Embed Size (px)
Citation preview

An International Journal of Marine Sciences
Number 28 (2) July 2012
Thalassas


ThalassasThalassas greek voice meaning...”of the sea”

THALASSAS is included in the following DATABASES:
THE BOWKER INTERNATIONAL SERIALS DATABASE (Ulrich’s International Periodicals Directory). USA.
ÍNDICE ESPAÑOL DE CIENCIA Y TECNOLOGÍA (I.C.Y.T.). SPAIN
FAO: FISHERY INFORMATION. DATA AND STATISTICS SERVICE ITALY
MS. MEDIA SERVICE GMBH. GERMANY
CATÁLOGO CSIC, SPAIN
LATINDEX, MÉXICO
SCOPUS
THOMSON REUTERS MASTER JOURNAL LIST
JOURNAL CITATION REPORTS: THOMSON-REUTERS WEB OF KNOWLEDGE
DIALNET
GEOREF
SCIENCE CITATION INDEX EXPANDED
ZOOLOGICAL RECORD
WEB PAGE: http://webs.uvigo.es/thalassas/Electronic submission of Manuscripts: http://recyt.fecyt.es/index.php/Thal
© Universidade de Vigo, 2012
Impresa: en Vigo. España
Edita: Servizo de Publicacións Universidade de Vigo. Campus das Lagoas, Marcosende 36310 Vigo. España.
I.S.S.N.: 0212-5919Dep. Leg.: C379-83Nº 28 (2) - 2012
Cover photograph: Effect of desalination plants on the marine environment along the Red Sea, Egypt. (Case Study)

Volume 28(2)
THALASSASAN INTERNATIONAL JOURNAL OF MARINE SCIENCES


EDITORIAL BOARD
Editor-in-Chief
Scientific Committee
MANUEL J. REIGOSA ROGERDepartament of Plant Biology and Soil Science
University of Vigo, Spain
ALFREDO ARCHE MIRALLESInstituto de Geología Económica.C.S.I.C., Madrid, Spain
ANTONIO CENDRERO UCEDAD.C.I.T.T.Y.M. Facultad de Ciencias.University of Cantabria, Santander, Spain
CARLOS SOUTOFaculty of Marine SciencesUniversity of Vigo, Spain
CÁSTOR GUISANDEFaculty of Marine SciencesUniversity of Vigo, Spain
DANIEL REYFaculty of Marine SciencesUniversity of Vigo, Spain
FEDERICO ISLACentro de Geología de CostasUniversity of Mar del Plata, Argentina
FEDERICO VILAS MARTÍNFaculty of Marine SciencesUniversity of Vigo, Spain
FRANCISCO RAMILFaculty of Marine SciencesUniversity of Vigo, Spain
GUILLERMO FRANCÉSFaculty of Marine SciencesUniversity of Vigo, Spain
GABRIEL ROSÓNFaculty of Marine SciencesUniversity of Vigo, Spain
JESÚS SOUZA TRONCOSOFaculty of Marine SciencesUniversity of Vigo, Spain
JOHN L. LARGIERBodega Marine LaboratoryDepartment of EnvironmentalScience and Policy,University of California, Davis
LUÍS GONZÁLEZDepartament of PlantBiology and SoilScienceUniversity of Vigo, Spain
LUISA ANDRADEDepartament of PlantBiology and SoilScienceUniversity of Vigo, Spain
MIGUEL Á. NOMBELAFaculty of Marine SciencesUniversity of Vigo, Spain
M. RUFUS KITTOMarine BiologyDepartment, Faculty of Marine SciencesKing AbdulazizUniversity, Jeddah, Saudi Arabia
NORBERT P. PSUTYCenter forCoastal and EnvironmentalStudiesUniversity of New Jersey, USA
TAKESHI YASUMOTODepartment of Chemistry, AgriculturalFaculty,University of Tohoku, Japan
TOMOHIRO KAWAGUCHIDepartment of EnvironmentalHealthSciencesThe Norman J. ArnoldSchool of PublicHealthUniversity of South Carolina, USA

Papers should be submitted using the web-based application. If you want to add some materials (software, extended maps, additional material) please contact [email protected] for instructions. The application is accessed from the Journal web page (http://webs.uvigo.es/thalassas). There you should select “Send an article” and follow the instructions. Alternatively, you can directly access the application in the following address: http://recyt.fecyt.es/index.php/Thal
Thalassas publishes papers related to all fields of marine sciences. Bothregular papers, short notes and review papers are accepted. You can also contact previously with the Editor-in-Chief (Manuel Reigosa, [email protected]) this is especially encouraged before submitting review papers or letters.
ArticlesThalassas is an international journal that accepts original papers, review papers and short notes about every aspect of marine sciences,especially when a multidisciplinary approach is followed. Language accepted is English. The journal will provide also a summary in Spanish.Authors are allowed to post their accepted papers in their own Web pages. Thalassas will, in any case, provide free to all the scientificcommunity, a version of the published papers to download from the Thalassas Web page.Revision of papers will be done using electronic facilities (that is, referees would receive by e-mail the papers under revision and should answerno later than two weeks after receiving the article by e-mail or fax).Authors can apply for a picture or graphics to be used as a full colour cover image for the paper version. Please state it when you submit your paper.
Full-length papers:Those are original previously unpublished works about any aspect of marine sciences. The title should be indicative of contents, and no longer than 60characters. The first page should include the names of authors and complete affiliations, including e-mail addresses and Web page addressesif any. They will include an abstract (100 - 300 words), followed by less than 15 keywords (both included in the abstract andadditional). Materials and Methods sections will be followed by Results and Discussion. Those sections can be put together if this fits the contentof the Manuscript. Manuscripts should be written in simple sentences, conforming to accepted Scientific Standard English. Texts should be clearconsidering the great scope of the audience (this is not a very specialised journal, covering a broad range of disciplines, although always relatedto marine or coastal ecosystems).
After those sections, Acknowledgements and References should follow. The style of citation will be as follows:
Journal articles:Author AA, Author BB (year). Title of article, Complete name of Journal, number: pages
Book articles:Author AA, Author BB (year). Title of article. In: A Editor, B Editor, eds, Title of Book, Ed, number, Vol number, Publisher, City, pages.Theses Author AA (year) Title of thesis.University, City.The citations should be arranged in the text from earliest to most recent year, alphabetised by name within the same year. In the references list,order by author (s) name, after by year.Finally, figures, tables and captions for figures and tables should be included.
Review papers:Those papers will be published mainly by invitation. But suggestions are also welcome. If you feel that you can contribute with a review, pleasecontact Editor-in-Chief by e-mail.
Technical papers:These papers are especially welcome for the electronic version, but if the editors appreciate their interest, they can also be published in the paperversion. The structure of the article should follow the same recommendations as full-length papers.
Letters:Correspondence prepared for publication in the paper version should not exceed two printed pages. For the electronic version (that will be thepreferred for letters because the speed of publication and the possibility of several responses) no page limit is applied, although the shorter thebest.
Meeting reports and Conference Proceedings:For meeting reports of Conferences about Marine Sciences, the coordinator of the Congress who wishes an abstract to appear in Thalassas (eitherin electronic or paper versions) should contact Editor-in-Chief ([email protected]). Congress Proceedings could also be published as specialnumbers of the journal.
INSTRUCTIONS FOR THE AUTHORS

7Thalassas, 28(2) · July 2012
INDEX
Aseer Manilal, Sugathan Sujith, Joseph Selvin, Mamkoottahil Velayudhan Nataraja Panikkar& Shiney GeorgeAnticoagulant potential of polysaccharide isolated from the Indian red alga, Asparagopsis taxiformis(Delile) Trevisan.
Tarek Abdel-Aziz A. MohammedCoral reefs quantitatively assessment along the egyptian Red Sea Coast.
Mohamed Abd El Wahab & Amr Zakria HamodaEffect of desalination plants on the marine environment along the Red Sea, Egypt. (Case Study)
V. Rajasree, S. Satheesh & S.G. Prakash VincentAntifouling activity of seaweed associated bacterium.
Leandro Fainburg, María Trassens, Julián Bastida, Marcelo Farenga, Federico Isla & Ricardo BastidaNearshore benthic communities and bioengineers from the macrotidal San Jorge Gulf: Patagonia, Argentina.
Craig W. Steele, Patrick R. Pontzer & John AntonelliDistribution of inland silversides Menidia beryllina (COPE) as a function of wave crest velocity at Pelican Island, Chincoteague Inlent, Virginia, USA.
9-15
17-26
27-36
37-43
45-56
57-63
Cover Photograph:Effect of desalination plants on the marine
environment along the Red Sea, Egypt. (Case Study).


ANTICOAGULANT POTENTIAL OF POLYSACCHARIDE ISOLATED FROM THE INDIAN RED ALGA, Asparagopsis taxiformis (Delile) Trevisan
*Dr. Aseer Manilal (corresponding author)Department of Biotechnology, Presentation College of Applied Sciences, Puthenvelikara 683594, South India
Email: [email protected] - Alternate email: [email protected]
(1,5)Department of Biotechnology, Presentation College of Applied Sciences, Puthenvelikara 683594, South India (1,2,3) Department of Bioinformatics, Bharathidasan University, Tiruchirappalli 620 024, South India(4) Department of Botany and Biotechnology, Sree Narayana College, Kollam 691001, South India.
Thalassas, 28(2) · July 2012: 9-15An International Journal of Marine Sciences
Key words: Anticoagulant, Asparagopsis taxiformis, red algae, seaweeds, sulfated polysaccharide
Palabras clave: Anticoagulante, Asparagopsis taxiformis, alga roja, alga, polisacárico sulfatado
ASEER MANILAL(*1), SUGATHAN SUJITH(2), JOSEPH SELVIN(3),MAMKOOTTAHIL VELAYUDHAN NATARAJA PANIKKAR(4) & SHINEY GEORGE(5)
9Thalassas, 28(2) · July 2012
ABSTRACT
A sulfated polysaccharide with anticoagulant activity was purified from the red alga Asparagopsis taxiformis (Delile) Trevisan in Bonnemaisoniaceae family. The molecular mass of the purified polysaccharide ranged between 60 and 500 KD. The anticoagulant activity of the purified compound was assayed using APTT (Activated partial thromboplatin time) and PT (Prothrombin time) assays. Commercial heparin was used as standard to determine the relative potency of the sulfated polysaccharide isolated from A. taxiformis. The purified polysaccharide had a relative clotting factor of 28.57 (>1000s ± 3.6) at 48 μg/mL, the value was comparable with heparin at 60 μg/mL. Statistical analysis revealed that there was no significant difference (p>0.05) between the anticoagulant activity of the purified polysaccharide and commercial heparin. Based on the present findings, it was envisaged that the polysaccharide from A. taxiformis might be capable of inhibiting both intrinsic and extrinsic pathways of blood coagulation. The relative clotting factor assayed by PT was within the range of the oral anticoagulant agent. Therefore, the sulfated polysaccharide of A. taxiformis could be developed as a potential oral anticoagulant agent.
RESUMEN (Potencial anticoagulante de un polisacárido aislado a partir el alga roja de la India Asparagopsis taxifor-mis (Delile) Trevisan)
Un polisacárido sulfatado con actividad anticoagulante fue purificado a partir del alga roja Asparagopsistaxiformis (Delile) Trevisan perteneciente a la familia Bonnemaisoniaceae. La masa molecular del polisacárido purificado osciló entre 60 y 500 KD. La actividad anticoagulante del compuesto purificado se ensayó midiendo APTT (tiempo de tromboplastina parcial activado) y PT (tiempo de protrombina). Se utilizó heparina comercial como estándar para determinar la potencia relativa del polisacárido sulfatado aislado de A taxiformis. El polisacárido purificado tuvo un factor de coagulación relativo de 28,57 (> 1000 ± 3,6) a 48 mg / mL, siendo el valor comparable con la heparina comercial a 60 mg / ml. El análisis estadístico reveló que no hubo diferencia significativa (p> 0,05) entre la actividad anticoagulante de la heparina comercial y el polisacárido purifi-cado. Basándose en los resultados encontrados, se propone que el polisacárido de A. taxiformis podría ser capaz de inhibir las rutas de coagulación sanguínea tanto intrínsecas como extrínsecas. El factor de coagulación relativa ensayado mediante PT estuvo dentro del rango del agente anticoagulante oral. Por lo tanto, el polisacárido sulfatado de A. taxiformis podría ser desarrollado como un agente anticoagulante oral potencial.
ASEER MANILAL, SUGATHAN SUJITH, JOSEPH SELVIN, MAMKOOTTAHIL VELAYUDHAN NATARAJA PANIKKAR & SHINEY GEORGE
INTRODUCTION
The disorders caused by the cardiovascular system have been identified and reported as a major cause of death. Heparin, a commercial anticoagulant was used for more than 50 years. Recently, several side effects of heparin have been identified such as the development of thrombocytopenia, hemorrhagic effect, ineffectiveness in congenital or acquired antithrombin deficiencies, ineffectiveness towards thrombin bound to fibrin, and more (Zoysa et al., 2007). Furthermore, heparin is available in very low concentrations in porcine intestine or bovine lungs from where it was primarily extracted (Pereira et al., 2005). Therefore, search for alternate sources of anticoagulants has been increased with increasing demand for a safe anticoagulant therapy. The search for bioactive compounds from natural resources especially from marine organisms has been steadily increasing during the past few years. The sulfated polysaccharides from marine sources received greater attention as bioactive compounds. The sulfated polysaccharides occur in a wide variety of marine organisms including marine algae, sea urchins etc. In marine algae, they occur as sulfated fructose and sulfated galactans (Painter, 1983). In recent years, sulfated polysaccharides from marine algae have been demonstrated to have many biological activities including anticoagulant (Shanmugam & Mody, 2000) antioxidant (Ruperez et al., 2002) and antihypertension
activities (Caceres et al., 2000; Carlucci et al., 1997). Anticoagulant and antithrombic activities are the most widely studied properties of sulfated polysaccharides. Anticoagulant activity of sulfated polysaccharides has been identified from several brown seaweeds such as Padina gymnospora Kuetzing (Dictyotaceae) (Silva et al., 2005), Dictyota mensteralis Hoyt (Dictyotaceae) (Albuquerque et al., 2004), Sargassum stenophyllum J. Agardh (Sargassaceae) (Duarte et al., 2001), Spatoglossum schroederi Mertens (Dictyotaceae) (Leite et al., 1998) and the red seaweed Gigartina skottsbergii Setchell & Gardner (Gigartinaceae) (Carlucci et al., 1997).
Most of the available reports focus on the anticoagu-lant properties of brown and green algae. In addition to the available reports, fucoidan prepared from a brown algae F. vesiculus, is commercially available at present. This fucoidan is composed of 44.1% fucose, 26.3% sulfate and 31.1% ash and little aminoglucose. It was found that the main component unit was 1,2-α-fucose and most of the sulphate groups was located at the position C-4 of the fucose units (Conchie & Percival, 1950; O’neill, 1954). On the contrary, there are no reports on the anticoagu-lants from Indian red algae. The present study was con-ducted to purify and characterize an anticoagulant from the red seaweed, particularly Asparagopsis taxiformis (Delile) Trevisan (Bonnemaisoniaceae) collected from the southwest coast of India.
10 Thalassas, 28(2) · July 2012
0 10 20 30 40 50-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6O
D 5
25nm
fractions
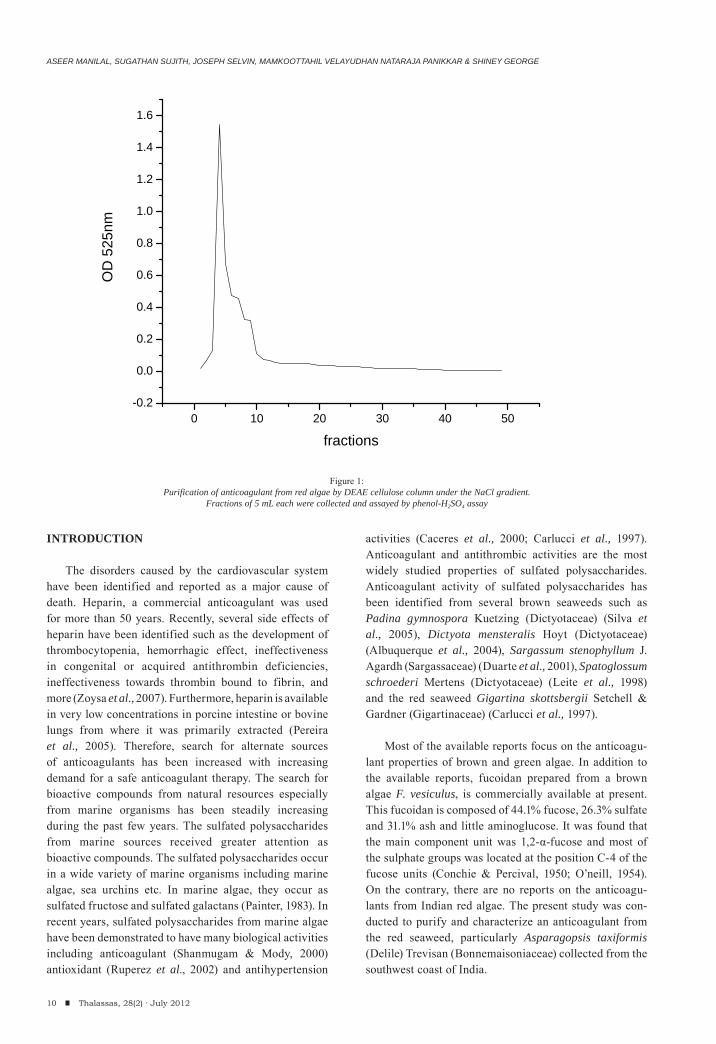
Figure 1:Purification of anticoagulant from red algae by DEAE cellulose column under the NaCl gradient.
Fractions of 5 mL each were collected and assayed by phenol-H2SO4 assay

ANTICOAGULANT POTENTIAL OF POLYSACCHARIDE ISOLATED FROM THE INDIAN RED ALGA, Asparagopsis taxiformis (Delile) Trevisan
MATERIALS AND METHODS
Collection of seaweeds
Five species of red algae (Asparagopsis taxiformis, Hypnea valentiae (Turner) Montagne, Champia compressa Harvey, Herposiphonia insidiosa (Greville ex J. Agardh) and Gelidium micropterum Kützing) growing exclusively on the intertidal rocky substratum were collected with utmost care to avoid contamination of other algae (Manilal et al., 2010; 2011). The seaweeds were collected during different seasons (April 2010 to March 2011) during the ebb tide from the following reefs along southwest coast of India, Kollam (08º 54’ N & 76º 38’ E) area. Epiphytic and extraneous matters were removed by washing the seaweeds first in seawater and then in fresh water (deionised water). The algae were transported to the laboratory in plastic bags under ice. Voucher specimens were stored in formalin solutions for taxonomic identification and future reference. A part of the collected samples were frozen at -20˚C for further extraction procedures.
Identification of the seaweeds
The morphological and anatomical characteristics of the collected seaweeds were studied using microscopic techniques. The fine microtome sections were stained in saffranin and analyzed under light microscopy (Optica). Based on the morphological and anatomical features, the taxonomic identification and classification was carried out. The characteristics considered for the identification includes length and diameter of the rhizome, its internal structure, distribution of rhizome length, width of the assimilator, colour, diameter of the peltate head, length of the plant, number and length of the lateral branches, size and shape of primary, secondary, and tertiary leaves, size
and shape of air bladders, branching and length of recep-tacles, shape and size of hold fast, reproductive characters like spermatia, tetra sporangia, and cystocarp.
Extraction of polysaccharides
The shade-dried algal samples were dried completely at 50˚C under ventilation in an oven (Neolabs), ground in to a fine powder and stored in capped glass vials for analysis. The finely powdered algal sample was incu-bated with acetone (1:5) to eliminate lipids and pigments. Acetone was separated from the algal residues by cen-trifugation at 6000 x g for 10 min at 30˚ C (Eppendorf) followed by evaporation. The algal residue was extracted twice at 50˚C for 6 h with 0.1N HCl (20 g algal powder/L). The filtered extracts were combined and neutralized with aqueous NaOH. Salts and small molecules were removed by dialysis (MWCO ca. 14 kDa). The aliquot was con-centrated under reduced pressure in a rotary evaporator (Yamato, Japan). The polysaccharides were precipitated by addition of 4 fold volume of 95% (v/v) ethanol, and washed twice with absolute ethanol, then dried at 60˚C to obtain crude polysaccharide.
Chemical analysis
Protein content The extracted polysaccharide was hydrolyzed with
10% trichloroacetic acid (Merck). The TCA was added to the extract so as to make a final concentration of 3% TCA. The total protein that precipitated was estimated according to Lowry et al. (1951).
Total polysaccharide Total polysaccharide was estimated by phenol-H2SO4
method (Dubois et al., 1956) using glucose as standard.
11Thalassas, 28(2) · July 2012
Red algae Clotting time (s)*
Herposiphonia insidiosa 55 ± 5.2 Gelidium micropterum 77 ± 2.8
Champia compressa 57 ± 7.6
Hypnea valentiae 45 ± 9.1
Asparagopsis taxiformis 240 ± 5.5
Deionized water (control) 35 ± 2.5 *The activity was expressed by the time (s) taken to clot the human plasma. Water was used as a control. All the values were the mean of triplicates ± Standard error.
Table 1:Anticoagulant activity of all the five red algae as determined by APTT assay
ASEER MANILAL, SUGATHAN SUJITH, JOSEPH SELVIN, MAMKOOTTAHIL VELAYUDHAN NATARAJA PANIKKAR & SHINEY GEORGE
Briefly, 100 μl sample was mixed with 1 mL of 5% phenol and 5 mL of 96% sulfuric acid. The absorbance was read at 490 nm in a UV/Vis double beam scanning spectropho-tometer (Thermospectronic) after 30 min of incubation at room temperature (32˚C).Sulfate content
Total sulfate content was measured according to Dodgson, (1961) using K2SO4 as standard. 0.2 mL polysaccharide solution was added to 3.8 mL of 4% tri-chloroacetic acid followed by 1.0 mL of the BaCl2-gelatin reagent. After mixing, the aliquot was incubated for 20 min at room temperature. A reagent blank was prepared by adding 0.2 mL of water instead of sulfate containing solution. The absorbance of the solution was measured at 360 nm.
Anticoagulant assay
Human blood was collected from 10 healthy donors and pooled into conical tubes with 2.5% sodium citrate solution (9:1 v/v). The plasma was separated by centrifuging the blood samples at 6000 x g at 4˚C for 20 min. The plasma was stored at -70˚C for further analysis. Anticoagulant activity was deter-mined by Activated Partial Thromboplastin Time (APTT) and Prothrombin Time (PT) assays according to the manufacturer’s guidelines (Fisher Scientific Company, USA). The activity was compared with heparin, a commercial anticoagulant. The difference in the anticoagulant activity was statistically analyzed
using ANOVA with tukey test for a multiple compari-son by MINITAB statistical software (MINITAB inc., Version 15, PA, USA).
Fractionation and purification of sulfated polysaccharide
The crude polysaccharide sample (200 mg) was purified by anion exchange chromatography using DEAE-cellulose column. The sample was applied to a DEAE-cellulose column which was pre-equilibrated with 50 mM sodium acetate (pH 5.0) and washed with 200 mL of the same buffer containing 0.2 M NaCl. The column was eluted by a linear gradient prepared by mixing 150 mL of 50 mM sodium acetate (pH 5.0) containing 0.2 M NaCl with 150 mL of 2 M NaCl in the same buffer with the f low rate of 60 mL/h. Then, fractions collected were analyzed for total polysaccha-ride, sulfate and protein contents as described earlier. Coagulation time was checked for all polysaccharide positive fractions by APTT assay. The fractions with high anticoagulant activity were selected for further investigations.
Analysis of purity of the polysaccharide
The purity of the polysaccharide was determined by 0.5% agarose gel electrophoresis according to Pereira et al. (1999). Brief ly, 50 μg of the purified polysac-charide was applied to a 0.5% agarose gel in 0.05 M sodium acetate buffer (pH 9.0) and electrophoresed for 1 h at 100 V. The polysaccharide in the gel was fixed
12 Thalassas, 28(2) · July 2012
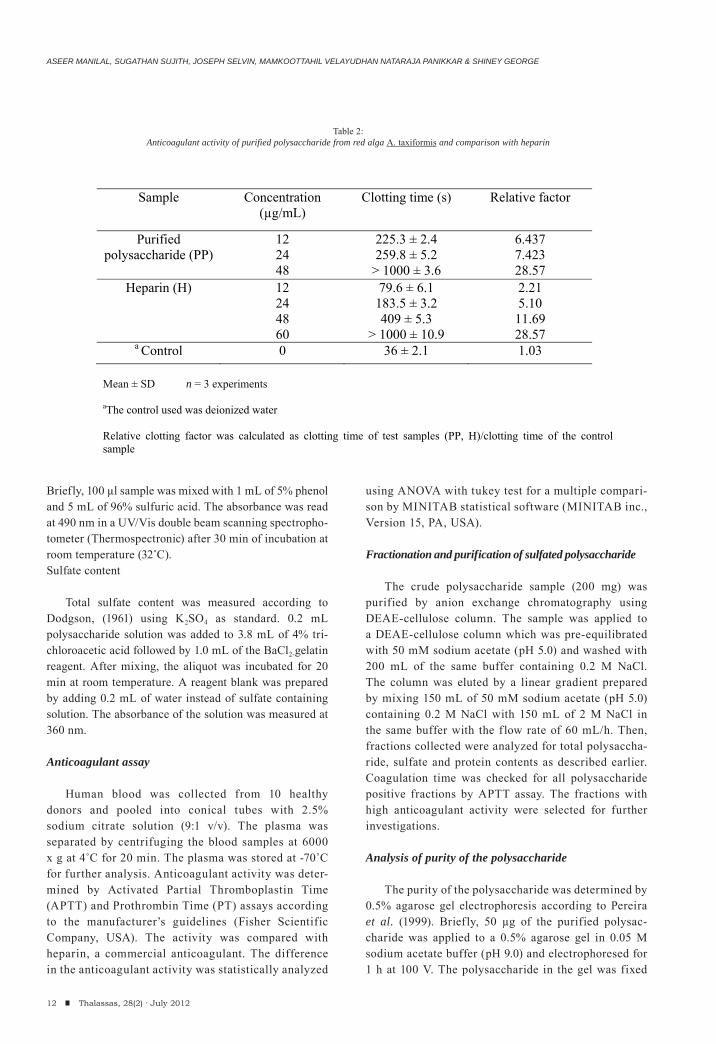
Sample Concentration (μg/mL)
Clotting time (s) Relative factor
Purified polysaccharide (PP)
12 24 48
225.3 ± 2.4 259.8 ± 5.2
> 1000 ± 3.6
6.437 7.423 28.57
Heparin (H) 12 24 48 60
79.6 ± 6.1 183.5 ± 3.2 409 ± 5.3
> 1000 ± 10.9
2.21 5.10 11.69 28.57
a Control 0 36 ± 2.1 1.03 Mean ± SD n = 3 experiments
aThe control used was deionized water Relative clotting factor was calculated as clotting time of test samples (PP, H)/clotting time of the control sample
Table 2:Anticoagulant activity of purified polysaccharide from red alga A. taxiformis and comparison with heparin

