Embed Size (px)
Citation preview

. .. . . . ~ .. .
Plant Cell Reports (1997) 16: 444-449 0 Springer-Verlag 1997
N. K. Konan - C. Schöpke - R. Cárcamo - R. N. Beachy dpdb C.(Fauquet
An ~~~~~~~~~ mass propagation system for cassava ( ~ a ~ j ~ ~ ~ esculenta Craaatz) based on nodal explants and axilllary bud4erived meristems
Received: 23 September 1996 / Revision received: 12 November 1996 / Accepted: 4 December 1996
Abstract Nodes from 3- to 5-week-old in vitro plants of different cassava cultivars were cultured for 2-3 days on solid Murashige and Skoog basal medium supplemented with cytokinin to induce the enlargement of axillary buds. Subculture of these buds on the same medium resulted in multiple shoot formation within 4-6 weeks. Of the four cy- tokinins tested (6-benzylaminopurine (BAP), thidiazuron (TDZ), zeatin, and kinetin), BAP induced shoot develop- ment most efficiently. The best results were obtained with cultivar TMS 30555, in which 63% of the explants each produced at least 25 shoots on medium with 10 mg/l BAP. In cultivars that did not produce shoots, the addition of the surfactant Pluronic F-68 (2% wt/vol) raised the percentage of explants forming at least 5 shoots from O to 20-60%. Axillary buds were also used to dissect meristems and test their ability to regenerate into shoots. Shoot formation from meristems of six different cultivars was observed af- ter preculture on medium with 5 mgll BAP followed by transfer to 10 mg/l BAP.
Key words Micropropagation. Pluronic F-68 Multiple shoots
Abbreviations MS Murashige and Skoog *
BAP 6-Benzylaminopurine . TDZ Thidiazuron
Introduction
Cassava (Manihot esculentu Crantz) is used in the tropics as a staple food by more than 500 million people (Cock
Communicated by I. K. Vasi1 N. K. Konan . C. Schöpke . R. Cárcamo . R. N. Beachy C. Fauquet (a) International Laboratory for Tropical Agricultural Biotechnology (ILTAB/ORSTOM-TSRI), Division of Plant Biology-BCC206, The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA
1985). However, its production is severely limited by a wide variety of viral and bacterial diseases. Recent ad- vances in genetic engineering of plants hold considerable potential to improve resistance as well as other agronomic characteristics of cassava. A prerequisite for the applica- tion of these methods is the availability of efficient in vi- tro plant regeneration systems for this crop.
Regeneration of cassava, both via organogenesis and somatic embryogenesis, has been reported previously. Shoot formation through organogenesis was achieved from stem callus (Tilquin 1979) and mesophyll protoplast cul- tures (Shahin and Shepard 1980). However, these results were not reproducible. On the other hand, regeneration through somatic embryogenesis is well established and has been used successfully by many workers (Stamp and Hen- shaw 1982, 1987; Szabados et al. 1987; Mathews et al. 1993; Raemakers et al. 1993; Konan et al. 1994a). Re- cently, the development of embryogenic suspension cul- tures was described (Taylor et al. 1996), as was their use for the genetic transformation of cassava and the produc- tion of the first transgenic cassava plants (Schöpke et al. 1996). Although regeneration through embryogenesis is now a standard procedure for some cassava cultivars, it is highly genotype dependent. Alternative regeneration systems might increase the number of cassava cultivars ac- cessible to regeneration and transformation protocols. In cassava this is especially important, since this vegetatively propagated crop is represented by at least five thousand varieties (estimate based on Iwanaga and Iglesias 1994; Ng et al. 1994).
Shoot meristem culture techniques were developed some 20 years ago for cassava (Kartha et al. 1974) and are being used for the production of virus-free plants of elite genotypes (Guo and Liu 1995; Mabanza et al. 1995). A method to produce multiple shoots from axillary buds and from bud-derived meristems has been previously reported for cassava (Konan et al. 1994b). Here we describe an improvement of this culture method, which is applicable to cultivars in which the standard protocol was ineffective.
