Embed Size (px)
Citation preview

John F. Lehm!Ulhl, USDA Forest Service, Pacific NcrJiwest Research Staiion, Wenatchee, Washington 98801
Stephen D, West, Ccl!ege of Forest ResQlJrces, University of Washington, Box 352100, Seattle, Washington 98195
Carel L Chambers, Department of Forest Resources, Oregon State University, Corvallis, Oregon 97331' . . . Wllllarn C. McComb, Department or Fo~t Science, Oregon State University, Corvallis, Oregon 973312
David A. Manuv.ia1, Gcllege cf Forest Resources, Univ.ersity of W~hingtcn, Sox 352100, Seattle, Washingtcn 98195
_Keith B. Aubry, USDA Forest Service, Pacific Northwest Research Station, Olympia, Washington 98512 ·
Janet L Erlci<Son, Robert A. Gitzen, and Matthias Leu, College of Forest Reso·urces, University cf Washington Box 352100, Seattle, Wasciington 98195 · . '
An Experiment for Assessing Vertebrate Response to Varying Levels and Patterns of Green-tres Retention
Abstract
The emphasis of forest ~~eat in the Pacific Northwest bas sbil'ted recently from the prodllction of timber resolllCeS t01be ma:in~=c~ or resiorarion of biological ~ and ecosys'.em func:iOlling. New standan:ls aDd gwrlelin;:s for managerm:nt emphasize the retention of forest stroctun:s (lfre trees, logs, and snags} to te1lnce logging impaas, to earich reestablishe<i stmds with impo11:w sl;:UctDral feat=, and to enhance collll<Cti'l'ity across forest landscapes. However, .little is known aboi.·nhe eff:e,.:ts on wikilife of ya.rymg the level and spatia! distribution of :retained stmctn,-cs in f= of wesiem O,:egon and Was~n. Repli=<i and controlled experiments within the Demonstration of Ecosystem~= Options (DElY,!OJ stndy are beginniQg r.o quantify the effecrs of varying the Je,-e! and spatial aggxegation of green-tree xetention ilnting forest harvest on a variety of ecosys= components·(c.g., '""""O!]lleS. invertebrates, vegetation. fungi), as well as snow hydrology and socful perteptions of these types of ~eneration harvests. Eigl!t ~li= bloclll; of six cxpf:rimcntal rreatmerus have been established Cit the U.ropqoa Nation.al Forest in Oregon, and on the GifIOid Pinchot Nanon.al fo,:est and Capitol S121e Fo<tstin W~n. 'Die objec::i= of the wildlife stnc:lies are to <JU3lll!fy pattems of speci!'5 richness, eV10ImeSS, a.,d relativ,, abimriaoce of birds. sm.,n mammals, bl,ts. anci amphibians before and after ha.-vest to 'examine short-<=~ responses.. ~en,sampling bas b=i completed -on all sires, b;,rvest =tmerus are in progress, and post-==t sampling bas begun. In mis P"J!C!, we present an overview of oar hypotheses lllld methods, aDd ci=m.."J)t tlle o=e and relllltive abundance of speqies prior to MJ:Vest of the smdy blocks.
·Long-teem stadles of vmebr.!te n:sponse, habit!t associations, and trophic in=clions a:re pianned. Results will ittfbnn managers on the consequences of alternative forest man.agcmeot strai=gies.
Introduction
Forest:mana,,<>ementin thePaci:ficNortliwestduring most of the last 50 years has emphasiz....od the production of timber resources. Traditional practices of clearcut logging and artificial regeneration typica:riy produce simplified stand structures that differ from the structural, compositional, and.functional complexity of naturally regenerated fotestS, primarily in the absence of a significant legacy of large live and dead trees and down woody debris (Franklin et al. 1997). The loss of the old-growth
1 Cuaenc address: School ofFarestzy, North=Arizona Uni-ve,siiy, Flagstaff. Arizona.86011-SO!l! ·
'Current address: Department of Forestry and WildliJe Managemem, University of Massachusetts. Amherst, Massacb.asetls 01003
legacy and attendant forest complexity has negatively affected the viability of associated species (Hams 1984, Mannan andMeslow 1984,J f'hmknbl and Ruggiero 1991, Ruggiero et al 1991, FEMAT 1993) and has spurred the development of-new forest management strategies for species conservation (e.e:,, USDA and USDI 1994). - .
New approaches for manag.in!; public forest lands seek to remedy problems associated v,.ith past management practices by integrating the maintenance of ecological values with commod:ityproduction (DeBeiletal.1997, Kohmandfunklin 1997, Tappeiner et al 1997). Among these techDiques are green-tree o~v.iriableretention systems. Retention of green trees in harve~ stands bas often been translated. in\o the retention of a fe.w
Northwest Science, Vo). 73, Special Issue, 1999· 45

scattered "wildlife" trees, with Utt!e apparentben- . efitto v.ildlife (McComb et aL 1993b ). More sophisticated retention techniques that V?rJ the level aod spatial pattern of retained trees have been proposed to mimic the effects of -nan:u-a.I forest distllrbance, in p¥(icular the retention of large structurnl elements including live~. snags, and down wood (FEMAT 1993, Franklin et al 1997). T'ne objectives of these tecllniques are to .maintain ecosystem structure and function, emich re- · established stands with structural features $at would otherv.-ise be absent, and enhance connectivity across the landscape (Ftanldin et al. 1997). . Presumably, vertebrate species that persisted
within disrurbance regimes of the pre-European . · settlement period may be ·expected to persist in managed forests if those regimes are mimicked. However, it is unclear how species will respond to our attempts to mimic natural disturbance regimes, as there ·are few data on the relationships berween the levels and spatial patterns of greentree retention and the short-term pe~tence or the long-term recolonization of species in harvested stands (McComb et aL 1993a, Chambers 1996, Han.sen and Hounihan 1996, Franklin et aL 1997). Rettospectiveresearch.basfocusedon
. species found iJ;i. unmanaged. lale-sera! and youngey forests (e.g.,Raphael 1988, Ruggieroet al.1991, Rosenberg and Anthony 1993, McGarigal and McComb l995;Gomez and Anthony 1996, Hagar et aL 1996), recent cleaxcuts (Medin 1986), or stands with evenly-distributed partial retention of green trees (Vega 1993,Hansen et al 1995,Hansen andHounihan 1996) .. Retainingmore~n trees might maintain components of S'.and structure or function that will allow some species to persist ·in haryested areas, but iµay result in poorer persistence o;f other species (Hansen et al 1995, Hansen and Hollllihan 1996, Chambers et aL in press): Retaining trees in undisturbed refngial patches Jilrewise might =ult m smaller initial ·. impacts or faster long-term recovery, depending on the life history and population structure of the species. Although the results of relevant e;,cperimental steelies with pl'!'l- and post-treatment data are beginning to emerge (e.g., Chambers et al. in press), no studies have been published that examine the effects on vertebrate comml:mities of .simultaneously varying both the level and spatial pattern of green-tree retei:J.tidn (Franklin etal. 1997).
As a component of theDemonstraticn of Ecosystem Management Options .(DENIO) study
46 Lehmkuhl et al.
(Aubry et al 1999), we have been ·studying responses of forest vertebrate species to, expetimen
. tally manipulated levels {percentage of basal area) and patterns ( dispersed vs. aggregated) of greentreeretenticn. Ourprimary goals are to: (1) quantify short-term changes in community composition and abundance ofbirds, smaJl mammals, bats, and amplu'bians; (2) identify the vegetation and other habitat attn'bntes associated with observed changes iii species occmrence and abundance; and (3) quantify functional ttopb.ic relationships (e.g., fungivory) of select species groups.
In this paper we proviclt: an overview of our study area, describe our experimental and sampling designs, briefly review the ecology and habitat relationships of vertebrate groups targeted for stndy, describe hypothetical short-term (5-10 yr) responses of vertebrates to treatments and the analytical techniques to test these hypotheses; briefly summarlze pre-treatment results; and describe the challenges and limitations of implementing an experimental study of this scope on 'vertebrates. This -paper does not contain a detailed analysis .of pre, treatmcnt data. Hypothetical responses of vertebrates to the retention treacnents are based on current knowledge of species' ecologies ancj !he hypothesized responses of habitat elements to these treatments as descnoed by Halpern.et al (1999). Aubry et aL (1.999) provide detailed di:· scriptions of overall project goals, ex-~enral design and harvest treatments,. and the full scope of DElvIO research (vegetation, fungi, invertebrates, social perceptions, sn~w hydrology).
Methods
Study Area
Study sites occ_ur in Douglas-fir (Pseudotsuga .menziesiz) dominated forests on the Umpqua National Forest in westem Oregon, and on the Gifford Pinchot National Forest and Capitol State Forest .in westem Washington (see Aubry et aL 1999). The eight study. biocks (four in each state) provide a broad geographical and e.cological scope ofinference(AnbryetaL 1999). Within each. block, six study units were selected from the same or adjacent watersheds to reduce differences in physjographic and ,stand characteristics, and to reduce differences re~-ulting .from past management li1 contrast, blocks vary v..idely in biophysical set
. ting and past management Variation within and

among blocks is accounted for in the samoling design and analytical tedmiques (see belo;), · . .
.A,,mOJJg tjie blocks, study sit.es range from 200-1700 m in elevation, slopes vary from steep to gentle, and nearly all aspects. are represented. Stands encompass a variety of disturbance histo-
. .ries, ages, and successional ·stages, but all have developed beyond the stem-exclusion stage of · forestdevelopment(sensuO!iverandlazson 1990). All stands on the Umpqua Nati0I1al I:orest hav:e been thinned or salvaged to some degree, whereas · th9se on the Gifford Pinch9t National Forest are allruttnral nnmaoagedforests. Two blocks (Layng ·Creek, Oregon and Capitol Stat.e Forest, Washington) are second growth as aresitlt.of previous harvest. Forest understories are compositionally diverse, but many blocks share the same dominant herbaceous and woody species. Halpern ·et al. (1999) provide detailed site descriptions am! an overview of _the variation among blocks.
Our results sho~d be applicable to portions of the Wes!em Cascade.and Coast Ranges with similar biophysical conditions and species distributions. Many, but ·not all, of the veiteb.J:ate species of .interest occur throughout both regions. Where biogeograpbic variation in species distcibutions and responses to treatments may· occur ( e.g., Carer and_Johnson 1995), the block design of'the experiment will help to control for some regional differences in treatment effects.
Experimental Design ·
A randomized block design allows us to control for biophysical differences across the .regi,onal study area At eac)l. of eight study bl~ks (replicates) five harvest treatments and a control were randomly· assigned to 13-ba treatment units. After extensive survey of candidate stands across the potential study area, 13 ha proved to be the iµaximum stand size that could accommodate the mizµmum s~ling grid (descn'bed below) and provide for homogenous within-stand 'biophysical conditions. Treatments vary in tb.e level of retention of live trees (15-100% basal area) and in the pattern of retention (trees uniformly dispersed vs. aggregated). The six treatments are: 100% retention (control); 75% aggregatedretenrion (trees harvested in three circular, I-ha gaps); 40% dispersed retention throughout the stand; 40% aggregated retention ( as five undisturbed, circular, 1-hapatches); 15% dispersed retention; and 15%
aggregated retention (as two und.istmbed, circular, 1-ha patches). All snags will be retamed in forest aggregates. In harvested portions of treatmentnnits, existing snags wi)Ibe i:etairied where possible,. and 65 sna,,as/ba will be created from live trees (see Aubry et al. [1999] for details).
Sampling.Design and AnalytlcaJ Methods
Sampling for vertebrates and. vegetatio~ v,itbin each treatment nnit OCCUIIed on a permanently marked 8 x 8 or 7 x 9 grid, depending on the shape ofthestand, with40-mspacingbetweengridpoints and a 40-m buffer to the edge of the treatment unit. This grid configuration was chosen to conform to sampling methods for arboreal rodents, which reqnfre 'the largest sampling area among all species stodiea, and to allow for systematic sampling of treatments. Carey et al. (1991) recommend 10 x 10 grids with 40-m spacing (16 ha) to estimate the density of northern flying squirrels (Glaucomys sabrinu.s), butindicatethatsmaller grids·(e.g., 8 x 8 or7 x9, [13 haD are adequate if .the goal is to calculate relative abundance indi.ces Iljther than density (see $o Carey et al. [1996}). Be----ause lamer !llids could not be accommodated - ~ .
. given tf?.e sizes of intact stands; we accepted the smaller grid (see Abbott eral. [19991 Aubry et al !1999] for additional details): .
Field methods w~re design,ed to inde" habitat use by numbers ·of detections or individuals captured. We assume fua1 use will be strongly correlated with changes in abundance for most' spe-· cies ormterest. forsomespecies w:ithlarge:ranges of movement relative to the area·of treatment units ( e.g., bats and some birds), nse may not be closely ·tied to clianges in abundance. Thus,. the most ecologically meaningfnlinformation will be gained from species that occur at moderate to bigb densities and that e--.Jn'bit local patterns of movement relative to the size of treatment units.
Data were collected for 2 yr prior to treatment to quantify species occurrences and relative abundances. Comparison of pre- with post-treatment data in testing treatment responses will allow us to controf for spatial variation in vertebrate occurrence and abundance among ti:eattnent units within blocks. Comnarison of databetween treatment and control units will control for temporal variation. Although comparisons ofpre-and posrtreatment data from manipulated and control stands enable us to separate annual variation iii species
Ve.rtebrate Response to Green~tree Retention 47

