Embed Size (px)
Citation preview

lable at ScienceDirect
European Journal of Soil Biology 62 (2014) 60e65
Contents lists avai
European Journal of Soil Biology
journal homepage: http: / /www.elsevier .com/locate/ejsobi
Original article
Allocation of photosynthetic carbon to nodules of soybean in threegeographically different Mollisols
Yuan Chen a,b, Zhenhua Yu b, Jifeng Wang c, Xingyi Zhang a,b,*
a Forestry Academy, Northeast Forestry University, Harbin 150040, ChinabKey Laboratory of Mollisols Agroecology, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Harbin 150081, ChinacDaxing’anling Academy of Agriculture and Forestry, Jiagedaqi, Heilongjiang Province 165000, China
a r t i c l e i n f o
Article history:Received 16 August 2013Received in revised form30 January 2014Accepted 17 February 2014Available online 17 March 2014Handling editor: Kristina Lindström
Keywords:Carbon contentMollisolsPhotosynthetic 13CRootSoybean nodule
* Corresponding author. Chinese Academy of SciGeography and Agroecology, Key Laboratory of M150081, China. Tel.: þ86 0451 86602926; fax: þ86 04
E-mail address: [email protected] (X. Zha
http://dx.doi.org/10.1016/j.ejsobi.2014.02.0071164-5563/� 2014 Elsevier Masson SAS. All rights res
a b s t r a c t
The development of legume nodules and N2-fixation demand fair proportion of photosynthetic carbon(C). Soil as a determinant of nodulation may influence C allocation to nodule and subsequent under-ground C fates. However, there is little information on the photosynthetic C allocation to the nodule ofsoybean grown in Mollisols in Northeast China. Such research is crucial for optimizing the strategies ofC/N management to improve nodulation and productivity in soybean farming systems. With a potexperiment, soybean plants were grown in three Mollisols sampled from low- to high-latitude inNortheast China and labeled with 13CO2 at the R4 (full pod) and R5 (initial pod filling) stages. The nodulecharacteristics and underground 13C distribution were investigated. The nodule number and noduledensity were in the order of Mid- > High- > Low-latitude Mollisol, resulting in 980, 578 and 252 nodulesper plant, and 11.1, 8.2 and 2.7 nodules per m of root length, respectively. The 13CO2 pulse-chase labelingshowed that higher proportion of 13C was recovered in Mid-latitude Mollisol at R4, and in both Mid- andHigh-latitude at R5. Moreover, a 53.8% of increase on C-growth rate of nodule was also found in the twosoils compared to Low-latitude. This suggests that nodules in high-latitude Mollisols have strongerC-sink activity than low-latitude ones, which attribute to nodule number and C-growth rate of nodule.The nodules accounted for 15.7%, 28.0% and 11.5% of net underground 13C in Low-, Mid- and High-latitude Mollisols, respectively. Therefore, the extent of nodule-driving underground C allocation var-ied with Mollisols from geographically different regions.
� 2014 Elsevier Masson SAS. All rights reserved.
1. Introduction
Nodulation and N2 fixation are closely related to carbohydratesupply by photosynthesis [1,2]. When nodule develops, it graduallybecomes a strong sink of photosynthate for the energy source. Itoet al. [3] reported that the carbon (C) allocation to the inoculatedroots did not increase during the early stages of nodule formation,while it markedly increased after emergence of the nodules.Therefore, the great demand of C in nodule may occur when noduleenters linear growth stages.
Soybean as a legume species is the major crop in NortheastChina with the sown acreage accounting for 33% of the nation’stotal [4e6]. Mollisols is the main farming soil type there andamounts to more than 4 million hectares [4]. However, the chem-ical and biological properties of Mollisols such as organic C, pH and
ences Northeast Institute ofollisols Agroecology, Harbin51 86603736.ng).
erved.
microbial activity, are various with geographic locations due tobiotic (plants) and abiotic factors (temperature and rainfall) [4].These discrepancies on soil properties may influence the nodula-tion. Lindström et al. [7] indicates that the nodulation varies whenlegumes grown in different soils. Studies found that the diversity ofrhizobium species isolated from bean nodules in Austrian, Mexicanand French soils were different, which affected the infectioneffectiveness and nodule characteristics [8,9]. Soils from differentlocalizations where the temperature and rainfall were different,had different communities of rhizobium [10,11]. Therefore, soiltypes or soils from separated locations may affect biomass andnumber of nodules and N2 fixation. Consequently, the demand of Cfor the development and function of nodules may change withsoils, especially when nodules are at fast growing stages. To ourknowledge, the information on the soybean photosynthetic Callocation to nodule in geographically different Mollisols is poorlyunderstood, which is essential for how to optimize N2 fixation bysoybean to save fossil N in the perspective of sustainable agricul-ture in Northeast China.

Y. Chen et al. / European Journal of Soil Biology 62 (2014) 60e65 61
The 13CO2 labeling techniques include continuous labeling andpulse labeling are widely used to trace photosynthetic C [12e19].With the labeling techniques, most studies focused on the under-ground fate of photosynthates in agricultural cereals and pasturespecies [20], while much less is known about N2-fixed soybeanplants grown inMollisols. Since pulse-labeling can be implementedto determine the seasonal dynamics of assimilate partitioning[21,22], and the residence time in various carbon pools [23,24], the13CO2 pulse-labeling was used in this experiment to quantitativelyanalyze the photosynthetic 13C distribution to nodule. The aims ofthis study are (1) to determine the nodule characteristics in threeMollisols from low- to high-latitude of the soil distributed region,and (2) subsequently quantify the differentiation of C allocation tothe nodules in soils.
