Embed Size (px)
Citation preview

Plant Cell Reports (1984) 3: 193-195 Plant Cell Reports © Springer-Verlag t984
Amino acid pools and protein synthesis in germinating maize embryos
Raul Aguilar and Estela Sanchez de Jiminez
Plant Biochemistry Department, Facultad de Quimica, Universidad Nacionat Autonoma de Mexico, 04510, Mexico, DF
Received February 17, 1984/Revised version received August 1, 1984 - Communicated by O. L, Gamborg
ABSTRACT Amino acid pools were analyzed in maize
(Zea mays L.) axes germinated fo r O, 6 and 24 h. Prote ins synthes ized at e a r l y (0-6 h) and l a t e (18-24 h) stages were charac te r i zed by gel e l e c t r o p h o r e s i s and f luorography a f - t e r e i t h e r 14C-leucine or 14C- lys ine pulses. An increase of amino acid i nco rpo ra t i on af ter 18-24 h was nbserved, as wel l as changes in the p ro te in pa t te rns of the corresponding f luorographs . Ana lys is of the endogenous ami- no acid pools showed major changes in con- tents of p r o l i n e , a lan ine , i s o l e u c i n e , v a l i - ne, l euc ine and l y s i n e . A s e l e c t i v e increase of l y s i n e i nco rpo ra t i on i n to p ro te ins dur ing the l a t e stage was detected.
INTRODUCTION Cereal embryos r e - e s t a b l i s h metabo l ic ac-
t i v i t y f o l l o w i n g i m b i b i t i o n (Bewley and Black 1978). Pro te in syn thes is is one of the major molecular events tak ing place in t h i s per iod (Brooker et a l . 1977). Several authors have shown tha t the p ro te ins synthes ized during germinat ion are e s s e n t i a l f o r f u r t h e r growth and p l a n t l e t development (Jendr isak 1980, Datta et a l . 1983). Among these, the r i boso- mal p ro te ins have been i d e n t i f i e d as being synthes ized e a r l y dur ing wheat germinat ion (Lane and Tumait is-Kennedy 1981), wh i le h i s - tones seem to be produced l a t e r in maize em- bryos (Van de Walle and Del tour 1974). On the other hand, there is c o n f l i c t i n g in forma- t ion regard ing the s i m i l a r i t y or d i f f e r e n c e s between e a r l y and l a t e p ro te ins produced dur- ing germinat ion (Brooker et a l . 1978, Caers et a l . 1979). These c o n t r a d i c t o r y observa- t i ons might be the r e s u l t of the s p e c i f i c developmental programs fo r RNA synthes is that seem to cha rac te r i ze d i f f e r e n t kinds of seeds (Van de Walle et a l . 1983). There fore , data obtained in one species might not apply to another. Furthermore, d i f f e r e n c e s in the en- dogenous amino acid pools might regu la te the ra te of p ro te in syn thes is dur ing seed germi- nat ion (Marsh et a l . 1982).
1Under the Academic Colaborat ion Program be- tween the Univers idad Nacional Aut6noma de MExico and the Colegio de Postgraduados, Chapingo, MExico.
The aim of the present paper is to i n q u i - re about s p e c i f i c changes in p ro te in synthe- s is tak ing place in maize embryos concomitant to the germinat ion process and t h e i r r e l a t i o n to t h e i r f ree amino acid contents . Two stages of germinat ion were chosen fo r t h i s purpose: 0-6 h, when metabol ic a c t i v i t y is beg inn ing, and 18-24 h, before r a d i c l e p ro tus ion is ob- served (23.5 h average) and c e l l d i v i s i o n is i n i t i a t e d .
MATERIALS AND METHODS A hybr id v a r i e t y of maize (Zea mays L.)