ANTICOAGULANT POTENTIAL OF POLYSACCHARIDE ISOLATED FROM THE INDIAN RED ALGA, Asparagopsis taxiformis (Delile) Trevisan
with 0.1% cetyl trimethyl ammonium bromide (CTAB) solution. After 12 h, the gel was dried and stained with 0.1% toluidine blue in acetic acid:ethanol:water (0.1:5:5 v/v) and destained with the same solution without tolui-dine blue.
Determination of the molecular mass of the purified polysaccharide
The molecular mass of the purified polysaccharide was determined using 12% PAGE. In this method, 50 μg of the purified polysaccharide was applied to a 12% pol-yacrylamide gel resolved in 0.5 M Tris buffer (pH 8.6) for 45 min at 100 V. After electrophoresis, the gel was silver stained. Different molecular weight markers including 500 kDa (Dextran sulfate), 60 kDa (Chondroitin 6 sulfate from shark cartilage), 20 kDa (Chondroitin sulfate B) and 8 kDa (Dextran sulfate from Leuconostoc species) were used to determine the molecular mass of the purified polysaccharide.
RESULTS AND DISCUSSION
Extraction of polysaccharide
In the present study, five red algae collected from the southwest coast (Kollam coast) of India were screened for anticoagulant activity. Acidic treatment of (0.1 N HCl for 6 h) algae yielded a crude polysaccharide extract (CPE). The anticoagulant activities of algae are presented in Table 1. A. taxiformis showed pronounced anticoagulant activity with an extended clotting time of 240 seconds. Thus the crude polysaccharide extract was taken into account for further purification and characterization procedures.
Chemical composition of crude polysaccharide
The chemical composition of CPE obtained from A. taxiformis include 59.15 mg/mL of total polysaccharides, 58.59 mg/mL of sulphated polysaccharides and 10.48 mg/mL of protein. The total polysaccharide and sulfate content was found to be higher when compared to
that of protein content. Rogers et al. (1990) observed the anticoagulant activity was directly proportional to the carbohydrate and sulfate contents and inversely proportional to the protein content. The pH of the purified polysaccharide was found to be 3.8 predicting it to be an acidic polysaccharide.
Fractionation and purification of polysaccharide
The sample containing 162.4 mg of total polysac-charide was used as a starting material for DEAE cellulose column. The elution profile of polysacchari-des on DEAE showed a single distinct peak at the NaCl gradient 0.2-0.4 M (fractions F3- F10) (Fig. 1). Fraction F4 had the highest polysaccharide concentration (24 μg/mL) with a highest coagulation time of 259.8 sec.
Characterization of the purified polysaccharide
The purity was determined by studying the migration pattern of the algal polysaccharide on an agarose gel. Based on the migration pattern, the molecular weight of the purified polysaccharide was determined between 60 to 500 kDa. The band of the algal polysaccharide lies between Chondroitin 6 sulfate (60 kDa) and dextran sul-fate (500 kDa). The anticoagulant sulfated galactan from green alga Codium pugniformis had a molecular weight of 100–500 kDa (Matsubara et al., 2000). The purified polysaccharide resulted as a broad band on PAGE. It suggested that the purified polysaccharide anticoagulant of A. taxiformis behaved as a heterogeneous system, similar to the polysaccharides identified from red alga B. occidentalis Borgesen (Rhodymeniaceae) (Farias et al., 2000), Gelidiuim crinale Turner (Gelidiaceae) (Pereira et al., 2005), Lomentaria catenata Harvey (Lomentariaceae) (Pushpamali et al., 2008) and G. cornea (Melo et al., 2002). The similar behavior of the sulfated galactans has also been reported in marine invertebrates (Pavao et al., 1989; Yamada et al., 2000). High molecular weight carrageenans with high sulfur content showed higher anticoagulant activity than those with low molecular weight with low sulfur content (Shanmugam & Mody, 2000).
13Thalassas, 28(2) · July 2012
Coagulation Assay Clotting time (s) Relative clotting factor Control Polysaccharide ( g/mL)
APTT 36.0 259.8 7.22 PT 13.7 44 3.22
Relative clotting factor was calculated as clotting time of test samples /clotting time of the control sample
Table 3:Comparison of anticoagulant activity of purified polysaccharide by different coagulation assays of APTT and PT

ASEER MANILAL, SUGATHAN SUJITH, JOSEPH SELVIN, MAMKOOTTAHIL VELAYUDHAN NATARAJA PANIKKAR & SHINEY GEORGE
Anticoagulant activity of the purified polysaccharide
The potency of the purified polysaccharide was determined by detecting the anticoagulant activity (APTT assay) for varied concentration of the polysaccharide. The clotting time of the sample was compared with increasing concentration series of the standard heparin (Table 2). The clotting time was found to be increased with increasing concentration of both sample and heparin. The purified polysaccharide at a concentration of 48 μg/mL showed a clotting time over 1000 sec. Whereas standard heparin showed similar activity at a concentration of 60 μg/mL. Statistically, the difference between the clotting time of the purified polysaccharide and heparin did not differ significantly (p= 0.373). Thus the purified polysaccharide has a similar anticoagulant property to that of commercial heparin. The inhibition pathway of the purified polysac-charide from A. taxiformis was determined by APTT and PT assays (Table 3). The addition of polysaccharide prolonged the coagulation time in both APTT and PT assays envisaging the inhibition of both intrinsic and extrinsic pathways of coagulation. Furthermore, the rela-tive clotting factor by PT assay was found to be within the range of normal recommended oral anticoagulant agent (Manufacturer’s guidelines). Caporiccio et al. (1984) reported that armatan, an acid sulphated polysaccharide isolated from a red algae, Asparagopsis armata (Harv.), increases the coagulation time of the rat plasma in vivo assays. The anticoagulant activity of Asparagopsis was more or less similar to those reported in other algal spe-cies (Mao et al., 2009; Pushpamali et al., 2008). Based on the present findings, the seaweed A. taxiformis might be a natural source for developing anticoagulant in future. Therefore, bioassay guided fractionation to identify the anticoagulant principles of A. taxiformis are needed in the subsequent investigation.
CONCLUSION
The blood coagulation is an extremely complicated mechanism. To determine the exact anticoagulation site, further studies have to be carried out. The present study could be considered as an attempt to propose anticoagu-lant activity in red seaweeds, which are less extensively studied for their anticoagulant activity. However, the A. taxiformis polysaccharide from Indian coast has seldom been explored for anticoagulant activity. In conclusion, the present study comes up with a new insight for the development of novel anticoagulant agent from A. taxi-formis.
ACKNOWLEDGMENT
The author Aseer Manilal is gratefully acknowled-
ged to Council of Scientific and Industrial Research for providing Senior Research Fellowship (File No. 09/475 (0149)/2010EMR-I).
REFERENCES
Albuquerque IRL, Queiroz KCS, Alves LG, Santos EA, Leite EL, Rocha HAO (2004). Heterofucans from Dictyota menstrualis have anticoagulant activity. Brazilian Journal Medicinal Biological Research, 37: 167–171.
Caceres PJ, Carlucci, MJ, Damonte EB, Matsuhiro B, Zuinga EA (2000). Carrageenans from chilean samples of Stenogramme interrupta: Structural analysis and biological activity. Photochemistry, 53: 81–86.
Caporiccio B, Braun M, Vignaud M, Chalet M, Codomier L, Teste J, Catayée G (1984). Effect of an acidic sulfated polysaccharide on whole blood coagulability. In vivo study in rats. Comptes Rendus des Seances de la Societe de Biologie et des ses Filiales (Paris) 178(6):691-696.
Carlucci MJ, Pujol CA, Ciancia M, Noseda MD, Matulewicz MC, Damonte EB, Cerezo AS (1997). Antiherpetic and anticoagulant properties of carrageenans from the red seaweed Gigartina skottsbergii and their cyclized derivatives: correlation between structure and biological activity. International Journal of Biological Macromolecules, 20:97–105.
Conchie J, Percival EGV (1950). Fucoidin part II. The hydrolysis of a methylated fucoidin prepared from Fucus vesiculosus. Journal of Chemical Society, 827–833.
Dodgson KS (1961). Determination of inorganic sulphate in studies on the enzymic and non-enzymic hydrolysis of carbohydrate and other sulphate esters. Biochemical Journal, 78: 312-319.
Duarte MER, Noseda DG, Noseda MD, Tulio S, Pujol CA, Damonte EB (2001). Inhibitory effect of sulphated galactans from the marine algae Bostrychia montagnei on herpes simplex virus replication in vitro. Phytomedicine, 8: 53–58.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956). Calorimetric method for determination of sugars and related substances. Analytical Chemistry, 28: 350–356.
Farias WRL, Valente AP, Pereira MS, Mourao PAS (2000). Structure and anticoagulant activity of sulfated galactans. Isolation of a unique sulfated galactan from the red algae Botryocladia occidentalis and comparison of its anticoagu-lant action with that of a sulfated galactans from inverte-brates. Journal of Biological Chemistry, 275: 29299–29307.
Leite EL, Medeiros MGL, Rocha HAO, Farias, GGM, Silva LF, Chavante SF, Abreu LRD, Dietrich CP, Nadir HB (1998). Structure and pharmacological activities of sulphated xylo-fucoglucuronan from the alga Spatoglossum schroederi. Plant Science, 132: 215–228.
Lowry OH, Rosebrough N, Farr A, Randall R (1951). Protein measurement with Folin phenol reagent. Journal of Biological Chemistry, 193: 265–275.
14 Thalassas, 28(2) · July 2012

ANTICOAGULANT POTENTIAL OF POLYSACCHARIDE ISOLATED FROM THE INDIAN RED ALGA, Asparagopsis taxiformis (Delile) Trevisan
Manilal A, Sujith S, Kiran GS, Selvin J, Shakir C, Lipton AP (2010). Antimicrobial potential of marine organisms col-lected from the southwest coast of India against multiresist-ant human and shrimp pathogens. Scientia Marina, 74 (2): 287-296.
Manilal A, Thajuddin N, Selvin J, Idhayadhulla A, Surendra Kumar R, Sujith S, (2011). In vitro Mosquito Larvicidal Activity of Marine Algae Against the Human Vectors, Culex quinquefasciatus (Say) and Aedes aegypti (Linnaeus) (Diptera: Culicidae). International Journal of Zoological Research, 7: 272-278.
Mao W, Li H, Li Y, Zhang H, Qi X, Sun H, Chen Y, Guo S (2009). Chemical characteristic and anticoagulant activity of the sulfated polysaccharide isolated from Monostroma latis-simum (Chlorophyta). International Journal of Biological Macromolecules, 44 (1): 70-74.
Matsubara K, Matsuura Y, Hori K, Miyazawa K (2000). An anti-coagulant proteoglycan from the marine green alga, Codium pugniformis. Journal of Applied Phycology, 12: 9–14.
Melo MRS, Feitosa JPA, Freitas ALP, De Paula, RCM (2002). Isolation and characterization of soluble sulfated polysaccha-ride from the red seaweed Gracilaria cornea. Carbohydrate Polymers, 49: 491– 498.
O’neill AN (1954). Degradative studies on fucoidan. Journal of American Chemical Society, 76: 5074–5076.
Painter TJ (1983). Algal polysaccharides. In Aspinall GO (Ed). The polysaccharides (Vol 2), Academic Press, New York, 195–285.
Pavao MS, Albano RM, Lawson AM, Mourao PA (1989). Structural heterogeneity among unique sulfated L-galactans from different species of ascidians (tunicates). Journal of Biological Chemistry, 264: 9972–9979.
Pereira MG, Benevides NMB, Melo MRS, Valente AP, Melo FR, Mourao PA, S (2005). Structure and anticoagulant activity of a sulfated galactan from the red alga, Gelidium crinale. Is there a specific structural requirement for the anticoagulant action? Carbohydrate Research, 340: 2015–2023.
Pereira MS, Mulloy B, Mourao PAS (1999). Structure and anti-coagulant activity of sulphated fucans: Comparison between the regular, repetitive and linear fucans from echinoderms with the more heterogenous and branched polymers from brown algae. Journal of Biological Chemistry, 274: 7656–7667.
Pushpamali WA, Nikapitiya C, Zoysa MD, Whang I, Kim SJ, Lee J (2008). Isolation and purification of an antico-agulant from fermented red seaweed Lomentaria catenata. Carbohydrate Polymers, 73: 274–279.
Rogers DJ, Jurd KM, Bluedan G, Paoletti S, Zanetti F (1990). Anticoagulant activity of a proteoglycan in extracts of Codium fragile spp. Atlanticum. Journal of Applied Phycology, 2: 357–361.
Ruperez P, Ahrazem O, Leal JA (2002). Potential antioxidant capacity of sulphated polysaccharides from the edible brown seaweed Fucus vesiculosis. Journal of Agricultural Food and Chemistry, 50: 840-845.
Shanmugam S, Mody KH (2000). Heparinoid active sulphated polysaccharides from marine algae as potential blood anti-coagulant agents. Current Science, 79:1672– 1683.
Silva TMA, Alves LG, De Queiroz KCS, Santos MGL, Marques CT, Chavanta SF, Rocha HAO, Leite EL (2005). Partial characterization and anticoagulant activity of a heterofucan from the brown seaweed Padina gymnospora. Brazilian Journal Medicinal Biological Research, 38: 523–533.
Yamada T, Ogamo A, Saito T, Uchiyama H, Nakagawa Y (2000). Preparation of O-acylated low-molecular-weight carrageen-ans with potent anti-HIV activity and low anticoagulant effect. Carbohydrate Polymers, 41:115–120.
Zoysa MD, Nikapitiya C, Jeon YJ, Jee Y, Lee J (2007). Anticoagulant activity of sulphated polysaccharide isolated from fermented brown seaweed Sargassum fulvellum. Journal of Applied Phycology, 20: 67–74.
15Thalassas, 28(2) · July 2012
(Received: December, 16, 2011; Accepted: March, 7, 2012)


CORAL REEFS QUANTITATIVELY ASSESSMENT ALONG THE EGYPTIAN RED SEA COAST
(1) Associated Prof. of marine invertebrates, National Institute of Oceanography and Fisheries,
Red Sea Branch – Hurghada, Red Sea, Egypt.
Tel/Fax. 002 065 3500103 / E-mail: [email protected]
Thalassas, 28(2) · July 2012: 17-26An International Journal of Marine Sciences
Key words: Coral assessment, diversity, community distribution, evenness index, Red Sea, Egypt.
Palabras clave: distribución de corales, diversidad, distribución de la comunidad, índice de uniformidad, Mar Rojo, Egipto.
TAREK ABDEL-AZIZ A. MOHAMMED(1)
17Thalassas, 28(2) · July 2012
ABSTRACT
Coral assessment and distribution have been studied along the Egyptian Red Sea coast from north Hurghada with 5km to Shalateen illustrating the most important factors that affect the coral distribution and abundance at the selected sites. The cover percentage of the coral reef community was estimated at each locality by using the standard method (the line intercept transect. During the present investigation, 68 coral species were recorded at seven coastal sites along the Egyptian Red Sea; forty-nine species of them were hard corals and the other 19 species were soft corals. North Hurghada site (NIOF) recorded the least cover of the living coral (66.23%) while Abu-Dabab area recorded the maximum coverage percent (91.50%). In spite NIOF site recorded the least cover, it measured the maximum species diversity (3.54) due to the maximum recorded number of species (48 species); while Shalateen recorded the least diversity (1.97) due to the least number of coral species (24 species).
The highest hard coral cover Pocillopora damicornis (15.6%) at El Sharm El-Bahari and the highest soft coral is Sarcophyton glaucum (10.18%) at North Qula’an. Some environmental characteristic variations and biological interaction between benthos, the anthropogenic activities, overfishing, tourism developments, as well as petroleum and phosphate pro-duction, Sedimentation processes, bottom topography and geomorphology are the main controlling factors of coral distribu-tion at the studied areas.
RESUMEN (Evaluación cuantitativa de arrecifes de coral a lo largo de la costa egipcia del Mar Rojo)
Se ha estudiado la distribución de corales a lo largo de la costa egipcia del Mar Rojo desde el norte de Hurghada hasta 5 km de Shalateen, relacionándolo con los factores mas importantes que afectan la distribución y abundancia de corales en los sitios seleccionados. El porcentaje de cobertura de los corales en la comunidad de arrecifes se estimó en cada localidad mediante el uso del método estándar (transecto de interceptación). Los resultados arrojaron 68 especies de coral en siete sitios a lo largo de la costa egipcia del Mar Rojo, siendo cuarenta y nueve especies de corales duros mientras se encontraron 19 especies de corales blandos. El lugar de muestreo del Norte de Hurghada (NIOF) registró la menor cobertura de coral vivo (66,23%) mien-tras que el área de Abu-Dabab registró la cobertura máxima en porcentaje (91,50%). Aunque la cobertura mínima se encontró en NIOF, apareció la máxima diversidad específica (3,54) debido a la cantidad máxima registrada de especies (48 especies); mientras que Shalateen registró la menor diversidad (1,97) debido al menor número de especies de corales (24 especies).
La mayor cobertura de coral duro Pocilloporadamicornis (15,6%) se registró en Sharm El-Bahari y la más alta de corales blandos fue Sarcophytonglaucum (10,18%) en el Norte de Qula’an. Los principales factores que controlan la distribución de los corales en el área de estudio fueron, además de algunas variaciones ambientales, las interacciones bentónicas, efectos antropogénicos como la pesca excesiva, el incremento del turismo, la producción de petróleo y la liberación de fosfato, ade-más de procesos sedimentarios, la topografía del fondo y la geomorfología.
TAREK ABDEL-AZIZ A. MOHAMMED
18 Thalassas, 28(2) · July 2012
INTRODUCTION
The Red Sea is still one of the most important areas that contains beautiful coral communities and are widespread throughout the tropical Indo-Pacific area. The abundance and the ecology of the hard and soft corals have been studied by many authors in the Red Sea and the Indo-Pacific regions (Cray, 1931 and Crossland, 1938) and in the central Great Barrier Reef (Diensen, 1983; Dai, 1990 and Fabricius, 1997). Moreover, the coral distribution in some localities of the Red Sea have been studied generally referring to the community structure of coral reefs (Ammar & Nawar, 1998; Ammar, 2003 and 2004), ecology and biology (Loya, 1972; Kotb, 1996; Kotb et al., 2001;
Mohammed, 2003 and 2006), the interaction of many factors that affecting the distribution and affect the coral bleaching (Mohammed and Mohamed, 2005), the affecting factors as sedimentation, overfishing, tourist activities, as well as petroleum and phosphate production (Mohammed et a.l, 2009), geographical relationship and geomorphological observations of coral reefs at the northern Red Sea (Scheer, 1971) and the basis of topographical characteristics of the reef (Loya, 1972). However, all of these factors had significant influencing on the distribution of corals among coral reefs at the studied areas. On the other hand, the biology and ecology of soft corals have been shown by Gohar (1940), Fishelson (1970 & 1973) Benayahu and Loya (1981) and Merganer & Schumacher (1981).
Figure 1:The selected studied sites along the Red Sea coast. (Red points)
CORAL REEFS QUANTITATIVELY ASSESSMENT ALONG THE EGYPTIAN RED SEA COAST
19Thalassas, 28(2) · July 2012
Mohammed (2006) and Mohammed et al. (2009) concluded that, many factors could affect the distribution of coral reefs and their structure and abundance such as the biological interaction between the benthos fauna, the bottom topography and geomorphology (Adjeroud et al., 2000; Kotb et al., 2001; Ouillon et al., 2004; Andréfouët and Guzman, 2005) as well as the physical factors and anthropogenic activities (Ammar and Nawar, 1998 and Mohammed, 2003). These activities include phosphate shipping smoothers and navigation activities, landfill and dredging, mining operations and overfishing (Daby 2003). These areas are controlled by; water depth, temperature variation (Rogers, 1990), tidal range and the degree of exposing, salinity and water mixing (Babcock and Davies, 1991), light penetration, geographic occurrence, the geomorphologic nature (Kotb, 2001; Abou Zaid and Kotb, 2000) and bottom sediment nature, turbidity and terrestrial inputs.
The present work aims to evaluate, assess quantitatively and compare the corals distribution, diversity and abundance along the Egyptian Red Sea Coast during January 2009 to February 2010. However the study will illustrate the different ecological factors that affect the coral diversity and richness as well as to explore the human threats on coral communities at each site.
MATERIALS AND METHODS
Area of study
During the present investigation seven sites were selected northern Red Sea along the Egyptian coast to evaluate and calculate the coral community, diversity and abundance referring to the most important factors affecting the coral diversity and distribution (Figure 1). These sites are highly influenced by different factors and activities;
such as phosphate shipment at Safaga and El-El-Hmrawin; overfishing at Shalateen; touristic activities (diving and snorkelingat) at NIOF, Sharm El-Naqa, El-Sharm El-Bahari and Abu-Dabab; coastal leveling and landfilling at NIOF; the effect of an active valley at Qula’an and. The features and characteristics of the selected stations, as well as the oceanographic parameters were listed in table (1). The program of samples collection is based on the NIOF field trips, starting in the June 2009 and ended March 2010, covering seven different distributed areas located at the Egyptian coast of Red Sea.
Methods
Studied sites were surveyed using the line intercept transect (LIT) methods (English et al., 1997) to evaluate the percentage cover of corals in the area relative to the other benthos using SCUBA diving equipments. Each transect has 20 m length and 2 m gap between the neighbor transects. Three replicate transects were counted and averages were calculated at sub-equal depths from 3 to 7 meters for all the selected sites. A total of 24 transects were surveyed from all of the studied sites, where the percentage cover and number of soft and hard corals were estimated. Also, the living corals (soft, hard) and dead corals were calculated. The percentage covers of other taxa including algae, sponges, gorgonians, sea anemones and sand with rocks were also estimated. The coral samples were brought to the laboratory for identification. They were preserved in 4% formalin in seawater, rinsed in fresh water after 24 h, and then transferred to 70% ethyl alcohol. Sclerites or spicules (endoskeleton) were obtained by dissolving soft coral tissues in 10% sodium hypochlorite. The soft corals (Alcyonacea) were identified according to Macfadyen (1929), Thomson & Dean (1931), Verseveldt (1982), and Fabricius & Alderslade (2001). Moreover, the hard corals
05
101520253035404550
MBS
Sharm El-N
aga
El-Hmraw
in
Sharm El-B
ahari
North Qula'a
n
Abu-Daba
b
Shlateen
Num
ber o
f spe
cies
0.0010.0020.0030.0040.0050.0060.0070.0080.0090.00
% c
over
NIOF
Sharm
El-N
aga
El-Hmraw
in
Sharm
El-B
ahari
North Q
ula'an
Abu-D
abab
Shlatee
n
Hard coralsSoft corals
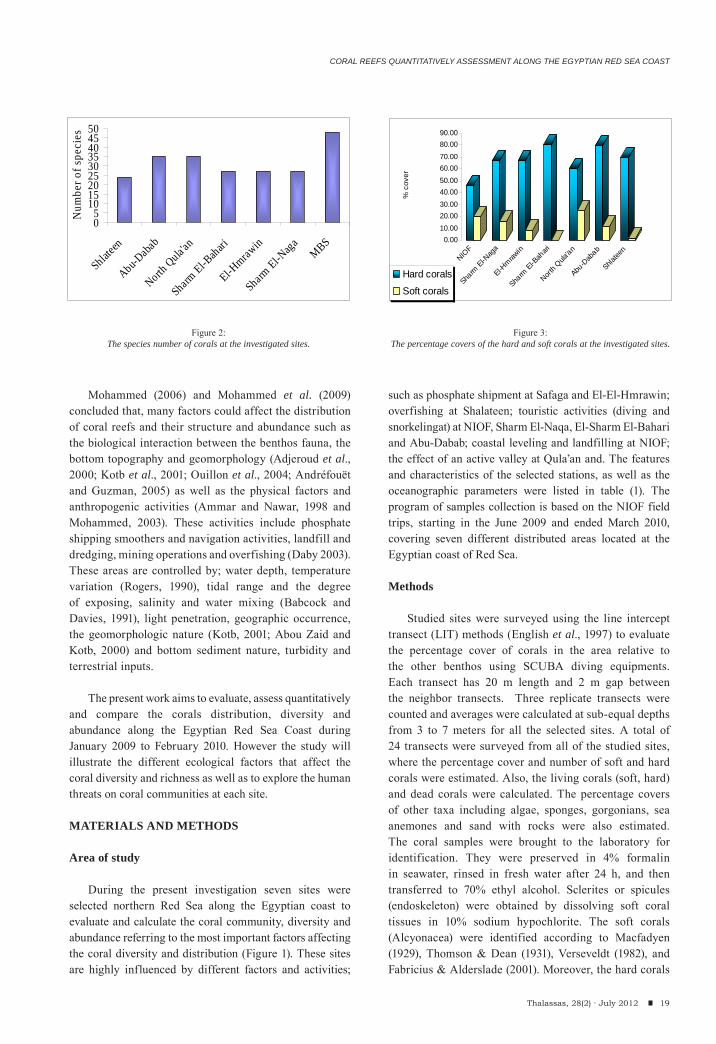
Figure 2:The species number of corals at the investigated sites.
Figure 3:The percentage covers of the hard and soft corals at the investigated sites.

