Embed Size (px)
Citation preview

PECKIANA • Volume8(2012) • pp.245–254
ISSN 1618-1735
New findings of the genus Altomiramys (Mammalia, Gliridae) in the Lower Miocene (Agenian, Ramblian and Aragonian) of the Ebro and Ribesalbes-Alcora Basins (Spain)
Vicente D. crespo, Francisco J. ruiz-sánchez, samuel mansino, mar González-parDos, maría ríos, eFrén colomina, Xabier murelaGa, Juan cruz larrasoaña, plinio montoya & matthiJs FreuDenthal
AbstractAltomiramys (Gliridae, myomiminae) is a very uncommon glirid in the rodent assemblages from the
lower part of the lower miocene (agenian to ramblian) of the iberian peninsula. only one species, Altomiramys daamsi, has been described from localities of ramblian age (loranca-1 and ramblar-1). an unpublished second species, Altomiramys aff. daamsi, is mentioned in the locality of cetina de aragón, of agenian age. new findings of this genus in the ebro and ribesalbes-alcora basin have allowed to expand the time range for this taxon to the later part of the early miocene (aragonian). this increase of the material leads to a modification of the diagnosis of the genus.
Keywords: early miocene, iberian peninsula, biostratigraphy
1. Introductionthe diversity of dormice (glirids) in europe during the miocene (23-5.3 ma) reaches
28 genera, with a maximum in the lower miocene (16.5 ma). at the end of the miocene, 23 of these genera were extinct (Daams 1999). a strong reduction of diversity came with the immigration of the modern cricetids (16.6 ma) and the murids (9.8 ma). the entry of these faunas coincided with a dramatic reduction in both number of species and relative abundance of the glirids in the rodent assemblages of europe.
Altomiramys Díaz-molina & lópez-martínez, 1979 (Gliridae, myomiminae) is a very uncommon endemic glirid in the rodent assemblages from the lower miocene (agenian and ramblian mn1-mn3, 23.0-16.6 ma) of the iberian peninsula. so far, only one species, Altomiramys daamsi Díaz-molina & lópez-martínez, 1979, has been described from localities of this age (loranca-1; agenian, upper part of mn2, 20 ma, Díaz-molina & lópez-martínez, 1979 and ramblar-1; lower ramblian, lower part of mn3, 19.4-19.9 ma, Daams, 1989) but an unpublished second species, Altomiramys aff. daamsi (Daams 1974, 1989, 1999), is mentioned from cetina de aragón, an older site from the early part of the early miocene (mn1). therefore, the fossil record of this genus covers the span between mn1 and the lower part of mn3 (Fig. 1). the remains of Altomiramys are extremely scarce in the paleontological record, with only one specimen recorded in its type locality (loranca-1) and 10 specimens in ramblar-1. in this paper we describe new remains of this genus from the localities of cuesta agujeros-2 (ca-2) and cuesta agujeros-4 (ca-4) (ebro basin), and mas de antolino-5 (mab-5, ribesalbes-alcora basin).

Vicente D. crespo et al.246
2. Material and methods2.1. Stratigraphical settingthe non-recurrent compositional and evolutionary changes of the european mammal fauna
history have been extensively used as a relative-age tool for the continental sediments in which they are found (van der meulen et al. 2011). thaler (1966) was the first to propose a modern biozonation based on mammals for paleogene and neogene. a comprehensive framework for mammal history was provided by the mn zones. this system is based on neogene mammal assemblages from europe and north africa (mein 1975, 1979, 1990). according to this system, the neogene is divided into seventeen units. it is ideally applicable, independent of regional factors or biogeography, and has been widely used during the past decades. since its proposition, the mn-system has been used for correlations to different scales. the recognition of the mn units in several geographic areas has allowed the establishment of different ages from the mn boundaries (see bibliography in van der meulen et al. 2011). many of these proposals have been calibrated to the time scale with paleomagnetic and/or radiometric calibrations. For the time interval represented in the localities studied in this paper, we will use the calibrations contained in larrasoaña et al. (2006), van der meulen et al. (2012) and ruiz-sánchez et al. (in press).