2. Materials and methods
2.1. Soil and plant materials
Three soils used in this study were taken from low- to high-latitude regions of Mollisols (named as Low-, Mid- and High-latitude Mollisol, respectively) in Northeast China. The soil fromeach location was air-dried, sieved through a 4 mm sieve, andmixed. The general chemistry characteristics of soil samples wereshowed in Table 1.
A soybean cultivar, i.e. Suinong 14 was used in this experiment.It has beenwidely grown over 2million ha and producedmore than937 million kg in China [25,26].
2.2. Plant culture
A pot experiment was conducted at Northeast Institute of Ge-ography and Agroecology (45�41.80 N, 126�38.10 E), Chinese Acad-emy of Sciences, Harbin, China. The experiment was completelyrandomized block design with three Mollisols, and each receivedsix pots for labeling and another six as control. Twelve kilograms ofa soil/sand mixture (1:1 w/w) was filled into each container(29.0 cm in height and 25.5 cm in diameter). The reason for mixingsoil with sand in this studywas to facilitate root washing at harvest.Basal nutrients were applied at the following rates (mg kg�1): 217urea, 219 KH2PO4, 167 CaCl2.2H2O, 43 MgSO4.7H2O, 9 Fe-EDTA, 6ZnSO4 and 5 CuSO4, 0.7H3BO3, 6.7 MnSO4.H2O, 10 ZnSO4.7H2O, 2CuSO4.5H2O, 0.3 CoSO4.7H2O, 0.2 Na2MoO4.2H2O [27]. Nutrientswere thoroughly mixed with the soil and sand. Nine seeds weresown to each pot on May 10, 2009, and the plants were thinned tothree per pot on the 14th day after sowing. The plants were grownin a glasshouse with night temperature of 16e20 �C and day tem-perature of 24e28 �C during the experiment. Water content in soilwas well controlled at 80 � 5% of field water capacity.
2.3. Labeling process and measurements
Soybean plants were labeled with 13CO2 on the R4 (full pod)stage when nodules rapidly grow, and the R5 (initial pod filling)stage when nodules reach maximum biomass. On labeling dates,
Table 1General chemical characteristics of three geographically different Mollisols in Northeast
Mollisols Sites Locations Corg (%)
Low-latitude Yushu, Jilin 43�200 N, 124�300E 1.04Mid-latitude Hailun, Heilongjiang 47�320 N, 126�570 E 2.90High- latitude Beian, Heilongjiang 48�170 N, 127�150 E 5.05
plants were moved into an airtight glass chamber (area130 � 80 cm2, height 130 cm). Before labeling, CO2-free air waspumped through the chamber for 30 min to eliminate 12CO2 inside.After that, five hundred ml of pure 13CO2 (Chemical purity � 99.9%)was injected with a syringe through a rubber gasket into thechamber [28]. The average 13CO2 concentration maintained about350e400 ppm. Plants were labeled for 6 h in the chamber where afan was installed to homogenize the atmosphere inside. After la-beling, plants in three pots were harvested at intervals of 0 and 12days, respectively. Plants without 13C labeling were harvested ascontrol at the same time. The entire root system was carefullyremoved from the pot after shoots were cut off at the soil surface.The roots were washed with tap water to remove soil and sandadhering roots. Nodules on root system were picked off, countedand weighed. The measurement of root length was performed withWin-RHIZO version 2004a (Regent Instruments, Quebec, Canada).All plant samples were dried at 70 �C for 72 h and then groundbefore further analysis. Soil samples were took at the same time,air-dried and ground.
The C concentrations and stable 13C isotope ratios of all sampleswere determined with an isotope ratio mass spectrometer (Delta-plus, Finnigan MAT GmbH, Bremen, Germany). The natural abun-dance of 13C isotopes was presented as ratio per thousands relativeto the international standard VPBD using delta units (d).
2.4. Calculation
Atomic 13C% was expressed as:
Atomic 13C% ¼ ½½ðd13Cþ 1000Þ � RVPBD�=½ðd13Cþ 1000Þ�� RVPBD þ 1� � 100
where R ¼ 13C/12C and RVPBD ¼ 0.0111802 [29].The net photosynthetic 13C in roots, nodules and soils was the
difference between 13C contents of the labeled samples and those ofthe nonlabeled samples as follows:
Csample ¼ �ðatomic 13C%Þl � ðatomic 13C%Þnl�sample � TCsample
� 100
where “l” and “nl” indicate labeling and nonlabeling; “TC” is thetotal C content of sample.
By immediate investigation after labeling (0 day), the sum of the13C amounts in shoots, roots, nodules and soils was assumed as thetotal 13C photosynthetic by plants during 6-h 13C labeling [30].
The percentage distribution of 13C at each harvesting time wasestimated as [30]:
Distribution% ¼ 13Csample=net13C assimilation
The underground 13C allocation was sum of 13Croot, 13Cnodule,and 13Csoil.
The C-growth rates of root and nodule were calculated accord-ing to the C accumulation of those during the interval of the two-harvest at each growth stage which was divided by 12 days.
China.
