Embed Size (px)
Citation preview

Amino Acid Production by Corynebacteriumglutamicum
Masato Ikeda and Seiki Takeno
Contents
1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 108
2 Amino Acid Fermentation Processes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112
3 Overall Strategies for Strain Improvement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 113
3.1 Terminal Pathways . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 114
3.2 Central Metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 115
3.3 NADPH Regeneration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 116
3.4 Amino Acid Transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
3.5 Respiratory Energy Efficiency . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 118
3.6 Global Regulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 118
3.7 Stress Responses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
3.8 Feedstock Utilization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
4 Recent Advances in Amino Acid Production . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 120
4.1 Glutamate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121
4.2 Lysine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
4.3 Arginine and Citrulline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
4.4 Branched-Chain Amino Acids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 128
4.5 Alanine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
4.6 Serine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 130
4.7 Methionine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 132
4.8 Cysteine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 134
5 Conclusions and Outlook . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 136
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 136
Abstract During the half century following its discovery, the L-glutamate-producing
microorganism Corynebacterium glutamicum has played a leading role in the amino
acid fermentation industry. Due to its importance as an amino acid producer,
M. Ikeda (*) • S. Takeno
Faculty of Agriculture, Department of Bioscience and Biotechnology, Shinshu University,
8304 Minami-minoma, Nagano 399-4598, Japan
e-mail: [email protected]
H. Yukawa and M. Inui (eds.), Corynebacterium glutamicum,Microbiology Monographs 23, DOI 10.1007/978-3-642-29857-8_4,# Springer-Verlag Berlin Heidelberg 2013
107

C. glutamicum is also one of the best-investigated microorganisms, evidenced by the
extensive body of relevant literature and patents. In the past quarter century, various
genetic engineering tools and global analysis techniques for this bacterium have been
developed and successfully applied, giving a thorough understanding of its physiology
and permitting the development of efficient production strains. The advances enhanc-
ing the usefulness of this bacterium for amino acid production over the last decade can
be summarized in five points: (1)Metabolic engineering strategies are expanding from
the core biosynthetic pathways to include central metabolism, cofactor-regeneration
systems, uptake and export systems, energy metabolism, global regulation, and stress
responses; strain improvement is bound to thereby optimize entire cellular systems.
(2) Systems biology for this bacterium is almost capable of predicting targets to be
engineered and metabolic states that will yield maximum production; these
developments should allow rational metabolic design. (3) Rapid strides in genome
analysis have revolutionized strain improvement methodology, allowing
reengineering of more efficient producers through knowledge of the mutations that
have accumulated over years of industrial strain development. (4) The spectra of both
products and assimilable carbon sources of this bacterium have expanded, leading to
the development of, e.g., production strains of serine and methionine that could not be
produced effectively from glucose and strains that can utilize alternative feedstocks
that do not compete with human food or energy sources. (5) Recent identification of a
putative mechanosensitive channel as a possible glutamate exporter has provided
valuable insight into the glutamate production mechanism which had long been the
central question concerning the industrial biotechnology of C. glutamicum. Thischapter describes advances in the production of amino acids by C. glutamicum, withspecial focus on the technology and strategies for molecular strain improvement.
1 Introduction
Amino acids have a wide variety of characteristics in terms of nutritional value,
taste, medicinal action, and chemical properties, and thus have many potential uses,
e.g., in food additives, feed supplements, pharmaceuticals, cosmetics, polymer
materials, and agricultural chemicals. As each new use is developed, demand for
that type of amino acid grows rapidly and is followed by the development of mass
production technology for that amino acid. The annual world production of amino
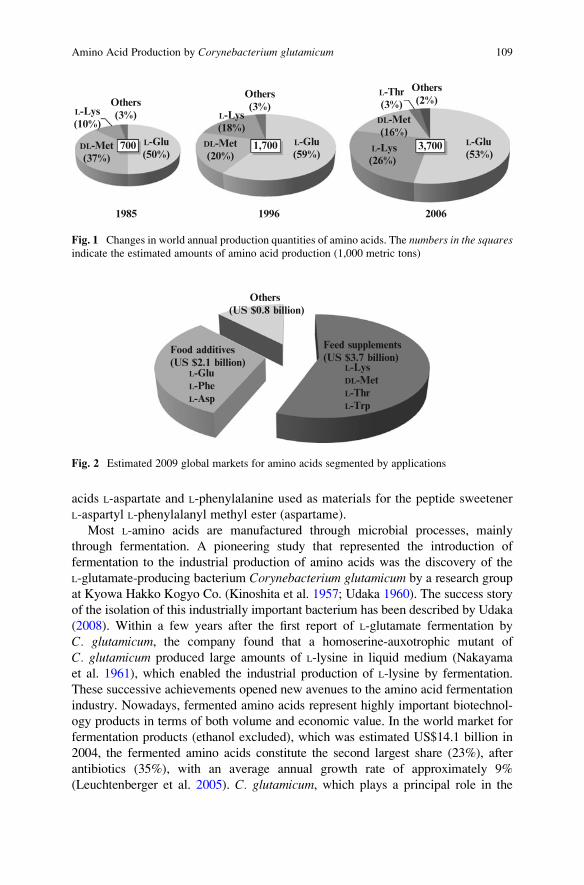
acids has increased year by year (Fig. 1) and is currently estimated at more than 3.7
million metric tons (Ikeda 2003; Hermann 2003; Ajinomoto 2007). According to a
recent market research report (McWilliams 2010) and other relevant publications
(Leuchtenberger et al. 2005; Ajinomoto 2007), the global market for amino acids is
estimated to be approximately US$6.6 billion in 2009 and is growing at an annual
rate of 8–10%. Figure 2 shows the estimated global markets for amino acids of
different applications in 2009. So-called feed amino acids L-lysine, DL-methionine,
L-threonine, and L-tryptophan have the largest share of the market, generating US
$3.7 billion in 2009. The second largest share belongs to food additives, which are
comprised mainly of the flavor-enhancer monosodium glutamate and the amino
108 M. Ikeda and S. Takeno
acids L-aspartate and L-phenylalanine used as materials for the peptide sweetener
L-aspartyl L-phenylalanyl methyl ester (aspartame).
Most L-amino acids are manufactured through microbial processes, mainly
through fermentation. A pioneering study that represented the introduction of
fermentation to the industrial production of amino acids was the discovery of the
L-glutamate-producing bacterium Corynebacterium glutamicum by a research group
at Kyowa Hakko Kogyo Co. (Kinoshita et al. 1957; Udaka 1960). The success story
of the isolation of this industrially important bacterium has been described by Udaka
(2008). Within a few years after the first report of L-glutamate fermentation by
C. glutamicum, the company found that a homoserine-auxotrophic mutant of
C. glutamicum produced large amounts of L-lysine in liquid medium (Nakayama
et al. 1961), which enabled the industrial production of L-lysine by fermentation.
These successive achievements opened new avenues to the amino acid fermentation
industry. Nowadays, fermented amino acids represent highly important biotechnol-
ogy products in terms of both volume and economic value. In the world market for
fermentation products (ethanol excluded), which was estimated US$14.1 billion in
2004, the fermented amino acids constitute the second largest share (23%), after
antibiotics (35%), with an average annual growth rate of approximately 9%
(Leuchtenberger et al. 2005). C. glutamicum, which plays a principal role in the
1985
L-Glu(50%)
DL-Met(37%)
L-Lys(10%)
Others(3%)
700 L-Glu(59%)
L-Lys(18%)
DL-Met(20%)
Others(3%)
1996
1,700
2006
L-Glu(53%)
L-Lys(26%)
DL-Met(16%)
Others(2%)
L-Thr(3%)
3,700
Fig. 1 Changes in world annual production quantities of amino acids. The numbers in the squaresindicate the estimated amounts of amino acid production (1,000 metric tons)
Feed supplements(US $3.7 billion)
L-LysDL-MetL-ThrL-Trp
Food additives(US $2.1 billion)
L-GluL-PheL-Asp
Others (US $0.8 billion)
Fig. 2 Estimated 2009 global markets for amino acids segmented by applications
Amino Acid Production by Corynebacterium glutamicum 109
process of amino acid fermentation, is therefore also highly important, as
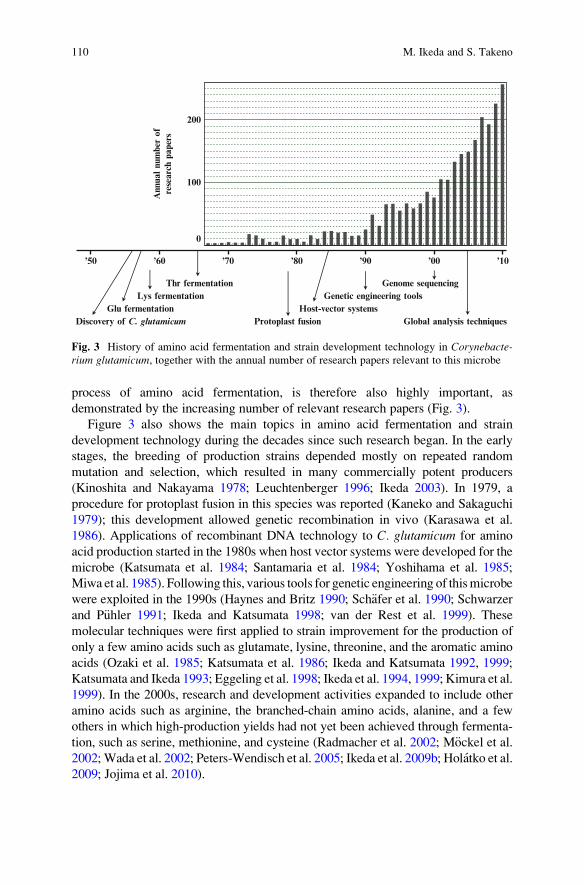
demonstrated by the increasing number of relevant research papers (Fig. 3).
Figure 3 also shows the main topics in amino acid fermentation and strain
development technology during the decades since such research began. In the early
stages, the breeding of production strains depended mostly on repeated random
mutation and selection, which resulted in many commercially potent producers
(Kinoshita and Nakayama 1978; Leuchtenberger 1996; Ikeda 2003). In 1979, a
procedure for protoplast fusion in this species was reported (Kaneko and Sakaguchi
1979); this development allowed genetic recombination in vivo (Karasawa et al.
1986). Applications of recombinant DNA technology to C. glutamicum for amino
acid production started in the 1980s when host vector systems were developed for the
microbe (Katsumata et al. 1984; Santamaria et al. 1984; Yoshihama et al. 1985;
Miwa et al. 1985). Following this, various tools for genetic engineering of thismicrobe
were exploited in the 1990s (Haynes and Britz 1990; Sch€afer et al. 1990; Schwarzerand P€uhler 1991; Ikeda and Katsumata 1998; van der Rest et al. 1999). These
molecular techniques were first applied to strain improvement for the production of
only a few amino acids such as glutamate, lysine, threonine, and the aromatic amino
acids (Ozaki et al. 1985; Katsumata et al. 1986; Ikeda and Katsumata 1992, 1999;
Katsumata and Ikeda 1993; Eggeling et al. 1998; Ikeda et al. 1994, 1999; Kimura et al.
1999). In the 2000s, research and development activities expanded to include other
amino acids such as arginine, the branched-chain amino acids, alanine, and a few
others in which high-production yields had not yet been achieved through fermenta-
tion, such as serine, methionine, and cysteine (Radmacher et al. 2002; M€ockel et al.2002;Wada et al. 2002; Peters-Wendisch et al. 2005; Ikeda et al. 2009b; Holatko et al.
2009; Jojima et al. 2010).
’50 ’60 ’70 ’80 ’90 ’00
Discovery of C. glutamicum
Host-vector systems
Genome sequencing
Glu fermentationGenetic engineering tools
Protoplast fusion
Thr fermentationLys fermentation
Global analysis techniques
0
100
200
Ann
ual nu
mbe
r of
rese
arch
pap
ers
’10
Fig. 3 History of amino acid fermentation and strain development technology in Corynebacte-rium glutamicum, together with the annual number of research papers relevant to this microbe
110 M. Ikeda and S. Takeno

Most fermented amino acid production processes currently rely on modified
C. glutamicum and Escherichia coli. Almost all amino acids can be produced
technologically by either organism, but C. glutamicum is used for the industrial
production of relatively large-scale bulk amino acids such as glutamate and
lysine as well as glutamine and arginine, while E. coli is predominantly used for
the production of threonine and the branched-chain amino acids. Although
C. glutamicum has several industrially important characteristics such as its high
growth yield even under conditions of high sugar concentration, it has one draw-
back: its optimal growth temperature is around 30 �C, which is lower than that of
E. coli. For this reason, the use of C. glutamicum may be economically disadvanta-
geous, especially in tropical regions, because of the substantial cost of the utilities
necessary to maintain the optimum fermentation temperature. In an attempt to
address this issue, Corynebacterium efficiens, a related species with an optimal
growth temperature near 40 �C, has been isolated and examined for the relevant
genetic traits and glutamate production ability at higher temperatures (Fudou et al.
2002; Nishio et al. 2003; Kimura 2005). Comparative genome sequence analysis
between C. glutamicum and C. efficiens, as well as amino acid substitution analysis,
has suggested that three types of amino acid substitutions in C. efficiens (K to R,
S to A, and S to T) are important for its thermotolerance (Nishio et al. 2003).
In general, commercially potent producers have been developed by the stepwise
accumulation of beneficial genetic and phenotypic characteristics in one back-
ground through classical mutagenesis and/or recombinant DNA technology. Such
improvements involve strains capable not only of producing amino acids at higher
yields but also of producing lower quantities of by-products, as the removal of
by-products dominates the costs of downstream processing (Ikeda 2003;
Marienhagen and Eggeling 2008). The current production yields toward sugar
(w/w %) can be estimated as follows: lysine hydrochloride, 45–55; glutamate,
45–55; glutamine, 35–45; arginine, 30–40; threonine, 40–50; isoleucine, 20–30;
valine, 30–40; alanine, 45–55; serine, 30–35; methionine, 15–20; tryptophan,
20–25; phenylalanine, 20–25; and histidine, 20–30.
During the last decade, genomic and other “omics” data have accumulated for
C. glutamicum, profoundly affecting strain development methods and providing a
global understanding of this microbe (Wittmann and Heinzle 2002; Ikeda and
Nakagawa 2003; Kalinowski et al. 2003; Strelkov et al. 2004; Yukawa et al.
2007). This work has revealed new regulatory networks and functions that had
not previously been identified in this bacterium. A novel methodology that merges
genomics with classical strain improvement has been developed and used to
rationally reconstruct classically derived production strains (Ikeda et al. 2006,
2009b). In the present chapter, the processes by which amino acids are produced
by C. glutamicum are first briefly described, and strategies for molecular strain
improvement are subsequently briefly summarized and illustrated with relevant
examples. Lastly, recent advances in amino acid production by this microbe are
provided from both basic and applied perspectives. The most common approaches
to strain improvement including conventional mutagenesis and screening have been
Amino Acid Production by Corynebacterium glutamicum 111

omitted because their descriptions can be found in many other publications
(Kinoshita and Nakayama 1978; Leuchtenberger 1996; Ikeda 2003).
2 Amino Acid Fermentation Processes
Fermentation processes typically comprise three steps: cultivation of amino acid-
producing strains, purification of amino acids from fermented broth, and wastewater
treatment. The economy of these processes depends mainly on the cost of the carbon
source, the fermentation yield, purification yield, and the productivity of the overall
process. To date, many technologies have been developed in attempts to establish
economically competitive processes.
Industrial amino acid fermentations are usually performed by means of batch or
fed-batch processes using aerated agitated tank fermentors or airlift tank fermentors
in the 50- to 500-kL size range. Although batch processes are easy to run and do not
require additional tanks for feeding nutrients, industrial processes predominantly
adopt fed-batch, mainly because fed-batch processes provide improved overall
productivity by increasing yields and reducing fermentation periods, especially
when a high or changing concentration of a certain nutrient affects the yield or
productivity of the process. By lowering the initial concentration of sugar and using
subsequent feeding, the total culture period can be shortened, especially the lag
time, and in some cases the yield can also be increased. In a process where
an auxotrophic strain is employed, the yield can be maximized by growing the
auxotrophic strain in a limited amount of the required nutrient through feeding
that nutrient at a controlled rate. An example of this is tryptophan fermentation
by a phenylalanine- and tyrosine-auxotrophic strain of C. glutamicum (Ikeda
and Katsumata 1999). Furthermore, the fed-batch techniques allow substrate
concentrations to be kept low enough to prevent oxygen limitation which causes
a decreased yield with concomitant acid by-production (Ikeda 2003). In these cases,
glucose-limited fed-batch cultures are commonly employed in industrial processes.
To improve overall productivity further, it is possible to extend fed-batch
fermentation by drawing out part of the broth one or more times during the process
and refilling it through nutrient feeding (semi-continuous fermentation), or shifting
the fermentation from batch to continuous culture, where fresh medium containing
all nutrients is fed into a fermentor at a specific rate while the same quantity of broth
with a portion of the microorganisms is continuously taken from the fermentor, thus
maintaining a constant culture volume. Continuous fermentation was investigated
with a C. glutamicum lysine-producing mutant (Hirao et al. 1989); the strain in that
study yielded stable lysine production for more than 300 h, with a maximum lysine
concentration of 105 g/L and a maximum volumetric productivity value of
5.6 g/L/h. Though this strain was able to produce 100 g/L lysine within 48 h in a
fed-batch process, its productivity did not exceed 2.1 g/L/h. This means that the
productivity seen in the continuous fermentation is more than 2.5 times higher than
that seen in the fed-batch culture. The feasibility of the continuous fermentation
112 M. Ikeda and S. Takeno

