Embed Size (px)
Citation preview

Copyright C Blackwell Munksgaard 2002Traffic 2002; 3: 452–460
Blackwell Munksgaard ISSN 1398-9219
Review
Amphiphysins: Raising the BAR for Synaptic VesicleRecycling and Membrane Dynamics
Bing Zhang1,2,3,4,* and Andrew C. Zelhof5
1Section of Neurobiology, 2Section of Molecular Cell and
Developmental Biology, 3 Institute for Cellular and
Molecular Biology, and 4 Institute for Neuroscience,
University of Texas, Austin, TX 78712, USA5Department of Biology, University of California, San Diego,
La Jolla, CA 92093, USA
* Corresponding author: Bing Zhang,
Amphiphysins, members of the BAR (Bin-Amphiphys-in-Rvsp) protein super family, have been postulated toplay a key role in clathrin-mediated endocytosis of syn-aptic vesicles (SVs). This review focuses on recentgenetic studies of the role of amphiphysins in SV re-cycling and membrane morphogenesis. In the mouse,brain-specific amphiphysin I and II regulate, but arenot essential for, SV recycling. The role of this regula-tion appears important, as mice deficient in these pro-teins have seizures and are deficient in learning andmemory. In the fruit fly Drosophila melanogaster, am-phiphysin is found in muscles and is enriched at post-synaptic membranes of neuromuscular junctions(NMJs); however, it does not play a role in SV recyc-ling. Rather, amphiphysin in fly muscles appears toregulate the organization and structure of the muscleT-tubule system and possibly the subsynaptic retic-ulum. Amphiphysin is also involved in membrane or-ganization in both neurons and non-neuronal cells inDrosophila. These studies reveal pleiotropic functionsfor amphiphysins in clathrin-mediated endocytosis andthe regulation of membrane dynamics, perhapsthrough the actin cytoskeleton.
Key words: actin cytoskeleton, clathrin, Drosophila,endocytosis, morphogenesis, muscle T-tubules, photo-receptor, synapse
Received 1 April 2002, revised and accepted for publi-cation 16 April 2002
Efficient signaling is of fundamental importance to all func-tions of the nervous system, including behavior and cog-nition. The major way neurons signal to their targets (neuronsor muscles) is through chemical transmission across the syn-apse. Chemical transmission at the presynaptic terminal in-volves release of transmitter through exocytosis of synapticvesicles (SVs) followed by the recycling of SVs. The latterstep is crucial for the continuation of synaptic transmission
452
because the number of quanta released during a short burstof intense nerve activity is much greater than the total num-ber of synaptic vesicles present in a nerve terminal (1–4).Although newly synthesized vesicle proteins can be trans-ported from the cell body to the nerve terminal, axonal trans-port occurs too slowly to refill the vesicle pool at the requiredrates. Hence, the only way to rapidly replenish SVs is to re-cycle them locally.
In most nerve terminals, SV recycling is accomplishedthrough clathrin-mediated endocytosis, a pathway com-monly used for nutrient and receptor internalization on theplasma membrane as well as for protein sorting and vesiculartrafficking in the trans-Golgi network in all eukaryotes [forrecent reviews, see (5,6)]. Current evidence suggests that re-cycling of SVs involves highly coordinated sequential stepsof protein–protein and protein–lipid interactions. These stepsinclude: (a) the formation of clathrin-coated vesicles; (b) fis-sion of coated vesicles; (c) uncoating of clathrin and ad-aptors; (d) reloading nascent vesicles with neurotransmitter;and (e) translocation of vesicles to vesicle pools [reviewed in(7,8)]. Amphiphysins have been implicated in vesicle fission,but their exact role is not known.
The initial, and perhaps the most dramatic, demonstration ofthe requirement of a single gene product in vesicle fissioncomes from studies of the Drosophila shibirets (shits) mu-tants. shits flies display temperature-sensitive paralysis dueto conditional impairment of the GTPase activity of the cyto-plasmic protein dynamin (9–12). At elevated temperatures,SV recycling at nerve terminals in shits mutants is completelyblocked, leading to vesicle depletion and the accumulationof endocytic invaginations on the plasma membrane (13,14).These endocytic invaginations are ringed by an electron-dense ‘collar’, which likely contains the mutant shits protein,suggesting that dynamin is essential for vesicle fission. Cellbiological and biochemical studies of mammalian dynamin 1,the shi homolog, support the idea that dynamin forms ringsat the neck of budding vesicles and detaches coated vesiclesfrom the plasma membrane at the step of vesicle fission[(15–18); reviewed in (19)].
How does dynamin find the neck region of coated vesiclesprior to fission? To accomplish this task, dynamin must inter-act with the plasma membrane and clathrin simultaneously.Although dynamin can associate with plasma membranethrough its PH domain, it does not directly interact withclathrin triskelia. A number of endocytic accessory proteins

Genetic Dissection of the Function of Amphiphysins
have been identified and one of these, amphiphysin, mayhelp recruit dynamin to the collar of coated vesicles. In thisreview, we will briefly summarize previous data implicatingamphiphysin as an essential factor in synaptic vesicle endo-cytosis. We will then focus on experimental evidence fromrecent genetic studies in the mouse and Drosophila revealingthe different functions for amphiphysins. In mice, amphiphys-ins do not play an essential role in vesicle recycling but onlyfine-tune the kinetics of the recycling process. In fruit flies,amphiphysin regulates muscle T-tubule structural organiza-tion and membrane morphogenesis and plays no role in ves-icle recycling.
Amphiphysins and Their BAR Friends
Amphiphysins belong to the expanding BAR (Bin-Amphiphy-sin-Rvsp) family proteins, which include the mammalianbridging-integrators (Bin1 and Bin2), amphiphysins, and theyeast Rvs161p and Rvs167p (Figure 1). Amphiphysin I, the
Figure1: Amphiphysins and the BAR family. Schematic illustration of the domain structures of vertebrate amphiphysin I, amphiphysinII/BIN1, BIN2, Drosophila Amphiphysin (dAmph), Caenorhabditis elegans amphiphysin (ceAmph), and their yeast homologs, Rvs167p andRvs161p. Only the N-terminal BAR (BIN/amphiphysin/Rvsp) domain is conserved among these proteins. Most members also have a highlyconserved C-terminal SH3 domain, whereas the central domain is most variable. Even though they belong to two different gene families,amphiphysin I shares the highest homology with the brain-specific amphiphysin IIa. They both have a proline-arginirie-rich domain (PRD)and a clathrin-AP-2 binding domain, which we name the CLAP domain. PRD and CLAP domains are both absent in all other BAR proteins.With the exception of the presence of an MYC-interacting site and a nuclear localization signal (NLS) in the muscle-specific amphiphysinIIb/BIN, the remaining members do not have any conserved central domains. The ubiquitous BIN1 isoform, dAmph, and ceAmph are closelyrelated to Rvps176p. BIN2 is similar to Rvs171p; both lack the C-terminal SH3 domain.