pH (1:5 v/v) Total (g kg�1) Available (mg kg�1)
N P K N P K
7.1 0.80 0.31 16 75 24.0 1007.1 1.72 0.78 16 154 37.1 1606.1 3.65 0.92 12 266 32.5 130
tnalpreprebmun
eludoN
0
200
400
600
800
1000
1200
1400
1600Low-latitude MollisolMid-latitude MollisolHigh-latitude Mollisol
htgneltoorfomrepreb
muneludo
N 0
2
4
6
8
10
12
14
tnalpg(
WFel udo
N-1
)
0
1
2
3
4
5
6
Growth stagesR4 R5
tnalpg
m(WF
elu do neludi vi dnI
-1)
02468
1012141618
A
B
C
D
c
a
b
b
aa
c
a
b
b
a a
b
a
b
b
aa
a
bc
a
bb
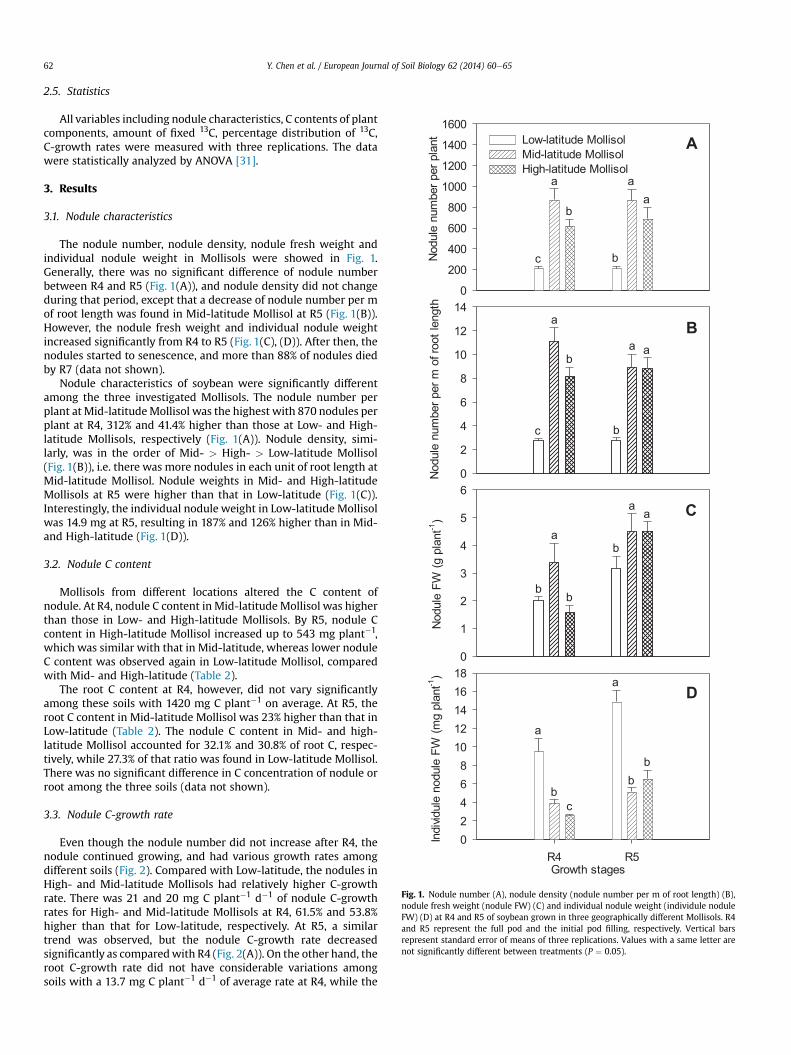
Fig. 1. Nodule number (A), nodule density (nodule number per m of root length) (B),nodule fresh weight (nodule FW) (C) and individual nodule weight (individule noduleFW) (D) at R4 and R5 of soybean grown in three geographically different Mollisols. R4and R5 represent the full pod and the initial pod filling, respectively. Vertical barsrepresent standard error of means of three replications. Values with a same letter arenot significantly different between treatments (P ¼ 0.05).
Y. Chen et al. / European Journal of Soil Biology 62 (2014) 60e6562
2.5. Statistics
All variables including nodule characteristics, C contents of plantcomponents, amount of fixed 13C, percentage distribution of 13C,C-growth rates were measured with three replications. The datawere statistically analyzed by ANOVA [31].
3. Results
3.1. Nodule characteristics
The nodule number, nodule density, nodule fresh weight andindividual nodule weight in Mollisols were showed in Fig. 1.Generally, there was no significant difference of nodule numberbetween R4 and R5 (Fig. 1(A)), and nodule density did not changeduring that period, except that a decrease of nodule number per mof root length was found in Mid-latitude Mollisol at R5 (Fig. 1(B)).However, the nodule fresh weight and individual nodule weightincreased significantly from R4 to R5 (Fig. 1(C), (D)). After then, thenodules started to senescence, and more than 88% of nodules diedby R7 (data not shown).
Nodule characteristics of soybean were significantly differentamong the three investigated Mollisols. The nodule number perplant at Mid-latitudeMollisol was the highest with 870 nodules perplant at R4, 312% and 41.4% higher than those at Low- and High-latitude Mollisols, respectively (Fig. 1(A)). Nodule density, simi-larly, was in the order of Mid- > High- > Low-latitude Mollisol(Fig. 1(B)), i.e. there was more nodules in each unit of root length atMid-latitude Mollisol. Nodule weights in Mid- and High-latitudeMollisols at R5 were higher than that in Low-latitude (Fig. 1(C)).Interestingly, the individual nodule weight in Low-latitude Mollisolwas 14.9 mg at R5, resulting in 187% and 126% higher than in Mid-and High-latitude (Fig. 1(D)).
3.2. Nodule C content
Mollisols from different locations altered the C content ofnodule. At R4, nodule C content in Mid-latitude Mollisol was higherthan those in Low- and High-latitude Mollisols. By R5, nodule Ccontent in High-latitude Mollisol increased up to 543 mg plant�1,which was similar with that in Mid-latitude, whereas lower noduleC content was observed again in Low-latitude Mollisol, comparedwith Mid- and High-latitude (Table 2).
The root C content at R4, however, did not vary significantlyamong these soils with 1420 mg C plant�1 on average. At R5, theroot C content in Mid-latitude Mollisol was 23% higher than that inLow-latitude (Table 2). The nodule C content in Mid- and high-latitude Mollisol accounted for 32.1% and 30.8% of root C, respec-tively, while 27.3% of that ratio was found in Low-latitude Mollisol.There was no significant difference in C concentration of nodule orroot among the three soils (data not shown).