Fonds Documentaire ORSTOM

Materials and methods
Plant material
The cassava cultivar TMS 30555 was provided by the Institut des Savannes (IDESSA, Ivory Coast). All other cultivars were from the i n vitro collection at the International Laboratory for Tropical Agri- cultural Biology, including Aipin Valenca, MBra 769, CMC 76, MCub 51, MCub 58, MMex 55, MPar 133 from South America, TMS 60444 Red, TMS 30395, TMS 50395, TMS 60142, TMS 83350, TMS 84537, TMS 90059, TMS 90553 from Africa, and Mpira Red from India. The material was maintained by monthly sub- culture as in vitro shoots, cultured on MS medium without growth regulators and solidified with 0.7% Difco Bacto agar. The cultures were kept at 27°C in a 16-hphotoperiod (90-1 10 pmol mb2 s-I; cool- white fluorescent lights Philips F40 CW).
Multiple shoot formation from axillary buds
Nodal explants from in vitro shoot cultures (3-5 weeks old) were cut and placed horizontally on solidified Murashige and Skoog (1962) (MS) medium (0.7% Difco Bacto agar) supplemented with 10, 15 or 20 mg/l of each of BAP, kinetin, zeatin, and TDZ. Culture conditions were the same as for shoot cultures (see above). Enlarged axillary buds that formed after 2-3 days were removed from the nodal ex- plants with a scalpel and were subcultured on the same medium as before to induce shoot formation. This protocol was applied to 16 different cassava cultivars (see above; 20-40 explants per cultivar), using the conditions that gave the best results with cv. TMS 30555, i.e., 10 mg/l BAP both in the medium for the culture of nodal ex- plants and for shoot development from axillary buds.
Effect of surfactants on shoot regeneration
The surfactants Tween 20, Triton X-100, and Pluronic F-68 (all from Sigma, St. Louis, Mo.) were added at different concentrations to shoot induction medium containing 10 mg/l BAP. Pluronic F-68 was added before, and Tween 20 and Triton after autoclaving of the cul- ture medium. Axillary buds from nodal explants of cv. TMS 60444 Red that had been cultured for 3 days on medium with 10 mg/l BAP were transferred for 2 weeks to medium with 10 mg/l BAP contain- ing a surfactant, and then subcultured on the same medium without surfactant. Using the treatment that was found to be optimal for the induction of shoot development (Pluronic F-68 at 2.0%), the experi- ment was repeated with three different cultivars, CMC 76, MPira Red, and MCub 58, using TMS 60444 Red as a control.
Shoot regeneration from meristems of axillary buds
Nodal explants were cultured on MS medium containing 10 mg/l BAP. Meristems from axillary buds were dissected after 4-6 days and transferred to different culture media. These media were com- posed of various basal salt mixtures and supplemented with MS vi- tamins, 10 mg/l BAP, and 2% (wthol) sucrose. In another experi- ment, meristems were transferred to MS medium with 1,5, or 10 mg/l BAP. The viability of explants 2 weeks after culture initiation was assessed visually. Explants that were green and had increased in size were considered viable.
Plant regeneration
Shoots 1-2 cm long that had been regenerated from axillary buds or from axillary-bud-derived meristems were transferred to hormone- free MS medium supplemented with 3% sucrose. Once the shoots had reached 5-6 cm and had a well-developed root system, they were
445
potted in a mixture of 50% vermiculite and 50% sand and transferred to a greenhouse. During the, first 2 weeks they were covered loose- ly with plastic foil to increase humidity.
Histology
Explants were fixed for 16-24 h in Histochoice (Amresco, Solon, Ohio) before dehydration in increasing concentrations of ethanol (30, 50,70,90,95 and loo%, vol/vol). Tissues were embedded using the JB-4 Plus Embedding Kit (Polysciences, Warrington, Penn.). Sec- tions of 7 pm were cut and stained essentially as described in Schnei- der (1981). Sections were incubated for 20 min in a solution of 0.5% (wthol) periodic acid, followed by 20 min in Schiff's reagent. Af- ter rinsing, sections were placed for 1-2 min in a solution of 2% (wt/vol) sodium bisulfite, washed in running water for 5-10 min and transferred for 1-2 min to a solution containing 1% Amido Black 10B (no. N 3005, Sigma) in 7% acetic acid. As a result of this stain- ing, polysaccharides appear red, and proteins bluish black. Stained sections were embedded in Permount (Fisher Scientific, USA) prior to microscopy.