abundances from the effects of har-vest treatments , . an understanding oflong-term (> 5 yr) responses· will require repeated measurements over longer time frames. ;
We analyzed the power of the experimental design to detect treatment effects on a subset of
· bird-ai,d small mammal species. We chose a fo- . cal species (norihem flying squirrel), four common and one wide-ranging bird species, and one moderately abundant terrestrial small roaroma)
· species that we knew to be well S3JJID!ed and could be anticipated to be either ~atly or minimally · affected.by treatments . . We used pre-treatment data to .estimate mean and variance· of captures. Treatment response was lzypothesized10 be at least a 40% (birds) or 50% (small roru:uroa)s) reduction in caotures between the controi'and one treatment-4e 40% disp~ed retention treatmentbased on the interactions of indhicfua) species life histories, habirat relationships, and the level of canopy removal. Nearly all tteatmeriteffects are expected to result in greater declines; thus, the test understates the expected magnitude and fre.. quency of treatment effects and gives conserva-tive power estimates ·
Treatment effects on.individual species abun. dances (for spe,.,'ies meeting minimum criterla tor occurrence or abundance in tr~ent units) and on colillllUllity attributes of species richness and evenness will be rested as the difference between
· pre- and post-treatment valnes among treatment and control units using randomized block analysis of variance CANOVA) (Skalski and Robson 1992) ornonparametricanalogs. Repeated-mea-· sures ANOVA will be 11Sro to test for long-t= treatment effects. Si.riJiJ.arity in faunal associations among and within· tr:"...atments will be examined by clustering or ordination techniques. Relationships between vegetation or. bther habitat a~'butes (e.g., volume of woody debris) and species presence or-abundance will be examined across all replicates primarily by regressfori. analysis ..
Hypotheses, Methods, and Prellmina.'1' Results · ·
Birds
Review
Some bird populations are declining in the Pacific Northwest (Sharp 1990, 1992), in pan as a result of timber harvesting that shifts tree age-
43 Lehroknhl et al.
class clistdbutions from oid to young and alters natural disturl:lance regunes (Franklin and Forman 1987, Spies and Franklin 1989, FElYLA..T 1993). There are concerns for cavity-nesting birds. (Morrison et al. 1986, Chambers et al_ 1997) neotropical'mig:rants (Eagan and Johnston 1992' Martin and Finch 1995), the north.em spotted owi (Strix occidentalis caurina) (Forsman et al. 1996); and marbl~d murrelet (Brachyramphus mamwratus) (Carter and Morrison 1992; Nelson and Sealy 1995, Ralph et aL 1995) that have fueled public.debate over forest manaaement in the Pacific Northwest ·· "
Significant research 1).as been devo!e:d to bird . populations in llllilll:al, unmanaged Douglas-fir ·forests of west.em Washington and Oregon (Carey et al 1991b, Gilbert andAllwine 1991a, Haff et al 1991, Manuwal 199.1), and there have ~ some retrospective studies of birds in managed forests of the region (Artman 1990, McGarigal and McComb 1992, Hansen etaL 1995, Ha!?aret
. b al.1996, Hlmsenand.Houniban 1996). However, replicatedmanipulationsthatincludepre-andpostharvest sampling (e.g., Bosakowsld 1997, C'nambers and McComb 1997, Chambers et at in press) have been limited in size and scope.
Bird species associated with mid- to late-successional forests that use large trees for foraging or as nest sites (Brown 1985) typically decline in response to harvest (Franzreb ·1977, Keller and Anderson 1992, Chambers 1996, Hansen and Hollllihan 1996), although. intensity qf ha;-vest · affects responses differently. Compared with clea:rcutting, 25.%retention of ~n-treebasal area offers benefits (foraging and nesting habitat) to some of the species associated with old-growth forests (Chambers 1996, Chambers eta!. inpres.s).
Neither Chambers, (1996) no/Vega (1993) detected a si.::,anificant change in bird species richness amongretention-treatment, late-successfonal forest, and cl~t stands 2:8 hain area, although the composition of the bird community changed after harvest However, ret.ention stands provided bi:eeding habitat both for birds associated with early seral stages and for some species associated with late-sUC1:eSsional for"...St Tbe retention of large trees apparently proVIded foraging substrates and in some cases breeding sites for some species associated with old~growth forest However, less intensive harvesting (approximately 30% reduction in wood volume) resulted in decline or loss of fewer bird species associated with late-

successional forests (Medin and Booth 1989, Chambers 1996, Chaµiliers etal. in press). Thus, a variety of stand treatments may be necessary to maint.µn habitat for different species.
Hypotheses
Hvoothesis 1-.-4.bundance of canopy-dwelling . birds will decline with'decreasi.ng levels of tree retention; aggregation of retention11,ill reduce this effect (Table I). Species whose _primary habitat is closed:-canopy forest, such as heonit/Ibwnsend's warbler (Dendroica occide!z,talis/D. tQwnsendi') and the golden-<:rowned kinglet (Regulus satrapa), likely will decline .in abundance with the. loss of canopy habitat Aggregation ofpatches will allow for some persistence m treatment units . .
~.danceofbirds.associatea with JIJUierstory vegetation will decline ·with decreasing levels of tree retention in the short tenn (<5 yr); 'abu:r.dances will.increas~ in the longer
· term. Aggregation of retenfion ·will ameliorate the effects of decreasing retention (Table I). Some vegetation layers, understory s~ in particular, are predicted to decline in the short-term after ba.-vest (Halpern et al. 19~), but will recover or be replaced by other undetstory species over lo~g periods of time (>5 yr).
Hypothesis }-Primary and secondary cavity-nesting birds will decline wirhdecreasing levels of tree retention (l]'.d by dispersing tl>.e pattern of retention (Table I). Increased levels of harvest will result in marked reductions in snags in treatment units with s;40% retention (Halpern et al . . 1999). Old decayed snags, which are valuable for ·some ,cavity-nesting birds (nortbem. flicker [ Colaptes aur~ J, red-breasted nuthatch [Sitta canadensis]), will be lost in treated areas, and creation of replacement snags from sound trees
(see Aubry et al. 1999) will have little effec~ in the short-term. Aggregation of retained trees in undisturbed refugia will likely ameliorate the effect of declining retention on cavity-nester$ that nse open canopy habitat (e.g., chesmut-backed · chickadees [Paros nifescens]).
Methods
Bird community composition, species richness, and relativ~ abundance were estimated in each stand from four point-count stations using the modified variable circular plot method described
by Reynolds et al. (1980). ·spot mapping was conducted daring point counts, following proce- -dllres b)( Ralph et al. (1993), to examine locations of territories of selected species ('Table· 2) relative to canopy and forest floor characteristics before and after harvest.
Bi.rd surveys began in late April and ended by early-·to mid-July . . Each treatment unit was "visited six times ouring the breeding seas~n. Surveys were evenly spaced throughotrt the breed-
. in,g season to capture variation in breeding phenology among bird species. Abundance was expressed as a detection rate (mean number of birds detected Within 75 m per visit) to facilitate comparison of species abundance among stmds. Count stations were~ least 160 m apart, thus 75 m was chosen: as a conservative maximum detection distance to avoid recounting indivi~ birds. We combined ·detections of hermit warblers and Tovrosend'swarblers because of extensive hybrid- ·
. ization between the two species (Rohw~andWocd 19_98). We later will as~ habitat qoality anq verify'breecli:ng stttus of all birds by using.a reproductive index (Vickery et al. 1992) to determine whether species are nesting in the study stands.
Power analyses for four common species showed adequate power in the point-count sampling design to oetect changes in detection rates o( heµnit!Townsend's warblers (0.99), goldencrownedkinglers (0.99), dl!I:k-eyedjuricos (Junco hyemalis) (Q.97), and bro~ creepex. (Certhia americana) (0.96). 'Powerwas inadequate (0.52) for the less ~ommon. and wider-ranging ·gray jay (Perisoreus ca:nadensis).
Summary of Pre-treatment Results . . Using the point-count.data, we selected those species for which some analyses might be pos-sible in one or more sites. Criteria for selection were species that were observed within 75 m of the observer and species that had 30 or more detections in a block in either of the two pre-treatment sampling sessions. A threshold of 30 detections was evident in the frequency distribution of pre-treatment detections and was thus used to separate common from 1lllCOmmon species. In Washington, 15 of 51 species detected fitthe criteria for analyses and in Oregon, 12 of70 species (Table 2). There should pe adequate pow.er to
Vertebrate' Response to Green-tree Retention 49
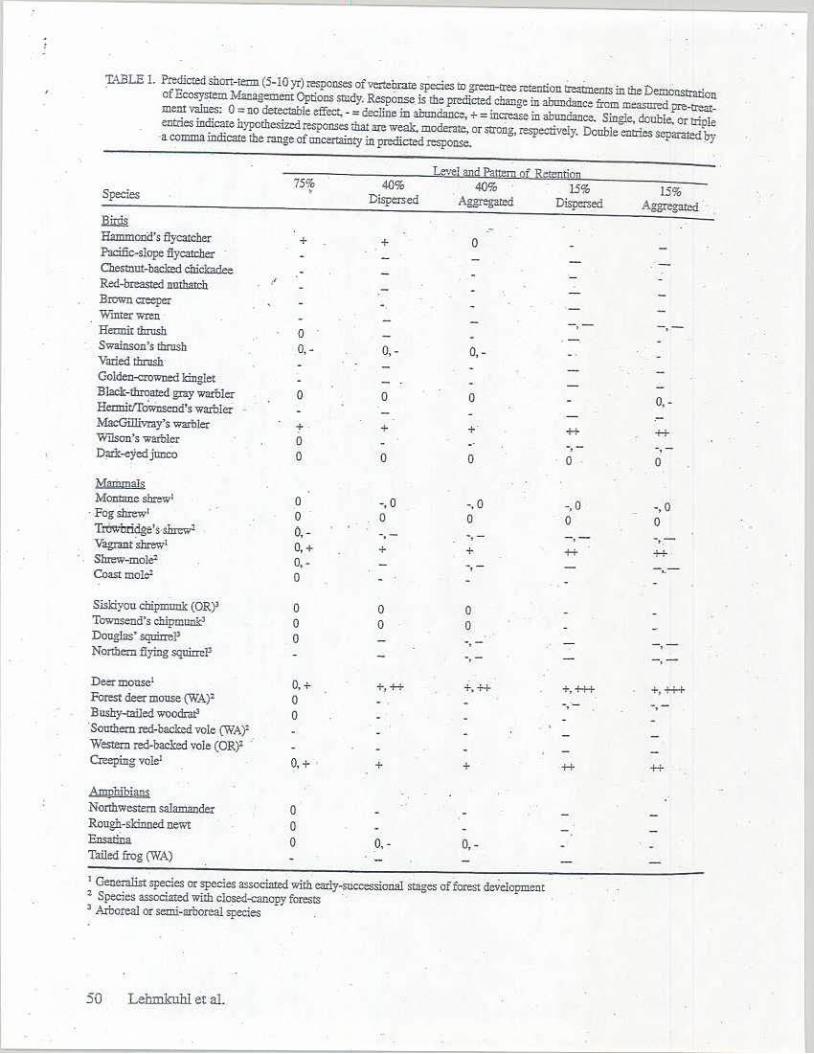
'I.'\BLE 1. Predicted shon-t= (5-IO yr) responses of Vtr<.C~ species !o green-tree =ntion ireaimerus in the Demonstration · of Ecosysrem lvfanagemen, Opti,ons smdy. Response.is tile predicted change in abtmdance from =med pre-trcar
mem valDes: 0 = no deteclable effect, - = clec)ine iD abtmdance, + = in=se in aburuiance. Single, double. or lriDle entries indie&e h)'JlCtb.es:ized respcnscs dlat ,ue weak. modera~ or strong, n,spective!y. Double entries separated )ry -a comma indicare the range of aneo...rtainty in predicted response.
J&:i,l !llld Pa~m Qf R<:tenli!l!I 75% 40% 40% 15% 1.5" , . Species • Dispened Aggregated Dispexsed Aggregated
~ --Ha:mmonl!'s flycarcher + + 0 .P-Jcific-s!ope flycatcher Chestnut-haded chickadee ·-Red-breasted nnlhaiclt l
Brown creeper Winter v,ren Hei:mit thrush 0
-.- -.-Swainson's dirosb Varied thrnsh·
0,- 0, - 0, -
C-olden-<:rowned kinglet Black-1fm:ated gray warolc:r 0 Heonit!!ownscoo's warbler
0 0 0,-.-MacGillivray's warnJer t + + ++ ++ W'ilson's warbler 0 -.- -.-.D;rlc-eyedjuoco 0 0 0 0 0
Mammals Momane sm:ew1 0 -. 0 -, 0 -,0 -,0 · Fog shrew' 0 0 0 0 0 Tom:!ridge's·shrew' 0,- -. - -. - -.- .. ,.-Vagrant shrew' O, + + + ++ ++ Shrew-mole' 0, - -. - ,. Coo,strnole' 0
Siskiyoucbipm,mk(ORY 0 0 0 Townsend's chipmunk' 0 0 0 Douglas' sgaim,l' 0 -.-Nortbem flying squi,rtl' -.- -.-O,....e:rmouse1 O, + +,++ +.+4 +,+++ 4,+i-+ Forest deer IDOllSe (WA)2 0 -. - -.-BllShy-<ailed woodrat' 0 · Sombero red-l>acked vole {W.A;)' Western red-backed vole (OR)' · Cre--ping vole' O. + + + ++ ++
AmprtJblans Northwestern salamander 0 Rough-skinned newt 0 Ensatina 0 Toiled frog (WA)
o. - 0, -
1 Generalist si:ecies er species associared with early-suc::,:ssional stages of fore.~ de;clopment • Species as.;,ciated wi1b closed~opy forests · ' .A.xboreal er semi-nrooreal species
50 Lehmkuhl et al_
f r f 6
fr ~ ' w r El. Iii v, .....
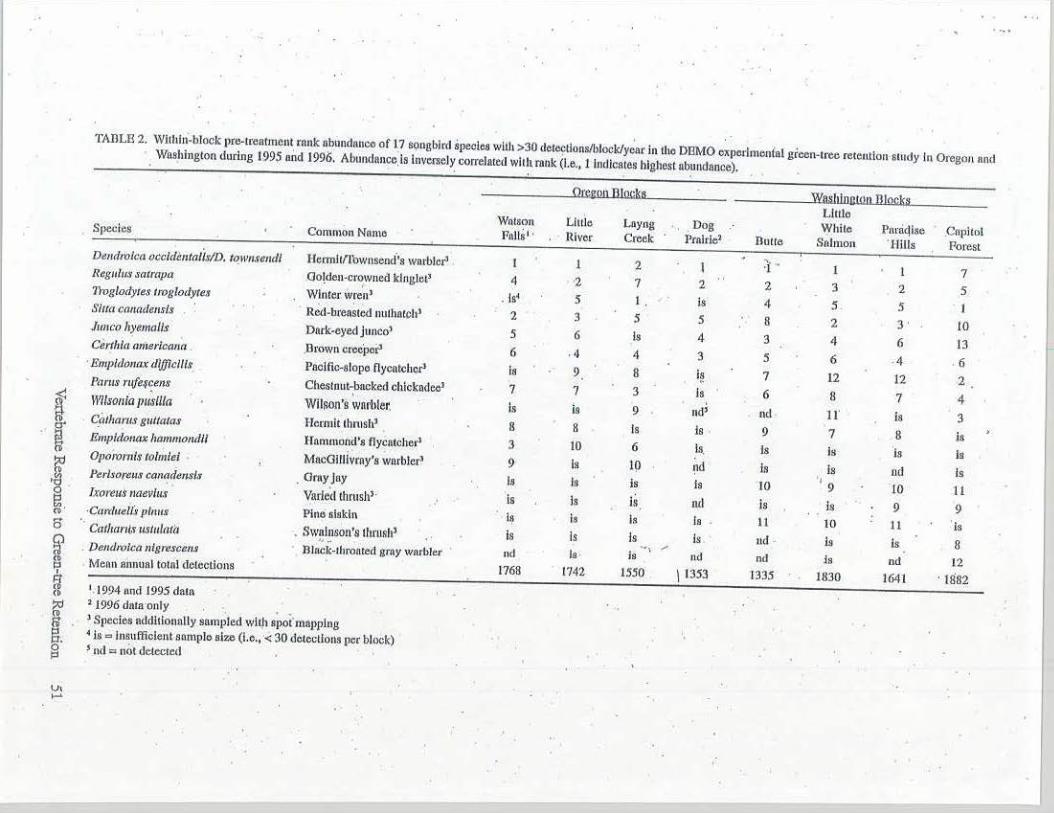
TABLE 2. Wllhhi-block pre-lreaimcnt rank nbundanco of 17 songbird species with >30 del~llons/block/y.;., ln lhc DBMO experlmonlat gioen-1ree rereutlon srudy In OregQn nnd Washinglon during t.995 und 19%. Abund1111ce_ls lnvecsel~ correlaced whit mnk (I.e., 1 lndicaces highest nb~udance).
·.aawu Illa~ · YiJWtlnnton mock• Lllllo Wntson Little Layng Dog While Parriqlse ,CF1phol Species Common Name Pulls I . River Creek Prnlrle' Dulle Salmon ' IHlls Porest
Dem/mica occldimtnlistD. to._wu.re11d/ Hermll/fownsond's warbler' . 1· ., l 1 2 l I I 7 Regulus satrapa .Oo{den-c-r~wned kinglet' 4 z 7 2 2 3 2 5 1lvg/otlyres troglotlytes Winter. wrenl . Is' 5 l . ls 4 5 5 I Siua ca,i(u/e11sls . : Red-brea;ted nulhntch' 2 3 5 5 B 2 3 ' 10 J1111co ltyema//s Dark-eyed Junco' 5 6 Is 4 3 4 6 13 C~rthltt ,miericafitJ .Drown crccpor' 6 , 4 4 3 5 6 .4 , 6 · Emp/tlolllu: tllj]lc/11,t Paclfic-slopo flycatc~cr' is 9 8 i§ 7 12 12 2 P11r11s riifer~ens Chestnut-backed chickadee' 7 7 3 ls 6 8 7 4 11'/lso,iia JIUSll/a Wilson's wart1ler. i-s is 9 nd> nd 11' is 3 C.f, thnrus gultatas Hermit thnish' 8 8 Is Is . 9 7 8 is Emp/douax luunmo,1dJ/ Hnmmon?'• flycntcher' '.l to 6 Is. Is Is is ls Oporom/s tolmlei MacGilllvrny's warbler' 9 Is 19 nd Is ls nd Is Is is ' 1
·10 PerU·oreHs can'!den#r . Omy Jay I• Is IO 9 11 b:oreu.t naevills Varied lhrush'· Is ls is nd is Is 9 9 ·C4rrluell:S pllw.r PincS siskin 1, i• ls is • II IO 11 ' · IS · Cathnnts ustulati'I . Swalnson's thrush' ls Is ls Is 11d is ts 8 ,, - ~, , . Dendrcl,,;a nlgrescc11s lllack-1hronred gmy warbler nd Is · Is ud nd Js nd 12 Mean aimual total detections 1768 1742 1550 \ 1353 1335 1830 !Ml · 1882 1• I 994 ond I 995 data ' . '1996 dnra·only . . . 'S[lecies nddltlooolly sampled with spoi mapping • l• = insufficient somplo size (l.o., < 30 deteclious per block) s nd ::: n01 detected