(H-30) donated by Dr. Joaqu~n Or t i z -Cereceres Colegio de Postgraduados, Chapingo, M~xico, was the source of the b i o l o g i c a l m a t e r i a l . Embryonic axes -w i thou t scu te l lum- were ex- cised from dry seeds and used in a l l the ex- per iments. Incubat ion system. Batches of I00 to 150 mg of axes were incubated at 25°C in an incuba- t i on medium (50 mM T r i s -HC l , pH 7.6, 50 mM KCI, 10 mM MgCI2 and 2% sucrose) . A m ix tu re made of 0.03 mM of each of the f o l l o w i n g ami- no acids was suppl ied to the incubat ion sys- tem, except f o r the experiments where the i n - t e rna l amino acid pools were determined: a l a - n ine, v a l i n e , p r o l i n e , a r g i n i n e , g l y c i n e , i s o l e u c i n e , th reon ine , se r i ne , t ryp tophan, c y s t e i n e , g lu tamate, aspa r t a te , g lu tamine, asparagine, t y r o s i n e , h i s t i d i n e , meth ion ine, pheny la lan ine , and e i t h e r leuc ine or l y s i n e depending on the 14C-amino acid supp l ied . 20 ~Ci of e i t h e r 14C-leucine (314 mCi/mmol) or 14C- lys ine (285 mCi/mmol) (New England) were app l ied fo r a 6 h per iod at e i t h e r 0 or 18 h of i ncuba t ion . A f t e r the incubat ion per iod , the t i ssues were washed, f rozen and homoge- nized in bu f f e r (50 mM Tr i s pH 7.6, 50 mM KCI, 10 mM MgCI 2, 5 mM ~-mercaptoethanol ) w i th a mortar and pes t l e and cen t r i f uged at 15,000 g f o r 30 min. The supernatant was again cen t r i f uged at 190,000 g f o r 80 min.
Pro te ins were p r e c i p i t a t e d from the su- pernatant w i th t r i c h l o r o a c e t i c acid ( f i n a l concen t ra t ion of 5%), cen t r i f uged and resus- pended in water , n e u t r a l i z e d wi th 0.5 M KOH and t h i s process was repeated tw ice. Pro te ins were assayed by the Lowry method (Lowry et al. 1951) using bovine serum albumin (Sigma) as the standard. R a d i o a c t i v i t y was determined in 100 ~I of p ro te in samples w i th 10 ml of
194
2 0 0
190
8O 09
U
2 0 '
0 Asp Glu Pro Vol Tyr Gly AIG Phe I le Leu Lys Arg Met Thr
÷
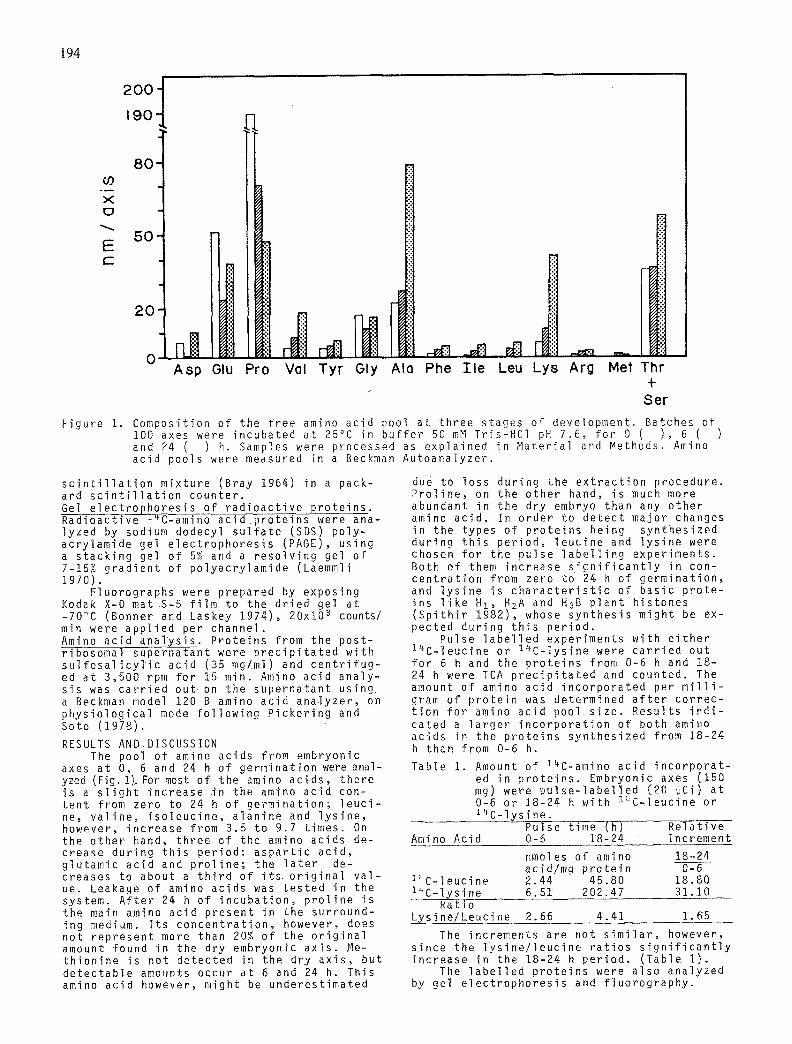
Ser Figure 1. Composition of the f ree amino acid pool at three stages of development. Batches of
100 axes were incubated at 25°C in bu f f e r 50 mM Tr is-HCl pH 7.6, fo r 0 ( ) , 6 ( and 24 ( ) h, Samples were processed as expla ined in Mater ia l and Methods. Amino acid pools were measured in a Beckman Autoanalyzer .