process is governed chiefly by the genetic stability of a production strain as well as
by the conduciveness of the process to the maintenance of the purity of the culture
over the long term. The former problem has been dealt with in the continuous
fermentation of arginine by C. glutamicum, formally classified as Corynebacteriumacetoacidophilum (Azuma and Nakanishi 1988; Azuma et al. 1988); the original
arginine producer gave rise to mutated variants with reduced productivity during
continuous culture, but one derivative isolated from the continuous culture broth
no longer caused the appearance of such variants, resulting in stable arginine
production for more than 250 h. The derivative strain was found to be less sensitive
to arginine with respect to both growth and production than the original producer,
though the genetic element(s) responsible for the change has not yet been identified.
The production profiles of amino acid fermentation processes can be growth
dependent or growth independent. Lysine and arginine are generally produced by
C. glutamicum in a growth-dependent manner, for example, while glutamate and
glutamine are produced by C. glutamicum in a growth-independent manner.
In growth-independent processes, continuous culture may not afford a higher
productivity than batch or fed-batch processes do because cells are always kept in
the growth phase. The productivity of such fermentation is likely to be improved by
incorporating cell recycling techniques in a fed-batch or continuous fermentation
process (Ishizaki et al. 1993).
Other fundamentals of amino acid fermentation processes, such as fermentation
operations, raw materials, downstream processing, and waste liquor treatment, are
thoroughly described in other publications (Leuchtenberger 1996; Ikeda 2003;
Hermann 2003).
3 Overall Strategies for Strain Improvement
The past quarter century has seen rapid developments in strain development
technology. Metabolic engineering has repeatedly led to successful yield
improvements, especially in the field of amino acid production by C. glutamicum(Eggeling and Bott 2005; Wendisch 2007; Burkovski 2008). In this latter field, the
targets of metabolic engineering have expanded beyond the core biosynthetic
pathways leading to products of interest and now include central metabolism,
cofactor-regeneration systems, uptake and export systems, energy metabolism,
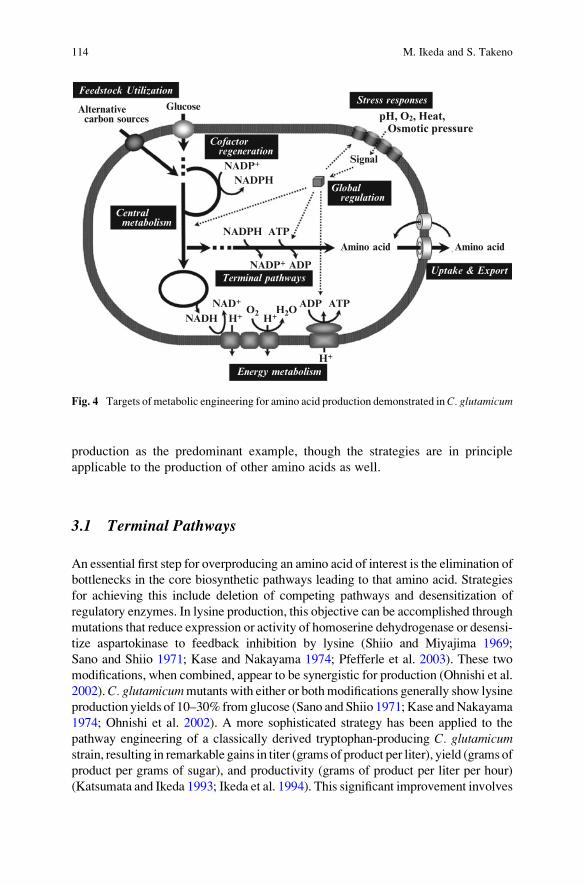
global regulation, and stress responses (Fig. 4). This means that strain development
is beginning to achieve the optimization of entire cellular systems. In addition,
the product spectrum of C. glutamicum also has been expanded, and metabolic
engineering has been applied to the production of amino acids that could not be
produced effectively from glucose, such as serine and methionine. Furthermore,
strains are being engineered with environmental concerns in mind: some utilize
alternative feedstocks, such as whey, lignocellulose-derived xylose and arabinose,
and glycerol, which do not compete with human food or energy sources. The
strategies of rational strain improvement are broadly described here with lysine
Amino Acid Production by Corynebacterium glutamicum 113
production as the predominant example, though the strategies are in principle
applicable to the production of other amino acids as well.
3.1 Terminal Pathways
An essential first step for overproducing an amino acid of interest is the elimination of
bottlenecks in the core biosynthetic pathways leading to that amino acid. Strategies
for achieving this include deletion of competing pathways and desensitization of
regulatory enzymes. In lysine production, this objective can be accomplished through
mutations that reduce expression or activity of homoserine dehydrogenase or desensi-
tize aspartokinase to feedback inhibition by lysine (Shiio and Miyajima 1969;
Sano and Shiio 1971; Kase and Nakayama 1974; Pfefferle et al. 2003). These two
modifications, when combined, appear to be synergistic for production (Ohnishi et al.
2002).C. glutamicummutants with either or bothmodifications generally show lysine
production yields of 10–30% fromglucose (Sano and Shiio 1971;Kase andNakayama
1974; Ohnishi et al. 2002). A more sophisticated strategy has been applied to the
pathway engineering of a classically derived tryptophan-producing C. glutamicumstrain, resulting in remarkable gains in titer (grams of product per liter), yield (grams of
product per grams of sugar), and productivity (grams of product per liter per hour)
(Katsumata and Ikeda 1993; Ikeda et al. 1994). This significant improvement involves
Energy metabolism
Glucose
H2OH+
O2
NADPH
ADP ATP
NADP+
Amino acid
NADPH
NADP+
ATP
ADP
Amino acid
Stress responsesAlternativecarbon sources
Feedstock Utilization
Uptake & Export
Signal
H+NADHNAD+
Centralmetabolism
Cofactorregeneration
Globalregulation
Terminal pathways
H+
Fig. 4 Targets of metabolic engineering for amino acid production demonstrated inC. glutamicum
114 M. Ikeda and S. Takeno

not only systematic genetic modifications that efficiently channel carbon toward
tryptophan via plasmid-mediated amplification of eight genes in all, but also the
construction of a plasmid stabilization system based on the presence of the serine-
biosynthetic gene on the plasmid and that gene’s absence from the chromosome.
3.2 Central Metabolism
Once the core pathway is optimized, further incremental gains can be attained by
increasing precursor supply. This task usually begins with pathway analysis, which
includes laying out all the possible routes from glucose to a desired amino acid,
calculating the theoretical yield of the amino acid from glucose for each route, and
determining the most efficient route including optimal flux distributions at key
branch points. For calculation of the theoretical yield, the energy and redox
balances should be included to balance ATP and the reducing power.
Within the central metabolism, the junction between glycolysis and the tricar-
boxylic acid (TCA) cycle is particularly important for directing metabolic fluxes to
desired biosynthetic pathways, because the junction in C. glutamicum consists of
several enzyme reactions carrying fluxes that connect phosphoenolpyruvate with
oxaloacetate or pyruvate, pyruvate with oxaloacetate, and pyruvate with malate.
Several strategies have proved beneficial in this regard. For example, increased
carbon flux from pyruvate to oxaloacetate by overexpression of the pyruvate
carboxylase gene or by deletion of the phosphoenolpyruvate carboxykinase gene
resulted in significantly increased production of lysine (Petersen et al. 2001; Peters-
Wendisch et al. 2001; Riedel et al. 2001). Increasing the availability of pyruvate by
decreasing or abolishing pyruvate dehydrogenase activity can also improve lysine
production (Shiio et al. 1984; Blombach et al. 2007b). On the other hand, decreased
activity of the TCA cycle enzyme isocitrate dehydrogenase was shown to improve
lysine production, probably by means of a flux shift from the TCA cycle toward
anaplerotic carboxylation (Becker et al. 2009).
Unlike lysine production, glutamate production induced by biotin limitation
depends solely on the phosphoenolpyruvate carboxylase-catalyzed anaplerotic reac-
tion (Sato et al. 2008). In this case, increased carbon flux from phosphoenolpyruvate
to oxaloacetate by overexpression of the phosphoenolpyruvate carboxylase gene
or by deletion of pyruvate kinase gene effectively improves glutamate production
(Sato et al. 2008; Sawada et al. 2010).
Replacement of a phosphoenolpyruvate-dependent sugar phosphotransferase
system (PTS) by a PTS-independent sugar uptake system can be an alternative
strategy for increasing the availability of phosphoenolpyruvate, as has been
demonstrated in aromatic production by E. coli (Flores et al. 1996). InC. glutamicum,the PTS had long been the only known system to uptake glucose, but very recently,
potential glucose uptake systems that function as alternatives to the PTS have been
identified in this microbe (Ikeda et al. 2010, 2011). These include the iolT1 and iolT2gene products, both known as myo-inositol transporters (Krings et al. 2006).
Amino Acid Production by Corynebacterium glutamicum 115

Expression of the iolT1-specific glucose uptake bypass instead of the native PTS
resulted in approximately 20% increased lysine production (Ikeda et al. 2011).
3.3 NADPH Regeneration
Amino acid biosynthesis from sugar typically uses cofactor NADPH as the reducing
power. NADPH supply is therefore an important consideration along with pathway
engineering to direct carbon toward a desired amino acid. Availability of NADPH
is crucial especially for the production of certain amino acids which require large
quantities of NADPH for their biosynthesis, such as lysine, methionine, and arginine.
InC. glutamicum, NADPH supply has been augmented by engineering the redirection
of carbon from glycolysis into the pentose phosphate pathway through methods
such as disruption of the phosphoglucose isomerase gene (Marx et al. 2003),
overexpression of the fructose 1,6-bisphosphatase gene (Becker et al. 2005) or the
glucose 6-phosphate dehydrogenase gene (Becker et al. 2007), and introduction of a
mutant allele of the 6-phosphogluconate dehydrogenase gene encoding an enzyme
that is less sensitive to feedback inhibition (Ohnishi et al. 2005).
The importance of the pentose phosphate pathway for lysine production is
particularly obvious when fructose is used as a carbon source: only 14.4% of
carbon is channeled through the pentose phosphate pathway on fructose, in contrast
to 62.3% when glucose is used (Kiefer et al. 2004). A reason for the lower flux
channeling through the pentose phosphate pathway could be the entry point of
fructose into glycolysis: fructose mostly enters glycolysis at the level of fructose
1,6-bisphosphate which requires gluconeogenetic fructose 1,6-bisphosphatase
activity in order to direct its carbon into the pentose phosphate pathway.
This problem also has some relevance for sucrose, because the fructose unit of
sucrose follows the same metabolic fate as free fructose. To solve this problem,
direct phosphorylation of intracellular fructose produced by sucrose hydrolysis has
been attempted through the heterologous expression of the fructokinase gene from
Clostridium acetobutylicum (Moon et al. 2005). This is expected to shift the
entry point of fructose from fructose 1,6-bisphosphate to its upstream fructose
6-phosphate and thus increase the flux through the pentose phosphate pathway.
In terms of carbon yield, however, supplying carbon through the pentose phos-
phate pathway is less advantageous than supplying it via the glycolytic pathway
because the former pathway inevitably involves the release of 1 mol of carbon
dioxide (CO2) accompanied by the oxidation of 1 mol of hexose. To solve this
dilemma, an attempt was recently made to engineer a functional glycolytic pathway
in C. glutamicum supplying NADPH through a new route. In this study, endogenous
NAD-dependent glyceraldehyde 3-phosphate dehydrogenase of C. glutamicum was
replaced with nonphosphorylating NADP-dependent glyceraldehyde 3-phosphate
dehydrogenase (GapN) of Streptococcus mutans, which catalyzes the irreversible
oxidation of glyceraldehyde 3-phosphate to 3-phosphoglycerate and the cor-
responding reduction of NADP+ to NADPH; the result was a C. glutamicum strain
116 M. Ikeda and S. Takeno

with an NADPH-generating glycolytic pathway. A lysine producer derived from the
engineered strain produced considerably more lysine than the reference strain,
exceeding the reference strain’s production levels by ~70% on glucose, ~120% on
fructose, and ~100% on sucrose (Takeno et al. 2010). As an alternative to this
method, expression of the membrane-bound transhydrogenase genes from E. coli inC. glutamicum provided an alternate source of NADPH (Kabus et al. 2007a, b).
3.4 Amino Acid Transport
For certain amino acids, the biosynthetic pathways are subject to multiple
regulations at several steps. In such cases, it is not easy to completely remove all
regulatory controls existing in the pathways, but this difficulty can largely be
overcome by reducing the intracellular pool of the amino acid to a level at which
feedback control does not operate. Since the intracellular pool of an amino acid is
assumed to depend on the uptake rate of the amino acid accumulated extracellularly
as well as the efflux rate, preventing amino acid re-uptake would serve this purpose.
The impact of such transport engineering on amino acid production was first shown
for tryptophan production by C. glutamicum (Ikeda and Katsumata 1994, 1995).
A modification leading to a decreased rate of tryptophan uptake in a tryptophan-
producing mutant increased production, while plasmid-mediated amplification of
the transporter gene drastically decreased production.
Sometimes the export step is critical for achieving efficient amino acid produc-
tion in C. glutamicum (Morbach et al. 1996; Burkovski and Kr€amer 2002). In such
cases, the intrinsic capacity of this bacterium for excreting a desired amino acid
becomes the barrier to improving productivity. Recently, the discovery of active
export systems for several kinds of amino acids has made it possible to compensate
for the limited capacity of amino acid efflux through recombinant DNA technology.
The exporters so far identified for C. glutamicum are LysE, which exports the basic
amino acids lysine and arginine (Vrljic et al. 1996); ThrE, which exports threonine
and serine (Simic et al. 2001); and BrnFE, which exports the branched chain amino
acids and methionine (Kennerknecht et al. 2002; Tr€otschel et al. 2005).
Overexpression of the lysE gene resulted in a fivefold increase in the excretion
rate for lysine compared to the rate of the control strain (Vrljic et al. 1996). More
recently, the NCgl1221 gene product, a mechanosensitive channel homolog, was
identified as a possible glutamic acid exporter (Nakamura et al. 2007).
The functions of such exporters also can be transferred to heterologous bacterial
species. For example, a mutant allele of the C. glutamicum lysE gene has been
successfully used to improve lysine production in the methylotroph Methylophilusmethylotrophus (Gunji and Yasueda 2006). Likewise, the limited capacity of
C. glutamicum for threonine production was improved not only by overexpression
of the endogenous thrE gene (Simic et al. 2002) but also by heterologous expression
of an E. coli threonine exporter (Diesveld et al. 2008).
Amino Acid Production by Corynebacterium glutamicum 117