show treatment effects for at least the fou:i: species found in significant numl:iers in all blocks. '.
. . . (Forsman et al.1984). Some arlJoreaJ and terres
. trial rodents consume· the sporocaxps of ectomycorrhizal fun~ and disperse fungal spores· and nili?gen-fi.'ting bacteria (Fogel and Trappe 1978, Li et al.. 198G)_ and ihus represent an important link i:n the nutrient cycling 2nd produc-
I:te:rorit/I'owns~'s warblers, golden-crowned
ti'(ity of foresrs. · .
· ki.ngle1S, red-breasted nuthatches, and brown creepers were fuir/.Y abundant on all blocks in both stares (Table 2). As ei;:pe..."ted, species that occupy large tecitories(e.g.,groose, woodpeck::rs;andcoivids) . ,. generally were detected in small nllI!lbers andare . not included in the lL<t. Of the 15 species in Washington bloc.ks; four were either not detected or detected only in small nmnbers on some blocks. · This was true for seven species in 'Oregon. .
¥orea!~semi-arborealspeciesinihestudy areas are ~y found in closed<anopyforest
. . that has developed beyond the stem-exclUSion · stage: This pattem is generally attnontable to the
• • • J •
Some elevatioiial. trends in species abundance also were evident. For example, winter wrens (Troglodytes troglodytes), Wtlson's warblers
.. (Wzlsoma pusiila), and Pacific-slope flycatchers (Empif;ionax difficilis) generally were most abundantin lov;:;-el~on blocks (Capitol Forest,. Layng Creek). ·Resulting regional and elevationai. dif.
· . · ferences in bird species cpmposition and abun. dail.ce between blocks in Oregon and Washington will make fumre analysis over the total array of treatments potentially difficult for some species.
Arboreal and Terrestrial Small Mammals
Review ·
Th~ small mammal comronnity of the Pacific · Northwest is one of the richest in North • .<\merica (Hams 1984, Raphael 1990). Several species,: such as die Trow bridge's shrew (Sorextrowbridgtz), shrew-mole (Neurotrichus gibbsit), forest deer mouse (Peromyscus keem),'westem red-backed. vole (Clethriorwm:ys califomicus), and red tree vole ·(Phe=orrrys longfcaudui), occur .only in western British _Colnlnbia; Washiilgton, Oregon, or northern CaJifOIIlia (P..a.gmeier and S~ 1964, Si:mpsQJl 1964). These. species are well adapted to older forests (Harris 1984, Aubry et aL 1991) and typically cccur in greatest abnndanca in natnrally regen.exated mature forests (Carey and Johnson 1995). : .
·Arboreal and tetrestrial small mammals play important ecological roles in forest ecosy5\CIDS. They form key links in many food webs. as 'consumers and prey, and are important ~ersers of plant propagules (Hanis 1984). For example, northern flying sqnmel.s,- red tree voles, and wooo.-ars (Neotoma spp.) are primary prey for the northern spotted owl and . other fo;est owls
. .
52 Lebmknbl et al
. greater compositional and StructJJral dwamyfonud • ino1dfores:tageclasses(Carey 1991, 1995). These S!)ecies can, however, be relatively abundant in. younger stands with div~e understory composition and woody _debris legacies (D?yle 1990, ~osenberg and Anthony 1993, Carey 1995). Howev:er, tbey·are heavily inflnen~ by traditional forest management practices that 'remove part or all oftbeforestcanopy(Carey 1991, 1995). . . The diversity of teirestrial small mammals in the study~ precludes a s.imple generalization aboutabnndancein.relationtoforestageandhabita!
. featureS. ·Many investigators have studied small :tnammals in old-growth forests, in naturally re"
.. generated younge:. stands; and in ~ntclearcnts (<20 yr old) (Hooven and Black 1976, Ramirez
. and Homccker 1981, Martell 1983, Van Home 1983,R.apb3e.l 1988, Moirison andAntbony -1989, Ruggiero et al. 1991, Carey 1995, ~y and 'Johnson 1995). Several species (red-backed voles, Trowbridge's shrew, shrew-moles) are most almn·dantin closed.:Canopy fore,sts tbatbave developed beyond the stem-exclusion stage, when;, sparse herb a:nd shrub'Jayers provide limited reso~ for these species. Several other species are habitat generalis~ (deer mouse [Peromyscl!S malliada.tus],mcintanesbrew [Sorexmo11iicolusD, or are most abundaiitfu qense, ground-level vegetation.characteristic of early-successional communities and,streamside areas (creeping vole
. [Microtus o~gom],Pacificjumpingm.01.1..se [2Apus_ ' trinptatus D- Clearcut logging produces anfu<;<?r
!lble habi:tat for c1osed-canopy species until.canopy . closare. Long-term adverse llll!:)acts on persis
tence (over several rotations) may occur if harvest techniques e!iroinat.o inputs of coarse woody
· debris and other resources. Conversely, high-intens:iry. di.stllrba:nces will lead to an increase of early-successiorutl small roamm•i species. Generalist species· also may increase in abUBdance
. with distorlJance as a direct effect of habitat changes