s c i n t i l l a t i o n mix ture (Bray 1964) in a pack- ard s c i n t i l l a t i o n counter . Gel e l ec t r opho res i s of r a d i o a c t i v e p ro te i ns . R a d i o a c ~ ~ n o a c i ~ ' n s were a-na- lyzed by sodium dodecyl s u l f a t e (SDS) po ly - acry lamide gel e l ec t r opho res i s (PAGE), using a s tack ing gel of 5% and a r eso l v i ng gel of 7-15% grad ien t of po lyacry lamide (Laemmli 1970).
Fluorographs were prepared by exposing Kodak X-O mat S-5 f i l m to the dr ied gel at -70°C (Bonner and Laskey 1974), 20x103 counts/ min were app l ied per channel. Amino acid a n a l y s i s . Prote ins from the post- ~s-s-oma7 supernatant were p r e c i p i t a t e d wi th s u l f o s a l i c y l i c acid (35 mg/ml) and c e n t r i f u g - ed at 3,500 rpm fo r 15 min. Amino acid ana ly - s is was ca r r i ed out on the supernatant using a Beckman model 120 B amino acid ana l yze r , o p h y s i o l o g i c a l mode f o l l o w i n g P icker ing and Soto (1978).
RESULTS AND DISCUSSION The pool of amino acids from embryonic
axes at O, 6 and 24 h of germinat ion were anal- yzed (Fig. 1).For most of the amino ac ids , there is a s l i g h t increase in the amino acid con- ten t from zero to 24 h of germina t ion ; l e u c i - ne, v a l i n e , i s o l e u c i n e , a lan ine and l y s i n e , however, increase from 3.5 to 9.7 t imes. On the other hand, three of the amino acids de -~ crease dur ing t h i s per iod: a s p a r t i c ac id , g lu tamic acid and p r o l i n e ; the l a t e r de- creases to about a t h i r d of i t s o r i g i n a l v a l - ue. Leakage of amino acids was tes ted in the system. A f t e r 24 h of i ncuba t i on , p r o l i n e is the main amino acid present in the surround- ing medium. I t s concen t ra t i on , however, does not represent more than 20% of the o r i g i n a l amount found in the dry embryonic ax i s , Me- t h i on ine is not detected in the dry a x i s , but de tec tab le amounts occur at 6 and 24 h. This amino acid however, might be underest imated
due to loss dur ing the e x t r a c t i o n procedure. P r o l i n e , on the other hand, is much more abundant in the dry embryo than any other amino ac id . In order to detect major changes in the types of p ro te ins being synthes ized during t h i s per iod , leuc ine and l y s i n e were chosen fo r the pulse l a b e l l i n g exper iments. Both of them increase s i g n i f i c a n t l y in con- c e n t r a t i o n from zero to 24 h of germina t ion , and l y s i n e is c h a r a c t e r i s t i c of basic p ro te - ins l i k e H1, HzA and H3B p lan t h istones ( S p i t h i r 1982), whose syn thes is might be ex- pected during t h i s per iod .
Pulse l a b e l l e d experiments w i th e i t h e r 14C-leucine or 1~C- lys ine were ca r r i ed out f o r 6 h and the p ro te ins from 0-6 h and 18- 24 h were TCA p r e c i p i t a t e d and counted. The amount of amino acid incorpora ted per m i l l i - gram of p ro te in was determined a f t e r cor rec- t i on f o r amino acid pool s i ze . Results i n d i - cated a l a rge r i nco rpo ra t i on of both amino acids in the p ro te ins synthes ized from 18-24 h than from 0-6 h.