453Traffic 2002; 3: 452–460
founding member of the BAR family, was first identified asan SV-associated protein in chicken-brain extracts (20) andas an autoantigen in Stiff-man syndrome associated withbreast cancer (21,22). Amphiphysin I is enriched in presynap-tic terminals of mammalian brains (23,24). Amphiphysin IIhas at least three different splice variants, encoding a brain-specific protein (also called amphiphysin IIa) (24,25), a skel-etal muscle-specific protein (also called amphiphysin IIb)(24,26), and two ubiquitous proteins (27,28). AmphiphysinIIa is enriched in axon initial segments and nodes of Ranvierin the brain, but not in synaptic terminals (24). AmphiphysinIIb, one of the three isoforms of BIN1s, was discovered inde-pendently as a tumor suppressor that interacts with c-Myconcoprotein (26) and is enriched around T-tubules in skeletalmuscles. In Drosophila (dAmph) and Caenorhabditis ele-
gans (ceAmph) there is a single amphiphysin gene. dAmphis found in neurons, muscles, and epithelial cells (29–31),whereas no study has been reported on the nematodehomolog. The newest member of the BAR proteins is BIN2(32).

Zhang and Zelhof
All members of the BAR family share a highly conserved N-terminal BAR domain, which has predicted coiled-coil struc-tures required for amphiphysin dimerization and plasmamembrane interaction (33). With the exception of BIN2 andRvs161p, all BAR members also share a conserved C-terminalSrc homology 3 (SH3) domain, which mediates their interac-tions with dynamin and synaptojanin 1 in vertebrates (23,34–36) and with actin in yeast (37,38). The central region of allthese proteins is most variable. In mammals, the central re-gion of amphiphysin I and amphiphysin IIa contains a pro-line–arginine-rich domain (PRD) for endophilin binding (34)and a CLAP domain, for binding to clathrin (24,39) and AP-2 (23,33,40). The muscle-specific amphiphysin IIb and theubiquitous BIN1 isoform lack the PRD and CLAP domains.However, they have an Myc-interacting domain and a nuclearlocalization signal. Amphiphysin isoforms in Drosophila, C.
elegans, and the yeast Rvsps do not have any of these con-served domains in their central region.
In vitro perturbation studies of amphiphysins in
clathrin-mediated endocytosis
A role for amphiphysins in clathrin-mediated endocytosiswas suggested by genetic studies of the two yeast proteins,Rvs.167p and Rvs.161p. The genes coding these two proteinswere identified in a screen for mutations causing reducedviability upon starvation (hence RVS) in stationary phase(41,42). The growth arrest is most likely caused by defects inseveral cellular processes related to the actin cytoskeleton,including receptor-mediated endocytosis.
Several other lines of evidence also argue strongly for arole for amphiphysins in clathrin-mediated endocytosis ofSVs in complex animals [reviewed in (43)]. The multido-mains present in amphiphysins, their biochemical prop-erties, and subcellular localizations implicate amphiphysinsin clathrin-mediated endocytosis. First, mammalian amphi-physin I and brain-specific amphiphysin IIa both interactwith other key endocytic proteins, such as clathrin, AP-2,endophilin, dynamin, and synaptojanin. Second, amphiphy-sin I has been shown to be enriched and colocalized withclathrin, dynamin, and AP-2 at presynaptic terminals in thebrain (44). Third, amphiphysin I has been shown to bindto lipid bilayers and cause deformation of the lipid mem-brane in vitro (45), suggesting a possible role in vesicleformation or budding.
Among these protein–protein interactions, the one betweenamphiphysin and dynamin has received special attention. TheC-terminal SH3 domain of amphiphysin I interacts with thePRD of dynamin with high affinity in vitro (35). This interac-tion has been postulated to be essential for the recruitment ofdynamin from cytosol to the neck region of budding coatedvesicles. In an elegant set of experiments conducted in thelamprey reticulospinal synapse, Shupliakov and colleagues(1997) directly tested this hypothesis by attempting to disruptthe amphiphysin–dynamin interaction (46). Microinjection ofa peptide containing the human amphiphysin I SH3 domaininto the nerve terminal results in an activity-dependent block
454 Traffic 2002; 3: 452–460
of SV endocytosis and an accumulation of budding vesiclesalong the presynaptic plasma membrane (Figure 2B). Injec-tion of a peptide containing the PRD from dynamin also leadsto a similar endocytic arrest. It is interesting to note that inboth cases these budding vesicles are reminiscent of endo-cytic pits arrested in shits mutant synapses but, remarkably,lack the dynamin collars commonly observed in shits prepara-tions. These results indicate that amphiphysin I–dynamin in-teraction is essential for SV endocytosis. They further indicatethat amphiphysin recruits dynamin to the neck region of bud-ding vesicles (46,47).