and mcfuectly if the disturbance adversely affects . collll'etitors. ·
Although the general changes in the composition.of sma.11 mammal <'Ommlllli.ties can be predicted following clearcutting, (Rooven and Black 1976, Moaison and Anthony 1989) responses to Iess·mtensive harvest are not clear; Recent studies have examined small mammal response to green-tree retention harvests (Warers and Zabel 1995,·Chmnbers 1996, Von Treba et al. 1998), but !hese stuclies were not designed to provide broad inference for Pacifi.cNorthwest species. In particular, little information is ava:ilable to compare effects of dispersed vs. aggregated retention, or t(j predict patterns of response across a gradi.c ent of increasing disturbance intensity. Similarly,
. the ability. of retention patches to serve as refugia for closed-canopy species is tlllknowa,
Hypotheses
HYpOrlJesis 1-Abwui.ance of arboreal rodents will decline with decreasing levels of retenlion CJ/ green trees, although aggregation of retention will redµce rhis effect (Tal:ile I). Decreasing retel).tion. will result in macked reduction in arboreal canopy habitat and den snags in !Ieat!nentunits wiih <40% retention (Halpem et al. 1999). Old decayed snags, which. are most valuable fqr denning sites, will be lost in treated areas, and creation of replacement snags fi;om sonnd trees will have little benefit m the short-renn. Similar declining trends in the number of vegetation layers and the cover of the dominant understory shrubs (e.g'.; Acer.· circinatum, Berberis nervosa, Gaultheria shallon and Vaccinium spp.)-especially ericaceous shrubs-will conttibute to the hypothesized re-spoose (Carey 199 5). . ·
Northemfiying squirrels likely wilt-show g=ter responses tQ treatments than chipmunks (Tamias spp.), whi~h ~ semi-arboreal (Carey 1991): However; aggregation of.retained 1rees .in llll~
turbed refugia likely will ameliorate these effects to some extent. Declining abundances· of arbo-
. real rodents also will be associated with declines in the diversity and abundance of ectomyc0n:hiza1
· fungi as a consequence of canopyremoval (Cazares ·et al.1.999). Northern flying squi,.-i:els, which are obligate fungivores, are ex~d to experience greater treannent effects than chipmunks, wbicli have more diverse diets.
Hypothesis 2-Terrestrial small mammals asso~ed with closed canopy (post stem-exclusion) foresrs will decline with increasing levels of disturbance ·(Table 1); tr~atment-level abundances will notdiffer between dispersed and aggregated harvests of the same intensity. ·we expect clumped populations of these species will be found in retention patches of aggregate treatment units, with few individuals occupying the harvested matrix.
. In clispersedretention patches, these species will be found in low abundance throughout the unit.
· Hvpothesis · 3-Small mammdl species asso-. dated.with eariy-successional. habitats and habitat
generalists will increase or lza;,e no. change in abundance with dis.fl:lrbance /nrensity .:. The relative abundance of these species will increase with the proportion of basal area removed. Patt.."IDS of clispersion for these species will be the ilrverse of the patterns· hypothesized for closed-canopy .species. ·
Methods ··
Methods for live-trapping arboreal rodents were modmed from. Carey et al (1991a). Tomahawk 2011 traps (Tomahawk Live Trap Company, Toma-· hawk, W1$consin) were used to sample ·an 8 x 8 or 7 X 9 sampling'grid within <each treatment unit .Sampling occmred in the fall to estimate reproductive status, relative abundances, and consumption of hypogeous fungi by flying squirrels and chipmunks. Captures were reported as individu-· als captured per 100 trap nights, adjusted· by 0.5
. trap nights. for sprung traps; however, maik-recap.ture poprilation estimates will be .maile if fur- · cher arialysis shows that capture data can meet the proper ;issumptions.
Sampling for 2 wk on all !lI1its during '1994 and 1995 indicated that flying squirrel ablllJdances were too low (averaging 0-1 squirrel per grid) in three of eight blocks for .future analysis of treatment effects, and that an additional 2 wk of sam-· . pling resulted m. about 25% more captnres of inclividual squirrels. J,....s a result, we disoonti:nued sampling arboreal rodents in the £oar blocks with low to marginal abundance, but douoled the s~pling period to 4 wk at the remaining four blocks. Power analysis showed that doubling the trapping sessions to 4 wk on four blocks would result in power ~ .90 for cletecting a 50% reduction in abundance.
Vertebrate Response to Green-tree Retention 53·

To estimate the species and quantities of fungi . consumed, fecal pellets were collected from fl.y- · ·mg squirrels, chipmunks; and red-backed v6les on the WaiSonFalls bloc.kin Oregon and.the Butte block m Washington. Frequency of occurrence and relative abundance of fungal spores hi feces will be compared to the conCUI:rentfreqriency and abimdance offuzigisampledas partoftheDElvIO mycological studies (Cazares et al. 1999).. · . Pitihll traps were used to sample tern:strialsmaTI
mammals at ea,ch point in the grid. . Only-pitfall traps were used because 'they sample those species whose habitat associations are least 1llld&stood, ie., the insectivores. ·Most other small ·rnaroroal species, although less efficiently captured by pitfall traps, are ge~y canghtin sufficient numbei:s for statistical analyses (Ai;bry et al. 1991); power analysis of preii:m.i:qai-y pitf.ll.l captures for deer-mice (Peromyscus spp.) showed power to be ~.99 for detecting 50% reductions in abtilldai:lce. Power to .estimate effects. 011 Trowbridge's shrew and vagrant shrew (Sorex vagrar..s), which were more abundant than deer mice, is expected to be similar ·
: Pitfall traps were operated as removal traps to provide data comparable to previous studies in
. unmanaged forests (Ruggiei:o et al. 1'991)' and to ongoµig studies in managed forests in the Pacific
Northwest (K. B .. Aubry, pei:s. comm.). Traps were partially filled with waier as recommended by the. .A..merica:o Society of Mammalogists (1987), opened far 28 days in each trean:nent unit, -and· checked once a wee.k: between September and November. Numbers canght were reported as an tlI!correC".ed catch per llllit effort index ( capturcS/ 10.0 trap nights).
' Sut:nmaif of Pre-treatment Results
Ten rodent species were captured using arbqreal ·rodent.trapping methods, during 1995-96 (Table 3). Clripmunks were the most :freqoentlf captured group. ·TOW!)Send's chipmonk. (Tamias
. · townsendi!)wasabmidaruonallWashingtonblocks · · and on the Layng Creek bloc.k: in Oregon; The . Siskiyou. chipmt!Dk (! siskiyou) was th<; most
, abundant. species on the Watson ralls b~ fu - Oregon. Altogether,. 568 northem flying squir
rels were captnred. Small numbers ofbushy-tailed wooch-ats (Nedtoma cinerea) were captured on several blocks. Douglas' squirrels (Tainia.sciur.is doug!asii) werecaptmed:in:freqnently en all ~locks, but. this species is probably nnde~nted .in the data!itJe to trap avpidance (Carey ~al.199la).
Captnres of teII°--strial small mammals were similar in bregon and Washington, with shrews (Sora spp.), deer mice (Peromyscus spp.), and
1:~LE 3. Wrthin-blcck pre-~nr rank abandance of rodeo! species. capmred in TOlll3hawldive traps in the DEMO e:xperi.memal gr=-;ree re!l?lllicn stndy in Oregcu and Washington daring 1995 and 1996, C:!p!!ln: ranks (l = lrigl,cst abundance) were de,""1llin-,.d from the lOt3l mnnber of individnals c,xptnred for "'1cb. s;,cci,:s wl!liin a block. . .
Tamias to·.vnsmdii
Tamias siskiyou
. G',;,,,;.,,rrys sabrinus Tamiasci,u,.ts daugiasii
SpermofhilUS_imerolis .Spemu;philus beecheyi
}feotoma cfrk."t'ea
Spermcpiri!us satw'atuJ· Tam/as torJJenus •
Sciurus can,lill.ensis Mean annna1 rota! individnals
Coimnon Name
Townsend's cbipm,ink' Siskiyou c:hipmnm:1 _
Northern B:img ~ Douglas' sq1liirel .
C-olden-mantled groll!td squincl
California groll!ld sqomel
Bushy-l:>i!ed woodtar .·
· C=rle golden-mantled ground sqail:rel Yellow-pine cbipIDllllk
Eastern zray sqnim,J
Watson Leyl\g-Folls Creek .
na' I
' I l!a,,
2 2
3 3
4 ..)
s ~
6 4
na 1lll
251 286
Wasnin$Jm 'Blocks . Capitol
Bmte Forest
I .
oa 2 5
3
4
6
321
oa. ·2 :
3
na
4.
153 1 Distributions ofTO\'YuSend's and Siskiyou chiplllllllks in ~on ae,cording to Vcrts and Cttrawey (1998).
1 na ~ species• r.mge outside of sample blocks 3
- = not captnrn!
54 J P.hm]mhJ et al.
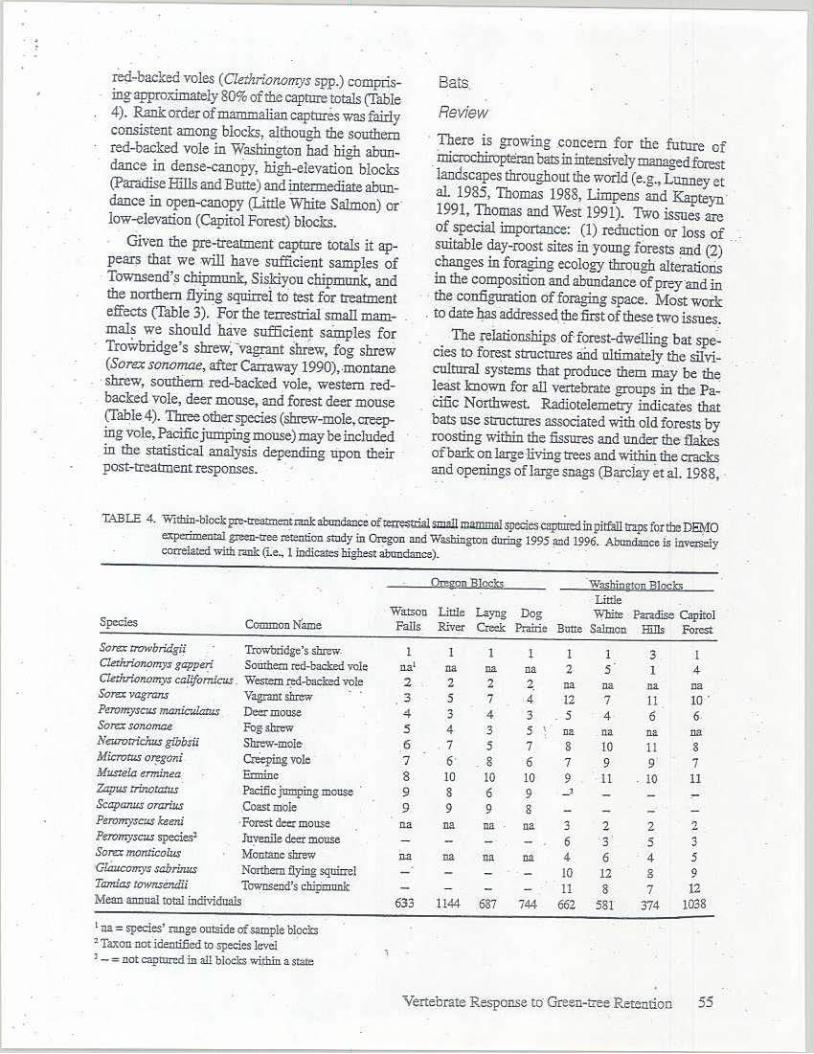
red-backed voles ( C/ethrionmeys spp.) comprising approximately 80% of the capture t.otius (Table 4). Rank order of roamroa)fan capta:res was fairly consistent among blocks, although the southern red-backed vole in Washington had high abundance in dense-canopy, high-elevation blocks (Paradise Hills and Butte) and jntermediate abundance in open-canopy (Little Wlri.te Salmon) or· low-elevation (Capitol Forest) blocks.
Sais,
Review
There is growing .concern for the future of . inicrochiropteran ba!sin intensiveiy managed forest landscapes tbroughoru the world ( e.g., Lunney et al. 1985, Thomas 1988, Llmpens and Kapteyn: 1991, Tnomas and West 1991). Two issues are of special importance: (1) reduction or loss of . suitable day-roost sites in young forests _and (2) · · changes in foraging ecology through alterations
Given the pre-treatment capture totius it appear~ that we will have sufficient samples of Townsend's chipmun1;:, Siskiyou chiDIIlunk, and the northern flying sqnmei to test f;r treatment effects (Table 3). · For the terrestrial small roam- . mals we should have sufficie~ samples for Trowbridge's shrew, -vagrant shrew, fog shrew (Sora sonomae, after Carraway 1990),-montane
in the composition and abundance of prey--3JJ.d :in the configuration of foraging space. Most work
. to date has ·addressed the first oftbesetwo issues.
The ~lationships ~f-forest-dwelling bat spe~ cies to forest Structures and ultimately the silvicultura! systems that produce them may be the least kno,wn for all vertebrate groups :in the Pacific Northwest Radiotelemetry indicare.s that bats 11Se structures associated with old forests_ by roosting within the fissures and under the flakes ofbatk on lame living trees and within the cracks and open:ings~oflargesnags (Barciay et al. 1988,
· shrew, soutliem red-backed vole, western redbacked vole, deer mouse, and forest deer mouse (Table 4). Three other species (shrew-mole, creeJr ing vole, Pacific junipi.ng mouse) may be included in the statistical analysis depending upon their post-treatment responses.
1:<\.BLE 4. Wilhin-blockpre-treannemr.mk abundance o(te=llialsmzll lllammal species captmed in pitfiill II:ips for the D~O experimental g,.=-ttee rei.ation Stlldy in On:gon and Washington du.ring 1995 !llld 1996. Aband.1nce is invezseiy COl'.l'elall:d wnh r.ink (i.e, l iDdicaies b(.smest aband3nce). ·
~onB]gcJI. Wa.~hinli!2JJ :SJ~~ Little
Wmoo Liule Layng Dog White Paradise Capitol Species CommonN'ame Falls River Creek Ptairic Butte Salmon Hills Forest Sorex rrowbridgii Trowbridge's shrew. I I 1 l I 3 l Cktr.r'.o~mys gappei;i Somhem n:d-backed vole n.a' na rut na 2 5 1 4 Ckthrionomy, caUfonuc-.s . Western rtd-backed vole 2 2 2 2. •• na _na na Sora vagrans ' V~ shrew · · 3 5 7 -4 12 7 11 10 . P"101n"JSCu.s mamc-.datus Deer mouse .. 3 4 3 . 5 4 6 6-~
Sore:;; sonorr.a.e. Fogs.hi= 5 4 3 5 na na n• lJ3 NeurorricJw.s gibbsu Shiew-mo!e 6 7 s 7 8 10 JI 8 Microw.s Ol'f'gOlli Creeping vole 7 6· . 8 6 7 9 9· 7 Musutla enmnt!a Eanine 8 10 JO JO 9 11 IO 11 Zap,4$ rrinotatus Paci.fie jumpiDg mouse 9 8 6 9 -' Scapa,u,s oninus Coast mole 9 9 9 8 Pm,myscus keeni · Forest de:r mouse na llll na oa 3 2 2 2 Peromyscu.s species' Juvenile deer mouse 6 3 5 3 Sore:r mar.lice/us Monmne shrew n:a na na Oll 4 6 4 5 C-llntcomys sabr'.nus Nartbern flying squirrel lO 12 8 9 Tamiizs Ulwnsendli T~d's clripmunk ll 8 7 12 Mean annual total individuals 633 1144 687 744 662 581 374 1038
1 na = species' :range outside of =pie blocks 'Taxon not identified to species level > - = not cap1llnd in all bl~ within a st:!le
Vertebrate Re~-ponse to Green-tree Retention 55