Results and discussion
Multiple shoot production from axillary buds
In nodal explants cultured on solidified MS medium con- taining cytokinin, the axillary buds enlarged to a diameter of 1-2 mm within 2-3 days (Fig. IA). BAP, TDZ, zeatin, and kinetin at concentrations of 10 and 15 mg/l stimulated the formation of these enlarged axillary buds at frequen- cies ranging from 70 to 90% for the cultivar TMS 30555. Concentrations of 20 mg/l cytokinin caused the nodal ex- plants to turn brown.
Enlarged axillary buds initiated on nodal explants of cassava cultivar TMS 30555 were subcultured on MS re- generation medium containing 10 mg/l cytokinin. Two days after the transfer, the explants started to expand, the covering leaflets opened, and the shoot meristems were ex- posed (Fig. 1B). Within the 2nd week of culture, secon- dary meristems (Fig. IC) arose in the form of protuber- ances. These protuberances could also appear on the sur- face of the expanded primary meristem. Some 4-6 weeks after culture initiation, clumps of microshoots (Fig. ID) developed.
The effect of four different cytokinins on the formation of multiple shoots from enlarged axillary buds of cassava cultivar TMS 30555 is summarized in Table 1. Variable rates of shoot proliferation were observed 5-6 weeks after culture initiation. The highest number of shoots per explant was obtained with BAP. At 10 mg/l BAP, the percentage of explants forming shoots was about seven times higher than with 15 mg/l BAP (73% vs. 9%). With 10 mg/l TDZ, the number of explants producing shoots was higher than with BAP (87%), but the average number of shoots per ex- plant was low. In addition, TDZ led to the development of hyperhydric shoots. Both kinetin and zeatin were less ef- ficient with regard to the total number of shoots produced compared to BAP and TDZ.
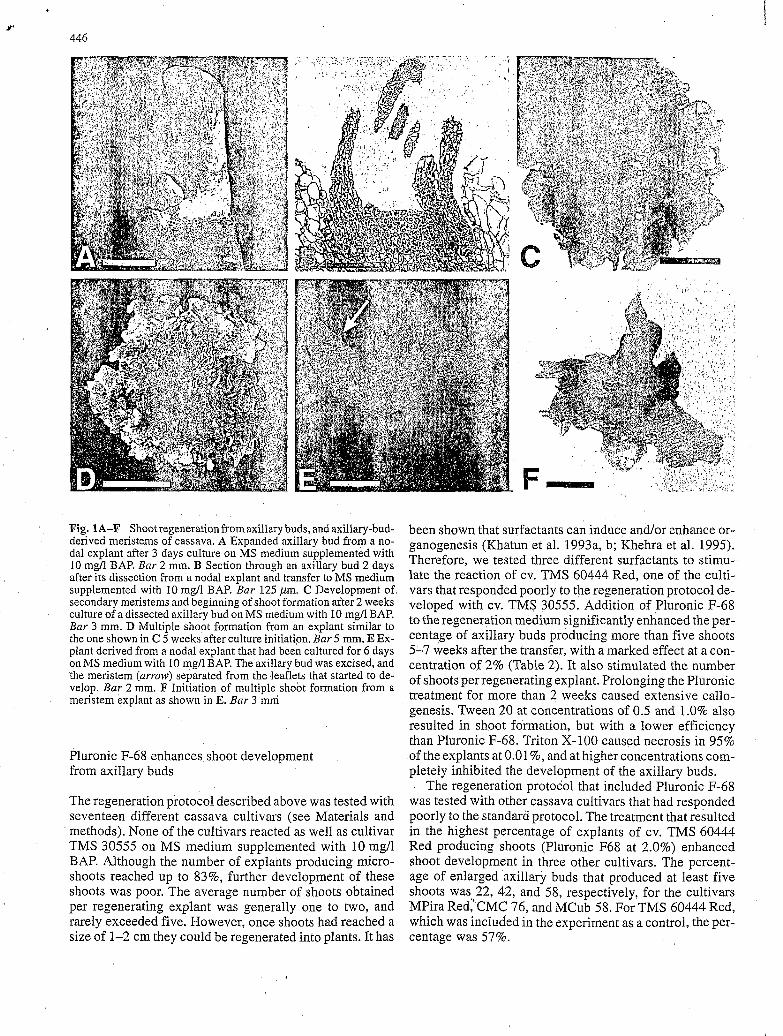
Fig. 1A-F Shoot regeneration from axillary buds, and axillary-bud- derived meristems of cassava. A Expanded axillary bud from a no- dal explant after 3 days culture on MS medium supplemented with 10 mgfi BAP. Bar 2 mm. B Section through an axillary bud 2 days after its dissection from a nodal explant and transfer to MS medium supplemented with 10 mgll BAP. Bar 125 ,um. C Development of secondary meristems and beginning of shoot formation after2 weeks culture of a dissected axillary bud on MS medium with 10 mg/l BAP. Bar 3 mm. D Multiple shoot formation from an explant similar to the one shown in C 5 weeks after culture initiation. Bar 5 mm. E Ex- plant derived from a nodal explant that had been cultured for 6 days on MS medium with 10 mg/l BAP. The axillary bud was excised, and the meristem (arrow) separated from the leaflets that started to de- velop. Bar 2 mm. F Initiation of multiple shoot formation from a meristem explant as shown in E. Bar 3 mm