This conclusion is well supported by parallel studies on re-ceptor-mediated endocytosis in cultured cells. Overexpres-sion of the SH3 domain from amphiphysin I blocks receptor-mediated endocytosis. Importantly, SH3 domains from otherproteins (such as Grb2 and phospholipase Cg) that also inter-act with dynamin in vitro do not affect receptor-mediatedendocytosis (48). In addition, the SH3 domain from spectrin,which does not interact with dynamin, has no effect on endo-cytosis (47,49). The SH3 domain-induced endocytosis blockcan be rescued by coexpression of dynamin (49), indicatingthat amphiphysin I SH3 domains specifically disrupt amphi-physin–dynamin interactions. Furthermore, a peptide con-taining the PRD fragment from dynamin also inhibits recep-tor-mediated endocytosis (49). These effects appear to bespecific to clathrin-mediated endocytosis of membrane sur-face proteins, such as transferrin and the epidermal growthfactor, but they do not affect fluid uptake via pinocytosis (48),though this has not been measured quantitatively. Finally,overexpression of the CLAP domain from amphiphysin I leadsto a defect in receptor-mediated endocytosis (40,50), sug-gesting that amphiphysin interactions with clathrin and AP-2are also important. The SH3 domain of amphiphysin II hasalso been shown to inhibit receptor-mediated endocytosis infibroblasts (51). Taken together, these studies support amodel in which amphiphysin I and II are required for the en-docytosis of both SVs and receptors, perhaps by recruitingdynamin to budding vesicles by interacting with clathrin-coated vesicles and dynamin simultaneously.
Although interpretations of the dominant negative experi-ments described above are most logical, other explanationsare plausible that may not support the role of amphiphysin inrecruiting dynamin to endocytic sites. For example, the ex-cess SH3 domains may absorb most, if not all, native dynam-ins in the cytosol, and consequently deleting dynamin frombudding vesicles. In addition, the amphiphysin SH3 domainhas been shown to inhibit dynamin multimerization and ringformation (51); the formation of these rings is thought to beessential for the fission step (19,45). This may explain theabsence of collars on invaginating vesicles as well as theblock of endocytosis. This may be the most likely scenario,since SH3 domains from other proteins that do not interferewith dynamin ring formation have no effect on receptor-me-diated endocytosis (51). Finally, the presence of high concen-trations of the dynamin proline-rich domain or the amphiphy-sin CLAP domain may interfere with the activity of dynamin

Genetic Dissection of the Function of Amphiphysins
Figure2: Schematic illustration of synaptic functions of amphiphysins. A. Proposed model of synaptic vesicle (SV) recycling inpresynaptic terminals. Following the formation of clathrin-coated vesicles, amphiphysin (green) is thought to recruit dynamin (red) to theneck region of a budding vesicle. Upon scission by dynamin activity, SVs enter the cytosol followed by a rapid removal of their clathrin coats.B. Microinjection of the SH3 domain of amphiphysin I into lamprey ‘giant’ terminals leads to a use-dependent blockade of SV endocytosisand accumulation of budding coated vesicles along the plasma membrane (46). Note that dynamin rings are absent from the buddingvesicles. C. When amphiphysin I is knocked out from the mouse, SV recycling is remarkably normal, presumably dynamin-dependent vesiclefission remain normal as well (56). D. Amphiphysin in Drosophila (dAmph) is not found at presynaptic membrane, but rather it is enrichedpostsynaptically with glutamate receptors and scaffold proteins such as Disc large (DLG, red) and Lethal giant larvae (LGL, blue) (29–31). E.In dAmph mutants, SV recycling is normal. However, LGL and DLG are delocalized from the postsynaptic area to the muscle surface. WhileDLG is still found in the postsynaptic area, LGL is absent (30).
or other endocytic proteins. It is conceivable that the dynaminPRD peptide could interfere with the functions of other SH3domain proteins, such as DAP160/Intersectin (52), syndapin(53), and F-actin binding protein 1 Abp1 (54). All of theseproteins contain an SH3 domain that interacts with dynaminvia the PRD and are implicated in clathrin-mediated endo-cytosis (55).
455Traffic 2002; 3: 452–460
Genetic dissection of the in vivo function ofamphiphysins in synaptic vesicleendocytosis
To further investigate the in vivo functions of amphiphysinsin SV endocytosis, several laboratories have turned to geneticapproaches to delete or ‘knockout’ amphiphysin genes. Re-

Zhang and Zelhof
cent genetic studies in the mouse and in Drosophila haverevealed unexpected aspects of the role of amphiphysins inSV recycling as well as other novel functions in neurons andmuscles. In mice, the removal of amphiphysin I also resultsin a reduction of amphiphysin IIa protein levels (56). Theknockout does not affect amphiphysin IIa transcription, nordoes it affect muscle-specific amphiphysin IIb, suggestingthat dimerization of amphiphysin I and IIa may be importantfor protein stability in the brain. The mechanism for this sta-bility remains unknown at present. Despite the complete ab-sence of amphiphysin I and a severe reduction of amphiphys-in IIa in the brain, the mutant mouse is remarkably viable andreproduces normally. Furthermore, the ultrastructure of thesynapse, including the total number of SVs, appears normal.In particular, there is no accumulation of budding vesiclesalong the plasma membrane, which is the hallmark for de-fects revealed in the amphiphysin-dominant negative experi-ments. These studies indicate that synaptic vesicle endo-cytosis is largely normal in the knockout mouse.
Detailed electrophysiological analysis shows that knockout ofamphiphysin I results in slight reductions in the kinetics ofvesicle endocytosis, vesicle translocation, and/or priming. Inaddition, the association of clathrin and AP-2 with the mem-brane is reduced, confirming a role for the central CLAP do-main in recruiting clathrin and AP-2 to the plasma membrane(33). Unexpectedly, the interaction of dynamin with AP-2 andclathrin is impaired by the deletion of amphiphysin I in themutant mouse, suggesting the amount of dynamin availablefor the fission step is reduced. All these factors may accountfor the mild defect in SV endocytosis in the knockout animal.The rather mild phenotype could also result from redundantfunctions of amphiphysin IIa. However, amphiphysin IIa is notusually present in presynaptic terminals (24), arguing againsta redundant role for it in synaptic function. Furthermore, SVsdo not seem to be recycled through an alternative, clathrin-independent, ‘kiss and run’ mechanism because SVs can bestained and destained quite well with FM dyes (56,57).Therefore, it is reasonable to conclude that amphiphysins donot play a key role in clathrin-mediated endocytosis of SVs;rather, they fine-tune the kinetics of this process.
Behavioral analysis of amphiphysin I knockout mice revealednovel roles for amphiphysin I and IIa in seizures, learning, andmemory. Knockout mice have a lower survival rate and exhibitsudden death due to seizures. Most interestingly, the knock-out mice display both spatial and fear-based learning andmemory defects. These behavioral and learning deficits maynot be directly linked to the minor defect in SV recycling. Theseizure observed in the knockout animal, for example, maybe related to an impairment of action potential propagation,as amphiphysin IIa is located near the spike initiation zoneand the node of Ranvier. However, changes in axonal mor-phology and action potential propagation have not beenexamined in these mice.