I I ! I I I I I I
Christy 1993, Frazier 1997). When smdied in forests with continuous canopy r:lllging from 55 to 700 yr~f-age, echolccaiion calls were 2.7 to 5.7 times more frequent in older (>210 yr) than inyo1m3"rforest(ThomasandWestl991). When smdied over the much smaller range of stand ages found on privare, in1ensi\:cly ~aed fu.rest lands (from c!ei)I'Cllt ~ to about 65 yr-of-a,,,<>e ), echolocation calls were about cwo times mere frequent over clearcut areas than in yollilg forest (Erickson and West 1996).
Concomitant with greater echolocation call frequency over clearcut areas is a shift in composition of calls to a higher proportion of larger bodied; npn-Myotis species, a pattern also found by Hayes andAcfam (1996). While there is little doubt that intensive forest management C3Il lead to loss of forest sttnctmes that are important to blllS and couid result in bat population decline,. the effects pf partial harvests and green-tree reteuti.on on bat use of forested areas are not clear (Fenton et al. 1998). Given the provision of suit
. able day roosts, bat populations lilllY do quire "."ell. Critical work on the cha:racteristics of roosts is very recent for this region (Campbell 1993, Frickson l993, Ormsbee 1996,Frazier 1997). No smdies have addressed bat habitat use on a largescale experimental basis.
Hypotheses
Hypothesis l 'Use of green-tree mention 111".its · will in.crease withharvesi wensity fornon-Myotis . species. As flightspacefor.larger-bodiedandfuster
flying bats is. created, echolocation calls of silver-Mired (Lasidrrycteris Mctivagaiis), hoary (Lasiurus cinereus), andTownseixi's big~ bals (Corynorhinus rownsDlllll") will be detected at higher mxmbeis. '!bf' mnni:.,-nf echolocalicn calls from Myotis species will no!: change. Thus, the number of echolocaiion calls SlllllID,ed across bat .. species will increase with harvest intensity.
flxpothesis 2 Feeding rateswill inCTf!llSe with hmyest intensity. Cl.earcttt areas function as commuting space and as .marginal areas for feeding. Closed-canopy s1l!Ilds fanction primarily for roosting. As harvest intensity increases, the'frequency of"feerling buzzes" (a chaiacteri.stic.cail indicat-ing prey~) should increase. ·
Methods
Indices of bat acµvity were estimated wi1h automated di'Vide-by-N detectors (Anabatlldetectors
56 I ~broknbl et al
and delay switches, Titley Electtonics, Ballina, N.S.W.,Anstralia). Tne~'IVeresettorecord at dusk and shut down 8-9 hr later. Several detectors were operated simnltaneou.sly at blocks
. within eacb state. Each bl eek was snrveyed for a 2--day period each mouc!L Sampling for echolocati,on cails began in late June or early July depending upon the timing of wann weather. We fOtIDd tilJ\t .a millimum of 6 aighrs_ of sampling per site (excladi.ng windy or rainy nig!rts) was necessary to accollllt for the mgb variation in re-. corde;d calls among aightS. Although it is genei-ailynotpossihle to distinguish repeated calls made by 3Il ind..~dnal batfromindividnal cails of many bats, ih,e technique offers a very good method for cvtermining pnesen...,,,, relati.Ye use, and behavior (feeding ·vs .. passover) of bats in an area. . ·
. To proyide infmmation on age, sex ratios, re- · productive condition: and species identification of Myotis species, we captnred bats near habitual
· fl yw:iys, streams, and ponds with mist nets and collapSiole harp traps (Kunz and Knrta 1988). Upon release, echolocation calls of caplll!ed bats
. were reconled tc augment our hor.ay of refer. ence calls. '
Summajy of Pre-treatment Results . .
Atpresentweca:nideatifytllefuilowingjivespecies or species groups (Erickson and West 1996): (1) big brown bat, (2) lloa:ry bat, (3) silver-haired bat, (4) Townsend's big~ bat, and (5)Myotisspecies (M. evotis, M. volans, M.. keenii, llL ciliolabnon, M califomicus, a,,.d M. lucifagus ).
· A tctal of3,924 calls wexe detected during the pre-treatment -=ipling. In Washington, tb.e.Myo~ . group accounted for 9.8% of the 2,102 rea:iroed detections (Table 5). Other species were,l"Ul'ely detected. Detections at tile Lltrle White Sahn~ block accounted for-over half ((>2%) ofall detec·
. ti.ens in Washington and 33% of calls for both states. .In Oregon, the Myotis species group acco~ for 97% of the 1,822 recorded calls.
Niiht!Y activity was bi~h~ OD ~Iittle White Salmon block, which has relatively open-canopy stands, with an a..erage of ~17 calls per night. This r&n!re of activity is as high as that found in old-growth (Thomas and West 1991). The Little River block was the second most active sire with just over eight cails pet aight. The least active blocks were the relatively closed-canopy Butte and Par-..dise Hills blocb with <3 calls pet night.
- - -

TABLE. 5. Pre-~ni mean c!e~--ticn ~ \ecbo!ccatlon ~~t), and &:mdaro = of ~ for all t,8!5, all Myotis
· species, and all non-Myou.r specie:; m D~O e.penmen!ai gre..-n-t= ~OD blocks .in Oregon (1995 and 1996) andWasJiin~.OD (1995). .
T013i. Bleck x(se)
~ Wa!Son Falls 7.89 (11.22) .Llttle River 8.21 (10.39),
. Layng Creek 4.92 (Q.:13) Dog Prairie 4.'06 (9.97)
· Wa$bingmn Btttte /' 2.53 (11.()4) Linle White Salmon· 17.~l (29.58) Paradise IDl1s · 1.56_(2.86)
. Capitol Forest 6.90(14.87)
As expected, feeding activey was low within an · blocks. ·of the 3,924 detections,. only 35 were identified as feeding activity. Lltt1e White Salmon bad the highest number of feeding buzzes, ac- · · COllllting for 66% of the feeding activir<J, ·
Arnphibians
Review
In regions west of the Cascade crest, amplnoi.ans dommatethePacificNorthwestherpeto.fanoa. Not only are th= more species of amphi'bians than reptiles,' bi:tt they are more distinctive in their endem.ism (Nnssb~um et al. 1983). In headwater s~. and~ l!l'eas of the Pacific Northwest, amphibian adul~ and.larvae are the top predators, easily exceajing the nru:nbers and biomass. of other foDDS (Bury 1988). Densities of ariipln'bians can be very high, up to 12 per in2 in moist areas {Leonani°et al _1993).·
Sevfil<!l amplnoi.an taxa are bejng revised and a number of cryptic species have been described (Good 1989, Good and Wake 1992, Green et al. 1997). Recent identification of new,~~es underscores tbe disjunct, nature of amphibian dis-1noutions and the potential for extinction. Al-
. though there have been studies 'of habitat relationships_ :in unmanaged fores1;5 for the fysteJlllltically stable species of the region (Raphael 1988,Aubry and;Hall 1991, ComandBmy 1991, GilbertandAilwine 1991b), weknowlittleabout habit.at associations for many of the newly iden-· tined species in unn:ianagP.d forests. Information ,in managed forests is particul.arly sparse' forinost
. MJ,-olis Non-MJotis. · x(se) . x(se)
7.71 (11.20) 0.11 (0.72) 8.14(1032} 0.00 (0.30) 4.43-(5.66) 0.45 (2.55) 4.06(9.97} 0
2.45 (lo.98) 0.06 (0.23) 17.26 (29.16) 0.51 (2.99) 1 . .5$ {2.96) 0.04 (0.19) 6.90 (14.87) 0
species. Well-replicated studies of amphibian occm:rence in managed forest are needed to investigate the ~.s of several commODly employed silvicu)~ practices, including shon-rotation harvest, thinning, ap.d green-cree retention. Sev-eral SP.eci,es have special stams m Oregon and Washington: Cas~frog (Rana =cadae), tailed
.. frog (Ascaplws truez), spottedfrog(Ranaprw,sa), clouded salaroawier (Aneides ferreus), Oregon·
. slender salamander (Barrachoseps wrig7!tz), tor-. . rent saJamamlers (Rhyacotrtton. cascadae and R. variegatus), . Larcli Mountain saJamantler (Plethodanlaiseik),Dwm's.salamaader(P.. dunm.), ~d Yan Dyke's salamander (.R l/anayke?). Infor-
. mation from: our stuaies will be import:ru:tt for · conservatiOI! of these sensitive species.
Hypotheses
Hypothesjs' I-'.Abundance of ami,hibians will · decline wiih harvest~- 'we -expect a general decline for all s,Pecies with ~g har
·vest intensity, ;.itb, the exception of.the westem red-backed saJail)ander (Pletlwdon vehiculum) m Waslrington wli.ere the species has shown an ability to persist in clearcut aro--as. We expect a sharp decline in species richness, but. this may <iccar over a longer time frame~ the initial 2-yrpost-
. harvest sampling period. Several ai:npbi'bian spe-
. ci~ are long lived and are capable of withstanding adverse conditions far prolon,,_oed periods. Thus, we may· see a lag response for this pattern, an-. der:scoring the need toresample these sites in the future(pemaps at5 and lOyraftertbe i:nitiarpost- . harvest sampling).
Vertebr..te Response to' Green~treeRetention 57