3.3. Nodule C-growth rate
Even though the nodule number did not increase after R4, thenodule continued growing, and had various growth rates amongdifferent soils (Fig. 2). Compared with Low-latitude, the nodules inHigh- and Mid-latitude Mollisols had relatively higher C-growthrate. There was 21 and 20 mg C plant�1 d�1 of nodule C-growthrates for High- and Mid-latitude Mollisols at R4, 61.5% and 53.8%higher than that for Low-latitude, respectively. At R5, a similartrend was observed, but the nodule C-growth rate decreasedsignificantly as comparedwith R4 (Fig. 2(A)). On the other hand, theroot C-growth rate did not have considerable variations amongsoils with a 13.7 mg C plant�1 d�1 of average rate at R4, while the
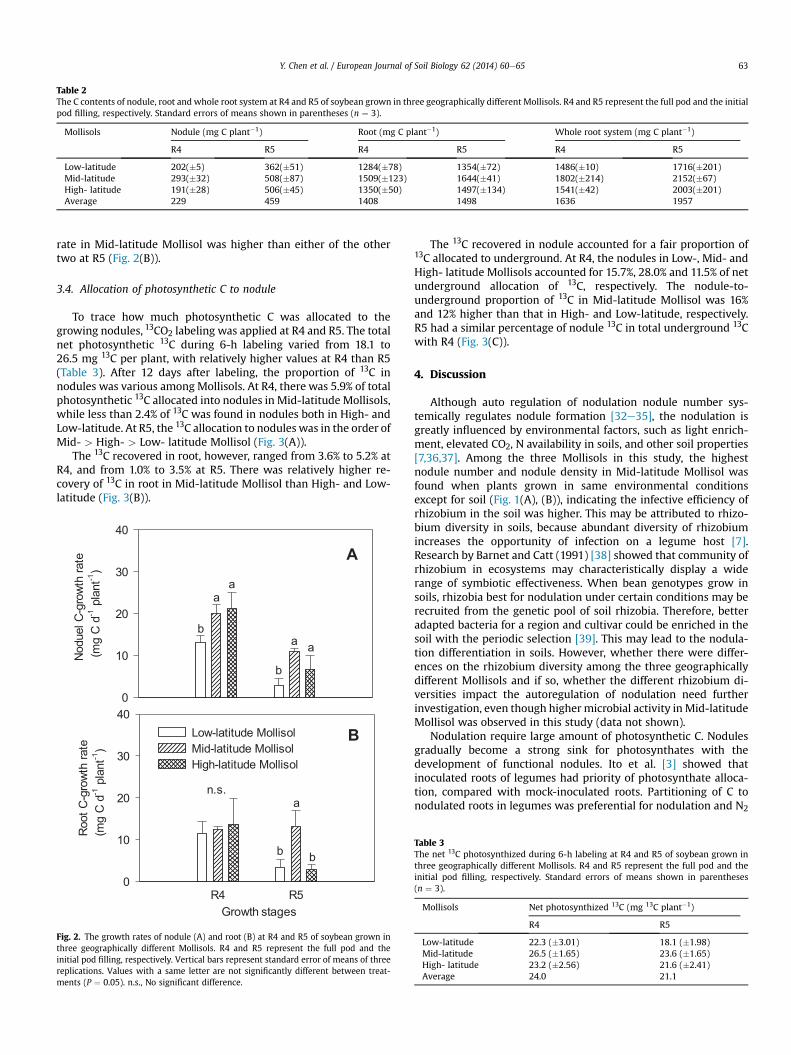
Table 2The C contents of nodule, root and whole root system at R4 and R5 of soybean grown in three geographically different Mollisols. R4 and R5 represent the full pod and the initialpod filling, respectively. Standard errors of means shown in parentheses (n ¼ 3).
Mollisols Nodule (mg C plant�1) Root (mg C plant�1) Whole root system (mg C plant�1)
R4 R5 R4 R5 R4 R5
Low-latitude 202(�5) 362(�51) 1284(�78) 1354(�72) 1486(�10) 1716(�201)Mid-latitude 293(�32) 508(�87) 1509(�123) 1644(�41) 1802(�214) 2152(�67)High- latitude 191(�28) 506(�45) 1350(�50) 1497(�134) 1541(�42) 2003(�201)Average 229 459 1408 1498 1636 1957
Y. Chen et al. / European Journal of Soil Biology 62 (2014) 60e65 63
rate in Mid-latitude Mollisol was higher than either of the othertwo at R5 (Fig. 2(B)).