Pluronic F-68 enhances shoot development from axillary buds
The regeneration protocol described above was tested with seventeen different cassava cultivars (see Materials and methods). None of the cultivars reacted as well as cultivar TMS 30555 on MS medium supplemented with 10 mg/l BAP. Although the number of explants producing micro- shoots reached up to 83%, further development of these shoots was poor. The average number of shoots obtained per regenerating explant was generally one to two, and rarely exceeded five. However, once shoots had reached a size of 1-2 cm they could be regenerated into plants. It has
been shown that surfactants can induce and/or enhance or- ganogenesis (Khatun et al. 1993~1, b; Khehra et al. 1995). Therefore, we tested three different surfactants to stimu- late the reaction of cv. TMS 60444 Red, one of the culti- vars that responded poorly to the regeneration protocol de- veloped with cv. TMS 30555. Addition of Pluronic F-68 to the regeneration medium significantly enhanced the per- centage of axillary buds producing more than five shoots 5-7 weeks after the transfer, with a marked effect at a con- centration of 2% (Table 2). It also stimulated the number of shoots per regenerating explant. Prolonging the Pluronic treatment for more than 2 weeks caused extensive callo- genesis. Tween 20 at concentrations of 0.5 and 1 .O% also resulted in shoot formation, but with a lower efficiency than Pluronic F-68. Triton X-100 caused necrosis in 95% of the explants at 0.0 1 %, and at higher concentrations com- pletely inhibited the development of the axillary buds.
The regeneration protocol that included Pluronic F-68 was tested with other cassava cultivars that had responded poorly to the standard protocol. The treatment that resulted in the highest percentage of explants of cv. TMS 60444 Red producing shoots (Pluronic F68 at 2.0%) enhanced shoot development in three other cultivars. The percent- age of enlarged axillary buds that produced at least five shoots was 22, 42, and 58, respectively, for the cultivars MPira Red,'CMC 76, and MCub 58. For TMS 60444 Red, which was included in the experiment as a control, the per- centage was 57%.