Table 1. Amount of 14C-amino acid i n c o r p o r a t - ed in p ro te i ns . Embryonic axes (150 mg) were p u l s e - l a b e l l e d (20 ~Ci) at 0-6 or 18-24 h wi th l~C- leuc ine or 14C- lys ine .
Pulse t ime (~) Re la t i ve Amino Acid 0-6 18-24 Increment
nmoles of amino 18-24 acid/mg p ro te in 0-6
1~C-leucine 2.44 45.80 18.80 l ~ 4 C - l y s i n e 6.51 202.47 31..10
Ratio Lys ine/Leuc ine 2.66 4.41 1.65
The increments are not s i m i l a r , however, since the l y s i n e / l e u c i n e r a t i o s s i g n i f i c a n t l y increase in the 18-24 h per iod. (Table 1).
The l a b e l l e d p ro te ins were also analyzed by gel e l ec t r opho res i s and f l uo rography .
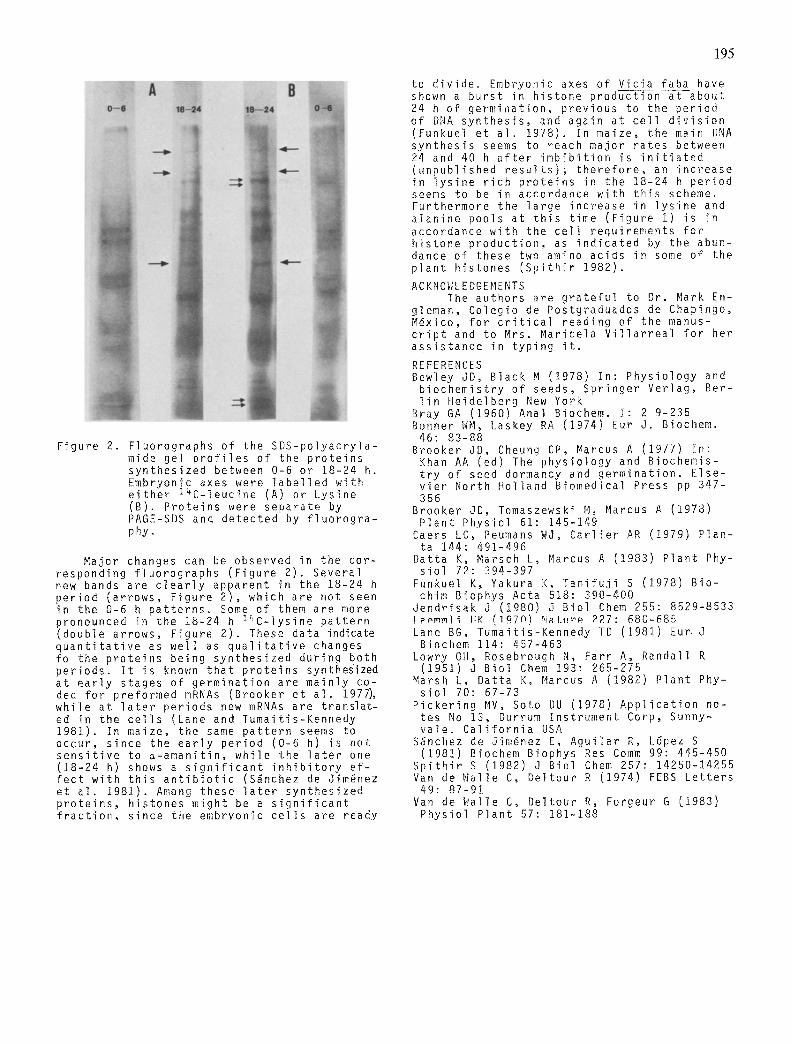
Figure 2. Fluorographs of the SDS-po lyacry la- mide gel p r o f i l e s of the p ro te ins synthes ized between 0-6 or 18-24 h. Embryonic axes were l a b e l l e d wi th e i t h e r 14C-leucine (A) or Lysine (B). Prote ins were separate by PAGE-SDS and detected by f l u o r o g r a - phy.