, Hypothe.5is2--A7rrphfbi=willpernrtinforest
· patches in aggregated retention W?irs; but· mdsr $pecies will decline in harvested areas wtthi:n aggregated retauion units 1111d in dispersed retention units. We expect that the retaineq forest patches may be sufficient to pei;mit sa:rvival of a portion. of the original amplnoian community, perliaps until the forest canopy closes ~oain. We expect the numbers of amphibians persisting" in the forest patches to be positively co.crelat.ed with the total area remaining in patches."
HY1?othesis }--:-Individuals CarJtw'ed in. the han:estedporf.onsofthe'aggregatedretentionuntts andin.thel5%dispersedret.entionlO!itswillsJww poorer b_ody condition compared to individuals found withi7! tr.e fo~ patches o'r in the 40% di.spened -retention unfr.s. Adverse-conditions in harvest areas should lead to poor body condition. for individuals residing there. We would expect reduced reproduction· and lower than expected values for. bi,'dy-condition indices (sqch as a re-
. gression of weight vs. snout-vent length). ·
Methods·
Amplubians were samp]edconcoo:entlywith tei:restrial -small mammals during fall usi:ng pitfall traps. 'Iraps were opened after the onset of fall Iains wh~ amplu'bian si:n:face activity increases
·. fromtherelativei:nact±v:ity-ofsmmner.·.Afreridentifying species and recording info.rmation on body
· dimensions, mass, sex and reproductive condition. amphi'bians were toe-dipped and released or held under cool, moist conc!itions for.the remainder of the sampling periods theJ! returned to their point of capture. As with small mammals, nlllJlbe:is caught were expressed l!S an uncorrected · catch of individnals per 1lllit effort index _( captures/100 !rap nights).
Pitfall trapping is a good techniqne for captaring surface7active amph!.oian.s, bm is not very effective at sampling species thatlimittheirmovements to sm:face cover or the interior of.large woody debris. Fortunately, only two species limit their surface activity, the clouded salamander-and ihe Oregon slender salamander, both of which occur only in 'Oregon. TllI!e-constrained searnhes and cover-board sampling techniques to targ~t these species initially were planned, In.it were not conduct.,<>d becanse of budget constraints and the localized diitnoutions of the species.
58 Lehmkuhl et al.
Summary of Pre-treatment Results
TheiaI!k ordering of amphibian captures sh~ed considerable variation among blocks and between states (Table 6). Am:pJ:iibian abundance and diversity w_ere about two times higher in Washington than m Oregon. Five species were common to all blocks-in Was!µngton, but only two species were present on all blocks in Oregon. In Washingto_n; species composition varied considerahlv among blocks (Table 6). This high variability will make statistical detectiou of post-treatment effects difficult. However, the ensatiiia (E11Satma eschscholtz:ii);-:;vhich comprised 54% of all am.-
. phibian capnne~ across the ~ight blocks should be a suitable sp~es for detecting treatment. effects across both ~res. In Washiµgton, the ~orthwes_tera saiarnander(Ambjstomagracile) and the taile(! frog were also cornm_on enough to sugg~ that treatment effects Will be detectable.
Limitations and Challenges
In an undertaking of this kind and .magnitude one expectS challenges. We hav~ experienced several difficulties thus fui; many were expected and others we.re not. For example, we anticipated tbat ,tlJe pa..'chiness of species' distributions wooldrnake o:¢:form replicates impossible. There are species in each vertebrate group that are found only in suosets of the study blocks.· Tnis will result in lower power .fur treatment comparisons. In addition, the scale of silvicultural op,,"'.rations does not always mesh well with ~e scales·of species' biology. Given the small size ?fll+trvest uni.~ .elatiye to the homer,mges of larger-bowed species, . some 5P.ecies will not respond e:!:Clusiveiy to th~ effecrs of the experimental treatments. They may show partial responses to the treatmenis, but may beinfluencedby conditions adjace~tto the treatment aro--as. Our cboices. of species' for the stuay
· were made partly with this in mmd. We chose those spetjes for which we had the best chance of measuring a response annlrutab!e to the treatments, ie~ thpse species with limited movement er those tied to small areas by seasonal territoriality. Th.is aimroach cannot be entirely successful, and whlle'the sca!i:ng will be acceptable for smlill roammak amphibians and several avian ' ' . . species, it may be ansatisf-aetozy for others socb as the larger cavity-nesting birds.
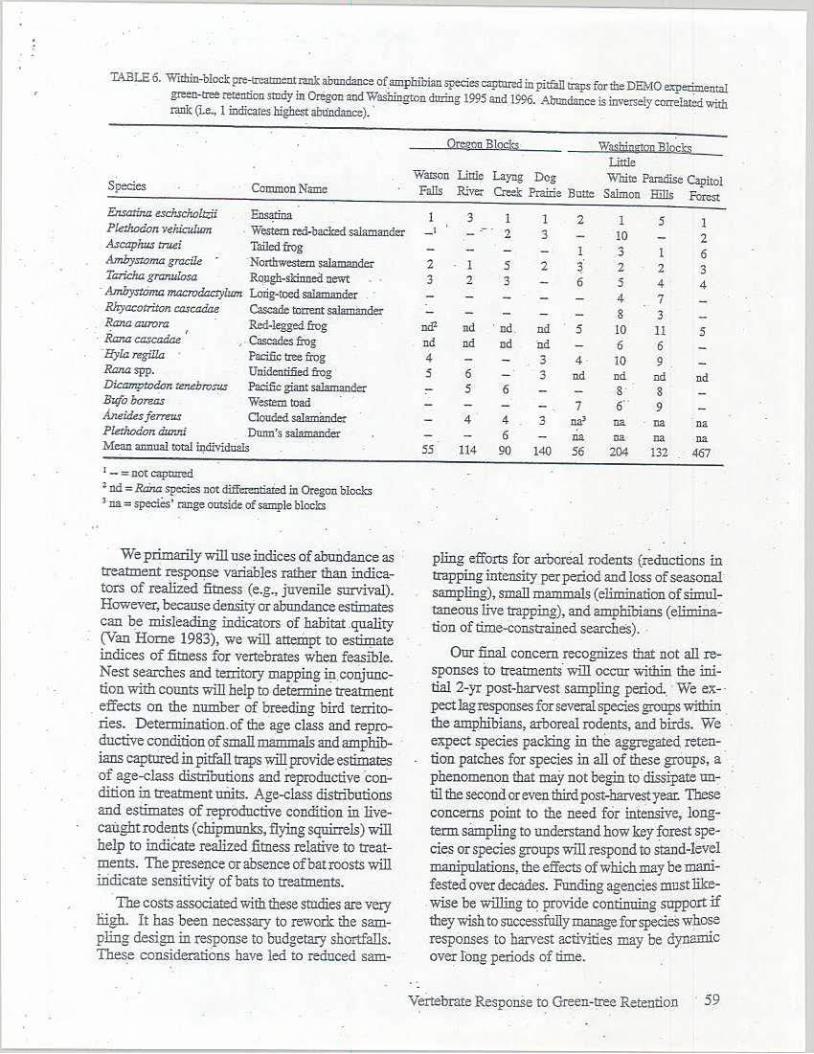
TABLE 6. Within-blocl: pre-lr,'3!mentrank ab1llld"1lce of_amphibian species captll.n:d in pitfall tt3ps for the DEMO e:,pemnental green-tree retention study in Oregan and Waslringt0n during 1995 and. 1996. Abundsr.ce is inversely correl:ued with Illilk (Je., I indicaces highest abundance). ·
Species CommonNamc
Ensar.,,.a eschscholJ:,;ii Ens¢na Pfethodcn Yeiucub:m . Westem red-baclred sal.amandzr Ascaph,,, tT'.tei Tailedfrog Amlrjsroma grad.le .
·Nonhwestem salamander Ta:richa gro:r.ulosa Rollgh-skinnoi! Oewt
· Ambystoma macrodacty/um Lorig-toe.rl salarnancier Rhyacotriton ca.:;cadae Cascade toaent sa-1e:mander Rana .aurora Red-legged frog Ranacascaaae ' ,. Cascades frog
'Hyla regiila Paci.fie tree frog Ranaspp. Unidentified frog Dicam:ptodcm ter.ebro..'7!3 Pacific giant salmnaodc:r Bufoboreas Wesrem toaJI A.neides ferm,.s Oouded salarnande: Pletlu:xkm dunJri Dunn's salamander Mean annual total wdi\idll2ls
1 - = not captured • nd = Rana species not ~din Oregon blocks •·na = species' range oll!Side of sample bloclcs
We primarily will use indices of abundance as treatment resporpe variables .rather than indicators of realized fitness {e.g., juvenile survival). However, because density or abundance estimates can be misreading indicators of habitat .quality (Van H=e 1983), we will attempt to esfiI!late indices of fitness for vertebrates when feasible. Nest searches and ten:itory mapping in. conjunction with counts will help to determine treatment
. effects on the number of breeding bird tenitories. Determination.of the a.,ae class and reproductive condition of small mammals 'llld amphibians captured in pilfall traps will provide P-Stimates of age-class distnoutions and reprodnctive condition ill treatment mii.ts. Age-class distributions and estimates of reprodnctive condition in livecaughtrodents ( chipmunks, flying squirrels) will help to indicate realized :fitness relative to trea1-p:1ents. The presence or absence ofbatroosts will indicate sensitivity of bats to tr"...atrnents.
The costs associated with these smdies are very liigh. It has been necessary to reworlc the sampling design in response to budgetary shortfalls. These considerations have led to reduced sam-
0rngQ!! B!gcls;; W~~WiltaQD B]Q~~ Little
Watsco Lialc Layng Dog White Paradise Capitol Falls Rivet Cr-..clc Prairie Butte Saimon Hills forest
I 3 l 1 2 1 5 l - ' .- . 2 3 HJ 2
1 3 l 6 2 l 5 2 ~ 2 2 3 3 2 3 6 5 4 4
4 7 8 3
ml' nd ' nd. ml 5 10 11 5 nd nd nd nd 6 6 4 3 4 10 9 5 6 3 nd nd nd nd ,- s 6 s· 8
7 6" 9 4 4 3 na• na na na
6 na ca na na 55 114 90 140 56 204 132 467
piing efforts for arboreal rodents· (reductions in trapping intensity per period and loss of seasonal' sampling), sma TI mammal.< (eliroinatinn of simultaneous live trapping), 3ll.d ampbilnans ( elimination of time-constrained searches).
Our final concern =o!!Ilizes that net all responses to treatments· will-occur within the initial 2-yr post-harvest sampling period., Wee~- pectlagresponses for several species groups within the amP.hibians, arboreal rodents, and birds. We expectspecies packing in the agg:regate4 reten- . ti.on patches for species in all of these groups, a phenomenon that may not begin to dissipate until the second or even third post-harvest year: These concerns point to the need for intensive, longterm sampling to understand how key forest speci~ or species gronps will respond to staud-Ievel manipulations, the effects of wbicbmay be manifested over decades. Funding agencies must like·wise be willing tq pro.,ide continuing supporrif tbeywishcosuccessfuilymaoageforspecies whose responses to harvest actrnties may be dy.namic over Iong periods of time.
v:errebrate Response to Greerr-iree Retention 59