TAREK ABDEL-AZIZ A. MOHAMMED
20 Thalassas, 28(2) · July 2012
were identified according to Sheppard & Sheppard (1991) and Veron (2000).
The percentage cover was calculated from the following formula:
Diversity (H`) and evenness index (J) was calculated in each lagoon according to Shannon-Wiener (1948) and Pielou (1966):
i) Shannon-Wiener species diversity (Hs).
s = Total species, (i) = Each species
ii) Pielou s evenness index (J).
where, s = number of species.
Some physical factors (temperature, salinity, and dissolved oxygen) were measured at each site directly by hydrolab instrument (model Surveyor 4, 1997).
RESULTS
Coral distribution along the coastal area:
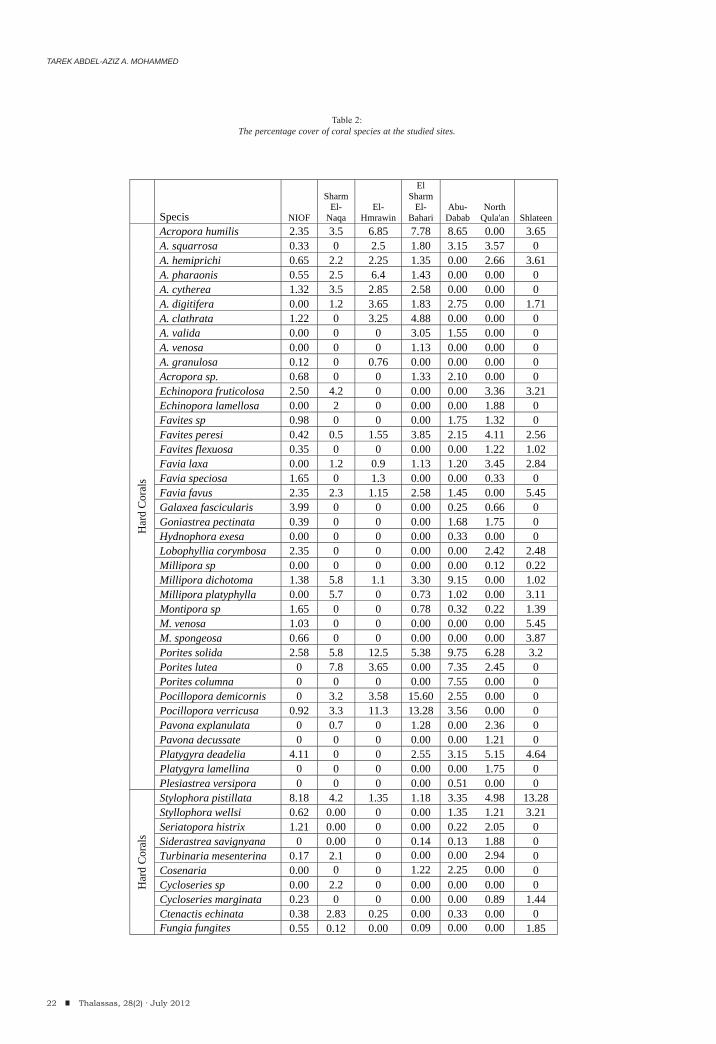
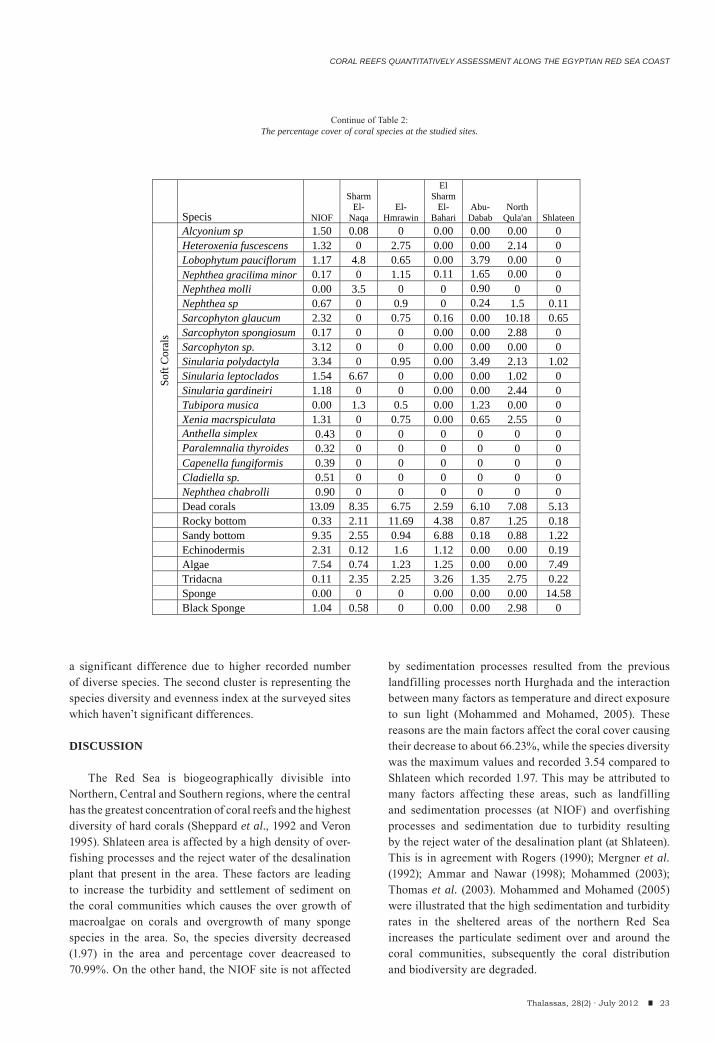
During the present study a total of 68 coral species belonging to 35 genera were surveyed during the present investigation where, 49 species of them were hard corals (belonging to 23 hard coral genera) and 19 species (belonging to 12 genera) were soft corals (Table 2) in addition to the other species that observed and not
intercepted in the line transects. Hurghada site recorded the highest number of Species (48 species), while the lowest number (24 species) was recorded at Shlateen (Figure 2). So, Hurghada illustrated the highest diversity than any other studied sites. The highest percentage cover of the hard corals has been recorded at El Sharm El-Bahari and reached about 80.25% whereas; the lowest percentage cover has been recorded at Hurghada in front of the NIOF Red Sea Branch (Marine Biological Station, MBS) with 45.87 %. On the other hand, El Sharm El-Bahari recorded the least cover of the soft corals (0.27%) and the highest value was recorded at North Qula’an (24.84%). The dead corals ranged between 2.59% at El Sharm El-Bahari and 13.09% at NIOF (Figure 3).
Pocillopora demicornis and Stylophora pistillata recorded the highest percentage cover of the hard coral species (15.60 at El Sharm El-Bahari and 13.28 at Shlateen respectively); moreover, Sarcophyton glaucum and Sinularia leptoclados recorded the highest soft coral species (10.18 and 6.67%) at North Qula’an and Sharm El-Naqa respectively (see table 2). Moreover, Acropora, Favites, Favia, Millipora, Porites, Pocillopora and Stylophora are the most frequent and common hard coral genera; while, Nephthea, Sarcophyton, Sinularia and Xenia are common and abundant soft coral genera.
Community structure and biodiversity of corals:
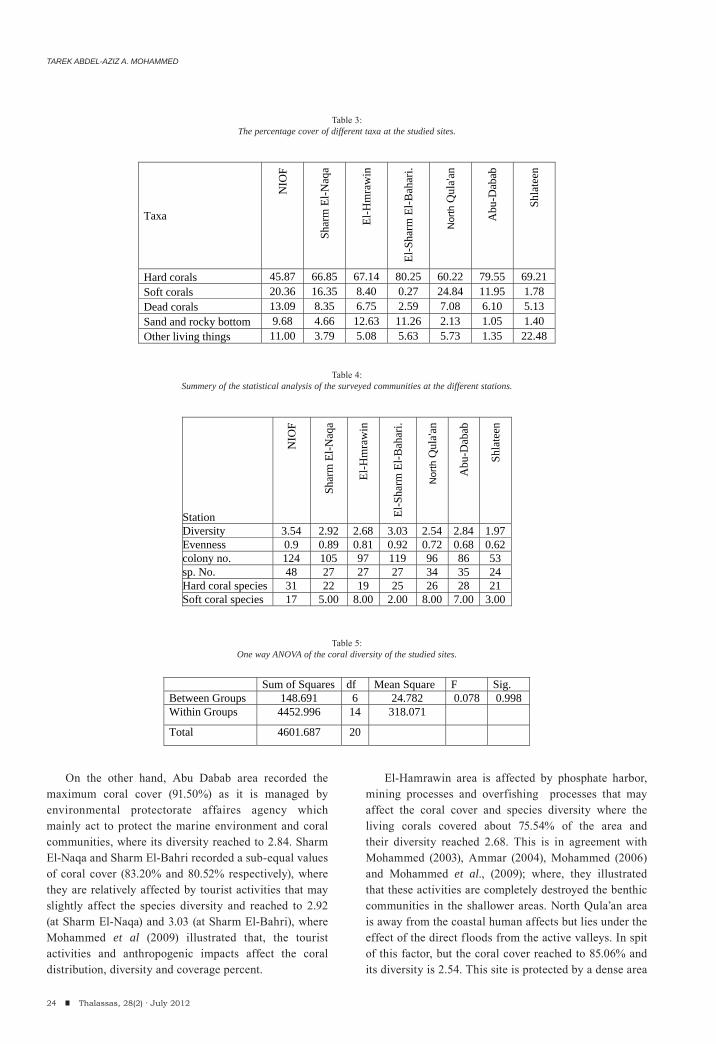
During the present investigation, the coral community recorded its highest cover of living corals (91.5%) at Abu-Dabab locality followed by North Qula’an, Sharm El-Naqa and El Sharm El-Bahari (85.06%, 83.20% and 80.52% respectively). While the lowest cover was demonstrated at NIOF (66.23%), which recorded the highest percent of dead corals (13.09%), while El Sharm El-Bahari has the lowest percent of dead corals and reached about 2.59% (Table 3 and figure 3). On the other hand, El Sharm El-Bahari recorded the highest
0102030405060708090
100
% c
over
NIOF
Sharm El-N
aga
El-Hmraw
in
Sharm El-B
ahari
North Q
ula'an
Abu-Daba
b
Shlateen
Living corals Dead corals
0.00
0.50
1.00
1.50
2.002.50
3.00
3.50
4.00
Diversity Evenness
NIOF Sharm El-Naga El-Hmrawin Sharm El-BahariNorth Qula'an Abu-Dabab Shlateen
Figure 4:The percentage cover of living (hard & soft) and dead corals.
Figure 5:The species diversity and evenness index at different sites.
- i=1ln Hs
s Pi Pi
colonies total of Number
(i) species colonies of Number Pi
sln H J
100 XlengthTransect lengthIntercept cover Percentage
CORAL REEFS QUANTITATIVELY ASSESSMENT ALONG THE EGYPTIAN RED SEA COAST
21Thalassas, 28(2) · July 2012
value of hard coral cover (80.25%) and the least soft corals (0.27%), But NIOF recorded the least hard corals (45.87%), while North Qula’an had the maximum soft coral cover that reached 24.84% (Table 3 & figure 4). NIOF and Shlateen sites recorded a high diversity of living organisms (11% and 22.48%) that associated with coral communities.
NIOF area recorded the highest species number and diversity (48 species and 3.54) followed by Abu-Dabab which recorded 35 coral species and its diversity reached 2.84 followed by North Qula’an (34 species and the diversity was 2.54. while Shlateen demonstrated the least diversity (1.97) and the recorded number of species was decreased to 24 (Table 4 and figure 5). On the other hand, the evenness index is the maximum vale at NIOF (0.9) and is related to coral diversity while the minimum value was detected at Shlateen and reached about 0.62.
Acropora humilis, Favites sp., Favia favus, Porites solida, Pocillopora sp. and Stylophora pistillata are the most frequent and repetitive hard species along the studied sites; while, Sarcophyton sp and Sinularia sp. are the most common soft corals.
Data analysis:
On the other hand, one way ANOVA illustrated that, there are no significant differences between the different sites and their diversity and evenness index (Table 5). Whenever, the cluster analysis illustrated that, there are two clusters, the first concerned with the number of species at the studied sites. This cluster (Figure 6) pointed out the similarity among El-Hmrawin, El Sharm El-Bahari, Sharm El-Naqa and Shlateen as they have an equal or sub-equal numbers. Another similarity between Abu-Dabab and North Qula’an; while NIOF has
Dendrogram using Average Linkage (Between Groups)
Rescaled Distance Cluster Combine
Figure 6:Growth of Heterosigma akashiwo exposed to various fractions of Skeletonema costatum filtrate fractionated by solid-phase extraction
and to corresponding negative controls (modified SWM-3).
Position Depth
Temperature
Salinity
pH Site Latitudes Longitudes
NIOF 27 17' 13" N 33 46' 43" E 3 29.32 41.7 7.89 Sharm El-Naqa 26 53' 39" N 33 58' 22" E 8 27.34 40.54 8.14 El-Hmrawin 26 15' 15" N 34 12' 10" E 4.5 27.11 40.24 8.09 El Sharm El-Bahari 25 52' 04" N 34 24' 57" E 6 31.54 40.74 8.3 Abu-Dabab 25 20' 19" N 34 44' 26" E 8 26.57 40.53 8.14 North Qula'an 24 21' 35" N 35 17' 47" E 5 32.84 41.23 8.3 Shlateen 23 09' 10" N 35 36' 58" E 4 30.26 41.12 8.2
Table 1:The latitude, longitude and the some oceanographic parameters at the studied sites.
TAREK ABDEL-AZIZ A. MOHAMMED
22 Thalassas, 28(2) · July 2012
Specis NIOF
Sharm El-
Naqa El-
Hmrawin
El Sharm
El-Bahari
Abu-Dabab
North Qula'an Shlateen
Har
d C
oral
s
Acropora humilis 2.35 3.5 6.85 7.78 8.65 0.00 3.65 A. squarrosa 0.33 0 2.5 1.80 3.15 3.57 0 A. hemiprichi 0.65 2.2 2.25 1.35 0.00 2.66 3.61 A. pharaonis 0.55 2.5 6.4 1.43 0.00 0.00 0 A. cytherea 1.32 3.5 2.85 2.58 0.00 0.00 0 A. digitifera 0.00 1.2 3.65 1.83 2.75 0.00 1.71 A. clathrata 1.22 0 3.25 4.88 0.00 0.00 0 A. valida 0.00 0 0 3.05 1.55 0.00 0 A. venosa 0.00 0 0 1.13 0.00 0.00 0 A. granulosa 0.12 0 0.76 0.00 0.00 0.00 0 Acropora sp. 0.68 0 0 1.33 2.10 0.00 0 Echinopora fruticolosa 2.50 4.2 0 0.00 0.00 3.36 3.21 Echinopora lamellosa 0.00 2 0 0.00 0.00 1.88 0 Favites sp 0.98 0 0 0.00 1.75 1.32 0 Favites peresi 0.42 0.5 1.55 3.85 2.15 4.11 2.56 Favites flexuosa 0.35 0 0 0.00 0.00 1.22 1.02 Favia laxa 0.00 1.2 0.9 1.13 1.20 3.45 2.84 Favia speciosa 1.65 0 1.3 0.00 0.00 0.33 0 Favia favus 2.35 2.3 1.15 2.58 1.45 0.00 5.45 Galaxea fascicularis 3.99 0 0 0.00 0.25 0.66 0 Goniastrea pectinata 0.39 0 0 0.00 1.68 1.75 0 Hydnophora exesa 0.00 0 0 0.00 0.33 0.00 0 Lobophyllia corymbosa 2.35 0 0 0.00 0.00 2.42 2.48 Millipora sp 0.00 0 0 0.00 0.00 0.12 0.22 Millipora dichotoma 1.38 5.8 1.1 3.30 9.15 0.00 1.02 Millipora platyphylla 0.00 5.7 0 0.73 1.02 0.00 3.11 Montipora sp 1.65 0 0 0.78 0.32 0.22 1.39 M. venosa 1.03 0 0 0.00 0.00 0.00 5.45 M. spongeosa 0.66 0 0 0.00 0.00 0.00 3.87 Porites solida 2.58 5.8 12.5 5.38 9.75 6.28 3.2 Porites lutea 0 7.8 3.65 0.00 7.35 2.45 0 Porites columna 0 0 0 0.00 7.55 0.00 0 Pocillopora demicornis 0 3.2 3.58 15.60 2.55 0.00 0 Pocillopora verricusa 0.92 3.3 11.3 13.28 3.56 0.00 0 Pavona explanulata 0 0.7 0 1.28 0.00 2.36 0 Pavona decussate 0 0 0 0.00 0.00 1.21 0 Platygyra deadelia 4.11 0 0 2.55 3.15 5.15 4.64 Platygyra lamellina 0 0 0 0.00 0.00 1.75 0 Plesiastrea versipora 0 0 0 0.00 0.51 0.00 0 p q Q
Har
d C
oral
s
Stylophora pistillata 8.18 4.2 1.35 1.18 3.35 4.98 13.28 Styllophora wellsi 0.62 0.00 0 0.00 1.35 1.21 3.21 Seriatopora histrix 1.21 0.00 0 0.00 0.22 2.05 0 Siderastrea savignyana 0 0.00 0 0.14 0.13 1.88 0 Turbinaria mesenterina 0.17 2.1 0 0.00 0.00 2.94 0 Cosenaria 0.00 0 0 1.22 2.25 0.00 0 Cycloseries sp 0.00 2.2 0 0.00 0.00 0.00 0 Cycloseries marginata 0.23 0 0 0.00 0.00 0.89 1.44 Ctenactis echinata 0.38 2.83 0.25 0.00 0.33 0.00 0 Fungia fungites 0.55 0.12 0.00 0.09 0.00 0.00 1.85
Table 2:The percentage cover of coral species at the studied sites.
CORAL REEFS QUANTITATIVELY ASSESSMENT ALONG THE EGYPTIAN RED SEA COAST
23Thalassas, 28(2) · July 2012
a significant difference due to higher recorded number of diverse species. The second cluster is representing the species diversity and evenness index at the surveyed sites which haven’t significant differences.
DISCUSSION
The Red Sea is biogeographically divisible into Northern, Central and Southern regions, where the central has the greatest concentration of coral reefs and the highest diversity of hard corals (Sheppard et al., 1992 and Veron 1995). Shlateen area is affected by a high density of over-fishing processes and the reject water of the desalination plant that present in the area. These factors are leading to increase the turbidity and settlement of sediment on the coral communities which causes the over growth of macroalgae on corals and overgrowth of many sponge species in the area. So, the species diversity decreased (1.97) in the area and percentage cover deacreased to 70.99%. On the other hand, the NIOF site is not affected
by sedimentation processes resulted from the previous landfilling processes north Hurghada and the interaction between many factors as temperature and direct exposure to sun light (Mohammed and Mohamed, 2005). These reasons are the main factors affect the coral cover causing their decrease to about 66.23%, while the species diversity was the maximum values and recorded 3.54 compared to Shlateen which recorded 1.97. This may be attributed to many factors affecting these areas, such as landfilling and sedimentation processes (at NIOF) and overfishing processes and sedimentation due to turbidity resulting by the reject water of the desalination plant (at Shlateen). This is in agreement with Rogers (1990); Mergner et al. (1992); Ammar and Nawar (1998); Mohammed (2003); Thomas et al. (2003). Mohammed and Mohamed (2005) were illustrated that the high sedimentation and turbidity rates in the sheltered areas of the northern Red Sea increases the particulate sediment over and around the coral communities, subsequently the coral distribution and biodiversity are degraded.
Specis NIOF
Sharm El-
Naqa El-
Hmrawin
El Sharm
El-Bahari
Abu-Dabab
North Qula'an Shlateen
Soft
Cor
als
Alcyonium sp 1.50 0.08 0 0.00 0.00 0.00 0 Heteroxenia fuscescens 1.32 0 2.75 0.00 0.00 2.14 0 Lobophytum pauciflorum 1.17 4.8 0.65 0.00 3.79 0.00 0 Nephthea gracilima minor 0.17 0 1.15 0.11 1.65 0.00 0 Nephthea molli 0.00 3.5 0 0 0.90 0 0 Nephthea sp 0.67 0 0.9 0 0.24 1.5 0.11 Sarcophyton glaucum 2.32 0 0.75 0.16 0.00 10.18 0.65 Sarcophyton spongiosum 0.17 0 0 0.00 0.00 2.88 0 Sarcophyton sp. 3.12 0 0 0.00 0.00 0.00 0 Sinularia polydactyla 3.34 0 0.95 0.00 3.49 2.13 1.02 Sinularia leptoclados 1.54 6.67 0 0.00 0.00 1.02 0 Sinularia gardineiri 1.18 0 0 0.00 0.00 2.44 0 Tubipora musica 0.00 1.3 0.5 0.00 1.23 0.00 0 Xenia macrspiculata 1.31 0 0.75 0.00 0.65 2.55 0 Anthella simplex 0.43 0 0 0 0 0 0 Paralemnalia thyroides 0.32 0 0 0 0 0 0 Capenella fungiformis 0.39 0 0 0 0 0 0 Cladiella sp. 0.51 0 0 0 0 0 0 Nephthea chabrolli 0.90 0 0 0 0 0 0
Dead corals 13.09 8.35 6.75 2.59 6.10 7.08 5.13 Rocky bottom 0.33 2.11 11.69 4.38 0.87 1.25 0.18 Sandy bottom 9.35 2.55 0.94 6.88 0.18 0.88 1.22 Echinodermis 2.31 0.12 1.6 1.12 0.00 0.00 0.19 Algae 7.54 0.74 1.23 1.25 0.00 0.00 7.49 Tridacna 0.11 2.35 2.25 3.26 1.35 2.75 0.22 Sponge 0.00 0 0 0.00 0.00 0.00 14.58 Black Sponge 1.04 0.58 0 0.00 0.00 2.98 0
Continue of Table 2:The percentage cover of coral species at the studied sites.
TAREK ABDEL-AZIZ A. MOHAMMED
24 Thalassas, 28(2) · July 2012
On the other hand, Abu Dabab area recorded the maximum coral cover (91.50%) as it is managed by environmental protectorate affaires agency which mainly act to protect the marine environment and coral communities, where its diversity reached to 2.84. Sharm El-Naqa and Sharm El-Bahri recorded a sub-equal values of coral cover (83.20% and 80.52% respectively), where they are relatively affected by tourist activities that may slightly affect the species diversity and reached to 2.92 (at Sharm El-Naqa) and 3.03 (at Sharm El-Bahri), where Mohammed et al (2009) illustrated that, the tourist activities and anthropogenic impacts affect the coral distribution, diversity and coverage percent.
El-Hamrawin area is affected by phosphate harbor, mining processes and overfishing processes that may affect the coral cover and species diversity where the living corals covered about 75.54% of the area and their diversity reached 2.68. This is in agreement with Mohammed (2003), Ammar (2004), Mohammed (2006) and Mohammed et al., (2009); where, they illustrated that these activities are completely destroyed the benthic communities in the shallower areas. North Qula’an area is away from the coastal human affects but lies under the effect of the direct floods from the active valleys. In spit of this factor, but the coral cover reached to 85.06% and its diversity is 2.54. This site is protected by a dense area
Table 3:The percentage cover of different taxa at the studied sites.
Table 4:Summery of the statistical analysis of the surveyed communities at the different stations.
Table 5:One way ANOVA of the coral diversity of the studied sites.
Taxa
NIO
F
Shar
m E
l-Naq
a
El-H
mra
win
El-S
harm
El-B
ahar
i.
Nor
th Q
ula'a
n
Abu
-Dab
ab
Shla
teen
Hard corals 45.87 66.85 67.14 80.25 60.22 79.55 69.21 Soft corals 20.36 16.35 8.40 0.27 24.84 11.95 1.78 Dead corals 13.09 8.35 6.75 2.59 7.08 6.10 5.13 Sand and rocky bottom 9.68 4.66 12.63 11.26 2.13 1.05 1.40 Other living things 11.00 3.79 5.08 5.63 5.73 1.35 22.48
Station
NIO
F
Shar
m E
l-Naq
a
El-H
mra
win
El-S
harm
El-B
ahar
i.
Nor
th Q
ula'a
n
Abu
-Dab
ab
Shla
teen
Diversity 3.54 2.92 2.68 3.03 2.54 2.84 1.97 Evenness 0.9 0.89 0.81 0.92 0.72 0.68 0.62 colony no. 124 105 97 119 96 86 53 sp. No. 48 27 27 27 34 35 24 Hard coral species 31 22 19 25 26 28 21 Soft coral species 17 5.00 8.00 2.00 8.00 7.00 3.00
Sum of Squares df Mean Square F Sig. Between Groups 148.691 6 24.782 0.078 0.998 Within Groups 4452.996 14 318.071
Total 4601.687 20

CORAL REEFS QUANTITATIVELY ASSESSMENT ALONG THE EGYPTIAN RED SEA COAST
25Thalassas, 28(2) · July 2012
of mangrove plant which protects the coral communities from the flood water of the valley.
The difference among the studied sites may be affected by many other natural factors as the difference in the geographic distribution as well as bottom topography (Kotb et al., 2001; Ouillon et al., 2004) and geomorphology of the sites (Bak, 1975; Adjeroud et al., 2000; Andrefouet and Guzman, 2005) and the interaction between physical and biological factor (Mohammed, 2006) that influenced the distribution, zonation and diversity of corals, as well as the interaction between physical and biological factors influences the identity, distribution and abundance of coral species and macro-benthic organisms in the area; moreover, longitudes and latitudes may be another factor affect the coral distribution and diversity. Finally, the differences in coral diversity and evenness index between the different localities can be attributed to the interactions between the environmental conditions in the different sites such as surface temperature, salinity, dissolved oxygen and turbidity (Sheppard et al., 1992), where the temperature is ranging between 27°C -29°C at different sites and lies in the range 26°C -32°C pointed out by Klein et al. (1997). There are no significance differences between the diversity in the different sites using ANOVA test.
CONCLUSION
1. Anthropogenic activities (landfilling and over-fishing) are responsible for decreasing the coral community’s distribution, diversity and number of species at most localities as NIOF and Slateen.
2. The bottom topography, geomorphology, geographic distribution, longitudes and latitudes are major factors controlling the coral distribution and differences in diversity of species and their numbers.
3. Competition as well as complex interaction between biotic and abiotic factors are another factors influencing the corals distribution and diversity.
4. Acropora humilis, Favites sp., Favia favus, Porites solida, Pocillopora sp., Stylophora pistillata, Sarcophyton sp and Sinularia sp. are the most common and frequent coral species along the Red Sea Coast.
ACKNOWLEDGEMENTS
I’d like to appreciate all my colleges who helped me in the trip and Strategy of the NIOF Red Sea Branch for completing this work.
REFERENCES
Abou Zaid, M.M. and Kotb, M.M.A. (2000). Human and natural induced impacts to the Egyptian Red Sea reefs. Int. Symp.
on the extent of coral reef bleaching 5-9 February, Riyadh, Saudi Arabia. Pp. 1-10.
Adjeroud, M; Andréfouët, S.; Payri, C. and Orempüller, J. (2000): Physical factors of differentiation in macrobenthic communities between atoll lagoons in the central Tuamotu Archipelago (French Polynesia). Mar. Ecol. Prog. Ser. (196): 25-38.
Ammar, M. S. A. (2003): Quantitive assessment of coral communities in two different lagoons near Hurghada, Red Sea, Egypt. J. Egypt. Acad. Soc. Environ. Develop., (D-Environmental studies). 4 (1): 1-18.
Ammar, M. S. A. (2004): Zonation of coral communities and environmental sensitivity offshore a resort site at Marsa Alam, Red Sea, Egypt. Egypt. J. Zool., 42: 76-81.
Ammar, M. S. A. and Nawar, A. H. (1998): Quantitative study for the distribution of reef-building corals at Abu-Galawa, Hurghada, Red Sea. International conference protection is a must. Euro-Arab cooperation center. Inter. Scie. Assoc. 222-233.
Andréfouët, S. and Guzman, H. M. (2005): Coral reef distribution, status and geomorphology-biodiversity relationship in Kuna Yala (San Blas) archipelago, Caribbean Panama. Coral Reefs. 24: 31-42.
Babcock, R. and Davies, P. H. (1991): Effect of sedimentation on settlement of Acropora millepora. Coral Reefs. (9): 205-208.
Bak, R. P. M. (1975): Ecological aspects of the distribution of reef corals in the Netherlands Antilles. Bijdragen Tut de Dierkunde, 45 (2), 57-61.
Benayahu, Y. and Loya, Y. (1981): Competition for space among coral-reef sessile organisms at Eilat, Red Sea. Bull. Mar. Sci. 31: 514-521.
Cray, L.R. (1931): Studies on the coral reefs of Tutuila, American Samoa with special reference to the Alcyonaria. Pap. Dept. Mar. Biol. Carnegie Inst. Wash 27: 53-98.
Crossland, C. (1938): The coral reef at Ghardaqa, Red Sea. Proc. Zool. Soc. London Ser. A 108: 513-523.
Daby, D. (2003): Effects of seagrass bed removal for tourism purposes in a Mauritian bay. Environ. Poll. 125: 313-324.
Dai, C. F. (1990): Interspecific competition between Taiwanese corals with special reference to interactions between alcyonaceans and scleractinians. Mar. Ecol. Prog. Ser. 60: 291-297.
Dinesen, Z. D. (1983): Patterns in the distribution of soft corals across the Central Great Barrier Reef. Coral Reefs, 1: 229-236.
English, S.; Wilkinson, C. and Baker, V. (1997): Survey manual of tropical marine resources. 2nd Edition, Australian institute of Marine Science, Townsville. 385pp.
Fabricius, K. E. (1997): Soft Coral abundance on the Central Great Barrier Reef: effects of Acanthaster planci, space availability, and aspects of the physical environment. Coral Reefs, 16: 159-167.
Fabricius, K. and Alderslade, P. (2001): Soft corals and sea fans: A comprehensive guide to the shallow-water genera