3.4. Allocation of photosynthetic C to nodule
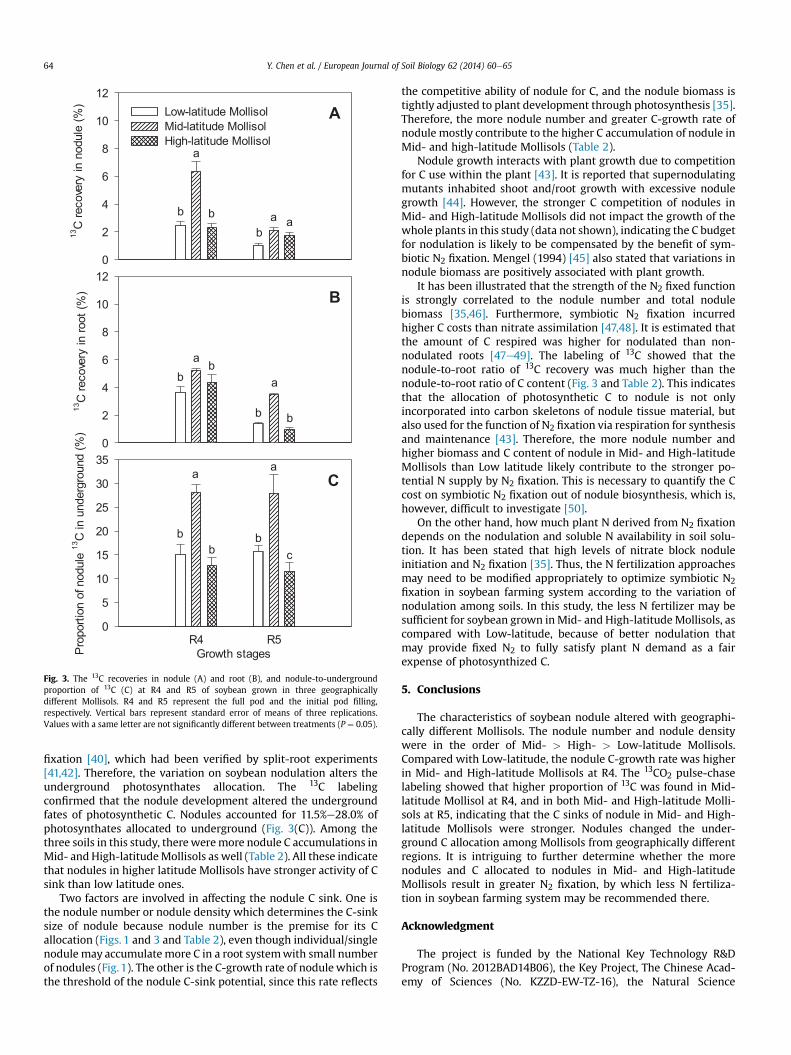
To trace how much photosynthetic C was allocated to thegrowing nodules, 13CO2 labeling was applied at R4 and R5. The totalnet photosynthetic 13C during 6-h labeling varied from 18.1 to26.5 mg 13C per plant, with relatively higher values at R4 than R5(Table 3). After 12 days after labeling, the proportion of 13C innodules was various among Mollisols. At R4, there was 5.9% of totalphotosynthetic 13C allocated into nodules in Mid-latitude Mollisols,while less than 2.4% of 13C was found in nodules both in High- andLow-latitude. At R5, the 13C allocation to nodules was in the order ofMid- > High- > Low- latitude Mollisol (Fig. 3(A)).
The 13C recovered in root, however, ranged from 3.6% to 5.2% atR4, and from 1.0% to 3.5% at R5. There was relatively higher re-covery of 13C in root in Mid-latitude Mollisol than High- and Low-latitude (Fig. 3(B)).
Growth stagesR4 R5
etarht
worg-Cto o
Rd
Cg
m(-1
tnalp-1
)
0
10
20
30
40
etarht
worg-Cleu do
Nd
Cg
m(-1
t nal p-1
)
0
10
20
30
40
Low-latitude MollisolMid-latitude MollisolHigh-latitude Mollisol
A
B
b
aa
b
a a
b
a
b
n.s.
Fig. 2. The growth rates of nodule (A) and root (B) at R4 and R5 of soybean grown inthree geographically different Mollisols. R4 and R5 represent the full pod and theinitial pod filling, respectively. Vertical bars represent standard error of means of threereplications. Values with a same letter are not significantly different between treat-ments (P ¼ 0.05). n.s., No significant difference.
The 13C recovered in nodule accounted for a fair proportion of13C allocated to underground. At R4, the nodules in Low-, Mid- andHigh- latitude Mollisols accounted for 15.7%, 28.0% and 11.5% of netunderground allocation of 13C, respectively. The nodule-to-underground proportion of 13C in Mid-latitude Mollisol was 16%and 12% higher than that in High- and Low-latitude, respectively.R5 had a similar percentage of nodule 13C in total underground 13Cwith R4 (Fig. 3(C)).
4. Discussion
Although auto regulation of nodulation nodule number sys-temically regulates nodule formation [32e35], the nodulation isgreatly influenced by environmental factors, such as light enrich-ment, elevated CO2, N availability in soils, and other soil properties[7,36,37]. Among the three Mollisols in this study, the highestnodule number and nodule density in Mid-latitude Mollisol wasfound when plants grown in same environmental conditionsexcept for soil (Fig. 1(A), (B)), indicating the infective efficiency ofrhizobium in the soil was higher. This may be attributed to rhizo-bium diversity in soils, because abundant diversity of rhizobiumincreases the opportunity of infection on a legume host [7].Research by Barnet and Catt (1991) [38] showed that community ofrhizobium in ecosystems may characteristically display a widerange of symbiotic effectiveness. When bean genotypes grow insoils, rhizobia best for nodulation under certain conditions may berecruited from the genetic pool of soil rhizobia. Therefore, betteradapted bacteria for a region and cultivar could be enriched in thesoil with the periodic selection [39]. This may lead to the nodula-tion differentiation in soils. However, whether there were differ-ences on the rhizobium diversity among the three geographicallydifferent Mollisols and if so, whether the different rhizobium di-versities impact the autoregulation of nodulation need furtherinvestigation, even though highermicrobial activity in Mid-latitudeMollisol was observed in this study (data not shown).
Nodulation require large amount of photosynthetic C. Nodulesgradually become a strong sink for photosynthates with thedevelopment of functional nodules. Ito et al. [3] showed thatinoculated roots of legumes had priority of photosynthate alloca-tion, compared with mock-inoculated roots. Partitioning of C tonodulated roots in legumes was preferential for nodulation and N2
Table 3The net 13C photosynthized during 6-h labeling at R4 and R5 of soybean grown inthree geographically different Mollisols. R4 and R5 represent the full pod and theinitial pod filling, respectively. Standard errors of means shown in parentheses(n ¼ 3).