I
Acknow}sdgements
We thank the vital and tLn;less work ofpe<Somiel on the Umpqua and Gifford Pinchot National Forests :i:nd the Capitol State Forest: Brenda Wcodard, Rick Abbott, Cindy I.a~oudakis, Barbai"a Fontaine, and Wayne. Kleckller (Um:{'qua National Forest); run White, Debbie Couche, Sue MacMeeken, Jon Nakae, and Ed Tompkins (Gifford Pincho'\ National Forest); and Richard Bigley (Capitol S~Forest). We could not have accomplished the collection and analysjs of data without the assistance of Chris Ma.,auire, Tom Manning, Jeff Feen, Rebecca Thompson, and
Litsratura Cited
. Abbott, R S, 1. D. White, and B. L Wocdard. [99g. The bettefits and cilailenges-of large.scale si!vicallmal "'Jlerimems: perspectives from forest """'2gers on !he Derocnstraiion of:EcosystemManagemem OptiO!)S (DEMO)Stlldy. Nonnw. Sci. 73(Spee.·Iss.):118-125.
",roeric:anSccietyofMammalogjsts. 1987. ~le6.e1d methods in marnrnaJogy: prelimma,y gmdeiines approved by the Amcri= Society of Mamma!ogist:s. Supplem~ to volllme 68(4). !8 p;
Artman. V. L 1990. B=ding bird populations and veget!· tion c~rislics in commercially tirinned and anthinned we.stem bemlcclc forests of Wasbing10,L
· Ullivernty ofW~ Sean!e. M.S .. Theru. Aobry, K. B, and P. A. Hall. 1991. T=n:ia! zmpltibi.an
commmntics in the southern Wasiringron casca~ R:a,,ge. 171 I. f. Ruggi=, K. B. Anbcy, A.. B. Ca:tey,. and M. Et Him (tecli. ~o,ds.). Wildlife and vege;ation of mmi.mage:i Douglas-fir-forests. USDA For. Serv" Gen. Tech. Rep. PNW-OIR-28.S. Pp. 327-338.
Aubry, .K. B, M. J. Crites, and S. D West. 1991. R~oiw • patt?:rnsof S!IlJlllmaurrn,J al:andan~andcommm,ity
C<>mpo_sition in Oregon and Washingron. In· I. F. Ruggiero, K. B. Anbcy, A B._ C-;, and .l,,L H. Raff (fe!;b. ccords.). Wildlife and ~eblion nf anmacag,,d Docglas-fir iimsts.·USDA Fer. Serv. C-en. Teclt. Rep: PNW-G'IR-285. Pp. 285-294.
Aubry, K. B., M. P. Amararubus, C. B. Halpern, J. D. W.qite; . B. L Wooda.'<i, C..E. Pe=cn. C.A. ~"<>nrlakis, and
A J. Horron. 1999. Evaluating !!le effects of varying levels and patterns of green-tree retention: e:qietin:.u:nt:tl design of tile DEMO stndy. Ncnhw. Sci. 73 (Spec. l'ss.):12-26.
Batclay, R. M. ~ P. A. Faure, D. R Fm, and R t)anieL 1988. Roosting behaviour and roost selection by mi-. grating silver-haired baJs (µ,simrycteris noctavagar.s)._ J. Mammal. '69:821-825.
Bosalrowski. '.I: 1997. B=<iing bird abnndance and habitat re!.ltionships on a privaleindustcial£cres in 1he westem· Washln,,aton casc3des. Normw. Sci. 71:87,%.
60 Lehmkuhl et al
Tiffany Chnrcli Lai:;:y Bednar did the power analy~ · ses . . rnn White .and three anonymous referees provided critical reviews of this paper. We ben
. efited greatly from the insightful editorial. comments of 1'f..artin Raphael and Charles Halpem. This is a product of the Demonstration of Ecosyst~m ~ement Options (DEMO) study; a joinr e_ffort of the USDA Forest Service Region 6 and Pacific Northwest Researcli Sration. Research partners incinde the {!o.iversity of Waspington, Oregon State University, University of Oregon, Gifford Pinchot and Urnpqua National Forests; aitd the Washington State Department of Natural Resources.
Brown, E.R. (tech. ed.). 1985. Managcmentofwilillife"!'d fishbabitlm,,infortsts of wes1em Oregon and Washingron.' Pon ~P!'endices. USDA Fat Serv, .R6-F&WL-!92-1985: USDA Forest Service, Pacific
... Nonhwest ~on. Pcrdand, OR 302 p. Bory, R. B. 1983. Eabitai relationshios snd ecological im
pottance of amphibians andreptiles. 1n K. r. Raedeki (ed.). Proc. S:;,nl£. st=msiden:ianagem.em: ripar:iru, wildlife and forest,:y int=ctions. Institute of Forest ~= u~ ofWashingt<lll, Seanie. Pp. 61-76. •
Campbell, LA. 1993. Bot div=icy ·aad habitd use in a ~lld != Wa.shington Stare Universiiy, Pnllma,,.. MS. Them. .
Carey, A. B. 1991. The biology of 3IOOiea! rodents in Douglas-fir foresis. ·USDA Fm: Ser:, Gen. Toch. Re;,. PNW-GTR2i6. .
· 1995. Sciurids iD Pacific Northwest Illllllllged - -an~ d old-gromb forests. Ecol Appl 5:648-¢61. · . Ca:tey; A. B., and M. L Jollllson. 1995. Sm,,n maT1T1Dals in
managed, iwux.illy young, and old-gzpwtb. forests.. Ecol Appl 5:336-352 . . .
Carey,A.B, B. I. Biswe!l. andJ. w. ·w·ttt. 199la. Methods for measuring Dopulations of atborea!'mdellls. USDA
. Fe~ Serv, C-en. Tech. lt.-p. PNW-GTR:2u. · Carey,A. B, M. :!>i Har-Jt. S..P. Honon, andll. Biswell. 1991b.
Spring bird COlllll!anitles in the Oregon Coa.,,-t RJID,,~ 171 L F. Ruggiero, K. B. Aubry, A.. B. Carey, and }.i
H. Hnif (r.cli. coords.). Wildlife a;ui'vegeution oi • tlDlJlmt3g<><l Docglas-fir;foresrs. us_DA Fee Scrv
7 Gen..
Teci!.. R.eo. PNW-OI'R-285. Pl>. 123-144. · Carev, A. B, D.R. Thyscl]. L J. vin;, T. M. Wilson, S. :!>~.
• W-J]son, J. ~1 T.i;q,pe, W. eo1gan; II!, E. R: Ingilam. and M. ·Holmes. 1996. Foundations of biodi:=sity in, ~ed Do~-fir forests. b: D. L PeteISon and C. V..Klimas (eds.). Tne role of ~on in, ecosystem ~cement. Society for Ecologjc:al Res-toration. Madison, WI. Pp. 68-82. ·
Cu:raway, L N. 1990. A ll)orphologic .and tnorphomettic a.ualy,is of the • Sora vagrw,s species comple:(' ll! the Pacffic ·ecast ~on. Special PubL 32 Tne :Mosewn of Texas Tech Unive,sity. Texas Tech. Unlversity Ptess, Lubbcck.

Car'~, R C0 and M. L. Mo:c:iso11 (eds.). l992. Starns and · ccnservation of Ille matbied m=.let in North Amerita
Proc. We,.<t. found. Y= Zoo!. S(l). .
Forsman..E. D, E. C. Mes!ow, and H. lvL Wight. 1984. Distriborioa and biology of !he spomed owl in Oregon.
Cazares, E, D. L Luoma, M. P. Amaranthus, C. L. Cbambe.s. and J. F.Lehml:uhl. 1999. Inrer:lction of fun. gal S!)OrOcaIJl prodDction with srnll m3Tllll)al 3bundance and diet in Dcnglas-fir stmds of the sou.rhem Cascade Range. Northw. Sci 73 (Spcc.Iss.):64-76. -
Cllambers, C. L. 1996. Respocse ofteri:eslrial ver-.ebi:ates to three si!vic:ul!nra!.n:cannCD.tS in die central Oregon Coast Range. Oregon State University, Corvallis.
-Ph.D. Dissertation. ' Chambets, C. L., and W. C. McComb. 1~97. Effects of sil
vic:ulnJml .treatm.e:us on winmriag bird communities· in the Oregon Coast Range. 1'iorihw. Sci 7.J:298-304.
· Chambez., C. L, Y,. C.M:Co.mb, 'andJ. C. Tappeiner, IlI. In press. B=ding bird community responses to three silviculnual tteattneuts in the Oregon Coa..<t Range. EcoLApp~ ..
Chambers, C. L., T. Caaigan, T. Sabin, J. Tappeiner. and W. McComb. 1997. Use of arti:fi..'ially created Do11glas:fir snags by cavity-nesting birds. Wesc I. AppL For. . 12:93-97.
Cbristy, R. E.. 1993. Ridiotr.lcl:iog Myotis bais 0
0n Long · Island. Washington. Unr.mty of Wasb.i.ngt0n, So
attle: M.S. Thesis. Com, P. S, and R. B. Bury. 1991. Te:resttial ampltfcian
communities in die Oregon Coast Range. 1n L. F. Ruggiero, K.. B. Anbty. A. B. Carey, and M. H. Hoff (rech.cootds.), 'i\--ildlifeandvegetationofnnmaoaged Douglas-fir forests. USDAFor. Sert, Gen. Te--...h. Rep. PNW-G:rR-285. Pp. 305-317. .
DeBeU. D. S., R. 0. Curtis, C. A. H:uri:ngtcn. and J. C. Tappeino,:. 1997. Shapi!lg stand developmem t!]rongh silvicoltme p.ractices. In K. A. Kohm. andJ. F. Franklin (eds.). CreatiDgaFo.restzyforthe2lstCe;nury: The Science of Ecosystem ManagemeoL !s!and Press, W2Sbmgn,o, D.C. Pp. 141-150.
Doyle, A. T. 1990. Use of rl~ and upland babitats by small OOillIUDals J. Mammal 71:14-23.
Erickson. l. L. 1993. Bat activity in· managed forests of ihe scmhwesrem Cascade ~oe. Ulliver.,ity ofWashingtoo, Seattle. M.S. Thesis.
Erickson, J. L, ands, D. West 1996. Maru,ged forests m the western Cascades:. the effi:cts of seral Sll¾,o-e oo bath2bitat use patterns. In RM R. Barday,'1Jld R. M. Brigham (eds.). Bats and fOf".sts symposium. Brit. . Col Min. For, Victoria, B.C. Pp. 215-227.
FEMAT. 1993. ~ecosystemmanagemcnt anecologi0
cal, economic, and social. assessment. report of lhe Forest Ecosystem Management Assessment Team. USDA For. Serv.; USDCNat. Ocean. Atmos. Admin. and Nat. Mar. Fish Ser,.; USDIBnr._LandManage., Fish Wtldl Scrv., and NaL.Parli: Serv.; and US -EPA, Wash.ingmn, D.C.
Fenton, M. B, D. H. M. Cumming, L L Rmlloobacl,. G. S. · . Cnmmiog, M. S. Cwmnil1g, G. Fo.rd, RD. Taylor, J.
Dunlop, I\IL D.Hovo,xa,D. S. Johnston, C. V. P-Jrtfors, M. C. Kalcounls, and Z. Mahlanga. 1998. Bats and the loss of 1ree canopy inAfric:m wcodland$. Conserv. Biol. 12:399-407.
Fcge~ R.. and J. M. Tta!'!J". 1978. Fungus con.<nmption (mycophagy) by small D1llllllllili. Northw. Sci. 52!1-30.
WiklL MO!JO!,l: ip:J-64, . Fors=, E. D, S. DeStefuno, :M. G, RapbaeL R. J: Gutierrez.
· 1996. Demographyoftheaonbem-spottedowL Stud-· les in Avian Biol.17:1-122.
Franklin, J. F., and RT. Fom:an. l98i. Cresting landscape pattern., by forest cutting: eeologicai consequences and principles. LandscapeEcol l:5-18. _.
Frnnk:lin, J. F,, D. R Be,g, D. A. Thornbwgl!, and r. C. Tappeiner; 1997. Allernative silvicoltnral approaches to timber harvesting: vm:iable retention barvest sys.
· rems. bt.K.A. Kohm,andJ. F.FtankJin(eds.). ere. ati:.ng a Forestry for tho 21st Centmy: Tiu, Science of Ecosystem Management Tsland Press, Wasjrington, D.C. Pp. 1!1·139.
Frnn:ueh, K. 1:. W77. Bird popttlat:ion changes after timber ha..estingofamixedconiferfo=inArizooa. USDA FOL Serv, Res. Pap, .RM-184.
Frazier, M. W. 1997. R00:5t site chatactcristics of the lon_gleggedMyotis (,1dyotis valtm.s) m ihe Teanaway River Valley of Wasbingion. Uo.ivemty of Washmgtoc, Seattle. MS. Thesis.
Gilb->...r,,F. F,aruiRAilwfue. 1991a. Spring~irocommunities in the Oregon Clisc:ade Range. bt L F. Ruggiero, K. B. Au:),ry, A. B. C=r, and M. a Haff (tech. . COCids.). '11-iidlife and vegetation of nn:maoaged Douglas-fir foreso;. USDA For. Serv., c;,,,,: Tech. Rep. PNW-OTR-285. Pp .. 145-160.
- --~- 1991b. Tem:strlal amphibian CQll!mllllilies in cl!e Oregon Cascade Ran,,ae. JnL. F. .Ruggiero, K. B. Aubry, A. B. Carey, and M Ft Huff (tech. coc.td,;.). Wildlife ai:ld "Vegetatjoa of UllllWlaged Douglas-fir forests. l'SDA Fa,:Setv, Gen. Tech.¾>. PNW-GTR-2&5. Pp. 31~324.
C-oznez. D. lvl..1Uld R. G. Anthony. 1996. ¥JPhibian arui reptil~ abundance intiparian and upslope~ cf f!ve · forest types in Western Oregon. Nartbw. Sci. 70-. I 09-119.
Good, D. f',.. 1989. Hybridization and cryptic species in Dicamptca,m (Canliata: DiCillllptcdontidae). Evolu- . tion 43:728-744.
Good, D. A., and D. B. Wake. 1992. Geographic variation and spe,."iation ill; the tozreru salamanders _of the genus RJ,yco:r.trm. (Caudata: Rhycotritontidae). Umv. Calif. Pnbl. ZooL 126:1-86.
.G=, D:M., H.Kai=, T.F. Shamel, i. Kerasley, andK. R. McAllister. 1997. Cryptic species of SJ)llttro frogs, Rona preriosa com~ in wes'.em N\}rth America Copeia 1997:1:.g.
Hagan. J. A., m, and D, Jolulston (eds.). 1992. Ecology and Con=vatioa of Neotropical-Migrant Landbirds. Smithsonian Instimtiool'ress, Washinaaton, D.C.
Hagar, J. ~w. 9- Wa:Comb, and w. H. Emmingbam. 1996. · Bird co.minllllities in commercially thinned and
Ulltb.inned Doul!!as-fir stands of western Oregon. · Wild!. Soc . .Bu!L 24:353-366. ·
Hal!llleier. E. M., :md C.D. Sllllts, i964. A numeridtl aoaiy- sis of the distnoutiona! patterns· of North ,•,,nerican
ma.w.mals. Syst. Zoot 13: 125-155.
Vertebrate Response to G-reen-tree Retention 61