the material from mab-5, ca-2 and ca-4 was collected during paleontological excavations carried out in the past five years, using the standard methodology for collecting microvertebrate fossils (hibbard 1950, thaler 1966, Freudenthal et al. 1976, Daams & Freudenthal 1988). the terminology of dental morphology used to describe the glirid remains is taken from Freudenthal (2004) (Fig. 2). the lower case denotes lower teeth, upper case denotes upper teeth (m1, m2, m1, m2). the measurements are given in tenths of millimetres, and were taken on a leica mz75 binocular microscope by means of displacement of a mechanical stage connected to a sony magnescale measuring device.
the tooth of Altomiramys from mab-5 is kept at the mGuV (museu de Geologia de la universitat de València) with the acronym mab5-, and the five teeth from locality ca-2 and one from ca-4 are housed in the Departamento de estratigrafía y paleontología de la universidad del país Vasco (upV/ehu) with the acronyms ca2- and ca4-.
Lower Miocene
Agenian Ramblian Ar.
Series
Continental Stage
MN Zone
Localities
MN
1
MN
2
MN
3
MN
4M
AB
-5
CA
-4
Ram
blar-1
Loranca-1
CA
-2
Cetina de
Argón
Altomiramys daamsi
Altomiramys sp.
aff
aff
Fig. 1 stratigraphic distribution of Altomiramys daamsi and Altomiramys sp. in different spanish localities through the lower miocene (after Daams 1999 and this paper).

new findings of the genus Altomiramys (mammalia, Gliridae) 247
3. Results 3.1. Geological settingthe ribesalbes-alcora basin is located in the east of spain (castellón province) and is
situated in the southern maestrazgo, between the sierra de espadán and fallen blocks of the Desierto de las palmas (agustí et al. 1988). the deposits of this basin have been divided into five units (a, b, c, D and e) (anadón 1983), and they consist of yellow and grey shales with banks of sandstone, dolomite and limestone beds. agustí et al. (1988) in the section of mas de antolino, and after that ruiz-sánchez et al. (2010) and Furió et al. (2010) in the section of mas de antolino b, both in unit c of anadón (1983), found several assemblages of mammalian remains. these localities contain faunal associations of early aragonian age (mn4). one of these localities, the rich association from the locality mab-5, contains among other rodents, Megacricetodon cf. primitivus (Freudenthal, 1963) and Ligerimys ellipticus Daams, 1976, which allow to date this site as early aragonian (mn4) (tab. 1).
on the other hand, the two other localities with Altomiramys remains analyzed in this paper are situated in the ebro basin. From the late eocene until the late miocene, this basin was an endorrheic depression. in the north-central area of this basin, the tudela Formation has been defined, where the localities ca-2 and ca-4 are located. this Formation is composed of 650 meters of fluvial distal, palustrine and lacustrine sediments, accumulated during the early and middle miocene (larrasoaña et al. 2006) and it crops out extensively in the so-called bardenas reales of navarra. the fossil sites presented in this work are located in the section of cuesta de agujeros (ca-). the presence of Armantomys daamsi (álvarez-sierra et al. 1991) in ca-2 and Armantomys cf. jasperi Daams, 1991, in ca-4 allow an age attribution of these localities to the late agenian (local zone y, mn2) and late ramblian (local zone a, mn3) (ruiz-sánchez et al. in press).
3.2. Systematicsaccording to the original diagnosis of the genus, Altomiramys is a glirid with upper molars
with subrounded outline, a low crown and concave occlusal surface. the ridges are thin and well-separated, with precentroloph only, which joins the anteroloph in an arch-shaped ridge. the labial part of the posteroloph ends free (Díaz-molina & lópez-martínez 1979).
the material of this genus is very scarce, restricted to one tooth in the type locality, loranca-1, four in ramblar-1, and an indeterminate low number of unpublished material in cetina de aragón; to this now should be added five teeth in ca-2, one in ca-4 and another one in mab-5.
Fig. 2 nomenclature of parts of the cheek teeth, adapted after Freudenthal (2004).
Vicente D. crespo et al.248
Tab. 1 Faunal list of the site mab-5 (ruiz-sánchez et al. 2010, Furió et al. 2010).