447
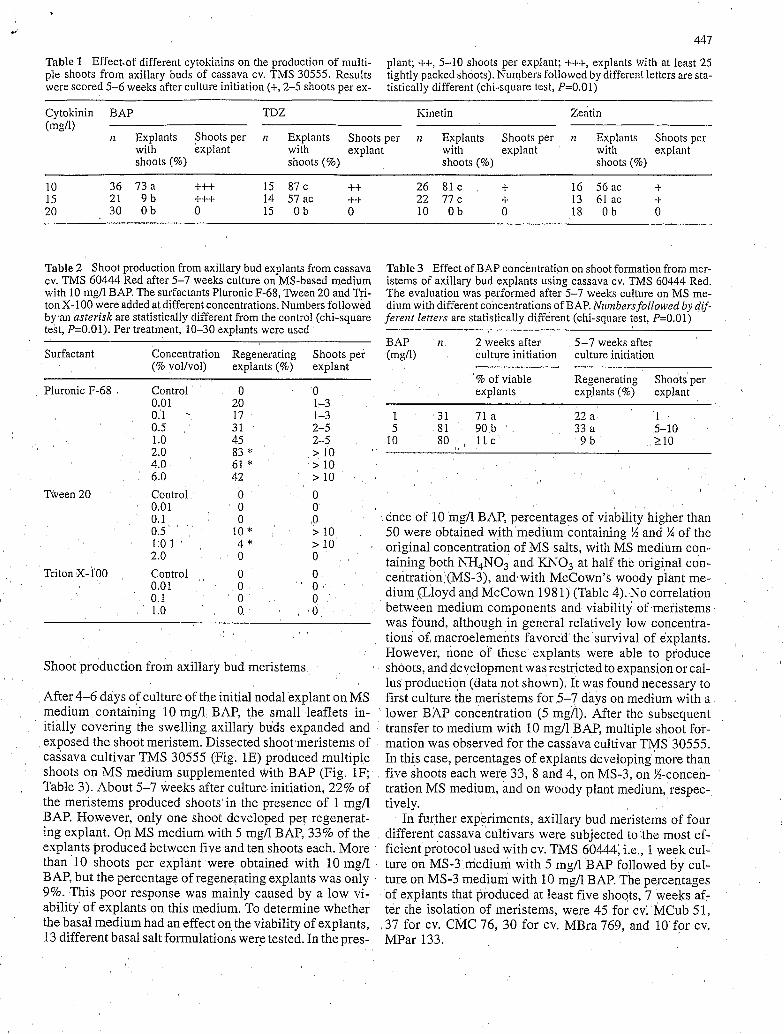
plant; ++, 5-10 shoots per explant; +++, explants with at least 25 tightly packed shoots). Numbers followed by different letters are sta- tistically different (chi-square test, P=O.Ol)
Table 1 Effect.of different cytokinins on the production of multi- ple shoots from axillary buds of cassava CY. TMS 30555. Results were scored 5-6 weeks after culture initiation (+, 2-5 shoots per ex-
~ ~~~ ~
~
Cytokinin BAP TDZ Kinetin Zeatin (mgll)
I I Explants Shoots per n Explants Shoots per 12 Explants Shoots per I I Explants Shoots per with explant with explant with explant with explant shoots (%) shoots (%) shoots (%) shoots (%)
10 36 7 3 a t++ 15 8 7 c ++ 26 81 c + 16 56 iic t 15 21 9 b +t+ 14 57ac ++ 22 7 7 c + 13 61 ac t 20 30 Ob O 15 Ob O 10 Ob O 18 Ob O
Table 2 Shoot production from axillary bud explants from cassava cv. TMS 60444 Red after 5-7 weeks culture on MS-based medium with 10 mg/l BAP. The surfactants P h o n i c F-68, Tween 20 and Tri- ton X-100 were added at different concentrations. Numbers followed by an asterisk are statistically different from the control (chi-square test, P=O.Ol). Per treatment, 10-30 explants were used
Surfactant Concentration Regenerating Shoots per
P h o n i c F-68 Control O O
(% vol/vol) explants (%) explant
0.01 20 1-3 0.1 - 17 1-3 0.5 31 2-5 1 .o 45 2-5 2.0 83 * > 10 4.0 61 * > 10 6.0 42 > 10
Tween 20 Control O O
Table 3 Effect of BAP concentration on shoot formation from mer- istems of axillary bud explants using cassava cv. TMS 60444 Red. The evaluation was performed after 5-7 weeks culture on MS me- dium with different concentrations of BAP. Nciinbers followed by dif- ferent letters are statistically different (chi-square test, P=O.Ol)
BAP it 2 weeks after 5-7 weeks after ( m m culture initiation culture initiation
% of viable Regenerating Shoots per explants explants (%) explant
1 31 71 a 22 a 1 5 81 9 0 b 33 a 5-10
10 80 l l c 9 b 2 10
I
0.01 O O o. 1 O O ence of 10 mg/l BAP, percentages of viability higher than 0.5 50 were obtained with medium containing !4 and K of the 1.0 1 4 " > 10 original concentration of MS salts, with MS medium con- 2.0 O O taining both NH,NO, and KNO, at half the original con-
Triton X-100 Control O O centration (MS-3), and with McCown's woody plant me- 0.0 1 O O dium (Lloyd and McCown 1981) (Table 4). No correlation o. 1 O O between medium components and viability of meristems 1 .o O , o
was found, although in general relatively low concentra- tions of, macroelements favored the survival of explants. However, none of these explants were able to produce shoots, and development was restricted to expansion or cal- lus production (data not shown). It was found necessary to first culture the meristems for 5-7 days on medium with a lower BAP concentration (5 mg/l). After the subsequent transfer to medium with 10 mg/l BAP, multiple shoot for- mation was observed for the cassava cultivar TMS 30555. In this case, percentages of explants developing more than five shoots each were 33, 8 and 4, on MS-3, on !4-concen- tration MS medium, and on woody plant medium, respec- tively.