,
Eaipem, C. B., S. A Evans, C.R. Nelsen, D. McKenzie, DUguori, D. E. Hibbs, and M._ G. Halaj. 1999. Response of fon:st vegetation to v11rying levels and pattems of green-tree retention: an oven-i.ew of a-long~ term "-'1>erimem. Nonhw. &i 73 (Spec. Iss.):27-44.
lfansen, A J, and P. Hounih:m.. 1996. Canopy tree retention and avian diversity in the Oregon Cascades. In R. C. Szaro, and D. W. Johnson (eds.). Biodlvemcy in Managed LruJdscapes: The<ity mi Practice. Oxford Univmicy Press, New Yorlc.
Hansen, A. J, W. C. McComb, R. Vega, M. G. Raphael, ·and · M. Hunter. 1995. Biro habiw re!atiOilShi:os in natu
ral and managed forest in the wesr cascades of Oregon. F.coL App[ S:555--569.
Haris, L. D. 1984. 'IbeFragmented Forest. The University of Chicago Press, Chicago. •
Hayes, J. P, and M. D. Adam. 199'6. The inflD= oflogging riparian areas on babilat lltilizatiOll by bal5 in wesrem Oxegon. In R. lvL 1t Ba,cmy, and R. .M. Brigham (eds:). Bais and forests symposlllllL Brit. C:oL Min. For.., Vietoria,B.C. Pp. 21.5-227.
Hooven. E. F., and H. C. Black- 1976. Effects of some cleat=ting practices on S!Jlall-=na.! populations · in Western Oregon. Northw. Sci 50:189-208.
Haff,M.H.,D.AManuwaJ,and-J.A.Pmera.1991. Wmrer bird communities in 1.~e soathem Washington Cascade Range. Jr. L F. Rilggiao,_K. B. Anbzy, A B. Ca.-ey, and M. H. Half (tech. coords.). Wildlife and vegetation of nomSIJaged Dougtas---fir fetests.. USDA For. Serv .• Gea Tech. R,:p. PNW-OTR-285. Pp. 207-220. -·
Keller. M. E., and S. a Anderson. 1992. Avian use Qfllat>i· ll!! configurations ~ by furest etttting in sombeas= Wyomi,tg. Condor 94:5~;
Kobm.. K. A, and I . F. Franklin (eds.). 1997. Creating a Forestry for the 21st Cen!Dr)o The~ of Ecosystem Management. Island Press, W~<>tan, D.C.
Kunz, T. E., and A. Kmta. 1988: Capture IJlcihods snd holdmg dev',ces_ In T. H. Kunz (ed.). Ecological and Be.b3vioral Methods for the Study ofBat.s. Smithsonian lnstitmion Press, Wa.shlA.,"lOn, D.C. Pp. l-29.
Lebmkubl, r_ F., and L F. Rllggiero_ 1991. Forest fragmentation and iis poremial dfectS on wildlife in !be l'a
:ci:fic .~orthwest In. L F. Rllggicro, ·K. B. Anbzy, A B. ·earey, and M H. Huff (t.ecll. coards.). Vftldliie and vegetation of tllllnallaged Dongl,is-.fir fcnests. USDA For. Serv, Gen. Tuch. Rep. PNW-Gl'R-285. Pp.35-46.
UODard, W. P.,H.A. Brown.LL C.Jones,K. R.McAllistcr, and R. M. -Swm,_ 1993. Ampln'bians oiWasbington and Oxegon. Seattle Audubon Society, Seattle.
Li, C. Y, C. Maser, Z. Maser, and B. A. Caldwe!L 1986. Role of three rodents in forest nitrogen fixation in western Oregon: anothe:raspectofmammaI-mycOIIbizalfungus-tree mutua!ism.. G=t Basin Nat 46:411-414.
Llmpens, H. I. G. A., and K. KapteY!J. 1991. Bais, their behaviour and linear landscai:e eieme.us. Myotis 29:39-48. • .
Lanney, D., J. Barl:et, and D. PtiddeL 1985. Movcmenis and day roosts of the cbccolate wattled bat ChaliM/obu.smorio (Gray) (Microchh--oprem: Vespertilionidae) in a logged fcx-.st. Au;t Mammal. 8:313-317.
62 Lebmkulil et al
Mannan, R. W, and E- C. Meslow. 191!4. Bird population and vegetation chanu:leristics in maru,goo and oid growth forests, northeastern Oregon. J. WildL Man
. age. 48:1219-1238. Mannwal,D.A.1991. Sptingbirdcommllllities in theSoutbe::n
· Washingt0n Ca.seode Range. Ir. L F. RllggiCIQ, .K. B. Aubcy, A. B. Carey, and Jl,L H. Hun (tech. COords.). Wildlife and vegetation of nomzmagetl Dougias,.:frt forests. USDAFoi: Serv, Gen. Te::b. Rep. PNW-GTR-
. 285. Pi,. 161-176 . . Mar.ell, A. M: · 1983. Changes in small mammal <OI!Jmtmi
ties after logging in llOrth-«ntraj Ontario. Cao. J. ZooL 61:970-980. .
Martia,:r. E:, andD. M. Finch (eds.). 1995. Ecology and Management of Neotropical Migruo;y Birds. Ox:-
. ford Ulliversity Press, New Yorl<. McComb, W. C., C..L Cb.mibers, "'1d M. Newton. 1993a.
Small mammal ~d ~phibian cnmmnnities and habitat associations m :te<!,aldcr stands, Cem:raJ Oregon Coa..<tRaQgc. Nonhw. Sci. 67:181-188.
McComb, W. C., K. McGarigaL and R. G. Anthony. 1993b. Small mamm:tl and mnpllioirul abundznce in·stream.side and upslope habitats of manm,Donglas-fir stands, w=m Oregon. Northw. Sci. 67:7-15. · . .
McGmgaL K., and W. McComb. 1992. Streamside ve::sus upslope breeding bird COmmnnities.in tj,e connal Oregon Coast.Range. J. Wild!. Manage. 56:10-21.
- - - ~ 1995. Relationships lietween landscape stnJc. • IUic and lneding birds in tile Oregon Coast Raot;c. 5:oL Monog,: 65:235-260.
Medin, D. E. 19&5. Small mammal ""SOODSes to diametercut' logging in an Idaho ~Lit f=i. USDA For. Serv., Gea Tc:,;h. Rep. INT--GTR-362.
Medin, D. E, and G. D. Booth. 1989. ~1>cnses of bil:ds and sm.aJl mammaJs to single~tree selection loggfug in Idaho. R.es. Bull., InteancantainR.se=h Stllt:ioo, Ogdeq, Utah. . .
Momsen, M. L, and R. G. A.mhony. 1989.' Habitat ose by small mammals· on early-growth ciear:CUnings fu wesiem Oregon. Can. J. ZooL 67:805-811 •
Momsen, M. L,.M. R. Dedon, M. G. Raphael, M. P. Yc_del. 1986. Snag requirements of cavity-nesting wrds: are USDA Forest Service gnidelines being met? West. J. Appl FOL l:38-40.
Nelson,S.K.,andS.G.Scaly(eds.).1995. Bialogyofmsri>led mmrelets-illland and at sea. Notthw. Nat. 76.
Nussbamn, R:A.; E. D. Brooi~Jr., andR.M. Stam,. 19.83. · Amplnoians and R,epliles of the Pacific Northwest.
Onivrzily Press ofldaho, Moscow. Oijver, C. D~ and B. C. Larson. 1990. Forest Stand Dynam
ics. Jobl! WJ!ey and Sons, Lid., New York. Otmsbee, P. C. 19%. Cba.-:icmi4.s.ics, nse, and distnouticn
of day roosts selected by iemaleMyox£; volans. b.-R. M. R llarcl,iy, and R.1/L Btigham (eds.). Brus and forests.symposium. Brit. CcL Min. FoL, VlC!Oria, B.C. _ Pp. 124-13!. '
Ralph. C. J., G. R. Geupel, ;!'. Pyle, T. E. Martin, and D: F. DeSsnte. 1993. Handbook offie!dmethods fur momtociD.g landbirds. USDA For. Serv., Gen. Tech. Rep. PSW-GIR-144.
Ralph, C. J., G. L. Hunt, k, M. G . .Raphael, and J. F. Piatt (eds.). ·1995, .Ecology and conservation of tile marbled mlliiclet. USDA Fer. Serv .. C-en. Tech. Rep. FSWGTR-152.

Ramirez, Jr., P, and M. Homocke:r. 1981. Small ~ populations in different-aged cl=tS in nonhw6t
. em Molllana. J. Mammal.. 62:400-403. Raphaei, M. G. 1988. Lor.g-tenn lreilds in abnndance of
amplu'bians, reptiles, and mammals in Douglas-fir fo:rtsts of ncmheastx,rn California. In R. C. Szaro. K. E. Siev~on, andD. R. Patton (tech. coor<is.). Mznagement of an,pluoians, reptiles, and small mammals in NonhAmerica. USDA For. Serv, G<:n. Tecb_ Rep. R.¼-GTR-166. P.p. 23-31.
- - -=-· 1990. Mammals of the anciem til.-est. In E. A.. Nme (ed.). Anciem Forests of the Pacific Northwest Islruid Press, Washingron, D.C.
Reynolds, R. T., i. M. Scott, and R. A. NllSSbamu. 1980. A variable circular-plot m¢iod for estimalulg birds. Condor 82:309-313. ·
Rohwe:t. S., and C. Wood. 1998. Three hybrid zones between hermit and Townsend's· warnlers in Washing• ton and Oregon. Auk 115:284-310.
Rosc:nbe,g, D. K.; and R. G. Anthony. 1993: Differences in TOWDselld's chipmunk populations~ second· and o.ld:l,rowth forests in ,..estem Oregon. r. Wild!.
· Mulage.-57:365-373. . Rnggiero, L. F, K. B. Anb,y, A. B. Ctrey, and M. H. Haff
(rech. coords.). 1991. Wildlife and vegetation of tmmaJJaged Doagjas-:fir forest;. USDA For. Sciv, Gen. Tech.Rep. PNW-OTR-,85. . .
Sharp, B. 1990. Population ll'Cnds of Oregon's neotropical migrants. OregonBi:rds 16:27-42. ·
--- 1992. Neotropical migrants on national fore,.s: a compilalion of t:.<istii)g infozm:ll:ion. Prepared for the USDA rorestSemce by B. E. Sba.-p, Ecological Perspeetives, 2234 NE 9" Avenue; Portland, Oregon 97212. ·.
Simpson. G. G. 1964. Species density of North American rteem roarnroals Syst Zoo!. 13:57-73.
Skalski, J. R., and D. S. Robson. 199.2. Teclmiqncs fur Viildlife Inveslig;,tions: .DesignandAna!ysisofa,ptnreDara. Academic PI=, San Diego.
Spies, T. A., and J. F. Fr.mkiin. 1989. Gap~ and vegetation response in coniferous forestS of the Pacific northwest. Ecology 70:543"545.
Endnote 1 The u.sc of trade or fum =es is for re:ider infonnalion cnly and does nor imply endo=ent by the U.S. Depa,tmenc of Agriculture of any prodncc or service.
Tappciner, J. C., D. Lavender, J. WalSlad. R.. 0. Cunis, 3l!d D.S. DeBcD. 1997. Silviculmral S}-stems and regeneration .methods: current practices and = alren-.a' tives. hiK.A.Koinn;.and J.F.Fr"'1klin (eds.). Creating a Forestry for the 21st Centmy: Th,: Science of Eco.system Management. Island Press, Wasbmgton., D.C. PD. 151-164.
Thomas, D. w'. 1988. The disln'bution of bats in different a,,oes ofDouglas-fuforest:;. J. Wildl. Mllruge.52:619-626.
Thmnas, D. W, and S. D. West. 1991. Forest ~e associations ofbal; in tbe Washington Cascades and On,gon Coast Ranges. In L F. Ruggiero, X:. B. Anb,y, A. B. Carey, and M. H. Huff (tech. CQOids.). Wildlife and vegetation of nnrom,aged Douglas-fir fOI"...sts. USDA Foi: Serv. c-,,n. Tech. ~-PNW-OTR-285. Pp. 295-303.
USDAandUSDl 1994. Recordofdecisionforamendincm,s to Forest Sc:i:vlce.and Bu:roau ·of l.aod l'>,fanageznent planning docnments wi1bin.tbl, range afthe northem soottcd owl. USDA FoL Sciv., 1'artl:md. Orelll)n.
Van Horiie, B. 1983. Density as a'misieedin2: md:icator of babi=quality. J. Wtldl. Manage. 47:893-901.
Vega, R. M. S. 1993. Bini colllilnmities m man,,,,oed conifer stands .in the Oregon Cascades: habitat assoc'.ations
· and nest predation. Oregon Slate Uttiv=ity, Corvallis. MS. Thesis. ·
v=, B. J, and L N. C=way. 1998. Land Marornsis of °""-"On. University of California Press. Jlcrl:eley.
. V~, P. D .. ML E!uni.r, and J. V. Wells. 1992. Use of a n,:w ceprod,,ctive i:ridex to ev:llna!e relationships betwe<:n habitat quality and breeduig :;uccess. Aok 109~97-705.
Von Treba, C .. D. P. Lavender. and T. P. Sullivan. 1998. Relalions of small !llall!lllcl popalaliollS to evi,n-aged shelteiwood syswns in sul>-boreal spmo, farest. J. Wildl. Mmiage. 62:630-642.
W=. J. R.. and C. J. Zabel. 1995. Nor.hem flyi:ng_sqwrtel dci'-~es in fir forests of northeastern California. i. Wtldl. Manage. 59:858--866.
Vertebrate Response to Green-tree Retention 63





























