Embed Size (px)
Citation preview

Neurotoxicology and Teratology, Vol. 11, pp. 35-38. © Pergamon Press plc. 1989. Printed in the U.S.A. 0892-0362/89 $3.00 + .00
Alterations in Offspring Behavior Induced by Chronic Prenatal Cocaine Dosing
R O B E R T F. S M I T H , K A T H L E E N M. MATTRAN, l M A U R A F. K U R K J I A N 2 A N D S T E V E N L. K U R T Z
Department o f Psychology, George Mason University, Fairfax, VA 22030
R e c e i v e d 28 D e c e m b e r 1987
SMITH, R. F., K. M. MATTRAN, M. F. KURKJIAN AND S. L. KURTZ. Alterations in offspring behavior induced by chronic prenatal cocaine dosing. NEUROTOXICOL TERATOL 11(1) 35-38, 1989.--Sperm-positive female Long-Evans hooded rats were dosed subcutaneously with 10 mg/kg/day cocaine or an equal volume of vehicle (0.9% sterile saline) from gestation day 4 (GD4) through GD 18. Offspring were assessed for development of negative geotaxis, righting reflex, spontaneous alternation, and open field activity, and for adult behaviors including DRL-20 acquisition, water maze, visual discrimination, barbiturate sleep time, shuttlebox avoidance, footshock sensitivity, and tail flick latency. Cocaine dosing produced no significant effects on dam weight gain, any measure of litter size and weight, or early postnatal behavioral tests, but there were significant drug effects on development of spontaneous alternation, development of open field activity, DRL-20 acquisition, water maze performance, tail flick, and footshock sensitivity. These data suggest that chronic administration of a modest dose of cocaine during gestation in the rat alters a number of behaviors in the offspring.
Prenatal cocaine Pregnancy Behavioral development Locomotor activity Pain sensitivity Schedule-controlled behavior
WHILE cocaine is a common drug of abuse, possible effects of prenatal administration on neurobehavioral development of the offspring have received little study. Chasnoff et al. (I) reported that infants born to cocaine-abusing mothers had some significant alterations in scores on the Brazelton Neonatal Behavioral Assess- ment Scale, primarily including poorer state organization and increased irritability. More recent work (2, 14, 20) has confirmed these findings, as well as reporting a high risk of Sudden Infant Death Syndrome in offspring of cocaine-using mothers, and an elevated risk of labor and delivery problems for these infants. Abnormal ventilatory patterns have been reported in infants born to cocaine addicts, as well as in other infants born to mothers addicted to opiates or to PCP (21).
There are extremely limited data on whether cocaine is a conventional teratogen. The drug has been variously reported as having no detectable teratogenic effects in mice or rats (5) (although rats administered very high doses during gestation experienced some consequent weight reduction), and as producing some teratogenic effects in mice (9). One human study (8) reported no teratology, although others (1,14) have reported evidence of urogenital malformations.
Although there are no published animal data on whether prenatal cocaine alters offspring behaviors, data on related sub- stances suggest the likelihood of such an effect. For example, we (18) have reported that a single 6 mg/kg dose of lidocaine on GD11 in the rat alters a number of offspring behaviors, and have since found (17) that administration on GD18 alters more behav- iors than administration on GD11. Middaugh and Zemp (10) have
summarized data on the consequences of prenatal administration of dopamine agonists and antagonists, and a number of studies [e.g., (22)] support the notion that cocaine is a D2 dopamine agonist.
Since human data on neurobehavioral effects of prenatal cocaine exposure are extremely limited, and subject to interpreta- tional problems of a correlational study in which subjects also used other drugs, we performed a brief assessment of a number of offspring behaviors in animals dosed with 10 mg/kg/day cocaine or an equal volume of saline from GD4 through GD18, as a preliminary determinant of whether cocaine appears to act as a neurobehavioral teratogen in the rat.
METHOD
Subjects
Subjects were offspring of Long-Evans derived hooded rats bred in our laboratory. Proven female breeders were housed overnight with proven male breeders until vaginal smear was positive for sperm. From GD4 through GD18 [GD1 =day found sperm positive; termination at GD18 was to avoid premature parturition suggested by human data- -e .g . , (2)], each dam was dosed subcutaneously with 10 mg/kg/day cocaine hydrochloride (Sigma Chemical Co., St. Louis, MO, administered as 10 mg/ml solution) in 0.9% sterile saline, or with an equal volume of vehicle. Following the completion of all dosing, dams were transferred to individual breeding boxes. Prior to GD18, dams were housed individually in hanging wire cages, as were offspring
1Present address: Hazleton Laboratories America, 1335-B Piccard Drive, Rockville, MD 20850. 2present address: Department of Psychology, University of Maryland, College Park, MD 20742.
