Embed Size (px)
Citation preview

371OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANSRevista Chilena de Historia Natural77: 371-409, 2004
An overview of interactions among oceanography, marine ecosystems,climatic and human disruptions along the eastern margins
of the Pacific Ocean
Una visión general de las interacciones entre la oceanografía, los ecosistemas marinos y lasdisrupciones climáticas y antrópicas en los márgenes este del Océano Pacífico
PATRICIA M. HALPIN1,2, P. TED STRUB3, WILLIAM T. PETERSON4
& TIM R. BAUMGARTNER5
1Department of Zoology, Oregon State University, Corvallis, Oregon 97331, USA Current address2Marine Science Institute, University of California Santa Barbara,
Santa Barbara, California 93106, USA; e-mail: [email protected] of Oceanic and Atmospheric Sciences, Oregon State University, Corvallis, Oregon 97331, USA
4National Marine Fisheries Service, Northwest Fisheries Science Center,2030 South Marine Science Drive, Newport, Oregon 97365, USA
5Centro de Investigación Científica y Educación Superior de Ensenada, División de Oceanología,kilómetro 107, Carretera Tijuana-Ensenada, Ensenada, BC México
ABSTRACT
Physical processes, biotic processes and human activities all act to shape marine ecosystems. Futuremanagement of these valuable ecosystems requires an integrative approach that takes into accountcomplicated organismal interactions, oceanographic processes that span large scales of time and space, andhow anthropogenic effects interact with the natural environment. The temperate west coasts of North andSouth America represent a unique opportunity for comparison, as many of the same oceanographic processes,particularly upwelling, occur off both coasts. Additionally, a number of marine ecosystems in both areas havesurprising parallels. Here we present an overview of the oceanographic processes that dominate the coastlines,and give examples of ecosystems and the effects that oceanography, human activities and their interactionhave on the communities.
Key words: eastern boundary currents, anthropogenic effects, marine community ecology, fisheries,interdecadal variability.
RESUMEN
Los procesos físicos y biológicos y las actividades humanas determinan los ecosistemas marinos. El manejofuturo de estos valiosos ecosistemas requiere de una aproximación integrativa que considere las complejasinteracciones entre individuos, los procesos oceanográficos que se desarrollan a gran escala espacial ytemporal, y la interacción de los efectos antropogénicos con el medio ambiente. Las costas templadas deloeste de Norteamérica y Sudamérica representan una oportunidad única para efectuar comparaciones ya quemuchos procesos oceanográficos, particularmente la surgencia costera, ocurren en ambas costas.Adicionalmente, numerosos ecosistemas marinos en ambas áreas presentan sorprendentes paralelos. Lapresente revisión ofrece una visión general de los procesos oceanográficos dominantes en ambas costas yofrece ejemplos de ecosistemas y los efectos que la oceanografía, las actividades humanas y las interaccionesentre individuos tienen sobre las comunidades marinas.
Palabras clave: corriente de borde oriental, efectos antrópicos, ecología de comunidades marinas,pesquerías, variabilidad interdecadal.
INTRODUCTION
The western coastal regions off North and SouthAmerica contain some of the most productiveoceanic ecosystems in the world. In bothhemispheres, many livelihoods depend on avariety of marine products derived from fish,invertebrates, and algae. These products
contribute significantly to local and internationaleconomies, as well as local cultures.Considering only the fisheries, the combinedannual landing of pelagic fish off Peru and Chileduring good years ranges from 12-18 millionmetric tons (mmt), representing 15-20 % of theglobal total of approximately 80-90 mmt. Thisdirectly adds $0.5 to $1.0 billion (USD) to the

372 HALPIN ET AL.
economies of each country. Indirect or inducedimpacts on their economies increase the benefitof these resources by a factor of 5-10 (FAO1993, 1996). Although the catch from the UnitedStates west coast is much smaller(approximately 1 mmt), the United StatesDepartment of Commerce estimates its indirectimpact on local economies as approximately$4.0 billion USD (figures for 1991). Clearly,marine resources are important in bothhemispheres.
The functioning of coastal ecosystems isinextricably linked to oceanographic processesthat can vary across enormous spatial andtemporal scales. Scientists are only beginningto appreciate how larger scale ocean basinprocesses influence community dynamics.Additionally, while the community dynamics ofvery nearshore benthic ecosystems have beenactively researched, the dynamics of pelagicecosystems are generally unknown apart fromsome basic food chain structures and broadresponses to the physical environment. A poorunderstanding of how oceanographic processesinteract with ocean ecosystems in conjunctionwith generally ill advised fishery practices hasled to devastation of some fisheries. Recoveryand management of fisheries along thesecoasts, as well as a deeper comprehension ofhow these ecosystems function, must take intoaccount the oceanographic processes thatunderpin the biotic interactions.
Marine ecosystems must endure large-scalenatural perturbations in addition to humaninduced disturbance. Important among these areEl Niño and La Niña events (the two extremes
of the El Niño Southern Oscillation (ENSO)“cycle”), tidal and current changes induced bythe “Metonic” 18.6 year lunar cycle, the Pacificdecadal oscillation (PDO), and long termclimate change stemming from natural andanthropogenic sources (Fig. 1). All of theseprocesses alter the physical conditions underwhich marine communities function, yet wehave very incomplete information on theirimpacts. Management schemes that fail to takeinto account such cycles risk overexploitingpopulations during periods when naturalenvironmental stresses are at a maximum, tothe point where viable population size cannotbe maintained in the face of otherenvironmental stresses.
Here, we review the predominantoceanographic processes that characterize thewestern coasts of North and South Americaincluding a discussion of interannual andinterdecadal variability, where known. Next wediscuss rocky shores, kelp forests and pelagicfisheries as examples of how oceanic andoceanographic processes and humanexploitation impact coastal marine ecosystems.We examine several examples of the interactionbetween marine resource use and the naturalcycles of variability in populations. Althoughthe exact details of these disruptions are still amatter of debate and ongoing research, theirsevere socioeconomic consequences are clearand unquestionable. We conclude with adiscussion of the need to integrateoceanographic and ecologic perspectives andmethodology, in order to better understand,preserve and use marine resources.
Fig. 1: Diagram of some important oceanographic cycles and events from 1900 to 2000. Dark barsindicate El Niño, severe events are marked with an asterisk; open bars indicate La Niña. Sine waveindicates the phase of the lunar 18.6 year Metonic cycle. Sections marked “warm” versus “cold”indicate shifts in the Pacific interdecadal oscillation. Arrow symbolizes long term global climatechange and its recent acceleration due to anthropogenic sources.Diagrama de algunos ciclos y eventos oceanográficos de importancia desde 1900 a 2000. Las barras oscuras indican ElNiño, los eventos severos se marcan con asteriscos; las barras claras indican eventos La Niña. Las ondas de seno indican lafase del ciclo Metónico lunar de 18.6 años. Las secciones marcadas “cálido” versus “frío” indican cambios en la oscilaciónPacífica interdecadal. Las flechas simbolizan el cambio climático Global de largo plazo y su reciente aceleración debido afuentes antropogénicas.

373OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
OCEANOGRAPHY
Background: description of the systems
Along the western coasts of North and SouthAmerica the climate and environment isdominated by the California and the Peru-Chilecurrents, respectively, which form wide,meandering near-surface flows directed towardthe equator (recent reviews of the Californiaand Peru-Chile current systems are presented inHickey (1989) and Strub et al. (1998)). At thesurface, these two eastern boundary currents(EBCs) transport cooler, fresher, nutrient-richwaters toward the warmer, saltier and nutrient-depleted waters of the tropics. Polewardundercurrents occur in both systems,transporting warm, salty, oxygen-poor andnutrient-rich water to higher latitudes. Thelarge scale dynamics of coastal waters, drivenby coupling between the ocean and theatmosphere over the Pacific basin, determineregional and local dynamics of these currents.Upwelling of colder, nutrient-rich water, afundamental process within these currents,drives the high productivity of nearby coastalecosystems. Upwelling occurs at the coast andalso in the offshore regions of the currents(within several hundred kilometers of thecoast), driven by winds and the internaldynamics of meanders and eddies within thecurrents. In turn, upwelling affects the large-scale winds through the interplay among theocean surface temperature, land surfacetemperature, and the overlying atmosphericwind and pressure systems. These easternboundary currents thus play a significant role incontrolling the large-scale climatic conditionsthat affect both the oceans and the continents ofNorth and South America. Because of theirproductivity, they contribute a significantportion of the world’s supply of protein andother marine products, through the harvest oftheir living marine resources.
The basin-scale circulation features thatimpact EBCs include equatorial currentsystems at low latitudes, west wind driftcurrents at mid to high latitudes and deep oceaneddies along the offshore edge of the systems atmid latitudes. In the California current, theseasonal EBC itself is believed to create theseeddies (Strub & James 2000) which may helpto retain zooplankton and fish larvae near thecoast. Much of the variability in the Peru-Chilecurrent system, may be derived from coastallytrapped waves with an equatorial origin.
In all EBCs a poleward undercurrent movesover the continental slope (Neshyba et al.
1989). Direct measurements of the polewardundercurrent off North America show that itflows continuously from southern California(33º N) to the British Columbia coast (51º N)(Pierce et al. 2000). The undercurrent off SouthAmerica has been traced using water masscharacteristics from off Peru (5º S) to southernChile (48º S) (Silva & Neshyba 1979).
The undercurrent potentially performsseveral roles in population regulation. It isoften considered the source of the nutrientsdrawn to the surface by upwelling, providingthe basis for new production. Zooplankton andfish larvae can also use the poleward flow tomaintain their latitudinal positions in anotherwise equatorward surface flow via verticalmigration (Peterson 1998). In contrast, thepoleward undercurrent off Peru and Chiletransports water with extremely lowconcentrations of oxygen which may harmmarine life when brought to the surface next tothe coast during strong upwelling events.
The physical environment and coastalecosystems in the Pacific EBCs respond stronglyto climatic changes on a variety of scales.Responses to the basin-scale ENSO fluctuationsare well documented (Chelton 1982, Barber &Chavez 1983, Wooster & Fluharty 1985, Castillaet al. 1993, Lubchenco et al. 1993). Recently,evidence for the sensitivity of these systems tointerdecadal variability, with time scales of 50-60 years, has become equally convincing. This isparticularly clear for the north Pacific basin,where the well observed basin-wide climaticshift in the mid-1970s resulted in reorganizationof heat storage in the ocean (Hahn & Denny1989, Miller et al. 1994, Miller 1996), in thecirculation in the atmosphere, and thesubsequent wind-driven circulation in thesurface ocean affecting the oceanic ecosystemsat trophic levels from phytoplankton to salmon(Polovina et al. 1995, Brodeur et al. 1996,Mantua et al. 1997). The eastern Pacific marginexperienced a warming that stretched along thecoast from southern California into the Gulf ofAlaska, with a concurrent cooling in the centralnorth Pacific. Graham’s (1994) analysis suggeststhat this decadal scale change originated in thetropics and reached the North Pacific throughatmospheric interconnections. Similaritiesbetween the surface temperature and atmosphericpressure patterns associated with ENSO and withinterdecadal variability in the North Pacific havebeen noted by several studies (Graham 1994,Mantua et al. 1997, Enfield & Mestas-Nunez1999).
Both of these EBC systems are forced by acombination of “local” winds and basin-scale

374 HALPIN ET AL.
currents. By “local” winds, we mean the large-scale wind fields directly over the EBCs, notingthat currents at a given location are actuallyforced by wind stress at locations 300-500 kmaway in the equatorward direction (Gill &Clarke 1974, Denbo & Allen 1987). At mid-latitudes, these winds result from offshore high-pressure systems found over the eastern sides ofthe ocean basins that drive equatorward,upwelling-favorable winds along the coast. Inspring and summer, the high pressure systemsare larger and stronger and often interact with“thermal” low-pressure systems over thecontinents to create a seasonal intensificationand expansion of the equatorward wind regime.At lower latitudes, the trade wind systemassociated with the seasonally migrating inter-tropical convergence zone (ITCZ) is moreimportant in the northern Peru-Chile current,creating an austral winter maximum inupwelling off Peru. At higher latitudes, stormsdrive poleward (downwelling-favorable) windsthat peak in winter in each hemisphere.
The 18.6 year lunar cycle or Metonic cycle,has been scrutinized as a long periodoceanographic cycle that may be biologicallyrelevant (Royer 1993, Beamish 1995). TheMetonic cycle is driven by the change in theangle of the lunar orbit with the plane of theearth’s rotation about the sun. The result is an18.6 year cycle affecting tidal currents. Duringtimes of low declination, the differencebetween extreme, “spring,” high and low tidesis greater and the difference between neap tidesis less (Denny & Paine 1998). While themagnitude and effects of the lunar cycle remainunclear and controversial, the cycle has beenimplicated as an influence on a number ofclimactic (Maksimov & Smirnov 1967, Loder& Garrett 1978, Currie 1993) and biotic (Currie1991, 1992, Beamish 1995) variables.Particularly controversial is the interaction oftropical systems with the lunar cycle. Tropicalatmospheric mechanisms such as the SouthernOscillation seem to drive much of the globalatmospheric and oceanic variation, but lunarinfluences may most strongly affect higherlatitudes (Ware 1995). The extent to whichcelestial phenomena influence long-periodoceanographic events and how thesemechanisms interact with each other remain tobe investigated.
Northeast Pacific: the California currentsystem
The California current system is the mostthoroughly studied of the EBCs (Hickey 1979,
Chelton 1981, Chelton et al. 1982, Lynn &Simpson 1987, Hickey 1989, 1998, Strub &James 2000). There are still questions, however,about even the large-scale circulation patterns(Brink & Cowles 1991, Strub et al. 1991).
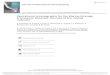
The mid-summer circulation of the EBC offthe coast of western North America oftenconsists of a convoluted, meandering,equatorward jet that flows from the Canadianborder to the southern California bight (Fig. 2,Lynn & Simpson 1987, Strub & James 1995,Strub & James 2000). The equatorward jet firstappears in spring, offshore of cold water thatoften forms cyclonic vortices in satellite SSTimages. These colder patches of upwelled waterappear to be separately created at the prominentcapes off northern California in the spring(April-May), joining later to form a scallopedband all along the coast (Fig. 2A). The mannerin which the system evolves from isolatedupwelling centers to a continuous band ispoorly understood. By June, the scalloped bandextends northward to central Oregon andbecomes a very narrow band next to the coast(Fig. 2B), consistent with the narrow (10-40km) spring-summer upwelling system offOregon and Washington documented in the1970’s. By July, the band becomes continuousas far north as Canada, separating from thecoast near Cape Blanco, 43º N (Barth & Smith1998, Barth et al. 2000), before meanderinghundreds of kilometers offshore of California.
In boreal fall and winter the jet continuesmoving offshore, losing its continuity aspoleward flow (the Davidson current) replacesthe equatorward current next to the coast (Fig.2A and 2D). In winter the jet leaves an offshoreeddy field that dissipates partially by spring,when the cycle begins again. Eddies from theprevious year are still present offshore whenthe jet moves into that region in mid-summer,providing some interaction between theambient eddy field and the evolving jet.
The seasonal jet also transports nutrientsand biological materials far offshore. Strub etal. (1990) used satellite color (CZCS) data toshow that the surface pigment field movesoffshore in fall and winter with the jet and eddyfield, leaving a wide region of moderately highchlorophyll concentrations. Whether offshorephytoplankton populations are primarilyconsumed by higher trophic levels or simplyadd carbon to ocean sediments remains an openquestion (Thomas et al. 1994).
The southern California bight creates abreak in the California current, caused by thechange in coastal orientation. Winds andupwelling weaken in the bight, where a
375OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
Fig. 2: A conceptual diagram of the seasonal evolution of the circulation in the California currentsystem (after Strub & James 2000).Diagrama conceptual de la evolución estacional de la circulación en el sistema de la corriente de California (en base aStrub & James 2000).

376 HALPIN ET AL.
cyclonic (counter-clockwise) circulation formsin summer. This cyclonic gyre, coupled withstrong stratification and calm winds, creates aretention center that favors larval fish survivaland is used as a “nursery” area for manyspecies of fish. Off Baja California, upwellingfavorable winds blow for the entire year,although weaker than summer winds off centraland northern California. The California currentturns offshore between 10-20º N, into thewestward North Equatorial current (NEC).Flow along Central America is poleward in theCosta Rica current and at a maximum insummer.
Although several mechanisms contribute tointerannual variability in the California currentsystem, much of the variability is related to theENSO cycle in the equatorial Pacific. El Niñoaffects the basin circulation directly throughpropagation along the equatorial and coastalwave guides, followed by Rossby wavepropagation away from the coasts. This is oftencalled the “oceanic pathway” of ENSO effectson the Northeast Pacific circulation. The ENSOcycle also affects the mid latitude oceansindirectly through changes in strength andposition of the mid and high lati tudeatmospheric pressure systems and the windsthey drive directly over the California current.This is often called the “atmospheric pathway”for ENSO effects.
Variation in the strength of the Californiacurrent affects the biota off western NorthAmerica. A series of analyses of the first 30years of California Cooperative OceanicFisheries Investigations (CalCOFI) data (1950-1980) found that zooplankton biomass in thesouthern California current system waspositively correlated with the strength of theequatorward transport. Periods of reducedtransport, and so reduced zooplankton biomass,were primarily coincident with warm phases ofthe ENSO cycle (El Niño) in the easternEquatorial Pacific (Chelton 1981, Chelton et al.1982). Advection of nutrients from the Alaskangyre via the California current may allowincreased primary productivity, which in turnsupports the increased zooplankton biomassobserved during periods of stronger flow(Wickett 1967). This hypothesis stemmedinitially from observations that the water wasoften cooler and fresher than locally upwelledwater during periods of greater equatorwardadvection during the cool phase of the ENSOcycle, the La Niña periods.
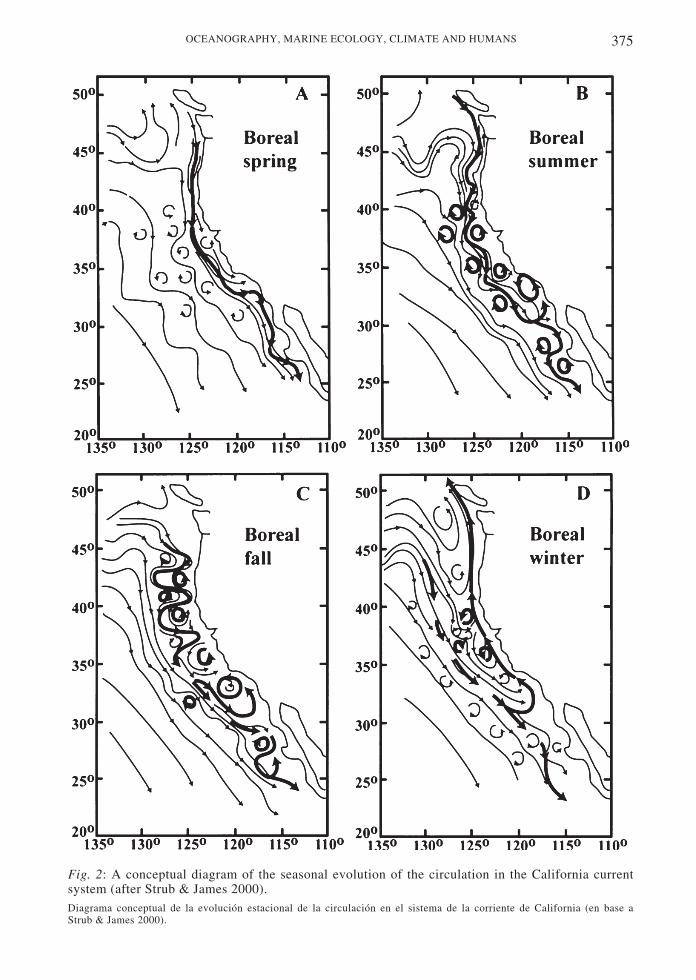
Changes in the strength of the transport inthe California current were hypothesized to berelated to changes in the position of the west
wind drift. Chelton & Davis (1982) used tidegauge sea level height data to show thatvariations in sea level and in alongshore coastalcurrents covaried in phase from southernCalifornia around the Gulf of Alaska. Since thecurrents in the eastern margin of the Alaskagyre and California current flow in oppositedirections, the strengths of the boundarycurrents in the two gyres were hypothesized tocovary out of phase, with the poleward flow ofthe Alaska gyre exhibiting increased flow whenthe equatorward California current exhibiteddecreased flow. Years with reducedequatorward/increased poleward flowcorresponded to warm phases of the ENSOcycle. Chelton & Davis (1982) hypothesizedthat this relation was due to interannual shiftsin the position of the west wind drift, whichfeeds both boundary currents, increasing theflow into one or the other of these systems in agiven year (Fig. 3). Thus, hypothetically,changes in the flow, water mass characteristicsand zooplankton biomass in the Californiacurrent depends on variability in the position ofthe west wind drift, with a connection to theinter-annual ENSO cycle through atmosphericforcing.
An alternate source of the cooler, fresherwater found in some years off southernCalifornia is the seasonal jet (Fig. 2)connecting the flow along the coasts of Oregon,Washington, and British Columbia (Strub et al.1991, Smith 1992, Strub & James 1995, Strub& James 2000, 2002a, 2002b). Upwelled waterin the coastal region of the Pacific Northwest iscold, rich in nutrients, and fresher than wateroff southern California, due to discharges outof the Columbia River, the Juan de FucaStraights and the Gulf of Alaska coast.Altimeter and drifter data show a directconnection between the coastal ocean offBritish Columbia and the Pacific Northwest,providing an even fresher source of water at thenorthern end of the seasonal jet shown in Fig.2. The jet moves offshore around 42º N (Barth& Smith 1998, Barth et al. 2000) and feedsdirectly into the core of the California current,while drifters released in the west wind driftgenerally stay offshore of the core of thecurrent (Mcnalley 1981). Thus the source of theanomalous water noted by Chelton et al. (1982)and others may be local to the boundarycurrents, upstream of the observations, withlittle connection to the west wind drift .Interannual variability in the seasonal jet isaffected by winds over the California current,which again are linked to interannual ENSOvariability.
377OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
A third source of interannual variability inthe California current is the wind-drivenonshore Ekman flow at the surface. Duringsome El Niño years, boreal winter stormactivity strengthens along the coast, drivingmore water onshore from the deep ocean,raising sea levels, deepening the mixed layerand warming the surface temperatures of theCalifornia current (Simpson 1984). A fourthsource of variation potentially arises from thedirect connection between the Californiacurrent, the equatorial ocean, and the ENSOcycle via the coastal waveguide along themargins of the Pacific, the oceanic pathway(Chelton & Davis 1982). Tide gauge dataclearly shows the propagation of interannualvariability in sea level from the equator to theCalifornia current and beyond (Enfield & Allen1980, Chelton & Davis 1982). Pares-Siera &O’Brien (1989) demonstrate through a
numerical model that much of the interannualvariability in the flow in the California currentpropagates into the region along the coast fromthe south, with an equatorial source linked tothe ENSO cycle.
The difficulty in determining the relativestrengths of these four processes in creating theobserved interannual variabili ty in theCalifornia current lies in their correlation withthe ENSO variability. This creates difficultiesin separating their importance using linearstatistical analyses. During the 1997-1998 ElNiño, however, altimeter data, wind fields andin situ measurements clearly document thetemporal development of surface circulationvariability during one strong event.
Two periods of high sea level occurred atthe equator, one in May-July 1997 and theother in October-January 1997-1998. High sealevels and poleward surface transports quickly
Fig. 3: Winter mean sea level pressure. Alternate states of oceanic circulation patterns in the NorthPacific Ocean (after Emery & Hamilton 1985, Hollowed & Wooster 1992, de Brodeur et al. 1996).Promedio de invierno de la presión a nivel del mar. Estados alternativos de los patrones de circulación oceánica en el nortedel Océano Pacifico (en base a Emery & Hamilton 1985, Hollowed & Wooster 1992, de Brodeur et al. 1996).