Mollisols Net photosynthized 13C (mg 13C plant�1)
R4 R5
Low-latitude 22.3 (�3.01) 18.1 (�1.98)Mid-latitude 26.5 (�1.65) 23.6 (�1.65)High- latitude 23.2 (�2.56) 21.6 (�2.41)Average 24.0 21.1
13)
%(eludon
niyrevocer
C
0
2
4
6
8
10
12Low-latitude MollisolMid-latitude MollisolHigh-latitude Mollisol
13)
%(toorni
yr evocerC
0
2
4
6
8
10
12
Growth stagesR4 R5
eludonfono itropor
P13
)%(
dn uorgrednuni
C
0
5
10
15
20
25
30
35
A
B
C
b
a
bb
a a
ba
b
b
a
b
b
a
bb
a
c
Fig. 3. The 13C recoveries in nodule (A) and root (B), and nodule-to-undergroundproportion of 13C (C) at R4 and R5 of soybean grown in three geographicallydifferent Mollisols. R4 and R5 represent the full pod and the initial pod filling,respectively. Vertical bars represent standard error of means of three replications.Values with a same letter are not significantly different between treatments (P ¼ 0.05).
Y. Chen et al. / European Journal of Soil Biology 62 (2014) 60e6564
fixation [40], which had been verified by split-root experiments[41,42]. Therefore, the variation on soybean nodulation alters theunderground photosynthates allocation. The 13C labelingconfirmed that the nodule development altered the undergroundfates of photosynthetic C. Nodules accounted for 11.5%e28.0% ofphotosynthates allocated to underground (Fig. 3(C)). Among thethree soils in this study, thereweremore nodule C accumulations inMid- and High-latitudeMollisols as well (Table 2). All these indicatethat nodules in higher latitude Mollisols have stronger activity of Csink than low latitude ones.
Two factors are involved in affecting the nodule C sink. One isthe nodule number or nodule density which determines the C-sinksize of nodule because nodule number is the premise for its Callocation (Figs. 1 and 3 and Table 2), even though individual/singlenodule may accumulatemore C in a root systemwith small numberof nodules (Fig. 1). The other is the C-growth rate of nodule which isthe threshold of the nodule C-sink potential, since this rate reflects
the competitive ability of nodule for C, and the nodule biomass istightly adjusted to plant development through photosynthesis [35].Therefore, the more nodule number and greater C-growth rate ofnodule mostly contribute to the higher C accumulation of nodule inMid- and high-latitude Mollisols (Table 2).
Nodule growth interacts with plant growth due to competitionfor C use within the plant [43]. It is reported that supernodulatingmutants inhabited shoot and/root growth with excessive nodulegrowth [44]. However, the stronger C competition of nodules inMid- and High-latitude Mollisols did not impact the growth of thewhole plants in this study (data not shown), indicating the C budgetfor nodulation is likely to be compensated by the benefit of sym-biotic N2 fixation. Mengel (1994) [45] also stated that variations innodule biomass are positively associated with plant growth.
It has been illustrated that the strength of the N2 fixed functionis strongly correlated to the nodule number and total nodulebiomass [35,46]. Furthermore, symbiotic N2 fixation incurredhigher C costs than nitrate assimilation [47,48]. It is estimated thatthe amount of C respired was higher for nodulated than non-nodulated roots [47e49]. The labeling of 13C showed that thenodule-to-root ratio of 13C recovery was much higher than thenodule-to-root ratio of C content (Fig. 3 and Table 2). This indicatesthat the allocation of photosynthetic C to nodule is not onlyincorporated into carbon skeletons of nodule tissue material, butalso used for the function of N2 fixation via respiration for synthesisand maintenance [43]. Therefore, the more nodule number andhigher biomass and C content of nodule in Mid- and High-latitudeMollisols than Low latitude likely contribute to the stronger po-tential N supply by N2 fixation. This is necessary to quantify the Ccost on symbiotic N2 fixation out of nodule biosynthesis, which is,however, difficult to investigate [50].
On the other hand, how much plant N derived from N2 fixationdepends on the nodulation and soluble N availability in soil solu-tion. It has been stated that high levels of nitrate block noduleinitiation and N2 fixation [35]. Thus, the N fertilization approachesmay need to be modified appropriately to optimize symbiotic N2fixation in soybean farming system according to the variation ofnodulation among soils. In this study, the less N fertilizer may besufficient for soybean grown inMid- and High-latitudeMollisols, ascompared with Low-latitude, because of better nodulation thatmay provide fixed N2 to fully satisfy plant N demand as a fairexpense of photosynthized C.
5. Conclusions
The characteristics of soybean nodule altered with geographi-cally different Mollisols. The nodule number and nodule densitywere in the order of Mid- > High- > Low-latitude Mollisols.Compared with Low-latitude, the nodule C-growth rate was higherin Mid- and High-latitude Mollisols at R4. The 13CO2 pulse-chaselabeling showed that higher proportion of 13C was found in Mid-latitude Mollisol at R4, and in both Mid- and High-latitude Molli-sols at R5, indicating that the C sinks of nodule in Mid- and High-latitude Mollisols were stronger. Nodules changed the under-ground C allocation among Mollisols from geographically differentregions. It is intriguing to further determine whether the morenodules and C allocated to nodules in Mid- and High-latitudeMollisols result in greater N2 fixation, by which less N fertiliza-tion in soybean farming system may be recommended there.
Acknowledgment
The project is funded by the National Key Technology R&DProgram (No. 2012BAD14B06), the Key Project, The Chinese Acad-emy of Sciences (No. KZZD-EW-TZ-16), the Natural Science

Y. Chen et al. / European Journal of Soil Biology 62 (2014) 60e65 65
Foundation of Heilongjiang Province (No. C200902), and theOversea-Scholar-Return Fund (2011-1568).
References
[1] P.M. Gresshoff, G. Caetano-Akao, Systemic regulation of nodulation in le-gumes, in: P.M. Gresshoff (Ed.), Plant Biotechnology and Development, CRCPress, Boca Raton, FL, U.S.A, 1991, pp. 87e100.