In further experiments, axillary bud meristems of four different cassava cultivars were subjected to the most ef- ficient protocol used with cv. TMS 60444, i.e., 1 week cul- ture on MS-3 medium with 5 mg/l BAP followed by cul- ture on MS-3 medium with 10 mg/l BAP. The percentages of explants that produced at least five shoots, 7 weeks af- ter the isolation of meristems, were 45 for cv. MCub 51, 37 for cv. CMC 76, 30 for cv. MBra 769, and 10 for cv. MPar 133.
10 * > 10
Shoot production from axillary bud meristems
After 4-6 days of culture of the initial nodal explant on MS medium containing 10 mg/l BAP, the small leaflets in- itially covering the swelling axillary buds expanded and exposed the shoot meristem. Dissected shoot meristems of cassava cultivar TMS 30555 (Fig. 1E) produced multiple shoots on MS medium supplemented with BAP (Fig. 1F; Table 3). About 5-7 weeks after culture initiation, 22% of the meristems produced shoots in the presence of 1 mg/l BAP. However, only one shoot developed per regenerat- ing explant. On MS medium with 5 mg/l BAP, 33% of the explants produced between five and ten shoots each. More than 10 shoots per explant were obtained with 10 mg/l BAP, but the percentage of regenerating explants was only 9%. This poor response was mainly caused by a low vi- ability of explants on this medium. To determine whether the basal medium had an effect on the viability of explants, 13 different basal salt formulations were tested. In the pres-

448
Table 4 The effect of different basal salt mixtures 011 the viability of meristems excised from expanded axillary buds of cassava cv. TMS 30555. For each treatment, 20-40 explants were placed on cul- ture medium containing different basal salt mixtures and 10 mg/l BAP. After 2 weeks, viability was assessed. An explant was scored as viable when it had stayed green and expanded in size
Viability Macroelements of culture medium Ionic concen- (%I tration of
macroelemen ts ( M O
O 5 6
12 14 14 20 31 33 56 59 72
74
De Greef and Jacobs (1979) 82.23 GresshofF and Doy (1974) 54.92 MS; NH4N03.replaced by NaN03 95.75 Schenk and Hildebrandt (1972) 67.17 Gamborg et al. (1968) 61.89 MS 95.75 Kao and Michayluk (1975) 80.25 MS; X concentration 71.81 MS; without NH4N0, 54.53 MS; !4 concentration 47.87 MS; !4 concentreation 23.94 MS; NH4N03 and KNO, at 56.53 !4 concentration Lloyd and McCown (1981) 44.01
Plant regeneration
Once shoots had reached a size of 1-2 cm, they were trans- ferred to hormone-free MS medium supplemented with 3 % sucrose. Independent of the type of cytokinin with which they had been cultured, and regardless of their origin (ax- illary buds or axillary-bud-derived meristems), all shoots developed into normal pla,nts with roots after a further transfer to the same medium. The survival rate of plants transferred to the greenhouse was in the range of 90-100%. No apparent differences between regenerants and mother plants were observed.