Faunal list of MAB-5order rodentia
Family GliridaeGlirudinus cf. modestusMicrodyromys sp.Simplomys cf. juliiAltomiramys sp.
Family cricetidaeMegacricetodon cf. primitivusDemocricetodon/Fahlbuschia sp.
Family eomyidaeLigerimys ellipticus
Family sciuridaesciuridae indet.
order insectivorainsectivora indet.
Family DimylidaePlesiodimylus sp.
infraclass metatheria Family herpetotheriidae
Amphiperatherium frequens
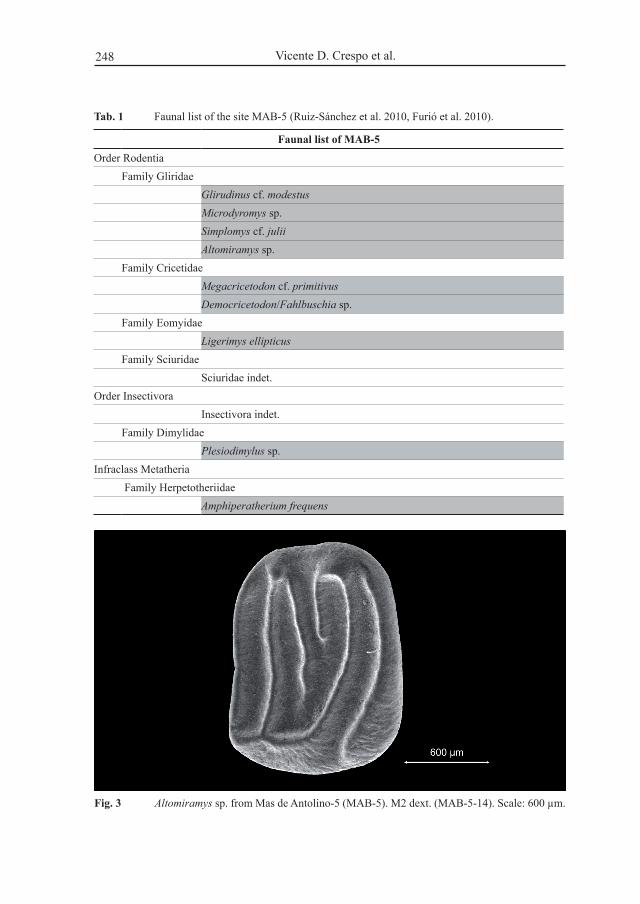
Fig. 3 Altomiramys sp. from mas de antolino-5 (mab-5). m2 dext. (mab-5-14). scale: 600 µm.

new findings of the genus Altomiramys (mammalia, Gliridae) 249
order: rodentia bodwich, 1821Family: Gliridae muirhead, 1819subfamily: myomiminae Daams, 1981attributed genera: Miodyromys Kretzoi, 1943, Prodryomys mayr, 1979, Myomimus ognev,
1924, Peridyromys stehlin & schaub, 1951, Pseudodryomys (emended diagnosis by García-paredes et al., 2009), Ramys García-moreno & lópez-martínez, 1986, Simplomys García-paredes et al., 2009, Plesiodyromys Werner, 1994 and Vasseuromys baudelot & De bonis, 1966.
Genus Altomiramys: Díaz-molina & lópez-martínez, 1979
Emended diagnosisa glirid with low-crowned molars and concave occlusal surface. the ridges are thin and
well-separated. the upper molars have precentroloph and/or postcentroloph. the presence of extra ridges in the upper and lower molars is variable. the labial portion of the posteroloph ends free. there is no continuous endoloph.
Differential diagnosisthe genus Altomiramys differs from other miocene low-crowned myomininae in: wide
valleys, thin and slanting ridges; it differs from Myomimus, Peridyromys, Pseudodryomys, Ramys and Vasseuromys in: the complexity of dental pattern; it differs from Simplomys in: upper molars with more rounded outline and anterior valley that extends along the lingual border to the protocone; it differs from Peridyromys in: the metalophid reaching the metaconid; it differs from Plesiodyromys in: sharper edges, not reduced central structures and lack of the cingulum.