3.5 Respiratory Energy Efficiency
Like the redox balance, the energy balance is critical for efficient amino acid
production. For this reason, improving the efficiency of ATP synthesis is another
strategy for increasing amino acid production. In C. glutamicum, two terminal
oxidases are positioned in a branched respiratory chain (Bott and Niebisch 2003).
One branch is composed of the cytochrome bc1-aa3 supercomplex, which has a
threefold higher bioenergetic efficiency than the other cytochrome bd branch.
Disruption of the inefficient cytochrome bd branch caused increased lysine produc-
tion with no marked effect on growth or glucose consumption (Kabus et al. 2007a, b).
Recently, it has been shown that C. glutamicum can grow anaerobically by
means of nitrate respiration (Nishimura et al. 2007; Takeno et al. 2007). In the
presence of nitrate, lysine and arginine production occurred anaerobically, though
at a very low level, indicating the potential of this bacterium for anaerobic amino
acid production (Takeno et al. 2007).
3.6 Global Regulation
The determination of the whole genome sequence of C. glutamicum and the
development of global analysis techniques such as DNA microarray have permitted
the identification of a variety of global regulators (Burkovski 2008). These include
GlxR (Kim et al. 2004), SugR (Engels et al. 2008), RamA (Cramer et al. 2006), and
RamB (Gerstmeir et al. 2004) controlling carbon metabolism, AmtR (Beckers et al.
2005) controlling nitrogen metabolism, PhoR (Schaaf and Bott 2007) controlling
phosphorus metabolism, McbR (Rey et al. 2003, 2005) and SsuR (Koch et al. 2005)
controlling sulfur metabolism, DtxR (Wennerhold and Bott 2006) controlling iron
homeostasis, and FarR (H€anbler et al. 2007) and LtbR (Brune et al. 2007)
controlling amino acid metabolism. The existence of these regulators indicates
that amino acid biosynthesis in C. glutamicum is directly or indirectly subject to
both pathway specific and global regulation (Brockmann-Gretza and Kalinowski
2006; Kr€omer et al. 2008). Thus, global regulation is also important in strain
improvement. Another interesting finding is that the global induction of amino
acid biosynthesis genes occurs in a classically derived industrial lysine-producing
strain of C. glutamicum (Hayashi et al. 2006b): the lysC gene, encoding the key
enzyme aspartokinase, was up-regulated several fold in this strain, though a repres-
sion mechanism for lysine biosynthesis is not known in C. glutamicum. Althoughthe genetic elements responsible for these changes have not yet been identified,
it has been demonstrated that the introduction of a mutant allele of the leuC gene
into a defined lysine producer triggered a stringent-like global response and thereby
led to a significant increase in lysine production (Hayashi et al. 2006a).
Engineering of global regulation has also been demonstrated to successfully
improve valine production by C. glutamicum. A pyruvate dehydrogenase-deficient
valine producer exhibited reduced glucose metabolism and a concomitant
nonproduction phenotype in the presence of acetate which was required for its
118 M. Ikeda and S. Takeno

growth. This drawback has been overcome by inactivating the global regulator SugR
which is responsible for acetate-mediated repression of the PTS (Blombach et al.
2009).
3.7 Stress Responses
Considerable heterogeneity exists within large-scale fermentors, especially with
respect to fed sugar, pH, and oxygen, due to the decrease in mixing efficiency
associated with the increase in the scale of an operation (Einsele 1978; Buckland
and Lilly 1993). Therefore, in large-scale industrial fermentations, the ambient
conditions vary considerably depending on the location of cells within the fermen-
tor. Thus, a cell’s tolerance to various stresses, referred to as cell robustness, is one
of the important characteristics which should be retained in a production strain.
Cells are assumed to possess a variety of mechanisms that allow them to adapt to
stressful conditions. A common stressor is external variation in pH, against which
each cell must maintain the internal pH of its cytoplasm. Although the mechanisms
of pH homeostasis in C. glutamicum are poorly understood, recent studies have
shown critical involvement of a putative transporter of the cation diffusion facilita-
tor family and a potential potassium channel (CglK) in alkaline and acidic pH
homeostasis, respectively (Takeno et al. 2008; Follmann et al. 2009a). Very
recently, transcriptome and proteome studies have revealed that C. glutamicumcan exhibit a variety of stress responses when subjected to acidic or alkaline pH
conditions (Follmann et al. 2009b).
Similarly,C. glutamicum is likely to possess somemeans of adaptation to conditions
of limited oxygen. Recently, the relevant genes have been isolated from the C.glutamicum genome by genetic complementation of mutants that have lost the ability
to grow under conditions of low oxygen (Ikeda et al. 2009a). Putative functions
encoded by these genes include SigD, Ferredoxin, Siderophore, and Cytidylate kinase;
there is also a set of membrane proteins which have not yet been assigned functions
even tentatively. Some of these genes have been demonstrated to show cross-
complementation of different mutants under oxygen limitation, suggesting the utility
of the genes for improving growth and production in industrial fermentation.
Examples of other stress response proteins of C. glutamicum include SigH
(Kim et al. 2005a) and WhcE (Kim et al. 2005b), which are involved in heat and
oxidative stress responses; and BetP (Peter et al. 1996), EctP (Peter et al. 1998),
ProP (Peter et al. 1998), LcoP (Steger et al. 2004), MtrB (M€oker et al. 2007), andMtrA (M€oker et al. 2007), all of which are involved in osmotic stress response.
These are potential targets for engineering in the future.
3.8 Feedstock Utilization
The main feedstocks for industrial amino acid fermentation by C. glutamicum are
sugars from agricultural crops, such as cane molasses, beet molasses, and starch
Amino Acid Production by Corynebacterium glutamicum 119

hydrolysates (glucose) from corn and cassava, but it is becoming increasingly
necessary to engineer the use of alternative raw materials, especially those that do
not compete with human food or energy sources. Wild-type C. glutamicum cannot
utilize lactose, galactose, starch, glycerol, xylose, or arabinose for growth, but
strains that do utilize these carbon sources have recently been engineered.
For example, heterologous expression of both lacYZ from Lactobacillus delbrueckiisubsp. bulgaricus and galMKTE from Lactococcus lactis subsp. cremoris in a
lysine-producing strain of C. glutamicum has resulted in a strain that is able to
produce lysine at up to 2 g/L when fed whey, which contains lactose and galactose
(Barrett et al. 2004). Another lysine-producing C. glutamicum strain has been
engineered to express the a-amylase gene from Streptomyces griseus, which allowsit to utilize soluble starch for lysine production, albeit at an efficiency lower
than that obtained using glucose (Seibold et al. 2006). More efficient lysine
production from soluble starch by C. glutamicum has been achieved by displaying
the a-amylase from Streptococcus bovis on the cell surface. As the anchor protein,
PgsA from Bacillus subtilis was fused to the N terminus of the a-amylase. A lysine
producer displaying this fusion protein on its cell surface produced 6 g/L of lysine
with a conversion yield of 18.9% on starch; this titer and yield are higher than those
obtained in glucose medium (Tateno et al. 2007). A similar cell surface display of
a-amylase has also enabled C. glutamicum to produce glutamate from starch (Yao
et al. 2009).
Glycerol, the main by-product of biodiesel production, is also a potential carbon
source for biotechnological processes. C. glutamicum has been engineered to
express the E. coli glycerol utilization genes glpF, glpK, and glpD so that it can
grow on glycerol. This allowed the production of glutamate and lysine from
glycerol with yields of 11% and 19%, respectively (Rittmann et al. 2008).
The use of lignocellulose as a feedstock is limited in part by a poor catabolism of
the xylose component. A xylose-utilizing C. glutamicum strain has been
constructed that expresses the xylA and xylB genes from E. coli on a high-copy
plasmid. It is interesting that the E. coli xylB gene contributed to improved growth
performance on xylose despite the existence of a functional xylB gene in the
C. glutamicum wild-type genome (Kawaguchi et al. 2006). Similarly, heterologous
expression of the E. coli arabinose-utilizing pathway in C. glutamicum resulted in a
strain that is able to grow on arabinose, another component of lignocellulose
(Kawaguchi et al. 2008).
4 Recent Advances in Amino Acid Production
Recently, various genetic engineering tools and global analysis techniques for
C. glutamicum as well as high-throughput genomic analysis technologies have
been successfully applied and have contributed both to the understanding of the
molecular mechanisms underlying high-level production and to the development of
more efficient production strains of this microbe. For example, DNA arrays have
120 M. Ikeda and S. Takeno

been used to find engineering targets expected to result in improved valine produc-
tion (Lange et al. 2003) and to identify mutations that confer traits conducive to
high-level production in lysine-producing strains through random mutagenesis and
screening programs (Hayashi et al. 2006a; Sindelar and Wendisch 2007). Here,
recent advances in amino acid production by C. glutamicum are highlighted, with a
focus on the amino acids whose production methods have been significantly
advanced in the 2000s. These include glutamate, lysine, arginine (citrulline), the
branched-chain amino acids, alanine, serine, and the sulfur-containing amino acids
methionine and cysteine. Production technology of other industrially important
amino acids such as threonine and the aromatic amino acids has been omitted
because it is discussed in other publications (Ikeda 2003; Willis et al. 2005;
Sprenger 2007; Rieping and Hermann 2007; Dong et al. 2011).
4.1 Glutamate
Since the discovery of C. glutamicum as a producer of the food flavoring
monosodium glutamate, commercial production of glutamate has been conducted
exclusively by this microbe. In 2006, the global demand for monosodium glutamate
amounted to almost two million metric tons (Fig. 1), and the market is expected to
continue its gradual expansion at an average annual rate of 3–4% (Ajinomoto
2007). Glutamate production by C. glutamicum is induced by biotin limitation or
by treatment with certain fatty acid ester surfactants or with b-lactam antibiotics
such as penicillin. Although the induction treatment is the core technology involved
in industrial glutamate production processes, the molecular basis of the induction of
glutamate secretion was long unknown. In recent years, however, a valuable insight
into the secretion mechanism has been gained in the form of the identification of the
NCgl1221 gene product as a possible glutamate exporter (Nakamura et al. 2007).
An intriguing finding is that only a specific point mutation in the NCgl1221 gene
resulted in glutamate secretion without any induction treatments. It has also been
shown that amplification of the wild-type NCgl1221 gene increases glutamate
secretion while its disruption substantially abolishes secretion accompanied by an
increase in the intracellular glutamate pool under the induction conditions men-
tioned earlier. The gene in question encodes the YggB protein which has been
described as a putative mechanosensitive channel (Nottebrock et al. 2003). Based
on the possible function as a mechanosensitive channel, the following mechanism
has been proposed: the induction conditions, such as biotin limitation and penicillin
treatment, alter membrane tension by inhibiting lipid or peptidoglycan synthesis.
This triggers conformational changes in the NCgl1221 gene product, which in turn
enables the protein to export glutamate (Fig. 5). To confirm this model, one must
determine how energy is supplied into the glutamate secretion process because
carrier-mediated glutamate secretion by C. glutamicum was shown to be energy
dependent (Gutmann et al. 1992).
The observation mentioned earlier raises the question of how the new model
is congruent with the accepted notion that a decrease in the activity of the
Amino Acid Production by Corynebacterium glutamicum 121

2-oxoglutarate dehydrogenase complex (ODHC) is crucial for glutamate production
(Shingu and Terui 1971; Kawahara et al. 1997; Kimura 2003; Asakura et al. 2007;
Kim et al. 2009a, b). Although the new model seems to explain the basics of the
mechanism underlying the induction of glutamate secretion, it is probably not
sufficient to explain the entire process of glutamate production by C. glutamicum.Recently, a possible connection at a molecular level has been uncovered between
ODHC activity and glutamate production (Fig. 5). A novel 15 kDa protein OdhI was
identified as a regulator of ODHC (Niebisch et al. 2006). The unphosphorylated
form of OdhI binds to the OdhA protein, one of the subunits of ODHC, and inhibits
the ODHC activity. This inhibition can be prevented by the PknG-catalyzed
phosphorylation of OdhI. A phospho-serine/threonine protein phosphatase
responsible for dephosphorylation of OdhI has also been identified (Niebisch et al.
2006). Interestingly, disruption of the odhI gene was shown to abolish glutamate
production even under the induction conditions (Schultz et al. 2007), suggesting a
close relationship between the regulator protein and the reduction of the ODHC
activity that occurs during glutamate production. It is also worth noting that
proteome analyses have revealed a significant increase in the OdhI protein upon
penicillin treatment, which has become a conventional industrial method to trigger
glutamate production (Kim et al. 2009a, b). These findings have confirmed the
existence of a connection between ODHC activity and glutamate production, but it
should be noted that an ODHC-activity-reducing metabolic change alone is not
sufficient to induce glutamate production (Kim et al. 2009a, b).
These results taken all together, the evidence to date suggests a link between
the induction treatments, such as biotin limitation and penicillin treatment, and
Glucose
Glutamate
Phosphoenol -pyruvate
Pyruvate
Acetyl-CoA
Biotin
Fatty acid
Oxaloacetate
OxoglutarateYggB
?
AccBCAccBCDtsRDtsR
PknGPknG
OdhIOdhI
ODHCODHC
Fig. 5 Possible mechanism triggering glutamate overproduction under the induction conditions in
C. glutamicum. In this model, proteins AccBC and DtsR form the biotin-dependent acetyl-CoA
carboxylase complex required for fatty acid biosynthesis; this biotin–enzyme complex is thought
to be the primary target of biotin limitation and surfactant addition
122 M. Ikeda and S. Takeno

glutamate production. In our proposed mechanism, the induction treatments
enhance the synthesis of the regulator protein OdhI in its unphosphorylated form
and thereby inhibit ODHC activity. This causes a metabolic shift at the branch point
of 2-oxoglutarate, which channels carbon toward glutamate. Intracellularly
accumulated glutamate is then secreted into the medium via the NCgl1221 gene
product, a possible glutamate exporter, which has been activated in response to
altered membrane tension (Fig. 5). Questions for the future include why and how
the OdhI protein is overexpressed in response to the induction treatment and what
conditions are required for the phosphorylation and dephosphorylation of OdhI.
Continuous efforts have been made not only to understand glutamate production
but also to improve the process. In addition to the general approaches, in which
metabolic fluxes are directed into glutamate (Kimura 2003; Sato et al. 2008;
Sawada et al. 2010), an innovative metabolic design allowing an increased
maximum theoretical yield has recently been reported (Chinen et al. 2007).
Glutamate biosynthesis from glucose in C. glutamicum is inevitably associated
with the release of CO2 in the pyruvate dehydrogenase reaction, but the creation
of a novel metabolic route by installing the phosphoketolase pathway of
Bifidobacterium animalis allowed the CO2-releasing pyruvate dehydrogenase
reaction to be bypassed, and thereby led to increased glutamate production
coupled with the suppression of CO2 emission. On the other hand, expression
of the Vitreoscilla hemoglobin gene vgb under a tac promoter in a wild-type
C. glutamicum strain has been shown to increase glutamate production in both
shake-flask and fermentor cultivations (Liu et al. 2008), probably due to the
enhancement of respiration by the hemoglobin (Webster 1987; Kallio et al. 1994;
Zhang et al. 2007a, b).
4.2 Lysine
Lysine, one of the essential amino acids for animals, has a significant commercial
value as a feed additive to promote the growth of animals including swine and
poultry, and thus is the second-ranking amino acid after glutamate in terms of
worldwide annual production. The scale of the lysine market in 2006 has been
estimated at approximately 960,000 metric tons (Fig. 1), and the market is expected
to grow continuously at an annual rate of 8–10% (Ajinomoto 2007). Because of the
growing market for lysine, exhaustive studies have been undertaken in an attempt to
engineer the metabolism of C. glutamicum for lysine production. These studies
have resulted in several effective strategies for rational strain improvement, includ-
ing engineering of terminal pathways, central metabolism, cofactor-regeneration
systems, export systems, energy metabolism, and global regulation. Typical
examples of these have been discussed earlier.
Recently, a genome-scale model of the C. glutamicum metabolic network has
been constructed, based on the annotated genome, available literature, and various
“omic” data (Kjeldsen and Nielsen 2009). The constructed metabolic model
Amino Acid Production by Corynebacterium glutamicum 123