Unlike yeast and mammals, Drosophila has only one geneencoding an amphiphysin family member that, via alternative
456 Traffic 2002; 3: 452–460
splicing, produces four isoforms. Alternative splicing only af-fects the nonconserved central domain; each isoform still hasthe conserved N-terminal BAR domain and the C-terminalSH3 domain. The unifying theme of three recent reports isthat dAmph is not involved in synaptic vesicle endocytosis atthe neuromuscular junction (29–31). First, as mentionedearlier, the central domain of dAmph lacks both the PRD andthe CLAP domain for interactions with endophilin, clathrin,and AP-2. Second, the SH3 domain of dAmph does not in-teract with Drosophila dynamin, even though it is capable ofinteracting with vertebrate dynamin, presumably due to theabsence of the amphiphysin SH3 binding motif (PxRPxR) onDrosophila dynamin. Third, the distribution of dAmph islimited to the postsynaptic side of type I boutons at the larvalneuromuscular junction (Figure 2D). Finally, damph mutantsare viable and show no sign of endocytic defects at the NMJin both single and prolonged stimulation protocols (Figure2E). dAmph is expressed in the CNS but its role in SV recyc-ling there has not been examined. However, there is little rea-son to believe that it plays a major role in SV recycling inthe brain, given that dAmph does not interact with knownendocytic proteins. In sum, these genetic studies in bothmice and fruit flies do not support a significant role for am-phiphysins in SV, and perhaps receptor endocytosis.
Amphiphysin Flexes Muscles
If dAmph does not play a major role in endocytosis, whatother functions does it have? Some of the answers to thisquestion were revealed by further analysis of dAmph mutantflies. One curious observation of dAmph mutants is that al-though they are viable and reproduce normally, larvae exhibitsluggish locomotion and adult flies are flightless. These be-havioral dysfunctions appear to be caused by a dramatic de-fect in the muscle transverse (T) tubules. Razzaq et al. (2001)demonstrated that not only does dAmph localize to the NMJsynapse but also to T-tubules, specialized invaginations ofthe plasma membrane that aid in the rapid propagation ofthe action potential (58). In the absence of dAmph, the T-tubules are altered in size and severely disorganized [see Fig-ure 5 in (31,58)]. The mutant T-tubules and flightlessness canbe rescued by expressing wild-type dAmph back intomuscles (31). Therefore, the locomotive defects in amph mu-tants can be attributed to the defective organization of the T-tubules, essential machinery for the coordinated contractionof muscle fibers. Interestingly, amphiphysin IIb has a similarpattern of expression in rat skeletal muscles. Amphiphysin IIbis colocalized with the actin cytoskeleton protein ankyrin3 inT-tubules (24). In addition, clathrin is also present but it doesnot overlap with amphiphysin IIb. These differences in thesubcellular localization of amphiphysin IIb and clathrin as wellas the biochemical property of dAmph raise doubts about thepossible involvement of amphiphysin IIb/dAmph in coatedvesicle fission in the muscle T-tubules.
At least four mechanisms by which amphiphysins may play arole in the T-tubule organization and function are summarized
Genetic Dissection of the Function of Amphiphysins
here. Razzaq and colleagues (2001) proposed that the BARdomain of dAmph, which can induce tubule formations fromliposomes similar to vertebrate amphiphysins (31,45), mayparticipate in the formation of T-tubules. Further, dAmph mayfunction together with Disc Large (DLG), a PDZ family proteinknown to function as a scaffold protein that organizes ionchannels and cell-adhesion proteins in postsynaptic density(59,60), to organize the protein matrix on T-tubules. Addition-ally, dAmph may regulate the interaction between the actincytoskeleton and the T-tubule, given that the yeast Rvspshave been shown to extensively interact directly with actin(37,38) and the actin-binding protein 1 (Abp 1p), (61). Finally,the T-tubule system arises from either invagination of theplasma membrane and/or the addition of new membrane viaa process similar to exocytosis. Hence, dAmph could also beinvolved in exocytotic vesicle trafficking to target secretoryvesicles to the T-tubule membrane, and thus participate in T-tubule membrane construction or maintenance. A precedentfor such a function in cell membrane integrity has been re-cently demonstrated for the yeast Rvsps (62,63).