Altomiramys sp.locality: mas de antolino 5 (mab-5)material: 1 m2 (mab-5-14, Fig. 3; size in tab. 2)Description: the only specimen of Altomiramys sp. from mab-5 is an m2 which has
an anteroloph that ends below the protocone at the lingual border, disconnected from the protocone. the precentroloph is labially connected to the base of the paracone, extending almost halfway into the central valley. the postcentroloph is longer than the precentroloph, and parallel to this ridge, ending near the protocone. the metaloph and protoloph join at the lingual border. the posteroloph is connected to the protocone at the lingual border and labially not connected to the metacone.
locality: cuesta agujeros 4 (ca-4)material: 1 m2 (ca-4-1/3, Fig. 4F; size in tab. 2)Description: this very scarce material is described in ruiz-sánchez et al. (in press). the
tooth from ca-4 has narrow and well-separated ridges, with a long anteroloph, not connected to the protocone. the anteroloph and protoloph are connected to the paracone. a long precentroloph and a short-medium postcentroloph are present. between precentroloph and protoloph a short prototrope is present.
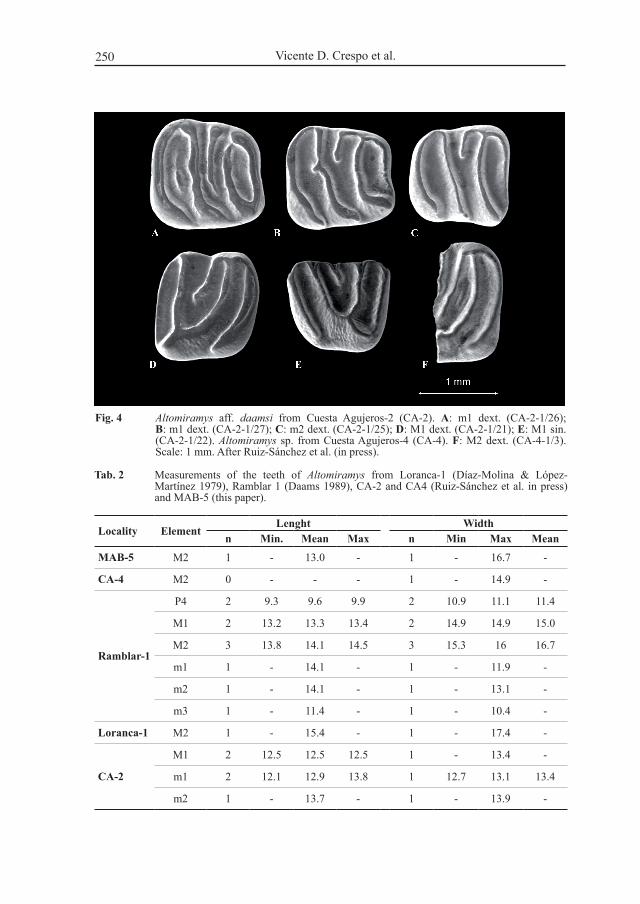
Altomiramys aff. daamsi Díaz-Molina & López-Martínez, 1979locality: cuesta agujeros 2 (ca-2)material: 2 m1 (ca-2-1/26 Fig. 4a, ca-2-1/27, Fig. 4b), 1 m2 (ca-2-1/25, Fig. 4c)
and 2 m1 (ca-2-1/21, Fig. 4D, ca-2-1/22, Fig 4e); sizes in table 2.Description: Five teeth are described in ruiz-sánchez et al. (in press) from this site. the
lower molars have a low crown, narrow crests and broad valleys, with a long anterolophid,
Vicente D. crespo et al.250
Fig. 4 Altomiramys aff. daamsi from cuesta agujeros-2 (ca-2). A: m1 dext. (ca-2-1/26); B: m1 dext. (ca-2-1/27); C: m2 dext. (ca-2-1/25); D: m1 dext. (ca-2-1/21); E: m1 sin. (ca-2-1/22). Altomiramys sp. from cuesta agujeros-4 (ca-4). F: m2 dext. (ca-4-1/3). scale: 1 mm. after ruiz-sánchez et al. (in press).
Tab. 2 measurements of the teeth of Altomiramys from loranca-1 (Díaz-molina & lópez- martínez 1979), ramblar 1 (Daams 1989), ca-2 and ca4 (ruiz-sánchez et al. in press) and mab-5 (this paper).