consists of 446 reactions and 411 metabolites; the predicted metabolic fluxes during
lysine production and growth under various conditions are highly consistent with
experimental values. The ability to predict the metabolic state associated with
maximum production yield can be used to guide strain engineering. This strategy
has been proven through the rational design of high lysine-producing strains of
C. glutamicum (Kr€omer et al. 2004; Becker et al. 2005; Wendisch et al. 2006).
In addition to such metabolic engineering approaches, a genome-based reverse
engineering approach has been employed to create a more robust and efficient
lysine producer (Ohnishi et al. 2002, 2003; Ikeda and Nakagawa 2003; Ikeda et al.
2006). Reverse engineering is so called because it traces backwards to an existing
classical producer. First, the genome sequence of an industrial lysine producer was
compared with a corresponding wild-type sequence to identify the mutational
differences. The mutations were then sequentially introduced by allelic replace-
ment into the wild-type genome (Ikeda et al. 2005). Mutations in the relevant
terminal pathways were introduced first, followed by those in central metabolism,
and finally those in genes involved in global regulation. Each of the strains thus
constructed was evaluated to determine the contribution of each mutation to
production. When the mutation was beneficial, the resulting strain was used as
the parent to which further mutations were introduced for evaluation. This iterative
cycle has led to a minimally mutated strain having only useful mutations. It should
be noted that the particular host strain used at the beginning of the process can have
a significant impact on the ultimate outcome. When reverse engineering was
applied to lysine production, lysC311, a key mutation that confers high-level lysine
production on wild-type C. glutamicumwas used to screen various wild-type strains
of C. glutamicum to identify the best background with which to begin the process
(Ohnishi and Ikeda 2006).
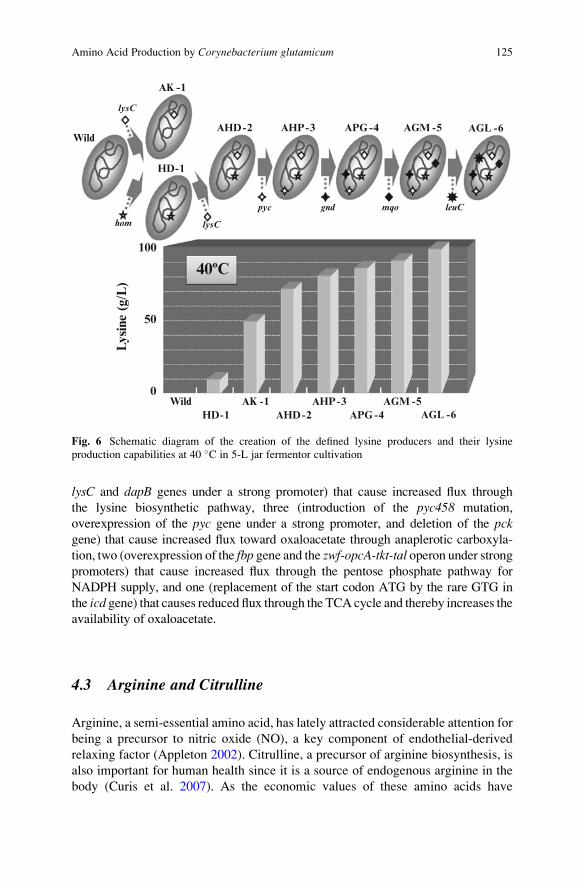
Among the useful mutations identified through the reverse engineering process
were two (hom59 and lysC311) that are located in the terminal pathway to lysine
(Ohnishi et al. 2002), three (pyc458, gnd361, and mqo224) involved in central
metabolism (Ohnishi et al. 2002, 2005; Mitsuhashi et al. 2006), and one (leuC456)causing global induction of the amino acid-biosynthetic genes and thereby further
increasing production (Hayashi et al. 2006a). The subsequent assembly of these six
useful mutations into the industrially robust wild-type strain chosen as the best
background was shown to substantially improve producer performance, resulting in
a final titer of 100 g/L after only 30 h of 5-L jar fermentor cultivation at a
suboptimal temperature of 40 �C (Fig. 6, Ikeda et al. 2006).
Most recently, the cumulative body of knowledge on lysine production was
combined with metabolic flux profiling and modeling technologies and systemized
to predict a combination of genetic modifications that would lead to the theoretically
best flux scenario for optimum lysine production (Becker et al. 2011). Ultimately,
the process has identified 12 steps of modifications to a wild-type genome leading to
the lysine hyper-producer, LYH-12 (Fig. 7), which can achieve a final titer of 120 g/L
with a conversion yield of 55% on glucose after 30 h of 5-L jar fermentor cultivation
at 30�C. Among the twelve modifications were six (introduction of the lysC311 and
hom59 mutations, duplication of the ddh and lysA genes, and overexpression of the
124 M. Ikeda and S. Takeno
lysC and dapB genes under a strong promoter) that cause increased flux through
the lysine biosynthetic pathway, three (introduction of the pyc458 mutation,
overexpression of the pyc gene under a strong promoter, and deletion of the pckgene) that cause increased flux toward oxaloacetate through anaplerotic carboxyla-
tion, two (overexpression of the fbp gene and the zwf-opcA-tkt-tal operon under strongpromoters) that cause increased flux through the pentose phosphate pathway for
NADPH supply, and one (replacement of the start codon ATG by the rare GTG in
the icd gene) that causes reduced flux through the TCA cycle and thereby increases the
availability of oxaloacetate.
4.3 Arginine and Citrulline
Arginine, a semi-essential amino acid, has lately attracted considerable attention for
being a precursor to nitric oxide (NO), a key component of endothelial-derived
relaxing factor (Appleton 2002). Citrulline, a precursor of arginine biosynthesis, is
also important for human health since it is a source of endogenous arginine in the
body (Curis et al. 2007). As the economic values of these amino acids have
lysC
hom
pyc gnd mqo
lysC
AK -1
AHD-2 AHP-3 APG -4 AGM -5Wild
HD-1
leuC
AGL -6
Wild AK -1AHD-2
AHP-3APG -4
AGM -5HD-1
50
100
0
AGL -6
Lys
ine
(g/L
)
40ºC
Fig. 6 Schematic diagram of the creation of the defined lysine producers and their lysine
production capabilities at 40 �C in 5-L jar fermentor cultivation
Amino Acid Production by Corynebacterium glutamicum 125

increased, considerable attention has recently been given to the development of
more efficient production strains, as well as to the analysis of arginine metabolism
in microbes (Utagawa 2004; Glansdorff and Xu 2007; Lee et al. 2010). The latest
research in the field of production technology involves the reengineering of an
arginine and citrulline-producing strain of C. glutamicum; in this experiment,
positive mutations derived from three different lines of classical producers were
systematically assembled into a single wild-type background (Fig. 8) (Ikeda et al.
2009b). The procedure and impact of this advanced approach are summarized
as follows:
1. The first step was to identify the basic mutation(s) causing arginine and citrulline
overproduction in wild-type C. glutamicum. For this purpose, three independentlyderived industrial arginine and citrulline producer strains were sequenced and
compared to their wild-type ancestors. This identified a variety of mutations
potentially associated with arginine biosynthesis. Among these, five specific
mutations (argB26, argB31, argR123, argG92up, and argG45) located within
arg operons were examined in a wild-type background for their relevance to
arginine and citrulline production. argB26 and DargR (argR123-derived deletion
mutation) were found to be the basic mutations.
2. The second step was to screen for the wild-type background giving the best
performance. The two basic mutations, argB26 and DargR, were introduced into
Glucose
Phosphoenolpyruvate
Pyruvate
Oxaloacetate
Oxoglutarate
Glucose 6-P
Fructose 6-P
Fructose1,6-BP
Glyceraldehyde 3-P
Ribulose 5-P
Xylulose 5-P Ribose 5-P
Erythrose 5-P Sedoheptulose 7-P
Aspartate
Aspartyl-P
Aspartatesemialdehyde
Threonine
Lysine
Diaminopimelate
Piperideinedicarboxylate
Lysine
Isocitrate
zwf pgl
tkttkt
tal
icd
pyc
lysC
dapB
ddh
lysA
fbp
hom
pck
Overexpression
Attenuation
opcA
Fig. 7 Schematic diagram of genetic modifications to a wild-type genome leading to the lysine
hyper-producer C. glutamicum LYS-12
126 M. Ikeda and S. Takeno
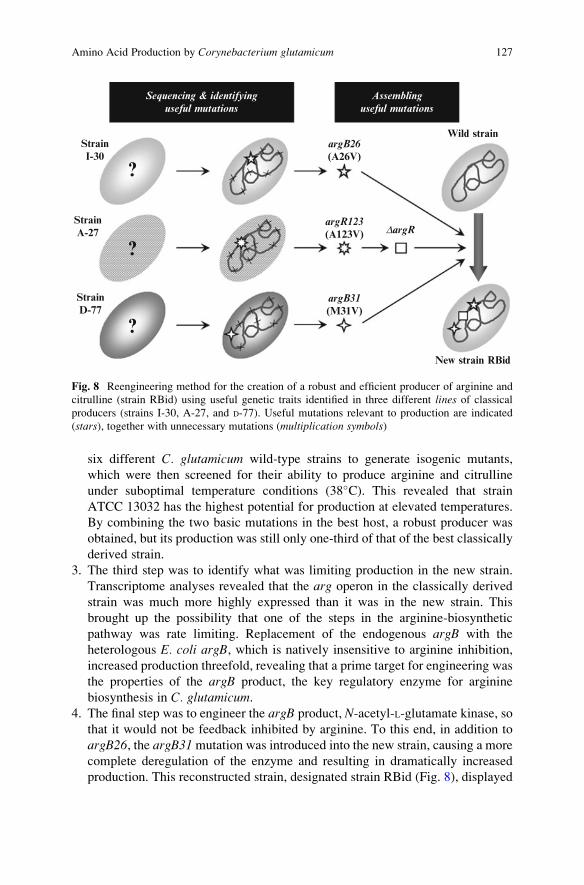
six different C. glutamicum wild-type strains to generate isogenic mutants,
which were then screened for their ability to produce arginine and citrulline
under suboptimal temperature conditions (38�C). This revealed that strain
ATCC 13032 has the highest potential for production at elevated temperatures.
By combining the two basic mutations in the best host, a robust producer was
obtained, but its production was still only one-third of that of the best classically
derived strain.
3. The third step was to identify what was limiting production in the new strain.
Transcriptome analyses revealed that the arg operon in the classically derived
strain was much more highly expressed than it was in the new strain. This
brought up the possibility that one of the steps in the arginine-biosynthetic
pathway was rate limiting. Replacement of the endogenous argB with the
heterologous E. coli argB, which is natively insensitive to arginine inhibition,
increased production threefold, revealing that a prime target for engineering was
the properties of the argB product, the key regulatory enzyme for arginine
biosynthesis in C. glutamicum.4. The final step was to engineer the argB product, N-acetyl-L-glutamate kinase, so
that it would not be feedback inhibited by arginine. To this end, in addition to
argB26, the argB31mutation was introduced into the new strain, causing a more
complete deregulation of the enzyme and resulting in dramatically increased
production. This reconstructed strain, designated strain RBid (Fig. 8), displayed
StrainI-30
?
?
?
StrainA-27
StrainD-77
argB26
(A26V)
argR123
(A123V) DargR
argB31
(M31V)
New strain RBid
Wild strain
Sequencing & identifying
useful mutations
Assembling
useful mutations
Fig. 8 Reengineering method for the creation of a robust and efficient producer of arginine and
citrulline (strain RBid) using useful genetic traits identified in three different lines of classical
producers (strains I-30, A-27, and D-77). Useful mutations relevant to production are indicated
(stars), together with unnecessary mutations (multiplication symbols)
Amino Acid Production by Corynebacterium glutamicum 127
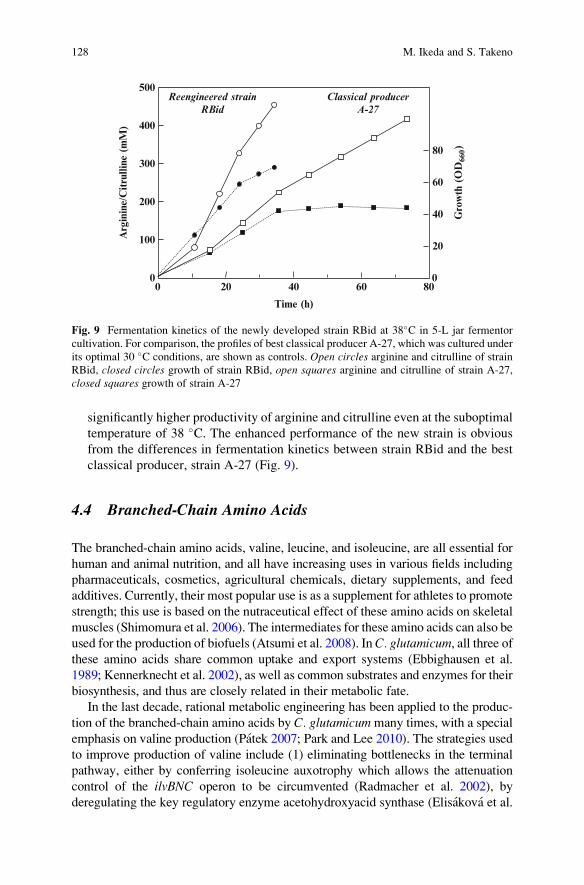
significantly higher productivity of arginine and citrulline even at the suboptimal
temperature of 38 �C. The enhanced performance of the new strain is obvious
from the differences in fermentation kinetics between strain RBid and the best
classical producer, strain A-27 (Fig. 9).
4.4 Branched-Chain Amino Acids
The branched-chain amino acids, valine, leucine, and isoleucine, are all essential for
human and animal nutrition, and all have increasing uses in various fields including
pharmaceuticals, cosmetics, agricultural chemicals, dietary supplements, and feed
additives. Currently, their most popular use is as a supplement for athletes to promote
strength; this use is based on the nutraceutical effect of these amino acids on skeletal
muscles (Shimomura et al. 2006). The intermediates for these amino acids can also be
used for the production of biofuels (Atsumi et al. 2008). InC. glutamicum, all three ofthese amino acids share common uptake and export systems (Ebbighausen et al.
1989; Kennerknecht et al. 2002), as well as common substrates and enzymes for their
biosynthesis, and thus are closely related in their metabolic fate.
In the last decade, rational metabolic engineering has been applied to the produc-
tion of the branched-chain amino acids by C. glutamicum many times, with a special
emphasis on valine production (Patek 2007; Park and Lee 2010). The strategies used
to improve production of valine include (1) eliminating bottlenecks in the terminal
pathway, either by conferring isoleucine auxotrophy which allows the attenuation
control of the ilvBNC operon to be circumvented (Radmacher et al. 2002), by
deregulating the key regulatory enzyme acetohydroxyacid synthase (Elisakova et al.
0 20 40 60
Gro
wth
(O
D66
0)
Arg
inin
e/C
itru
lline
(m
M)
Time (h)
40
80
100
0
500
400
300
200
0
60
20
80
Reengineered strain
RBid
Classical producer
A-27
Fig. 9 Fermentation kinetics of the newly developed strain RBid at 38�C in 5-L jar fermentor
cultivation. For comparison, the profiles of best classical producer A-27, which was cultured under
its optimal 30 �C conditions, are shown as controls. Open circles arginine and citrulline of strain
RBid, closed circles growth of strain RBid, open squares arginine and citrulline of strain A-27,
closed squares growth of strain A-27
128 M. Ikeda and S. Takeno