[2] K.B. Walsh, K.J. Vessey, D.B. Layzel, Carbohydrate supply and N2 fixation insoybean. The effect of varied daylength and stem girdling, Plant Physiol. 85(1987) 137e144.
[3] S. Ito, N. Ohtake, K. Sueyoshi, T. Ohyamaall, Allocation of photosyntheticproducts in soybean during the early stages of nodule formation, Soil. Sci.Plant Nutr. 52 (2006) 438e443.
[4] J.L. Wang, Q.K. Yang, Z.P. Wu (Eds.), Soybean in Northeast China, HeilongjiangScience Press, Harbin, China, 2000, pp. pp.8e58 (In Chinese).
[5] X.B. Liu, S.J. Herbert, Fifteen years of research examining cultivation ofcontinuous soybean in Northeast China, Field Crops Res. 79 (2002) 1e7.
[6] B. Liu, X.B. Liu, C. Wang, J. Jin, S.J. Herbert, M. Hashemi, Responses of soybeanyield and yield components to light enrichment and planting density, Int. J.Plant Prod. 4 (2010) 1e9.
[7] M. Lindström, M. Murwira, A. Willems, N. Altier, The biodiversity of beneficialmicrobe-host mutualism: the case of rhizobia, Res. Microbiol. 161 (2010)453e463.
[8] A. Sessitsch, H. Ramírez-Saad, G. Hardarson, A.D.L. Akkermans, W.M. de Vos,Classification of Austrian rhizobia and the Mexican isolate FL27 obtained fromPhaseolus vulgaris L. as Rhizobium gallicum, Int. J. Syst. Bacteriol. 47 (1997)1097e1101.
[9] N. Amarger, V. Macheret, G. Laguerre, Rhizobium gallicum sp. nov. andRhizobium giardinii sp. nov., from Phaseolus vulgaris nodules, Int. J. Syst. Bac-teriol. 47 (1997) 996e1006.
[10] P.H. Graham, K.J. Draeger, M.L. Ferrey, M.J. Conroy, B.E. Hammer, E. Martínez,S.R. Aarons, C. Quinto, Acid pH tolerance in strains of Rhizobium and Bra-dyrhizobium, and initial studies on the basis for acid tolerance of Rhizobiumtropici UMR1899, Can. J. Microbiol. 40 (1994) 198e207.
[11] J. Michiels, C. Verreth, J. Vanderleyden, Effects of temperature stress on bean-nodulating Rhizobium strains, Appl. Environ. Microbiol. 60 (1994) 1206e1212.
[12] A. De Neergaard, J. Magid, Influence of the rhizosphere on microbial biomassand recently formed organic matter, Eur. J. Soil. Sci. 52 (2001) 377e384.
[13] R.I. Griffiths, M. Manefield, N. Ostle, N. McNamara, A.G. O’Donnell, M.J. Bailey,A.S. Whiteley, 13CO2 pulse labeling of plants in tandem with stable isotopeprobing: methodological considerations for examining microbial function inthe rhizosphere, J. Microbiol. Methods 58 (2004) 119e129.
[14] Y. Kuzyakov, W. Cheng, Photosynthesis controls of rhizo-sphere respirationand organic matter decomposition, Soil. Biol. Biochem. 33 (2001) 1915e1925.
[15] W. Merbach, E. Mirus, G. Knof, R. Remus, S. Ruppel, R. Russow, A. Gransee,J. Schulze, Release of carbon and nitrogen compounds by plant roots and theirpossible ecological importance, J. Plant Nutr. Soil. Sci. 162 (1999) 373e383.
[16] J.M. Whipps, Carbon loss from the roots of tomato and pea seedlings grown insoil, Plant Soil. 103 (1987) 95e100.
[17] C. Nguyen, Rhizodeposition of organic C by plants: mechanisms and controls,Agronomie 23 (2003) 375e396.
[18] Y. Kuzyakov, K. Schneckenberger, Review of estimation of plant rhizodepo-sition and their contribution to soil organic matter formation, Arch. Agron.Soil. Sci. 50 (2004) 115e132.
[19] M. Werth, Y. Kuzyakov, Root-derived carbon in soil respiration and microbialbiomass determination by 14C and 13C, Soil. Biol. Biochem. 40 (2008) 625e637.
[20] J. Pausch, Y. Kuzyakov, Photoassimilate allocation and dynamics of hotspots inroots visualized by 14C phosphor imaging, J. Plant Nutr. Soil. Sci. 174 (2011)12e19.
[21] J.K. Martin, J.R. Kemp, The measurement of C transfers within the rhizosphereof wheat grown filed plots, Soil. Biol. Biochem. 18 (1986) 103e107.
[22] J. Swinnen, J.A. Van Veen, R. Merckx, Rhizosphere carbon fluxes in field-grownspring wheat: model calculations based on 14C partitioning after pulse-la-beling, Soil. Biol. Biochem. 26 (1994) 171e182.
[23] N.K. Ruehr, C.A. Offermann, A. Gessler, J.B. Winkler, J.P. Ferrio, N. Bunchmann,R.L. Barnard, Drought effects on allocation of recent carbon: from beech leavesto soil CO2 efflux, New. Phytol. 184 (2009) 950e961.
[24] E.M. Powers, J.D. Marshall, Pulse labeling of dissolved 13C-carbonate into treexylem: developing a new method to determine the fate of recently fixedphotosynthate, Rapid Commun. Mass Spectrom. 25 (2010) 33e40.
[25] J. Qin, R.Q. Yang, C.X. Jiang, W.B. Li, Y.H. Li, R.X. Guan, R.Z. Chang, L.J. Qiu,Discovery and transmission of functional QTL in the pedigree of an elitesoybean cultivar Suinong 14, Plant Breed. 129 (2010) 235e242.