35

36 SMITH ET AL.
after weaning on PND21. Food and water were available ad lib, except when food restriction was required for particular behavioral tests. Animals were maintained on a 12:12 LD schedule (0700- 1900 ON); dosing occurred at about 1000 each day, and testing occurred between 1000 and 1600.
Design
All pups were tested for development of negative geotaxis and righting reflex. Litters were then divided randomly into four sex pairs, with each sex pair receiving a different testing schedule. Data for analysis were litter means for those measures on which all pups in a litter were tested, and data for a single pup of each sex from each litter for those tests on which a single sex pair was tested from each litter. Data analysis was by analysis of variance (BMDP), with repeated measures where appropriate. All initial ANOVAs on offspring included both drug and sex in a factorial design; where drug × sex interactions occurred, subsequent analyses were performed separately for each sex.
Procedures
Descriptions for most tests have been previously reported (18). References for procedures not included in that paper are included in the following very brief description of our procedures.
Birth and growth. Within 24 hours of birth, all pups were weighed, sexed, and culled to four males and four females. Litters with fewer than four of either sex were culled to a total of eight.
For the first four weeks of life, weekly measures were taken of body weight of each offspring. After that point, body weight was taken on a biweekly basis, except that, for animals food deprived for a particular behavioral test, weight measures ceased at the beginning of food deprivation.
Preweaning behaviors. Righting reflex was assessed daily from postnatal day 2 (PND2) through PND8 (PND1 is defined as GD22, although births occurred on either GD22 or 23). Negative geotaxis was assessed daily from PND4 through PNDI0.
Postweaning behaviors. Sex pair I. Subjects were tested for developmental patterns of activity in the open field for 30 min on PNDs 25, 30, 35, and for adult activity on PND91. On PND94, they were food deprived, shaped to bar press, given three days of continuous reinforcement (100 per day), and 30 days of 30 min per day DRL-20 sec training, using methods previously reported (9). Subjects were given three day of water maze testing on PND137-139. Sex pair I1. On PNDs 25, 30, 35, 40, and 45, subjects were given two unreinforced trials per day in a T-maze, to test for develop- ment of spontaneous alternation [see (3) for procedures]. On PND90, they were food deprived, shaped to bar press, and given continuous reinforcement experience as described above for DRL.They were then trained on a visual discrimination task for 10 days, which consisted of 30 rain/day of light cued continuous reinforce- ment, alternating at semirandom intervals (mean = 30 sec, s.d. = 10 sec) with dark-cued extinction. On PND111 they were given a single intraperitoneal injection of 30 mg/kg sodium pentobarbital, and time from loss to regain of the righting reflex was manually timed. Sex pair III. These animals received five days of 30 Sonalert-cued two-way avoidance trials per day in a shuttle box on PNDs 92-96, followed by tail flick testing (immersion in 55 degree C water) and (one hour later) footshock sensitivity on PND101, Briefly, foot- shock sensitivity consisted of a series of ten ascending intensities following by the same ten in descending order (ISI = 10 sec), then a second entire series after a two rain wait. For each trial, subject movement was monitored by Lafayette 86010 activity monitor; the
TABLE l
PREGNANCY AND BIRTH MEASURES
Cocaine Control
M F M F
Number of 6.54-+0.72 5.63 -+0.41 6.09+0.55 5.55 -+0.47 pups
Pup birth 6.74+0.21 6.34-+0.20 6.86-_0.13 6.35-+0.09 weight (g)
Maternal 82.9 +- 2.51 83.4 -~ 2.56 weight gain (g)
dependent measure was pen deflection on an Esterline-Angus miniservo. The analysis of variance for response magnitudes included within-subject factors of shock intensity, ascending vs.descending order, and series. Sex pair IV. Subjects received footshock sensitivity and tail flick testing as described for Series III above on PND101, but no shuttle box testing.
For automated data collection, we used the Apple-based OPN system (19), instead of electromechanical control reported in previous work.
RESULTS
Eleven cocaine dosed dams (of 13 dosed) delivered litters; eleven control clams (of 11) delivered. Due to nonresponders (e.g., animals who exceeded the maximum of 300 sec ceiling repeatedly in the water maze were not included in latency data) and a few short litters (too small for four complete sex pairs), group sizes for each of the tasks reported here were 8-11/sex/treatment group. Cocaine dosing produced no differences in maternal weight gain for the dosing days GD4-18, birth weight or sex ratio of litters (see Table 1), or early developmental tests (righting reflex and negative geotaxis). Developmental weight gain was highly similar for cocaine-dosed and saline-closed offspring.