2005), or by overexpressing the gene set responsible for valine biosynthesis
(Radmacher et al. 2002; Blombach et al. 2007a; Bartek et al. 2010a, b); (2) increasing
the availability of precursor pyruvate, either by blocking pantothenate synthesis
(Radmacher et al. 2002; Bartek et al. 2008) or by inactivating pyruvate dehydroge-
nase, pyruvate carboxylase, and pyruvate:quinine oxidoreductase (Blombach et al.
2007a, 2008, 2009); and (3) increasing NADPH supply by inactivating phos-
phoglucose isomerase (Blombach et al. 2008; Bartek et al. 2010a, b). These
modifications have mostly been achieved through plasmid-mediated amplification
and/or deletion of the targeted genes, possibly leading to perturbations of the natural
homeostatic mechanisms of the cell. To alleviate such side effects on cell physiology,
the desired metabolic engineering has been achieved through purposeful mutagenesis
of promoters of the chromosomal genes involved in the valine biosynthesis pathway
and in competing pathways (Holatko et al. 2009). The resulting plasmid-free valine
producer was auxotrophic to pantothenate and bradytrophic to isoleucine, carried
a feedback-resistant acetohydroxy acid synthase, and expressed the genes ilvD and
ilvE from strong mutant promoters. This new type strain with all mutations
constructed within the chromosome has been shown to produce 136 mM valine
from 4% glucose after 48 h of flask cultivation.
An H+-ATPase defect has also been shown to be effective for accelerating sugar
metabolism and increasing valine production in C. glutamicum (Wada et al. 2008).
Proteomic analysis has revealed various metabolic responses to this defect, including
upregulated expression of 6-phosphofructokinase and pyruvate kinase in the glycolytic
pathway (Li et al. 2007). These increases may contribute to the enhanced glycolysis
observed in the mutant, and thus to the increased availability of precursor pyruvate.
For the production of the branched-chain amino acids which are mainly used for
pharmaceutical purposes and are therefore required to have the highest degree of
purity, it is desirable to minimize by-production of other amino acids to a level at
which supplementary purification of the desired amino acid is not necessary.
By-production of alanine occurs during valine production, but the enzymes
involved in alanine formation from pyruvate by C. glutamicum had not been
identified until recently. Double knockout mutants of the alanine aminotransferase
gene (alaT, NCgl2747) and the alanine-valine transaminase gene (avtA, NCgl2510)were shown to be auxotrophic for alanine, revealing that the two gene products are
the only aminotransferases involved in alanine biosynthesis by C. glutamicum(Marienhagen and Eggeling 2008). Deletion of the alaT gene, which is primarily
responsible for alanine formation, in a valine producer actually reduces the extra-
cellular alanine accumulation, thereby facilitating cost-effective downstream
processing (Marienhagen and Eggeling 2008).
4.5 Alanine
Alanine, a nonessential amino acid, is used mainly as a sweetener in dairy products,
a natural moisture balancer in cosmetics, and an ingredient in infusion solutions and
pharmaceutical products. Unlike most amino acids, which are currently produced
Amino Acid Production by Corynebacterium glutamicum 129

by means of fermentative processes, alanine, the simplest optically active amino
acid, is produced commercially through the enzymatic decarboxylation of aspartate
(Shibatani et al. 1979) which is synthesized from fumarate and ammonia with
immobilized cells expressing aspartase. However, since the initial substrate, fuma-
rate, is produced primarily from petroleum, a fermentation process that uses
renewable feedstocks to produce alanine has attracted increasing attention.
Several alanine-producing microorganisms have long been known, including
C. glutamicum and E. coli (Kitai 1972), but alanine production at a useful level was
first demonstrated in a natural isolate, Arthrobacter oxydans HAP-1 (Hashimoto and
Katsumata 1993, 1998). It has since been determined that alanine hyperproduction
results from the combination of the presence of the glucose-nonrepressive, NADH-
dependent alanine dehydrogenase and decreases in the activities of pyruvate dehydro-
genase and NADH oxydase in the stationary phase, which allows balanced coupling of
redox potential between glycolysis and the dehydrogenase pathway (Hashimoto and
Katsumata 1999). Aside from the successful example utilizing the intrinsic
characteristics of A. oxydans, metabolic engineering has enabled several other
microorganisms such as Zymomonas mobilis, Lactococcus lactis, andE. coli to producehigh levels of alanine from sugar (Uhlenbusch et al. 1991; Hols et al. 1999; Smith et al.
2006; Zhang et al. 2007a, b). The most common strategies for alanine production
involve the heterologous expression of NADH-dependent alanine dehydrogenase in
hosts, some of which already possess defects in alanine racemase and/or the pathways
that compete with alanine biosynthesis. C. glutamicum has also been demonstrated to
be a useful host for alanine production (Katsumata and Hashimoto 1996). As in many
other microorganisms, biosynthesis of alanine in C. glutamicum depends on the
transamination reaction from pyruvate and glutamate. Expression of alanine dehydro-
genase from A. oxydans in an alanine racemase-deficient C. glutamicum strain has
resulted in hyperproduction of L-alanine isomer under oxygen limitation (Fig. 10).
The process of alanine production by C. glutamicum is economically significant,
but has the drawback of relatively low productivity due to decreased glucose
metabolism under oxygen-limited conditions. Very recently, it has been shown by
a bioprocess using growth-arrested packed cells that homologous overexpression of
the glyceraldehyde 3-phosphate dehydrogenase gene in a C. glutamicum strain
expressing alanine dehydrogenase from Lysinibacillus sphaericus and simulta-
neously deficient in the genes associated with by-production of organic acids can
dramatically improve glucose metabolism and thereby also alanine productivity
under conditions of oxygen deprivation (Fig. 10, Jojima et al. 2010). Further inacti-
vation of alanine racemase has led to the production of L-alanine isomer with a chiral
purity greater than 99.5%. This bioprocess has achieved a final titer of 98 g/L of L-
alanine with a conversion yield of 83% on glucose after 32 h.
4.6 Serine
Serine is likewise a nonessential amino acid, but one that is important in metabo-
lism because it participates in the biosynthesis of many important metabolites such
130 M. Ikeda and S. Takeno

as purines, pyrimidines, cysteine, and tryptophan. Although the amount of serine
produced each year worldwide is relatively small compared with the amounts of the
other amino acids mentioned earlier, serine is of significant commercial value as an
ingredient in pharmaceuticals such as infusion solutions, a natural moisturizing
material in cosmetics, and a feed additive.
Serine is among the few amino acids for which high-production yields were not
achieved by fermentation directly from sugar until recently. Classical mutagenesis
and screening for a strain producing serine from glucose had not resulted in
a practical production method. Recently, however, an attempt at systematic
metabolic engineering resulted in a C. glutamicum strain that produces considerable
amounts of serine, with help in the form of interventions at several points in the
complicated metabolism of the amino acid (Fig. 11, Peters-Wendisch et al. 2005).
Initial overexpression of serine biosynthesis genes (serAD197, serC, and serB)coding for deregulated enzymes did not lead to significant serine accumulation,
nor did the further deletion of the serine dehydratase gene (sdaA) that catalyzesserine degradation to pyruvate. Only by reducing the glyA-encoded serine
hydroxymethyltransferase (SHMT) activity was considerable serine accumulation
ultimately achieved. Since SHMT, being the unique route to the C1 supply, is
essential for growth of C. glutamicum, reducing the activity of this enzyme by
exactly the right amount was key. This was initially accomplished by replacing the
glyA promoter with the tac promoter, which reduced glyA expression in the absence
of isopropyl-thio-b-D-galactopyranoside. As it turned out, this method of glyAcontrol was unstable: since mutations in lacIq restored expression. As an alternate
method of glyA expression control, a more reliable physiological method was
developed in which the strain is made auxotrophic for folate by deleting the
Glucose
NADH
NAD+
Hypoxic conditions
L-Alanine L-AlaninePyruvateLactate
Glutamate Oxoglutarate
D-Alanine
NH4+
AlaATAlaAT
GAPDHGAPDH
AlaDHAlaDHLDHLDH
ALRALR
Fig. 10 Outline of metabolic engineering for L-alanine production inC. glutamicum. AlaDH alanine
dehydrogenase from A. oxydans or L. sphaericus, ALR alanine racemase, AlaAT alanine aminotrans-
ferase, GAPDH glyceraldehydes 3-phosphate dehydrogenase, LDH lactate dehydrogenase
Amino Acid Production by Corynebacterium glutamicum 131

pabABC genes so that SHMT activity can be controlled by the availability of
5,6,7,8-tetrahydrofolate (THF). This resulted in an accumulation of 345 mM of
serine in a 20-L controlled fed-batch culture (Stolz et al. 2007).
4.7 Methionine
Methionine, another essential amino acid for animals, has a great deal of commer-
cial value as a feed additive. For this purpose, methionine is produced exclusively
by chemical synthesis in D, L-forms, as this amino acid is considered to have a
similar effect on animal nutrition in both L- and D, L-forms. Nowadays, however,
there is an increasing interest in the development of environmentally friendly
fermentation methods using renewable feedstocks to produce methionine.
In the hope of discovering a method for the rational construction of a methionine
producer, methionine biosynthesis and its regulation are being studied in
C. glutamicum. Recently, C. glutamicum was shown to possess both transsulfuration
and direct sulfhydrylation pathways, in contrast to most microorganisms, including
E. coli, which utilize only one of these two pathways (Lee and Hwang 2003; Hwanget al. 2007). Metabolic engineering to redirect carbon from the lysine pathway into
the methionine pathway led to a C. glutamicum strain that produced 2.9 g/L of
methionine, together with 23.8 g/L of lysine (Park et al. 2007). Overexpression of
the homologous metX and metY genes in another lysine-producing C. glutamicumstrain was reported in a patent byM€ockel et al. (2002) to result in a final titer of 16 g/Lof methionine. Two regulatory genes in C. glutamicum have been identified as being
relevant to methionine biosynthesis: mcbR (cg3253) and NCgl2640. Inactivation of
Glucose
Serine3-Phosphoglycerate
Pyruvate
GlycineMethylene-
THF
THF
Plasmid
Serine
Folate
DsdaADsdaA
serACBserACB
SHMTSHMT
DpabABCDpabABC
Fig. 11 Schematic diagram of genetically engineered serine-producing C. glutamicum strain
132 M. Ikeda and S. Takeno

either in wild-type C. glutamicum results in increased methionine production
(Mampel et al. 2005; Rey et al. 2003, 2005).
Very recently, an attempt at systematic metabolic engineering resulted in an
E. coli strain that produces methionine at an industrially useful level (Figge et al.
2009). The key to success here was achieving a balanced supply of three important
precursors for methionine biosynthesis: O-succinylhomoserine, cysteine, and a C1
carbon, methyl-tetrahydrofolate (CH3-THF). An imbalanced supply of these
precursors causes the formation of undesired by-products, such as homolanthionine
and isoleucine, through the involvement of certain methionine-biosynthetic
enzymes themselves. The procedure and impact of the metabolic engineering
involved in the production of this strain can be summarized as follows (Fig. 12):
1. The first step was to eliminate bottlenecks in the core pathways leading to three
precursors. For this purpose, the metF, metH, and cysE genes, as well as mutant
alleles of the metA and thrA genes that encodes enzymes less sensitive to
feedback inhibition, were overexpressed in an E. coli strain deficient in the
methionine repressor gene metJ. This engineering resulted in methionine pro-
duction at the yield of 6.7% on glucose in 50-mL batch culture.
2. The second step was to make more sulfur available for cysteine biosynthesis.
For this purpose, the cysPUWAM and cysJIH gene clusters were overexpressed,
increasing the yield up to 8.5%.
Glucose
3-Phospho-glycerate
Phosphoenol-pyruvate
Pyruvate
Serine
H2S
CysteineTHF
CH2-THF CH3-THF
Glycine
Oxaloacetate
Aspartate HomoserineO-succinyl-homoserine
γ-Cystathioine
HomocysteineMethionine
COOH
S-adenosyl-methionine
O-Acetylserine
Asp-P Asp-SA
Threonine
Formyl-THFHomo-
lanthionine
Isoleucine
metB
metC
metB
metE
metL metL
Lysine
succinate
HistidinePurines
ppc
cysPUWA
cysJIH
cysM
metF
metA*
metH
serACB cysE
glyA
thrA*thrA*
ΔpykA
ΔpykF
ΔpurU
metA
metB
metC metEmetF
ΔmetJOverexpression Disruption
gcvTHP
NADPH
metB
Metabolic pathway
Overexpression
Feedback inhibition
S2O3
S2O3
Fig. 12 Schematic diagram of genetically engineered methionine-producing E. coli strain.
Asterisked genes encode enzymes less sensitive to feedback inhibition
Amino Acid Production by Corynebacterium glutamicum 133

3. The third step was to increase the availability of phosphoenolpyruvate for
biosynthesis of the precursor O-succinylhomoserine. Deletion of two pyruvate
kinase genes, pykA and pykF, further increased the yield up to 10.2%.
4. The fourth step was to channel more carbon toward a C1 source, in this case,
methyl-tetrahydrofolate CH3-THF, via serine and glycine. Overexpression of the
relevant genes including serACB, glyA, gcvTHP, andmetF, as well as deletion ofthe purU gene, further boosted the yield to 12.9%.
The engineered E. coli strain has achieved a yield of 19.9% after 50 h in
fed-batch fermentation without the formation of any detectable undesirable by-
products. Based on this yield, the methionine titer is estimated at more than 35 g/L.
Though some progress has been made toward creating improved methionine
producers, methionine yields still remain low compared with those attained for
other amino acids. Metabolic pathway analysis has been used to evaluate the
theoretical maximum yields of methionine production on the substrates glucose,
sulfate, and ammonia in C. glutamicum and E. coli (Kr€omer et al. 2006). The
theoretical yield (mol-C methionine per mol-C glucose) of C. glutamicumwas 0.49,
while that of E. coli was somewhat higher at 0.52. This analysis also showed that
introduction of the E. coli glycine cleavage system into C. glutamicum as an
additional C1 source and replacing sulfate with thiosulfate or sulfide, thereby
avoiding the need for reduction of oxidized sulfur, would increase the theoretical
maximal methionine yields in C. glutamicum to 0.57 and 0.68, respectively.
Furthermore, when methanethiol (also known as methylmercaptan) is used as a
combined source for a C1 carbon and sulfur in C. glutamicum, the theoretical yieldwas estimated to reach its highest potential value at 0.91 (Kr€omer et al. 2006).
Most recently, the potential utilization of methanethiol and its dimeric form
dimethyldisulfide as both the C1 source and the sulfur source has been experimen-
tally verified in C. glutamicum (Bolten et al. 2010). Isotope experiments have
revealed that the S-CH3 group is entirely added to O-acetylhomoserine, directly
yielding methionine (Fig. 13). It has also been shown that the reaction is catalyzed
by MetY, creating a shortcut for methionine biosynthesis. The problem in this case
would be the toxicity of these sulfur compounds to cells. A delivery system using a
beaded macroporous polystyrene resin has been suggested to be a way of alleviating
the toxic effects (Bolten et al. 2010).
4.8 Cysteine
Cysteine, the other sulfur-containing amino acid, is a nonessential amino acid but
has a crucial function in metabolism as a precursor of sulfur-containing compounds
such as methionine, thiamine, biotin, lipoic acid, and coenzyme A. In addition to its
biological significance, cysteine is important commercially because of its various
applications in the pharmaceutical, cosmetic, food, and livestock industries. Due to
the lack of an efficient method of producing cysteine through fermentation, its
134 M. Ikeda and S. Takeno
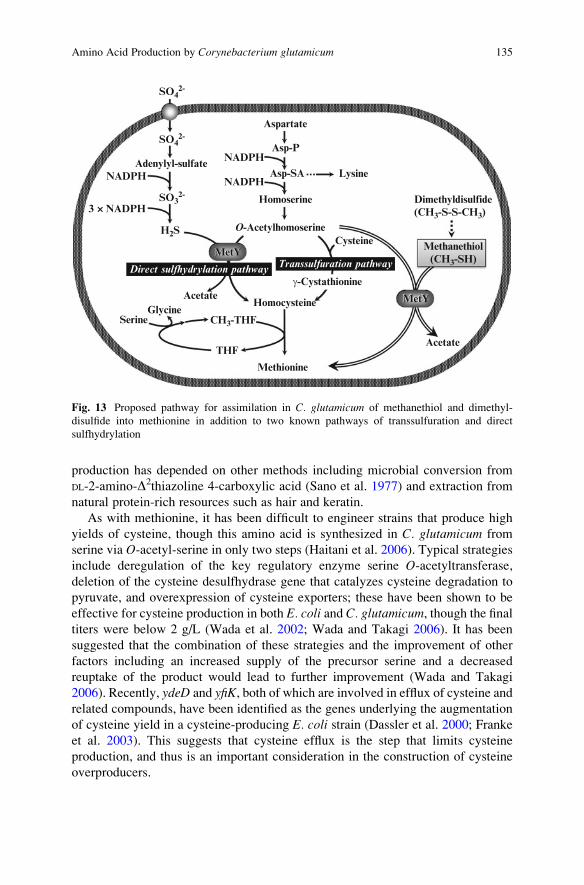
production has depended on other methods including microbial conversion from
DL-2-amino-D2thiazoline 4-carboxylic acid (Sano et al. 1977) and extraction from
natural protein-rich resources such as hair and keratin.
As with methionine, it has been difficult to engineer strains that produce high
yields of cysteine, though this amino acid is synthesized in C. glutamicum from
serine via O-acetyl-serine in only two steps (Haitani et al. 2006). Typical strategiesinclude deregulation of the key regulatory enzyme serine O-acetyltransferase,deletion of the cysteine desulfhydrase gene that catalyzes cysteine degradation to
pyruvate, and overexpression of cysteine exporters; these have been shown to be
effective for cysteine production in both E. coli and C. glutamicum, though the finaltiters were below 2 g/L (Wada et al. 2002; Wada and Takagi 2006). It has been
suggested that the combination of these strategies and the improvement of other
factors including an increased supply of the precursor serine and a decreased
reuptake of the product would lead to further improvement (Wada and Takagi
2006). Recently, ydeD and yfiK, both of which are involved in efflux of cysteine andrelated compounds, have been identified as the genes underlying the augmentation
of cysteine yield in a cysteine-producing E. coli strain (Dassler et al. 2000; Franke
et al. 2003). This suggests that cysteine efflux is the step that limits cysteine
production, and thus is an important consideration in the construction of cysteine
overproducers.
SO42-
SO42-
SO32-
Adenylyl-sulfate
H2S
NADPH
3 × NADPH
CH3-THF
THF
Homocysteine
Methionine
Aspartate
O-Acetylhomoserine
NADPH
NADPH
Cysteine
Acetate
SerineGlycine
Homoserine
Asp-P
Asp-SA Lysine
γ-Cystathionine
Methanethiol(CH3-SH)
Dimethyldisulfide(CH3-S-S-CH3)
Acetate
MetYMetY
MetYMetY
Direct sulfhydrylation pathwayTranssulfuration pathway
Fig. 13 Proposed pathway for assimilation in C. glutamicum of methanethiol and dimethyl-
disulfide into methionine in addition to two known pathways of transsulfuration and direct
sulfhydrylation
Amino Acid Production by Corynebacterium glutamicum 135