TAREK ABDEL-AZIZ A. MOHAMMED
26 Thalassas, 28(2) · July 2012
of the Central West Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science, Townsville, Australia. P. 264.
Fishelson, L. (1970): Littoral Fauna of the Red Sea; the Population of non- Scleractinian Anthozoans of shallow waters of the Red Sea, Marine Biology 6: 106-116 P.
Fishelson, L. (1973): Ecological and Biological phenomena influencing coral-species composition on the reef table at Eilat (Gulf of Aqaba, Red Sea). Mar. Biol. 19: 183-196.
Gohar, H. A. F. (1940): Studies on the Xeniidae of the Red Sea. Publ. Mar. Biol. Stn. Ghardaqa, Red Sea, Egypt 2: 25-118.
Klein, R.; Tudhope, A. W.; Chilcott, C. P.; Patzold, J.; Abdulkarim, Z.; Fine, M.; Fallick, A. E.; and Loya, Y. (1997): Evaluating southern Red Sea corals as a proxy record for the Asian monsoon. Earth Planet. Sci. Lett. 148: 381-394.
Kotb, M. M. A. (1996): Ecological and biological studies on the coral reefs at southern Sinai coasts, Red Sea, Egypt. Ph. D. Thesis., Faculty of Science, Suez Canal Univ., 174pp.
Kotb, M. M. A. (2001): Growth rates of three reef-building coral species in the northern Red Sea, Egypt. J. Aquat. Biol. And Fish. Vol. 5 (4): 165-185.
Kotb, M.M.A.; Abou Zeid, M. M. and Hanafy, M. H. (2001): Overall evaluation of coral reef status along the Egyptian Red Sea Coast. Biol. Mar. medit. 8 (1): 15-32.
Loya, Y. (1972): Community structure and species diversity of hermatypic corals at Eilat, Red Sea. Int. J. of life in Oceans and coastal waters. Marine Biology. 13, 100-123.
Macfadyen, L. M. (1929): Great Barrier Reef Expedation, Alcyonaria, volume (V) No 1, 19-71 P.
Mergner, H. and Schuhmacher, H. (1981): Quantitative analyse der korallenbesiedlung eines vorriffareals bei Aqaba (Rotes Meer). Helgol Wiss Meeresunters, 34: 337-354.
Mergner, H.; Schuhmacher, H. and Kroll, D. K. (1992): Long-term changes in the coral community of a fore reef area near Aqaba (Red Sea): 1987-1989. Proc. 7th Int. Coral Reef Symp. Guam. 1: 104-113.
Mohammed, T. A. A. (2003): Study of growth and reproduction of some corals at Hurghada region with reference to the effect of some pollutants in the area. Zool. Dept. Faculty of Science. Suez Canal University. 204pp.
Mohammed, T. A. A. (2006): Evaluation, distribution and the
coral diversity in some coastal lagoons, Red Sea, Egypt. Egyptian Journal of Aquatic Research. 32, special issue: 180-195.
Mohammed, T. A. A. and Mohamed, M. A. (2005): Some Ecological Factors Affecting Coral Reef Assemblages Off Hurghada, Red Sea, Egypt. Egyptian Journal of Aquatic Research. Vol. 31 (1): 133-145.
Mohammed, T. A.; Shoukr, F. A.; El-Komi, M. M.; Ezz El-Arab, M. A. H. (2009): Distribution and diversity of alcyonacean soft corals and scleractinian hard corals in the northern Red Sea, Egypt. J. Egyp. German Soci. Zool. Vol. 58D: 67- 83.
Ouillon, S; Douillet, P. and Andréfouët, S. (2004): Coupling satellite data with in situ measurements and numerical modeling to study suspended-sediment transport: a study for lagoon of New Caledonia. Coral Reefs. 23: 109-122.
Pielou, E. C. (1966): The measurement of diversity in different types of biological collections. J. Theor. Biol. 13: 131-144.
Rogers, C. S. (1990): Responses of coral reefs and reef organisms to sedimentation. Mar. Ecol. Prog. Ser. , 62 (1-2): 185-202.
Scheer, G. (1971): Coral reef and coral genera in the Red Sea and Indian Ocean. Symposia of Zoology Society of London. 28: 329-367.
Shannon, C.E. and Wierner, W. (1948): The mathematical theory of communication. Illinois Univ., Urbana, 117pp.
Sheppard, C. R. and Sheppard, A. L. S. (1991): Corals and coral communities of Arabia. II. Fauna of Saudia Arabia, 12, 170 pp.
Sheppard, C.; Price, A. and Roberts, C. (1992): Marine ecology of the Arabian Region. Academic Press, New York. 359 pp.
Thomas, S.; Ridd, P.V. and Day, G. (2003): Turbidity regimes over fringing coral reefs near a mining site at Lihir Island, Papua New Guinea. Mar. Poll. Bull., 46: 1006–1014.
Thomson, J. A. and Dean, L. M. I. (1931): Alcyonacea of the SIBOGA Expedition, volume (XIIId) 1-227 P.
Veron, J. E. N. (1995): Corals in space and time. Cornell University Press, Ithaca, NY, 321Pp.
Veron, J. E. N. (2000): Corals of the World. (3 parts), 477 P. Verseveldt, J. (1982): A Revision of the Genus Sarcophyton Lesson
(Octocorallia, Alcyonacea), Zoologische Verhandelingen Leiden, No 192:1- 91.
(Received: March, 25, 2011; Accepted: April, 9, 2012)

EFFECT OF DESALINATION PLANTS ON THE MARINE ENVIRONMENT ALONG THE RED SEA, EGYPT.
(CASE STUDY)
ABSTRACTMost coastal areas of the Red Sea have a rapid touristic and urban growth; this development is sustained by an increasing
number of seawater desalination plants in the regions which satisfy the growing demand for fresh water. Al Yosr desalination plant was situated at Hurghada and considered a one of the main sources of Hurghada’s growing demand for fresh water. This desalina-tion plant depends on sea water with salinity value varies between 40‰ and 41‰ to produce potable water with total dissolved salts between 300-500ppm, which means that the reject water has salinity rate from 50‰ to 68‰.
The reject water is characterized by increased salinity and high temperature. It additionally contains substantial amounts of chemical pollutants, such as chlorine (which is used for biofouling control in the plants), antiscalants (which are used for scale inhibition) and heavy metals (which are present due to corrosion). So, the heavy metals tend to enrich in sea water than in the sediments, also the carbonate content decreases with increase organic matter. It is estimated that the reject water resulting from this plant will cause an increase of up to 1.5% in mortality for planktonic larvae in an area extending as far away as discharge pipelines. The severity of these effects differs in different areas according to: a) the hydrogeological nature of the marine body (bathymetry, depth, tides, waves, currents); b) the biological sensitivity of the marine habitat; c) the type of desalination plant, its size, the required secondary structures and infrastructure.
The effluent from desalination plant is a multi-component waste, with multiple effects on water, sediment and marine organ-isms. The discharge point must be extends to the end of the back reef zone and below the reef slope (water depth will reaches more than 20m), away from the navigation paths, and away from any benthic communities. It is necessary that all relevant issues, including the seawater intakes, the concentrate and chemical discharges should be continuously monitoring in order to investigate and minimize negative impacts.
(1) National Institute of Oceanography and Fisheries, Red Sea Branch, Egypt. [email protected]
(2) National Institute of Oceanography and Fisheries, Alexandria Branch, Egypt. [email protected]
Thalassas, 28(2) · July 2012: 27-36An International Journal of Marine Sciences
Key words: Desalination plants, Brine water, Marine organisms, Environmental impact, Red Sea.
MOHAMED ABD EL WAHAB(1) & AMR ZAKRIA HAMODA(2)
27Thalassas, 28(2) · July 2012
Palabras clave: Planta desalinizadora, agua salada, organismos marinos, impacto medioambiental, Mar Rojo.
RESUMEN (Efecto de plantas de desalinización sobre el ambiente marino a lo largo del Mar Rojo, Egipto -estudio de caso-)La mayor parte de las zonas costeras del Mar Rojo presentan rápido crecimiento urbano y desarrollo turístico, lo que hace necesario un número creciente de plantas de desalinización de agua de mar en esta región para satisfacer la creciente demanda de agua dulce. La planta desalinizadora Yosr se ubicó en Hurghada, siendo una de las principales fuentes para satisfacer la creciente demanda de agua dulce de esta zona. Este planta de desalinización se surte de agua de mar cuya salinidad varía entre el 40 ‰ y 41 ‰ para producir agua potable agua con sales disueltas entre 300-500ppm, lo que significa que el agua de desecho tiene tasas de salinidad entre 50 ‰ y 68 ‰. El agua de desecho se caracteriza por aumento de la salinidad y alta temperatura. Además contiene cantidades sustanciales de con-taminantes químicos como el cloro (Que se utiliza para control de biofouling en las plantas), antiincrustantes (que se utilizan para la inhibición de la incrustación) y metales pesados (que están presentes debido a la corrosión). Así, los metales pesados tienden a enriquecerse en el agua de mar más que en los sedimentos, también disminuye el contenido de carbonato con aumento de la materia orgánica. Se estima que el agua de desecho resultante de esta planta causará un aumento de hasta el 1,5% en la mortalidad de larvas planctónicas en una zona que se extiende tan lejos como las tuberías de descarga. La gravedad de estos efectos difiere en diferentes áreas de acuerdo a: a) la naturaleza hidrogeológica de los cuerpos de agua (batimetría, profundidad, mareas, olas, corrientes), b) la sensibilidad biológica del hábitat marino; c) el tipo de la planta desalinizadora, su tamaño, las estructuras secundarias necesarias y la infraestructura.El efluente de la planta desalinizadora es un desecho con múltiples componentes, con múltiples efectos sobre el agua, los sedimentos y los organismos marinos. El punto de descarga debe extenderse hasta el extremo de la zona de arrecife posterior y por debajo de la pendiente del arrecife (la profundidad del agua llega a más de 20 m), lejos de las rutas de navegación, y lejos de cualquier comunidad bentónica. Es necesario que toda la información pertinente a estas cuestiones, incluyendo las tomas de agua de mar, el concentrado y las descargas de compuestos químicos deben ser objeto de continua vigilancia con el fin de conocer y minimizar los impactos negativos.

MOHAMED ABD EL WAHAB & AMR ZAKRIA HAMODA
INTRODUCTION Getting access to drinking water is a daily challenge
for more than one billion people in the world; there are more than 7,000 desalination plants worldwide. The Egyptian Red Sea coast extends to about 2025 Km, of this, 1080 Km is Red Sea coast and 945 Km is the coastline of the gulfs of Suez and Aqaba. Most of cities and tourist resorts on the coasts of the Red Sea owned desalination plants discharge their effluent into the sea, which lead to considerable local damage to marine life. At the present time, the Red Sea governorate has 207 hotels and resorts. It is estimated that the discharge of all desalination plants in the Red Sea coast amounts to a waste water flow of about 1,000 m3 per second (Information bank of Red Sea Governorate). Water desalination processes offer various environmental benefits (related to sanitation, water softening, quality of sewage effluents), but the process is also accompanied by adverse environmental effects on the most biologically diverse and productive habitats.
Al Yosr desalination plant was set up as a solution to Hurghada’s growing demand for fresh water. It is one of the largest plant in the Red Sea, and the largest in the Hurghada city. Hurghada city is depending on this desalination plant, which provides about 1/3 of the fresh water requirements for Hurghada.
The impact on the marine environment takes place mainly in the vicinity of the concentrated brine discharge pipelines. Even though the concentrated brine contains natural marine ingredients, its high specific weight causes it to sink to the sea floor without prior mixing. In addition, chemicals, which are administered to the water in the pre-treatment stages of the desalination process, may harm the marine life in the vicinity of the pipe’s outlet. The actual placement of the discharge pipelines may also damage sensitive marine communities.
Recently research studies on the Red Sea environment, and tourism projects and their impact on coastal zone are based on the ecological analysis were done by several
28 Thalassas, 28(2) · July 2012
32° 34°22°
24°
26°
28°
30°
0 1 2
150 Km
Hurghada
Safaga
Qusier
Marsa Alam
Sinai
Suez
Ras Mohamed
Shalatin
Al Yosr Desalination Plant
Scale 1: 500000
Figure 1: Location map of Al Yosr desalination plant, Hurghada, Red Sea.

EFFECT OF DESALINATION PLANTS ON THE MARINE ENVIRONMENT ALONG THE RED SEA, EGYPT. (CASE STUDY)
authors, such as Hawkins and Roberts (1997); UNEP (1997); GEF (1998); Reeve et al. (1998); Reigl and Luke (1998); Hodgson (1999); Jameson et al. (1999); Shackley (1999); Kotb et al. (2001); Abd El-Wahab and El-Sorogy (2003); Cesar (2003); Wielgus et al. (2004); Dar and Abd El Wahab (2005); Farouk (2005); Kondolf et al. (2005); and Shaalan (2005).
This paper throws light on the effects of brine on the marine organisms inhabiting the sandy substratum in the marine area off the desalination plant and discusses the potential impacts of seawater desalination activity on the marine environment.
MATERIALS AND METHODS
The present study was carried out in the marine area off Al Yosr desalination plant in Hurghada City during July 2006. This is the largest seawater desalination facility in the Red Sea, producing 5000 m3/day of potable water. Ten surface sediment samples were collected by Scuba diving, while five sea water samples were collected using water sampler. Also, data on physico-chemical and biological aspects were measured and collected during field survey using Hydrolab Instrument (Surveyor 4).
In the laboratory; the sediment samples were washed with fresh water several time to remove any soluble salts and then dried in the sunlight to avoid any change in their physical and chemical properties. The grain size analysis was done using sieves arrange at 1Φ interval according to (Folk and Ward, 1957).
Ten grams of each sample were ground using agate mortar, passed through a 80 mesh sieve to determinate total carbonate content, (Basaham and El-Sayed 1998) and total organic matter, (Bernner and Binford 1988). Also, about 1.5 liters of each water sample were filtrated using 0.45 μm Millipore filter to determine the trace metal contents in the seawater.
The heavy metals (Cu, Pb, Zn, Cd, Mn & Fe) in sediment and sea water samples were determined using the AAS technique by GBC-932 Ver 1.1. In the sediment samples; the concentrations of the different cations have been determined after the complete digestion of 0.5gm of the powdered samples in a 10ml of hot HF:HNO3:HOCl acid mixture (Chester et al. 1994). While in sea water samples the heavy metals were determined according to Martin (1972), where this technique depends on catching the cations within ammonium pyrrolidine dithiocarbamate (APDC) and methyl isobutyl ketone (MIBK) complex and then extracted using 6N HNO3 acid. The extracted solution is digested on hot plate to volatilize the acid and then solved in about 10 ml of de-ionized water for the heavy metal measurements. The measurements accuracy was checked by applying three replicates in each one and the digestion procedures were tested with the reference materials.
RESULTS AND DISCUSSION
General conditions
Al Yosr desalination plant is considered a one of the vital projects for the human settlements and tourist industry in Hurghada City. Therefore, it was built since 1986 on the land belong to Red Sea Governorate between the Hurghada shipyard from the north and new touristic harbour from the south at Sakkala area, (Fig. 1). The desalination plant depends on sea water with salinity value varies between 40‰ and 41 ‰ and produce potable water with total dissolved salts between 300-500ppm, which means that the reject water has salinity rate from 50‰ to 68‰.
The desalination plant consists of 4 osmosis units; each of them includes 6 element membranes. Each membrane contains 20 glass tubes. So, the feed zone is supplying sea water throughout 4 pipelines with 4 inches diameter, which extend through the back reef zone of the
29Thalassas, 28(2) · July 2012
S. No. Depth in m.gravel sand mud
Carbonates % Total organic matter %
1 5 5.00 95.00 0.0 12.74 10.56 2 10 10.73 89.26 0.012 10.97 7.69 3 15 9.75 84.25 6.001 22.31 11.36 4 20 9.22 85.84 4.940 35.00 5.36 5 25 0.01 95.04 4.951 33.23 3.60 6 30 3.17 93.01 3.816 65.32 4.06 7 35 0.75 96.17 3.072 66.56 3.09 8 40 0.13 93.38 6.488 76.43 4.83 9 45 0.31 91.08 8.613 69.35 4.86
10 50 0.25 92.52 7.235 70.25 3.90 Avg. ------ 3.93 91.56 4.51 46.22 5.93
Sediment Types (%)
Table 1: Grain size analysis, carbonate content and total organic matter of samples collected from marine area off Al Yosr desalination plant.

MOHAMED ABD EL WAHAB & AMR ZAKRIA HAMODA
30 Thalassas, 28(2) · July 2012
sea to about 10m depth. The feed water is pumping by three suction pumps with 70 bar pressure. Each pump has suction capacity 156 m/h. The brine water was discharged throughout 5 pipelines at the beach face.
Geological setting
The beach has inclination angle about 3o with about 4m width, followed by muddy sand tidal flat with gently slope, (Pl. 1), then wide back reef zone covered with biogenic sand. Ten sediment samples were collected and analyzed to give an idea about their texture, conditions of deposition, the nature of the sediments constituents and the main forces controlling the sedimentation processes in the site, (Table1). However, the area is mainly muddy sand bottom with grain size of the sand grades from mud in deeper part to very fine sand near the shoreline.
Biological issue
The marine area off Al Yosr desalination plant can be biologically divided into three zones according to the density and diversity of fauna and flora, (Fig. 2). The first zone lies directly next to the shore and extends for about 25m seaward. It is a very shallow area with the depth ranging from 0.3m to 1.5m and sometimes exposed during the low tide. Its bottom is mostly very fine sand and completely covered with green algae (Enteromorpha). This species covers this entire zone and forms condensed layer over the bottom, (Pl. 2). The fauna are represented with few types of dead gastropods; also this zone is characterized by presence of the five outflow pipelines with diameters 6 inches, (Pl. 3).
The second zone extends to about 150m towards the sea and its depth increase from 1.5m to 15m. Seagrass spots were recorded in this zone in addition to some patches of soft corals.
The third zone is the large extending to about 500m, with max. depth 50m and rich with different fishes type, and soft coral (Sarcophyton hrenbergi and S. eleelegans) and hard one (Acropora eurystoma) Also, several types of invertebrates such as dead mollusks (Tridacna squamosa), echinoderms (Diadema setosum & Holothuria atra.) and sponge (black spongy). The marine flora in this zone is mainly represented by brown and blue green algae and some seagrass spots. Algae is mainly e.g. Chlorophyceae, Digenea simplex and Padina pavonia which cover wide area. Generally, the scarcity of the living corals in the area may be attributed to high sedimentation rate and high percentage cover of filamentous algae due to the high temperature, (Pls. 4-6). It is estimated that the plant will
cause an increase of up to 1.5% in mortality for planktonic larvae in an area extending as far away as desalination plant, (VDP, 2008).
Physico-chemical features
Temperature
In front of Hurghada, the water temperature was ranging between 18.9o C (during January) and 28.9o C (during July), affecting by heating effect of the Sun (El Sman 2000). In extreme shallow areas, higher values were recorded due to solar radiation (Morcos, 1970). Temperature of seawater at the feeding zone off the plant showed moderate fluctuations, whereas surface water temperature was 29o C and bottom water temperature at 50m depth was 26.2o C (Table 2). The area being relatively shallow, the difference between the surface and bottom water temperatures was less than 3o C and the water body was unhomogenous throughout the day because of the continuous movement of water towards the sea.
Salinity
Salinity is increased in the brine stream of all processes, but elevated temperature values are characteristic of desalination plant effluents only. The RO brine has a higher density than seawater as a result of its increased salinity and will mostly affect benthic communities, while desalination plant discharges tend to float on the surface and interfere with productivity in the pelagic community. The positive buoyancy of desalination plant discharges is mostly due to the discharge of large volumes of cooling water, which are blended with the brine. The salinity value around the discharge point recorded 60.0‰ at 5m depth and 50.5‰ at 50m depth (Table 2), while in the open sea the average salinity reaches 40.31‰, 40.06 ‰, 40.58 ‰ and 40.24 ‰ in winter, spring, summer and autumn respectively, (Maine Waters, Red Sea Studied, Report III, 1981).
Specific conductivity
The specific conductivity values in the feeding zone showed narrow fluctuations, its variation over marine area was ranged from 62.0 ms/cm to 62.9 ms/cm, in that the values were the highest at 50m depth and the lowest at 5m depth (Table 2), while the specific conductivity values in Safaga bay fluctuates from 61.5 to 61.73 ms/cm, (Helal & Abd El Wahab, 2004). The conductivity values observed now were converted to salinity. Generally, surface salinity is generally high in shallow areas because of excess evaporation in relation to precipitation, and poor rainfall.

EFFECT OF DESALINATION PLANTS ON THE MARINE ENVIRONMENT ALONG THE RED SEA, EGYPT. (CASE STUDY)
31Thalassas, 28(2) · July 2012
pH
The pH of seawater in the feeding zone remained always alkaline, whereas it ranged from 8.0 to 8.8 (Table 2), while in the open sea it varies from 8.08 to 8.37 (El-Mamony, 1986).
Dissolved Oxygen
The dissolved oxygen concentration in the feeding zone seawater varies between 5.6 and 6.8 mg/L (Table 2). In the open sea, the dissolved oxygen is fluctuated between 6.36 mg/L in winter and 5.28 mg/L in summer (El- Mamony, 1986). With increasing temperature and salinity, oxygen becomes less soluble in seawater. However, oxygen levels are deliberately reduced in desalination plants by physical deaeration and addition of oxygen scavengers like sodium bisulfite to inhibit corrosion. Oxygen depletion is also a problem of the RO brine, as sodium bisulfite is commonly used as a neutralizing agent for chlorine. The lack of dissolved oxygen could be toxic to marine organisms and aeration is recommended prior to oceanic discharge.
Total Dissolved Salts (TDS)
TDS value in feed water of the Al Yosr plant was lowest (39.7 g\L) at 5m depth and highest 40.1 g\L at 50m depth (Table 2). High TDS value is considered as an index of water pollution and it is one of the important water quality criterions for which environmental limits are prescribed by regulatory agencies in many countries. Reason for this elevation can be attributed to the shallow area conditions, the re-suspension of fine grained particles by wave action and the uprooting of seagrass due to consequent turbulence in the sea.
The suspended particle load in the area off the plant greatly influenced by accumulation of garbage and fine particles coming from both the shipyard at the north
ward and tourstic harbour from the south ward. Also, the most important contribution to TDS comes from marine organisms as decomposable organic matter which is continually coming from the navigation of tourstic boats. Therefore, sheltered coastal areas off the plant collect organic matter in the form of detritus which in return decomposes and releases nutrients which subsequently become available to the plankton. There is a strong relationship between suspended particles and marine food chain. The coarse particles in seawater are colonized by fungi, because of larger surface area offered by them, while the finer particles are colonized by bacteria, (Smith, 1985). Macro algae have been cited as an important source of detritus suspended solids in the area, as well as heaps of seagrass were observed in the feeding area during periods of survey.
Carbonates
Carbonate sediments are usually concentrated in warm tropical or subtropical seawater carbonate secreting plants and animals are dominant (El-Mamony, 1986). The total carbonate content in the investigated sediments varies between 10.97% and 76.43% averaging 46.22% (Table 2). Distribution of the carbonate content through the studied sediments show a lesser value of carbonate concentrations relative to the other locations. Strong relation is detected between the carbonate content and organic matter of the sediments in the investigated area, whereas the carbonate content decreases with increase organic matter (Table 2).
Total Organic Matter
Organic matter affects the aquatic ecosystem by interacting with inorganic matter to form complex compounds, which include in its structure several elements. Free carbon dioxide and hydrogen sulfide may be released and affect the composition of the sediments even more
S. N.
Depth (m) Temp. oC DO mg\l Sal. ‰ pH
SP.C. ms\cm TDS g\l
1 5 28.4 5.7 68.0 8.8 62.0 39.8 2 10 27.9 5.8 58.6 8.5 62.1 39.7 3 15 26.8 5.6 56.9 8.4 62.3 39.9 4 20 25.6 6.5 54.8 8.4 62.4 40.0 5 25 25.3 6.6 53.1 8.3 62.4 40.0 6 30 24.7 6.7 52.7 8.3 62.5 40.1 7 35 24.5 6.7 52.3 8.2 62.6 40.1 8 40 24.3 6.7 51.7 8.0 62.7 40.1 9 45 23.8 6.7 51.1 8.0 62.8 40.1 10 50 23.2 6.8 50.5 8.0 62.9 40.1
oC = Temperature, Do = Dissolved oxygen, Sal. ‰ = Salinity, pH = Hydrogen ion concentration, Sp. C. = Specific conductivity, TDS = Total dissolved salts.
Table 2: Physical parameters of water masses off Al Yosr desalination plant, Hurghada.
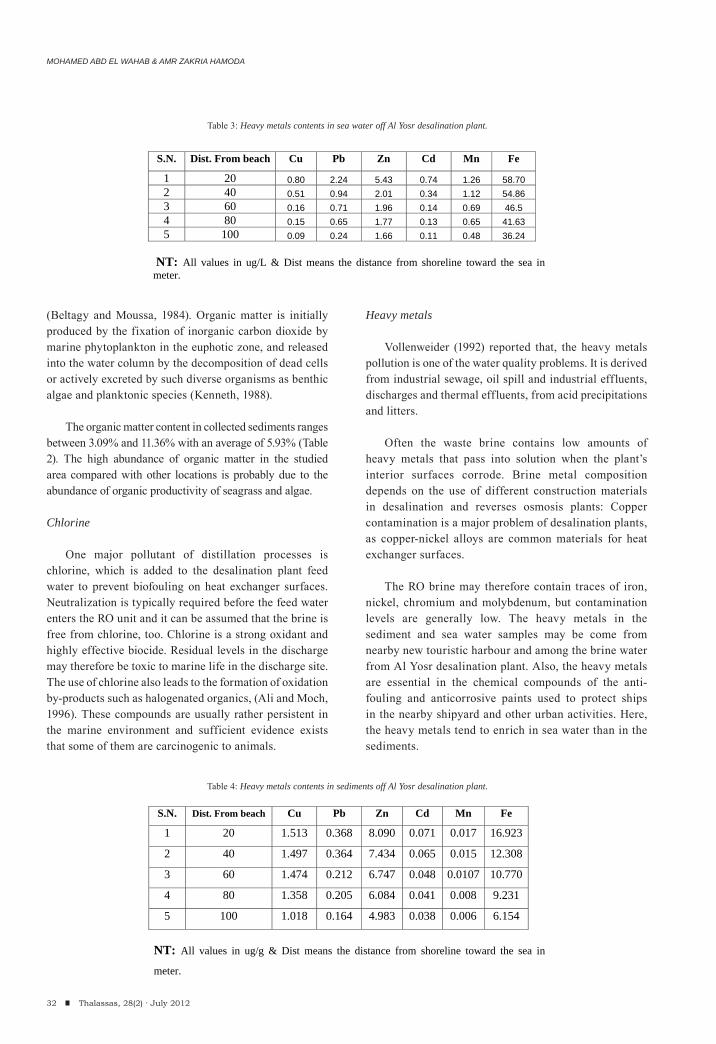
MOHAMED ABD EL WAHAB & AMR ZAKRIA HAMODA
32 Thalassas, 28(2) · July 2012
(Beltagy and Moussa, 1984). Organic matter is initially produced by the fixation of inorganic carbon dioxide by marine phytoplankton in the euphotic zone, and released into the water column by the decomposition of dead cells or actively excreted by such diverse organisms as benthic algae and planktonic species (Kenneth, 1988).
The organic matter content in collected sediments ranges between 3.09% and 11.36% with an average of 5.93% (Table 2). The high abundance of organic matter in the studied area compared with other locations is probably due to the abundance of organic productivity of seagrass and algae.
Chlorine
One major pollutant of distillation processes is chlorine, which is added to the desalination plant feed water to prevent biofouling on heat exchanger surfaces. Neutralization is typically required before the feed water enters the RO unit and it can be assumed that the brine is free from chlorine, too. Chlorine is a strong oxidant and highly effective biocide. Residual levels in the discharge may therefore be toxic to marine life in the discharge site. The use of chlorine also leads to the formation of oxidation by-products such as halogenated organics, (Ali and Moch, 1996). These compounds are usually rather persistent in the marine environment and sufficient evidence exists that some of them are carcinogenic to animals.
Heavy metals
Vollenweider (1992) reported that, the heavy metals pollution is one of the water quality problems. It is derived from industrial sewage, oil spill and industrial effluents, discharges and thermal effluents, from acid precipitations and litters.
Often the waste brine contains low amounts of heavy metals that pass into solution when the plant’s interior surfaces corrode. Brine metal composition depends on the use of different construction materials in desalination and reverses osmosis plants: Copper contamination is a major problem of desalination plants, as copper-nickel alloys are common materials for heat exchanger surfaces.
The RO brine may therefore contain traces of iron, nickel, chromium and molybdenum, but contamination levels are generally low. The heavy metals in the sediment and sea water samples may be come from nearby new touristic harbour and among the brine water from Al Yosr desalination plant. Also, the heavy metals are essential in the chemical compounds of the anti-fouling and anticorrosive paints used to protect ships in the nearby shipyard and other urban activities. Here, the heavy metals tend to enrich in sea water than in the sediments.
S.N. Dist. From beach Cu Pb Zn Cd Mn Fe
1 20 0.80 2.24 5.43 0.74 1.26 58.70 2 40 0.51 0.94 2.01 0.34 1.12 54.86 3 60 0.16 0.71 1.96 0.14 0.69 46.5 4 80 0.15 0.65 1.77 0.13 0.65 41.63 5 100 0.09 0.24 1.66 0.11 0.48 36.24
NT: All values in ug/L & Dist means the distance from shoreline toward the sea in meter.
S.N. Dist. From beach Cu Pb Zn Cd Mn Fe
1 20 1.513 0.368 8.090 0.071 0.017 16.923
2 40 1.497 0.364 7.434 0.065 0.015 12.308
3 60 1.474 0.212 6.747 0.048 0.0107 10.770
4 80 1.358 0.205 6.084 0.041 0.008 9.231
5 100 1.018 0.164 4.983 0.038 0.006 6.154
NT: All values in ug/g & Dist means the distance from shoreline toward the sea in
meter.
Table 3: Heavy metals contents in sea water off Al Yosr desalination plant.
Table 4: Heavy metals contents in sediments off Al Yosr desalination plant.