Beyond Endocytosis: A Yeast and ActinConnection
The endocytic defects discovered in the yeast Rvsp mutantshave often been taken as supporting evidence for a role of am-phiphysins in clathrin-mediated endocytosis. However, the role
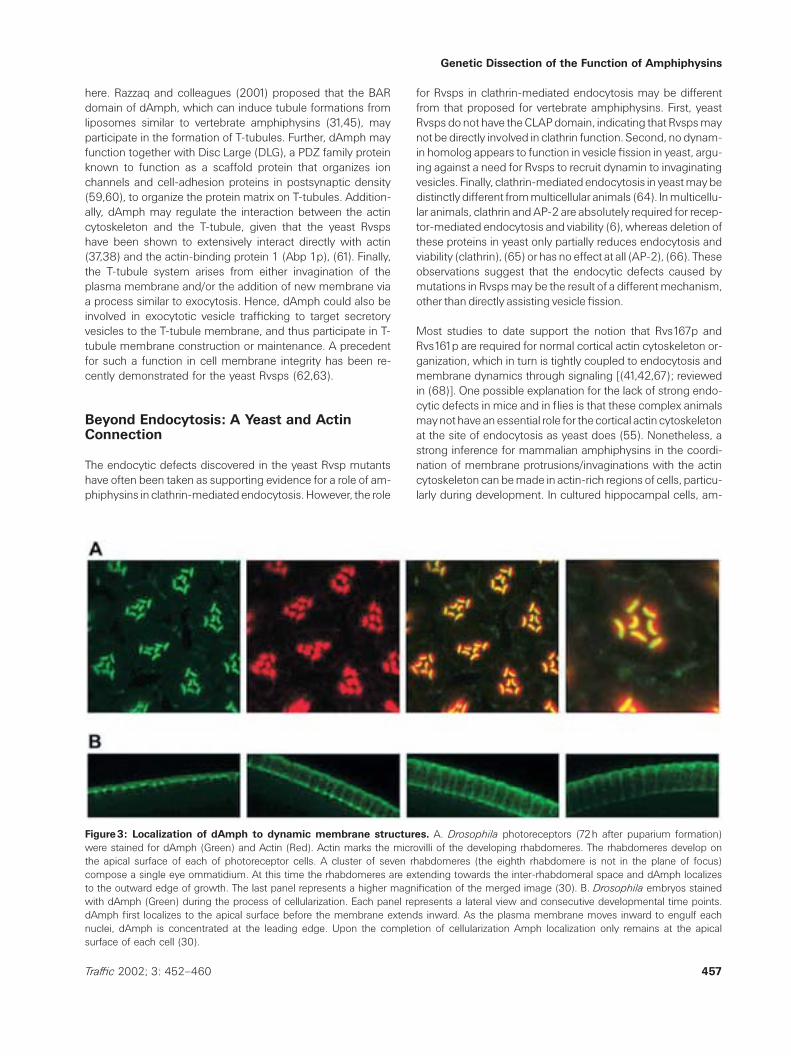
Figure3: Localization of dAmph to dynamic membrane structures. A. Drosophila photoreceptors (72h after puparium formation)were stained for dAmph (Green) and Actin (Red). Actin marks the microvilli of the developing rhabdomeres. The rhabdomeres develop onthe apical surface of each of photoreceptor cells. A cluster of seven rhabdomeres (the eighth rhabdomere is not in the plane of focus)compose a single eye ommatidium. At this time the rhabdomeres are extending towards the inter-rhabdomeral space and dAmph localizesto the outward edge of growth. The last panel represents a higher magnification of the merged image (30). B. Drosophila embryos stainedwith dAmph (Green) during the process of cellularization. Each panel represents a lateral view and consecutive developmental time points.dAmph first localizes to the apical surface before the membrane extends inward. As the plasma membrane moves inward to engulf eachnuclei, dAmph is concentrated at the leading edge. Upon the completion of cellularization Amph localization only remains at the apicalsurface of each cell (30).
457Traffic 2002; 3: 452–460
for Rvsps in clathrin-mediated endocytosis may be differentfrom that proposed for vertebrate amphiphysins. First, yeastRvsps do not have the CLAP domain, indicating that Rvsps maynot be directly involved in clathrin function. Second, no dynam-in homolog appears to function in vesicle fission in yeast, argu-ing against a need for Rvsps to recruit dynamin to invaginatingvesicles. Finally, clathrin-mediated endocytosis in yeast may bedistinctly different from multicellular animals (64). In multicellu-lar animals, clathrin and AP-2 are absolutely required for recep-tor-mediated endocytosis and viability (6), whereas deletion ofthese proteins in yeast only partially reduces endocytosis andviability (clathrin), (65) or has no effect at all (AP-2), (66). Theseobservations suggest that the endocytic defects caused bymutations in Rvsps may be the result of a different mechanism,other than directly assisting vesicle fission.
Most studies to date support the notion that Rvs167p andRvs161p are required for normal cortical actin cytoskeleton or-ganization, which in turn is tightly coupled to endocytosis andmembrane dynamics through signaling [(41,42,67); reviewedin (68)]. One possible explanation for the lack of strong endo-cytic defects in mice and in flies is that these complex animalsmay not have an essential role for the cortical actin cytoskeletonat the site of endocytosis as yeast does (55). Nonetheless, astrong inference for mammalian amphiphysins in the coordi-nation of membrane protrusions/invaginations with the actincytoskeleton can be made in actin-rich regions of cells, particu-larly during development. In cultured hippocampal cells, am-

Zhang and Zelhof
phiphysin I and F-actin colocalize to growth cones (69). Treat-ment with antisense amphiphysin I oligonucleotides collapsesthe growth cone and disrupts actin organization without affect-ing receptor-mediated endocytosis. This suggests that amphi-physins could have functions independent of clathrin-me-diated endocytosis.
In Drosophila, the spatial and temporal expression of dAmphcorresponds with changes in plasma membrane shape andthe accumulation/organization of actin in cells that actively un-dergo either remodeling or morphogenesis (30). For example,dAmph localizes to the apical surface of photoreceptor cellsprior to the appearance of the actin-based microvilli projec-tions that will give rise to the rhabdomere. Later, dAmph marksthe leading edge of the elongating projections of developingphotoreceptors (Figure 3A); but it is absent subsequently inadult photoreceptor cells. In dAmph mutants, rhabdomeresare more closely packed compared to wild-type, and occasion-ally fused. Overexpression of dAmph results in the delocaliz-ation of endogenous dAmph and actin, leading to either a lossof rhabdomeres or an appearance of ectopic rhabdomeres(30). These studies demonstrate that dAmph plays a regula-tory role in the morphogenesis of rhabdomeres.
In addition, dAmph localization corresponds with the inser-tion of new membrane between daughter cells during em-bryonic cellularization (Figure 3B). Such temporal and spatialrestriction is also seen in the developing trachea, hindgut,and esophagus (30). At the NMJ, type-1 boutons (the onlyNMJ synapse that shows dAmph localization) are also char-acterized by an extensive and highly folded membrane, thesubsynaptic reticulum. In the absence of dAmph, there is aloss of a putative exocytotic and targeting protein, Lethalgiant larvae (Lgl), from the postsynaptic density area, the siteof the subsynaptic reticulum (Figure 2E). Hence, dAmph maybe involved in protein assembly or trafficking during synapsedevelopment. Another potential function of dAmph is to as-sist plasma membrane extension and protrusion during or-ganogenesis and synaptogenesis through dynamic reorgan-ization of the actin cytoskeleton. This would require dAmpheither to directly deform membranes or to act as a signalingmolecule to coordinate other proteins to change the mem-brane. dAmph can tubulate lipid membranes but has notbeen studied in cell signaling (31). However, increasing evi-dence supports the signaling hypothesis. Yeast Rvsps andvertebrate amphiphysins have been shown to interact withvarious actin-activation kinases, including nonreceptor tyro-sine kinase abelson (70) and the cyclin-dependent kinase(cdk) 5 (71–73). Phosphorylation of amphiphysin by thesekinases may play an important role in organization of actincytoskeleton during cell development.