5 Conclusions and Outlook
The determination of the complete genome sequence of C. glutamicum was
obviously an important milestone in the history of amino acid fermentation. The
subsequent rapid progress in C. glutamicum genomics and so-called post-genome
technologies has opened up new avenues for the development of various global
analysis techniques, which have led to our current understanding of whole cellular
metabolism and systems in this microbe. Furthermore, these advances have dra-
matically transformed our approaches toward strain development. For example, in
silico modeling and simulation approaches are nowadays being used routinely to
help identify new targets for further engineering and strain improvement.
The power of such systems-level approaches will surely increase as modeling is
combined with the ever-accumulating “-omics” data. Meanwhile, the availability of
high-throughput DNA sequencing has made it feasible to decode the genomes of
classical industrial producers and thereby to identify important genetic traits that
distinguish them from their wild-type ancestors. As a result, the conventional style
of selecting improved strains by phenotypes, formerly the standard practice in
industry, is rapidly being replaced by the new method of reengineering strains by
assembling desirable genotypes. The reconstructed strains can be more robust, give
higher fermentation yields in less time, and resist stressful conditions better than
classical industrial producers.
In addition, these technologies can help us meet new and possibly unforeseen
challenges in the future: the amino acid industry is now beginning to consider
sustainable and environmentally friendly manufacturing systems in response to the
continuing crisis of global warming. There is a powerful drive to develop highly
efficient fermentation processes, especially for methionine and cysteine, the sulfur-
containing amino acids, using feedstocks that are renewable and that do not
compete with human food or energy sources. The development of strains that
enable the reduction in effluents and wastes generated during fermentation and
purification processes is also expected. These remain important themes for future
engineering.
References
Ajinomoto (2007) Fact sheets: amino acids business and feed-use amino acids business. http://
www.ajinomoto.com/ar/i_r/pdf/fact/Aminoacids-Oct2007.pdf http://www.ajinomoto.com/ar/
i_r/pdf/fact/Feed-useAA-Oct2007.pdf
Appleton J (2002) Arginine: clinical potential of a semi-essential amino acid. Altern Med Rev
7:512–522
Asakura Y, Kimura E, Usuda Y, Kawahara Y, Matsui K, Osumi T, Nakamatsu T (2007) Altered
metabolic flux due to deletion of odhA causes L-glutamate overproduction in Corynebacteriumglutamicum. Appl Environ Microbiol 73:1308–1319
Atsumi S, Hanai T, Liao JC (2008) Non-fermentative pathways for synthesis of branched-chain
higher alcohols as biofuels. Nature 451:86–89
136 M. Ikeda and S. Takeno

Azuma T, Nakanishi T (1988) Factors affecting L-arginine production in the continuous culture of
an l-arginine producer of Corynebacterium acetoacidophilum. J Ferment Technol 3:285–290
Azuma T, Nakanishi T, Sugimoto M (1988) Isolation and characterization of a stable L-arginine
producer from continuous culture broth of Corynebacterium acetoacidophilum. J Ferment
Technol 3:279–284
Barrett E, Stanton C, Zelder O, Fitzgerald G, Ross RP (2004) Heterologous expression of lactose-
and galactose-utilizing pathways from lactic acid bacteria in Corynebacterium glutamicum for
production of lysine in whey. Appl Environ Microbiol 70:2861–2866
Bartek T, Makus P, Klein B, Lang S, Oldiges M (2008) Influence of L-isoleucine and pantothenate
auxotrophy for L-valine formation in Corynebacterium glutamicum revisited by metabolome
analyses. Bioprocess Biosyst Eng 31:217–225
Bartek T, Blombach B, Z€onnchen E, Makus P, Lang S, Eikmanns BJ, Oldiges M (2010a)
Importance of NADPH supply for improved L-valine formation in Corynebacteriumglutamicum. Biotechnol Prog 26:361–371
Bartek T, Z€onnchen E, Klein B, Gerstmeir R, Makus P, Lang S, Oldiges M (2010b) Analysing
overexpression of L-valine biosynthesis genes in pyruvate-dehydrogenase-deficient Coryne-bacterium glutamicum. J Ind Microbiol Biotechnol 37:263–270
Becker J, Klopprogge C, Zelder O, Heinzle E, Wittmann C (2005) Amplified expression of
fructose 1,6-bisphosphatase in Corynebacterium glutamicum increases in vivo flux through
the pentose phosphate pathway and lysine production on different carbon sources. Appl
Environ Microbiol 71:8587–8596
Becker J, Klopprogge C, Herold A, Zelder O, Bolten CJ, Wittmann C (2007) Metabolic flux
engineering of L-lysine production in Corynebacterium glutamicum—over expression and
modification of G6P dehydrogenase. J Biotechnol 132:99–109
Becker J, Klopprogge C, Schr€oder H, Wittmann C (2009) Metabolic engineering of the tricarbox-
ylic acid cycle for improved lysine production by Corynebacterium glutamicum. Appl EnvironMicrobiol 75:7866–7869
Becker J, Zelder O, H€afner S, Schr€oder H, Wittmann C (2011) From zero to hero: design-based
systems metabolic engineering of Corynebacterium glutamicum for L-lysine production.
Metab Eng 13:159–168
Beckers G, Str€osser J, Hildebrandt U, Kalinowski J, Farwick M, Kr€amer R, Burkovski A (2005)
Regulation of AmtR-controlled gene expression in Corynebacterium glutamicum: mechanism
and characterization of the AmtR regulon. Mol Microbiol 58:580–595
Blombach B, Schreiner ME, Holatko J, Bartek T, Oldiges M, Eikmanns BJ (2007a) L-valine
production with pyruvate dehydrogenase complex-deficient Corynebacterium glutamicum.Appl Environ Microbiol 73:2079–2084
Blombach B, Schreiner ME, Moch M, Oldiges M, Eikmanns BJ (2007b) Effect of pyruvate
dehydrogenase complex deficiency on L-lysine production with Corynebacterium glutamicum.Appl Microbiol Biotechnol 76:615–623
Blombach B, Schreiner ME, Bartek T, Oldiges M, Eikmanns BJ (2008) Corynebacteriumglutamicum tailored for high-yield L-valine production. Appl Microbiol Biotechnol
79:471–479
Blombach B, Arndt A, Auchter M, Eikmanns BJ (2009) L-valine production during growth of
pyruvate dehydrogenase complex-deficient Corynebacterium glutamicum in the presence of
ethanol or by inactivation of the transcriptional regulator SugR. Appl Environ Microbiol
75:1197–1200
Bolten CJ, Schr€oder H, Dickschat J, Wittmann C (2010) Towards methionine overproduction in
Corynebacterium glutamicum: methanethiol and dimethyldisulfide as reduced sulfur sources.
J Microbiol Biotechnol 20:1196–1203
Bott M, Niebisch A (2003) The respiratory chain of Corynebacterium glutamicum. J Biotechnol104:129–153
Brockmann-Gretza O, Kalinowski J (2006) Global gene expression during stringent response in
Corynebacterium glutamicum in presence and absence of the rel gene encoding (p)ppGpp
synthase. BMC Genomics 7:230
Amino Acid Production by Corynebacterium glutamicum 137

Brune I, Jochmann N, Brinkrolf K, H€user AT, Gerstmeir R, Eikmanns BJ, Kalinowski J, P€uhler A,Tauch A (2007) The IclR-type transcriptional repressor LtbR regulates the expression of
leucine and tryptophan biosynthesis genes in the amino acid producer Corynebacteriumglutamicum. J Bacteriol 189:2720–2733
Buckland BC, Lilly MD (1993) Fermentation: an overview. In: Stephanopoulos G (ed) Biotech-
nology, vol 3, 2nd edn, Bioprocessing. VCH Verlagsgesellschaft mbH, Weinheim, pp 7–22
Burkovski A (2008) Corynebacteria: genomics and molecular biology. Caister Academic, Norfolk
Burkovski A, Kr€amer R (2002) Bacterial amino acid transport proteins: occurrence, functions, and
significance for biotechnological applications. Appl Microbiol Biotechnol 58:265–274
Chinen A, Kozlov YI, Hara Y, Izui H, Yasueda H (2007) Innovative metabolic pathway design for
efficient L-glutamate production by suppressing CO2 emission. J Biosci Bioeng 103:262–269
Cramer A, Gerstmeir R, Schaffer S, Bott M, Eikmanns BJ (2006) Identification of RamA, a novel
LuxR-type regulator of genes involved in acetate metabolism of Corynebacterium glutamicum.J Bacteriol 188:2554–2567
Curis E, Crenn P, Cynober L (2007) Citrulline and the gut. Curr Opin Clin Nutr Metab Care
10:620–626
Dassler T, Maier T, Winterhalter C, B€ock A (2000) Identification of a major facilitator protein
from Escherichia coli involved in efflux of metabolites of the cysteine pathway. Mol Microbiol
36:1101–1112
Diesveld R, Tietze N, F€urst O, Reth A, Bathe B, Sahm H, Eggeling L (2008) Activity of exporters
of Escherichia coli in Corynebacterium glutamicum, and their use to increase L-threonine
production. J Mol Microbiol Biotechnol 16:198–207
Dong X, Quinn PJ, Wang X (2011) Metabolic engineering of Escherichia coli and Corynebacte-rium glutamicum for the production of L-threonine. Biotechnol Adv 29:11–23
Ebbighausen H, Weil B, Kr€amer R (1989) Transport of branched-chain amino acids in Coryne-bacterium glutamicum. Arch Microbiol 151:238–244
Eggeling L, Bott M (2005) Handbook of Corynebacterium glutamicum. CRC, Boca Raton, FLEggeling L, Oberle S, Sahm H (1998) Improved L-lysine yield with Corynebacterium glutamicum:
use of dapA resulting in increased flux combined with growth limitation. Appl Microbiol
Biotechnol 49:24–30
Einsele A (1978) Scaling-up bioreactors. Proc Biochem 7:1–14
Elisakova V, Patek M, Holatko J, Nesvera J, Leyval D, Goergen JL, Delaunay S (2005) Feedback-
resistant acetohydroxy acid synthase increases valine production in Corynebacteriumglutamicum. Appl Environ Microbiol 71:207–213
Engels V, Lindner SN, Wendisch VF (2008) The global repressor SugR controls expression of
genes of glycolysis and of the L-lactate dehydrogenase LdhA in Corynebacterium glutamicum.J Bacteriol 190:8033–8044
Figge R, Soucaille P, Barbier G, Bestel-Corre G, Boisart C, Chateau M (2009) Increasing
methionine yield. International Patent Application WO 2009/043803 A2
Flores N, Xiao J, Berry A, Bolivar F, Valle F (1996) Pathway engineering for the production of
aromatic compounds in Escherichia coli. Nat Biotechnol 14:620–623Follmann M, Becker M, Ochrombel I, Ott V, Kr€amer R, Marin K (2009a) Potassium transport in
Corynebacterium glutamicum is facilitated by the putative channel protein CglK, which is
essential for pH homeostasis and growth at acidic pH. J Bacteriol 191:2944–2952
Follmann M, Ochrombel I, Kr€amer R, Tr€otschel C, Poetsch A, R€uckert C, H€user A, Persicke M,
Seiferling D, Kalinowski J, Marin K (2009b) Functional genomics of pH homeostasis in
Corynebacterium glutamicum revealed novel links between pH response, oxidative stress,
iron homeostasis and methionine synthesis. BMC Genomics 10:621
Franke I, Resch A, Dassler T, Maier T, B€ock A (2003) YfiK from Escherichia coli promotes export
of O-acetylserine and cysteine. J Bacteriol 185:1161–1166
Fudou R, Jojima Y, Seto A, Yamada K, Kimura E, Nakamatsu T, Hiraishi A, Yamanaka S (2002)
Corynebacterium efficiens sp. nov., a glutamic-acid-producing species from soil and
vegetables. Int J Syst Evol Microbiol 52:1127–1131
138 M. Ikeda and S. Takeno

Gerstmeir R, Cramer A, Dangel P, Schaffer S, Eikmanns BJ (2004) RamB, a novel transcriptional
regulator of genes involved in acetate metabolism of Corynebacterium glutamicum. J Bacteriol186:2798–2809
Glansdorff N, Xu Y (2007) Microbial arginine biosynthesis: pathway, regulation and industrial
production. In: Wendisch VF (ed) Microbiology monographs, amino acid biosynthesis –
pathways, regulation and metabolic engineering. Springer, Berlin, pp 219–257
Gunji Y, Yasueda H (2006) Enhancement of L-lysine production in methylotroph Methylophilusmethylotrophus by introducing a mutant LysE exporter. J Biotechnol 127:1–13
Gutmann M, Hoischen C, Kr€amer R (1992) Carrier-mediated glutamate secretion by Corynebac-terium glutamicum under biotin limitation. Biochim Biophys Acta 1112:115–123
Haitani Y, Awano N, Yamazaki M, Wada M, Nakamori S, Takagi H (2006) Functional analysis of
L-serine O-acetyltransferase from Corynebacterium glutamicum. FEMS Microbiol Lett
255:156–163
H€anbler E, M€uller T, Jessberger N, V€olzke A, Plassmeier J, Kalinowski J, Kr€amer R, Burkovski A
(2007) FarR, a putative regulator of amino acid metabolism in Corynebacterium glutamicum.Appl Microbiol Biotechnol 76:625–632
Hashimoto S, Katsumata R (1993) Overproduction of alanine by Arthrobacter strains with
glucose-nonrepressible L-alanine dehydrogenase. Biotechnol Lett 15:1117–1122
Hashimoto S, Katsumata R (1998) L-Alanine fermentation by an alanine racemase deficient
mutant of the DL-alanine hyperproducing bacterium, Arthrobacter oxydans HAP-1. J Ferment
Bioeng 86:346–351
Hashimoto S, Katsumata R (1999) Mechanism of alanine hyperproduction by arthrobacteroxydans HAP-1: metabolic shift to fermentation under nongrowth aerobic conditions. Appl
Environ Microbiol 65:2781–2783
Hayashi M, Mizoguchi H, Ohnishi J, Mitsuhashi S, Yonetani Y, Hashimoto S, Ikeda M (2006a) A
leuC mutation leading to increased L-lysine production and rel-independent global expressionchanges in Corynebacterium glutamicum. Appl Microbiol Biotechnol 72:783–789
Hayashi M, Ohnishi J, Mitsuhashi S, Yonetani Y, Hashimoto S, Ikeda M (2006b) Transcriptome
analysis reveals global expression changes in an industrial L-lysine producer of Corynebacte-rium glutamicum. Biosci Biotechnol Biochem 70:546–550
Haynes JA, Britz ML (1990) The effect of growth conditions of Corynebacterium glutamicum on
the transformation frequency obtained by electroporation. J Gen Microbiol 136:255–263
Hermann T (2003) Industrial production of amino acids by coryneform bacteria. J Biotechnol
104:155–172
Hirao T, Nakano T, Azuma T, Sugimoto M, Nakanishi T (1989) L-lysine production in continuous
culture of an L-lysine hyper- producing mutant of Corynebacterium glutamicum. Appl
Microbiol Biotechnol 32:269–273
Holatko J, Elisakova V, Prouza M, Sobotka M, Nesvera J, Patek M (2009) Metabolic engineering
of the L-valine biosynthesis pathway in Corynebacterium glutamicum using promoter activity
modulation. J Biotechnol 139:203–210
Hols P, Kleerebezem M, Schanck AN, Ferain T, Hugenholtz J, Delcour J, de Vos WM (1999)
Conversion of Lactococcus lactis from homolactic to homoalanine fermentation through
metabolic engineering. Nat Biotechnol 17:588–592
Hwang BJ, Park SD, Kim Y, Kim P, Lee HS (2007) Biochemical analysis on the parallel pathways
of methionine biosynthesis in Corynebacterium glutamicum. J Microbiol Biotechnol
17:1010–1017
Ikeda M (2003) Amino acid production processes. In: Faurie R, Thommel J (eds) Adv Biochem
Eng Biotechnol, vol 79, Microbial production of L-amino acids. Springer, Berlin, pp 1–35
Ikeda M, Katsumata R (1992) Metabolic engineering to produce tyrosine or phenylalanine in a
tryptophan-producing Corynebacterium glutamicum strain. Appl Environ Microbiol
58:781–785
Ikeda M, Katsumata R (1994) Transport of aromatic amino acids and its influence on overproduc-
tion of the amino acids in Corynebacterium glutamicum. J Ferment Bioeng 78:420–425
Amino Acid Production by Corynebacterium glutamicum 139