EFFECT OF DESALINATION PLANTS ON THE MARINE ENVIRONMENT ALONG THE RED SEA, EGYPT. (CASE STUDY)
33Thalassas, 28(2) · July 2012
So, the areas of restricted water exchange and soft bottom habitats could be affected by heavy metal accumulation. Many benthic shells are usually affected by metal accumulations in polluted areas and consequently are important food source (Abd El Wahab, 2003), with the risk that metals are enriched in their bodies and passed on to higher trophic levels.
Antiscalants
Scaling on heat exchanger surfaces, inside tubes, or on RO membranes impairs plant performance. Antiscalants are commonly added to the feed water in both desalination and RO plants to prevent scale formation. The main representatives of antiscalants are organic, carboxylic-rich polymers such as polyacrylic acid and polymaleic acid. Acids and polyphosphates are still in use at a limited scale but on the retreat, (Abdul Azis, et al. 2000). As antiscalants have a low toxicity, the acute environmental risk associated with their release into the marine environment is relatively low.
Coagulants and coagulant aids
Coagulants like ferric or aluminum chloride are used to improve filtration of suspended material from the RO feed water. Coagulant aids (organic substances with high molecular masses that bridge particles further together)
and pH control are supplementary methods to enhance coagulation. The filter backwash can be discharged to the sea, as toxic effects are not expected by coagulants and coagulant aids, (Abdul Azis, et al. 2000). However, possible impacts such as reduced primary production or burial of sessile organisms by increased turbidity in the discharge should be anticipated.
Antifoaming agents
To reduce foaming in desalination plants, antifoaming agents like polyglycols are added to the feed water, which are not toxic but poorly biodegradable, (Winters, 1997 and Jamaluddin, et al. 1999). Adverse effects are not to be expected with regard to a low dosage level and sufficient dilution following discharge.
Cleaning chemicals
Cleaning intervals have to be established for each desalination plant individually and are typically three to six months depending on the quality of the plant’s feed water. In RO plants, alkaline cleaning solutions (pH 11-12) are used for removal of silt deposits and biofilms, whereas acidified solutions (pH 2-3) remove metal oxides and scales. Further chemicals are often added to improve the cleaning process of RO membranes, such as detergents, oxidants, complexing agents or biocides for membrane
Figure 2: Map showing distribution of bottom facies off Al Yosr desalination plant at Hurghada.

MOHAMED ABD EL WAHAB & AMR ZAKRIA HAMODA
34 Thalassas, 28(2) · July 2012
Plate (1). Beach profile off Al Yosr desalination plant. Plate (2). Dense filamentous algae in the intertidal zone off plant.
Plate (3). Five outlets pipelines on the beach off Al Yosr plant.
Plate (4). Polluted filamentous algae in the end of intertidal zone.
Plate (5). Unhealthy soft corals in the marine area off the
desalination plant. Plate (6). High turbidity at the end of back reef zone.

EFFECT OF DESALINATION PLANTS ON THE MARINE ENVIRONMENT ALONG THE RED SEA, EGYPT. (CASE STUDY)
35Thalassas, 28(2) · July 2012
disinfection. In desalination plants, cleaning is typically very simple: Copper-nickel heat exchanger surfaces are washed with acidified warm seawater to remove alkaline scales, (Abdul Azis, et al. 2000). The acidic solution often contains a chemical inhibitor which is added to protect the plant from corrosion. Most of the named cleaning and disinfection chemicals may be hazardous to aquatic life.
POTENTIAL IMPACTS OF SEAWATER DESALINATION (PROBLEM SEVERITY)
The sea as a source of drinking water seems to be unlimited, but pollution by land-based activity often impairs the quality of coastal water bodies that serve as feed water for desalination plants. Hot spots of marine pollution are typically near coastal areas of intense human activity such as hotels, resorts, and harbours, which are also the areas where desalinated water is most needed for tourism activities.
The process of desalination is not preserve environmentally friendly and seawater desalination plants also contribute to the wastewater discharges that affect coastal water quality. This is mostly due to the highly saline brine that is emitted into the sea, which may be increased in temperature, contain residual chemicals from the pretreatment process, heavy metals from corrosion or intermittently used cleaning agents. The effluent from desalination plants is a multi-component waste, with multiple effects on water, sediment and marine organisms. The severity of these effects differs in different areas according to: a) the hydrogeological nature of the marine body (bathymetry, depth, tides, waves, currents); b) the biological sensitivity of the marine habitat; c) the type of desalination plant, its size, the required secondary structures and infrastructure.
PROBLEM RESOLUTION
For impact mitigation, some aspects should be taken in consideration:
1. Alternative biocides include for example ozone and monochloramine, while disinfection with ultraviolet light may be used instead of biocides to eliminate micro-organisms.
2. Several alternative pretreatment methods have been considered to replace chlorine in desalination plants to avoid environmental and health problems.
3. The disposal brine to the sea should be strictly regulated using neutralization of alkaline or acidic solutions and treatment of additional cleaning agents is recommended before discharge to the sea to remove any potential toxicity.
4. The backwash could be diluted, e.g. by continuous blending with the brine, or alternatively it could be removed from the filters and transported to a landfill.
5. The discharge point must be extends to the end of the back reef zone and below the reef slope (water depth will reaches more than 20m), away from the navigation paths, and away from any benthic communities. Therefore, this zone is wavy in all conditions that allow continuous mixing occur and disperses before reaching the marine communities.
6. Use the diffusers at the end of reject pipelines, which provide good mixing with the seawater and dispersing immediately by the waves and currents before reaching the nearest marine communities.
7. It is necessary that all relevant issues, including the seawater intakes, the concentrate and chemical discharges should be continuously monitoring in order to investigate and minimize negative impacts.
8. In contrast, non-metal equipment and stainless steels are typically used in RO plants.
9. Furthermore, we believe that desalination activity needs to become an integrated part of regional and national water management plans in order to identify the best water supply option under environmental, socio-economic, energy and human health criteria.
10. Environmental awareness and preliminary planning can minimize the adverse effects of the desalination process on the environment.
REFERENCES
Abd El-Wahab, M. (2003). Comparative geochemical study on heavy metals of some molluscan shells from Hurghada area, Egypt. Egyptian Journal of Geology, 47)(2): 1139-1151.
Abd El-Wahab, M. and El-Sorogy, A. S. (2003). Scleractinian corals as pollution indicators, Red Sea Coast, Egypt. N. Jb. Geol. Palaon.Mh., Stuttgart, :641-655.
Abdul Azis, P. K.; Al-Tisan, I.; Al-Daili, M.; Green, T. N.; Dalvi, A. I. and Javeed, M. A. (2000). Effects Of Environment On Source Water For Desalination Plants On The Eastern Coast Of Saudi Arabia1 Desalination Journal, 132: 29-40.
Ali, B.H., and Jr. Irving Moch (1996). Desalination and Water Re-use, 6: 40-45.
Basaham, A. S. and El-Sayed, M. A. (1998). Distribution and phase association of some major and trace elements in the Arabian Gulf sediments. Academic Press Limited, 46: 185-194.
Beltagy, A. I. and Moussa, Kh. A. (1984). The distribution of organic matter in the sediments of the Gulf of Suez. Inst. Oceanogr. & Fish., ARE. 10 : 99-109.
Bernner, M. and Binford, M. E. (1988). Relationships between concentrations of sedimentary variables and trophic state in Florida lakes. Can. J. Fish. Aquat. Sci., 45: 294-300.

MOHAMED ABD EL WAHAB & AMR ZAKRIA HAMODA
36 Thalassas, 28(2) · July 2012
Cesar, H. (2003). Economic valuation of the coral reefs of Egypt. Report prepared for the MVE-unit of EEPP, funded by USAID. Preliminary Version, January 2003 (mimeo).
Chester, R.; Lin, F.G. and Basaham, A.S. (1994). Trace metals solid state speciation changes associated with the down-column fluxes of oceanic particulates. Journal of the Geological Society of London. 151: 351-360.
Dar, M. A. and Abd El Wahab, M. (2005). The coastal alterations due to the artificial lagoons, Red Sea (Case Study). Egyptian Journal of Aquatic Research, Vol. 31, Special issue, : 57-68.
El-Mamony, M. H. (1986). Studies on the beach and offshore sediments of Jubal strait area at the entrance of Gulf of Suez, M.Sc. Thesis, Alexandria Univ. Egypt, 126P.
El Samn, M. I. (2000). Hydrographic studies of some lagoons near Hurghada, M.Sc Thesis, South Valley Univ., 156P.
Folk, R. L. and Ward, W. C. (1957). Brazos River bar: a study in the significance of grain size. Jour. Sed. Petrol. 27(1) :3.26, Tulsa, Oklahoma.
Farouk, M. A. (2005). Tourism projects and their impact on coastal zone. Regional Organization for the Conservation of the Environment of the Red Sea and Gulf of Aden. Sea to Sea, Second Regional Forum, 13-16 Feb. 2005, Cairo, :197-205.
Egyptian Red Sea Coast and Marine Resource Management Project (GEF 1998). Integrated coastal zone management action plan for the Egyptian Red Sea. September 1998, Report 6.
Hawkins, J. P. and Roberts, C. M. (1997). Estimating the carrying capacity of coral reefs for recreational scuba diving. In: proc. 8th Int. Coral Reef Symp. 2: 1923-1926.
Helal, S. A. and Abd El Wahab, M. (2004). Recent Ostracodes from Marine sediments of Safaga Bay, Red Sea, Egypt. Egypt. Jour. Paleontol., 4 :75-93.
Hodgson, G., (1999). A global assessment of human effects on coral reefs. Mar. Poll. Bull., 385: 345-355.
Jamaluddin, A.T.M.; Saeed, M.O.; Al-Rowaili, A.R.; Al-Tisan,I; Al-Rasheed, R.; Al-Amri M.M.; Lawrence, P.A. and Kamran, C. (1999). Technical Report No. 3807/APP 95006, SWCC, R&D Center
Jameson, S. C.; Ammar. M. S. A.; Sadalla, E.; Mostafa, H. M.and Riegle, B. (1999). A coral damage index and its application to diving sits in the Egyptian Red Sea . Coral reefs, 18 :333 .339 ـ
Kenneth, P. (1988). Sediment transport and depositional processes. Postgarduate Research Institute for Sedimentology, Univ. of Reading, Whiteknights, UK. 397P.
Kondolf, G. M.; Radke, J. and Ismail, I. (2005). Sea to Sea Second regional Form, 13-16 February 2005, Cairo Egypt.
Kotb, M. M.; Abu Zeid, M. H. and Hanafy, M. H. (2001). Overall evaluation of the coral reef status along the Egyptian Red Sea coast. Biol. Mar. Medit. 8 (1) :15-32.
Marine Waters, Red Sea Studies (1981). Investigation of level and effects of pollutants in saline lakes and littoral marine environments. Publ. Inst. Oceano. & Fish., Report No. III, 256 P.
Martin, D.F. (1972). Marine chemistry, New York: Marcel Dekker Inc.
Morcos, S.A. (1970). Physical and chemical oceanography of the Red Sea. Oceanography and Marine Biology, Annual review, (8) :73-202.
Riegl, B. and Luke, K. E. (1998). Ecological parameters of dynamited reefs in the northern Red Sea and their relevance to reef rehabilitation. Mar. Poll. Bull., 37(8-12): 488-498.
Reeve, S. M.; Jameson, S. C.; Abdel Fattah, R. S.; Riegl, B.; Hassan, R. and Newman, A. P. (1998). Best practices for tourism center development along the Red Sea coast. Tourism Development Authority, Cairo, 107P.
Shaalan, I. M. (2005). Sustainable tourism development in the Red Sea of Egypt threats and opportunities. Journal of Cleaner Production, 13 :83-87.
Shackley, M. (1999). Tourism development and environmental protection in southern Sinai. Tourism Management, 20 :543-548.
Smith, R. L. (1985), Elements of Ecology, Second Edition, Harper and Row Publishers, New York.
United Nations Environmental Program (UNEP) (1997). Regional Seas Reports and Studies No.166 United Nations Environment Program.
Victorian Desalination Project Environmental Effects Statement (2008). Operations impact assessment, (2) Ch 8. p13.
Vollenweider, R.A. (1992).Coastal marine eutrophication: Principles and control. Marine Coastal Eutrophication, Proceedings of an International conference, Bologna, Italy, :1-20.
Wielgus, J; Chadwic-Furman, N. E. and Dubinsky, Z. (2004). Coral cover and partial mortality on anthropogenically impacted coral reefs at Eilat, Northern Red Sea. Mar. Pollu. Bull., 48 :248-253.
Winters, H. (1997). Proc. Intern. Symp. on pretreatment of feed water for RO Desal. Plants, Kuwait, :143-146.
(Received: March, 30, 2011; Accepted: April, 13, 2012)

ANTIFOULING ACTIVITY OF A MARINE EPIBIOTIC BACTERIUM FROM THE SEAWEED Sargassum wightii
Thalassas, 28(2) · July 2012: 37-43An International Journal of Marine Sciences
V. RAJASREE, S. SATHEESH(1,*) & S.G. PRAKASH VINCENT(1)
37Thalassas, 28(2) · July 2012
Key words: Biofouling; Biofi lm; Seaweeds; Extracellular polymeric substance; surface-associated bacteria.
Palabras clave: Bio-incrustantes, biopelícula, algas macrófi tas, sustancia polimérica extracelular, bacterias asociadas a superfi cie.
ABSTRACT
The antifouling activities of a marine bacterium isolated from the surface of the seaweed Sargassum wightii is presented in this paper. The extracellular polymeric substance (EPS) of the bacterial strain showed inhibitory activity against the bacteria involving in biofilm formation. The EPS reduced the adhesion of biofilm forming bacteria on hard surface. The EPS was mixed with varnish and coated onto acrylic coupons. The adhesion assay showed that the number of cells adhered on the control coupons were higher than that the number of cells found on the slides coated with EPS. The antifouling assay conducted in the costal water showed that the EPS of the strain also have inhibitory activity against the settlement of biofouling organisms. Thin-layer chromatographic analysis indicated a single active compound in the EPS. In conclusion, results indicated that the bacteria associated with the seaweeds could provide leads for the development of natural product based antifouling agents.
RESUMEN (Actividad antiincrustante (antifouling) de una bacteria marina epibiótica del alga Sargassumwightii)
Se presentan las actividades anti-incrustantes de una bacteria marina aislada de la superficie de las algas Sargassumwightii. La sustancia extracelular polimérica (EPS) de la cepa bacteriana mostró una actividad inhibitoria contra las bacterias que implican en la formación de biopelículas. La EPS reduce la adherencia de las bacterias formadoras de biopelículas sobre una superficie dura. Las EPS se mezcló con el barniz y envueltas en muestras acrílicas. El ensayo de adhesión mostró que el número de células adheridas en las muestras de control fueron superiores a las que se fijaron en las muestras revestidas con EPS. El ensayo de antiincrustaciónllevado a cabo en el agua costera mostró que las EPS también tienen actividad inhibitoria contra la fijación de los organismos marinos incrustantes. Los análisis de cromatografía en capa fina mostraron un solo com-puesto activo en la EPS. En conclusión, los resultados indicaron que las bacterias asociadas con las algas podrían proporcionar pistas para el desarrollo de agentes antiincrustantes basados en productos naturales.
(1) Department of Marine Biology, Faculty of Marine Sciences, King Abdulaziz University, Post Box No. 80207, Jeddah-21589, Saudi Arabia.
*Corresponding author, e-mail: [email protected]
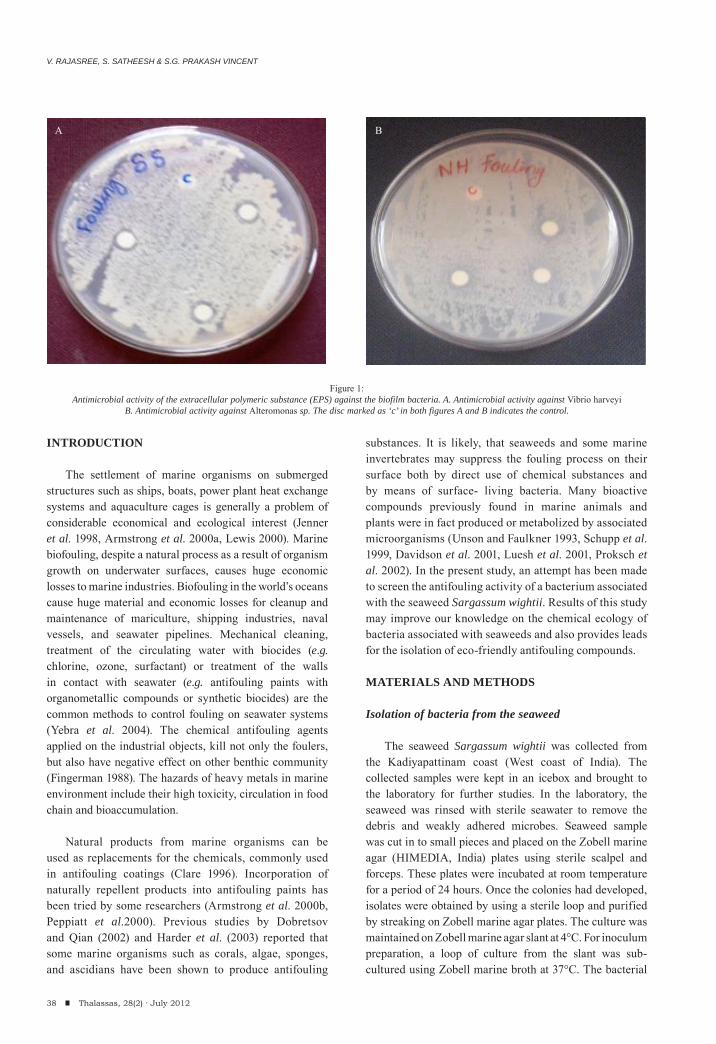
V. RAJASREE, S. SATHEESH & S.G. PRAKASH VINCENT
38 Thalassas, 28(2) · July 2012
INTRODUCTION
The settlement of marine organisms on submerged structures such as ships, boats, power plant heat exchange systems and aquaculture cages is generally a problem of considerable economical and ecological interest (Jenner et al. 1998, Armstrong et al. 2000a, Lewis 2000). Marine biofouling, despite a natural process as a result of organism growth on underwater surfaces, causes huge economic losses to marine industries. Biofouling in the world’s oceans cause huge material and economic losses for cleanup and maintenance of mariculture, shipping industries, naval vessels, and seawater pipelines. Mechanical cleaning, treatment of the circulating water with biocides (e.g. chlorine, ozone, surfactant) or treatment of the walls in contact with seawater (e.g. antifouling paints with organometallic compounds or synthetic biocides) are the common methods to control fouling on seawater systems (Yebra et al. 2004). The chemical antifouling agents applied on the industrial objects, kill not only the foulers, but also have negative effect on other benthic community (Fingerman 1988). The hazards of heavy metals in marine environment include their high toxicity, circulation in food chain and bioaccumulation.
Natural products from marine organisms can be
used as replacements for the chemicals, commonly used in antifouling coatings (Clare 1996). Incorporation of naturally repellent products into antifouling paints has been tried by some researchers (Armstrong et al. 2000b, Peppiatt et al.2000). Previous studies by Dobretsov and Qian (2002) and Harder et al. (2003) reported that some marine organisms such as corals, algae, sponges, and ascidians have been shown to produce antifouling
substances. It is likely, that seaweeds and some marine invertebrates may suppress the fouling process on their surface both by direct use of chemical substances and by means of surface- living bacteria. Many bioactive compounds previously found in marine animals and plants were in fact produced or metabolized by associated microorganisms (Unson and Faulkner 1993, Schupp et al. 1999, Davidson et al. 2001, Luesh et al. 2001, Proksch et al. 2002). In the present study, an attempt has been made to screen the antifouling activity of a bacterium associated with the seaweed Sargassum wightii. Results of this study may improve our knowledge on the chemical ecology of bacteria associated with seaweeds and also provides leads for the isolation of eco-friendly antifouling compounds.
MATERIALS AND METHODS
Isolation of bacteria from the seaweed
The seaweed Sargassum wightii was collected from the Kadiyapattinam coast (West coast of India). The collected samples were kept in an icebox and brought to the laboratory for further studies. In the laboratory, the seaweed was rinsed with sterile seawater to remove the debris and weakly adhered microbes. Seaweed sample was cut in to small pieces and placed on the Zobell marine agar (HIMEDIA, India) plates using sterile scalpel and forceps. These plates were incubated at room temperature for a period of 24 hours. Once the colonies had developed, isolates were obtained by using a sterile loop and purified by streaking on Zobell marine agar plates. The culture was maintained on Zobell marine agar slant at 4°C. For inoculum preparation, a loop of culture from the slant was sub-cultured using Zobell marine broth at 37°C. The bacterial
Figure 1:Antimicrobial activity of the extracellular polymeric substance (EPS) against the biofilm bacteria. A. Antimicrobial activity against Vibrio harveyi
B. Antimicrobial activity against Alteromonas sp. The disc marked as ‘c’ in both figures A and B indicates the control.
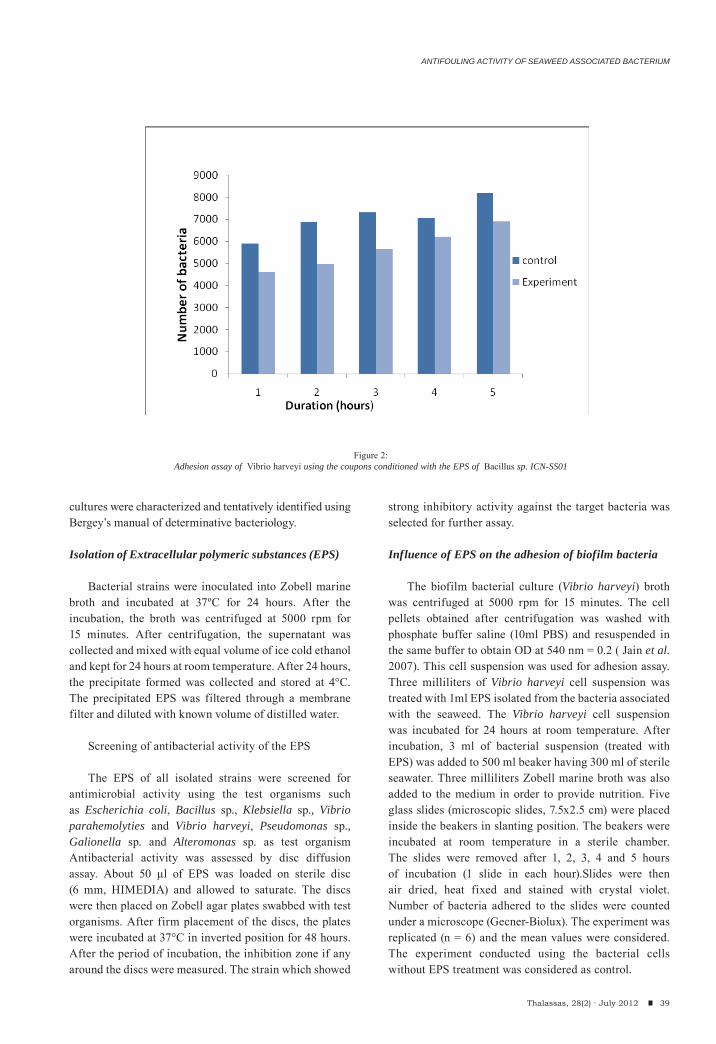
cultures were characterized and tentatively identified using Bergey’s manual of determinative bacteriology.
Isolation of Extracellular polymeric substances (EPS)
Bacterial strains were inoculated into Zobell marine broth and incubated at 37ºC for 24 hours. After the incubation, the broth was centrifuged at 5000 rpm for 15 minutes. After centrifugation, the supernatant was collected and mixed with equal volume of ice cold ethanol and kept for 24 hours at room temperature. After 24 hours, the precipitate formed was collected and stored at 4°C. The precipitated EPS was filtered through a membrane filter and diluted with known volume of distilled water.
Screening of antibacterial activity of the EPS
The EPS of all isolated strains were screened for antimicrobial activity using the test organisms such as Escherichia coli, Bacillus sp., Klebsiella sp., Vibrio parahemolyties and Vibrio harveyi, Pseudomonas sp., Galionella sp. and Alteromonas sp. as test organism Antibacterial activity was assessed by disc diffusion assay. About 50 μl of EPS was loaded on sterile disc (6 mm, HIMEDIA) and allowed to saturate. The discs were then placed on Zobell agar plates swabbed with test organisms. After firm placement of the discs, the plates were incubated at 37°C in inverted position for 48 hours. After the period of incubation, the inhibition zone if any around the discs were measured. The strain which showed
strong inhibitory activity against the target bacteria was selected for further assay.
Influence of EPS on the adhesion of biofilm bacteria
The biofilm bacterial culture (Vibrio harveyi) broth was centrifuged at 5000 rpm for 15 minutes. The cell pellets obtained after centrifugation was washed with phosphate buffer saline (10ml PBS) and resuspended in the same buffer to obtain OD at 540 nm = 0.2 ( Jain et al. 2007). This cell suspension was used for adhesion assay. Three milliliters of Vibrio harveyi cell suspension was treated with 1ml EPS isolated from the bacteria associated with the seaweed. The Vibrio harveyi cell suspension was incubated for 24 hours at room temperature. After incubation, 3 ml of bacterial suspension (treated with EPS) was added to 500 ml beaker having 300 ml of sterile seawater. Three milliliters Zobell marine broth was also added to the medium in order to provide nutrition. Five glass slides (microscopic slides, 7.5x2.5 cm) were placed inside the beakers in slanting position. The beakers were incubated at room temperature in a sterile chamber. The slides were removed after 1, 2, 3, 4 and 5 hours of incubation (1 slide in each hour).Slides were then air dried, heat fixed and stained with crystal violet. Number of bacteria adhered to the slides were counted under a microscope (Gecner-Biolux). The experiment was replicated (n = 6) and the mean values were considered. The experiment conducted using the bacterial cells without EPS treatment was considered as control.
ANTIFOULING ACTIVITY OF SEAWEED ASSOCIATED BACTERIUM
39Thalassas, 28(2) · July 2012
Figure 2:Adhesion assay of Vibrio harveyi using the coupons conditioned with the EPS of Bacillus sp. ICN-SS01

V. RAJASREE, S. SATHEESH & S.G. PRAKASH VINCENT
40 Thalassas, 28(2) · July 2012
Evaluation of antifouling activity in the coastal waters
The EPS was mixed with wood polish in the ratio of 1:1. This mixture was coated onto acrylic panels. The panels were dried and used for the antifouling trials in the natural environment. The panels were fitted onto a metal raft and submerged in the Colachel coastal waters (West coast of India) using sufficient weight and floats. The panels were retrieved from the metal raft after 10 days of submersion. Panels coated with wood polish were used as control. The recruitment of biofouling communities on the panels was analysed from the images.
Analysis of EPS by thin-layer Chromatography
The EPS was analysed by thin-layer chromatography using silica gel coated glass plates. The solvents used for the TLC were methanol, acetic acid and benzene in the ratio 3:1:1.
RESULTS
EPS isolated from the bacterial strain associated with the showed inhibitory activity against Klebsiella sp., Escherichia coli. Alteromonas sp., Galionella sp. Vibrio harveyi and Pseudomonas sp (Fig 1). The active bacterial strain isolated from seaweed Sargassum wightii was Gram positive, non–motile and rod shaped. It was
indole negative, methyl red positive and Voges–Proskauer negative. It did not utilize citrate and in triple sugar iron agar test, it showed acid positive and alkaline negative results. It showed negative result for gas production and positive result for catalase test. It did not hydrolyze gelatin and starch. It did not utilize glucose, sorbitol, lactose and fructose in the carbohydrate utilization test. It showed acid positive in urease hydrolysis test (Table 1). Based on the biochemical and morphological characters, the strain was tentatively identified as Bacillus sp. and designated as Bacillus sp. ICN-SS01.
The adhesion assay carried out using EPS isolated
from strain (Bacillus sp. ICN-SS01) against the target bacteria showed that the extracellular polymeric substance inhibits the adhesion of bacterial strain on glass surface. The number of Vibrio harveyi on the coupons conditioned with the EPS isolated from the strain Bacillus sp. ICN-SS01 was 4612 cm-2 after 1 hour, whereas on the control coupons number of Vibiro harveyi cells adhered was 5904 cm-2. After 5 hours of glass surface immersion, the number of Vibiro harveyi cells found on the coupons coated with EPS of strain Bacillus sp. ICN-SS01 was 6915 cm-2. On the control coupons 8205 cells cm-2 were adhered on the glass surface after 5 hours (Fig. 2). Student ‘s ‘t’ test showed significant variation in the adhesion of Vibrio harveyi on coupons treated with EPS isolated from the strain Bacillus sp. ICN-SS01 and control (t stat: 7.82; df: 4; P<0.05).
Figure 3:
Antifouling assay in the coastal water using panels coated with EPS. The panels were coated with EPS of strain Bacillus sp. ICN-SS01 and wood polish and submerged in the coastal waters for a period of 10 days. A.
Control panel coated with wood polish only. B. Experimental panel coated with EPS and wood polish.