Conclusions
Cassava is a highly heterozygous crop that is propagated almost exclusively by stem cuttings. From a mature cas- sava plant, 10-30 cuttings can be obtained per year. Thus the propagation rate is a limiting factor in the distribution of planting material, especially of improved cultivars for which there is a high demand. Although under experimen- tal conditions much higher propagation rates can be achieved (Kamalan et al. 1977: 323; Pateña and Barba 1979: 8400), for practical purposes a substantial improve- ment is desirable. In vitro culture has the potential to in- crease the propagation rate by orders of magnitude. Through shoot culture, one nodal explant can theoretically produce one million plantlets with four nodes each in 1 year. With our new system, this number could be further increased by at least a factor of ten.
The use of genetic engineering for the improvement of cassava requires efficient regeneration and transformation systems. Recently, genetic transformation of cassava was achieved both by microparticle bombardment (Schöpke et
al. 1996) and by Agrobnctkriuin (Li et al. 1996). In both cases, the tissue used for transformation was derived from somatic embryos. However, somatic embryogenesis in cas- sava is highly genotype dependent. As an example, two cassava cultivars, Clone 31 (Guyana) and TMS 30001 (Ni- geria), have thus far been shown to be recalcitrant to re- peated efforts to induce somatic embryogenesis (N. Tay- lor, personal communication). The regeneration system de- scribed here provides an alternative that could be used in cases where somatic embryogenesis as a regeneration system is not available or is not efficient enough for the purpose of genetic transformation. The meristematic struc- tures that develop 2 weeks after the transfer of expanded axillary buds from cassava shoots to culture medium with BAP (Fig. IC) could serve as target tissue both for trans- formation through microparticle bombardment and through Agrobncterium.
‘
References
Cock JH (1985) Cassava: new potential for a neglected crop. West- view Press, Boulder, Colo.
De Greef W, Jacobs M (1979) In vitro culture of sugarbeet: descrip- tion of a cell line with high regeneration capacity. Plant Sci Lett
Gamborg OL, Miller RA, Ojima K (1968) Nutritional requirements of suspension cultures of soybean root cells. Exp Cell Res
Gresshoff PM, Doy CH (1974) Derivation of a haploid cell line from Wtis vinifera and importance of the stage of meiotic development of anthers for haploid culture of this and other genera. Z Pflan- zenphysiol73:132-141
Guo YJ, Liu YQ (1995) Rapid propagation of cassava by tissue cul- ture and its application in rural districts in China. In: The cassa- va biotechnology network. Proceedings of the Second Interna- tional Scientific Meeting, Bogor, Indonesia, 22-26 August 1994. CIAT, Cali, vol 1, pp 183-189
Iwanaga M, Iglesias C (1994) Cassava genetic resources manage- ment at CIAT. In: Report of the First Meeting of the Internation- al Network for Cassava Genetic Resources, CIAT, Cali, Colom- bia, 18-23 August 1992. International Crop Network Series no. 10. IPGRI, Rome, pp 77-86
Kamalan P, Rajendran PG, Hrishi N (1977) A new technique for the rapid propagation of cassava (Manihot esculenta Crantz). Trop Agric (Trinidad) 54:213-217
Kao KN, Michayluk MR (1975) Nutritional requirements for growth of Vicia hajastana cells and protoplasts at a very low population density in liquid media. Planta 126: 105-1 10
Kartha KK, Gamborg OL, Constabel F, Shyluk JP (1974) Regener- ation of cassava plants from apical meristems. Plant Sci Lett
Khatun A, Davey MR, Power JB, Lowe KC (1993a) Stimulation of shoot regeneration from cotyledons cultured with non-ionic sur- factants and relationship to physico-chemical properties. Plant Cell Rep 13:49-53