EmBARking on a New Journey
Recent genetic studies reveal that:
1. Amphiphysins do not have a conserved function in SV re-
458 Traffic 2002; 3: 452–460
cycling. In vertebrates, amphiphysins may regulate, but arenot essential for clathrin-mediated endocytosis of SVs.However, in Drosophila amphiphysin is not involved at allin SV endocytosis;
2. Amphiphysins are implicated in seizures, learning, andmemory in the mouse;
3. Amphiphysin is required for T-tubule structure and exci-tation-contraction coupling in Drosophila muscles; and
4. Amphiphysin plays a role in membrane morphogenesis indeveloping photoreceptors and a variety of other cells.
These studies have extended observations in yeast and sig-nificantly advanced our knowledge of the biological functionsof amphiphysins in higher animals.
These studies have also raised more questions to be ad-dressed. Does amphiphysin IIa have redundant functions inSV recycling or are there developmental compensations forthe absence of amphiphysin I? What roles do amphiphysinsplay in learning and memory? What are the conserved func-tions of the BAR domain present in all amphiphysin familymembers? If the SH3 domain in both Drosophila and yeastdoes not bind dynamin, what other proteins may it interactwith? Would it also interact with Abp1 in Drosophila as itdoes in yeast? Such an interaction would suggest an import-ant tie for amphiphysins in the actin cytoskeleton dynamics.Therefore, elucidating the mechanisms by which amphiphys-ins interact with or signal to actin will likely become the majortask for cell biologists. We anticipate that genetic approachesin the mouse, Drosophila, and other model organisms willplay an even more prominent role in the new journey of dis-covering the mysteries of amphiphysins.
Acknowledgments
A.C.Z. is supported by an NIH NSRA Postdoctoral Fellowship (DC00432-02). Research in the laboratory of B.Z. is supported by a start-up fund fromthe University of Texas and by an NSF Career Award (IBN-0093170). B.Z.thanks Janice Fischer for stimulating discussions on eye development, andTerry O’Halloran, Wesley Thompson, Hong Bao, Richard Daniels and Ming-shan Xue for constructive comments. We apologize to our colleagueswhose contributions were not cited due to space limitations.
References
1. Bittner GD, Kennedy D. Quantitative aspects of transmitter release. JCell Biol 1970;47:585–592.
2. Clark AW, Hurlbut WP, Mauro A. Changes in the fine structure of theneuromuscular junction of the frog caused by black widow spidervenom. J Cell Biol 1972;52:1–514.
3. Ceccarelli B, Hurlbut WP, Mauro A. Depletion of vesicles from frogneuromuscular junctions by prolonged tetanic stimulation. J Cell Biol1972;54:30–38.
4. Heuser JE, Reese TS. Evidence for recycling of synaptic vesicle mem-brane during transmitter release at the frog neuromuscular junction. JCell Biol 1973;57:315–344.

Genetic Dissection of the Function of Amphiphysins
5. Kirchhausen T, Clathrin. Annu Rev Biochem 2000;69:699–727.6. Brodsky FM, Chen CY, Knuehl C, Towler MC, Wakeham DE. Biological
basket weaving: Formation and function of clathrin-coated vesicles.Annu Rev Cell Dev Biol 2001;17:517–568.
7. Zhang B, Ramaswami M. Synaptic vesicle endocytosis and recycling.In: Bellen H J, eds. Frontiers in Molecular Biology: NeurotransmitterRelease. London: Oxford University Press; 1999. pp. 389–431.
8. Brodin L, Low P, Shupliakov O. Sequential steps in clathrin-mediatedsynaptic vesicle endocytosis. Curr Opin Neurobiol 2000;10:312–320.
9. Grigliatti TA, Hall L, Rosenbluth R, Suzuki DT. Temperature-sensitivemutations in Drosophila melanogaster. XIV. A selection of immobileadults. Mol Gen Genet 1973;120:107–114.
10. Poodry CA, Edgar L. Reversible alteration in the neuromuscular junc-tions of Drosophila melanogaster bearing a temperature-sensitivemutation, shibire. J Cell Biol 1979;81:520–527.
11. van der Bliek AM, Meyerowitz EM. Dynamin-like protein encoded bythe Drosophila shibire gene associated with vesicular traffic. Nature1991;351:411–414.
12. Chen MS, Obar RA, Schroeder CC, Austin TW, Poodry CA, WadsworthSC, Vallee RB. Multiple forms of dynamin are encoded by shibire, aDrosophila gene involved in endocytosis. Nature 1991;351:583–586.
13. Kosaka T, Ikeda K. Possible temperature-dependent blockage of syn-aptic vesicle recycling induced by a single gene mutation in Droso-
phila. J Neurobiol 1983;14:207–225.14. Koenig JH, Ikeda K. Disappearance and reformation of synaptic ves-
icle membrane upon transmitter release observed under reversibleblockage of membrane retrieval. J Neurosci 1989;9:3844–3860.
15. Damke H, Baba T, Warnock DE, Schmid SL. Induction of mutant dyna-min specifically blocks endocytic coated vesicle formation. J Cell Biol1994;127:915–934.
16. Hinshaw JE, Schmid SL. Dynamin self-assembles into rings sug-gesting a mechanism for coated vesicle budding. Nature 1995;374:190–192.
17. Takei K, McPherson PS, Schmid SL, De Camilli P. Tubular membraneinvaginations coated by dynamin rings are induced by GTP-gamma Sin nerve terminals. Nature 1995;374:186–190.
18. Sweitzer SM, Hinshaw JE. Dynamin undergoes a GTP-dependentconformational change causing vesiculation. Cell 1998;93:1021–1029.
19. Sever S, Damke H, Schmid SL. Garrotes, springs, ratchets, and whips:Putting dynamin models to the test. Traffic 2000;1:385–392.
20. Lichte B, Veh RW, Meyer HE, Kilimann MW. Amphiphysin, a novelprotein associated with synaptic vesicles. EMBO J 1992;11:2521–2530.