Locality ElementLenght Width
n Min. Mean Max n Min Max MeanMAB-5 m2 1 - 13.0 - 1 - 16.7 -
CA-4 m2 0 - - - 1 - 14.9 -
Ramblar-1
p4 2 9.3 9.6 9.9 2 10.9 11.1 11.4
m1 2 13.2 13.3 13.4 2 14.9 14.9 15.0
m2 3 13.8 14.1 14.5 3 15.3 16 16.7
m1 1 - 14.1 - 1 - 11.9 -
m2 1 - 14.1 - 1 - 13.1 -
m3 1 - 11.4 - 1 - 10.4 -
Loranca-1 m2 1 - 15.4 - 1 - 17.4 -
CA-2
m1 2 12.5 12.5 12.5 1 - 13.4 -
m1 2 12.1 12.9 13.8 1 12.7 13.1 13.4
m2 1 - 13.7 - 1 - 13.9 -

new findings of the genus Altomiramys (mammalia, Gliridae) 251
which may be connected to the protoconid. in 1 out of 2 m1 a posterotropid (Fig. 4a), and a short posterior spur in the middle of the mesolophid (Fig. 4b) are present. the upper molars have an anteroloph, which is isolated from the protocone and a precentroloph connected to the paracone. protoloph and metaloph are connected, forming an oblique y.
4. Discussionthe very scarce material from the type locality, loranca-1, impedes to know the variability
of the diagnostic features of the genus. the study of the Altomiramys material from ramblar-1 (Daams 1989) and the readscription of Pseudodryomys simplicidens (Simplomys simplicidens in García-paredes et al. 2009) from cetina de aragón to Altomiramys aff. daamsi (Daams, 1989), allow to know its variability better.
the general shape of the teeth (upper molars with rounded outline; low crown, concave occlusal surface, ridges thin and well-separated) is very similar in all assemblages (loranca-1, ramblar-1, ca-2, ca-4, cetina de aragón and mab-5). however, the presence of a unique precentroloph, as mentioned in the original diagnosis, is variable. thus, the postcentroloph is absent in the only tooth from the type locality, loranca-1, in 1 out of 2 m2 from ramblar-1, in the only complete m1 from ca-2, and in all (37) specimens adscribed to A. aff. daamsi in cetina de aragón (Daams 1989).
the size of the material from ca-2 (mn2) is somewhat smaller than the material described by Daams (1989) from ramblar-1 (tab. 2). the specimens from ca-2 show some morphological features, different from those of the type locality: development of a short posterior spur in the middle of the mesolophid (Fig. 4b) in 1 out of 2 m1, and the presence in 1 out of 2 m1 of a posterotropid (Fig. 4a).
in cetina de aragón have been cited P. ibericus and P. simplicidens (Altomiramys aff. daamsi sensu Daams, 1989). among the m1 of glirids from cetina de aragón, 15 out of 65 specimens lack extra ridges, another 15 have only a posterotropid and 35 have anterotropid and posterotropid. on the other hand, in cetina de aragón, 21 out of 91 m2 lack extra ridges, 44 have only a posterotropid, and 26 have anterotropid and posterotropid. according to García-paredes et al. (2009), S. simplicidens lacks extra ridges. the absence of lower molars in the type locality of A. daamsi impedes to know the variability of this and other characters in the lower molars.
the general shape of the tooth from ca-4 (mn3) allows to adscribe this fragment to the genus Altomiramys. the precentroloph in the specimen from ca-4 is larger than in those from loranca-1 and ramblar-1. the presence of a short prototrope (Fig. 4F) is similar to that described in 1 out of 3 m2 from ramblar 1.
the tooth of Altomiramys sp. from mab-5 (mn4) is smaller than A. daamsi from the type locality, loranca-1, and similar to the material from ramblar-1 (tab. 2). the most striking feature of the m2 of Altomiramys from mab-5 is the presence of two centrolophs, of which the postcentroloph is longer than the precentroloph. as has been remarked before, the postcentroloph is absent in the type locality loranca-1 (one tooth) and in only 23 % of m1.2 from cetina de aragón and 1 out of 2 m2 from ramblar-1.