ANTIFOULING ACTIVITY OF SEAWEED ASSOCIATED BACTERIUM
41Thalassas, 28(2) · July 2012
The acrylic panels coated with EPS of the strain Bacillus sp. ICN-SS01 showed strong inhibitory activity against the settlement of fouling organism (Fig. 3). The control panel showed intense settlement of macroalgae during the 10 days submersion period. However the EPS coated panels showed less algae settlement than that of control. For the characterization of active compounds, the EPS were loaded on silica gel plates. The EPS of the bacterial strain showed a single distinct spot with the Rf value of 0.79 cm in the thin layer chromatogram (Fig. 4).
DISCUSSION
In the present study, a bacterial strain isolated from the surface of the seaweed Sargasum wightii showed antifouling activity against the bacteria involved in biofilm formation and invertebrates in the field assay. The strain was classified by using traditional biochemical and morphological methods and tentatively identified as Bacillus sp. ICN-SS01. The adhesion assay showed that EPS of the seaweed associated bacterial strain inhibited the adhesion of biofilm bacterial strains on to glass surface. Generally, marine bacteria are well known to produce a wide range of organic molecule. It is likely that bacteria adapted to surface could excrete metabolites that are not produced by the same cell in suspension. Production of these metabolites on a surface could be induced by other members of the bacterial community through
the intracellular communication. The search for these products has generally been focused on molecules that would have medical or industrial use. Exopolysaccharide (EPS) is a term first used by Sutherland (1982) to described high molecular weight carbohydrate polymers produced by many marine bacteria. Since that time, EPS also has been used to indicate a more broadly define extracellular polymeric substances (Wingender et al. 1999). In recent years, there has been a growing interest in isolating new exopolysaccharides (EPSs) - producing bacteria from marine environments, particularly from various extreme marine environments. Bacterial EPS can serve as antibacterial components to control bacterial attachment (Fletcher and Floodgate 1973) and benefit the survival of both the host and other organisms that live in the vicinity of the producer strain (Vincent et al. 1994, Szewzyk et al. 1991).
The antifouling compounds can be incorporated into the matrix at concentration higher than those in the natural environment without altering the physical characteristics of the settlement surface (Henrikson and Pawlik 1995). This technique is valuable for assaying the antifouling effects of secondary metabolites produced by marine organisms (Henrikson and Pawlik 1998). In
Sl. No.
Biochemical and
Physiological tests Results
1. Gram staining +
2. Morphology Rod
3. Motility Non- motile
4 Gelatin Hydrolysis -
5. Starch Hydrolysis -
6. Catalase Activity +
7. Citrate Utilization -
8. Indole production -
9. Methyl Red +
10. Voges Proskauer -
11. Glucose -
12. Fructose -
13. Lactose -
14. Sorbitol -
Figure 4:Thin-layer chromatographic analysis of the EPS isolated from the strain
Bacillus sp. ICN-SS01.
Table 1:Biochemical characteristics of the bacterial strain isolated from the
surface of the seaweed Sargassum wightii.

V. RAJASREE, S. SATHEESH & S.G. PRAKASH VINCENT
42 Thalassas, 28(2) · July 2012
the present study, the EPS was mixed with varnish and coated on test coupons. The EPS was added to the varnish at 1:1 ratio in order to maintain a higher concentration of the metabolite in the coating. Promising results were obtained in the field assay as the coupons considerably inhibited the settlement of microfouling and macrofouling communities found in the natural seawater. This is one of the major advantages of this study as most of the previous studies are mainly focusing on laboratory based screening and characterization of the compounds.
Thin layer chromatography analysis of the EPS showed a single spot indicating the presence of an active fraction. Exopolysaccharides generally consist of monosaccharides and some non-carbohydrate substituents (such as acetate, pyruvate, succinate, and phosphate). Thin-layer chromatography is an important analytical technique for the detection of bioactive compounds. However, further characterization of the active fraction using advanced techniques may lead to the identity and chemical structure of the compound being responsible for the antifouling activity.
In the conclusion, the present study showed that bacteria associated with seaweeds could be used as a potential source for the isolation of antifouling compounds. Because of the low content of active compounds in marine animals and plants as well as limitation of bio-resource supply, more and more researches have been focused on marine microorganisms as sustainable resources (Bultel-Ponoe et al. 1999). The EPS of the strain Bacillus sp. ICN-SS01 exhibited strong inhibitory activity against the fouling growth in field assay. Thus, the active compound present in the EPS of the Bacillus sp. ICN-SS01 is a topic of interest for future studies.
REFERENCES
Armstr ong E, Boyd KG, Burgess JG (2000a) Prevention of marine biofouling using natural compounds from marine organisms, Biotechnology Annual Review, 6: 221–241.
Armstrong E, Boyd KG, Pisacane A, Peppiatt CJ, Burges J G (2000b) Marine microbial natural products in antifouling coating, Biofouling, 16: 215–224.
Bultel-Ponce V, Berge JP, Debitus C, Nicola JL, Guyot M (1999) Metabolites from the sponge associated bacterium Pseudomonas species. Marine Biotechnology, 1: 384–390.
Clare AS (1996) Marine natural product antifoulants: status and potential. Biofouling, 9: 211–239.
Davidson SK, Allen SW, Lim GE, Anderson CM, Haygood MG (200) Evidence for the biosynthesis of bryostation by bacterial symbiont “Candidatus Endobugula sertula” of the bryozoan Bugula neritina. Applied and Environmental Microbiology, 67: 4531–4537.
Fingerman SW (1988) Environmental pollution by heavy metals from antifouling compounds. In: Thompson MF, Sarojini R, Nagabhushanam R.,eds, Marine Biodeterioration: Advanced techniques applicable to the Indian Ocean. Oxford & IBH. New Delhi 781-789.
Fletch er M, Floodgate GD (1973) An electron-microscopic demonstration of an acidic polysaccharide involved in the adhesion of a marine bacterium to solid surface. Journal of General Microbiology, 74: 325–334.
Harder T, Lau SC, Dobrestov S, Fang TK, Qian P-Y (2003) A distinctive epibiotic bacterial community on the soft coral Dendronephthya sp. and antibacterial activity of coral tissue extracts suggest of chemical mechanism against bacterial epibiosis. FEMS Microbiology Ecology, 43: 337–347.
Henrikson AA, Pawlik JR (1995) A new method of assaying extracts of marine organisms for antifouling properties. Journal of Experimental Marine Biology and Ecology, 194: 157–165.
Henrikson AA, Pawlik JR (1998) Seasonal variation in biofouling of gels containing extracts of marine organisms. Biofouling, 12: 245–255.
Jain A, Nishad KK, Narayan KK, Bhosle B (2007) Effects of DNP on the cell surface properties of marine bacteria and its implication for adhesion to surfaces. Biofouling, 23: 171–177.
Jenner HA, Whitehouse JW, Taylor CJL, Khalanski M (1998) Cooling water management in European power stations: biology and control Hydroecologie Applique e 1–2, Chatou, Paris: Electricite´ de France. 1–225.
Luesh H, Moore RE, Paul VJ, Mooberry SL, Corbett TH (2001) Isolation of dolastatin 10 from the marine cyanobacterium Symploca species VP 642 and total stereochemistry and biological evaluation of its analogue symplostatin 1. Journal of Natural Products, 64: 907–910.
Peppiatt CJ, Armstrong E, Pisacane T, Burguess JG (2000) Antibacterial activity of resin based coatings containing marine microbial extracts. Biofouling,16: 225–234.
Proksch P, Edrada RA, Ebel R (2002) Drugs from the seas-current status and microbiological implications. Applied Microbiology and Biotechnology, 59: 125–134.
Schupp P, Eder C, Proksch P, Warry VV, Schneider B, Herderich M, Paul VV (1999) Staurosporine derivatives from the ascidican Eudistoma toealensis and its predatory flatworm Pseudoceros sp. Journal Natural Products, 62: 959–962.
Sutherland IW (1982) Biosynthesis of microbial exopolysaccharides. Advances in Microbial Physiology, 23:79–150.
Szewzyk U, Holmstrom C, Wrangstadh M, Samuelsson MO, Maki JS, Kjelleberg S, (1991) Relevance of the exopolysaccharides of marine Pseudomonas sp strain sq for the attachment of Ciona intestinalis larvae. Marine Ecology Progress Series, 75: 259–265.
Unson MD, Faulkner DJ (1993) Cyanobacterial symbiont biosynthesis of chlorinated metabolites from Dysidea herbacea (Proifera). Experientia, 49: 349–353.

ANTIFOULING ACTIVITY OF SEAWEED ASSOCIATED BACTERIUM
43Thalassas, 28(2) · July 2012
Vincent P, Pignet, Talmont F, Bozzi L, Fournet B, Guezennee JG (1994) Production and characterization of an exopolysachharide excreted by a deep sea hydrothermal vent bacterium isolated from the polychaete annelid Alvinella pompejana. Applied and Environmental Microbiology, 60: 4134–4141.
Wingender J, Neu TR, Flemming H-C (1999) What are bacterial extracellular polymer substances? In: Wingender J, Neu TR, Flemming H-C, eds, Microbial Extracellular Polymer Substances. Springer, Berlin. 1-19.
Yebra DM, Kiil S, Dam-Johansen K (2004) Antifouling technology – past, present and future steps towards efficient and environmentally friendly antifouling coatings. Progress in Organic Coatings, 50: 75 – 104.
(Received: February, 24, 2012; Accepted: May, 2, 2012)


45Thalassas, 28(2) · July 2012
NEARSHORE BENTHIC COMMUNITIESAND BIOENGINEERS FROM THE MACROTIDALSAN JORGE GULF: PATAGONIA, ARGENTINA
(1) Universidad Nacional de Mar del Plata. Funes 3350 (7600) Mar del Plata, ARGENTINA
(2) Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) ARGENTINA
e-mail: [email protected]
Thalassas, 28(2) · July 2012: 45-56An International Journal of Marine Sciences
Key words: benthic communities, bioengineers, macrotides, San Jorge gulf, Patagonia.
Palabras clave: comunidades bentónicas, bioingenieros, macromareas, Golfo San Jorge, Patagonia.
LEANDRO FAINBURG(1), MARÍA TRASSENS(1,2), JULIÁN BASTIDA(1), MARCELO FARENGA(1),FEDERICO ISLA(1,2) & RICARDO BASTIDA(1,2)
45Thalassas, 28(2) · July 2012
ABSTRACT
San Jorge Gulf, located in northern Patagonia, is characterized by extensive soft bottoms and large tidal ranges. A marine bot-tom section of 3 x 2 km, located off Caleta Olivia (Santa Cruz Province, Argentina) was sampled intensively during August 2008, covering a total of 35 oceanographic stations, with depths ranging from 25 to 48 m. Despite the small size of the surveyed area, and the short distance between stations, the study showed important bottom diversity, with patchiness distribution and associated with different biological assemblages. Through the geological and biological samples analysis, and subsequent statistical treat-ment, four main types of substratum were defined in the area, from those with very fine dominant sedimentary fraction to others with gravel dominance and also rocky platforms. The study area, of high hydrological energy, impacts on sediment dynamics, thus clogging phenomena are observed on rocky bottoms benthic communities. This clogging effect is mitigated in part by the bioengineering action of the polychaete Eunice fraunfeldi, whose long leathery tubes allow the settlement of various epibenthic species in areas dominated by soft bottoms, and thus reflecting the importance of bioengineering species creating special sub-strate for epibenthic communities.
RESUMEN (Comunidades bentónicas costeras y bioingenieras macromareales del Golfo San Jorge: Patagonia, Argentina)
El Golfo San Jorge, ubicado en el norte de la Patagonia (Argentina), se caracteriza por extensos fondos blandos y amplios rangos mareales. Una porción del fondo marino de 3 x 2 km, ubicado costa afuera de Caleta Olivia (Provincia de Santa Cruz) fue muestreado según 35 estaciones oceanográficas en agosto de 2008, y a profundidades entre 25 y 48 m. A pesar del pequeño tamaño del área analizada, y la escasa distancia entre estaciones, se constató una significativa diversidad de fondos, con distribución según parches y asociados a diferentes conjuntos biológicos. A través de muestras geológicas y biológicas, y su tratamiento estadístico, cuatro tipos de sustratos fueron definidos para el área, desde los dominados por sedimentos muy finos a los compuestos por grava o plataformas rocosas. La alta energía de la zona impacta en la dinámica sedimentaria, y así se reconocieron fenómenos de sofocamiento de las comunidades bentónicas en fondos rocosos. Este efecto es mitigado en parte por la actividad del poliqueto Eunice fraunfeldi, cuyos largos tubos coriáceos permiten el asentamiento de varias especies epibentónicas en fondos blandos, y así reflejando la importancia de las especies bioingenieras en la creación de sustratos especiales para comunidades epibentónicas.

LEANDRO FAINBURG, MARÍA TRASSENS, JULIÁN BASTIDA, MARCELO FARENGA, FEDERICO ISLA & RICARDO BASTIDA
INTRODUCTION
Macroinvertebrates from subtidal environments are conditioned by the bottom composition and the hydrological dynamics. Epifauna dominates in rocky bottoms, while infaunal benthic communities dominate where soft bottoms are stable (Elías 1995; López Gappa & Sueiro, 2007). Sediment transport can favour the food supply of bottom organisms but it can also induce clogging of benthic communities, especially in reduced extension areas where both types of bottoms coexist. Epifaunal subtidal assemblages are less studied than infaunal communities, although the former- in many areas- are continuously subject to instability related to sediment dynamics (Nichols et al. 2007).
In regard to hydrological dynamics, bays with low tidal range respond in relation to wave/wind effects. On the other hand, in macrotidal bays the benthic community is subject to intense currents twice a day. Therefore, nearshore fauna has significant changes in its composition in relation to the stress induced by the erosion of the substrate or the episodic sedimentation of mud over the benthos (Moreira et al. 2010).
Besides the effects of the bottom type as a limiting factor, epibiosis consortium process is another important factor in the structure of benthic communities. Some species offer a hard biological substrate for other organisms through their exoskeletons, in order to avoid assemblages clogging by soft sediments. Although epibiosis processes have been very well known since long time ago, in the last decades they were associated directly with other more complex ecological mechanisms (Jones et al. 1994).
These biological substrates, in many cases, create mesocosmos environments above the benthic layer subject to more stress due to clogging. Some of the organisms may also take advantage of these conditions living above the original bottom, avoiding predator actions or having a better access to water column preys, and also increasing community biodiversity (Margalef 1962, 1974; Rützler 1970; Jones et al. 1994; Nybakken & Bertness 2004; Levington 2009).
This process is fundamental in the development of epibenthic communities, and it can be easily observed in all the marine regions of the world. In our days this phenomena is included in the bioengineering concepts (Margalef 1962, 1974; Bastida et al. 1977; Bastida & Lichtschein 1978; Jones et al. 1994; Sosa 2006).
In the present paper, a reduced bottom area (3 x 2 km), between 25 and 48 m depth, was analyzed in order to study the composition and distribution of benthic macroinvertebrates and their relationships with different substrates and also in terms of determining bioengineering processes (Fig. 1).
SETTING
Argentine continental shelf is dominated by sandy bottoms (Fray & Ewing 1963; Vozza et al. 1974; Bastida et al. 1981, 1992) that vary in response to tidal energy, distance to coast, bioclastic abundance and bioturbation activity. The initial surveys conducted on the continental shelf of Buenos Aires Province (Olivier et al. 1968, Cortelezzi et al. 1971) and throughout the continental shelf (Bastida et al. 1981, 1992; Fernández et al. 2003, 2007a, 2007b, 2008) confirm the clear dominance of
46 Thalassas, 28(2) · July 2012
Figure 1:Location map of San Jorge Gulf, with detail of sampling stations at Caleta Olivia (Santa Cruz).

NEARSHORE BENTHIC COMMUNITIES AND BIOENGINEERS FROM THE MACROTIDAL SAN JORGE GULF: PATAGONIA, ARGENTINA
soft bottoms. However, in many of these surveys rocky bottoms were also detected, although of reduced surface and low altitude, thus clogging processes in areas of high energy may be frequent (Brankevich, 1990). Also the interaction between hard and soft bottoms has been recorded photographically in different parts of the continental shelf (Bastida, unpublished) (Fig. 6).
The San Jorge Gulf (Fig.1) is characterized by a
significant amount of wave energy dispersed (Pousa et al. 1995).Tidal range increases towards the interior of the gulf to a macrotidal semidiurnal regime (Isla et al. 2002). Temperature fluctuates within the gulf between 7 and 13ºC; salinity is rather stable between 33.2 and 33.6 psu (Roux et al. 1995; Fernández et al. 2007a, 2008). The gulf is silted by mud, although fine, medium and coarse sand can be sampled from the shallow areas (Fernández et al. 2003). Gravel and rocky bottoms dominate towards
the northern and southern capes (Cabo Tres Puntas, Cabo Dos Bahías and Cabo Blanco) and surrounded by areas composed of fine sand (Roux et al. 1995). Fine silt dominates towards the southwest area of the gulf.
San Jorge Gulf is important for its biological and commercial activities. It is part of the nursery area of the hake (Merluccius hubbsi), the main fish resource from Argentina, and a the fishing ground of the shrimp (Pleoticus muelleri) the more valuable fishing species of the country (Roux et al. 1995; Boschi 1997; Fernández et al. 2003; Fernández et al. 2007b,. San Jorge Gulf is also a major reproductive area of the lobster krill (Munida gregaria), one of the most important prey for top predators as big fish, birds and marine mammals of Patagonia (Vinuesa & Varisco 2007; Fainburg, 2010) The Macrocystis pyrifera kelp forests are also frequent in the gulf and other coastal localities of the Patagonian region.
47Thalassas, 28(2) · July 2012
Figure 2:Dendrogram based on the Euclidean distance of the sediments grain size and bottom characteristics of studied stations.

LEANDRO FAINBURG, MARÍA TRASSENS, JULIÁN BASTIDA, MARCELO FARENGA, FEDERICO ISLA & RICARDO BASTIDA
METHODS
Bottom sampling spanned between 46º 28´ 02” S and 46º 29 54” S, and depths between 25 and 48 m in front of Caleta Olivia harbour (Fig. 1). A total of 35 stations were sampled at distances of 500 m between each other. Quantitative Picard dredges of 60 x 20 cm were used to collect benthic macroinvertebrates and sediment samples.
Sediment samples were sieved in lab between 1 and 0.1 cm mesh. Data obtained from sediment were included in tables indicating percentages of gravel, coarse-very coarse sand, medium sand, fine sand, very fine sand, silt and clay (Table 1). Cluster analyses based on Euclidean distances were performed by the mean of the SPSS 11.5 package.
48 Thalassas, 28(2) · July 2012
Group A1
Group A2
Group B
Sand
Gravel
Mud
Figure 3:Bottom substrate compositional triangle.
Figure 4: Dendrogram based on the Pearson correlation coefficient showing the main association between benthic species.

49Thalassas, 28(2) · July 2012
Biological samples were obtained from sieved sediment or fragmented rocks. Benthic organisms were classified to species or genus level and fixed in formalin and kept in alcohol 70%.
Based on the identified benthic species, a faunal inventory of the studied area was achieved (Table 2) and a basic data matrix was used for statistical handling.
In order to analyze the association among the most frequent benthic species, a dendrogram based in Pearson correlation R-mode was performed (Fig. 4) and a Q-mode dendrogram (Fig. 4) was designed to determine the biological affinity between stations.
RESULTS
Marine bottom characteristics.
Despite the small size of the studied area, there was a great diversity in its bottom substrates. Some sectors were dominated by rocky bottoms, others by gravel and sand, others exclusively by sand and finally bottom sectors with less sediment selection, composed by gravel, sands, silt and clay (Table1 and Fig. 8).
Dendrogram related with sea bottom characteristics, based on the Euclidean distance (Fig. 2), indicates two major groupings of stations (A and B). The subgroup A 1 consists of stations with a clear dominance of gravel representing 99% of the sample and directly linked to the platform rocky outcrops. In the subgroup A2 gravel is still the sediment dominant fraction, represented mostly as fragments of the rocky platform, but with values ranging between 45 and 70%; the sediment matrix is also integrated by other smaller grain size fractions.
Group B is characterized by stations with greater grain size variability, including different percentages of sand and mud; while few stations are composed by small size gravels.
The clustering shown in the last dendrogram is also expressed in the triangular graph (Fig. 3). In its upper angle we can observe the gravel dominance stations (A1), below them the medium sedimentary fractions (A2) and in the lower sector of the graph, the sand and pelites dominance in the sedimentary fraction (B).
Benthic community distribution
Benthic assemblages of the studied area are represented by 8 phyla, 35 genus and at least 37 species (Table 2). Stations biodiversity values (S) varies between
20 and 22. The Shannon index of ecological diversity is of 2.49, with H max= log S 3.178 and an evenness of 0.783.
The Pearson correlation dendrogram (Fig. 4) defines a total of three main groups. Group A concentrates all the fauna that colonize Eunice frauenfeldi polychaete tubes, a bioengineer species of this assemblage, which enables epizoites settlement of other species of small or medium size (Porifera 11.11%, Anthozoans and Hydrozoans 44.44%, Bryozoans 33.33% and Tunicates 11.11%) (Fig. 5).
Group B is an indicator of hard-bottom assemblage; its diversity is lower than in group A. It is dominated by sessile species such as Spirorbidae and Serpulidae polychaetes and the bivalve Aulacomya ater; between mobile species we can also mention the Echinoderms Arbacia dufresnei and Cosmasterias lurida.
Group C is poorly diversified and dominated by the filtering Pelecypod Tindaria striata, a soft bottom indicator species. The crustacean Munida gregaria is the subdominant species of the group, and because of its great mobility and food habits, it can be linked with both rocky and soft bottoms benthic assemblages (Group A and Group B, respectively) ( Fig. 4).
Annelida and Mollusca are the dominant phyla (Table 2) in the studied area. Most of these species are subject to stress induced by high-energy waves or clogging when mud becomes dominant on the bottom. Some polychaetes species, as Eunice frauenfeldi, are able to construct large and resistant tubes as refuge. These tubes constitute a three dimensional protruding substrate where other benthic species find stability and shelter (Fig. 6 and 7).
In order to define the affinity between the various benthic stations, a Q-mode dendrogram was performed, using the Pearson correlation test (Fig. 8). Based on this dendrogram, and despite the small size of the study area, three main clusters were differentiated.
Group A is formed by stations inhabited by soft bottoms organisms. It is mainly characterized by the presence of Tindaria striata - Munida gregaria and some species of polychaetes poorly represented.
Group B is composed by stations with bioengineering processes promoted by the presence of Eunice frauenfeldi and its leathery tubes.
Group C clusters rocky bottom stations, mainly colonized by calcareous polychaete tubes of the Spirorbidae and Serpulidae family, and the Mytilidae mollusk Aulacomya ater.
NEARSHORE BENTHIC COMMUNITIES AND BIOENGINEERS FROM THE MACROTIDAL SAN JORGE GULF: PATAGONIA, ARGENTINA
LEANDRO FAINBURG, MARÍA TRASSENS, JULIÁN BASTIDA, MARCELO FARENGA, FEDERICO ISLA & RICARDO BASTIDA
50 Thalassas, 28(2) · July 2012
Figure 5:Percentage of faunistic groups that act as epizoites of Eunice frauenfeldi polychaete tubes.
Figure 6:Underwater photograph of benthic assemblages colonizing polychaete tubes.
Figure 7:Detail of the bioengineer polychaete Eunice frauenfeldi; specimen and tube.
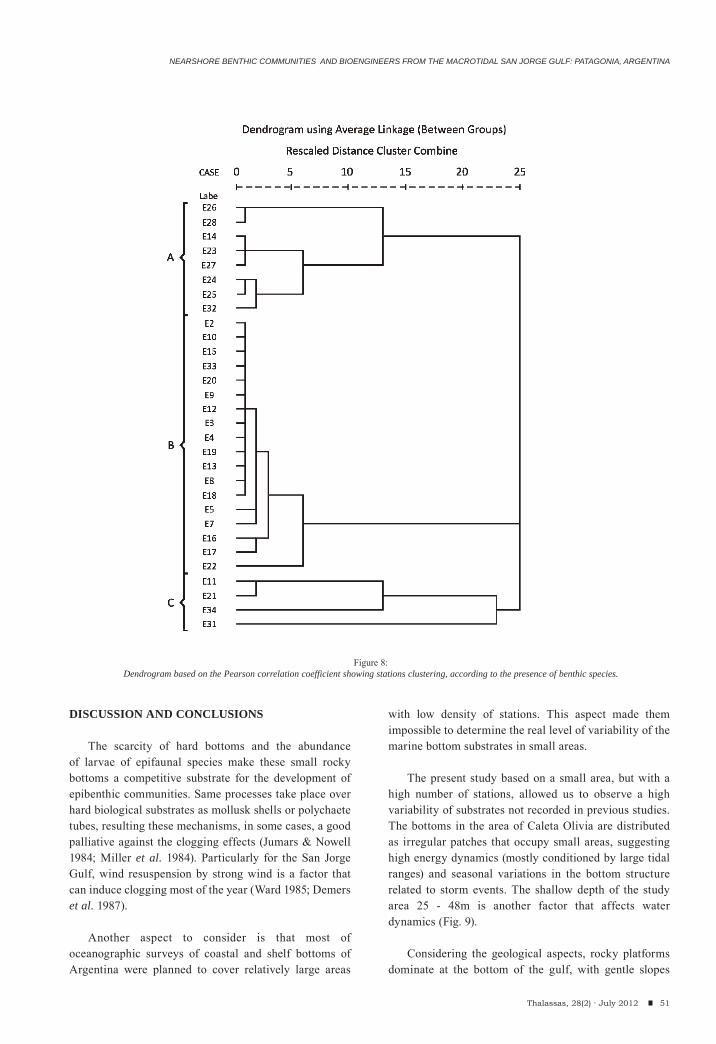
NEARSHORE BENTHIC COMMUNITIES AND BIOENGINEERS FROM THE MACROTIDAL SAN JORGE GULF: PATAGONIA, ARGENTINA
DISCUSSION AND CONCLUSIONS
The scarcity of hard bottoms and the abundance of larvae of epifaunal species make these small rocky bottoms a competitive substrate for the development of epibenthic communities. Same processes take place over hard biological substrates as mollusk shells or polychaete tubes, resulting these mechanisms, in some cases, a good palliative against the clogging effects (Jumars & Nowell 1984; Miller et al. 1984). Particularly for the San Jorge Gulf, wind resuspension by strong wind is a factor that can induce clogging most of the year (Ward 1985; Demers et al. 1987).
Another aspect to consider is that most of oceanographic surveys of coastal and shelf bottoms of Argentina were planned to cover relatively large areas
with low density of stations. This aspect made them impossible to determine the real level of variability of the marine bottom substrates in small areas.
The present study based on a small area, but with a high number of stations, allowed us to observe a high variability of substrates not recorded in previous studies. The bottoms in the area of Caleta Olivia are distributed as irregular patches that occupy small areas, suggesting high energy dynamics (mostly conditioned by large tidal ranges) and seasonal variations in the bottom structure related to storm events. The shallow depth of the study area 25 - 48m is another factor that affects water dynamics (Fig. 9).
Considering the geological aspects, rocky platforms dominate at the bottom of the gulf, with gentle slopes
51Thalassas, 28(2) · July 2012
Figure 8:Dendrogram based on the Pearson correlation coefficient showing stations clustering, according to the presence of benthic species.