21. De Camilli P, Thomas A, Cofiell R, Folli F, Lichte B, Piccolo G, MeinckHM, Austoni M, Fassetta G, Bottazzo G, Bates D, Cartlidge N, Soli-mena M, Kiliman MW. The synaptic vesicle-associated protein amphi-physin is the 128-kD autoantigen of Stiff-Man syndrome with breastcancer. J Exp Med 1993;178:2219–2223.
22. Folli F, Solimena M, Cofiell R, Austoni M, Tallini G, Fassetta G, BatesD, Cartlidge N, Bottazzo GF, Piccolo G. Autoantibodies to a 128-kdsynaptic protein in three women with the stiff-man syndrome andbreast cancer. N Engl J Med 1993;328:546–551.
23. David C, McPherson PS, Mundigl O, de Camilli P. A role of amphiphys-in in synaptic vesicle endocytosis suggested by its binding to dynaminin nerve terminals. Proc Natl Acad Sci USA 1996;93:331–335.
24. Butler MH, David C, Ochoa GC, Freyberg Z, Daniell L, Grabs D, Cre-mona O, De Camilli P. Amphiphysin II (SH3P:9; BIN1), a member ofthe amphiphysin/Rvs family, is concentrated in the cortical cytomatrixof axon initial segments and nodes of ranvier in brain and around Ttubules in skeletal muscle. J Cell Biol 1997;137:1355–1367.
25. Ramjaun AR, Micheva KD, Bouchelet I, McPherson PS. Identificationand characterization of a nerve terminal-enriched amphiphysin iso-form. J Biol Chem 1997;272:16700–16706.
459Traffic 2002; 3: 452–460
26. Sakamuro D, Elliott KJ, Wechsler-Reya R, Prendergast GC. BIN1 is anovel MYC-interacting protein with features of a tumour suppressor.Nat Genet 1996;14:69–77.
27. Tsutsui K, Maeda Y, Tsutsui K, Seki S, Tokunaga A. cDNA cloningof a novel amphiphysin isoform and tissue-specific expression of itsmultiple splice variants. Biochem Biophys Res Commun 1997;236:178–183.
28. Wechsler-Reya R, Sakamuro D, Zhang J, Duhadaway J, PrendergastGC. Structural analysis of the human BIN1 gene. Evidence for tissue-specific transcriptional regulation and alternate RNA splicing. J BiolChem 1997;272:31453–31458.
29. Leventis PA, Chow BM, Stewart BA, Iyengar B, Campos AR, BoulianneGL. Drosophila amphiphysin is a post-synaptic protein required fornormal locomotion but not endocytosis. Traffic 2001;2:839–850.
30. Zelhof AC, Bao H, Hardy RW, Razzaq A, Zhang B, Doe CQ. Drosophila
amphiphysin is implicated in protein localization and membrane mor-phogenesis but not in synaptic vesicle endocytosis. Development2001;128:5005–5015.
31. Razzaq A, Robinson IM, McMahon HT, Skepper JN, Su Y, Zelhof AC,Jackson AP, Gay NJ, O’Kane CJ. Amphiphysin is necessary for organ-ization of the excitation–contraction coupling machinery of muscles,but not for synaptic vesicle endocytosis in Drosophila. Genes Dev2001;15:2967–2979.
32. Ge K, Prendergast GC. Bin2, a functionally nonredundant member ofthe BAR adaptor gene family. Genomics 2000;67:210–220.
33. Ramjaun AR, Philie J, de Heuvel E, McPherson PS. The N terminus ofamphiphysin II mediates dimerization and plasma membrane target-ing. J Biol Chem 1999;274:19785–19791.
34. Micheva KD, Ramjaun AR, Kay BK, McPherson PS. SH3 domain-de-pendent interactions of endophilin with amphiphysin. FEBS Lett1997;414:308–312.
35. Grabs D, Slepnev VI, Songyang Z, David C, Lynch M, Cantley LC, DeCamilli P. The SH3 domain of amphiphysin binds the proline-rich do-main of dynamin at a single site that defines a new SH3 bindingconsensus sequence. J Biol Chem 1997;272:13419–13425.
36. Ringstad N, Nemoto Y, De Camilli P. The SH3p4/Sh3p8/SH3p13 pro-tein family. binding partners for synaptojanin and dynamin via a Grb2-like Src homology 3 domain. Proc Natl Acad Sci USA 1997;94:8569–8574.
37. Munn AL, Stevenson BJ, Geli MI, Riezman H. end5, end6, and end7:Mutations that cause actin delocalization and block the internalizationstep of endocytosis in Saccharomyces cerevisiae. Mol Biol Cell1995;6:1721–1742.
38. Amberg DC, Basart E, Botstein D. Defining protein interactions withyeast actin in vivo. Nat Struct Biol 1995;2:28–35.
39. McMahon HT, Wigge P, Smith C. Clathrin interacts specifically withamphiphysin and is displaced by dynamin. FEBS Lett 1997;413:319–322.
40. Slepnev VI, Ochoa GC, Butler MH, De Camilli P. Tandem arrangementof the clathrin and AP-2 binding domains in amphiphysin 1 and dis-ruption of clathrin coat function by amphiphysin fragments compris-ing these sites. J Biol Chem 2000;275:17583–17589.
41. Crouzet M, Urdaci M, Dulau L, Aigle M. Yeast mutant affected forviability upon nutrient starvation. characterization and cloning of theRVS161 gene. Yeast 1991;7:727–743.
42. Bauer F, Urdaci M, Aigle M, Crouzet M. Alteration of a yeast SH3protein leads to conditional viability with defects in cytoskeletal andbudding patterns. Mol Cell Biol 1993;13:5070–5084.
43. Wigge P, McMahon HT. The amphiphysin family of proteins and theirrole in endocytosis at the synapse. Trends Neurosci 1998;21:339–344.
44. Bauerfeind R, Takei K, De Camilli P. Amphiphysin I is associated withcoated endocytic intermediates and undergoes stimulation-depend-ent dephosphorylation in nerve terminals. J Biol Chem 1997;272:30984–30992.

Zhang and Zelhof
45. Takei K, Slepnev VI, Haucke V, De Camilli P. Functional partnershipbetween amphiphysin and dynamin in clathrin-mediated endocytosis.Nat Cell Biol 1999;1:33–39.