Ikeda M, Katsumata R (1995) Tryptophan production by transport mutants of Corynebacteriumglutamicum. Biosci Biotechnol Biochem 59:1600–1602
Ikeda M, Katsumata R (1998) A novel system with positive selection for the chromosomal
integration of replicative plasmid DNA in Corynebacterium glutamicum. Microbiology
144:1863–1868
Ikeda M, Katsumata R (1999) Hyperproduction of tryptophan by Corynebacterium glutamicumwith the modified pentose phosphate pathway. Appl Environ Microbiol 65:2497–2502
Ikeda M, Nakagawa S (2003) The Corynebacterium glutamicum genome: features and impacts on
biotechnological process. Appl Microbiol Biotechnol 62:99–109
Ikeda M, Nakanishi K, Kino K, Katsumata R (1994) Fermentative production of tryptophan by a
stable recombinant strain of Corynebacterium glutamicum with a modified serine-biosynthetic
pathway. Biosci Biotechnol Biochem 58:674–678
Ikeda M, Okamoto K, Katsumata R (1999) Cloning of the transketolase gene and the effect of its
dosage on aromatic amino acid production in Corynebacterium glutamicum. Appl Microbiol
Biotechnol 51:201–206
Ikeda M, Ohnishi J, Mitsuhashi S (2005) Genome breeding of an amino acid-producing Coryne-bacterium glutamicum mutant. In: Barredo JLS (ed) Microbial processes and products.
Humana, Totowa, pp 179–189
Ikeda M, Ohnishi J, Hayashi M, Mitsuhashi S (2006) A genome-based approach to create a
minimally mutated Corynebacterium glutamicum strain for efficient L-lysine production.
J Ind Microbiol Biotechnol 33:610–615
Ikeda M, Baba M, Tsukamoto N, Komatsu T, Mitsuhashi S, Takeno S (2009a) Elucidation of
genes relevant to the microaerobic growth of Corynebacterium glutamicum. Biosci BiotechnolBiochem 73:2806–2808
Ikeda M, Mitsuhashi S, Tanaka K, Hayashi M (2009b) Reengineering of a Corynebacteriumglutamicum L-arginine and L-citrulline producer. Appl Environ Microbiol 75:1635–1641
Ikeda M, Takeno S, Mizuno Y, Mitsuhashi S (2010) Process for producing useful substance.
International Patent Application WO 2010/024267 A1
Ikeda M, Mizuno Y, Awane S, Hayashi M, Mitsuhashi S, Takeno S (2011) Identification and
application of a different glucose uptake system that functions as an alternative to the
phosphotransferase system in Corynebacterium glutamicum. Appl Microbiol Biotechnol
90:1443–1451
Ishizaki A, Takasaki S, Furuta Y (1993) Cell recycled fermentation of glutamate using a novel
cross-flow filtration system with constant air supply. J Ferment Bioeng 76:316–320
Jojima T, Fujii M, Mori E, Inui M, Yukawa H (2010) Engineering of sugar metabolism of
Corynebacterium glutamicum for production of amino acid L-alanine under oxygen depriva-
tion. Appl Microbiol Biotechnol 87:159–165
Kabus A, Georgi T, Wendisch VF, Bott M (2007a) Expression of the Escherichia coli pntAB genes
encoding a membrane-bound transhydrogenase in Corynebacterium glutamicum improves
L-lysine formation. Appl Microbiol Biotechnol 75:47–53
Kabus A, Niebisch A, Bott M (2007b) Role of cytochrome bd oxidase from Corynebacteriumglutamicum in growth and lysine production. Appl Environ Microbiol 73:861–868
Kalinowski J, Bathe B, Bartels D, Bischoff N, Bott M, Burkovski A, Dusch N, Eggeling L,
Eikmanns BJ, Gaigalat L, Goesmann A, Hartmann M, Huthmacher K, Kr€amer R, Linke B,
McHardy AC,Meyer F,M€ockel B, PfefferleW, P€uhler A, Rey DA, R€uckert C, Rupp O, SahmH,
Wendisch VF, Wiegr€abe I, Tauch A (2003) The complete Corynebacterium glutamicum ATCC
13032 genome sequence and its impact on the production of L-aspartate-derived amino acids and
vitamins. J Biotechnol 104:5–25
Kallio PT, Kim DJ, Tsai PS, Bailey JE (1994) Intracellular expression of Vitreoscilla hemoglobin
alters Escherichia coli energy metabolism under oxygen-limited conditions. Eur J Biochem
219:201–208
Kaneko H, Sakaguchi K (1979) Fusion of protoplasts and genetic recombination of
Brevibacterium flavum. Agric Biol Chem 43:1007–1013
Karasawa M, Tosaka O, Ikeda S, Yoshii H (1986) Application of protoplast fusion to the
development of L-threonine and L-lysine producers. Agric Biol Chem 50:339–346
140 M. Ikeda and S. Takeno

Kase H, Nakayama K (1974) Mechanism of L-threonine and L-lysine production by analog-
resistant mutants of Corynebacterium glutamicum. Agric Biol Chem 38:993–1000
Katsumata R, Hashimoto S (1996) Process for producing alanine. US Patent 5 559 016
Katsumata R, Ikeda M (1993) Hyperproduction of tryptophan in Corynebacterium glutamicum by
pathway engineering. Biotechnology 11:921–925
Katsumata R, Ozaki A, Oka T, Furuya A (1984) Protoplast transformation of glutamate-producing
bacteria with plasmid DNA. J Bacteriol 159:306–311
Katsumata R, Mizukami T, Kikuchi Y, Kino K (1986) Threonine production by the lysine
producing strain of Corynebacterium glutamicum with amplified threonine biosynthetic
operon. In: Alacevic M, Hranueli D, Toman Z (eds) Genetics of industrial microorganisms.
Pliva Publishing Co., Yugoslavia, pp 217–226
Kawaguchi H, Vertes AA, Okino S, Inui M, Yukawa H (2006) Engineering of a xylose metabolic
pathway in Corynebacterium glutamicum. Appl Environ Microbiol 72:3418–3428
Kawaguchi H, Sasaki M, Vertes AA, Inui M, Yukawa H (2008) Engineering of an L-arabinose
metabolic pathway in Corynebacterium glutamicum. Appl Microbiol Biotechnol
77:1053–1062
Kawahara Y, Takahashi-Fuke K, Shimizu E, Nakamatsu T, Nakamori S (1997) Relationship
between the glutamate production and the activity of 2-oxoglutarate dehydrogenase in
Brevibacterium lactofermentum. Biosci Biotechnol Biochem 61:1109–1112
Kennerknecht N, Sahm H, Yen MR, Patek M, Saier MH Jr, Eggeling L (2002) Export of
L-isoleucine from Corynebacterium glutamicum: a two-gene-encoded member of a new
translocator family. J Bacteriol 184:3947–3956
Kiefer P, Heinzle E, Zelder O, Wittmann C (2004) Comparative metabolic flux analysis of lysine-
producing Corynebacterium glutamicum cultured on glucose or fructose. Appl Environ
Microbiol 70:229–239
Kim HJ, Kim TH, Kim Y, Lee HS (2004) Identification and characterization of glxR, a gene
involved in regulation of glyoxylate bypass in Corynebacterium glutamicum. J Bacteriol
186:3453–3460
Kim TH, Kim HJ, Park JS, Kim Y, Kim P, Lee HS (2005a) Functional analysis of sigH expression
in Corynebacterium glutamicum. Biochem Biophys Res Commun 331:1542–1547
Kim TH, Park JS, Kim HJ, Kim Y, Kim P, Lee HS (2005b) The whcE gene of Corynebacteriumglutamicum is important for survival following heat and oxidative stress. Biochem Biophys
Res Commun 337:757–764
Kim J, Fukuda H, Hirasawa T, Nagahisa K, Nagai K, Wachi M, Shimizu H (2009a) Requirement
of de novo synthesis of the OdhI protein in penicillin-induced glutamate production by
Corynebacterium glutamicum. Appl Microbiol Biotechnol. doi:10.1007/s00253-009-2360-6
Kim J, Hirasawa T, Sato Y, Nagahisa K, Furusawa C, Shimizu H (2009b) Effect of odhAoverexpression and odhA antisense RNA expression on Tween-40-triggered glutamate produc-
tion by Corynebacterium glutamicum. Appl Microbiol Biotechnol 81:1097–1106
Kimura E (2003) Metabolic engineering of glutamate production. In: Faurie R, Thommel J (eds)
Adv Biochem Eng Biotechnol, vol 79, Microbial production of L-amino acids. Springer,
Berlin, pp 37–58
Kimura E (2005) L-Glutamate production. In: Eggeling L, Bott M (eds) Handbook of Corynebac-terium glutamicum. CRC, Boca Raton, FL, pp 439–463
Kimura E, Yagoshi C, Kawahara Y, Ohsumi T, Nakamatsu T, Tokuda H (1999) Glutamate
overproduction in Corynebacterium glutamicum triggered by a decrease in the level of a
complex comprising DtsR and a biotin-containing subunit. Biosci Biotechnol Biochem
63:1274–1278
Kinoshita S, Nakayama K (1978) Amino acids. In: Rose AH (ed) Primary products of metabolism.
Academic, London, pp 209–261
Kinoshita S, Udaka S, Shimono M (1957) Studies on the amino acid fermentation, part I.
Production of L-glutamic acid by various microorganisms. J Gen Appl Microbiol 3:193–205
Amino Acid Production by Corynebacterium glutamicum 141

Kitai A (1972) Alanine. In: Yamada K, Kinoshita S, Tsunoda T, Aida K (eds) The microbial
production of amino acids. John Wiley and Sons, New York, NY, pp 325–337
Kjeldsen KR, Nielsen J (2009) In silico genome-scale reconstruction and validation of the
Corynebacterium glutamicum metabolic network. Biotechnol Bioeng 102:583–597
Koch DJ, R€uckert C, Albersmeier A, H€user AT, Tauch A, P€uhler A, Kalinowski J (2005) Thetranscriptional regulator SsuR activates expression of the Corynebacterium glutamicumsulphonate utilization genes in the absence of sulphate. Mol Microbiol 58:480–494
Krings E, Krumbach K, Bathe B, Kelle R, Wendisch VF, Sahm H, Eggeling L (2006) Characteri-
zation of myo-inositol utilization by Corynebacterium glutamicum: the stimulon, identification
of transporters, and influence on L-lysine formation. J Bacteriol 188:8054–8061
Kr€omer JO, Sorgenfrei O, Klopprogge K, Heinzle E, Wittmann C (2004) In-depth profiling of
lysine-producing Corynebacterium glutamicum by combined analysis of the transcriptome,
metabolome, and fluxome. J Bacteriol 186:1769–1784
Kr€omer JO, Wittmann C, Schr€oder H, Heinzle E (2006) Metabolic pathway analysis for rational
design of L-methionine production by Eschrichia coli and Corynebacterium glutamicum.Metab Eng 8:353–369
Kr€omer JO, Bolten CJ, Heinzle E, Schr€oder H, Wittmann C (2008) Physiological response of
Corynebacterium glutamicum to oxidative stress induced by deletion of the transcriptional
repressor McbR. Microbiology 154:3917–3930
Lange C, Rittmann D, Wendisch VF, Bott M, Sahm H (2003) Global expression profiling and
physiological characterization of Corynebacterium glutamicum grown in the presence of
L-valine. Appl Environ Microbiol 69:2521–2532
Lee HS, Hwang BJ (2003) Methionine biosynthesis and its regulation in Corynebacteriumglutamicum: parallel pathways of transsulfuration and direct sulfhydrylation. Appl Microbiol
Biotechnol 62:459–467
Lee SY, Shin HS, Park JS, Kim YH, Min J (2010) Proline reduces the binding of transcriptional
regulator ArgR to upstream of argB in Corynebacterium glutamicum. Appl Microbiol
Biotechnol 86:235–242
Leuchtenberger W (1996) Amino acids – technical production and use. In: Roehr M (ed)
Biotechnology, vol 6, 2nd edn, Products of primary metabolism. VCH Verlagsgesellschaft
mbH, Weinheim, pp 465–502
Leuchtenberger W, Huthmacher K, Drauz K (2005) Biotechnological production of amino acids
and derivatives: current status and prospects. Appl Microbiol Biotechnol 69:1–8
Li L, Wada M, Yokota A (2007) A comparative proteomic approach to understand the adaptations
of an H+-ATPase-defective mutant of Corynebacterium glutamicum ATCC14067 to energy
deficiencies. Proteomics 7:3348–3357
Liu Q, Zhang J, Wei XX, Ouyang SP, Wu Q, Chen GQ (2008) Microbial production of
L-glutamate and L-glutamine by recombinant Corynebacterium glutamicum harboring
Vitreoscilla hemoglobin gene vgb. Appl Microbiol Biotechnol 77:1297–1304
Mampel J, Schr€oder H, Haefner S, Sauer U (2005) Single-gene knockout of a novel regulatory
element confers ethionine resistance and elevates methionine production in Corynebacteriumglutamicum. Appl Microbiol Biotechnol 68:228–236
Marienhagen J, Eggeling L (2008) Metabolic function of Corynebacterium glutamicumaminotransferases AlaT and AvtA and impact on L-valine production. Appl Environ Microbiol
74:7457–7462
Marx A, Hans S, Mockel B, Bathe B, de Graaf AA (2003) Metabolic phenotype of phosphoglucose
isomerase mutants of Corynebacterium glutamicum. J Biotechnol 104:185–197McWilliams A (2010) Biotechnologies for medical applications: global markets. Report code:
BIO072A, BCC Research (http://www.bccresearch.com)
Mitsuhashi S, Hayashi M, Ohnishi J, Ikeda M (2006) Disruption of malate: quinone oxidoreduc-
tase increases L-lysine production by Corynebacterium glutamicum. Biosci Biotechnol
Biochem 70:2803–2806
142 M. Ikeda and S. Takeno