[26] J. Jin, X.B. Liu, G.H. Wang, L. Mi, Z.B. Shen, X.L. Chen, S.J. Herbert, Agronomicand physiologic contributions to the yield improvement of soybean cultivarsreleased from 1950 to 2006 in Northeast China, Field Crop Res. 115 (2010a)116e123.
[27] J. Jin, G.H. Wang, X.B. Liu, L. Mi, Y.H. Li, Y.X. Xu, S.J. Herbert, Geneticimprovement of yield shapes the temporal and spatial root morphology ofsoybean (Glycine max) grown in north-east China, N. Z. J. Crop Hortic. Sci. 38(2010b) 177e188.
[28] K.D. Subedi, B.L. Ma, B.C. Liang, New method to estimate root biomass in soilthrough root-derived carbon, Soil. Biol. Biochem. 38 (2006) 2212e2218.
[29] R.A. Werner, W.A. Brand, Referencing strategies and techniques in stableisotope ratio analysis, Rapid Commun. Mass Spectrom. 15 (2001) 501e519.
[30] Y.H. Lu, A. Watanabe, M. Kimura, Input and distribution of photosyntheticcarbon in a flooded rice soil, Global Biogeochem. Cycles 16 (2002) 321e328.
[31] SAS Institute, SAS User’s Guide: Statistics. Version 6.12, SAS Institute, Cary, NC,U.S.A, 1994.
[32] T.V. Bhuvaneswari, G.B. Turgeon, W.D. Bauer, Early events in the infection ofsoybean (Glycine max L. Merr.) by Rhizobium japonicum I. Localization ofinfectible root cells, Plant Physiol. 66 (1980) 1027e1031.
[33] M. Pierce, W.D. Bauer, A rapid regulatory response governing nodulation insoybean, Plant Physiol. 73 (1983) 286e290.
[34] R.M. Kosslak, B.B. Bohlool, Suppression of nodule development on one side ofa split root system of soybeans caused by prior inoculation of the other side,Plant Physiol. 75 (1984) 125e130.
[35] A.S. Voisin, N.G. Munier-Jolain, C. Salon, The nodulation process is tightlyadjusted to plant growth. An analysis using environmentally and geneticallyinduced variation of nodule number and biomass in pea, Plant Soil. 337 (2010)399e412.
[36] M. Bacanamwo, J.E. Harper, Response of a hypernodulating soybean mutant toincreased photosynthate supply, Plant Sci. 124 (1997) 119e129.
[37] G. Kaschuk, M. Hungria, P.A. Leffelaar, K.E. Giller, T.W. Kuyper, Differences inphotosynthetic behaviour and leaf senescence of soybean (Glycine max [L.]Merrill) dependent on N2 fixation or nitrate supply, Plant Biol. 12 (2010) 60e69.
[38] Y.M. Barnet, P.C. Catt, Distribution and characteristics of root-nodule bacteriaisolated from Australian Acacia spp, Plant. Soil. 135 (1991) 109e120.
[39] E. Martínez-Romero, Diversity of Rhizobium-Phaseolus vulgaris symbiosis:overview and perspectives, Plant. Soil. 252 (2003) 11e23.
[40] J.I. Hacin, B.B. Bohlool, P.W. Singleton, Partitioning of 14C-labelled photosyn-thate to developing nodulates and roots of soybean (Glycine max), New.Phytol. 137 (1997) 257e265.
[41] J.E. Olsson, P. Nakao, B.B. Bohlool, P.M. Gresshoff, Lack of systemic suppressionof nodulation in split root system of supernodulating soybean (Glycine max[L.] Merr.) mutants, Plant Physiol. 90 (1989) 1347e1352.
[42] P.W. Singleton, C.V. Kessel, Effect of localized nitrogen availability to soybeanhalf-root systems on photosynt-hate partitioning to roots and nodules, PlantPhysiol. 83 (1987) 552e556.
[43] A.S. Voisin, C. Salon, C. Jeudy, F.R. Warembourg, Root and nodule growth inPisum sativum L. in relation to photosynthesis: analysis using 13C-labelling,Ann. Bot. 92 (2003a) 557e563.
[44] M. Sagan, G. Duc, Sym28 and Sym29, two new genes involved in regulation ofnodulation in pea (Pisum sativum L.), Symbiosis 20 (1996) 229e245.
[45] R. Mengel, Symbiotic dinitrogen fixationdIts dependence on plant nutritionand its ecophysiological impact, Z. Pflanzenernahr Bodenkd 157 (1994) 233e241.
[46] A.S. Voisin, C. Salon, C. Jeudy, F.R. Warembourg, Symbiotic N2 fixation activityin relation to C economy of Pisum sativum L. as a function of plant phenology,J. Exp. Bot. 54 (2003b) 2733e2744.
[47] C.A. Atkins, J.S. Pate, G.J. Griffiths, S.T. White, Economy of carbon and nitrogenin nodulated and non nodulated (NO3
- grown) cowpea (Vigna unguiculata L.Walp.), Plant Physiol. 66 (1980) 978e983.
[48] J. Schulze, E. Adgo, W. Merbach, Carbon costs associated with N2 fixation inVicia faba L. and Pisum sativum L. over a 14-day period, Plant Biol. 1 (1999)625e631.
[49] J.S. Pater, D.B. Layzell, C.A. Atkins, Economy of carbon and nitrogen in anodulated and non nodulated (NO3
- grown) legume, Plant Physiol. 64 (1979)1083e1088.
[50] D.A. Phillips, Efficiency of symbiotic nitrogen fixation in legumes, Annu. Rev.Plant Physiol. 31 (1980) 29e49.





