Khatun A, Laouar L, Davey MR, Power JB, Mulligan JB, Lowe KC (1993b) Effects of pluronic F-68 on shoot regeneration from cul- tured jute cotyledons and on growth of transformed roots. Plant Cefi Tissue Organ Cult 3 4 133-140
Khehra M, Lowe KC, Davey MR, Power JB (1995) An improved micropropagation system for Clzrysantkemum based on Pluron- ic F-68-supplemented media. Plant Cell Tissue Organ Cult 41:87-90
KonanNK, SangwanRS, Sangwan-Norreel BS (19944 Somatic em- bryogenesis from cultured mature cotyledons of cassava (Mani- hot esculenta Crantz) - identification of parameters influencing
17 ~5.5-6 1
50:151-158
2: 107-1 13
3

449
the frequency of embryogenesis. Plant Cell Tissue Organ Cult
Konan NK, Sangwan RS, Sangwan-Norreel BS (1994b) Efficient in vitro shoot regeneration systems in cassava (Manihot escidenia
Lloyd G, McCown B (198 1) Commercially-feasible micropropaga- tion of mountain laurel, Kcilniia latifolia, by use of shoot-tip cul- ture. Int Plant Prop Soc Proc 30:421-427
Li H-Q, Sautter C, Potrykus I, Puonti-Kaerlas J (1996) Genetic trans- formation of cassava (Manihot escidenfa Crantz). Nat Biotech- no1 14:736-740
Mabanza J, Rodriguez-Andriyaniasi AV, Mahouka J, Boumba B (1995) Evaluation of cleaned cassava varieties in Congo. In: The cassava biotechnology network. Proceedings of the Second Inter- national Scientific Meeting, Bogor, Indonesia, 22-26 August 1994. CIAT, Cali, vol 1, pp 194-201
Mathews H, Schöpke C, CBrcamo R, Chavarriaga P, Fauquet C, Beachy RN (1993) Improvement of somatic embryogenesis and plant recovery in cassava. Plant Cell Rep 12:328-333
Murashige T, Skoog F (1962) A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant
Ng NQ, Asiedu R, Ng SYC (1994) Cassava genetic resources pro- gramme at the International Institute of Tropical Agriculture, Ib- adan. In: Report of the First Meeting of the International Net- work for Cassava Genetic Resources, CIAT, Cali, 18-23 August 1992. International Crop Network Series no. 10. IPGRI, Rome,
Pateiia LF, Barba RC (1979) Rapid propagation of cassava by leaf-
37:91-102
. Crantz). Plant Breed 113:227-236
15~473-497
pp 71-76 ’
bud cuttings. Philipp J Crop Sci 453-62
Raemakers CJJM, Bessembjnder J, Staritsky G, Jacobsen E, Visser RGF (1993) Induction, germination and shoot development of somatic embryos in cassava. Plant Cell Tissue Organ Cult
Schenk RU, Hildebrandt AC (1972) Medium and techniques for in- duction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can J Bot 50:199-204
Schneider H (1981) Plant anatomy and general botany. In: Clark C (ed) Staining procedures, 4th edn. Williams gL Wilkins, Balti- more, pp 323-324
Schöpke C, Taylor N, Cárcamo R, Konan NK, Marmey P, Henshaw GG, Beachy RN, Fauquet C (1996) Regeneration of transgenic cassava plants (Manihot esculenta Crantz) from microbombard- ed embryogenic suspension cultures. Nat Biotechnol14:73 1-735
Shahin EA, Shepard JF (1980) Cassava mesophyll protoplasts: iso- lation, proliferation and shoot formation. Plant Sci Lett 17:459-465
Stamp JA, Henshaw GG (1982) Somatic embryogenesis in cassava. Z Pflanzenphysiol105:97-102
Stamp JA, Henshaw GG (1987) Secondary embryogenesis and plant regeneration in cassava. Plant Cell Tissue Organ Cult 10:227-233
Szabados L, Hoyos R, Roca W (1987) In vitro somatic embryogen- esis and plant regeneration of cassava. Plant Cell Rep 6:248-251
Taylor NJ, Edwards M, Kiernan RJ, Davey C, Blakesley D, Henshaw GG (1995) Development of friable embryogenic callus and em- bryogenic suspension culture systems in cassava (Manilzof escu- lenta Crantz). Nat Biotechnol 14:726-730
Tilquin JP (1979) Plant regeneration from stem callus of cassava. Can J Bot 57:1761-1763
33:151-156





