LEANDRO FAINBURG, MARÍA TRASSENS, JULIÁN BASTIDA, MARCELO FARENGA, FEDERICO ISLA & RICARDO BASTIDA
52 Thalassas, 28(2) · July 2012
without significant irregularities, and with a thin cover of sediment, responsible of the clogging processes. At the area of this study, significant variations of the composition of the marine bottom were distributed into 4 types of substrates (Fig. 9).
From the biological point of view, the effect of clogging in the study area is clearly evidenced by the rocky bottom communities, which show low biodiversity or no colonization because they were covered by a sedimentary mantle until shortly before the time of this sampling. At some stations, through fragments of abiotic sedimentary rocks, it was possible to confirm historical clogging events by imprints and remains of calcareous organisms attached to the substrate (Polychaeta Serpulidae, Spirorbidae, Bryozoans Membraniporidae and Balanidae Cirripeds).
The statistical treatment of biological samples indicate the presence of different benthic assemblages or communities in the study area (Fig. 4) and each of these assemblages (Group A, B and C) is associated in turn with substrates of different features confirming the conditioning exerted by the substrate on the type and abundance of benthic communities (Fig 8).
The rocky bottom benthic assemblage (group B) shows low abundance of organisms and low diversity, as a result of the effect of clogging. In short, it is an impoverished community when compared with another assemblage of the Patagonian coast not affected by clogging phenomena (Olivier et al. 1966; Cuevas et al. 2006).
The group C has low diversity and is dominated by Tindari striata, a good indicator species of sandy sediments, however Munida gregaria (subdominant
species) is not a good indicator of substrates, because due to their high mobility and plasticity (Perez Barros 2004; Vinuesa & Varisco 2007) can be integrated into other assemblages and hence its links with groups A and B of the corresponding dendrogram (Fig. 8).
The group A assemblage is characterized by the dominance of epizoites species colonizing the leathery tube of the polychaete Eunice frauenfeldi, so that assemblage is not directly linked with the sediment but with the biological substrate. Eunice frauenfeldi may be present in variable-grained soft bottoms, but the organism must have certain percentage of gravel to allow the initial anchoring of the tube. With the passage of time and the growth of the polychaete, the tube can remain supported directly over the fine sediments surrounding the gravel initially colonized by the bioengineer polychaete.
Today, due to the rise of the concept of bioengineering, the phenomenon of epibiosis consortium has increased in consideration by ecologists as an important mechanism in structuring benthic communities and ecosystems.
The community controlled by bioengineers was not recognized in other macrotidal bays dominated by mud in the Patagonian coast, although bioengineering processes has been detected in certain soft bottom areas of the Argentine Continental Shelf (Olivier et al. 1968; Bastida et al. 1992).
The study of consortia, as a biological and ecological phenomena, has attracted the attention of naturalists since ancient times. However, the wide range of diverse consortia has not been addressed with similar dedication. Most of the efforts have been devoted to the knowledge
Figure 9: Classification of marine bottom substrates along the studied stations.
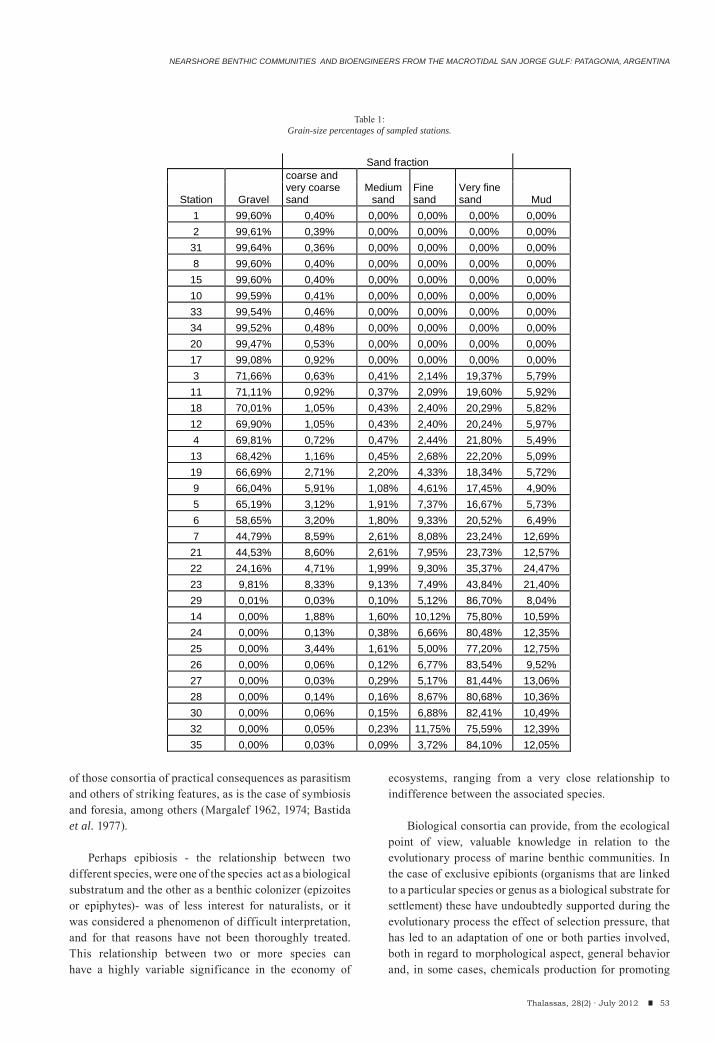
NEARSHORE BENTHIC COMMUNITIES AND BIOENGINEERS FROM THE MACROTIDAL SAN JORGE GULF: PATAGONIA, ARGENTINA
53Thalassas, 28(2) · July 2012
of those consortia of practical consequences as parasitism and others of striking features, as is the case of symbiosis and foresia, among others (Margalef 1962, 1974; Bastida et al. 1977).
Perhaps epibiosis - the relationship between two different species, were one of the species act as a biological substratum and the other as a benthic colonizer (epizoites or epiphytes)- was of less interest for naturalists, or it was considered a phenomenon of difficult interpretation, and for that reasons have not been thoroughly treated. This relationship between two or more species can have a highly variable significance in the economy of
ecosystems, ranging from a very close relationship to indifference between the associated species.
Biological consortia can provide, from the ecological point of view, valuable knowledge in relation to the evolutionary process of marine benthic communities. In the case of exclusive epibionts (organisms that are linked to a particular species or genus as a biological substrate for settlement) these have undoubtedly supported during the evolutionary process the effect of selection pressure, that has led to an adaptation of one or both parties involved, both in regard to morphological aspect, general behavior and, in some cases, chemicals production for promoting
Sand fraction
Gravel
coarse and very coarse sand
Medium sand
Fine sand
Very fine sand Mud Station
1 99,60% 0,40% 0,00% 0,00% 0,00% 0,00% 2 99,61% 0,39% 0,00% 0,00% 0,00% 0,00%
31 99,64% 0,36% 0,00% 0,00% 0,00% 0,00% 8 99,60% 0,40% 0,00% 0,00% 0,00% 0,00%
15 99,60% 0,40% 0,00% 0,00% 0,00% 0,00% 10 99,59% 0,41% 0,00% 0,00% 0,00% 0,00% 33 99,54% 0,46% 0,00% 0,00% 0,00% 0,00% 34 99,52% 0,48% 0,00% 0,00% 0,00% 0,00% 20 99,47% 0,53% 0,00% 0,00% 0,00% 0,00% 17 99,08% 0,92% 0,00% 0,00% 0,00% 0,00% 3 71,66% 0,63% 0,41% 2,14% 19,37% 5,79%
11 71,11% 0,92% 0,37% 2,09% 19,60% 5,92% 18 70,01% 1,05% 0,43% 2,40% 20,29% 5,82% 12 69,90% 1,05% 0,43% 2,40% 20,24% 5,97% 4 69,81% 0,72% 0,47% 2,44% 21,80% 5,49%
13 68,42% 1,16% 0,45% 2,68% 22,20% 5,09% 19 66,69% 2,71% 2,20% 4,33% 18,34% 5,72% 9 66,04% 5,91% 1,08% 4,61% 17,45% 4,90% 5 65,19% 3,12% 1,91% 7,37% 16,67% 5,73% 6 58,65% 3,20% 1,80% 9,33% 20,52% 6,49% 7 44,79% 8,59% 2,61% 8,08% 23,24% 12,69%
21 44,53% 8,60% 2,61% 7,95% 23,73% 12,57% 22 24,16% 4,71% 1,99% 9,30% 35,37% 24,47% 23 9,81% 8,33% 9,13% 7,49% 43,84% 21,40% 29 0,01% 0,03% 0,10% 5,12% 86,70% 8,04% 14 0,00% 1,88% 1,60% 10,12% 75,80% 10,59% 24 0,00% 0,13% 0,38% 6,66% 80,48% 12,35% 25 0,00% 3,44% 1,61% 5,00% 77,20% 12,75% 26 0,00% 0,06% 0,12% 6,77% 83,54% 9,52% 27 0,00% 0,03% 0,29% 5,17% 81,44% 13,06% 28 0,00% 0,14% 0,16% 8,67% 80,68% 10,36% 30 0,00% 0,06% 0,15% 6,88% 82,41% 10,49% 32 0,00% 0,05% 0,23% 11,75% 75,59% 12,39% 35 0,00% 0,03% 0,09% 3,72% 84,10% 12,05%
Table 1:Grain-size percentages of sampled stations.
LEANDRO FAINBURG, MARÍA TRASSENS, JULIÁN BASTIDA, MARCELO FARENGA, FEDERICO ISLA & RICARDO BASTIDA
54 Thalassas, 28(2) · July 2012
attachments. These aspects are well manifested in the epibionts with specialized adhesive organs, chemical receptors, or those that are set in certain parts of the body of the substrate organism in connection with some of the vital activities it develops (Wilson 1952; Crisp & Williams 1960; Margalef 1962; Williams 1964; Rützler 1970; Stebbing 1971, 1972; Bastida et al. 1977).
Experimental studies on epibenthic communities have shown in Argentine harbour areas the importance of the phenomenon of epibiosis during different stages of the ecological succession of these communities and provide also important knowledge for biofouling control systems (Rascio et al. 1969; Bastida 1971; Bastida & Torti, 1973; Piriz 1972; Bastida et al. 1977, 1978, 2011).
In the particular case of Eunice frauenfeldi, it plays a significant role in the ecology of the study area. Their leathery tubes can reach several meters in length and allow the development of epibenthic species typical of rocky bottoms on soft-bottom areas (Fainburg 2010).
ACKNOWLEDGEMENTS
This work could not have been possible without the help of YPF company and the crew of the research vessel “Mr. Bigg”. We also want to thank Dr. Juan Pablo Martin (UNPA) for his collaboration in the identification of polychaetes and Dr. Viviana Quse (IUCN- UNL) for the manuscript review.
REFERENCES
Bastida, R. (1971). Las incrustaciones biológicas en el puerto de Mar del Plata, Período 1966/1967. Revista del Museo de Ciencias Naturales. “Bernardino Rivadavia”, Hidrobiol., III (2): 203-285.
Bastida, R. & M. R. Torti. 1973. Estudios preliminares sobre las incrustaciones biológicas de Puerto Belgrano (Argentina). Corrosión y Protección, 5: 3-11.
Bastida, R., E. Spivak, S. L Hoste & H. Adabbo. 1977. Las incrustaciones biológicas de Puerto Belgrano. II. Estudio de los procesos de epibiosis registrados sobre paneles
Cellaria malvinensisPhylum Bryozoa Scrupocellaria scruposa puelcha
Amastigia benemunitaHarmotoinae indet. Lepidonotidae indet.Terebillides stroemii Nephtys magellanica
Phylum Annelida Clase Polychaeta Syllidae indet Maldanidae indet. Eunice frauenfeldiSerpulidae indet.Spirobidae indet.
Photinula coerulescensCalyptrea pileus
Clase Gasteropoda Calyptrea pileolusCalliostoma coppingeri
Phylum Mollusca Polinices patagonicusClase Bivalvia Aulacomya ater
Tindaria striataClase Polplacophora Tonicia chilensis
Munida gregariaPhylum Arthropoda Subphylum Crustacea Peltarion spinosulum
Anfípodo gamárido indet.Clase Asteroidea Platisteria sp.
Cosmasterias luridaPhylum Echinodermata Pseudechinus magellanicus
Clase Echinoidea Arbacia dufresnei Tripylaster philippi
Clase Ophiuroidea Ophiura sp .Ascidia sp.
Phylum Chordata Subphylum Urochordata Molgulidae indet.
Didemnum sp.
Table 2:List of the principal benthic species of the studied area.

NEARSHORE BENTHIC COMMUNITIES AND BIOENGINEERS FROM THE MACROTIDAL SAN JORGE GULF: PATAGONIA, ARGENTINA
55Thalassas, 28(2) · July 2012
mensuales. Corrosión y Protección 8:25-33Bastida, R. & V. Lichtschein. 1978. Las incrustaciones biológicas
de Puerto Belgrano. III. Estudio de los procesos de epibiosis registrados sobre paneles acumulativos. CIDEPINT Anales 1978, Ser. II. (368): 57-97.
Bastida, R., C. Urien, V. Lichtschein, A. Roux & P. Arias, 1981. Investigaciones sobre comunidades bentónicas. I. Características generales del sustrato (Campañas IV, V, X y XI del BIP Shinkai Maru).Contribución INIDEP 383: 318-339.
Bastida, R., A. Roux & D. Martinez, 1992. Benthic communities of the Argentine Continental Shelf. Oceanologica Acta 15 (6): 687- 698.
Bastida, R., M. Trassens, J. P. Martin & M. Grosso, 2011. El papel de los sedimentos en la formación y conservación de los sitios arqueológicos subacuáticos: el caso de la HMS Swift (1770). Sección Estudios Especializados: 27-57. En: Elkin, D., C. Murray, R. Bastida, M. Grosso, A. Argüeso, D. Vainstub, C. Underwood y N. Ciarlo. 2011. El naufragio de la HMS Swift (1770). Arqueología marítima en la Patagonia, Vázquez Mazzini Editores, Buenos Aires.
E. Boschi, 1997. Las pesquerías de crustáceos decápodos en el litoral de la República Argentina. Investigaciones Marinas. Valparaíso, 25: 19-40
Brankevich, G., A. Roux & R. Bastida, 1990. Relevamiento de un banco de pesca del besugo (Sparus pagrus) en la plataforma bonaerense. Características fisiográficas y aspectos ecológicos preliminares. Frente Marítimo 7, Sec. a: 75-86.
Cortelezzi, C., Colado, U, Mouzo, F. & Robbiano, J. 1971. Resultado de las campañas oceanográficas, Mar del Plata I-V, Carta Batilitológica de la región de Mar del Plata. Revista del Museo de La Plata, NS, 7 (59): 207-252.
Crisp, D. & Williams, G. 1960. Effect of extracts from fucoids in promoting settlement of epiphytic Polyzoa. Nature, 188: 1206-1207.
Cuevas, J. M., J. P Martin & R. Bastida, 2006. Benthic Community changes in a Patagonian Intertidal: a forty years later comparison. Thalassas, 22 (1):31-39. ISSN: 0212-5919.
Demers, S., Therriault, J. C., Bourguet, E. & Bah, A., 1987. Resuspension in the shallow sublittoral zone of a macrotidal estuarine environment: wind influence. Limnology and Oceanography 32 (2):327-329.
Elías, R., 1995. The subtidal macroinfauna from soft-bottom substrate of the Blanca Bay (Argentine). Thalassas 11, 73-86.
Fainburg, L. A., 2010. Relación entre los organismos bentónicos y sus sustratos: un caso de muestreo intensivo en un área restringida de la plataforma costera frente a Caleta Olivia (Santa Cruz-Argentina). Unpublished Thesis, Facultad de Ciencias Exactas y Naturales, Mar del Plata, 55 pp.
Fernández, M., Roux, A., Fernández, E., Calo, J., Marcos, A.& Aldacur, H., 2003. Grain-size analysis of surficial sediments from Golfo San Jorge, Argentina. Journal of the Marine Biology Association of the United Kingdom, 83: 1193-1197.
Fernández, M., D. A. Cucchi Colleoni, A. Roux, A. Marcos & E. Fernández, 2007a Caracterización físico-química del sistema bentónico en el sector sur del Golfo San Jorge, Argentina. Revista de Biología Marina y Oceanografía, 42 (2): 177-192.
Fernández, M., D. Hernández & A. Roux, 2007b Distribución espacial del langostino patagónico (Pleoticus muelleri, (Bate, 1888) y su relación con las variables ambientales, Golfo San Jorge, Argentina. Revista de Biología Marina y Oceanografía, 42(3): 335-344
Fernández, M., J. Mora, A. Roux, D.A. Cucchi Colleoni & J.C. Gasparoni, 2008., New contribution on spatial and seasonal variability of environmental conditions of the Golfo San Jorge benthic system, Argentina. Journal of the Marine Biology Association of the United Kingdom, 88 (2): 227-236.
Fray, Ch. & M. Ewing, 1963. Pleistocene sedimentation and fauna of the Argentine shelf.I. Wisconsin sea level as indicated in Argentine continental shelf series. Proc. Academy of Natural Sciences of Philadelphia, 115 (6): 113-126.
Isla, F. I., N. Iantanos & E. Estrada, 2002. Playas reflectivas y disipativas macromareales del Golfo San Jorge. AAS Revista, 9 (2): 155-164.
Jones, C.G., J.H. Lawton & M, Shachak., 1994. Organisms as ecosystem engineers. Oikos, 69: 373-386.
Jumars, P. A. & A.R Nowell,., 1984. Fluid and sediment dynamic effects on marine benthic community structure. Am. Zoologist 24: 45-55.
Levington, J., 2009. Marine Biology – Function, Biodiversity, Ecology. Oxford University Press. USA. 3rd Edition. ISBN 978-0-19-532694-9. 640 pp.
López Gappa, J. & M.C. Sueiro, 2007. The subtidal macrobenthic assemblages of Bahía San Sebastián (Tierra del Fuego, Argentina). Polar Biology 30: 679-687.
Margalef, R. 1962. Comunidades naturales. Instituto de Biología Marina. Universidad de Puerto Rico (publ. esp.), I-VII:1-468.
Margalef, R. 1974. Ecología. Ediciones Omega S.A., Barcelona.Miller, D.C., P.A. Jumars, & A.R. Nowell, 1984. Effects of
sediment transport on deposit feeding: scaling arguments. Limnology and Oceanography 29(6):1202-1217.
Moreira, J., A. Lourido & J. S Troncoso, 2010. Temporal dynamics of the benthic assemblage in the muddy sediments of the harbour of Baiona (Galicia, NW Iberian Peninsula). Thalassas 26 (2): 9-22.
Nicholls, R.J., P.P. Wong, V.R. Burkett, J.O. Codignotto, J.E. Hay, R.F. McLean, S. Ragoonaden & C.D. Woodroffe, 2007. Coastal systems and low-lying areas. Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, M.L. Parry, O.F. Canziani, J.P. Palutikof, P.J. van der Linden & C.E. Hanson, Eds., Cambridge University Press, Cambridge, UK, 315-356.
Nybakken, J.W. & M.D. Bertness, 2004. Marine Biology:

LEANDRO FAINBURG, MARÍA TRASSENS, JULIÁN BASTIDA, MARCELO FARENGA, FEDERICO ISLA & RICARDO BASTIDA
56 Thalassas, 28(2) · July 2012
An Ecological Approach. Sixth Edition. Published by Benjamin/ Cummings Pub. Co. 579 pp.
Olivier, S., R. Bastida & M. R. Torti, 1968. Resultados de las campañas oceanográficas Mar del Plata I - V. Contribución al trazado de una carta bionómica del área de Mar del Plata. Las asociaciones del Sistema Litoral entre 20 y 70m de profundidad. Boletín del Instituto de Biología Marina, 16: 1-85.
Olivier, S., R. Bastida & M. R. Torti, 1968. Sobre el Ecosistema de las Aguas Litorales de Mar del Plata. Niveles Tróficos y Cadenas Alimentarias Pelágico - Demersales y Bentónico - Demersales. Servicio de Hidrografía Naval H - 1025: 1-45.
Olivier, S., I.K. de Paternoster & R. Bastida, 1966. Estudios biocenóticos en las costas de Chubut (Argentina). I. Zonación biocenológica de Puerto Pardelas (Golfo Nuevo). Boletín del Instituto de Biología Marina, (Mar del Plata), 10: 1-74.
Pérez-Barros, P. 2004. Benthic decapod crustaceans associated with captures of Munida spp. (Decapoda: Anomura) in the Beagle Channel, Argentina. Scientia Marina, 68 (2): 2237-246
Piriz, M. 1972. Los organismos incrustantes de las costas argentinas. II. Estudio preliminar de la ficoflora y fauna asociada a Ulva lactuca L. (Clorophyta) en el puerto de Mar del Plata. LEMIT-Anales 4-1972: 73-98.
Pousa, J. L., W. C. Dragani, C. A. Mazio & N. W. Lanfredi, 1995. La energía oceánica en el Atlántico Sudoccidental. Posibilidades e impacto ambiental. Thalassas, 11: 59-72.
Rascio, V., J. Caprari & R. Bastida. 1969. Etudes sur les peintures sous-marines: Contribution a l’etude du comportament des peintures anti-salissures. II Influence de la concentration de toxiques. Pigments et Vernis, 45 (11): 724-735.
Roux, A.M, Fernández, M. & C. Bremec, 1995. Estudio preliminar de las comunidades bentónicas de los fondos
de pesca de langostino patagónico del Golfo San Jorge (Argentina). Ciencias Marinas 21 (3): 295-310.
Rützler, K. 1970. Spatial competition among Porifera: solution by epizoism. Oecologia, 5: 85-95.
Sosa, B., 2006. Efectos comunitarios y poblacionales de la ingeniería ecosistémica. El rol de la hormiga Atta vollenweideri en Sabanas arboladas del Litoral Oeste Uruguayo. Unpublished Thesis, 79 pp.
Stebbing, A. 1971, The epizoic fauna of Flustra foliacea (Bryozoa). J. Mar. Biol. Ass. U. K., 51: 283-300.
Stebbing, A. 1972. Preferential settlement of a bryozoan and serpulid larvae on the younger parts of Laminaria fronds. J. Mar. Biol. Ass. U. K., 52: 765-772.
Vinuesa, J. & M. Varisco, 2007. Trophic ecology of the lobster krill Munida gregaria in San Jorge Gulf, Argentina. Invest. Mar., Valparaiso, 35 (2):25-34.
Vozza, O., D.A. Sabio, , N.F. Randich, H.C. Martínez, A.S. Strada, S.M.C. Allevato, M. M. Etchegoin & R. Giussi, 1974. Sedimentología de la plataforma continental argentina. Texturas. Publicación del Servicio de Hidrografía Naval, H 669/1, Buenos Aires, 25 pp.
Ward, L. G., 1985. The influence of wind waves and tidal currents on sediment resuspensión in Middle Chesapeake bay. Geo-Marine Letters 5:71-75.
Williams, G. 1964. The effect of extracts of Fucus serratus in promoting settlement of larvae of Spirorbis borealis (Polychaeta). J. Mar. Biol. Ass. U. K., 44: 379-415.
Wilson, D. 1952. The influence of the nature of the substratum on the metamorphosis of the larvae of marine animals, specially the larvae of Ophelia bicornia. Annals Institute Oceangraphique Monaco, 27: 49-156.
(Received: January, 22, 2012; Accepted: April, 30, 2012)

