell line. Proc. Natl. Acad. Sci. USA 90:183-187.
4. Gassmann, G. S., E. Jacobs, R. Deutzmann, and U. B. Goibel.1991. Analysis of the Borrelia burgdorferi GeHo fla gene andantigenic characterization of its gene product. J. Bacteriol.173:1452-1459.
5. Jiang, W., B. J. Luft, W. Schubach, R. J. Dattwyler, and P. D.Gorevic. 1992. Mapping of the major antigenic domains of thenative flagellar antigen of Borrelia burgdorfen. J. Clin. Micro-biol. 30:1535-1540.
5a.Robinson, J. M., and J. C. Hunt. Unpublished results.6. Schneider, T., R. Lange, W. Ronspeck, W. Weigelt, and H. W.
Kolmel. 1992. Prognostic B-cell epitopes on the flagellar proteinof Borrelia burgdorferi. Infect. Immun. 60:316-319.
John M. RobinsonJ. C. HuntInfectious Disease and ImmunologyAbbott LaboratoriesOne Abbott Park RoadAbbott Park, Illinois 60064
Use of Selective Buffered Charcoal-Yeast Extract Mediumfor Isolation of Nocardiae from Mixed Cultures
I read with interest the recent letter submitted by Boquestand Tosolini (3) describing their observation that strains ofNocardia asteroides grew well on selective buffered char-coal-yeast extract agar (sBCYE). In fact, the observationwas first published by us in abstract form in 1990 (5), and,later, in 1992 (4) as a note in the Journal of ClinicalMicrobiology. We failed to cite the subsequent study ofBoquest and Tosolini (1) in our paper because the journal inwhich they published was not included in our literaturesearch. The careful systematic evaluation of sBCYE forrecovery of nocardiae conducted by our laboratory and byBoquest and Tosolini (2) provided convincing evidence thatsBCYE could be used for the selective isolation of both
Legionella spp. and Nocardia spp. We pointed out in ourpaper, as did Boquest and Tosolini in their letter, that thelimited clinical experience with the use of Legionella-selec-tive media for isolation of N. asteriodes from sputum re-ported by Vickers et al. (6) demonstrated the practicalimplications of our observations.Our study was prompted by the observation that sBCYE,
used as part of our routine culture of bronchoalveolar lavagefluid, effectively recovered N. asteroides from mixed culturesfrom two immunocompromised patients with pulmonary no-cardiosis. In routine practice, sBCYE continues to performwell in our laboratory for the selective isolation of nocardiae,and I urge other laboratories to use it for this purpose.
J. CLIN. MICROBIOL.