Miwa K, Matsui K, Terabe M, Ito K, Ishida M, Takagi H, Nakamori S, Sano K (1985) Construc-
tion of novel shuttle vector and a cosmid vector for the glutamic acid-producing bacteria
Brevibacterium lactofermentum and Corynebacterium glutamicum. Gene 39:281–286M€ockel B, Pfefferle W, Huthmacher K, R€uckert C, Kalinowski J, P€uhler A, Binder M, Greissinger
D, Thierbach G (2002) Nucleotide sequences which code for the metY gene. International
Patent Application WO 02/18613
M€oker N, Kr€amer J, Unden G, Kr€amer R, Morbach S (2007) In vitro analysis of the two-
component system MtrB-MtrA from Corynebacterium glutamicum. J Bacteriol
189:3645–3649
Moon MW, Kim HJ, Oh TK, Shin CS, Lee JS, Kim SJ, Lee JK (2005) Analyses of enzyme II gene
mutants for sugar transport and heterologous expression of fructokinase gene in Corynebacte-rium glutamicum ATCC 13032. FEMS Microbiol Lett 244:259–266
Morbach S, Sahm H, Eggeling L (1996) L-Isoleucine production with Corynebacteriumglutamicum: further flux increase and limitation of export. Appl Environ Microbiol
62:4345–4351
Nakamura J, Hirano S, Ito H, Wachi M (2007) Mutations of the Corynebacterium glutamicumNCgl1221 gene, encoding a mechanosensitive channel homolog, induce L-glutamic acid
production. Appl Environ Microbiol 73:4491–4498
Nakayama K, Kitada S, Kinoshita S (1961) Studies on lysine fermentation. I. The control
mechanism on lysine accumulation by homoserine and threonine. J Gen Appl Microbiol
7:145–154
Niebisch A, Kabus A, Schultz C, Weil B, Bott M (2006) Corynebacterial protein kinase G controls
2-oxoglutarate dehydrogenase activity via the phosphorylation status of the OdhI protein.
J Biol Chem 281:12300–12307
Nishimura T, Vertes AA, Shinoda Y, Inui M, Yukawa H (2007) Anaerobic growth of Corynebac-terium glutamicum using nitrate as a terminal electron acceptor. Appl Microbiol Biotechnol
75:889–897
Nishio Y, Nakamura Y, Kawarabayasi Y, Usuda Y, Kimura E, Sugimoto S, Matsui K, Yamagishi
A, Kikuchi H, Ikeo K, Gojobori T (2003) Comparative complete genome sequence analysis of
the amino acid replacements responsible for the thermostability of Corynebacterium efficiens.Genome Res 13:1572–1579
Nottebrock D, Meyer U, Kr€amer R, Morbach S (2003) Molecular and biochemical characteriza-
tion of mechanosensitive channels in Corynebacterium glutamicum. FEMS Microbiol Lett
218:305–309
Ohnishi J, Ikeda M (2006) Comparisons of potentials for L-lysine production among different
Corynebacterium glutamicum strains. Biosci Biotechnol Biochem 70:1017–1020
Ohnishi J, Mitsuhashi S, Hayashi M, Ando S, Yokoi H, Ochiai K, Ikeda M (2002) A novel
methodology employing Corynebacterium glutamicum genome information to generate a new
L-lysine-producing mutant. Appl Microbiol Biotechnol 58:217–223
Ohnishi J, Hayashi M, Mitsuhashi S, Ikeda M (2003) Efficient 40�C fermentation of L-lysine by a
new Corynebacterium glutamicum mutant developed by genome breeding. Appl Microbiol
Biotechnol 62:69–75
Ohnishi J, Katahira R, Mitsuhashi S, Kakita S, Ikeda M (2005) A novel gnd mutation leading to
increased L-lysine production in Corynebacterium glutamicum. FEMS Microbiol Lett
242:265–274
Ozaki A, Katsumata R, Oka T, Furuya A (1985) Cloning of the genes concerned in phenylalanine
biosynthesis in Corynebacterium glutamicum and its application to breeding of a phenylalanine
producing strain. Agric Biol Chem 49:2925–2930
Park JH, Lee SY (2010) Fermentative production of branched chain amino acids: a focus on
metabolic engineering. Appl Microbiol Biotechnol 85:491–506
Park SD, Lee JY, Sim SY, Kim Y, Lee HS (2007) Characteristics of methionine production by an
engineered Corynebacterium glutamicum strain. Metab Eng 9:327–336
Amino Acid Production by Corynebacterium glutamicum 143

Patek M (2007) Branched-chain amino acids. In: Wendisch VF (ed) Microbiology monographs,
amino acid biosynthesis – pathways, regulation and metabolic engineering. Springer, Berlin,
pp 129–162
Peter H, Burkovski A, Kr€amer R (1996) Isolation, characterization, and expression of the Coryne-bacterium glutamicum betP gene, encoding the transport system for the compatible solute
glycine betaine. J Bacteriol 178:5229–5234
Peter H, Weil B, Burkovski A, Kr€amer R, Morbach S (1998) Corynebacterium glutamicum is
equipped with four secondary carriers for compatible solutes: identification, sequencing, and
characterization of the proline/ectoine uptake system, ProP, and the ectoine/proline/glycine
betaine carrier, EctP. J Bacteriol 180:6005–6012
Petersen S, Mack C, de Graaf AA, Riedel C, Eikmanns BJ, Sahm H (2001) Metabolic
consequences of altered phosphoenolpyruvate carboxykinase activity in Corynebacteriumglutamicum reveal anaplerotic mechanisms in vivo. Metab Eng 3:344–361
Peters-Wendisch PG, Schiel B, Wendisch VF, Katsoulidis E, M€ockel B, Sahm H, Eikmanns BJ
(2001) Pyruvate carboxylase is a major bottleneck for glutamate and lysine production by
Corynebacterium glutamicum. J Mol Microbiol Biotechnol 3:295–300
Peters-Wendisch P, Stolz M, Etterich H, Kennerknecht N, Sahm H, Eggeling L (2005) Metabolic
engineering of Corynebacterium glutamicum for L-serine production. Appl Environ Microbiol
71:7139–7144
Pfefferle W, M€ockel B, Bathe B, Marx A (2003) Biotechnological manufacture of lysine. In:
Faurie R, Thommel J (eds) Adv Biochem Eng Biotechnol, vol 79, Microbial production of
L-amino acids. Springer, Berlin, pp 59–112
Radmacher E, Vaitsikova A, Burger U, Krumbach K, Sahm H, Eggeling L (2002) Linking central
metabolism with increased pathway flux: L-valine accumulation by Corynebacteriumglutamicum. Appl Environ Microbiol 68:2246–2250
Rey DA, P€uhler A, Kalinowski J (2003) The putative transcriptional repressor McbR, member of
the TetR-family, is involved in the regulation of the metabolic network directing the synthesis
of sulfur containing amino acids in Corynebacterium glutamicum. J Biotechnol 103:51–65Rey DA, Nentwich SS, Koch DJ, R€uckert C, P€uhler A, Tauch A, Kalinowski J (2005) The McbR
repressor modulated by the effector substance S-adenosylhomocysteine controls directly the
transcription of a regulon involved in sulphur metabolism of Corynebacterium glutamicumATCC 13032. Mol Microbiol 56:871–887
Riedel C, Rittmann D, Dangel P, M€ockel B, Sahm H, Eikmanns BJ (2001) Characterization,
expression, and inactivation of the phosphoenolpyruvate carboxykinase gene from Corynebac-terium glutamicum and significance of the enzyme for growth and amino acid production.
J Mol Microbiol Biotechnol 3:573–583
Rieping M, Hermann T (2007) L-Threonine. In: Wendisch VF (ed) Microbiology monographs,
amino acid biosynthesis – pathways, regulation and metabolic engineering. Springer, Berlin,
pp 71–92
Rittmann D, Lindner SN, Wendisch VF (2008) Engineering of a glycerol utilization pathway for
amino acid production by Corynebacterium glutamicum. Appl Environ Microbiol
74:6216–6222
Sano K, Shiio I (1971) Microbial production of L-lysine. IV. Selection of lysine-producing
mutants from Brevibacterium flavum by detecting threonine sensitivity or halo-forming
method. J Gen Appl Microbiol 17:97–113
Sano K, Yokozeki K, Tamura F, Yasuda N, Noda I, Mitsugi K (1977) Microbial conversion of
DL-2-amino-D2thiazoline-4-carboxylic acid to L-cysteine and L-cystine: screening of
microorganisms and identification of products. Appl Environ Microbiol 34:806–810
Santamaria R, Gil JA, Mesas JM, Martin JF (1984) Characterization of an endogenous plasmid and
development of cloning vectors and a transformation system in Brevibacteriumlactofermentum. J Gen Microbiol 130:2237–2246
Sato H, Orishimo K, Shirai T, Hirasawa T, Nagahisa K, Shimizu H, Wachi M (2008) Distinct roles
of two anaplerotic pathways in glutamate production induced by biotin limitation in Coryne-bacterium glutamicum. J Biosci Bioeng 106:51–58
144 M. Ikeda and S. Takeno

Sawada K, Zen-In S, Wada M, Yokota A (2010) Metabolic changes in a pyruvate kinase gene
deletion mutant of Corynebacterium glutamicum ATCC 13032. Metab Eng 12:401–407
Schaaf S, Bott M (2007) Target genes and DNA-binding sites of the response regulator PhoR from
Corynebacterium glutamicum. J Bacteriol 189:5002–5011Sch€afer A, Kalinowski J, Simon R, Seep-Feldhaus A-H, P€uhler A (1990) High-frequency conjugal
plasmid transfer from gram-negative Escherichia coli to various gram-positive coryneform
bacteria. J Bacteriol 172:1663–1666
Schultz C, Niebisch A, Gebel L, Bott M (2007) Glutamate production by Corynebacteriumglutamicum: dependence on the oxoglutarate dehydrogenase inhibitor protein OdhI and proteinkinase PknG. Appl Microbiol Biotechnol 76:691–700
Schwarzer A, P€uhler A (1991) Manipulation of Corynebacterium glutamicum by gene disruption
and replacement. Biotechnology 9:84–87
Seibold G, Auchter M, Berens S, Kalinowski J, Eikmanns BJ (2006) Utilization of soluble starch
by a recombinant Corynebacterium glutamicum strain: growth and lysine production.
J Biotechnol 124:381–391
Shibatani T, Kakimoto T, Chibata I (1979) Stimulation of L-asparate b-decarboxylase formation
by L-glutamate in Pseudomonas dacunhae and improved production of L-alanine. Appl
Environ Microbiol 38:359–364
Shiio I, Miyajima R (1969) Concerted inhibition and its reversal by end products of aspartate
kinase in Brevibacterium flavum. J Biochem 65:849–859
Shiio I, Toride Y, Sugimoto S (1984) Production of lysine by pyruvate dehydrogenase mutants of
Brevibacterium flavum. Agric Biol Chem 48:3091–3098
Shimomura Y, Yamamoto Y, Bajotto G, Sato J, Murakami T, Shimomura N, Kobayashi H,
Mawatari K (2006) Nutraceutical effects of branched-chain amino acids on skeletal muscle.
J Nutr 136:529S–532S
Shingu H, Terui G (1971) Studies on process of glutamic acid fermentation at the enzyme level.
Part I. On the change of a-ketoglutaric acid dehydrogenase in the course of culture. J Ferment
Technol 49:400–405
Simic P, Sahm H, Eggeling L (2001) L-Threonine export: use of peptides to identify a new
translocator from Corynebacterium glutamicum. J Bacteriol 183:5317–5324Simic P, Willuhn J, Sahm H, Eggeling L (2002) Identification of glyA (encoding serine
hydroxymethyltransferase) and its use together with the exporter ThrE to increase L-threonine
accumulation by Corynebacterium glutamicum. Appl Environ Microbiol 68:3321–3327
Sindelar G, Wendisch VF (2007) Improving lysine production by Corynebacterium glutamicumthrough DNA microarray-based identification of novel target genes. Appl Microbiol
Biotechnol 76:677–689
Smith GM, Lee SA, Reilly KC, Eiteman MA, Altman E (2006) Fedbatch two-phase production of
alanine by a metabolically engineered Escherichia coli. Biotechnol Lett 28:1695–1700Sprenger GA (2007) Aromatic amino acids. In: Wendisch VF (ed) Microbiology monographs,
amino acid biosynthesis – pathways, regulation and metabolic engineering. Springer, Berlin,
pp 93–127
Steger R, Weinand M, Kr€amer R, Morbach S (2004) LcoP, an osmoregulated betaine/ectoine
uptake system from Corynebacterium glutamicum. FEBS Lett 573:155–160
Stolz M, Peters-Wendisch P, Etterich H, Gerharz T, Faurie R, Sahm H, Fersterra H, Eggeling L
(2007) Reduced folate supply as a key to enhanced L-serine production by Corynebacteriumglutamicum. Appl Environ Microbiol 73:750–755
Strelkov S, von Elstermann M, Schomburg D (2004) Comprehensive analysis of metabolites in
Corynebacterium glutamicum by gas chromatography/mass spectrometry. Biol Chem
385:853–861
Takeno S, Ohnishi J, Komatsu T, Masaki T, Sen K, Ikeda M (2007) Anaerobic growth and
potential for amino acid production by nitrate respiration in Corynebacterium glutamicum.Appl Microbiol Biotechnol 75:1173–1182
Amino Acid Production by Corynebacterium glutamicum 145

Takeno S, Nakamura M, Fukai R, Ohnishi J, Ikeda M (2008) The Cgl1281-encoding putative
transporter of the cation diffusion facilitator family is responsible for alkali-tolerance in
Corynebacterium glutamicum. Arch Microbiol 190:531–538
Takeno S, Murata R, Kobayashi R, Mitsuhashi S, Ikeda M (2010) Engineering of Corynebacte-rium glutamicum with an NADPH-generating glycolytic pathway for L-lysine production.
Appl Environ Microbiol 76:7154–7160
Tateno T, Fukuda H, Kondo A (2007) Production of L-lysine from starch by Corynebacteriumglutamicum displaying a-amylase on its cell surface. Appl Microbiol Biotechnol 74:1213–1220
Tr€otschel C, Deutenberg D, Bathe B, Burkovski A, Kr€amer R (2005) Characterization of methio-
nine export in Corynebacterium glutamicum. J Bacteriol 187:3786–3794Udaka S (1960) Screening method for microorganisms accumulating metabolites and its use in the
isolation of Micrococcus glutamicus. J Bacteriol 79:754–755Udaka S (2008) The discovery of Corynebacterium glutamicum and birth of amino acid fermenta-
tion industry in Japan. In: Burkovski A (ed) Corynebacteria: genomics and molecular biology.
Caister Academic, Norwich, pp 1–6
Uhlenbusch I, Sahm H, Sprenger GA (1991) Expression of an L-alanine dehydrogenase gene in
Zymomonas mobilis and excretion of L-alanine. Appl Environ Microbiol 57:1360–1366
Utagawa T (2004) Arginine metabolism: enzymology, nutrition, and clinical significance. J Nutr
134:2854S–2857S
van der Rest ME, Lange C, Molenaar D (1999) A heat shock following electroporation of
Corynebacterium glutamicum with xenogeneic plasmid DNA. Appl Microbiol Biotechnol
52:541–545
Vrljic M, Sahm H, Eggeling L (1996) A new type of transporter with a new type of cellular
function: L-lysine export from Corynebacterium glutamicum. Mol Microbiol 22:815–826
Wada M, Takagi H (2006) Metabolic pathways and biotechnological production of L-cysteine.
Appl Microbiol Biotechnol 73:48–54
Wada M, Awano N, Haisa K, Takagi H, Nakamori S (2002) Purification, characterization and
identification of cysteine desulfhydrase ofCorynebacterium glutamicum, and its relationship tocysteine production. FEMS Microbiol Lett 217:103–107
Wada M, Hijikata N, Aoki R, Takesue N, Yokota A (2008) Enhanced valine production in
Corynebacterium glutamicum with defective H+-ATPase and C-terminal truncated
acetohydroxyacid synthase. Biosci Biotechnol Biochem 72:2959–2965
Webster DA (1987) Structure and function of bacterial hemoglobin and related proteins. In:
Eichhorn GC, Marzilli LG (eds) Advances in inorganic chemistry. Elsevier, New York, NY,
pp 245–265
Wendisch VF (2007) Microbiology monographs, amino acid biosynthesis – pathways, regulation
and metabolic engineering. Springer, Berlin
Wendisch VF, Bott M, Kalinowski J, Oldiges M, Wiechert W (2006) Emerging Corynebacteriumglutamicum systems biology. J Biotechnol 124:74–92
Wennerhold J, Bott M (2006) The DtxR regulon of Corynebacterium glutamicum. J Bacteriol
188:2907–2918
Willis LB, Lessard PA, Sinskey AJ (2005) Synthesis of L-threonine and branched-chain amino
acids. In: Eggeling L, Bott M (eds) Handbook of Corynebacterium glutamicum. CRC, BocaRaton, FL, pp 511–531
Wittmann C, Heinzle E (2002) Genealogy profiling through strain improvement by using meta-
bolic network analysis: metabolic flux genealogy of several generations of lysine-producing
Corynebacteria. Appl Environ Microbiol 68:5843–5859
Yao W, Chu C, Deng X, Zhang Y, Liu M, Zheng P, Sun Z (2009) Display of a-amylase on the
surface of Corynebacterium glutamicum cells by using NCgl1221 as the anchoring protein, and
production of glutamate from starch. Arch Microbiol 191:751–759
Yoshihama M, Higashiro K, Rao EA, Akedo M, Shanabruch WG, Folletie MT, Walker GC,
Sinskey AJ (1985) Cloning vector system for Corynebacterium glutamicum. J Bacteriol
162:591–597
146 M. Ikeda and S. Takeno

Yukawa H, Omumasaba CA, Nonaka H, Kos P, Okai N, Suzuki N, Suda M, Tsuge Y, Watanabe J,
Ikeda Y, Vertes AA, Inui M (2007) Comparative analysis of the Corynebacterium glutamicumgroup and complete genome sequence of strain R. Microbiology 153:1042–1058
Zhang L, Li Y, Wang Z, Xia Y, Chen W, Tang K (2007a) Recent developments and future
prospects of Vitreoscilla hemoglobin application in metabolic engineering. Biotechnol Adv
25:123–136
Zhang X, Jantama K, Moore JC, Shanmugam KT, Ingram LO (2007b) Production of L-alanine by
metabolically engineered Escherichia coli. Appl Microbiol Biotechnol 77:355–366
Amino Acid Production by Corynebacterium glutamicum 147





