DISTRIBUTION OF INLAND SILVERSIDES Menidia beryllina (COPE) AS A FUNCTION OF WAVE CREST VELOCITY AT
PELICAN ISLAND, CHINCOTEAGUE INLET, VIRGINIA, USA
Department of Biology and Health Services, Edinboro University, Edinboro, PA 16444, USA*Corresponding author: Tel: 814-732-2353; E-mail: [email protected]
Thalassas, 28(2) · July 2012: 57-63An International Journal of Marine Sciences
Key words: Surf zone, fi sh distribution, wave exposure, wave energy, sandy beach, estuaries.
Palabras clave: zona de rompientes, distribución de peces, exposición al oleaje, energía de las olas, playa de arena, estuario..
CRAIG W. STEELE*, PATRICK R. PONTZER & JOHN ANTONELLI
57Thalassas, 28(2) · July 2012
ABSTRACT
The present study examines the size distribution of inland silversides, Menidia beryllina, as a result of wave crest velocity around Pelican Island, a sand spit island located near the mouth of Chincoteague Inlet, Virginia, USA. The landward side of the island is separated from Wallops Island by Wallops Channel and is protected from wave action, while the seaward side faces Assateague Island (across the inlet mouth) and the Atlantic Ocean, with concomitant heavy surf action that decreases from the inlet mouth. Measurements were taken of wave crest height at each of six sampling site. Water temperature, salinity, and dissolved oxygen measurements were also taken at each sites (no significant differences among sites were found for any of these variables). Fish were collected using three seine hauls per site during each collecting period (n = 50 fish per site, minimum). All water sampling and fish collecting were conducted during three consecutive afternoon high tides. Results of linear regression and analysis of variance of crest velocity and fish length, and of crest velocity and fish weight indicate that larger inland silversides are found predominantly in areas of lesser wave energy, while smaller inland silversides are found predominantly in areas of greater wave energy.
RESUMEN (Distribución del pejerrey de río Menidia beryllina, en función de la velocidad en cresta de la ola en Pelican Island, ChincoteagueInlet, Virginia, EE.UU.)
El presente estudio examina la distribución de tamaños de pejerrey de río, Menidiaberyllina, como resultado de la velocidad de cresta de la ola en los alrededores de Pelican Island, un banco de arena cerca de la boca de entrada de Chincoteague, Virginia, EE.UU.. El lado de tierra de la isla está separada de la isla Wallops Island por el canal Wallops y está protegida de la acción de las olas, mientras que del lado del mar está frente a la Isla de Assateague (a través de la boca de entrada) y el Océano Atlántico, con la consiguiente exposición a oleaje fuerte que disminuye a medida que se aleja de la boca de entrada.Se midió la altura de cresta de la ola en seis puntos de muestreo. Además se midieron temperatura del agua, salinidad y oxí-geno disuelto en los mismos puntos, no apareciendo diferencias estadísticamente significativas entre los sitios para ninguna de estas variables. Los peces fueron recolectados a través de tres lances de cerco por lugar de muestreo durante cada período de recolección (n = 50 peces por sitio, como mínimo). Todos los muestreos de agua y recolección de los peces se llevaron a cabo durante tres mareas altas consecutivas por la tarde. Los resultados de la regresión lineal y análisis de la varianza de la velocidad de cresta y la longitud del pez, y de la velocidad de cresta y el peso de los peces indican que los peces grandes de pejerrey de río se encuentran predominantemente en áreas de menor energía de las olas, mientras que los más pequeños se encuentran predominantemente en las zonas de mayor energía de las olas.
CRAIG W. STEELE, PATRICK R. PONTZER & JOHN ANTONELLI
INTRODUCTION
Numerous studies have examined the effects of surf zone dynamics on fish assemblages, with results indicating that physical factors often have an overriding influence on fish distribution patterns (Clark, 1997; Fulton et al., 2005; Nanami, 2007; and e.g. see Romer, 1990; Clark et al., 1996a, 1996b; Harris & Cyrus, 1996). Layman (2000), however, insists that the shallow surf-zone (<0.4 m) is a little-studied micro-habitat of the ocean surf.
Surf zone fish assemblages are characterized by great variability at multiple scales in space and time as a consequence of abiotic and biotic factors (Gaelzer & Zalmon, 2008). For example, Gaelzer and Zalmon (2008) determined that increased fish biomass and species diversity were positively correlated with greater food availability and greater tide height among three Brazilian sandy beaches. Felix et al. (2007) also examined the effects of wave action in Brazilian sandy beach surf zones and its influence on species composition. They found that the ichthyofaunal composition of sheltered and moderately exposed beach samples were similar, with depth appearing to exert the major influence on ichthyofauna community structure, presumably by providing less turbulent waters that might have permitted shoals maintenance. Work by Layman (2000) indicates that, in some instances, increased species richness and
overall fish abundance suggests that fishes move parallel to the beach faces, seeking sheltered runnel habitats. Conversely, Felix et al. (2007) also determined that exposed beach samples differed largely from the other beaches and were influenced primarily by wave height and salinity differences.
According to Denny (1994), wave energy is a prominent physical factor in sub-tidal and intertidal marine ecosystems (as well as some aquaculture situations, e.g., Srivastava et al., 1991)) and correlations between wave energy and species distribution patterns are well documented. For example, Clark (1997), working on the South African coast, determined two important trends in the teleost catches: overall fish abundance increased significantly as wave exposure decreased; and greatest species richness and diversity (and lowest dominance) were seen at intermediate levels of exposure. Similarly, Wilbur et al. (2003) found that fish abundance and richness were greater closest to sheltering rock groins and Beyst et al. (2002) found that spatial variation in some fish assemblages is mainly correlated with local characteristics such as turbidity of the water and the morphodynamic features of both the beach and the adjacent subtidal area. Clark (1997) also discovered a positive relationship between fish size and exposure for at least some species. They attributed these trends primarily to the availability and abundance of food, and to physical disturbance from breaking waves. Fulton et
58 Thalassas, 28(2) · July 2012
Figure 1:Sampling sites, Pelican Island, Chincoteague Inlet, Virginia. Abbreviations: W.I. = Wallops Island; C.I. = Chincoteague Island; A.I. = Assateague Island.
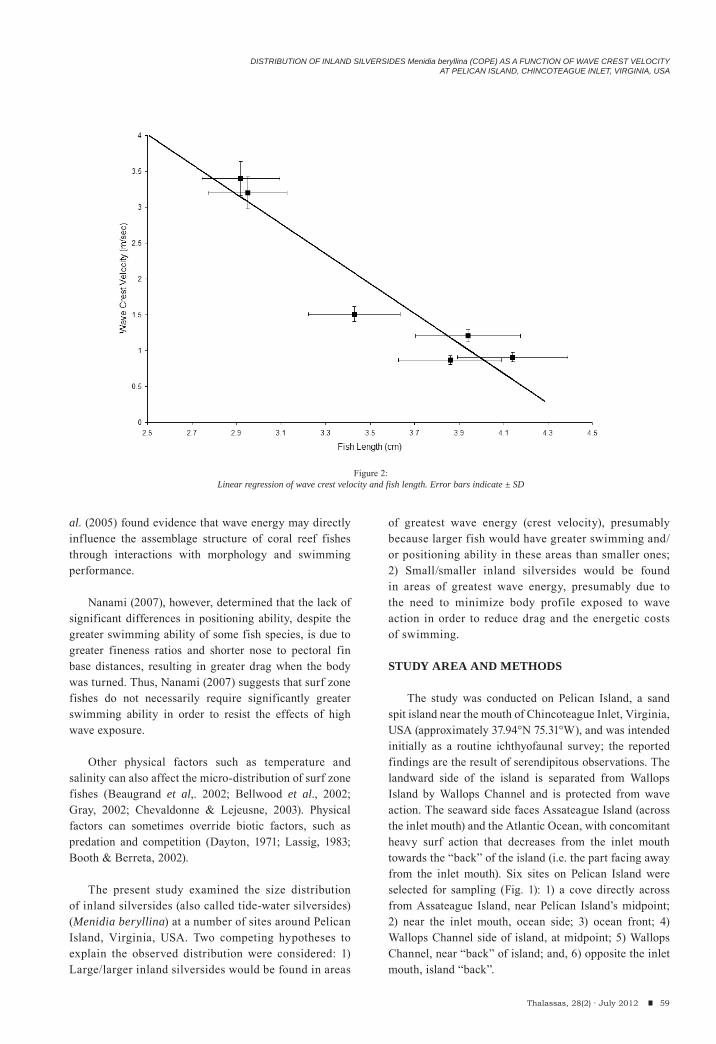
DISTRIBUTION OF INLAND SILVERSIDES Menidia beryllina (COPE) AS A FUNCTION OF WAVE CREST VELOCITYAT PELICAN ISLAND, CHINCOTEAGUE INLET, VIRGINIA, USA
al. (2005) found evidence that wave energy may directly influence the assemblage structure of coral reef fishes through interactions with morphology and swimming performance.
Nanami (2007), however, determined that the lack of significant differences in positioning ability, despite the greater swimming ability of some fish species, is due to greater fineness ratios and shorter nose to pectoral fin base distances, resulting in greater drag when the body was turned. Thus, Nanami (2007) suggests that surf zone fishes do not necessarily require significantly greater swimming ability in order to resist the effects of high wave exposure.
Other physical factors such as temperature and salinity can also affect the micro-distribution of surf zone fishes (Beaugrand et al,. 2002; Bellwood et al., 2002; Gray, 2002; Chevaldonne & Lejeusne, 2003). Physical factors can sometimes override biotic factors, such as predation and competition (Dayton, 1971; Lassig, 1983; Booth & Berreta, 2002).
The present study examined the size distribution of inland silversides (also called tide-water silversides) (Menidia beryllina) at a number of sites around Pelican Island, Virginia, USA. Two competing hypotheses to explain the observed distribution were considered: 1) Large/larger inland silversides would be found in areas
of greatest wave energy (crest velocity), presumably because larger fish would have greater swimming and/or positioning ability in these areas than smaller ones; 2) Small/smaller inland silversides would be found in areas of greatest wave energy, presumably due to the need to minimize body profile exposed to wave action in order to reduce drag and the energetic costs of swimming.
STUDY AREA AND METHODS
The study was conducted on Pelican Island, a sand spit island near the mouth of Chincoteague Inlet, Virginia, USA (approximately 37.94°N 75.31°W), and was intended initially as a routine ichthyofaunal survey; the reported findings are the result of serendipitous observations. The landward side of the island is separated from Wallops Island by Wallops Channel and is protected from wave action. The seaward side faces Assateague Island (across the inlet mouth) and the Atlantic Ocean, with concomitant heavy surf action that decreases from the inlet mouth towards the “back” of the island (i.e. the part facing away from the inlet mouth). Six sites on Pelican Island were selected for sampling (Fig. 1): 1) a cove directly across from Assateague Island, near Pelican Island’s midpoint; 2) near the inlet mouth, ocean side; 3) ocean front; 4) Wallops Channel side of island, at midpoint; 5) Wallops Channel, near “back” of island; and, 6) opposite the inlet mouth, island “back”.
59Thalassas, 28(2) · July 2012
Figure 2:Linear regression of wave crest velocity and fish length. Error bars indicate ± SD
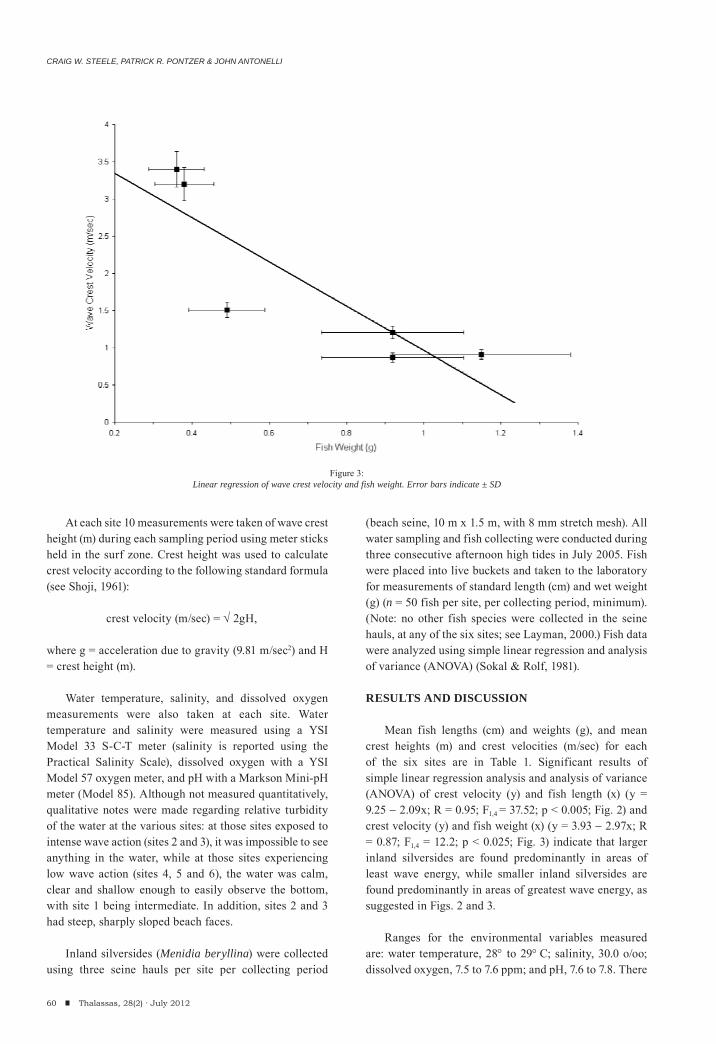
CRAIG W. STEELE, PATRICK R. PONTZER & JOHN ANTONELLI
At each site 10 measurements were taken of wave crest height (m) during each sampling period using meter sticks held in the surf zone. Crest height was used to calculate crest velocity according to the following standard formula (see Shoji, 1961):
crest velocity (m/sec) = 2gH,
where g = acceleration due to gravity (9.81 m/sec2) and H = crest height (m).
Water temperature, salinity, and dissolved oxygen measurements were also taken at each site. Water temperature and salinity were measured using a YSI Model 33 S-C-T meter (salinity is reported using the Practical Salinity Scale), dissolved oxygen with a YSI Model 57 oxygen meter, and pH with a Markson Mini-pH meter (Model 85). Although not measured quantitatively, qualitative notes were made regarding relative turbidity of the water at the various sites: at those sites exposed to intense wave action (sites 2 and 3), it was impossible to see anything in the water, while at those sites experiencing low wave action (sites 4, 5 and 6), the water was calm, clear and shallow enough to easily observe the bottom, with site 1 being intermediate. In addition, sites 2 and 3 had steep, sharply sloped beach faces.
Inland silversides (Menidia beryllina) were collected using three seine hauls per site per collecting period
(beach seine, 10 m x 1.5 m, with 8 mm stretch mesh). All water sampling and fish collecting were conducted during three consecutive afternoon high tides in July 2005. Fish were placed into live buckets and taken to the laboratory for measurements of standard length (cm) and wet weight (g) (n = 50 fish per site, per collecting period, minimum). (Note: no other fish species were collected in the seine hauls, at any of the six sites; see Layman, 2000.) Fish data were analyzed using simple linear regression and analysis of variance (ANOVA) (Sokal & Rolf, 1981).
RESULTS AND DISCUSSION
Mean fish lengths (cm) and weights (g), and mean crest heights (m) and crest velocities (m/sec) for each of the six sites are in Table 1. Significant results of simple linear regression analysis and analysis of variance (ANOVA) of crest velocity (y) and fish length (x) (y = 9.25 2.09x; R = 0.95; F1,4 = 37.52; p < 0.005; Fig. 2) and crest velocity (y) and fish weight (x) (y = 3.93 2.97x; R = 0.87; F1,4 = 12.2; p < 0.025; Fig. 3) indicate that larger inland silversides are found predominantly in areas of least wave energy, while smaller inland silversides are found predominantly in areas of greatest wave energy, as suggested in Figs. 2 and 3.
Ranges for the environmental variables measured are: water temperature, 28 to 29 C; salinity, 30.0 o/oo; dissolved oxygen, 7.5 to 7.6 ppm; and pH, 7.6 to 7.8. There
60 Thalassas, 28(2) · July 2012
Figure 3:Linear regression of wave crest velocity and fish weight. Error bars indicate ± SD
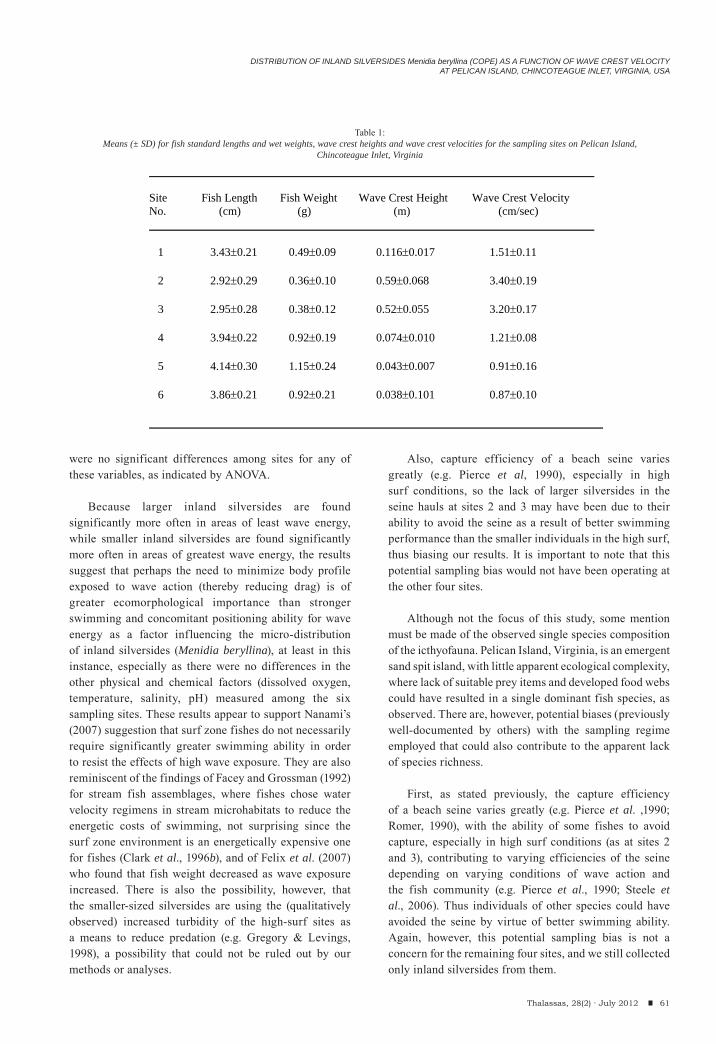
DISTRIBUTION OF INLAND SILVERSIDES Menidia beryllina (COPE) AS A FUNCTION OF WAVE CREST VELOCITYAT PELICAN ISLAND, CHINCOTEAGUE INLET, VIRGINIA, USA
61Thalassas, 28(2) · July 2012
were no significant differences among sites for any of these variables, as indicated by ANOVA.
Because larger inland silversides are found significantly more often in areas of least wave energy, while smaller inland silversides are found significantly more often in areas of greatest wave energy, the results suggest that perhaps the need to minimize body profile exposed to wave action (thereby reducing drag) is of greater ecomorphological importance than stronger swimming and concomitant positioning ability for wave energy as a factor influencing the micro-distribution of inland silversides (Menidia beryllina), at least in this instance, especially as there were no differences in the other physical and chemical factors (dissolved oxygen, temperature, salinity, pH) measured among the six sampling sites. These results appear to support Nanami’s (2007) suggestion that surf zone fishes do not necessarily require significantly greater swimming ability in order to resist the effects of high wave exposure. They are also reminiscent of the findings of Facey and Grossman (1992) for stream fish assemblages, where fishes chose water velocity regimens in stream microhabitats to reduce the energetic costs of swimming, not surprising since the surf zone environment is an energetically expensive one for fishes (Clark et al., 1996b), and of Felix et al. (2007) who found that fish weight decreased as wave exposure increased. There is also the possibility, however, that the smaller-sized silversides are using the (qualitatively observed) increased turbidity of the high-surf sites as a means to reduce predation (e.g. Gregory & Levings, 1998), a possibility that could not be ruled out by our methods or analyses.
Also, capture efficiency of a beach seine varies greatly (e.g. Pierce et al, 1990), especially in high surf conditions, so the lack of larger silversides in the seine hauls at sites 2 and 3 may have been due to their ability to avoid the seine as a result of better swimming performance than the smaller individuals in the high surf, thus biasing our results. It is important to note that this potential sampling bias would not have been operating at the other four sites.
Although not the focus of this study, some mention must be made of the observed single species composition of the icthyofauna. Pelican Island, Virginia, is an emergent sand spit island, with little apparent ecological complexity, where lack of suitable prey items and developed food webs could have resulted in a single dominant fish species, as observed. There are, however, potential biases (previously well-documented by others) with the sampling regime employed that could also contribute to the apparent lack of species richness.
First, as stated previously, the capture efficiency of a beach seine varies greatly (e.g. Pierce et al. ,1990; Romer, 1990), with the ability of some fishes to avoid capture, especially in high surf conditions (as at sites 2 and 3), contributing to varying efficiencies of the seine depending on varying conditions of wave action and the fish community (e.g. Pierce et al., 1990; Steele et al., 2006). Thus individuals of other species could have avoided the seine by virtue of better swimming ability. Again, however, this potential sampling bias is not a concern for the remaining four sites, and we still collected only inland silversides from them.
Site Fish Length Fish Weight Wave Crest Height Wave Crest Velocity No. (cm) (g) (m) (cm/sec)
1 3.43 0.21 0.49 0.09 0.116 0.017 1.51 0.11
2 2.92 0.29 0.36 0.10 0.59 0.068 3.40 0.19
3 2.95 0.28 0.38 0.12 0.52 0.055 3.20 0.17
4 3.94 0.22 0.92 0.19 0.074 0.010 1.21 0.08
5 4.14 0.30 1.15 0.24 0.043 0.007 0.91 0.16
6 3.86 0.21 0.92 0.21 0.038 0.101 0.87 0.10
Table 1:Means (± SD) for fish standard lengths and wet weights, wave crest heights and wave crest velocities for the sampling sites on Pelican Island,
Chincoteague Inlet, Virginia

CRAIG W. STEELE, PATRICK R. PONTZER & JOHN ANTONELLI
62 Thalassas, 28(2) · July 2012
Second, temporal variations in fish abundance and in fish assemblages, due to diel activity patterns (e.g. Ross et al., 1987; Romer, 1990; Clark et al., 1996b; Layman, 2000), tide condition (low versus high) (e.g. Harvey, 1998; Layman, 2000) could also be contributing to the observed single species “assemblage”. Diel variation in the composition of fish assemblages in shallow, nearshore zones is well known in both freshwater and marine systems (e.g. Morrison et al., 2002; Blackwell & Brown, 2005; Reid & Mandrak, 2009). For example, Layman (2000), in his study of Virginia barrier islands, found an increased species richness at night. A variety of reasons, including environmental factors (e.g. oxygen gradients, tidal condition), feeding patterns and predator avoidance (e.g. Layman, 2000; Koutrakis et al., 2005; see review by Reid & Mandrak, 2009) and/or susceptibility to sampling gear (e.g. Pierce et al,. 2001; Gries & Letcher, 2002; Steele et al., 2006) have been proposed to account for these observations.
Third, the intensity of sampling effort greatly affects the interpretation of the fish assemblage (e.g. Romer, 1990; Wilber et al., 2003; Reid & Mandrak, 2009). However, we were limited, logistically, in our access to the island and could not increase our sampling effort to assess this or the other potential biases in our sampling results. Their resolution must await further research.
ACKNOWLEDGEMENTS
This research was conducted while the authors were in residence at the Wallops Island Marine Science Center, Wallops Island, Virginia, USA. We thank Emily Steele and Ryan Steele for assistance with the drawing of Fig. 1.
REFERENCES
Beaugrand G, Reid PG, Ibanez F, Lindley JA, Edwards M (2002). Reorganization of North Atlantic marine copepod biodiversity and climate. Science 296: 16921694.
Bellwood DR, Wainwright PC, Fulton, CJ, Hoey, A (2002). Assembly rules and functional groups at global biogeographical scales. Functional Ecology 16: 557562.
Blackwell BG, Brown ML (2005). Comparison of day and night shoreline seine catches in two South Dakota glacial lakes. Jouornal of Freshwater Ecology 20: 7983.
Booth DJ, Beretta GA (2002.) Changes in a fish assemblage after a coral bleaching event. Marine Ecology Progress Series 245: 205212.
Beyst B, Hostens K, Mees J (2002). Factors influencing the spatial variation in fish and macrocrustacean communities in the surf zone of sandy beaches in Belgium. Journal of the Marine Biological Association of the UK 82(2): 181187.
Chevaldonne P, Lejeusne C (2003). Regional warming induced species shift in northwest Mediterranean marine caves. Ecological Letters 6: 371379.
Clark BM (1997). Variation in surf-zone fish community structure across a wave-exposed gradient. Estuarine Coastal and Shelf Science 44(6): 659674.
Clark BM, Bennett BA, Lamberth SJ (1996a). Factors affecting spatial variability in seine net catches of fish in the surf zone of False Bay, South Africa. Marine Ecology Progress Series 131: 1734.
Clark BM, Bennett BA, Lamberth SJ (1996b). Temporal variations in surf zone fish assemblages from False Bay, South Africa. Marine Ecology Progress Series 131: 3547.
Dayton PK (1971). Competition, disturbance, and community organization: the provision and subsequent utilization of space in a rocky intertidal community. Ecological Monographs 41: 351389.
Denny MW (1994). Roles of hydrodynamics in the study of life on wave-swept shores. In: PC Wainwright, SM Reilly, eds, Ecological Morphology: Integrative Organismal Biology, University of Chicago Press, Chicago, 169204.
Facey DE, Grossman GD (1992). The relationship between water velocity, energetic costs, and microhabitat use in four North American stream fishes. Hydrobiologia 239: 16.
Felix CF, Spach HL, Moro PS, Hackradt CW, Nogueria de Queiroz G MacL, Hostim-Silva M (2007). Ichthyofauna composition across a wave-energy gradient on southern Brazil beaches. Brazilian Journal of Oceanography 55(4): 281292.
Fulton CJ, Bellwood DR, Wainwright PC (2005). Wave energy and swimming performance shape coral reef fish assemblages. Biological Science 272(1565): 827–832.
Gaelzer LR, Zalmon IR (2008). Tidal influence on surf zone ichthyofauna structure at three sandy beaches, southeastern Brazil. Brazilian Journal of Oceanography 56(3): 165177.
Gray JS (2002). Species richness of marine soft sediments. Marine Ecology Progress Series 244: 285297.
Gries G, Letcher BH (2002). A night seining technique for juvenile Atlantic salmon in streams. North American Journal of Fisheries Management 22: 595601.
Gregory RS, Levings CD (1998). Turbidity reduces predation on migrating juvenile Pacific salmon. Transactions of the American Fisheries Society 127: 275285.
Harris SA, Cyrus DP (1996). Larval and juvenile fishes in the surf zone adjacent to the St Lucia estuary mouth, KwaZulu-Natal, South Africa. Marine and Freshwater Research 47: 465482.
Harvey C. (1998). Use of sandy beach habitat by Fundulus majalis, a surf-zone fish. Marine Ecology Progress Series 164: 307310.
Koutrakis ET, Tsikliras AC, Sinis AI (2005). Temporal variability of the ichthyofauna in a northern Aegean coastal lagoon (Greece). Hydrobiologia 543: 245257.
Layman CA (2000). Fish assemblage structure of the shallow

DISTRIBUTION OF INLAND SILVERSIDES Menidia beryllina (COPE) AS A FUNCTION OF WAVE CREST VELOCITYAT PELICAN ISLAND, CHINCOTEAGUE INLET, VIRGINIA, USA
63Thalassas, 28(2) · July 2012
ocean surf-zone on the eastern shore of Virginia barrier islands. Estuarine Coastal and Shelf Science 51: 201213.
Lassig BR (1983.) The effects of a cyclonic storm on coral reef fish assemblages. Environmental Biology of Fishes 9: 55-63.
Morrison MA, Francis MP, Hartill BW, Parkinson DM (2002). Diurnal and tidal variations in the abundance of the fish fauna of a temperate tidal mudflat. Estuarine Coastal and Shelf Science 54: 793807.
Nanami A (2007). Juvenile swimming performance of three fish species on an exposed sandy beach in Japan. Journal of Experimental Marine Biology and Ecology 346: 110.
Pierce CL, Corcoran AM, Gronbach AN, Hsia, S, Mullarkey BJ, Schwartzhoff AJ (2001). Influence of diel period on electrofishing and beach seining assessments of litoral fish assemblages. North American Journal of Fisheries Management 21: 918926.
Pierce CL, Rasmussen JB, Leggett WC (1990). Sampling littoral fish with a seine: corrections for variable capture efficiency. Canadian Journal of Fisheries and Aquatic Sciences 47: 10041010.
Reid SM, Mandrake NE (2009). Lake Erie beaches: diel variation in fish assemblage structure and implications for monitoring. Hydrobiologia 618: 139-148.
Romer GS (1990). Surf zone fish community and species response to a wave energy gradient. Journal of Fish Biology 36: 279287.
Ross ST, McMichael RH, Ruple DL (1987). Seasonal and diel variation in the standing crop of fishes and macroinvertebrates from a Gulf of Mexico surf-zone. Estuarine Coastal and Shelf Science 25: 391412.
Shoji D (1961). On the variations of the daily mean sea levels along the Japanese islands. Journal of the Oceanographic Society of Japan 3: 141152.
Srivastava RK, Brown JA, Allen J (1991). The influence of wave frequency and wave height on the behaviour of rainbow trout (Oncorhynchus mykiss) in cages. Aquaculture 97: 143153.
Sokal RR, Rolf FJ (1981). Biometry, 2nd edn, W.H. Freeman, San Francisco, 859p.
Steele MA, Schroeter SC, Page HM (2006). Experimental evaluation of biases associated with sampling estuarine fishes with seines. Estuaries and Coasts 29(6B): 11721184.
Wilber DH, Clarke DG, Burlass MH, Ruben H, Will RJ (2003). Spatial and temporal variability in surf zone fish assemblages on the coast of northern New Jersey. Estuarine Coastal and Shelf Science 56: 291304.
(Received: October, 17, 2011; Accepted: February, 15, 2012)

An International Journal of Marine Sciences
Number 28 (2) July 2012
Thalassas






















![Dictionary wild animals [salva miralles]](https://img.pdfslide.us/doc/110x75/5594d4741a28ab66798b46a6/dictionary-wild-animals-salva-miralles.jpg)
![Dictionary wild animals [s. miralles][april 2012]](https://img.pdfslide.us/doc/110x75/5594d3281a28ab83798b45bd/dictionary-wild-animals-s-mirallesapril-2012-55954e985373f.jpg)