Analysis of variance of data for development of spontaneous alternation revealed a number of significant drug effects and interactions involving drug condition. For the probability of alternating, there was a significant drug × sex interaction, F( 1,31) = 4.02, p<0.05. Further analyses indicated that for males, there was a significant main effect of drug, F(1,15)=6.61, p<0.05, with cocaine-dosed male offspring (mean= 20.8%) al- ternating less than saline-dosed offspring (mean = 40,7%). There were no significant effects of cocaine dosing on alternation probability in females.
For latency data in the spontaneous alternation test, analysis revealed a significant interaction of drug and the factor of age at testing, F(5,80)--2.28, p<0.05. This effect was most prominent in females, in which latencies were longer in the cocaine-dosed animals at all ages tested except PND35. For males, there was an additional interaction of drug with trial, F(1,15)= 4.38, p<0.05, due to vehicle control animals having substantially longer mean latencies on the second trial of each test day than on the first (167.5 +49.7 vs. 80+26.9 see), while cocaine animals showed no such increase in latencies (87 vs. 92 see).
Analysis of variance for development of open field activity demonstrated that there was a significant interaction of drug, age at testing, and time period during a testing session, F(18,324)= 1.67, p<0.05, for male animals, but not for females. Inspection of the data indicated a tendency for cocaine-dosed offspring to be less

PRENATAL COCAINE AND OFFSPRING BEHAVIOR 37
eo.
o
~J 50-
~ . 4 o -
a o - eaJD
~o- o
1 o -
c°n-~v//
coc -~/// c o c- ~vv//o
o I I ( i i I i i i i i i
.:~ .4 .6 .8 1.0 1.2 1.4 1.6 1.8 2.0
S h o c k I n t e n s i t y (MA)
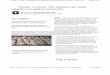
FIG. 1. Footshock sensitivity as a function of shock intensity, prenatal dosing history, and past exposure to footshock. Legend: coc = cocaine offspring, con =control offspring, w/= prior footshock experience in shuttle box, w/o = no prior footshock experience.
active than controls on middle time periods of GD30 and both early and late time periods of GD35, but significance tests for individual testing ages did not reveal any significant effect involving drug on either day. The triple interaction thus appeared to be a true interaction which was not dissociable into components, and which is therefore difficult to interpret.
For DRL-20 acquisition, there was a significant interaction of drug with the repeated measures factor (days), F(29,906)= 1.50, p<0.05. Although both groups obtained similar numbers of reinforcements on early days of testing (mean for first five days: cocaine = 16.9 ± 1.33, vehicle = 16.2 ± 1.29), they progressively diverged as testing proceeded (mean for last five days: cocaine = 33.6 -+ 3.52, vehicle = 26.0 ± 3.41).
For water maze latencies, there was a significant interaction between drug, day of testing and trial, F(18,396)=2.71, p<0.0005. The drug × trial interaction was also significant, F(9,198) = 2.48, p<0.05; these two interactions are due to cocaine- dosed offspring having longer latencies on early trials, particularly on the first day of testing• A nearly significant interaction between trial, drug, and sex, F(9,198)= 1.87, p<0.10, reflected that these effects were primarily attributable to the male offspring. Separate analyses for males and females indicated that for males there were significant effects of the drug x day interaction, F(2,20) = 6.53, p<0.001, the trial x drug interaction, F(9,90)= 5.46, p<0.001, and the triple interaction of drug, day, and trial, F( 18,180) = 5.76, p<0.001. These effects in males were primarily due to very long latencies in the cocaine-dosed offspring on early trials on the first test day. For example, the mean latency on the first trial of the first day was 158.1 ± 34.4 sec for cocaine animals, but 59.4 ± 20.4 sec for controls. Analysis of variance for females disclosed no significant drug effects on latency• No significant drug effects for either sex were discerned for number of errors.