5. Conclusionsa scarce material of Altomiramys has been found in the localities ca‐2, ca‐4 (ebro basin)
and mab‐5 (ribesalbes‐alcora basin). the presence of Armantomys daamsi suggests a late

Vicente D. crespo et al.252
agenian age (upper part of mn2) for ca-2; Armantomys cf. jasperi gives a late ramblian age (upper part of mn3) for ca-4 , and mab-5 is attributed an early aragonian age (mn4) because of the presence of Megacricetodon cf. primitivus and Ligerimys ellipticus. these new findings allow to extend the record of the genus Altomiramys until the early aragonian (mn4).
the new findings of Altomiramys in the ribesalbes-alcora and ebro basins, allow a better knowledge of the metrical and morphological variability of this genus. Whereas the general shape, characterized by upper molars with subrounded outline, low crown, concave occlusal surface and thin and well-separated ridges, is constant in all assemblages from mn2 until mn4, the presence and development of the precentroloph and postcentroloph is more variable than stated in the diagnosis of the genus. While in the type locality, loranca-1, (with only one tooth), the precentroloph is the only centroloph in the trigone, in the assemblages from cetina de aragón (mn2), ramblar-1, ca-4 (mn3), and mab-5 (mn4), the postcentroloph may also be present. in the same way, and despite the absence of lower molars in the type locality, loranca-1, in the oldest assemblages of the genus Altomiramys (ca-2), the posterotropid may be present too.
therefore, the diagnosis of the genus Altomiramys has been modified, including the possible presence of a postcentroloph in the upper molars and of a posterotropid in the lower molars.
6. Referencesagustí, J., anadón, p., Gingsburg, l., mein, p. & moissenet, e. (1988): araya et mira: nouveaux gisements
de mammifères dans le miocène inférieur-moyen des chaînes ibériques orientales et méditerranéennes. conséquences stratigraphiques et structurales. – paleontologia i evolució 22: 83–101.
anadón, p. (1983): características generales de diversas cuencas lacustres terciarias con pizarras bituminosas del ne de la península ibérica. – X congreso nacional de sedimentología 1: 9–12.
Daams, r. (1974): Pseudodryomys (Gliridae, rodentia, mammalia) from early and middle miocene deposits in spain. – proceedings Koninklijke nederlandse akademie van Wetenschappen, b 77/3: 201–240.
Daams, r. (1989): miscellaneous Gliridae from the miocene of the calatayud-teruel basin, aragón, spain. – scripta Geologica 89: 13–26.
Daams, r. (1999): Family Gliridae. – in: rössner, G. e. & heissig, K. (eds): the miocene land mammals of europe. – Verlag Dr. Friedrich pfeil: 301–318.
Daams, r. & Freudenthal, m. (1988): synopsis of the Dutch-spanish collaboration program in the neogene of the calatayud-teruel basin, 1976–1986. – in: Freudenthal, m. (ed.): biostratigraphy and paleoecology of the neogene micromammalian faunas from the calatayud-teruel basin (spain). – scripta Geologica, spec. issue 1: 3–18.
Díaz-molina, m. & lópez-martínez, n. (1979): el terciario continental de la Depresión intermedia (cuenca). bioestratigrafía y paleogeografía. – estudios Geológicos 35: 149–167.
Freudenthal, m. (2004): Gliridae (rodentia, mammalia) from the eocene and oligocene of the sierra palomera (teruel, spain). – treballs del museu de Geologia de barcelona 12: 97–173.
Freudenthal, m., meijer, t. & meulen, a. J. van der (1976): preliminary report on a field campaign in the continental pleistocene of tegelen (the netherlands). – scripta Geologica 34: 1–27.
Furió, m., ruiz-sánchez, F. J., crespo-roures, V. D., Freudenthal, m. & montoya, p. (2010): new didelphimorph (marsupialia, mammalia) and dimylid (eulipotyphla, mammalia) remains from the late early miocene of spain. – 70th meeting of society of Vertebrate paleontology. abstract book: 93.