46. Shupliakov O, Low P, Grabs D, Gad H, Chen H, David C, Takei K, DeCamilli P, Brodin L. Synaptic vesicle endocytosis impaired by disrup-tion of dynamin–SH3 domain interactions. Science 1997;276:259–263.
47. Shpetner HS, Herskovits JS, Vallee RB. A binding site for SH3 do-mains targets dynamin to coated pits. J Biol Chem 1996;271:13–16.
48. Wigge P, Vallis Y, McMahon HT. Inhibition of receptor-mediated endo-cytosis by the amphiphysin SH3 domain. Curr Biol 1997;7:554–560.
49. Volchuk A, Narine S, Foster LJ, Grabs D, De Camilli P, Klip A. Pertur-bation of dynamin II with an amphiphysin SH3 domain increasesGLUT4 glucose transporters at the plasma membrane in 3T3-L1 ad-ipocytes. Dynamin II participates in GLUT4 endocytosis. J Biol Chem1998;273:8169–8176.
50. Slepnev VI, Ochoa GC, Butler MH, Grabs D, Camilli PD. Role of phos-phorylation in regulation of the assembly of endocytic coat complex-es. Science 1998;281:821–824.
51. Owen DJ, Wigge P, Vallis Y, Moore JD, Evans PR, McMahon HT. Crys-tal structure of the amphiphysin-2 SH3 domain and its role in theprevention of dynamin ring formation. EMBO J 1998;17:5273–5285.
52. Roos J, Kelly RB. Dap160, a neural-specific Eps15 homology andmultiple SH3 domain-containing protein that interacts with Droso-
phila dynamin. J Biol Chem 1998;273:19108–19119.53. Qualmann B, Kelly RB. Syndapin isoforms participate in receptor-me-
diated endocytosis and actin organization. J Cell Biol 2000;148:1047–1062.
54. Kessels MM, Engqvist-Goldstein AE, Drubin DG, Qualmann B. Mam-malian Abp1, a signal-responsive F-actin-binding protein, links the ac-tin cytoskeleton to endocytosis via the GTPase dynamin. J Cell Biol2001;153:351–366.
55. Qualmann B, Kessels MM, Kelly RB. Molecular links between endo-cytosis and the actin cytoskeleton. J Cell Biol 2000;150:F111–F116.
56. Di Paolo G, Sankaranarayanan S, Wenk MR, Daniell L, Perucco E, Cald-arone BJ, Flavell R, Picciotto MR, Ryan TA, Cremona O, De Camilli P.Decreased synaptic vesicle recycling efficiency and cognitive deficitsin amphiphysin 1 knockout mice. Neuron 2002;33:789–804.
57. Ryan TA, Reuter H, Smith SJ. Optical detection of a quantal presynap-tic membrane turnover. Nature 1997;388:478–482.
58. Flucher BE. Structural analysis of muscle development: Transverse tu-bules, sarcoplasmic reticulum, and the triad. Dev Biol 1992;154:245–260.
59. Woods DF, Bryant PJ. The discs-large tumor suppressor gene ofDrosophila encodes a guanylate kinase homolog localized at septatejunctions. Cell 1991;66:451–464.
460 Traffic 2002; 3: 452–460
60. Koh YH, Gramates LS, Budnik V. Drosophila larval neuromuscularjunction: molecular components and mechanisms underlying synap-tic plasticity. Microsc Res Techn 2000;49:14–25.
61. Lila T, Drubin DG. Evidence for physical and functional interactionsamong two Saccharomyces cerevisiae SH3 domain proteins, an ad-enylyl cyclase-associated protein and the actin cytoskeleton. Mol BiolCell 1997;8:367–385.
62. Bon E, Recordon-Navarro P, Durrens P, Iwase M, Toh EA, Aigle M. Anetwork of proteins around Rvs167p and Rvs161p, two proteins re-lated to the yeast actin cytoskeleton. Yeast 2000;16:1229–1241.
63. Breton AM, Schaeffer J, Aigle M. The yeast Rvs161 and Rvs167 pro-teins are involved in secretory vesicles targeting the plasma mem-brane and in cell integrity. Yeast 2001;18:1053–1068.
64. Baggett JJ, Wendland B. Clathrin function in yeast endocytosis. Traffic2001;2:297–302.
65. Payne GS, Schekman R. A test of clathrin function in protein secretionand cell growth. Science 1985;230:1009–1014.
66. Huang KM, D’Hondt K, Riezman H, Lemmon SK. Clathrin functions inthe absence of heterotetrameric adaptors and AP180-related proteinsin yeast. EMBO J 1999;18:3897–3908.
67. Lombardi R, Riezman H. Rvs161p and Rvs167p, the two yeast amphi-physin homologs, function together in vivo. J Biol Chem 2001;276:6016–6022.
68. Schafer DA. Coupling actin dynamics and membrane dynamics dur-ing endocytosis. Curr Opin Cell Biol 2002;14:76–81.
69. Mundigl O, Ochoa GC, David C, Slepnev VI, Kabanov A, De Camilli P.Amphiphysin I antisense oligonucleotides inhibit neurite outgrowth incultured hippocampal neurons. J Neurosci 1998;18:93–103.
70. Kadlec L, Pendergast AM. The amphiphysin-like protein 1 (ALP1)interacts functionally with the cABL tyrosine kinase and may play arole in cytoskeletal regulation. Proc Natl Acad Sci USA 1997;94:12390–12395.
71. Lee J, Colwill K, Aneliunas V, Tennyson C, Moore L, Ho Y, AndrewsB. Interaction of yeast Rvs167 and Pho85 cyclin-dependent kinasecomplexes may link the cell cycle to the actin cytoskeleton. Curr Biol1998;8:1310–1321.
72. Rosales JL, Nodwell MJ, Johnston RN, Lee KY. Cdk5/p25 (nck5a)interaction with synaptic proteins in bovine brain. J Cell Biochem2000;78:151–159.
73. Floyd SR, Porro EB, Slepnev VI, Ochoa GC, Tsai LH, De Camilli P. Am-phiphysin 1 binds the cyclin-dependent kinase (cdk) 5 regulatory sub-unit p35 and is phosphorylated by cdk5 and cdc2. J Biol Chem 20011970;276:8104–8110.





