An overall analysis of variance on footshock sensitivity data disclosed no significant effects other than the expected sex effects•
However, two interactions involving shock history and the drug condition of the animal approached significance (p's<0.10), and individual analyses for cocaine dosed subjects and controls were performed. For control subjects, prior shuttle box history signifi- cantly affected shock sensitivity. The history x series interaction, F(1,33)=5.13, p<0.05, and the history × series × shock intensity interaction. F(9,297)= 2.25, p<0.05, were both signif- icant. As Fig. 1 indicates, control animals which had received footshock experience had higher magnitudes of reaction to shock• This effect was especially obvious at higher shock amplitudes, and on the second series of shocks (series not shown separately in figure). Essentially, in control animals prior shock history ap- peared to sensitize the subjects to additional shocks. In cocaine- dosed subjects, this effect was absent, with the Fs corresponding to those which were significant in control subjects all less than 1. This resulted in cocaine-dosed subjects appearing hyposensitive relative to controls, if they had a prior shuttle box history [shock intensity × drug × sex interaction, F(9,306)=7.74, p<0.001, effects more prominent in females], but not in the absence of prior shuttle box exposure.
A similar interaction occurred for tail flick to thermal stimuli. There was a significant interaction between drug and shuttlebox experience, F(1,70)=7.06, p<0.01. In this case, in animals without prior footshock experience, cocaine-dosed offspring had significantly shorter latencies than controls (2.00±0.093 vs. 2 .68±0.24); in subjects tested after footshock experience, off- spring of cocaine- and saline-dosed dams did not differ signifi- cantly (2.47±0.15 vs. 2 .16±0.17 sec).
There were no effects of cocaine dosing which approached significance on shuttle box avoidance learning, visual discrimina- tion, adult open field activity or barbiturate sleep time.
D I S C U S S I O N
Previous work has indicated that 20 mg/kg subcutuaneous

38 SMITH E T AL.
cocaine produces peak serum concentrations of 490 ng/ml in the rat (11). A 96 mg intranasal dose, which produced detectable euphoria in humans, yielded a peak serum concentration of 206 ng/ml (6). Extrapolating from the data of these studies, we would expect our 10 mg/kg/day dosage to produce peak serum levels similar to those accompanying modest euphoria in humans. Despite our relatively modest dosing, significant behavioral alter- ations resulted from 10 mg/kg/day cocaine.
Significant effects of prenatal cocaine dosing were found on performance in spatial learning tasks (latency in water maze and latency and alternation rate in spontaneous alternation), on activity in the open field (although the magnitude of effect was slight), on number of reinforcements obtained in the DRL-20 paradigm, and on the development of shuttlebox-induced sensitization to painful stimuli. While a number of neural systems are involved in each of these types of behavior, hippocampus has been reported to be involved in each of these [e.g., (3, 12, 15, 16)]. Hippocampal development has proven sensitive to other developmental drug exposure, and local anesthetics administered to adults have major effects on the limbic system [e.g., (13)]. Dow-Edwards et al. (4) have recently reported that early postnatal cocaine reduces 2- deoxyglucose uptake in limbic system (particularly hippocampus). Although our behavioral data hardly provide specific support for hippocampal alterations induced by early cocaine, we will inves- tigate the possibility that prenatal cocaine dosing affects hippo- campal development, while not ruling out other possibilities.
Two measures of reactivity to painful stimuli (footshock and tail flick) indicated that the reactivity of the subjects was a joint function of prenatal dosing condition and whether the subject had recent experience with painful stimuli (footshock). Further work will be required to determine whether cocaine alters development
of systems involved in stress-induced analgesia, and to character- ize the nature of the system(s) affected.
These data indicate that a number of changes in offspring behavior follow maternal cocaine dosing with 10 mg/kg/day during gestation, but the limited scope of the present study left important questions which require additional investigation. We did not investigate possible effects of other doses of cocaine, and did not examine whether effects are attributable to direct alteration of neural development, or are secondary to alterations in maternal competence which might be induced by cocaine. One possibility which appears unlikely is that cocaine alters offspring behavior by altering maternal food consumption. Body weight gain during dosing in cocaine-dosed dams was within 1% of that of saline- dosed dams. Although questions such as direct vs. indirect actions on the pups, and the effects of other doses of cocaine must be resolved by further work (which should include fostering to untreated dams), our data suggest that prenatal cocaine dosing in the rat alters neurobehavioral development, that a range of behaviors is altered by such dosing, and that effects persist into adulthood. Taken in conjunction with the reports on human infants born to cocaine-abusing mothers (1, 2, 14, 20) and the apparently signif- icant numbers of users, our data suggest that more detailed studies of the potential of cocaine for altering neurobehavioral develop- ment should be pursued.
ACKNOWLEDGEMENTS
This work was supported by a summer stipend from the Graduate School of George Mason University to R.F.S. The authors thank Ms. Regina James, Ms. Dawn Royall and Ms. Karen Alien for competent assistance with testing.