García-paredes, i., peláez-campomanes, p. & álvarez-sierra, m. a. (2009): Gliridae (rodentia, mammalia) with a simple dental pattern: a new genus a new species from the european early and middle miocene. – zoological Journal of the linnean society 157: 622–652.
hibbard, c. W. (1950): mammals of the rexroad Formation from Fox canyon, meade county, Kansas. – contributions from the museum of paleontology, university of michigan 8:113–192.
larrasoaña, J. c., murelaga, X. & Garcés, m. (2006): magnetobiochronology of lower miocene (ramblian) continental sediments from the tudela Formation (western ebro basin, spain). – earth and planetary science letters 243: 409–423.

new findings of the genus Altomiramys (mammalia, Gliridae) 253
mein, p. (1975): résultats du Groupe de travail des Vertébrés. – in: senes, J. (ed.): report on activity on the rcmns Working Groups. – bratislava, 1971–1975: 78–81.
mein, p. (1979): rapport d’activité du Groupe de travail de Vertébrés. mise à jour de la biostratigraphie du néogène basée sur les mammifères. – annales Géologiques des pays helléniques 3: 1367–1372.
mein, p. (1990): updating of mn zones. – in: lindsay, e. h., Fahlbusch, V. & mein, p. (eds): european neogene mammal chronology. – plenum press, new york: 73–90.
meulen, a. J. van der, García-paredes, i., álvarez-sierra, m. a., hoek-ostende, l.W. van den, hordijk, K., oliver, a., lópez-Guerrero, p., hernández-ballarín, V., peláez-campomanes, p. (2011): biostratigraphy or biochronology? lessons from the early and middle miocene small mammal events in europe. – Geobios, 44 (2-3): 309–321.
meulen, a. J. van der, García-paredes, i., álvarez-sierra, m.a., hoek-ostende, l. W. van den, hordijk, K. oliver, a. & peláez-campomanes, p. (2012): updated aragonian biostratigraphy: small mammal distribution and its implications for the miocene european chronology. – Geologica acta 10: 1–24.
ruiz-sánchez, F. J., crespo-roures, V. D., Furió, m., mansino, s., Freudenthal, m. & montoya, p. (2010): mab-5: un nuevo yacimiento de micromamíferos fósiles de la mn4 en la cuenca de ribesalbes-alcora (castellón). – in: moreno-azanza, m.,Díaz-martínez, i., Gasca, J. m., melero-rubio, m., rabal-Garcés, r. & sauqué, V. (eds): cidaris, número especial. – Viii encuentro de Jóvenes investigadores en paleontología, volumen de actas: 279–282.
ruiz-sánchez, F. J., murelaga, X., Freudenthal, m., larrasoaña, J. c. & Garcés, m. (in press): hypsodont myomiminae (Gliridae, rodentia) from five new localities in the lower miocene tudela Formation (bardenas reales, ebro basin, spain) and their bearing on the age of the agenian-ramblian boundary. – Geodiversitas.
thaler, l. (1966): les rongeurs fossiles du bas-languedoc dans leurs rapports avec l’histoire des faunes et la stratigraphie du tertiaire d’europe. – mémoires du museum national d’histoire naturelle, (c) 17: 295 pp.

Vicente D. crespo et al.254
accepted 20 august 2012
authors` addresses:
Vicente D. crespo*, Francisco J. ruiz-sánchez, samuel mansino, mar González-pardos, maría ríos, efrén colomina, plinio montoyaDepartament de GeologiaÀrea de paleontologiauniversitat de ValenciaDr. moliner 5046100 burjassot, spain
Xabier murelagaDepartamento de estratigrafía y paleontologíauniversidad del país Vascoaptdo. 64448080 bilbao, spain
Juan cruz larrasoañaárea de cambio Global, instituto Geológico y minero de españa (oficina de zaragoza)manuel lasala, 44, 9º b. 50006 zaragoza, spain
matthijs FreudenthalDepartamento de estratigrafía y paleontologíauniversidad de Granada, Granada (spain) and netherlands centre for biodiversitynaturalis2333 leiden, netherlands
*corresponding author: Vicente D. crespo (e-mail: [email protected])





