Major changes can be observed in the cor- responding f luorographs (F igure 2). Several new bands are c l e a r l y apparent in the 18-24 h per iod (arrows, Figure 2) , which are not seen in the 0-6 h pa t te rns . Some of them are more pronounced in the 18-24 h l ~ C - l y s i n e pa t te rn (double arrows, Figure 2). These data indicate q u a n t i t a t i v e as wel l as q u a l i t a t i v e changes fo the p ro te ins being synthes ized during both per iods . I t is known tha t p ro te ins synthesized at e a r l y stages of germinat ion are main ly co- ded fo r preformed mRNAs (Brooker et a l . 1977), wh i le at l a t e r per iods new mRNAs are t rans la t - ed in the c e l l s (Lane and Tumait is-Kennedy 1981). In maize, the same pa t te rn seems to occur, s ince the e a r l y per iod (0-6 h) is not s e n s i t i v e to ~ -aman i t i n , wh i le the l a t e r one (18-24 h) shows a s i g n i f i c a n t i n h i b i t o r y e f - f e c t w i th t h i s a n t i b i o t i c (S~nchez de Jim~nez et a l . 1981). Among these l a t e r synthes ized p r o t e i n s , h istor ies might be a s i g n i f i c a n t f r a c t i o n , s ince the embryonic c e l l s are ready
195
to d i v i d e . Embryonic axes of V ic ia faba have shown a burst in h is tone product ion at about 24 h of germina t ion , previous to the per iod of DNA syn thes i s , and again at c e l l d i v i s i o n (Funkuel et a l . 1978). In maize, the main DNA syn thes is seems to reach major rates between 24 and 40 h a f t e r i m b i b i t i o n is i n i t i a t e d (unpubl ished r e s u l t s ) ; t h e r e f o r e , an increase in l y s i n e r i ch p ro te ins in the 18-24 h per iod seems to be in accordance wi th t h i s scheme. Furthermore the large increase in l y s i n e and a lan ine pools at t h i s t ime (F igure 1) is in accordance w i th the c e l l requirements f o r h is tone produc t ion , as i nd i ca ted by the abun- dance of these two amino acids in some of the p lan t h is tones ( S p i t h i r 1982).
ACKNOWLEDGEMENTS The authors are g r a t e f u l to Dr. Mark En-
gleman, Colegio de Postgraduados de Chapingo, M#xico, fo r c r i t i c a l reading of the manus- c r i p t and to Mrs. Mar ice la V i l l a r r e a l fo r her ass is tance in typ ing i t .
REFERENCES Bewley JD, Black M (1978) In : Physio logy and
b iochemis t ry of seeds, Spr inger Ver lag, Ber- l i n Heidelberg New York
Bray GA (1960) Anal Biochem. I : 2 9-235 Bonner WM, Laskey RA (1974) Eur J. Biochem.
4 6 : 8 3 - 8 8 Brooker JD, Cheung CP, Marcus A (1977) In:
Khan AA (ed) The phys io logy and Biochemis- t r y of seed dormancy and germina t ion . Else- v i e r North Hol land Biomedical Press pp 347- 356
Brooker JD, Tomaszewski M, Marcus A (1978) Plant Physiol 61 :145-149
Caers LC, Peumans WO, C a r l i e r AR (1979) Plan- ta 144:491-496
Datta K, Marsch L, Marcus A (1983) Plant Phy- s i o l 72 :394-397
Funkuel K, Yakura K, T a n i f u j i S (1978) Bio- chim Biophys Acta 518:390-400
Jendr isak J (1980) J Bio l Chem 255:8529-8533 ! ~ m m l i II~ (197N) N@t~!re 227: 680-685 Lane BG, Tumait is-Kennedy TD (1981) Eur, J
Biochem 114:457-463 Lowry OH, Rosebrough N. Farr A, Randall R
(1951) J B io l Chem 193:265-275 Marsh L, Datta K, Marcus A (1982) Plant Phy-
s i o l 70: 67-73 P icker ing MV, Soto DU (1978) A p p l i c a t i o n no-
tes No 13, Durrum Inst rument Corp, Sunny- va le . C a l i f o r n i a USA
S~nchez de Jim~nez E, Agu i l a r R, L6pez S (1981) Biochem Biophys Res Comm 99 :445-450
S p i t h i r S (1982) J Biol Chem 257: 14250-14255 Van de Walle C, Del tour R (1974) FEBS Le t te rs
49: 87-91 Van de ~lal le C, Del tour R, Forgeur G (1983)
Physiol Plant 57 :181-188