REFERENCES
1. Chasnoff, I. J.; Bums, W. J.; Schnoll, S. H.; Bums, K. A. Cocaine use in pregnancy. N. Engl. Med. 313:666-669; 1985.
2. Chasnoff, I. J.; Bums, K. A.; Bums, W. J. Cocaine use in pregnancy: Perinatal morbidity and mortality. Neurotoxicol. Teratol. 9:291-293; 1987.
3. Douglas, R, J.; Isaacson, R. L. Spontaneous alternation and scopol- amine. Psychon. Sci. 4:283-284; 1966.
4. Dow-Edwards,D. L.; Fico, T. A.; Hutchings, D. E. Functional effects of cocaine given during critical periods of development. Teratol. 37:518; 1988.
5. Fantel, A. G.; MacPhail, B. J. The teratogenicity of cocaine. Teratol. 26:17-19; 1982.
6. Javaid, J. I.; Fishman, M. W.; Schuster, C. R.; Dekirmenjian, H.; Davis, J. M. Cocaine plasma concentration: relation to physiological and subjective effects in humans. Science 212:227-228; 1978.
7. McDonough, J. H.; Smith, R. F.; Smith, C. D. Behavioral correlates of soman-induced neuropathology: deficits in DRL acquisition. Neu- robehav. Toxicol. Teratol. 8:179-187; 1986.
8. Madden, J. D.; Payne, T. F.; Miller, S. Matemal cocaine abuse and effect on the newborn. Pediatrics 77:209-211;1986.
9. Mahalik, M. P.; Gautieri, R. F.; Mann, D. E. Teratogenic potential of cocaine hydrochloride in CF-1 mice. J. Pharm. Sci. 69:703-706; 1980.
10. Middaugh, L.D.; Zemp, J. W. Dopaminergic mediation of long-term behavioral effects of in utero drug exposure. Neurobehav. Toxicol. Teratol. 7:685-690; 1986.
11. Nayak, P. K.; Misra, A. L.; Mule, S. J. Physiological disposition and biotransformation of [3H]cocaine in acutely and chronically treated rats. J. Pharmacol. Exp. Ther. 196:556-569; 1976.
12. Olton, D.; Walker, J. A.; Gage, F. H. Hippocampal connections and spatial discrimination. Brain Res. 139:295-308; 1978.
13. Post, R. M.; Kennedy, C.; Shinohara, M.; Squillace, K.; Miyaoka,
M.; Suda, S.; Ingvar, D. H.; Sokoloff, L. Metabolic and behavioral consequences of lidocaine-kindled seizures. Brain Res. 324:295-303; 1984.
14. Ryan, L.; Ehrlich, S.; Finnegan, L. Cocaine abuse in pregnancy: Effects on the fetus and newborn. Neurotoxicol. Teratol. 9:295-299; 1987.
15. Schmaltz, L. W.; Isaacson, R. L. The effects of preliminary training conditions upon DRL performance in the hippocarnpectomized rat. Physiol. Behav. 1:175-182; 1966.
16. Smith, R. F. Mediation of footshock sensitivity by serotonergic projection to hippocampus. Pharmacol. Biocbem. Behav. 10:381- 388; 1978.
17. Smith, R. F.; Kurkjian, M. F.; Mattran, K. M.; Kurtz, S. L. Prenatal lidocaine exposure: effects of dosage and time on administration on behavioral sequelae in offspring. Teratology 37:521; 1988.
18. Smith, R. F.; Wharton, G. G.; Kurtz, S. L. Mattran, K. M.; Hollenbeck, A. R. Behavioral effects of mid-pregnancy administra- tion of lidocaine and mepivacaine in the rat. Neurobehav. Toxicol. Teratol. 8:61~8; 1986.
19. Spencer, D. G., Jr.; Emmett-Oglesby,M. W. Parallel processing strategies in the application of computers to the behavioral laboratory. Behav. Res. Methods Instrument. 17:294-300; 1985.
20. Wang, C. H.; Schnoll, S. H. Prenatal cocaine use associated with down regulation of receptors in human placenta. Neurotoxicol. Teratol. 9:301-304; 1987.
21. Ward, S. L. D.; Schuetz, S.; Krishna, V.; Bean, X.; Wingert, W.; Wachsman, L.; Keens, T. G. Abnormal sleeping ventilatory patterns in infants of substance-abusing mothers. Am. J. Dis. Child. 140: 1015-1020; 1986.
22. Woolverton, W. L. Effects of a DI and D2 dopamine antagonist on the self-administration of cocaine and pirebidil by rhesus monkeys. Pharmacol. Biochem. Behav. 24:531-535; 1986.