378 HALPIN ET AL.
propagated south (poleward) along the coast ofSouth America in May-June 1997, aided bypoleward winds during austral fall. The samesignals propagated north along the coast ofCentral America, but stalled near the mouth ofthe Gulf of California. During July, the firstsignal of raised sea levels appeared in thesouthern Gulf of California, then moved slowlyup Baja California, opposed by equatorialwinds during boreal summer. During thesecond period of equatorial high sea levels, thewinds reversed off North America, becomingpoleward (north) during boreal fall and winter.Subsequently, high sea levels and polewardtransports propagated quickly off the coast ofNorth America and around the Gulf of Alaskain October and November 1997. Thus, thesesignals had an equatorial origin but were alsoaided by local winds. As to connections to thewest wind drift , the poleward boundarycurrents intensified in fall, followed by anintensification of the entire Alaska gyreincluding the northern half of the west winddrift. This produced a circulation anomalysomewhat similar to that proposed by Chelton& Davis (1982). The sequence of events makesit clear, however, that the strengthened northernhalf of the west wind drift is a result of thebasin scale wind forcing and follows thechanges in the boundary currents, rather thandriving them (Strub & James 2002b).
The movement of the west wind drift hasalso been put forward as a possible mechanismfor creating the interdecadal changes in theboundary currents. Fig. 3 shows the generalpattern of hypothesized changes in atmosphericforcing (a stronger or weaker Aleutian lowpressure system) and oceanic response (moreflow into the California current or coastalAlaska current) conjectured to be responsiblefor interdecadal changes in zooplankton andfish abundance (Hollowed & Wooster 1992,Brodeur et al. 1996). This mechanism forinterdecadal change also remains a topic ofongoing research, using both historical data inretrospective studies and coupled atmosphere-ocean models to investigate the dynamics(United State GLOBEC 1996).
On smaller scales in nearshore ecosystems,an unknown but potentially importantcomponent driving nearshore ecosystemdifferences is variability at the “mesoscale”(i.e., tens to hundreds of kilometers). Menge etal. (1996, 1997a, 1997b) demonstrated thatchlorophyll “a” (Chl-a) concentrations differconsistently between onshore sites along theOregon coast. Sampling at spatial scales of 1-100 km, and temporal scales of hours to weeks,
they established that the 10-100 km scale wasthe most useful at explaining variability in Chl-a concentrations. Temporally, daily samplingwas sufficient to capture changes in waterquality. Onshore Chl-a concentrations peakafter upwelling events when phytoplanktonconcentrated at the upwelling front drift towardshore. Benthic diatoms may bloom in responseto upwelling and contribute to measured Chl-alevels. Study sites spaced 50+ km apartconsistently exhibited different concentrationswhen sampled from shore. Several years of dataindicate that either phytoplankton are deliveredonshore in different concentrations or thecondition for phytoplankton production at theoffshore upwelling front varies along the coastin a predictable fashion. In addition to Chl-a,recruitment of mussels varies along the coast ina similar, predictable fashion, providing furtherevidence of biologically importantoceanographic processes at work. The exactoceanographic mechanism of these onshoremesoscale differences remains to be elucidated.The biological ramifications of these conditionsare discussed in more detail below.
Southeast Pacific: the Peru-Chile currentsystem and its Equatorial connection
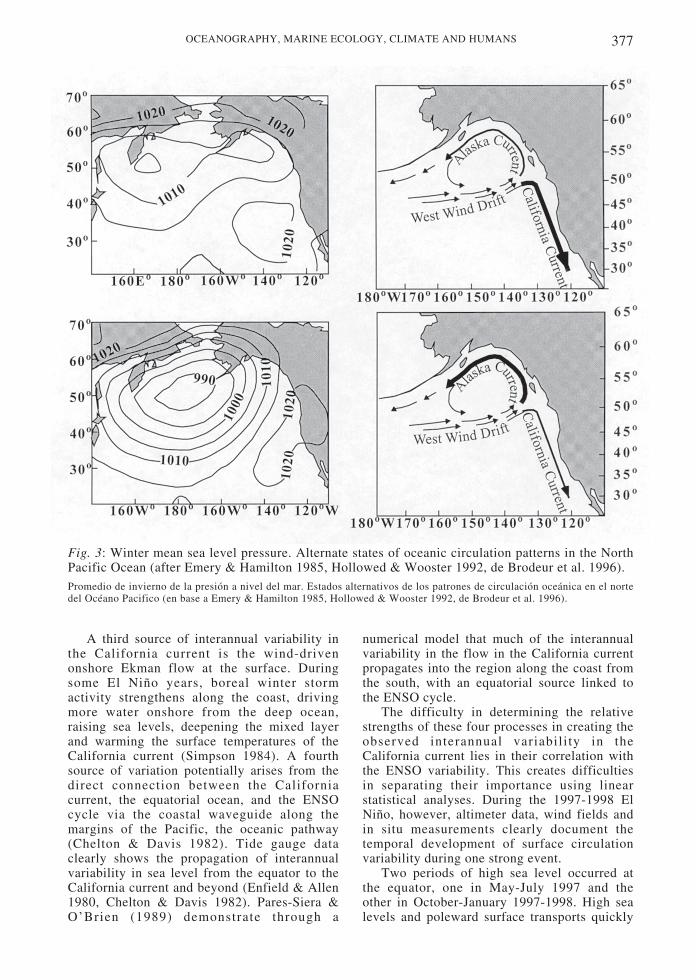
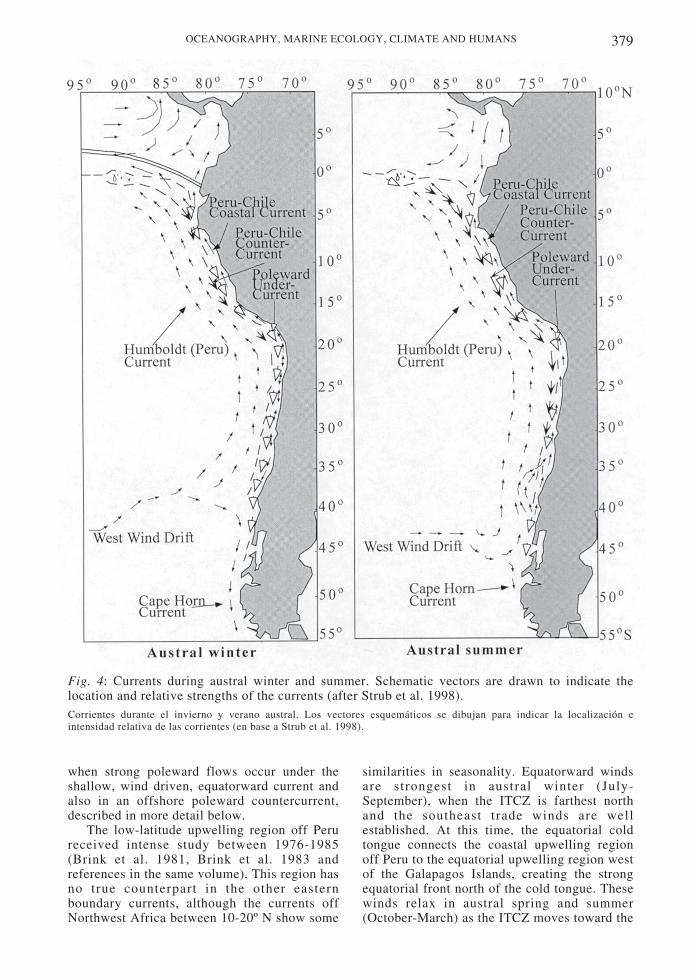
The Peru-Chile current system has not beenstudied in as much detail as the Californiacurrent, especially off Chile (Strub et al. 1998).This system has the greatest latitudinal extentof any EBC, stretching from the equator to 55˚S (Fig. 4). The equatorward-flowing portion ofthe current, however, only extends as far southas approximately 42-45o S in austral summerand 32-35o S in winter. South of 40-45o S, thepoleward Cape Horn current is found, driven bypoleward, downwelling-favorable winds andstrong buoyancy forcing by heavyprecipitation, similar to the coastal regions ofthe northeast Pacific along British Columbiaand Alaska.
The continuous equatorward current thatconnects the regions of Chile and Peru to thewestward south Equatorial current differs fromthe flow in the California current, which turnswestward far north of the equator. The directconnection between the equatorial currentsystems and the Peru-Chile current provides itsmost unique aspect-a prevalence of polewardflow that affects it from 5o S to 35o S or farthersouth. This is especially important during ElNiño, when higher sea levels and warmercoastal temperatures are found from north ofthe equator to approximately 33-36o S. Theconnection also influences normal periods,
379OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
Fig. 4: Currents during austral winter and summer. Schematic vectors are drawn to indicate thelocation and relative strengths of the currents (after Strub et al. 1998).Corrientes durante el invierno y verano austral. Los vectores esquemáticos se dibujan para indicar la localización eintensidad relativa de las corrientes (en base a Strub et al. 1998).
when strong poleward flows occur under theshallow, wind driven, equatorward current andalso in an offshore poleward countercurrent,described in more detail below.
The low-latitude upwelling region off Perureceived intense study between 1976-1985(Brink et al. 1981, Brink et al. 1983 andreferences in the same volume). This region hasno true counterpart in the other easternboundary currents, although the currents offNorthwest Africa between 10-20º N show some
similarities in seasonality. Equatorward windsare strongest in austral winter (July-September), when the ITCZ is farthest northand the southeast trade winds are wellestablished. At this time, the equatorial coldtongue connects the coastal upwelling regionoff Peru to the equatorial upwelling region westof the Galapagos Islands, creating the strongequatorial front north of the cold tongue. Thesewinds relax in austral spring and summer(October-March) as the ITCZ moves toward the

380 HALPIN ET AL.
equator, allowing the cold tongue to collapseand a warm tongue to develop that reachesfrom the equator to central Chile, offshore ofthe narrow upwelling region (Strub & James1995). At the same time (austral spring-summer), equatorward winds intensify offcentral Chile and upwelling occurs in episodeswith time scales of 7-10 days. Upwelling ismost intense at upwelling centers near smallcapes and bays, where filaments of cold waterexpand offshore and then retreat as the windpulses. The offshore size of these filaments isless than in the California current: 25-50 km offPeru (Brink et al. 1981) and 60-125 km offChile (Fonseca & Farias 1987).
The region most equivalent to the large,seasonal, meandering jet in the Californiacurrent system occurs between approximately35-44o S, remaining closer to the coast than offCalifornia. Surface salinities from a summercruise in 1960 (Brandhorst 1971) indicate alarge plume of less saline water extendingoffshore around 42-44o S, arcing back onshorenear 35o S. Inshore of this fresh plume is aregion of higher salinities next to the coastbetween 38-35o S, indicating the area ofupwelling and offshore Ekman transport. Aseparate tongue of salty water flows from thenorth to around 33-35o S. This may be thesouthern end of the Peru-Chile countercurrent.The arcing summer flow can be seen in Geosataltimeter height fields (Strub et al. 1998),appearing much smaller than that found offCalifornia. The strongest and most continuousfeature in the altimeter fields, however, is notthe expected equatorward flow, but thepoleward offshore countercurrent. Both griddedheights from Geosat and crosstrack geostrophiccurrents from the Geosat and TOPEX/POSEIDON altimeters show a band ofpoleward current, 100-300 km offshore, thatreaches from 6-8o S to 35o S (or farther),maximum in spring and minimum (or reversed)in fall (Strub et al. 1995). This offshore Peru-Chile countercurrent (Fig. 4) is hypothesized toconnect to the Equatorial undercurrent-(Tsuchiya 1985, Lukas 1986). It is also evidentbetween 100-300 km offshore in thehydrographic surveys of Huyer et al. (1991) offPeru. Off Chile, Fonseca (1989) refers to aband of poleward currents between 100-200 kmoffshore as one of the features found mostconsistently in repeated hydrographic surveys.Thus, although the altimeter cannot determineabsolute velocities, Strub et al . (1995)hypothesize the countercurrent to be presentmuch of the year and maximum in australspring.
The large “bight” off southern Peru andnorthern Chile (where the coastline changesorientation) may be the analog to the southernCalifornia bight. A recent climatology for theregion between 18-24o S (Blanco et al. 2001)shows that winds are weaker than in thesurrounding regions; the circulation is weak;there is an anticyclonic gyre in summer; andstratification is strong in the upper watercolumn. The weak, possibly recirculating, flowwould provide the mechanical means forretention of larval fish, as in the southernCalifornia bight. The large distribution of smallpelagic fish off northern Chile suggests that thepelagic ecosystems of the two areas may besimilar.
The narrowness of the South Americanupwelling system is supported by theclimatology of satellite and in situ SST and bysatelli te estimates of surface pigmentconcentrations (Thomas et al. 1994). Althoughsampling and algorithm problems may affectthe absolute values of satellite-derived surfacepigment concentrations in the Peru-Chilesystem, the spatial patterns are consistent withother evidence that implies a much narrowerupwelling system and a summer jet that stayswithin 200 km of the coast between 35-45o S.Upwelling-favorable wind forcing is alsoweaker off Chile than off California. Thus,some of the major unanswered questions aboutthe circulation concern the relation between thevarious physical components of this system: theoffshore countercurrent, the wind forcing, thewidth and alongshore extent of the upwellingsystem.
Much of the interannual variability of thissystem is driven directly by the ENSO cycle,especially north of 35o S (Enfield et al. 1987,and references in the same volume). Fonseca(1985) found a strong El Niño signal in 1982-1983 coastal SSTs to extend to 35o S but notfarther. Coastal tide gauges show the El Niñosignal to propagate to at least 35o S (Blanco etal. 2002). High sea levels and polewardcurrents found south of 35o S are likely to beinfluenced by winds associated with ENSOeffects (as in the northern hemisphere), but thishas not been systematically explored. Likewise,changes in the variability caused by changes inthe west wind drift are a possibility that has notbeen investigated. However, several recentstudies have identified coastal-trapped waveswith periods of approximately 50 days offnorthern Chile (Shaffer et al. 1997). Thesewaves arise equatorially and are hypothesizedto biologically important due to theirmodulation of coastal upwelling. Their effects

381OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
are currently under investigation. Theavailability of new satellite ocean sensors andthe increased sea-going capabilities of theChilean oceanographic community shouldclarify many of these issues over the nextdecade. Likewise, the presence of interdecadalvariabili ty in ocean conditions and fishpopulations has yet to be addressed in theSoutheast Pacific ocean. Inter- decadalvariability in fish catch is well documented, butis confounded by changes in effort to catchdifferent species.
Recent data sets from Chile demonstratenearshore variation in phytoplanktonconcentrations at the scale of 15-20 km.Despite large temporal variation, spatialvariation in concentrations appears dependenton position relative to upwelling regionscreated by local geography. Water at theupwelling center site was colder with lowerphytoplankton concentration (Wieters et al.2003).
Major unanswered questions remain aboutmarine productivity in North and SouthAmerica. Even when the Peruvian fishery isexcluded (there is no equivalent region offNorth America), the Chilean total fish catch islarger (approximately 6 mmt) than that offwestern North America (approx. 1-2 mmt). Inthe past several years (prior to the 1997-1998El Niño), northern Chile has averaged 2 mmt ofanchovy harvests, implying a much largerstanding stock than traditionally found offNorth America. Is this because of a greaterlevel of primary productivity, higher rates oflarval survival in the larger “bight,” or otherfactors? Initial estimates of phytoplanktonconcentrations from the CZCS satellite colorsensor indicated much lower levels of pigmentconcentrations off northern Chile than offCalifornia (Thomas et al. 1994). A great deal ofwork remains to be done to understand thedifferences between these pelagic ecosystems.
EXAMPLES OF MARINE ECOSYSTEMS AND THEIR
INTERACTIONS WITH HUMAN AND
OCEANOGRAPHIC IMPACTS
Rocky intertidal ecosystems: common features
Exposed rocky shores in the temperate regionsof both North and South America containextensive and actively researched benthicecosystems, due to their accessibility and thelow mobility of the organisms that make up thecommunity. Intertidal communities aregenerally space limited with invertebrate filter
feeders and macroalgae forming the primarycover. Barnacles and algae typically occupy thehighest intertidal areas, followed by mussels inthe mid-intertidal zone, and a combination ofinvertebrates and macroalgae in low to subtidalareas. However, the distribution and abundanceof primary cover changes with biotic andabiotic conditions of the area.
Both macroalgae and invertebrate primaryspace occupiers, usually barnacles or mussels,can be considered basal trophic species in theseecosystems, though filter feeders are notautotrophs (Menge et al. 1999). Since filterfeeders consume plankton and dissolvedorganic matter within the water column, they,like algae, transfer carbon from outside sourcesto within the community. In many intertidalareas they may be the principal agents of thistransfer. Also, as dominant space occupiers,benthic filter feeders provide the basic spatialstructure of the community in much the samefashion as terrestrial trees and subtidal giantkelps (Lohse 1993). Therefore, the productivityand biomass of rocky intertidal communitiesdepend in part on the properties of the waterflowing over both algae and invertebrates athigh tide, though the measure of water qualitywill be different for each group.
Generally, the animals in the upper trophiclevels exerting the strongest communityeffects are invertebrates such as seastars andmuricid gastropods (Paine 1966, 1974, 1976,Moreno et al. 1986, Castilla & Paine 1987).While birds and fish can alter the populationstructure of their prey, they have not beendemonstrated to inf luence communitystructure to the degree that invertebratepredators do (but see Frank 1982, Hahn &Denny 1989, Wootton 1997). The Chileanclingfish or pejesapo, Sicyases sanguinus, maybe an exception to this (Paine & Palmer 1978,Cancino & Castilla 1988). Pejesapos consumealgae and sessile invertebrates off rocks viascraping, and use specialized pelvic fins thatfunct ion as suct ion cups to remain inextremely exposed, wave-swept environments.Whether the Chilean clingfish can function asa keystone predator remains unknown, as thefish inhabit vert ical walls in extremelyturbulent wave surge channels that prohibitsafe experimentation. The primary grazers inintertidal communities are molluscs such aslimpets, snails and chitons, but also seaurchins in low intertidal to shallow subtidalareas.
Intertidal research has traditionallyoverlooked the influence that biologicaloceanographic conditions can have on

382 HALPIN ET AL.
community structure, primarily becausecompetition for space, predator-preyinteraction, and low tide abiotic conditionswere assumed to have much stronger effects oncommunity structure. Also, oceanographicconditions such as nutrient and phytoplanktonconcentrations were not known to vary inbiologically significant fashion in the nearshoreregion, because oceanographic measures aretypically taken from ships much furtheroffshore.
The importance of large-scaleoceanographic transport to larval recruitment inintertidal areas has been more widelyrecognized. As most organisms within thecommunity have limited or no adult mobilitybut broadcast their larvae into the water columnto develop, the larval input to the communitymay have little connection to the conditions atthe site. Additionally, spawners cannotinfluence the final destination of their larvae bymoving from an unfavorable current, e.g., onewhere their larvae will be transported to anunsuitable site or where the water conditionsare poor, to a favorable one. Therefore,populations can potentially function as sources,providing juveniles to other populations, or assinks, dependent on other populations forjuveniles but producing little on their own. Thecomplicated and oceanographically dependentlife cycle of marine plants and animals must betaken into account when designing managementstrategies. Therefore conservation plansdeveloped within terrestrial ecosystems, wherepopulations are typically more closed, cannotbe directly transferred to marine systems(Allison et al . 1998). New conservationstrategies have been developed in SouthAmerica, New Zealand, and North America thatcenter on preserving “source” populations intheir conservation plans.
Rocky intertidal ecosystems: North America
North American rocky shores in the easternPacific have not been exploited historically tothe extent of South American, though nativeAmericans did harvest intertidal resources andCalifornia rocky shores have recently begun tobe heavily harvested by southeast Asianimmigrants (Simenstad et al. 1978, Knudsen &Vogel 1997). To this end, we can considerNorth American rocky shore ecosystems asfunctioning a manner less directly impacted byhuman activities. On shores from Canadathrough California, the California mussel,Mytilus californianus, forms the dominantcover in the mid-intertidal zone. Mytilus
californianus is limited at low intertidal levelsby predation from the ochre seastar Pisasterochraceus and at the upper level by physicalfactors (Paine 1966, 1974, 1976). The musselachieves a size refuge from its other majorpredator, whelks in the genus Nucella at higherlatitudes, and at even larger sizes from P.ochraceus (Paine 1976, Sanford et al. 2003).The barnacle, Balanus glandula, also formsmajor cover in this zone when mussels areabsent. The upper zone is dominated by thebarnacles B. glandula and Chthamalus dalli, aswell as algae in the genera Fucus, Pelvetia, andMastocarpus.
Experimental work on western NorthAmerican rocky shores has elucidated many ofthe major physical and biotic interactions thatcontribute to community structure (Paine 1966,Dayton 1971, Paine 1974, 1980, Paine & Levin1981, Menge et al. 1994, Robles 1997). Fig. 5shows a generalized interaction web for lowerintertidal rocky shores in central Oregon.Pisaster ochraceus acts as a keystone predatorthroughout its range via its consumption of M.californianus and other invertebrates. Predatorywhelks can be important predators when P.ochraceus is absent (Navarrete & Menge 1996).In Southern California, spiny lobsters,Panulirus interruptus, may also prey heavily onmussels, acting as a keystone predator wheremussel recruitment rates are high (Robles1997). Herbivore grazing strongly affectsephemeral algae and the course of benthicsuccession, but their abili ty to controlmacroalgal abundance patterns remains unclear(Robles & Cubit 1981, Cubit 1984). Thecontrols of herbivore population sizes arepoorly understood, but oystercatchers havebeen shown to influence limpet populationstructure (Frank 1982, Hahn & Denny 1989,Wootton 1997). Space is a limiting resource forboth algae and sessile invertebrates. Thus,competition for space is a typically a stronginteraction both between and within species onexposed rocky shores. However, oceanographicfactors may help explain which sessile speciesdominates at a given site by altering growthand recruitment rates (discussed below).
The role of recruitment in structuring rockyintertidal communities has been a majorresearch focus since the early 1980s.Roughgarden et al. (1988) documented thatvariation in oceanographic conditions along theCalifornia coast influenced recruitment patternsin the barnacle, B. glandula, and that B.glandula populations were limited byrecruitment in many areas. They also suggestedthat recruitment patterns were tied to upwelling

383OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
patterns and that both patterns could bedisrupted by El Niño conditions. The authorshypothesized, and subsequent data havesupported, that larvae produced onshore areadvected offshore by upwelled water. Larvaeconcentrate at the boundary between coldupwelled and warmer offshore surface water aswell as in eddies of the California current (Fig.6). When upwelling winds relax, the larvae aretransported back onshore where theyconsequently settle out of the water column(Roughgarden et al. 1988, Farrell et al. 1991,Roughgarden et al. 1991, Wing et al. 1995).The “tattered curtain” hypothesis (Roughgardenet al. 1991), posits that the length of upwellingrelaxations in combination with the width ofthe upwelling band determine the strength ofrecruitment at a latitudinal scale. During briefrelaxation periods, the upwelling front wherelarvae concentrate may only reach land inregions where the upwelling band is narrowbefore upwelling resumes, driving the frontoffshore again. During the 1982-1983 El Niño,when upwelling decreased along the Californiacoast, intertidal barnacle recruitment increased(Roughgarden et al. 1988). Since the upwellingband is narrower in Oregon and Washingtonthan off of California, the hypothesis maypartially explain recruitment differencesbetween the two regions (Roughgarden et al.1988, Connolly & Roughgarden 1998).Recruitment rates in Oregon north of Cape
Fig. 5: Interaction web showing direct interactions in the low zone of exposed rocky shores ofOregon, USA. Minus signs indicate negative effects, plus signs indicate positive effects (afterMenge et al. 1994).Trama de interacciones mostrando las interacciones directas en la zona inferior de la costa rocosa expuesta de Oregon,USA. Los signos menos indican efectos negativos, los signos más indican efectos positivos (en base a Menge et al. 1994).
Fig. 6: Schematic diagram of California currentsystem next to the California coast during springupwelling. Circles with dots indicate southwardflow, circles with x indicates northward flow(after Roughgarden et al. 1988).Diagrama esquemático del sistema de la corriente de Cali-fornia cercano a la costa de California durante la surgenciade primavera. Los círculos con puntos indican flujos haciael sur, los círculos con x indican flujos hacia el norte (enbase a Roughgarden et al. 1998).
Blanco are an order of magnitude higher thatthey are in California (Connolly et al. 2001),highlighting the potential importance ofoceanographic features to recruitment.
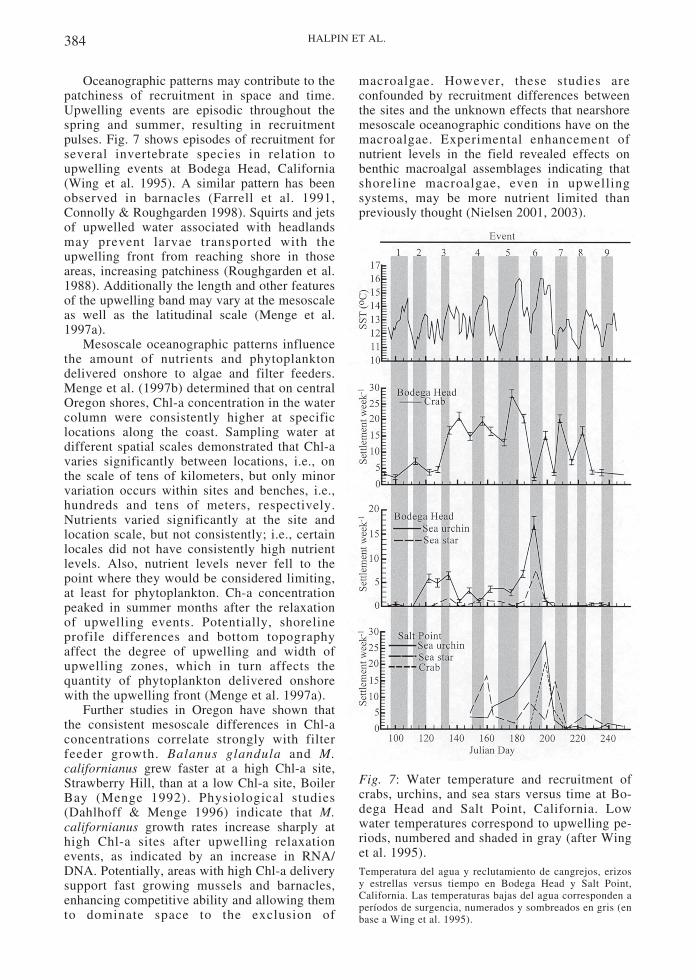
384 HALPIN ET AL.
Oceanographic patterns may contribute to thepatchiness of recruitment in space and time.Upwelling events are episodic throughout thespring and summer, resulting in recruitmentpulses. Fig. 7 shows episodes of recruitment forseveral invertebrate species in relation toupwelling events at Bodega Head, California(Wing et al. 1995). A similar pattern has beenobserved in barnacles (Farrell et al. 1991,Connolly & Roughgarden 1998). Squirts and jetsof upwelled water associated with headlandsmay prevent larvae transported with theupwelling front from reaching shore in thoseareas, increasing patchiness (Roughgarden et al.1988). Additionally the length and other featuresof the upwelling band may vary at the mesoscaleas well as the latitudinal scale (Menge et al.1997a).
Mesoscale oceanographic patterns influencethe amount of nutrients and phytoplanktondelivered onshore to algae and filter feeders.Menge et al. (1997b) determined that on centralOregon shores, Chl-a concentration in the watercolumn were consistently higher at specificlocations along the coast. Sampling water atdifferent spatial scales demonstrated that Chl-avaries significantly between locations, i.e., onthe scale of tens of kilometers, but only minorvariation occurs within sites and benches, i.e.,hundreds and tens of meters, respectively.Nutrients varied significantly at the site andlocation scale, but not consistently; i.e., certainlocales did not have consistently high nutrientlevels. Also, nutrient levels never fell to thepoint where they would be considered limiting,at least for phytoplankton. Ch-a concentrationpeaked in summer months after the relaxationof upwelling events. Potentially, shorelineprofile differences and bottom topographyaffect the degree of upwelling and width ofupwelling zones, which in turn affects thequantity of phytoplankton delivered onshorewith the upwelling front (Menge et al. 1997a).
Further studies in Oregon have shown thatthe consistent mesoscale differences in Chl-aconcentrations correlate strongly with filterfeeder growth. Balanus glandula and M.californianus grew faster at a high Chl-a site,Strawberry Hill, than at a low Chl-a site, BoilerBay (Menge 1992). Physiological studies(Dahlhoff & Menge 1996) indicate that M.californianus growth rates increase sharply athigh Chl-a sites after upwelling relaxationevents, as indicated by an increase in RNA/DNA. Potentially, areas with high Chl-a deliverysupport fast growing mussels and barnacles,enhancing competitive ability and allowing themto dominate space to the exclusion of
macroalgae. However, these studies areconfounded by recruitment differences betweenthe sites and the unknown effects that nearshoremesoscale oceanographic conditions have on themacroalgae. Experimental enhancement ofnutrient levels in the field revealed effects onbenthic macroalgal assemblages indicating thatshoreline macroalgae, even in upwellingsystems, may be more nutrient limited thanpreviously thought (Nielsen 2001, 2003).
Fig. 7: Water temperature and recruitment ofcrabs, urchins, and sea stars versus time at Bo-dega Head and Salt Point, California. Lowwater temperatures correspond to upwelling pe-riods, numbered and shaded in gray (after Winget al. 1995).Temperatura del agua y reclutamiento de cangrejos, erizosy estrellas versus tiempo en Bodega Head y Salt Point,California. Las temperaturas bajas del agua corresponden aperíodos de surgencia, numerados y sombreados en gris (enbase a Wing et al. 1995).

385OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
El Niño effects on rocky shore communitieshave been poorly understood, primarily due tothe lack of long term monitoring of intertidalbiota. Paine (1986) assessed disruptions to theintertidal community at Tatoosh Island,Washington, in the context of El Niño events.He found that while community disruptionsoccurred during El Niño years, the samedisruptions were also caused by extremeweather and oceanographic events notassociated with El Niño. For example, El Niñorelated storms removed large portions of themussel bed, but severe storms in other yearsalso caused extensive damage. Also, of the fourmajor sea urchin recruitment episodes of thepast 20 years, two occurred in El Niño years,and the other two occurred in non-El Niñoyears, though those years also had unusuallywarm waters. Elevated sea levels such as thosethat occur during El Niño rather thantemperature (Mysak 1986), appears to coincidewith strong urchin recruitment.
The water transported north during ENSOevents contains few nutrients, which candetrimentally affect growth of intertidal algae.Additionally, the thermocline and nutriclinecan deepen, making deep-water nutrients lessavailable (McGowan et al. 1998). Research hasindicated that nutrients for microalgae mightbecome limiting during El Niño summers(Wootton et al. 1996). Laminarian algal speciesappear to be the most susceptible (Gunnill1985). Algae weakened by the nutrient poorwater might then suffer severe damage duringsubsequent boreal winter storms. However, asPaine (1986) suggested for mussels, consistenteffects of El Niño on macroalgae in the rockyintertidal are difficult to separate from seasonaland interannual variability.
Dayton & Tegner (1990) speculated on whyintertidal communities might be resistant to thesevere perturbation that El Niño events have onother ecosystems. They hypothesized thatintertidal plants and animals have evolved inthe context of severe physical conditions suchas wave action and heat stress. Most intertidalalgae can regenerate from their holdfasts andcan recover from large loss of biomass due tostorm damage or nutrient starvation. Moreover,the elevated sea levels associated with ENSOsmay mitigate the increased temperature effectson intertidal organisms by decreasing exposureto low tide thermal stress.
The potential effects of global warming onNorth American rocky shores have just begunto be studied (Ray et al. 1992). Comparisonsbetween 1931-1933 and 1993-1994 census ofintertidal biota at a site in Monterey Bay,
California, indicate that biotic changes haveoccurred in the intervening time period (Barryet al. 1995, Sagarin et al. 1999). Significantly,there was a greater tendency for southernspecies to extend their range northward intoMonterey than for northern species to extendtheir range southward. Concurrently, mean andmaximum shoreline water temperaturesincreased by 0.75 and 2.2 oC, respectively.Therefore, one consequence of global warmingmay be latitudinal shifts in species ranges.However, Denny & Paine (1998) pointed outthat the two samples taken in the study were atdifferent phases of the Metonic cycle. Since theMetonic cycle influences the duration andtiming of low tide exposure, the study may beconfounded by variation in low-tide physicalstress caused by the cycle. Sagarin et al. (1999)attempted to account for the problemsassociated with a lack of continuous data at theMonterey site. While their analyses provide astrong inference that climate change isaffecting the community, they also point to theimportance of both sustained biologicalmonitoring and biologically relevant thermalmodeling. Recent data on mussel bodytemperatures modeled through thermal“mimics,” show no increase of maximal bodytemperature with latitude, and that the highestbody temperatures occurred during low tide(Helmuth et al. 2002). These data indicate thatterrestrial climatic conditions may impactorganismal body temperature more thanincreases in water temperature. In contrast,Sanford (1999) documented that the moderatewater temperature fluctuations (8-14 oC)associated with normal upwelling events onthe Oregon coast have dramatic behavioralimpacts on P. ochraceus. Since P. ochraceusis a keystone predator, even minor watertemperature changes or modif icat ion ofupwelling patterns can have wide communityeffects.
Climate change models also predict anincreased rate of sea level rise, through thermalexpansion of the water and melting of ice overthe Antarctic (Beatly 1991). Currently sea levelis rising at 0.9 ± 0.2 mm per year on thesouthern California coast (Roemmich 1992).The rise and fall of water with the tidesdetermine the distribution of intertidal animalsand plants. A change in sea level rise wouldimpact intertidal organisms significantly.Sudden changes of tidal height have beendocumented due to coseismic uplift in Chileand Alaska, where earthquakes raised intertidalareas in relation to sea level (Castilla 1988).These sudden catastrophic changes had severe

386 HALPIN ET AL.
impacts on the intertidal communities. Theability of communities to change in relation tomore gradual sea level rise is not known. Adetermining factor will probably be theavailability of suitable substrate in “new”intertidal zones.
Global warming has also been predicted toaffect the degree of upwelling along the NorthAmerican coast. Hypothetically, upwellingmight increase as climate warming increaseswind stresses along the coast (Bakun 1990) andthere is evidence indicating that upwelling hasincreased in the northeastern Pacific even asocean temperatures have increased (Schwing &Mendelssohn 1997). However, it has also beenposited that due to a deepened thermocline, theupwelled water will be warmer and less nutrientrich (Roemmich & McGowan 1995). Changes inupwelling patterns would effect recruitment,delivery of Chl-a, and average onshore watertemperatures during the spring and summer.Additionally, upwelling influences the onshoreterrestrial climate. During upwelling, foggy coolconditions prevail along coastlines, amelioratingpotentially stressful low tide conditions (Bakun1990). Alterations in upwelling patterns couldtherefore significantly alter the physicalconditions at both high and low tide forintertidal organisms.
The Metonic cycle affects watertemperature and the duration of aerial exposureby intertidal organisms (Denny & Paine 1998,Helmuth et al. 2002). During periods whenlunar declination is at a maximum, tidalfluctuations are smaller and onshore watertemperatures are lower than when lunardeclination is at a minimum. Calculatedexposure times for Monterey Bay organismsmay change by nearly 50 % depending on thephase of the Metonic cycle. On Tatoosh Island,Washington, the Metonic cycle helps to explainvariation in changes in the upper limit ofmussel distribution. Consistent long termmonitoring of intertidal sites is required tobetter understand the responses of intertidalcommunities to climate change, ENSOs, andlong period climactic fluctuations.Additionally, the effect of the PDO onnortheastern Pacific rocky shores are alsounknown, but is most l ikely to effectrecruitment patterns of species with planktoniclarvae via effects on the California current andlarval transport.
Rocky intertidal ecosystems: South America
South American intertidal communities sharemany qualities with North American systems.
For many trophic and tidal levels the majorphyla are the same (Broitman et al. 2001). Athigh to mid intertidal areas, barnacles such asJehlius cirratus, Balanus laevis, and Balanusflosculus dominate. Grazers such as fissurellidlimpets and chitons exert strong effects on thesystem (Jara & Moreno 1984). At slightlylower levels, chorito maico (mussel),Perumytilus purpuratus, persists when noteaten by the predaceous loco (muricidgastropod), Concholepas concholepas (Morenoet al. 1986, Durán & Castilla 1989). Thecrustose green alga Codium dimorphum alsoforms a major portion of the cover (Santeliceset al. 1981). In the low zone, the largemacroalgae chascón, Lessonia nigrescens, andconchayuyo, Durvillea antarctica, comprise theprimary cover, with L. nigrescens havinggreater abundance at more wave swept sitesthan D. antarctica (Santelices et al. 1980).Other important predators, include the seastarsStichaster striatus, Meyenaster gelatinosus, andparticularly the sol de mar (sun star), Heliasterhelianthus (Navarrete & Castilla 2003). Thepejesapo, S. sanguinus , consumes mostorganisms attached to the rocks in its grazingarea, including barnacles, mussels, and algae.
A generalized interaction web for the mid-intertidal in Chile is given in Fig. 8 (afterMoreno 1986, Moreno et al. 1986, Durán et al.1987, Durán & Castilla 1989, Godoy & Moreno1989). When not heavily fished by humans,locos exert strong top down control on musselsand barnacles (Moreno et al. 1986). Similarly,lapas (keyhole limpets, Fissurella spp.) grazeheavily on algae when humans do notdrastically reduce populations. In reserveswhere harvesting is prohibited, grazing byfissurellid limpets restrict the red alga, Irideaboryana , to refuge habitats (Moreno &Jaramillo 1983). In the absence of herbivory, I.boryana forms a dominant part of the cover(Jara & Moreno 1984). Fissurellid limpetscompete heavily with patellid l impets(Collisella spp.) and pulmonate gastropods(Siphonaria spp.) (Moreno & Jaramillo 1983,Godoy & Moreno 1989).
As indicated above, humans have haddramatic impacts on the structure of intertidalcommunities in Chile (Moreno 1986, Moreno etal. 1986, Durán et al. 1987, Durán & Castilla1989, Godoy & Moreno 1989, Castilla 1999).Locos, whelks, limpets, urchins, picorocos(large barnacle, Austromegalobalanuspsittacus), mussels, and several species ofseaweed in the genera Lessonia, Durvillea,Mazzaella, and Gelidium are all harvested(Durán et al. 1987, Santelices & Norambuena

387OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
1987, Hoffmann & Santelices 1997). Sixtythousand tons of Lessonia and 300-500 tons ofGelidium are harvested yearly for export ofalginate and agar respectively. Approximately430 tons of D. antarctica are harvested for foodyearly (Hoffmann & Santelices 1997).
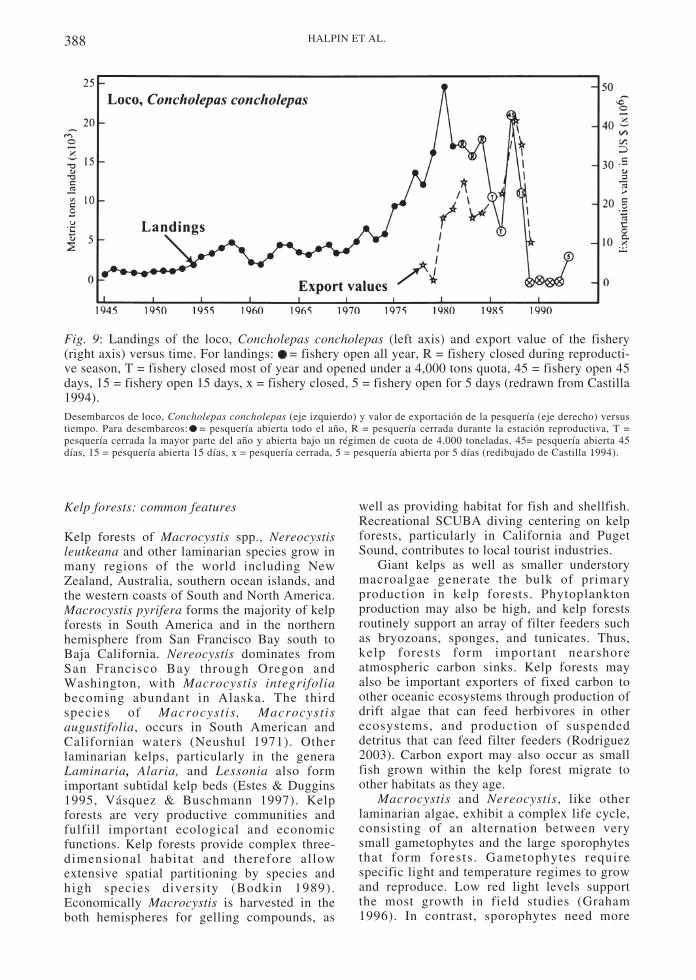
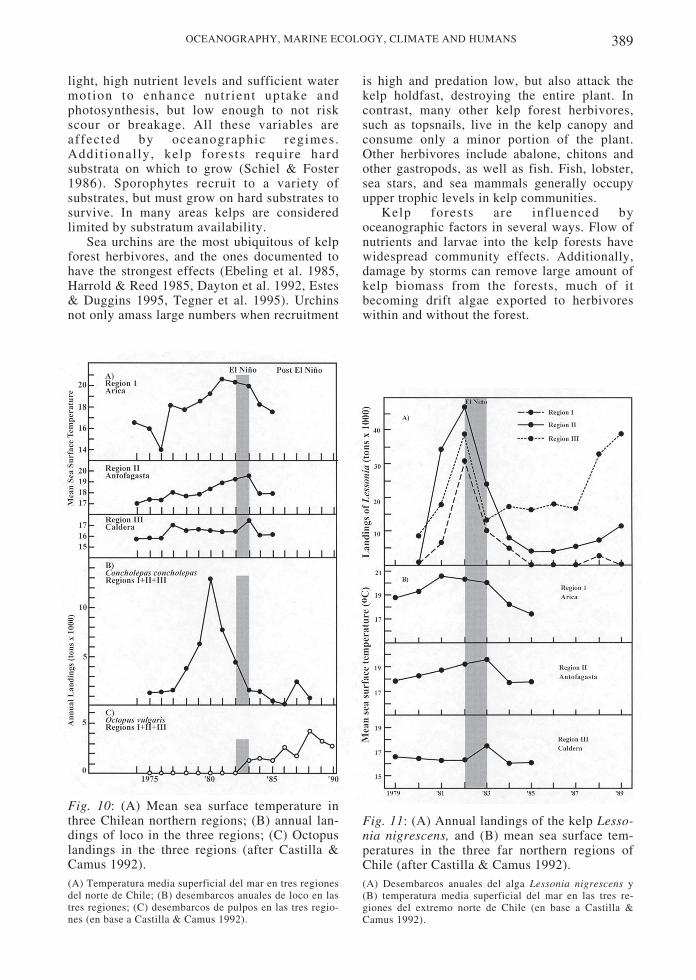
In the 1980s increased human harvesting,perhaps in combination with ENSO events,severely threatened the loco and Lessoniaharvests. In the case of the loco, increasedforeign demands caused prices to skyrocket,increasing fishing pressure (Fig. 9). Both locoand Lessonia populations declined sharply at thetime of the 1982-1983 El Niño (Fig. 10 and 11).However, the fishery was under severe humanpressure at the time and may have collapsed forthis reason alone (Castilla & Camus 1992). Dataindicate that loco populations exhibit reducedrecruitment when the ENSO index is eitherstrongly negative or positive (Moreno et al.1998). Upwelling events, rather than relaxationevents, may advect loco larvae onto shore due tomigration of the larvae into deeper waters atnight (Poulin et al. 2002a, Poulin et al. 2002b).Concurrent with the collapse of the loco, pulpo(octopus) landings increased dramatically (Fig.10). Although ENSO events have beendocumented to have severe effects on Peruvianand northern Chilean rocky shores (Soto 1985,Tomicic 1985, Arntz & Tarazona 1990), themore temperate rocky shores of central andsouthern Chile may be less directly effected(Navarrete et al. 2002).
The crash of the loco fishery led to theinstitution of new fishery practices in Chile.Two key elements are part of this plan: localcontrol of resources in coordination with centralgovernment and the establishment of marineprotected areas and reserves (Castilla 1994,Castilla & Fernández 1998, Castilla 2000,Castilla & Defeo 2001). Establishing localcontrol puts regulation of fisheries in the handsof those who both have a long term interest inthe fishery and are on site to enforce rules. Themarine protected areas establish larval sourcesfor outside populations that are exploited.Success hinges on establishing a sufficientnumber of enforced reserves to support asustainable fishery outside the protected areas(Castilla 2000, Fernández et al. 2000). Localartisanal fishing collectives manage reserves andenforce policies, in conjunction with nationalcoordination of the collectives.
“No take” reserves and protected areas havealso been a boon to experimental ecology inChile. Reserves that prohibit fishing havehelped document the severe impact that humanintervention has had outside the reserves. Inparticular, within reserve areas have lowermacroalgal and mussel cover due to greatlyincreased herbivory and predation (Moreno etal. 1986, Durán & Castilla 1989). Additionally,reserves can potentially seed non-reserve areasthrough the increased population size and bodysize of protected species, such as the loco(Manríquez & Castilla 2001).
Fig. 8: Interaction web for central Chilean mid rocky intertidal. Minus signs indicate negativeeffects, plus signs indicate positive effects (after Moreno et al. 1986).Trama de interacciones para el intermareal rocoso medio de Chile central. Los signos menos indican efectos negativos, lossignos más indican efectos positivos (en base a Moreno et al. 1986).
388 HALPIN ET AL.
Kelp forests: common features
Kelp forests of Macrocystis spp., Nereocystisleutkeana and other laminarian species grow inmany regions of the world including NewZealand, Australia, southern ocean islands, andthe western coasts of South and North America.Macrocystis pyrifera forms the majority of kelpforests in South America and in the northernhemisphere from San Francisco Bay south toBaja California. Nereocystis dominates fromSan Francisco Bay through Oregon andWashington, with Macrocystis integrifoliabecoming abundant in Alaska. The thirdspecies of Macrocystis, Macrocystisaugustifolia, occurs in South American andCalifornian waters (Neushul 1971). Otherlaminarian kelps, particularly in the generaLaminaria, Alaria, and Lessonia also formimportant subtidal kelp beds (Estes & Duggins1995, Vásquez & Buschmann 1997). Kelpforests are very productive communities andfulfill important ecological and economicfunctions. Kelp forests provide complex three-dimensional habitat and therefore allowextensive spatial partitioning by species andhigh species diversity (Bodkin 1989).Economically Macrocystis is harvested in theboth hemispheres for gelling compounds, as
Fig. 9: Landings of the loco, Concholepas concholepas (left axis) and export value of the fishery(right axis) versus time. For landings: = fishery open all year, R = fishery closed during reproducti-ve season, T = fishery closed most of year and opened under a 4,000 tons quota, 45 = fishery open 45days, 15 = fishery open 15 days, x = fishery closed, 5 = fishery open for 5 days (redrawn from Castilla1994).Desembarcos de loco, Concholepas concholepas (eje izquierdo) y valor de exportación de la pesquería (eje derecho) versustiempo. Para desembarcos: = pesquería abierta todo el año, R = pesquería cerrada durante la estación reproductiva, T =pesquería cerrada la mayor parte del año y abierta bajo un régimen de cuota de 4.000 toneladas, 45= pesquería abierta 45días, 15 = pesquería abierta 15 días, x = pesquería cerrada, 5 = pesquería abierta por 5 días (redibujado de Castilla 1994).
well as providing habitat for fish and shellfish.Recreational SCUBA diving centering on kelpforests, particularly in California and PugetSound, contributes to local tourist industries.
Giant kelps as well as smaller understorymacroalgae generate the bulk of primaryproduction in kelp forests. Phytoplanktonproduction may also be high, and kelp forestsroutinely support an array of filter feeders suchas bryozoans, sponges, and tunicates. Thus,kelp forests form important nearshoreatmospheric carbon sinks. Kelp forests mayalso be important exporters of fixed carbon toother oceanic ecosystems through production ofdrift algae that can feed herbivores in otherecosystems, and production of suspendeddetritus that can feed filter feeders (Rodriguez2003). Carbon export may also occur as smallfish grown within the kelp forest migrate toother habitats as they age.
Macrocystis and Nereocystis, like otherlaminarian algae, exhibit a complex life cycle,consisting of an alternation between verysmall gametophytes and the large sporophytesthat form forests. Gametophytes requirespecific light and temperature regimes to growand reproduce. Low red light levels supportthe most growth in field studies (Graham1996). In contrast, sporophytes need more
389OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
light, high nutrient levels and sufficient watermotion to enhance nutr ient uptake andphotosynthesis, but low enough to not riskscour or breakage. All these variables areaffected by oceanographic regimes.Addit ional ly, kelp forests require hardsubstrata on which to grow (Schiel & Foster1986). Sporophytes recruit to a variety ofsubstrates, but must grow on hard substrates tosurvive. In many areas kelps are consideredlimited by substratum availability.
Sea urchins are the most ubiquitous of kelpforest herbivores, and the ones documented tohave the strongest effects (Ebeling et al. 1985,Harrold & Reed 1985, Dayton et al. 1992, Estes& Duggins 1995, Tegner et al. 1995). Urchinsnot only amass large numbers when recruitment
Fig. 10: (A) Mean sea surface temperature inthree Chilean northern regions; (B) annual lan-dings of loco in the three regions; (C) Octopuslandings in the three regions (after Castilla &Camus 1992).(A) Temperatura media superficial del mar en tres regionesdel norte de Chile; (B) desembarcos anuales de loco en lastres regiones; (C) desembarcos de pulpos en las tres regio-nes (en base a Castilla & Camus 1992).
is high and predation low, but also attack thekelp holdfast, destroying the entire plant. Incontrast, many other kelp forest herbivores,such as topsnails, live in the kelp canopy andconsume only a minor portion of the plant.Other herbivores include abalone, chitons andother gastropods, as well as fish. Fish, lobster,sea stars, and sea mammals generally occupyupper trophic levels in kelp communities.
Kelp forests are influenced byoceanographic factors in several ways. Flow ofnutrients and larvae into the kelp forests havewidespread community effects. Additionally,damage by storms can remove large amount ofkelp biomass from the forests, much of itbecoming drift algae exported to herbivoreswithin and without the forest.
Fig. 11: (A) Annual landings of the kelp Lesso-nia nigrescens, and (B) mean sea surface tem-peratures in the three far northern regions ofChile (after Castilla & Camus 1992).(A) Desembarcos anuales del alga Lessonia nigrescens y(B) temperatura media superficial del mar en las tres re-giones del extremo norte de Chile (en base a Castilla &Camus 1992).
390 HALPIN ET AL.
Kelp forests: North America
The bulk of information on North Americankelp forests comes from studies in central tosouthern California waters and Alaska. Inparticular the kelp beds off of Monterey andSan Diego, California, where the HopkinsMarine Station and Scripps OceanographicInstitute, respectively, are located, and theAleutian Islands in Alaska have receivedconsiderable long term study. Less is knownabout kelp stands on the open coasts ofWashington and Oregon, possibly due to therelative lack of Macrocystis spp. as comparedto Nereocystis, and the logistical difficulty ofdiving research in these areas.
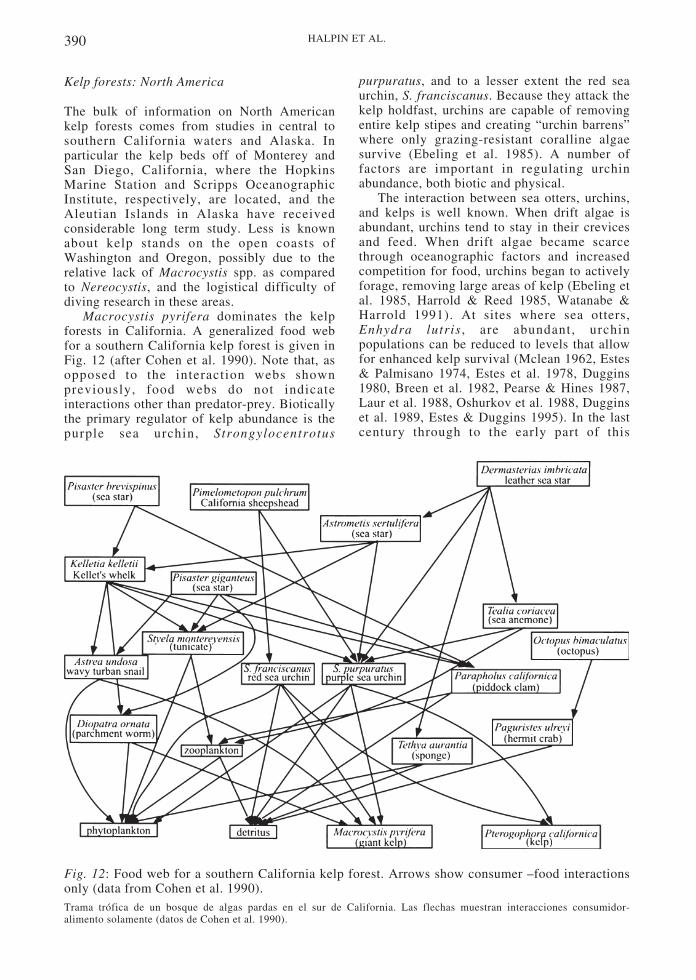
Macrocystis pyrifera dominates the kelpforests in California. A generalized food webfor a southern California kelp forest is given inFig. 12 (after Cohen et al. 1990). Note that, asopposed to the interaction webs shownpreviously, food webs do not indicateinteractions other than predator-prey. Bioticallythe primary regulator of kelp abundance is thepurple sea urchin, Strongylocentrotus
purpuratus, and to a lesser extent the red seaurchin, S. franciscanus. Because they attack thekelp holdfast, urchins are capable of removingentire kelp stipes and creating “urchin barrens”where only grazing-resistant coralline algaesurvive (Ebeling et al. 1985). A number offactors are important in regulating urchinabundance, both biotic and physical.
The interaction between sea otters, urchins,and kelps is well known. When drift algae isabundant, urchins tend to stay in their crevicesand feed. When drift algae became scarcethrough oceanographic factors and increasedcompetition for food, urchins began to activelyforage, removing large areas of kelp (Ebeling etal. 1985, Harrold & Reed 1985, Watanabe &Harrold 1991). At sites where sea otters,Enhydra lutris , are abundant, urchinpopulations can be reduced to levels that allowfor enhanced kelp survival (Mclean 1962, Estes& Palmisano 1974, Estes et al. 1978, Duggins1980, Breen et al. 1982, Pearse & Hines 1987,Laur et al. 1988, Oshurkov et al. 1988, Dugginset al. 1989, Estes & Duggins 1995). In the lastcentury through to the early part of this
Fig. 12: Food web for a southern California kelp forest. Arrows show consumer –food interactionsonly (data from Cohen et al. 1990).Trama trófica de un bosque de algas pardas en el sur de California. Las flechas muestran interacciones consumidor-alimento solamente (datos de Cohen et al. 1990).

391OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
century, sea otters were driven extinctthroughout most of their range on the NorthAmerican coast. The cessation of otter huntingallowed reintroduction of the otter to some ofits former range in California and Alaska.Subsequent regulation of the urchin populationsby sea otter predation has strong positiveeffects on kelp abundances in areas of otterreintroduction (Estes & Palmisano 1974, Estes& Duggins 1995).
Even with sea otter re-introduction, urchinbarrens may persist until storms, disease, andrecruitment failures reduce urchin numbers.These factors can also facilitate kelp forest re-establishment in urchin barrens outside ofcurrent sea-otter geographic range (Ebeling etal. 1985). Large and mesoscale oceanographicfactors have a strong influence on urchinrecruitment. Areas with strong offshore flowsuch as capes and headlands show low andunpredictable recruitment patterns (Ebert &Russell 1988, Wing et al. 1995). Areas betweenheadlands may receive strong annual pulses. ElNiño disrupts recruitment of sea urchins, eitherthrough changing current patterns or becausethe warm nutrient poor water associated with ElNiño is not conducive to larval survival.
The effects of El Niño on Macrocystis havebeen well documented (see Tegner & Dayton1987, Dayton & Tegner 1990 for review). Kelpforests are impacted directly through increasedstorm activity during the winter and throughdelivery of warm, nutrient-poor water insummer, and indirectly through the subsequenteffects on urchin populations (Dayton &Tegner 1984, Dayton et al. 1999). El Niñogenerated storms can remove large amounts ofkelp biomass from a forest or remove entirestands (Dayton & Tegner 1984, Dayton et al.1992, Stull 1995). During the summers of ElNiño years, kelps may undergo further die offsdue to nutrient limitation, particularly ofnitrogen (Dayton et al. 1999). The effect ofelevated temperature is not considered to be assevere as that of nutrient depletion, but there issome evidence that it may make the kelps morevulnerable to disease (North 1983). Very young“blade stage” Macrocystis have been observedto have enhanced growth rates during El Niño,perhaps because water clarity is greater andnutrients may be less limiting at the kelp forestbottom (Schroeter et al. 1995). In southernCalifornia, El Niño storms cause increases inthe pollutants delivered to coastal areas throughincreased runoff (Stull 1995, Maurer & Nguyen1997), though the effects of this are unknown.
The extensive biomass removal due to ElNiño effects within the kelp stands results in
lower drift algae available for urchins (Daytonet al. 1992). Consequently, urchins begin toforage outside of crevices and within the bed asdescribed above. Thus, even more kelp biomassis lost as the urchins feed. In this fashion,stable urchin barrens may be created (Dayton etal. 1992), although most sites recover quicklyfrom El Niño via subsequent kelp recruitment.Urchins are capable of maintaining barrensuntil the population is reduced by disease orother mortality, after which Macrocystis againdominates (Dayton et al. 1992). Other effectsof El Niño on kelp forest communities includedecreased recruitment of red abalone, Haliotisrufescens ; increased recruitment of pinkabalone, H. corrugata, spiny lobster, Panulirusinterruptus, and the California sheepsheadwrasse, Semicossyphus pulcher; and declines inthe abundances of sea stars species Patiriaminiata , P. ochraceus , and Dermasteriasimbricata. In Baja California, severe El Niñocorresponded to Macrocystis loss andrecruitment failure of abalone, perhaps becauseMacrocystis aids in local larval retention(Shepherd et al. 1998).
Human activities impacting kelp forests inNorth America include kelp harvesting;commercial fisheries for abalone, sea urchins,fish; sport fishing for lobster and fish; andsewage runoff. Historically, sea otter huntinghad devastating effects on kelp forests asmentioned above. In southern California,Macrocystis beds where sea otters are absent,urchin populations are heavily preyed upon bythe California sheepshead wrasse and spinylobster, both of which are fished by humans(Tegner & Dayton 1981). A number of otherupper trophic level species have been heavilyfished in and around kelp forests, such as rockfish, seabass, and sharks. But since baselinedata are limited and all change occurs withinthe context of larger scale oceanographicdynamics, full assessment of human impacts isdifficult (Dayton et al . 1998). Abalonepopulations of all species have been severelyoverfished, to near the point of extinction in thecase of white abalone, Haliotis sorensii, listedas endangered in 2001 (Davis et al. 1996,NOAA 2001, Rogers-Bennett et al. 2002a).
Kelp has been harvested off of NorthAmerica since the early part of the 20thcentury, primarily for potash. During WWIkelp forests were heavily exploited for potash,acetone, and charcoal. After the war the marketfor those kelp products collapsed and graduallyalgin became the primary commercial kelpproduct. The value of kelp algin production hashistorically rivaled that of the offshore fisheries

392 HALPIN ET AL.
(North 1971). Generally, kelp boats harvest thetop portion of kelp stipes, allowing kelpsurvival and maintenance of the threedimensional ecosystem structure. However,productivity is lost with kelp harvesting due totissue loss, particularly of the most heavilyphotosynthesizing parts, those at the surface.
While sea otter reintroduction is regardedboth as a highly successful recovery andcautionary tale about species exploitation, threecaveats must be added. One is that sea ottershave yet to occupy the full extent of theirformer range. The region from northernCalifornia to Vancouver Island still lacks viableotter populations. Secondly, sea otters arecurrently in decline in areas of Alaska fromheavy predation by killer whales, Orcinus orca.Killer whales have apparently changed diet aspopulations of their preferred prey, pinnipeds,have declined (Estes et al. 1998, Doroff et al.2003). Lastly, the fishery for red sea urchinsbecame highly profitable in the 1980’s due toAmerican and Japanese demand for the roe(Kato & Schroeter 1985, Pfister & Bradbury1996). The fishery quickly became over-fishedin California and has declined precipitouslysince 1988 (Botsford et al. 1999), though itremains the second-most top grossing fishery inCalifornia (Rogers-Bennett et al. 2002b). Seaotter reintroduction plans are often met withresistance by urchin fishers, who view theotters as competitors.
It is not known how global climate changewill impact kelp communities. As mentionedpreviously, some conceptual models of climatechange predict increased upwelling (Bakun1990). If, as Roemmich & McGowan (1995)suggest, upwelled water becomes warm and hasinsufficient nutrients to sustain the kelps,forests would be severely adversely affected.Conversely, if upwelled water is cool andcontains sufficient nutrients, the increasedupwelling may buffer other effects of climatechange. Climate change models also predictincreased storm activity, which would increasephysical kelp damage, and increased frequencyof El Niño, which would starve kelps ofnutrients. Additionally, California ChannelIsland black abalone populations have sufferedserious declines due to “withering syndrome”since the late 1980’s. Caused by aRickettsiales-like prokaryote, watertemperature appears to be the strongestenvironmental correlate with disease severity(Moore et al. 2002). Increased sea temperaturesdue to either global warming or frequent ElNiño would impact negatively on abalonepopulations through disease or other factors
(Harvell et al. 1999, Hobday & Tegner 2002,Moore et al. 2002).
Kelp forests: South America
The kelp forests off of the South Americanwest coast are not as extensive or diverse asthose off of North America. Nor have they beenas extensively exploited commercially,although they are harvested for alginate. Thismay be due to the more patchy distribution ofMacrocystis off South America, or becauseLessonia is more abundant and accessible.Lessonia forms highly diverse “forests” of itsown in the shallow subtidal and low intertidal.Vásquez & Buschmann (1997) provide areview of work on Chilean subtidal kelp beds.
Macrocystis forms beds all along theChilean and southern Peruvian coast, but ismost abundant south of 42º. Consequently, themajority of studies on Macrocystis have beenconducted in southern Chile. In many areas offSouth America, Macrocystis stands aredecimated on an annual basis, due to stormactivity (Moreno & Sutherland 1982), orunknown factors. At a site in southern Chile,the depth of the shallow edge of Macrocystisbeds appeared to be set through competitionwith Lessonia vadosa, and its deep edgethrough a combination of substrate availabilityand competition with Lessonia flavicans(Santelices & Ojeda 1984). The role of nutrientlimitation in general or in conjunction withENSO events has been little studied (Vásquez& Buschmann 1997).
Unlike their northern hemispherecounterparts, the South American otter, thesea cat Lontra felina, does not prey on seaurchins (Cohen et al. 1990) and is not akeystone predator. However, data on the roleof sea urchins in regulating South AmericanMacrocystis communities is equivocal (seeVásquez & Buschmann 1997 for review ofherbivory in Chilean Lessonia andMacrocystis beds). Moreno & Sutherland(1982) found that nei ther the urchinLoxechinus albus nor the snail Tegula atraplayed a strong role in regulating Macrocystis.Castilla & Moreno (1982)1 obtained similarresults in southern Chile. Both studies foundthat, similar to the northern hemisphere,urchins subsisted on abundant drift kelp and
1 CASTILLA JC & CA MORENO (1982) Sea urchinsand Macrocystis pyrifera: experimental test of theirecological relations in southern Chile. Lawrence JM (ed)International Echinoderms Conference: 552. AA Balkema,Rotterdam, The Netherlands.

393OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
evidently did not need to actively forage onstanding kelp. However no information isavailable on grazing activity when drift kelpbecome limiting.
In contrast, Dayton (1985) found strongeffects of L. albus between 54-46º S in waveexposed areas. Macrocystis stands developed inareas where urchins were harvested. Hehypothesized that south of 54º S recruitment ofL. albus was extremely low, and thepopulations never grow to the point wherekelps are eliminated. Two factors were putforward to possibly account for low urchinrecruitment. One is that the extensivefreshwater runoff from the coastal fjordscreates water conditions averse to urchinlarvae. Alternatively the west wind drift, whichhits the South American coast north of wherethe kelp forests become abundant, may carrythe majority of urchin larvae off the coasttowards Antarctica (Dayton 1985)
Clearly more research needs to be done onfactors affecting kelp communities in SouthAmerica and on interactions among factors.Further study of the interactions of nutrientavailability, drift kelp availability, stormdamage, and urchin grazing would beparticularly valuable. El Niño effects on kelpforests have also been little studied in SouthAmerica. Arntz & Tarazona (1990) reportedthat a Macrocystis stands in Peru weredevastated during the 1982-1983 ENSO, butother data are lacking.
Pelagic ecosystems
Intuitively, open water ecosystems would seemto be those most likely to be directly impactedby oceanographic changes. The water masswithin which the organisms exist provides thephysical conditions of their environment,including transport, nutrients, and food. Unlikebenthic communities, space is not consideredlimiting. However, due to the logisticaldifficulties of open water experimentation, littleexperimental work has been done to elucidateinteraction webs in these communities. Muchdata has been collected on plankton abundancesas they are oceanographically informative, andfisheries research provides considerable data onthe abundance and diet of commerciallyimportant fish.
Primary productivity in these communitiesderives from phytoplankton, while fish andmarine mammals generally occupy the toptrophic levels. Organisms at lower trophiclevels, such as plankton, are small with weakor no swimming ability, and tend to move with
the water mass. Lower t rophic levelpopulat ions change on an annual andinterannual scale in response to localenvironmental conditions, since these speciesare short lived and have limited mobility.Satellite color data from 1979-1986 showedthat El Niño negatively impact phytoplanktonpopulations, except in a narrow region next tothe coast, by creating a warmer, deeper mixedlayer. El Niño accounted for most of theinterannual variabil i ty in phytoplanktonconcentration (Fiedler 1984, Strub et al.1990). More recent direct measurements offcentral California following the 1992 El Niñogive essentially the same results (Chavez1996). As described above, CalCOFI dataindicate that zooplankton populations alsorespond to changes during and following ElNiño years (Chelton et al. 1982, Roemmich &McGowan 1995). The primary result from theCalCOFI studies was that lower equatorwardflow and warmer water during El Niño yearsresulted in lower zooplankton populations offcentral and southern California.
Fish species are longer lived and canmigrate long dis tances, making use ofgeographic differences in physical forcing andcirculation. Thus, they can respond to bothannual and interannual variabi l i ty inenvironmental conditions by spawning inareas best suited for larval survival, andsubsequently migrating to more productiveregions to feed. In the California current, thepreferred spawning area for many species isthe quiescent and thermally stratified southernCalifornia bight. Over most of the past severaldecades, observations have shown that bothsmall pelagic species, such as sardine andanchovy, and larger fish, such as hake,migrate to the bight to spawn. Adults andjuveniles then migrate into the more stronglyforced, turbulent and advective regions northof the bight, where upwelling produces muchhigher concentrations of phytoplankton andzooplankton inshore of the core of theCalifornia current.
Interannual variability in fish catch is notusually as clearly related to environmentalconditions during a given year as phytoplanktonand zooplankton abundances are. Although thismay be partially due to the confounding effectsof changing fishing pressure and strategies,other factors may be at work such as the longerlife span and higher mobility of fish mentionedabove. For example, a strong year-class from agood year before an El Niño can result in a largepopulation, some of which are caught during thepoorer El Niño conditions and others of which
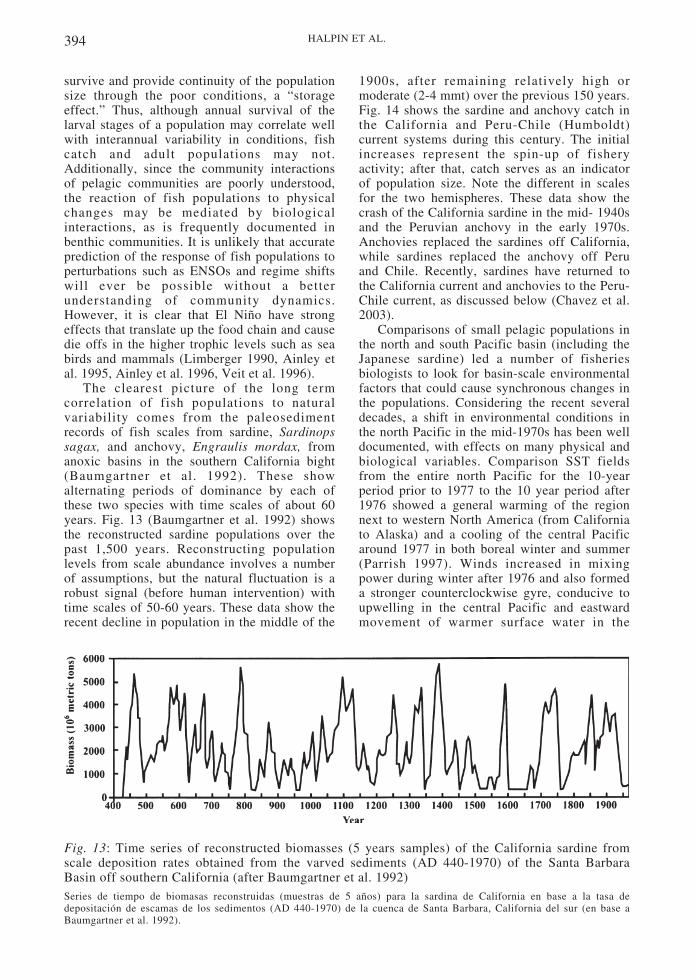
394 HALPIN ET AL.
survive and provide continuity of the populationsize through the poor conditions, a “storageeffect.” Thus, although annual survival of thelarval stages of a population may correlate wellwith interannual variability in conditions, fishcatch and adult populations may not.Additionally, since the community interactionsof pelagic communities are poorly understood,the reaction of fish populations to physicalchanges may be mediated by biologicalinteractions, as is frequently documented inbenthic communities. It is unlikely that accurateprediction of the response of fish populations toperturbations such as ENSOs and regime shiftswill ever be possible without a betterunderstanding of community dynamics.However, it is clear that El Niño have strongeffects that translate up the food chain and causedie offs in the higher trophic levels such as seabirds and mammals (Limberger 1990, Ainley etal. 1995, Ainley et al. 1996, Veit et al. 1996).
The clearest picture of the long termcorrelation of fish populations to naturalvariability comes from the paleosedimentrecords of fish scales from sardine, Sardinopssagax, and anchovy, Engraulis mordax, fromanoxic basins in the southern California bight(Baumgartner et al. 1992). These showalternating periods of dominance by each ofthese two species with time scales of about 60years. Fig. 13 (Baumgartner et al. 1992) showsthe reconstructed sardine populations over thepast 1,500 years. Reconstructing populationlevels from scale abundance involves a numberof assumptions, but the natural fluctuation is arobust signal (before human intervention) withtime scales of 50-60 years. These data show therecent decline in population in the middle of the
1900s, after remaining relatively high ormoderate (2-4 mmt) over the previous 150 years.Fig. 14 shows the sardine and anchovy catch inthe California and Peru-Chile (Humboldt)current systems during this century. The initialincreases represent the spin-up of fisheryactivity; after that, catch serves as an indicatorof population size. Note the different in scalesfor the two hemispheres. These data show thecrash of the California sardine in the mid- 1940sand the Peruvian anchovy in the early 1970s.Anchovies replaced the sardines off California,while sardines replaced the anchovy off Peruand Chile. Recently, sardines have returned tothe California current and anchovies to the Peru-Chile current, as discussed below (Chavez et al.2003).
Comparisons of small pelagic populations inthe north and south Pacific basin (including theJapanese sardine) led a number of fisheriesbiologists to look for basin-scale environmentalfactors that could cause synchronous changes inthe populations. Considering the recent severaldecades, a shift in environmental conditions inthe north Pacific in the mid-1970s has been welldocumented, with effects on many physical andbiological variables. Comparison SST fieldsfrom the entire north Pacific for the 10-yearperiod prior to 1977 to the 10 year period after1976 showed a general warming of the regionnext to western North America (from Californiato Alaska) and a cooling of the central Pacificaround 1977 in both boreal winter and summer(Parrish 1997). Winds increased in mixingpower during winter after 1976 and also formeda stronger counterclockwise gyre, conducive toupwelling in the central Pacific and eastwardmovement of warmer surface water in the
Fig. 13: Time series of reconstructed biomasses (5 years samples) of the California sardine fromscale deposition rates obtained from the varved sediments (AD 440-1970) of the Santa BarbaraBasin off southern California (after Baumgartner et al. 1992)Series de tiempo de biomasas reconstruidas (muestras de 5 años) para la sardina de California en base a la tasa dedepositación de escamas de los sedimentos (AD 440-1970) de la cuenca de Santa Barbara, California del sur (en base aBaumgartner et al. 1992).
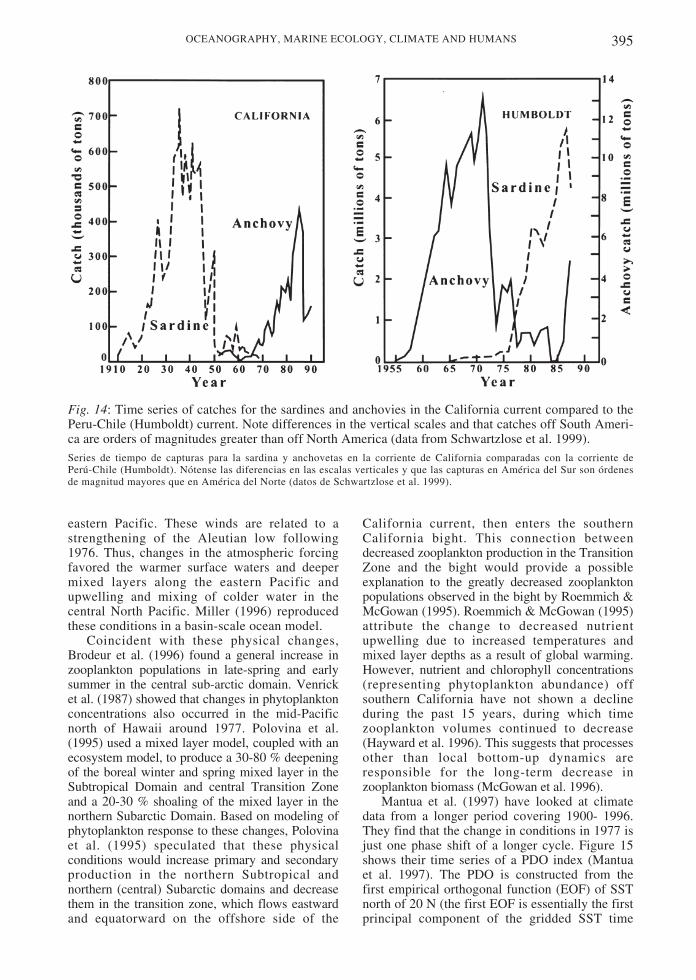
395OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
California current, then enters the southernCalifornia bight. This connection betweendecreased zooplankton production in the TransitionZone and the bight would provide a possibleexplanation to the greatly decreased zooplanktonpopulations observed in the bight by Roemmich &McGowan (1995). Roemmich & McGowan (1995)attribute the change to decreased nutrientupwelling due to increased temperatures andmixed layer depths as a result of global warming.However, nutrient and chlorophyll concentrations(representing phytoplankton abundance) offsouthern California have not shown a declineduring the past 15 years, during which timezooplankton volumes continued to decrease(Hayward et al. 1996). This suggests that processesother than local bottom-up dynamics areresponsible for the long-term decrease inzooplankton biomass (McGowan et al. 1996).
Mantua et al. (1997) have looked at climatedata from a longer period covering 1900- 1996.They find that the change in conditions in 1977 isjust one phase shift of a longer cycle. Figure 15shows their time series of a PDO index (Mantuaet al. 1997). The PDO is constructed from thefirst empirical orthogonal function (EOF) of SSTnorth of 20 N (the first EOF is essentially the firstprincipal component of the gridded SST time
Fig. 14: Time series of catches for the sardines and anchovies in the California current compared to thePeru-Chile (Humboldt) current. Note differences in the vertical scales and that catches off South Ameri-ca are orders of magnitudes greater than off North America (data from Schwartzlose et al. 1999).Series de tiempo de capturas para la sardina y anchovetas en la corriente de California comparadas con la corriente dePerú-Chile (Humboldt). Nótense las diferencias en las escalas verticales y que las capturas en América del Sur son órdenesde magnitud mayores que en América del Norte (datos de Schwartzlose et al. 1999).
eastern Pacific. These winds are related to astrengthening of the Aleutian low following1976. Thus, changes in the atmospheric forcingfavored the warmer surface waters and deepermixed layers along the eastern Pacific andupwelling and mixing of colder water in thecentral North Pacific. Miller (1996) reproducedthese conditions in a basin-scale ocean model.
Coincident with these physical changes,Brodeur et al. (1996) found a general increase inzooplankton populations in late-spring and earlysummer in the central sub-arctic domain. Venricket al. (1987) showed that changes in phytoplanktonconcentrations also occurred in the mid-Pacificnorth of Hawaii around 1977. Polovina et al.(1995) used a mixed layer model, coupled with anecosystem model, to produce a 30-80 % deepeningof the boreal winter and spring mixed layer in theSubtropical Domain and central Transition Zoneand a 20-30 % shoaling of the mixed layer in thenorthern Subarctic Domain. Based on modeling ofphytoplankton response to these changes, Polovinaet al. (1995) speculated that these physicalconditions would increase primary and secondaryproduction in the northern Subtropical andnorthern (central) Subarctic domains and decreasethem in the transition zone, which flows eastwardand equatorward on the offshore side of the
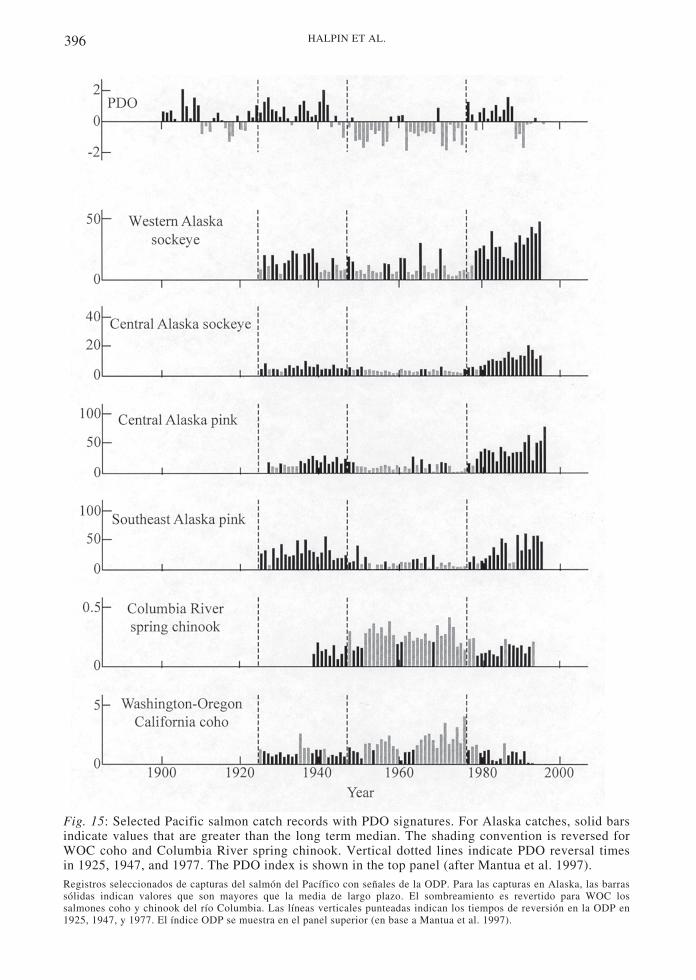
396 HALPIN ET AL.
Fig. 15: Selected Pacific salmon catch records with PDO signatures. For Alaska catches, solid barsindicate values that are greater than the long term median. The shading convention is reversed forWOC coho and Columbia River spring chinook. Vertical dotted lines indicate PDO reversal timesin 1925, 1947, and 1977. The PDO index is shown in the top panel (after Mantua et al. 1997).Registros seleccionados de capturas del salmón del Pacífico con señales de la ODP. Para las capturas en Alaska, las barrassólidas indican valores que son mayores que la media de largo plazo. El sombreamiento es revertido para WOC lossalmones coho y chinook del río Columbia. Las líneas verticales punteadas indican los tiempos de reversión en la ODP en1925, 1947, y 1977. El índice ODP se muestra en el panel superior (en base a Mantua et al. 1997).

397OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
series) (Preisendorfer 1988). The spatial patternassociated with this EOF is similar to the patterndescribed above: warm water along the easternboundary of the north Pacific and cold water inthe central gyre. The yearly value of the PDO isthe average of the time series of this EOF duringNovember-March. It thus represents the dominantSST pattern during the cold period of the year(when surface waters are well mixed and notdisguised by shallow warm layers). Mantua et al.(1997) use intervention analysis to argue for rapidchanges in conditions in 1925, 1947 and 1977.When SST and atmospheric pressure from theentire Pacific basin are regressed against thePDO, the SST pattern described above isrecovered, with a connection between the warmnortheast Pacific, a warm band along the equator,and another warm region in the southeast Pacificnext to South America, linking the environmentalvariability in the two Pacific eastern boundarycurrents. The atmospheric pressure patternassociated with the PDO consists primarily oflower pressures in the Alaska Gyre (a deeperAleutian Low). When the basinwide SST andatmospheric pressure signals are regressed againsta cold tongue index (CTI, a measure of the meanSST in the equatorial Pacific), a similar patternemerges, even though the CTI has ENSO timescales of 3-7 years and the PDO has time scalesof 30-50 years. The relationship between theinterannual and interdecadal changes is unknownand a subject of active research.
The PDO is correlated with a number ofother variables. We refer to periods of positivePDO as “warm periods,” focusing on the warmeastern boundaries of the north (and south)Pacific when the index is positive. Mantua etal. (1997) show that, during warm periods, airtemperature is also warmer around the easternbasin (from southern California to Alaska) andprecipitation is greater in the Alaska Gyre andless between northern California and BritishColumbia. The precipitation is reflected ingreater stream flow in the Alaskan Gyre andlower stream flow in the Pacific northwest.
Perhaps more striking than the correlation ofphysical factors with the PDO is thecorrespondence of warm phases of the PDO withhigher salmon catch in the Alaskan Gyre, withlower salmon catch in the Pacific Northwest, andthe seeming coincidence with increased survivalof sardine in the California current. In Fig. 15(Mantua et al. 1997) along with the PDO areplotted fish catch of four Alaska salmon stocksand two Pacific Northwest salmon stocks.Intervention analysis was used to fit step changesto the PDO and the salmon stocks. The fit to thePDO gives best results for the dates indicated
with vertical dotted line in Fig. 15. The fits forthe Alaska sockeye, Onchorhynchus nerka, stocksgive a step change 2-3 years after the PDOchange, while the fits for the Alaska pink, O.gorbuscha, give a step change one year after thePDO. Since pink salmon spend little time in thefresh water and only a year at sea, the differencein timing is consistent with a correlation betweensurvival and conditions during the early lifehistory. Pearcy (1992) used data from the PacificNorthwest to suggest that the critical period in theoceanic phase is during the first weeks or monthsafter juveniles enter the ocean from fresh water.The lags of the intervention analysis can beinterpreted to support this hypothesis. Off thePacific Northwest, Pearcy (1992) suggests thatpredation by warm-water fish (mackerel, hake)decreases survival of the juveniles during warmperiods, a hypothesis in need of testing.Mechanisms responsible for increases in thesalmon populations in the Gulf of Alaska duringthe warm water years are not known. The size ofcaught salmon in the Gulf of Alaska has declined;leading to the hypothesis that they have reachedor exceeded the carrying capacity of theirenvironment due to unusually low mortalitylevels in their earlier stages of life history and arenow food limited (United States GLOBEC 1996).
Gargett (1997), suggests a mechanismwhereby the productivity in the gulf of Alaskaand California current systems may be coupled.Using the concept of the “optimal environmentalwindow” she presents the idea that if the stabilityof the water column in the entire eastern coastalnorth Pacific increases, this will favor higherphytoplankton growth rates in the Gulf of Alaskabut lower growth rates in the California currentsystem. Conversely, decreases in the stabilityeverywhere will favor high phytoplankton growthrates in the California current but lower rates inthe Gulf of Alaska. Differences in resourcelimitation within the two current systems underliethe difference. Gulf of Alaska phytoplanktonstocks are more light limited, whereas Californiacurrent stocks are more nutrient limited. Shesuggests that if stability of the water columnvaries in phase over the entire eastern coastalNorth Pacific then secondary production andsalmon survival will vary out of phase.
Considering that the simple patterns shownin Fig. 14 and 15 for sardine, anchovy andsalmon suggest a connection between basin-scale conditions and the fish populations duringthis period, one might ask why the sardinepopulation did not rebound off California in themid-1970s. Although sardine populationsreached low values in 1960s and 1970s, enoughremained to estimate the rates of larval
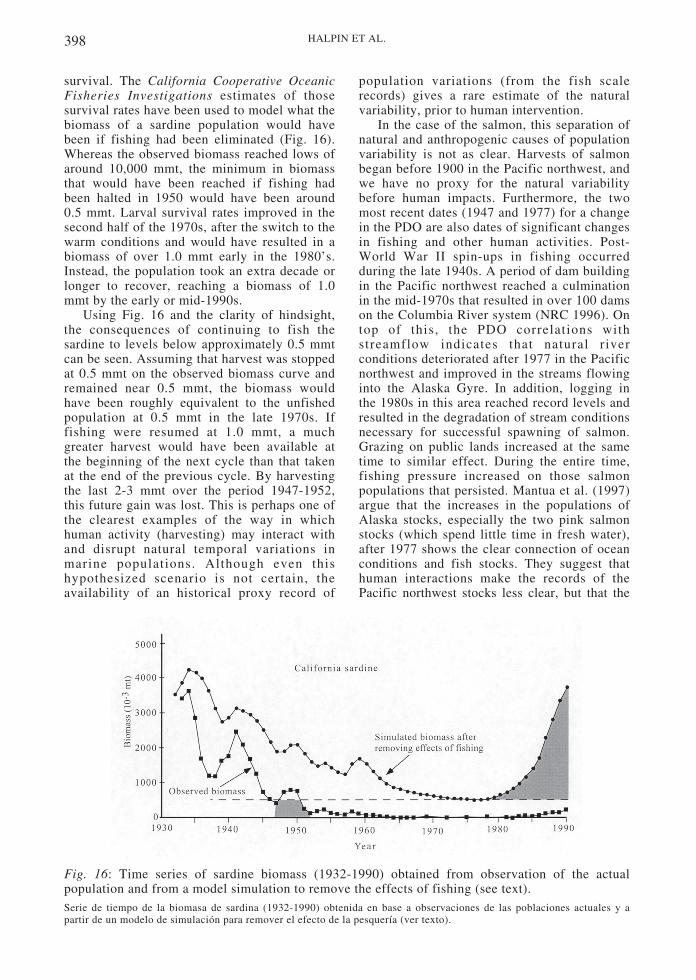
398 HALPIN ET AL.
survival. The California Cooperative OceanicFisheries Investigations estimates of thosesurvival rates have been used to model what thebiomass of a sardine population would havebeen if fishing had been eliminated (Fig. 16).Whereas the observed biomass reached lows ofaround 10,000 mmt, the minimum in biomassthat would have been reached if fishing hadbeen halted in 1950 would have been around0.5 mmt. Larval survival rates improved in thesecond half of the 1970s, after the switch to thewarm conditions and would have resulted in abiomass of over 1.0 mmt early in the 1980’s.Instead, the population took an extra decade orlonger to recover, reaching a biomass of 1.0mmt by the early or mid-1990s.
Using Fig. 16 and the clarity of hindsight,the consequences of continuing to fish thesardine to levels below approximately 0.5 mmtcan be seen. Assuming that harvest was stoppedat 0.5 mmt on the observed biomass curve andremained near 0.5 mmt, the biomass wouldhave been roughly equivalent to the unfishedpopulation at 0.5 mmt in the late 1970s. Iffishing were resumed at 1.0 mmt, a muchgreater harvest would have been available atthe beginning of the next cycle than that takenat the end of the previous cycle. By harvestingthe last 2-3 mmt over the period 1947-1952,this future gain was lost. This is perhaps one ofthe clearest examples of the way in whichhuman activity (harvesting) may interact withand disrupt natural temporal variations inmarine populations. Although even thishypothesized scenario is not certain, theavailability of an historical proxy record of
population variations (from the fish scalerecords) gives a rare estimate of the naturalvariability, prior to human intervention.
In the case of the salmon, this separation ofnatural and anthropogenic causes of populationvariability is not as clear. Harvests of salmonbegan before 1900 in the Pacific northwest, andwe have no proxy for the natural variabilitybefore human impacts. Furthermore, the twomost recent dates (1947 and 1977) for a changein the PDO are also dates of significant changesin fishing and other human activities. Post-World War II spin-ups in fishing occurredduring the late 1940s. A period of dam buildingin the Pacific northwest reached a culminationin the mid-1970s that resulted in over 100 damson the Columbia River system (NRC 1996). Ontop of this, the PDO correlations withstreamflow indicates that natural riverconditions deteriorated after 1977 in the Pacificnorthwest and improved in the streams flowinginto the Alaska Gyre. In addition, logging inthe 1980s in this area reached record levels andresulted in the degradation of stream conditionsnecessary for successful spawning of salmon.Grazing on public lands increased at the sametime to similar effect. During the entire time,fishing pressure increased on those salmonpopulations that persisted. Mantua et al. (1997)argue that the increases in the populations ofAlaska stocks, especially the two pink salmonstocks (which spend little time in fresh water),after 1977 shows the clear connection of oceanconditions and fish stocks. They suggest thathuman interactions make the records of thePacific northwest stocks less clear, but that the
Fig. 16: Time series of sardine biomass (1932-1990) obtained from observation of the actualpopulation and from a model simulation to remove the effects of fishing (see text).Serie de tiempo de la biomasa de sardina (1932-1990) obtenida en base a observaciones de las poblaciones actuales y apartir de un modelo de simulación para remover el efecto de la pesquería (ver texto).
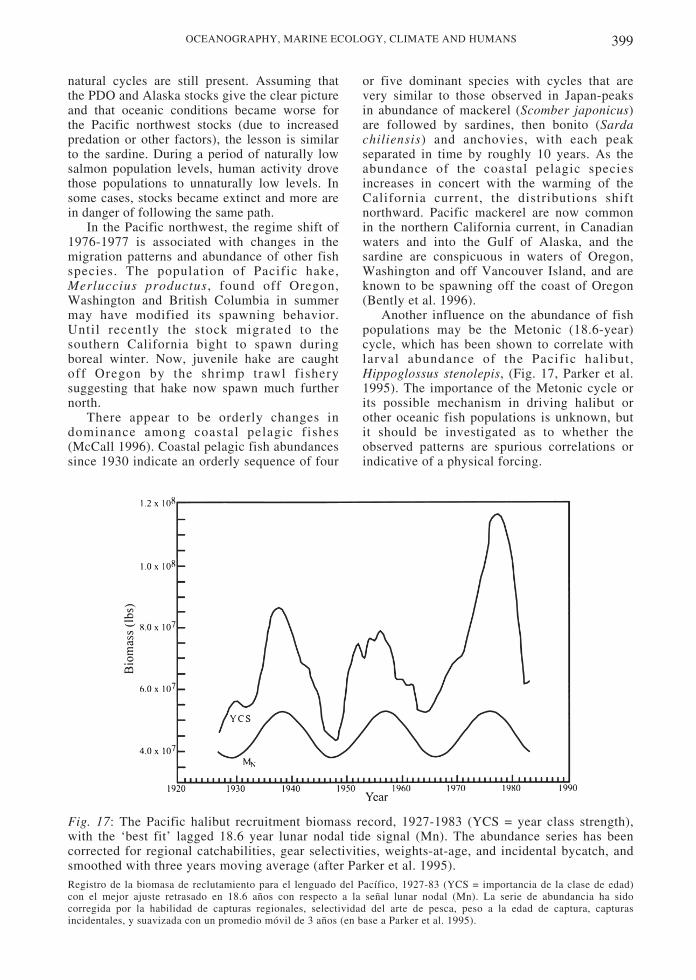
399OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
natural cycles are still present. Assuming thatthe PDO and Alaska stocks give the clear pictureand that oceanic conditions became worse forthe Pacific northwest stocks (due to increasedpredation or other factors), the lesson is similarto the sardine. During a period of naturally lowsalmon population levels, human activity drovethose populations to unnaturally low levels. Insome cases, stocks became extinct and more arein danger of following the same path.
In the Pacific northwest, the regime shift of1976-1977 is associated with changes in themigration patterns and abundance of other fishspecies. The population of Pacific hake,Merluccius productus, found off Oregon,Washington and British Columbia in summermay have modified its spawning behavior.Until recently the stock migrated to thesouthern California bight to spawn duringboreal winter. Now, juvenile hake are caughtoff Oregon by the shrimp trawl fisherysuggesting that hake now spawn much furthernorth.
There appear to be orderly changes indominance among coastal pelagic fishes(McCall 1996). Coastal pelagic fish abundancessince 1930 indicate an orderly sequence of four
or five dominant species with cycles that arevery similar to those observed in Japan-peaksin abundance of mackerel (Scomber japonicus)are followed by sardines, then bonito (Sardachiliensis) and anchovies, with each peakseparated in time by roughly 10 years. As theabundance of the coastal pelagic speciesincreases in concert with the warming of theCalifornia current, the distributions shiftnorthward. Pacific mackerel are now commonin the northern California current, in Canadianwaters and into the Gulf of Alaska, and thesardine are conspicuous in waters of Oregon,Washington and off Vancouver Island, and areknown to be spawning off the coast of Oregon(Bently et al. 1996).
Another influence on the abundance of fishpopulations may be the Metonic (18.6-year)cycle, which has been shown to correlate withlarval abundance of the Pacific halibut,Hippoglossus stenolepis, (Fig. 17, Parker et al.1995). The importance of the Metonic cycle orits possible mechanism in driving halibut orother oceanic fish populations is unknown, butit should be investigated as to whether theobserved patterns are spurious correlations orindicative of a physical forcing.
Fig. 17: The Pacific halibut recruitment biomass record, 1927-1983 (YCS = year class strength),with the ‘best fit’ lagged 18.6 year lunar nodal tide signal (Mn). The abundance series has beencorrected for regional catchabilities, gear selectivities, weights-at-age, and incidental bycatch, andsmoothed with three years moving average (after Parker et al. 1995).Registro de la biomasa de reclutamiento para el lenguado del Pacífico, 1927-83 (YCS = importancia de la clase de edad)con el mejor ajuste retrasado en 18.6 años con respecto a la señal lunar nodal (Mn). La serie de abundancia ha sidocorregida por la habilidad de capturas regionales, selectividad del arte de pesca, peso a la edad de captura, capturasincidentales, y suavizada con un promedio móvil de 3 años (en base a Parker et al. 1995).

400 HALPIN ET AL.
Coho salmon management as an example
Coho salmon reside in the ocean duringapproximately half of their life. Adults migrateinto their natal streams (or to their natalhatchery) in fall and lay eggs, which in turnproduce offspring that spend one year in freshwater before migrating to sea as 1+ year old fish.The fish then spend 1.5 years in the ocean beforereturning to freshwater. Most of the mortalityduring the oceanic phase is thought to take placeduring the first 6-8 weeks at sea (Pearcy 1992).
Reliable estimates of the abundance ofOregon-California coho salmon are availableonly from 1970 until present. Those data showthat abundances during the 1970s rangedbetween 2 and 4 million fish. Abundancesreached a peak of 4.5 million fish in 1976 andhave declined ever since. Two periods of rapiddecline occurred: between 1976 and 1977,numbers declined three-fold, from 4.5 to 1.5million, and between 1991 and 1992, numbersdeclined four-fold, from 2 million to 500,000fish. The rapid change in numbers in 1976 issimilar in scope to what was noted above forAlaska sockeye and pink salmon (Francis &Hare 1994). Abundances of coho declined in1976 in part because survival declined (Fig.18). The survival time series shows twodramatic drops: for coho that entered the oceanin spring 1976 and for those entering in 1991.Over the first period of decline, harvest levelsof coho remained high through the early 1980s(Fig. 19). Thus, given that the fish wereharvested at very high rates, the changes inabundance in the early 1980s were causedprimarily by both fishing and climate drivenchanges in ocean conditions. This is not to saythat watershed degradation and hydroelectricdams were not also contributing factors todeclines. For the 1990s, a different set ofcircumstances operated.
Wild coho salmon stocks in centralCalifornia, northern California/ southernOregon, and coastal Oregon are listed asthreatened species under the ESA, otherevolutionarily significant units are candidatesfor listing as threatened (1995, 1996, USFWS1997, USNMFS 1998). The 1983 El Niño eventawakened managers to the fact that the cohostocks were in trouble. Due to over fishing andpoor ocean conditions, coho abundances haddeclined greatly as compared to the 1960s and1970s. Managers implemented regulations atthe start of the 1984 fishing season that led toreduced catches, through a shortening of thelength of the fishing season. In addition,harvest quotas were established. Catches were
carefully monitored and the fishery terminatedwhen quotas were reached. The stocks began torebound in 1986 suggesting that controls onharvest were working. However, the “recovery”only lasted for about five years. Between 1991and 1993 exploitation rates were reducedgreatly and were reduced further in 1994 and1995 but the population did not respondpositively. Survival dropped below 1 % in1991, a level which could not tolerate anyfishing whatsoever. The abundance of wildcoastal coho may be less than 500,000 animals,although estimates of population size remaincontroversial. The cause of this recentcatastrophic decline may be due to the extendedEl Niño-like conditions that have persisted offOregon since 1992 and the severe El Niño of1997-1998. Thus, in the present situation, thedecline in abundance has been due to acombination of poor survival and poor oceanconditions as opposed to the case in 1976,when numbers declined as a result of overfishing along with an as yet unquantifiedchange in ocean conditions.
It is now over 20 years since the regime shiftof 1976-1977 and there are as yet no explanationsthat account for the response of the ecosystem tothe event. A large number of papers have beenpublished which document physical andbiological changes and show correlations amongchanges in biomass, migration patterns ordistribution of resource species with climate,atmospheric or physical oceanographic variables.Few published papers have suggestedmechanisms; few have demonstrated howchanges in physical, trophic factors oranthropogenic factors might lead to the observedchanges in the density or productivity of a givenresource. Our ignorance may remain if thesuggestions of Ware (1995) and McCall (1996)are true - they argue that we may now beexperiencing another regime shift. The mostextreme ENSO event of this century occurred in1997-1998, with dramatic impacts worldwide.Almost uninterrupted El Niño-like conditionshave occurred off Oregon since 1992, whichbodes poorly for coho. The scientific researchcommunity must educate the public about howclimate variability and climate change affectsresources. Managers of living marine resources atthe federal, state and local levels need to beinformed of recurring regime shifts because mostmanagement plans incorrectly assume the oceandoes not contribute significant variability tospawning stock biomass. A better understandingof how the ecosystem as a whole responds toregime shifts may be the key to understandingstock responses.
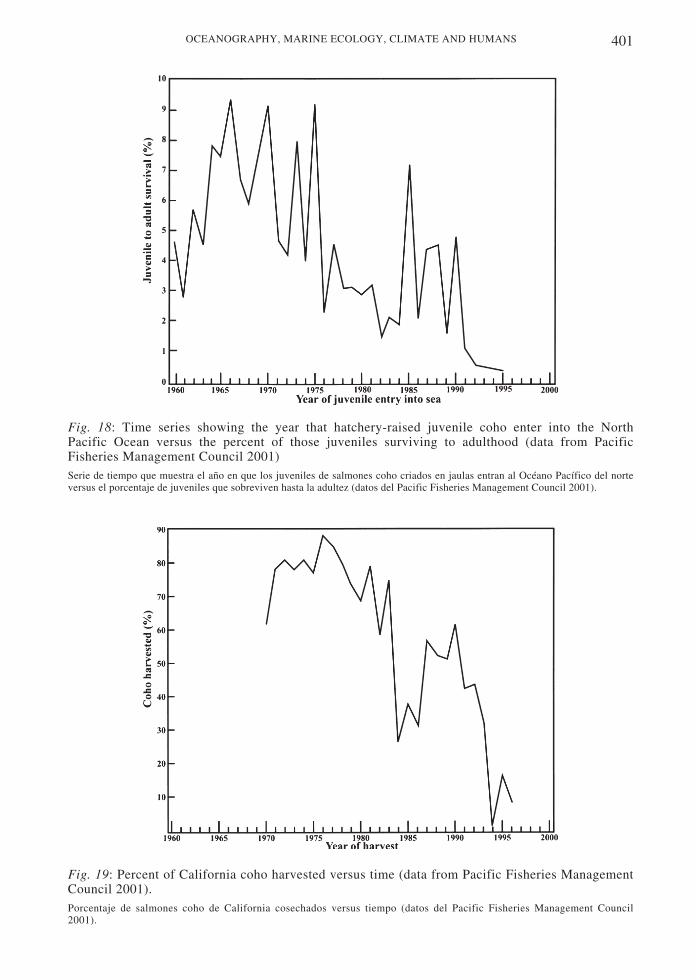
401OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
Fig. 19: Percent of California coho harvested versus time (data from Pacific Fisheries ManagementCouncil 2001).Porcentaje de salmones coho de California cosechados versus tiempo (datos del Pacific Fisheries Management Council2001).
Fig. 18: Time series showing the year that hatchery-raised juvenile coho enter into the NorthPacific Ocean versus the percent of those juveniles surviving to adulthood (data from PacificFisheries Management Council 2001)Serie de tiempo que muestra el año en que los juveniles de salmones coho criados en jaulas entran al Océano Pacífico del norteversus el porcentaje de juveniles que sobreviven hasta la adultez (datos del Pacific Fisheries Management Council 2001).

402 HALPIN ET AL.
TOWARD AN ECO-OCEANOGRAPHIC INTEGRATION
Evidence supports the existence of naturalfluctuations in basin-scale physicalenvironmental conditions that includeatmospheric pressure systems, wind forcing,precipitation, surface temperature and mixedlayer depth. In the oceanic ecosystems, changesin nutrient fluxes, phytoplankton andzooplankton concentrations are linked to thephysical changes. However, communities wherebiotic interactions have been well studied, suchas the rocky shore, and where bothoceanographic and biotic factors have beenstudied, such as kelp forests, demonstrate thatcommunity wide effects of physical parametersare always mediated by biotic interactions. Forexample, the oceanographic effects of El Niñounderlie dramatic losses in kelp biomass inNorth America. One could correctly correlatethe biomass loss with physical factors such asnutrient depletion and storms and incorrectlyconclude that nutrient depletion and stormsdirectly accounted for the total kelp die off. Indoing so the important role that urchins and seaotters play in kelp forest dynamics would becompletely overlooked. Conversely, withoutknowledge of oceanographic conditions, theframework that underlies biotic factors, such asthe loss of drift kelp forcing urchins to graze onstanding kelp, would be missed. Similarly, in the1960s, the collapse of the California sardinefishery was widely believed to be due solely tooverfishing. The evidence presented abovesuggests that it was due to continued fishing atprevious level while the population was in aperiod of natural decline.
The dominant time scales of changes inmany marine resources tends to be severaldecades, which have historically beenattributed to human exploitation alone. Asdescribed above, there is growing evidence ofnatural variability of the environment andpopulations on these time scales (PDO,Metonic cycle, ENSOs), although it is usuallydifficult to separate natural variability fromhuman influences. This raises the possibility ofinteractions between human actions and theselonger, and consequently harder to recognize,natural cycles. Failure to understand and detectthese cycles and modify harvesting pressuresmay result in substantial losses of harvestablemarine populations (as in the case of theCalifornia sardine), which would haverebounded if they had been allowed to remainat viable levels during their natural low phases.
Whatever lies ahead, it is now well establishedthat the productivity of eastern boundary currents
oscillates at frequencies of 20-60 years and islinked to decadal scale variability in theatmospheric circulation. These low frequencyoscillations in productivity explain thespectacular collapse of the Monterey sardine inthe late 1940s (and their recovery in the 1990s),the collapse of the Peruvian anchovy in the 1960s(and their “miraculous” recovery in the 1990s),and may partially explain the demise of the cohosalmon in the Pacific northwest. Such collapses/recoveries of commercially valuable fish stocksseem to be inevitable consequences of ourinadequate understanding of these ecosystems. Inorder to reduce the impact of boom-bust cycles onsocial structure and economies of localcommunities, we need improved resourcemanagement models based on understanding ofatmospheric and oceanic factors that underliethese oscillations, improved capability to predictthese regime shifts, and a better understanding ofcommunity dynamics. It is doubtful that adversefluctuations in the stocks can be avoided entirely,but if management were armed with the aboveknowledge and acted appropriately, it should bepossible to reduce the severity and duration of thedownturns and their resultant economic and socialhardships.
Clearly, to understand and manage benthicand oceanic ecosystems, an integration ofoceanography and ecology is required. Thisincludes an understanding of both small-scalebiotic interactions and large-scale climacticfluctuations. Unfortunately progress has beenthwarted by a historical separation of thedisciplines intellectually and in the case ofnearshore benthic ecology versus open oceanoceanography, a physical gap as well. Logisticfactors, such as the difficulty of investigatingbiotic interactions in the open ocean have alsoimpeded progress. However, integration ispossible with collaboration between ecologistsand oceanographers, and a better understandingof how marine communities respond to large-scale oceanographic fluctuations will result.
ACKNOWLEDGEMENTS
The authors wish to thank Bruce Menge, JangeLubchenco and the members of their laboratoryfor their input on the manuscript. Writing of hismanuscript was supported in part by the AndrewJ. Mellon Foundation and the David and LucilePackard Foundation. This is contribution number28 of the Partnership for InterdisciplinaryStudies of Coastal Oceans (PISCO): A Long-Term Ecological Consortium funded by theDavid and Lucile Packard Foundation.

403OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
LITERATURE CITED
AINLEY DG, WJ SYDEMAN & J NORTON (1995)Upper trophic level predators indicate interannualnegative and positive anomalies in the Californiacurrent food web. Marine Ecology Progress Series118: 69-79.
AINLEY DG, LB SPEAR & SG ALLEN (1996) Variationin the diet of Cassin’s auklet reveals spatial,seasonal, and decadal occurrence patterns ofEuphausiids off California, USA. Marine EcologyProgress Series, 137: 1-10.
ALLISON GW, J LUBCHENCO & MH CARR (1998)Marine reserves are necessary but not sufficient formarine conservation. Ecological Applications 8:S79-S92.
ARNTZ WE & J TARAZONA (1990) Effects of El Niño1982-1983 on benthos, fish and fisheries off theSouth American Pacific coast. In: Glynn PW (ed)Global ecological consequences of the 1982-1983El Niño Southern Oscillation (1989): 323-360.Elsevier, Amsterdam, The Netherlands.
BAKUN A (1990) Global climate change and theintensification of coastal ocean upwelling. Science247: 198-201.
BARBER RT & FP CHÁVEZ (1983) Biologicalconsequences of El Niño. Science 222: 1203-1210.
BARRY JP, CH BAXTER, RD SAGARIN & SE GILMAN(1995) Climate-related, long-term faunal changes ina California rocky intertidal community. Science267(5198): 672-675.
BARTH JA & RL SMITH (1998) Separation of a coastalupwelling jet at Cape Blanco, Oregon, USA. SouthAfrican Journal of Marine Science 19: 5-14.
BARTH JA, SD PIERCE & RL SMITH (2000) Aseparating coastal upwelling jet at Cape Blanco,Oregon, and its connection to the California currentsystem. Deep-Sea Research Part II 47: 783-810.
BAUMGARTNER TR, A SOUTAR & V FERREIRA-BARTRINA (1992) Reconstruction of the history ofpacific sardine and northern anchovy populationsover the past two millennia from sediments of theSanta Barbara Basin, California. CaliforniaCooperative Oceanic Fisheries InvestigationsReports 33: 24-40.
BEAMISH RJ (ed) (1995). Climate change and northernfish populations. Canadian Special Publications inFisheries and Aquatic Sciences. NRC Press,Ottawa, Canada. 739 pp.
BEATLY T (1991) Protecting biodiversity in coastalenvironments: introduction and overview. CoastalManagement 19:1-19.
BENTLY PJ, RJ EMMETT, NCH LO & HG MOSER(1996) Egg production of pacific sardine (Sardinopssagax) off Oregon in 1994. CoICOFI Repport.
BLANCO JL, AC THOMAS, ME CARR & PT STRUB(2001) Seasonal climatology of hydrographicconditions in the upwelling region off northernChile. Journal of Geophysical Research-Oceans106: 11451-11467.
BLANCO JL, ME CARR, AC THOMAS & PT STRUB(2002) Hydrographic conditions off northern Chileduring the 1996-1998 La Niña and El Niño events.Journal of Geophysical Research-Oceans: 107.
BODKIN JL (1989) Effects of kelp forest removal onassociated fish assemblages in central California.Journal of Experimental Marine Biology andEcology 117: 227-238.
BOTSFORD LW, LE MORGAN, DR LOCKWOOD & JEWILEN (1999) Marine reserves and management of
the northern California red sea urchin fishery.California Cooperative Oceanic FisheriesInvestigations Reports 40: 87-93.
BRANDHORST W (1971) Condiciones oceanográficasestivales frente a la costa de Chile. Revista deBiología Marina, Valparaíso (Chile) 14: 45-84.
BREEN PA, TA CARSON, JB FOSTER & EA STEWART(1982) Changes in subtidal community structureassociated with British Columbia sea otter transplants.Marine Ecology Progress Series 7: 12-20.
BRINK KH & TJ COWLES (1991) The coastal transitionzone program. Journal of Geophysical Research 96:14637-14647.
BRINK KH, D HALPERN, A HUYER & RL SMITH(1983) The physical environment of the Peruvianupwelling system. Progress in Oceanography 12:285-306.
BRINK KH, BH JONES, JCV LEER, CNK MOOERS,DW STUART, MR STEVENSON, RC DUGDALE& GW HEBURN (1981) Physical and biologicalstructure and variability in an upwelling center offPeru near 15º S during March 1977. In: RichardsFA (ed) Coastal upwelling: 473-495. AmericanGeophysical Union, Washington, Distr ict ofColumbia, USA.
BRODEUR RD, BW FROST, SR HARE, RC FRANCIS &WJ INGRAHAM JR. (1996) Interannual variationsin zooplankton biomass in the gulf of Alaska, andcovariation with California current zooplanktonbiomass. California Cooperative Oceanic FisheriesInvestigations Reports 37: 80-99.
BROITMAN BR, SA NAVARRETE, F SMITH & SDGAINES (2001) Geographic variat ion ofsoutheastern Pacific intertidal communities. MarineEcology Progress Series 224: 21-34.
CANCINO JM & JC CASTILLA (1988) Emersion behaviourand foraging ecology of the common Chilean clingfishSicyases sanguineus (Pisces: Gobiesocidae). Journalof Natural History 22: 249-261.
CASTILLA JC (1988) Earthquake-caused coastal upliftand its effects on rocky intertidal kelp communities.Science 242: 440-443.
CASTILLA JC (1994) The Chilean small-scale benthicshellfisheries and the institutionalization of newmanagement practices. Ecology InternationalBulletin 21: 47-63.
CASTILLA JC (1999) Coastal marine communities: trendsand perspectives from human-exclusionexperiments. Trends in Ecology and Evolution 14:280-283.
CASTILLA JC (2000) Roles of experimental marineecology in coastal management and conservation.Journal of Experimental Marine Biology andEcology 250: 3-21.
CASTILLA JC & RT PAINE (1987) Predation andcommunity organization on eastern Pacific,temperate zone, rocky intertidal shores. RevistaChilena de Historia Natural 60: 131-151.
CASTILLA JC & PA CAMUS (1992) The Humboldt-ElNiño scenario: coastal benthic resources andanthropogenic influences, with particular referenceto the 1982/1983 ENSO. In: Payne AIL, KH Brink,KH Mann & R Hilbom (eds) Benguela TrophicFunctioning: 703-712. South African Journal ofMarine Science.
CASTILLA JC & M FERNÁNDEZ (1998) Small-scalebenthic fisheries in Chile: on co-management andsustainable use of benthic invertebrates. EcologicalApplications 8: S124-S132.
CASTILLA JC & O DEFEO (2001) Latin Americanbenthic shellfisheries: emphasis on co-management

404 HALPIN ET AL.
and experimental practices. Reviews in FishBiology and Fisheries 11: 1-30.
CASTILLA JC, SA NAVARRETE & J LUBCHENCO(1993) Southeastern Pacific coastal environments:main features, large-scale perturbations, and globalclimate change. In: Mooney HA, ER Fuentes & BIKronberg (eds) Earth system responses to globalchange: 167-188. Academic Press, New York, NewYork, USA.
CHÁVEZ FP (1996) Forcing and biological impact ofonset of the 1992 El Niño in central California.Geophysical Research Letters 23: 265-268.
CHÁVEZ FP, J RYAN, SE LLUCH-COTA & MCNIQUEN (2003) From anchovies to sardines andback: multidecadal change in the Pacific ocean.Science 299: 217-221.
CHELTON DB (1981) Interannual variability of theCalifornia current: physical factors. CaliforniaCooperative Oceanic Fisheries InvestigationsReports 22: 130-148.
CHELTON DB (1982) Seasonal variability of alongshoregeostrophic velocity off central California. Journalof Geophysical Research 89: 3473-3486.
CHELTON DB & RE DAVIS (1982) Monthly mean sea levelvariability along the west coast of North America.Journal of Physical Oceanography 12: 757-784.
CHELTON DB, PA BERNAL & JA MCGOWAN (1982)Large-scale interannual physical and biologicalinteraction in the California current. Journal ofMarine Research 40: 1095-1125.
COHEN JE, F BRIAND & CM NEWMAN (eds) (1990)Community food webs: data and theory. Springer-Verlag, Berlin, Germany. 308 pp.
CONNOLLY SR & J ROUGHGARDEN (1998) A latitudinalgradient in northeast Pacific intertidal communitystructure: evidence for an oceanographically basedsynthesis of marine community theory. AmericanNaturalist 151: 311-326.
CONNOLLY SR, BA MENGE & J ROUGHGARDEN(2001) A latitudinal gradient in recruitment ofintertidal invertebrates in the northeast Pacificocean. Ecology 82: 1799-1813.
CUBIT JD (1984) Herbivory and the seasonal abundanceof algae on a high intertidal rocky shore. Ecology65: 1904-1917.
CURRIE RG (1991) Deterministic signals in tree-ringsfrom Tasmania, New Zealand, and South Africa.Annals Geophysicae 9: 71-81.
CURRIE RG (1992) Deterministic signals in tree-ringsfrom Europe. Annals Geophysicae 10: 241-253.
CURRIE RG (1993) Luni-solar 18-6- and solar cycle 10-11-year signals in USA air temperature records.International Journal of Climatology 13: 31-50.
DAHLHOFF EP & BA MENGE (1996) Influence ofphytoplankton concentration and wave exposure onthe ecophysiology of Mytilus californianus. MarineEcology Progress Series 144: 97-107.
DAVIS GE, PL HAAKER & DV RICHARDS (1996)Status and trends of white abalone at the CaliforniaChannel Islands. Transactions of the AmericanFisheries Society 125: 42-48.
DAYTON PK (1971) Competition, disturbance, andcommunity organization: The provision andsubsequent utilization of space in a rocky intertidalcommunity. Ecological Monographs 41: 351-389.
DAYTON PK (1985) The structure and regulation of someSouth American kelp communities. EcologicalMonographs 55: 447-468.
DAYTON PK & MJ TEGNER (1984) Catastrophic storms,El Niño, and patch stability in a southern Californiakelp community. Science 224: 283-285.
DAYTON PK & MJ TEGNER (1990) Bottoms beneathtroubled waters: benthic impacts of the 1982-1984El Niño in the temperate zone. In: Glynn PW (ed)Global ecological consequences of the 1982-1983El Niño-Southern Oscillation (1989): 433-472.Elsevier, Amsterdam, The Netherlands.
DAYTON PK, MJ TEGNER, PE PARNELL & PBEDWARDS (1992) Temporal and spatial patterns ofdisturbance and recovery in a kelp forestcommunity. Ecological Monographs 62: 421-445.
DAYTON PK, MJ TEGNER, PB EDWARDS & KLRISER (1998) Sliding baselines, ghosts, andreduced expectations in kelp forest communities.Ecological Applications 82: 309-322.
DAYTON PK, MJ TEGNER, PB EDWARDS & KLRISER (1999) Temporal and spatial scales of kelpdemography: The role of oceanographic climate.Ecology 69: 219-250.
DENBO DW & JS ALLEN (1987) Large-scale response toatmospheric forcing of shelf currents and coastalsea level off the west coast of North America: May-July, 1981-1982. Journal of Geophysical Research92: 1757-1782.
DENNY MW & RT PAINE (1998) Celestial mechanics,sea level changes, and intertidal ecology. BiologicalBulletin 194: 108-115.
DOROFF AM, JA ESTES, MT TINKER, DM BURN & TJEVANS (2003) Sea otter population declines in theAleutian archipelago. Journal of Mammalogy 84:55-64.
DUGGINS D (1980) Kelp beds and sea otters: anexperimental approach. Ecology 61: 447-453.
DUGGINS D, CA SIMENSTAD & JA ESTES (1989)Magnification of secondary production by kelp detritusin coastal marine ecosystems. Science 245: 170-173.
DURÁN LR, JC CASTILLA & D OLIVA (1987) Intensityof human predation on rocky shores at Las Crucesin central Chile. Environmental Conservation 14:143-149.
DURÁN R & JC CASTILLA (1989) Variation andpersistence of the middle rocky intert idalcommunity, of central Chile, with and withouthuman harvesting. Marine Biology 103: 555-562.
EBELING AW, DR LAUR & RJ ROWLEY (1985) Severestorm disturbances and reversal of communitystructure in a southern California kelp forest.Marine Biology 84: 287-294.
EBERT TA & MP RUSSELL (1988) Latitudinal variationin size structure of the west coast purple sea urchin:A correlation with headlands. Limnology andOceanography 33: 286-294.
EMERY WJ & K HAMILTON (1985) Atmosphericforcing of interannual variability in the northeastPacific ocean connections with El-Niño. Journal ofGeophysical Research-Oceans 90: 857-868.
ENFIELD D & JS ALLEN (1980) On the structure anddynamics of monthly mean sea level anomaliesalong the Pacific coast of North and South America.Journal of Physical Oceanography 107: 5757-5787.
ENFIELD D, M CORNEJO-RODRÍGUEZ, RL SMITH &PA NEWBERGER (1987) The equatorial source ofpropagating variability along the Peru coast duringthe 1982-1983 El Niño. Journal of GeophysicalResearch 92: 14335-14346.
ENFIELD DB & AM MESTAS-NÚNEZ (1999) Multiscalevariabilities in global sea surface temperatures andtheir relationships with tropospheric climatepatterns. Journal of Climate 12: 2719-2733.
ESTES JA & JF PALMISANO (1974) Sea otters: their rolein structuring nearshore communities. Science 185:1058-1060.

405OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
ESTES JA & D DUGGINS (1995) Sea otters and kelpforests in Alaska: generality and variation in acommunity ecological paradigm. EcologicalMonographs 65: 75-100.
ESTES JA, NS SMITH & JF PALMISANO (1978) Seaotter predation and community organization inwestern Aleutian Islands, Alaska. Ecology 59: 822-833.
ESTES JA, MT TINKER, TM WILLIAMS & DF DOAK(1998) Killer whale predation on sea otters: linkingoceanic and nearshore ecosystems. Science 282:473-476.
FARRELL TM, D BRACHER & J ROUGHGARDEN(1991) Cross-shelf transport causes recruitment tointert idal populations in central California.Limnology and Oceanography 36: 279-288.
FERNÁNDEZ M, E JARAMILLO, P MARQUET CMORENO, S NAVARRETE, P OJEDA, CVALDOVINOS & J VÁSQUEZ (2000) Diversity,dynamics and biogeography of Chilean benthicnearshore ecosystems: an overview and guidelinesfor conservation. Revista Chilena de HistoriaNatural 73: 797-830.
FIEDLER PC (1984) Satellite observations of the 1982-1983 El Niño along the US Pacific coast. Science224: 1251-1254.
FONSECA T (1985) Efectos físicos del fenómeno El Niño1982-1983 en la costa chilena. InvestigaciónPesquera (Chile) 32: 61-68.
FONSECA T (1989) An overview of the polewardundercurrent and upwelling along the Chilean coast.In: Neshyba SJ, CNK Mooers, RL Smith & RTBarber (eds) Poleward flows along eastern oceanboundaries: 203-228. Springer-Verlag, New York,New York, USA.
FONSECA T & M FARÍAS (1987) Estudio del proceso desurgencia en la costa chilena utilizando percepciónremota. Investigación Pesquera (Chile) 34: 33-46.
FOOD AND AGRICULTURE ORGANIZATION OF THEUNITED NATIONS (1993) FAO Yearbook ofFishery Statistics, 1991: 395. Food and AgriculturalOrganization of the United Nations, Rome, Italy.
FOOD AND AGRICULTURE ORGANIZATION OF THEUNITED NATIONS (1996) FAO Yearbook ofFishery Statistics, 1996: 179. Food and AgriculturalOrganization of the United Nations, Rome, Italy.
FRANCIS RC & SR HARE (1994) Decadal-scale regimeshifts in the large marine ecosystems of thenortheast Pacific: a case for historical science.Fisheries Oceanography 3: 279-291.
FRANK PW (1982) Effects of winter feeding on limpetsby black oystercatchers, Haematopus bachmani.Ecology 63: 1352-1362.
GARGETT AE (1997) The optimal stability “window”: amechanism underlying decadal fluctuations inNorth Pacific salmon stocks. FisheriesOceanography 6: 109-117.
GILL AE & AJ CLARKE (1974) Wind-induced upwelling,coastal currents and sea-level changes. Deep-SeaResearch 21: 325-345.
GODOY C & CA MORENO (1989) Indirect effects ofhuman exclusion from the rocky intertidal inSouthern Chile: a case of cross-linkage betweenherbivores. Oikos 54: 101-106.
GRAHAM MH (1996) Effect of high irradiance ofrecruitment of the giant kelp Macrocystis(Phaeophyta) in shallow water. Journal ofPhycology 32: 903-906.
GRAHAM NE (1994) Decadal scale variability in the1970’s and 1980’s: observations and model results.Climate Dynamics 10: 60-70.
GUNNILL FC (1985) Population fluctuations of sevenmacroalgae in southern California during 1981-1983 including effects of severe storms and an ElNiño. Journal of Experimental Marine Biology andEcology 85: 149-164.
HAHN T & M DENNY (1989) Tenacity mediatedselective predation by oystercatchers on intertidall impets and i ts role in maintaining habitatparti t ioning by Collisella scabra and Lottiadigitalis. Marine Ecology Progress Series 53: 1-10.
HARROLD C & DC REED (1985) Food availability, seaurchin grazing, and kelp forest communitystructure. Ecology 66: 1160-1169.
HARVELL CD, K KIM, JM BURKHOLDER, RRCOLWELL, PR EPSTEIN, DJ GRIMES, EEHOFMANN, EK LIPP, AD OSTERHAUS, RMOVERSTREET, JW PORTER, GW SMITH & GRVASTA (1999) Emerging marine diseases-climatelinks and anthropogenic factors. Science 285: 1505-1510.
HAYWARD TL, SL CUMMINGS, DR CAYAN, FPCHAVEZ, RJ LYNN, AW MANTYLA, PPNIILER, FB SCHWING, RR VEIT & ELVENRICK (1996) The state of the CaliforniaCurrent in 1995-1996: continuing declines inmacrozooplankton biomass during a period ofnearly normal circulation. California CooperativeOceanic Fisheries Investigations Reports 37: 22-37.
HELMUTH B, CDG HARLEY, PM HALPIN, MO’DONNELL, GE HOFMANN & CABLANCHETTE (2002) Climate change andlatitudinal patterns of intertidal thermal stress.Science 298: 1015-1017.
HICKEY BM (1979) The California Current system:hypotheses and facts. Progress in Oceanography 8:191-279.
HICKEY BM (1989) Patterns and processes of circulationover the continental shelf off Washington. In:Landry MR & BM Hickey (eds) Coastaloceanography of Washington and Oregon: 441-451.Elsevier, New York, New York, USA.
HICKEY BM (1998) Coastal oceanography of westernNorth America from the tip of Baja California toVancouver Island. In: Robinson AR & KH Brink(eds) The Sea: 345-394. John Wiley and Sons, Inc.,New York, New York, USA.
HOBDAY AJ & MJ TEGNER (2002) The warm and thecold: influence of temperature and fishing on localpopulation dynamics of red abalone. CaliforniaCooperative Oceanic Fisheries InvestigationsReports 43: 74-96.
HOFFMANN A & B SANTELICES (1997) Flora marinade Chile central. Ediciones Universidad Católica deChile, Santiago, Chile. 434 pp.
HOLLOWED AB & WS WOOSTER (1992) Variability ofwinter ocean conditions and strong year classes ofnortheast Pacific groundfish. ICES Marine ScienceSymposia 195: 433-444.
HUYER A, M KNOLL, T PALUSZKIEWICZ & RLSMITH (1991) The Peru undercurrent: a study invariability. Deep-Sea Research 38: S247-S271.
JARA HF & CA MORENO (1984) Herbivory andstructure in a midlittoral rocky community: a casein southern Chile. Ecology 65: 28-38.
KATO S & SC SCHROETER (1985) Biology of the red seaurchin, Strongylocentrotus franciscanus, and its fisheryin California. Marine Fisheries Review 47: 1-20.
KNUDSEN T & N VOGEL (1997) Sea creatures in tidepools occupy a precarious perch. In: San FranciscoSunday Examiner and Chronicle: B-6, SanFrancisco, USA.

406 HALPIN ET AL.
LAUR DR, AW EBELING & DA COON (1988) Effects ofsea otter foraging on subtidal reef communities offcentral California. In: Blaricom GRV & JA Estes(eds) The community ecology of sea otters: 151-167. Springer-Verlag, Berlin, Germany.
LIMBERGER D (1990) El Niño’s effect on SouthAmerican pinniped species. In: Glynn PW (ed)Global ecological consequences of the 1982-1983El Niño Southern Oscillation: 417-430. Elsevier,Amsterdam, The Netherlands.
LODER JW & C GARRETT (1978) The 18.6 year cycle ofsea surface temperature in shallow seas due tovariation in tidal mixing. Journal of GeophysicalResearch 83: 1967-1970.
LOHSE DP (1993) The effects of substratum type on thepopulation dynamics of three common intertidalanimals. Journal of Experimental Marine Biologyand Ecology 173: 133-154.
LUBCHENCO J, SA NAVARETTE, BN TISSOT & JCCASTILLA (1993) Possible ecological responses toglobal climate change: nearshore benthic biota ofnortheastern Pacific coastal ecosystems. In:Mooney HA, ER Fuentes & BI Kronberg (eds)Earth system responses to global change: contrastsbetween North and South America: 147-166.Academic Press, New York, New York, USA.
LUKAS R (1986) The termination of the equatorialundercurrent in the eastern Pacific. Progress inOceanography 16: 63-90.
LYNN RJ & JJ SIMPSON (1987) The California currentsystem: the seasonal variability of its physicalcharacteristics. Journal of Geophysical Research92: 2515-2534.
MAKSIMOV IV & NP SMIRNOV (1967) A contribution tothe study of the causes of long period variations inthe activity of the Gulf stream. Oceanology 5: 15-24.
MANRÍQUEZ PH & JC CASTILLA (2001) Significanceof marine protected areas in central Chile asseeding grounds for the gastropod Concholepasconcholepas. Marine Ecology Progress Series 215:201-211.
MANTUA NJ, SR HARE, Y ZHANG, JM WALLACE &RC FRANCIS (1997) A Pacific interdecadalclimate oscil lat ion with impacts on salmonproduction. Bullet in of the AmericanMeteorological Society 78: 1069-1079.
MAURER D & H NGUYEN (1997) The relationshipbetween on ocean outfall and sediment propertiesfrom the San Pedro shelf, California. BulletinSouthern California Academy of Sciences 96: 22-33.
MCCALL AD (1996) Patterns of low-frequency variabilityin fish populations of the California current.California Cooperative Oceanic FisheriesInvestigations Reports 37: 100-110.
MCGOWAN JA, DB CHELTON & A CONVERSI (1996)Plankton patterns, climate, and change in theCalifornia current. California Cooperative OceanicFisheries Investigations Reports 37: 45-69.
MCGOWAN JA, DR CAYAN & LM DORMAN (1998)Climate-ocean variability and ecosystem responsein the northeast Pacific. Science 281: 210-217.
MCLEAN JH (1962) Sublittoral ecology of kelp beds ofthe open coast near Carmel. Biological Bulletin122: 213-219.
MCNALLEY GI (1981) Satellite-tracked drift buoyobservations of the surface flow in the mid-latitudeNorth Pacific. Journal of Geophysical Research 86:8022-2030.
MENGE BA (1992) Community regulation: Under whatconditions are bottom up factors important in rockyshores? Ecology 73: 755-765.
MENGE BA, B DALEY & PA WHEELER (1996) Controlof interaction strength in marine benthiccommunities. In: Polis GA & R Winemuller (eds)Food webs: integration of pattern and process: 258-274. Chapman and Hall, New York, New York,USA.
MENGE BA, BA DALEY, PA WHEELER & PT STRUB(1997a) Rocky intert idal oceanography: anassociation between community structure andnearshore phytoplankton concentration. Limnologyand Oceanography 42: 57-66.
MENGE BA, EG BERLOW, CA BLANCHETTE, SANAVARRETE & SB YAMADA (1994) Thekeystone species concept: variation in interactionstrength in a rocky intertidal habitat. EcologicalMonographs 64: 249-286.
MENGE BA, BA DALEY, PA WHEELER, EDAHLHOFF, E SANFORD & PT STRUB (1997b)Benthic-pelagic l inks and rocky intert idalcommunities: bottom-up effects on top-downcontrol? Proceedings of the National Academy ofScience of the United States of America 94: 14530-14535.
MENGE BA, BA DALEY, J LUBCHENCO, ESANFORD, E DAHLHOFF, PM HALPIN, GHUDSON & JL BURNAFORD (1999) Top-downand bottom-up regulation of New Zealand rockyintertidal communities. Ecological Monographs 69:297-330.
MILLER AJ (1996) Recent advances in California currentmodeling: decadal and interannual thermoclinevariat ions. California Cooperative OceanicFisheries Investigations Reports 37: 69-79.
MILLER AJ, DR CAYAN, TP BARNETT, NE GRAHAM& JM OBERHUBER (1994) Interdecadal variabilityof the Pacific Ocean: model response to observedheat flux and wind stress anomalies. ClimateDynamics 9: 287-302.
MOORE JD, CA FINLEY, TT ROBBINS & CSFRIEDMAN (2002) Withering syndrome andrestoration of southern California abalonepopulations. California Cooperative OceanicFisheries Investigations Reports 43: 112-117.
MORENO CA (1986) Un resumen de las consecuenciasecológicas de la exclusión del hombre en la zonaintermareal de Mehuin -Chile. EstudiosOceanológicos (Chile) 5: 59-66.
MORENO CA & JP SUTHERLAND (1982) Physical andbiological processes in a Macrocystis pyriferacommunity near Valdivia, Chile. Oecologia 55: 1-6.
MORENO CA & E JARAMILLO (1983) The role ofgrazers in the zonation of intertidal macroalgae ofthe Chilean coast. Oikos 41: 73-76.
MORENO CA, KM LUNECKE & MI LÓPEZ (1986) Theresponse of an intertidal Concholepas concholepas(Gastropoda) population to protection from man insouthern Chile and the effects on benthic sessileassemblages. Oikos 46: 359-364.
MORENO CA, G ASCENSIO, WE DUARTE & VMARÍN (1998) Sett lement on the muricidConcolepas concholepas and its relationship withEl Niño and coastal upwellings in Chile. MarineEcology Progress Series 167: 171-175.
MYSAK LA (1986) El Niño, interannual variability andfisheries in the northeast Pacific Ocean. CanadianJournal of Fisheries and Aquatic Science 43: 464-497.
NATIONAL RESEARCH COUNCIL COPAMOPNAS(1996) Upstream: salmon and society in the PacificNorthwest. National Academy of Science Press,Washington, District of Columbia, USA. 452 pp.

407OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
NAVARRETE SA & BA MENGE (1996) Keystonepredation and interaction strength: interactiveeffects of predators on their main prey. EcologicalMonographs 66: 409-429.
NAVARRETE SA & JC CASTILLA (2003) Experimentaldetermination of predation intensity in an intertidalpredator guild: dominant versus subordinate prey.Oikos 100: 251-262.
NAVARRETE SA, B BROITMAN, EA WIETERS, GRFINKE, RM VENEGAS & A SOTOMAYOR(2002) Recruitment of intertidal invertebrates in thesoutheast Pacific: interannual variability and the1997-1998 El Niño. Limnology and Oceanography47: 791-802.
NESHYBA SJ, CNK MOOERS, RL SMITH & RTBARBER (eds) (1989). Poleward flows alongeastern ocean boundaries. Springer-Verlag, NewYork, New York, USA. 374 pp.
NIELSEN KJ (2001) Bottom-up and top-down forces intide pools: test of a food chain model in anintertidal community. Ecological Monographs 71:187-217.
NIELSEN KJ (2003) Nutrient loading and consumers:agents of change in open-coast macrophyteassemblages. Proceedings of the National Academyof Sciences of the United States of America 100:7660-7665.
NORTH W (1983) Separating the effects of temperatureand nutrients. Bascom W (ed). The effects of wastedisposal on kelp communities: 243-255. CaliforniaSea Grant College Program, California, USA.
NORTH WJ (1971) Introduction and background. In: NorthWJ (ed) The biology of giant kelp beds (Macrocystis)in California: 1-97. J Cramer, Lehre, USA.
OSHURKOV VV, AG BAZHIN, VI LUKIN & VFSEVOSTYANOV (1988) Sea otter predation andthe benthic community structure of the CommanderIslands. Biologia Morya (Vladivostok) 6: 50-60.
PACIFIC FISHERIES MANAGEMENT COUNCIL (2001)Review of the 2000 ocean salmon fisheries. PacificFisheries Management Council, Portland, Oregon,USA. 267 pp.
PAINE RT (1966) Food web complexity and speciesdiversity. American Naturalist 100: 65-76.
PAINE RT (1974) Intertidal community structure:experimental studies on the relationship between adominant competitor and its principal predator.Oecologia 15: 93-120.
PAINE RT (1976) Size-limited predation: an observationaland experimental approach with the Mytilus-Pisaster interaction. Ecology 57: 858-873.
PAINE RT (1980) Food webs: linkage, interaction strengthand community infrastructure. Journal of AnimalEcology 49: 667-685.
PAINE RT (1986) Benthic community-water columncoupling during the 1982-1983 El Niño. Arecommunity changes at high latitudes attributable tocause or coincidence? Limnology andOceanography 31: 351-360.
PAINE RT & AR PALMER (1978) Sicyases sanguineas: aunique trophic generalist from the Chilean intertidalzone. Copeia 1978: 75-81.
PAINE RT & SA LEVIN (1981) Intertidal landscapes:disturbance and the dynamics of pattern. EcologicalMonographs 51: 145-178.
PARES-SIERRA A & JJ O’BRIEN (1989) The seasonaland interannual variability of the California currentsystem: a numerical model. Journal of GeophysicalResearch 94: 3159-3180.
PARKER KS, TC ROYER & RB DERISO (1995) Highlatitude climate forcing and tidal mixing by the 18.6
year lunar nodal cycle and low frequencyrecruitment trends in Pacific halibut (Hippoglossusstenolepis). In: Beamish RJ (ed) Climate changeand northern fish populations: 447-459. CanadianSpecial Publications in Fisheries and AquaticSciences. NRC Press, Ottawa, Canada. 739 pp.
PARRISH RH (1997) Regime-scale fluctuations in thecirculation of the North Pacific. In: Emmett RL &MH Schiewe (eds) Estuarine and ocean survival ofnortheastern Pacific salmon: NMFS-NWFSC-29.95104 pp.
PEARCY WG (1992) Ocean ecology of North Pacificsalmonids. University of Washington Press, Seattle,Washington, USA. 179 pp.
PEARSE JS & AH HINES (1987) Long term populationdynamics of sea urchins in a central California kelpforest: rare recruitment and rapid decline. MarineEcology Progress Series 39: 275-283.
PETERSON W (1998) Life cycle strategies of copepods incoastal upwelling zones. Journal of Marine Systems15: 313-326.
PFISTER CA & A BRADBURY (1996) Harvesting red seaurchins: recent effects and future predictions.Ecological Applications 6: 298-310.
PIERCE SD, RL SMITH, PM KOSRO, JA BARTH & CDWILSON (2000) Continuity of the polewardundercurrent along the eastern boundary of the mid-latitude north Pacific. Deep-Sea Research 47: 811-829.
POLOVINA JJ, GT MITCHUM & GT EVANS (1995)Decadal and basin-scale variation in mixed-layerdepth and the impact on biological production inthe central and North Pacific, 1960-1988. Deep-SeaResearch 42: 1701-1706.
POULIN E, AT PALMA, G LEIVA, E HERNÁNDEZ, PMARTÍNEZ, SA NAVARRETE & JC CASTILLA(2002a) Temporal and spatial variation in thedistribution of epineustonic competent larvae ofConcholepas concholepas along the central coast ofChile. Marine Ecology Progress Series 229: 95-104.
POULIN E, AT PALMA, G LEIVA, D NARVÁEZ, RPACHECO, SA NAVARRETE & JC CASTILLA(2002b) Avoiding offshore transport of competentlarvae during upwelling events: The case of thegastropod Concholepas concholepas in CentralChile. Limnology and Oceanography 47: 1248-1255.
PREISENDORFER RW (1988) Principal componentanalysis in meteorology and oceanography.Elsevier, New York, New York, USA. 425 pp.
RAY GC, BP HAYDEN, AJ BULGAR JR& MGMCCORMICK-RAY (1992) Effects of globalwarming on the biodiversity of coastal marinezones. In: Peters RL & TE Lovejoy (eds) Globalwarming and biodiversity: 91-104. Yale UniversityPress, New Haven, Connecticut, USA.
ROBLES CD (1997) Changing recruitment in constantspecies assemblages: implications for predationtheory in intertidal communities. Ecology 78: 1400-1414.
ROBLES CD & J CUBIT (1981) Influence of bioticfactors in an upper intertidal community: dipteranlarvae grazing on algae. Ecology 62: 1536-1547.
RODRÍGUEZ SR (2003) Consumption of drift kelp byintertidal populations of the sea urchin Tetrapygusniger on the central Chilean coast: possibleconsequences at different ecological levels. MarineEcology Progress Series 251: 141-151.
ROEMMICH D & J MCGOWAN (1995) Climaticwarming and the decline of zooplankton in theCalifornia current. Science 467: 1324-1326.

408 HALPIN ET AL.
ROGERS-BENNETT L, PL HAAKER, TO HUFF & PKDAYTON (2002a) Estimating baseline abundancesof abalone in California for restoration. CaliforniaCooperative Oceanic Fisheries InvestigationsReports 43: 97-111.
ROGERS-BENNETT L, D BERGEN, P COLLIER, MERICKSON, P KALVASS, C KONG, J O’BRIEN,K ODA, J O’LEARY, D ONO, K O’REILLY, DOSORIO, K WALTERS, R LEOS, V TAYLOR, DTHOMAS, A VEJAR & D SWEETNAM (2002b)Review of some California fisheries for 2001:market squid, sea urchin, dungeness crab, lobster,prawn, abalone, groundfish, swordfish and shark,coastal pelagic finfish, ocean salmon, nearshorelive-fish, Pacific herring, white seabass, and kelp.California Cooperative Oceanic FisheriesInvestigations Reports 43: 13-30.
ROUGHGARDEN J, S GAINES & H POSSINGHAM(1988) Recruitment dynamics in complex life cyles.Science 241: 1397-1560.
ROUGHGARDEN J, JT PENNINGTON, D STONER, SALEXANDER & K MILLER (1991) Collisions ofupwelling fronts with the intertidal zone: the causeof recruitment pulses in barnacle populations ofcentral California. Acta Oecologica 12: 35-51.
ROYER TC (1993) High latitude oceanic variabilityassociated with the 18.6-year nodal tide. Journal ofGeophysical Research 98: 4639-4644.
SAGARIN RD, JP BARRY, SE GILMAN & CH BAXTER(1999) Climate-related change in an intertidalcommunity over short and long time scales.Ecological Monographs 69: 465-490.
SANFORD E (1999) Regulation of keystone predation bysmall changes in ocean temperature. Science 283:2095-2097.
SANFORD E, MS ROTH, GC JOHNS, JP WARES & GNSOMERO (2003) Local selection and latitudinalvariation in a marine predator-prey interaction.Science 300: 1135-1137.
SANTELICES B & FP OJEDA (1984) Populationdynamics of a coastal forest of Macrocystis pyriferain Puerto Toro, Isla Navarino, southern Chile.Marine Ecology Progress Series 14: 175-183.
SANTELICES B & R NORAMBUENA (1987) Aharvesting strategy for Iridaea laminarioides incentral Chile. In: Ragan MA & CJ Bird (ed)Hydrobiologia, Twelfth International SeaweedSymposium: 329-333. Dr. W. Junk Publishers,Dordrecht, The Netherlands.
SANTELICES B, S MONTALVA & P OLIGER (1981)Competitive algal community organization inexposed intertidal habitats from central Chile.Marine Ecology Progress Series 6: 267-276.
SANTELICES B, JC CASTILLA, J CANCINO & PSCHMIEDE (1980) Comparative ecology of Lessonianigrescens and Durvillaea antarctica (Phaeophyta) incentral Chile. Marine Biology 59: 119-132.
SCHIEL DR & MS FOSTER (1986) The structure ofsubtidal algal stands in temperate waters.Oceanography and Marine Biology Annual Review24: 265-307.
SCHROETER SC, TA DEAN, K THIES & JD DIXON(1995) Effects of shading by adults on the growthof blade stage Macrocystis pyrifera (Phaeophyta)during and after the 1982-1984 El Nino. Journal ofPhycology 31: 697-702.
SCHWARTZLOSE RA, J ALHEIT, T BAUMGARTNER,R CLOETE, RJM CRAWFORD, WJ FLETCHER,Y GREEN-RUIZ, E HAGEN, T KAWASAKI, DLLUCH-BELDA, SE LLUCH-COTA, ADMACCALL, Y MATSUURA, MO NEVIREZ-
MARTÍNEZ, RH PARRISH, C ROY, R SERRA,KV SHUST, NM WARD & JZ ZUZUNAGA (1999)Worldwide large-scale fluctuations of sardine andanchovy populations. South African Journal ofMarine Science 21: 289-347.
SCHWING FB & R MENDELSSOHN (1997) Increasedcoastal upwelling in the California current system.Journal of Geophysical Research 120: 3421-3438.
SHAFFER G, O PIZARRO, L DJURFELDT, S SALINAS &J RUTLLANT (1997) Circulation and low-frequencyvariability near the Chilean coast: remotely forcedfluctuations during the 1991-92 El Niño. Journal ofPhysical Oceanography 27: 217-235.
SHEPHERD SA, JR TURRUBIATES-MORALES & KHALL (1998) Decline of the abalone fishery at LaNatividad, Mexico: overfishing or climate change?Journal of Shellfish Research 17(3): 839-846.
SILVA N & S NESHYBA (1979) On the southernmostextension of the Peru-Chile undercurrent. Deep-SeaResearch 26: 1387-1393.
SIMENSTAD CA, JA ESTES & KW KENYON (1978)Aleuts, sea otters, and alternate stable-statecommunities. Science 200: 403-411.
SIMPSON JJ (1984) A simple model of the 1982-1983California “El Niño”. Geophysical Research Letters11: 237-240.
SMITH RL (1992) Modern ocean upwell ing. In:Summerhayes CP, WL Prell & KC Emeis (eds)Upwelling systems: evolution since the earlyMiocene: 9-28. Geological Society, London,Special Publication, London, United Kingdom.
SOTO R (1985) Efectos del fenómeno El Niño. 1982-1983El Niño ecosistemas de la primera región.Investigación Pesquera (Chile) 32: 199-206.
STRUB PT & C JAMES (1995) The large-scale summercirculation of the California current. GeophysicalResearch Letters 22: 207-210.
STRUB PT & C JAMES (2000) Altimeter-derivedvariability of surface velocities in the Californiacurrent system: seasonal circulation and eddystatistics. Deep-Sea Research 47: 831-870.
STRUB PT & C JAMES (2002a) Altimeter-derived surfacecirculation in the large-scale northeast Pacificgyres: seasonal variabil i ty. Progress inOceanography 53: 163-183.
STRUB PT & C JAMES (2002b) Altimeter-derivedsurface circulation in the large-scale northeastPacific gyres: 1997-1998 El Niño anomalies.Progress in Oceanography 53: 185-214.
STRUB PT, JM MESÍAS & C JAMES (1995) Satelliteobservations of the Peru-Chile countercurrent.Geophysical Research Letters 22: 211-214.
STRUB PT, C JAMES, AC THOMAS & MR ABBOTT(1990) Seasonal and nonseasonal variability ofsatellite-derived surface pigment concentration inthe California current. Journal of GeophysicalResearch 95: 11501-11530.
STRUB PT, PM KOSRO, A HUYER & CTZCOLLABORATORS (1991) The nature of the coldfilaments in the California current system. Journalof Geophysical Research 96: 14743-14768.
STRUB PT, JM MESÍAS, V MONTECINO, JRUTLLANT & S SALINAS (1998) Coastal oceancirculation off western South America. In:Robinson AR & KH Brink (eds) The Sea: 273-314.John Wiley & Sons, Inc, New York, New York,USA.
STULL JK (1995) Two decades of marine biologicalmonitoring, Palos Verdes, California, 1972 to 1992.Bulletin Southern California Academy of Sciences94: 21-45.

409OCEANOGRAPHY, MARINE ECOLOGY, CLIMATE AND HUMANS
TEGNER MJ & PK DAYTON (1981) Populationstructure, recruitment and mortality of two seaurchins (Strongylocentrotus franciscanus and S.purpuratus) in a kelp forest. Marine EcologyProgress Series 5: 255-268.
TEGNER MJ & PK DAYTON (1987) El Niño effects onsouthern California kelp forest communities.Advances in Ecological Research 17: 243-279.
TEGNER MJ, PK DAYTON, PB EDWARDS& KL RISER(1995) Sea urchin cavitat ion of giant kelp(Macrocystis pyrifera C. Agardh) holdfasts and itseffects on kelp mortality across a large Californiaforest. Journal of Experimental Marine Biology andEcology 191: 83-99.
THOMAS AC, F HUANG, PT STRUB & C JAMES (1994)A comparison of the seasonal and interannualvariability of phytoplankton pigment concentrationsin the Peru and California current systems. Journal ofGeophysical Research 99: 7355-7370.
TOMICIC JJ (1985) Efectos del fenómeno El Niño 1982-1983 El Niño las comunidades litorales de laPenínsula de Mejillones. Investigación Pesquera(Chile) 32: 209-213.
TSUCHIYA M (1985) The subthermocline phosphatedistribution and circulation in the far easternequatorial Pacific Ocean. Deep-Sea Research 32:299-313.
UNITED STATES DEPARTMENT OF COMMERCENATIONAL MARINE FISHERIES SERVICE(1998) Endangered and threatened species;threatened status for the Oregon coast evolutionarysignificant unit of coho salmon. United StatesFederal Register 63(153): 42587-42591.
UNITED STATES DEPARTMENT OF COMMERCENATIONAL OCEANIC AND ATMOSPHERICADMINISTRATION (2001) Endangered andthreatened species; endangered status for whiteabalone. United States Federal Register 66(103):29046-29055.
UNITED STATES DEPARTMENT OF THE INTERIORFISH AND WILDLIFE SERVICE (1995)Endangered and threatened species; proposedthreatened status for three contiguous ESUs of cohosalmon ranging from Oregon through CentralCalifornia. United States Federal Register 62:38011-38030.
UNITED STATES DEPARTMENT OF THE INTERIORFISH AND WILDLIFE SERVICE (1996)Endangered and threatened wildlife and plants;listing of the central California coast coho salmonas threatened in California. United States FederalRegister 661: 59028-59029.
UNITED STATES DEPARTMENT OF THE INTERIORFISH AND WILDLIFE SERVICE (1997)Endangered and threatened wildlife and plants;threatened status for the southern Oregon/northern
California coast evolutionary significant unit ofcoho salmon. United States Federal Register 62:33038-33039.
UNITED STATES GLOBAL OCEAN ECOSYSTEMSDYNAMICS (GLOBEC) (1996) United StateGLOBEC northeast Pacific implementation plan.United State GLOBEC Scientif ic SteeringCommittee Coordinating Office, Berkeley,California, USA. 60 pp.
VÁSQUEZ JA & AH BUSCHMANN (1997) Herbivore-kelp interactions in Chilean subtidal communities: areview. Revista Chilena de Historia Natural 70: 41-52.
VEIT RR, P PYLE & JA MCGOWAN (1996) Oceanwarming and long term change in pelagic birdabundance within the California current system.Marine Ecology Progress Series 139: 11-18.
VENRICK IL, JA MCGOWAN, DR CAYAN & TLHAYWARD (1987) Climate and chlorophyll a:long-term trends in the central North Pacific Ocean.Science 238: 70-72.
WARE DM (1995) A century and a half of change in theclimate of the Northeast Pacific. FisheriesOceanography 4: 267-277.
WATANABE JM & C HARROLD (1991) Destructivegrazing by sea urchins Strongylocentrotus spp. in acentral California kelp forest potential roles ofrecruitment, depth, and predation. Marine EcologyProgress Series 71: 125-141.
WICKETT WP (1967) Ekman tanspot and zooplanktonconcentration in the North Pacific. Journal of theFisheries Research Board Canada 24: 581-594.
WIETERS EA, DM KAPLAN, SA NAVARRETE, ASOTOMAYOR, J LARGIER, KJ NIELSEN & FVÉLIZ (2003) Alongshore and temporal variabilityin chlorophyll a concentration in Chilean nearshorewaters. Marine Ecology Progress Series 249: 93-105.
WING SR, JL LARGIER, LW BOTSFORD & JF QUINN(1995) Sett lement and transport of benthicinvertebrates in an intermittent upwelling region.Limnology and Oceanography 40: 316-329.
WOOSTER WS & DL FLUHARTY (1985) El Niño North:Niño effects in the eastern subarctic Pacific Ocean.Universi ty of Washington Press, Seatt le,Washington, USA. 312 pp.
WOOTTON JT (1997) Estimates and tests of per capitainteraction strength: diet, abundance, and impact ofintertidally foraging birds. Ecological Monographs67: 45-64.
WOOTTON JT, ME POWER, RT PAINE & CAPFISTER (1996) Effects of product ivi ty ,consumers, competitors, and El Niño events onfood chain pat terns in a rocky inter t idalcommunity. Proceedings of the National Academyof Sciences of the United States of America 93:13855-13858.
Associate Editor: Pablo Marquet