Embed Size (px)
Citation preview

104 Biochimica et Biophvsica Acta, 758 (1983) 104 113 Elsevier
BBA 21483
A F R I C A N T R Y P A N O S O M E S C O N T A I N CALMODULIN WHICH IS DISTINCT FROM HOST CALMODULIN
LARRY RUBEN, CHARLES EGWUAGU and CURTIS L. PATTON
Department of Epidemiology and Public Health, Yale University, 60 College Street, New Haven, CT 06510 (U.S.A.)
(Received January 5th, 1983) (Revised manuscript received March 22nd, 1983)
Key words: Calmodulin; Ca -binding protein," (Trypanosome)
Studies were initiated to determine whether African trypanosomes utilize Ca 2 + fluxes to coordinate complex morphological and biochemical life cycle changes. We have identified the ubiquitous intracellular Ca 2+ receptor, calmodulin, in two developmental stages of Trypanosoma brucei rhodesiense. The transition from rapidly dividing, slender bloodstream trypomastigotes to slow growing procyclics in axenic culture was accompanied by changes in specific calmodulin content (3 t tg /mg cell protein to 1 I~g/mg cell protein, respectively) and a shift in intracellular calmodulin distribution, Trypanosome calmodulin is physically and functionally distinct from that of host tissues, including bovine brain and rat erythrocytes. It is similar to but distinct from Tetrahymena calmodulin. Comparisons among these proteins isolated from the four sources were made using the following criteria: (1) mobility on sodium dodecyl sulfate discontinuous polyacrylamide gels; (2) Ca 2 +-induced conformational changes; (3) CNBr-cleavage fragments; (4) activation of bovine brain cyclic nucleotide phosphodiesterase in both a Ca2+-dependent and calmodulin-dependent manner; (5) activation of human erythrocyte (Ca2++ Mg2+)-ATPase; and (6) inhibition of calmodulin activity by trifluoperazine and penfluridol. Trifluoperazine but not trifluoperazine suifoxide was cytotoxic to trypano- somes in vitro. Half maximal effect occurred at 15 ttM. We conclude that calmodulin is a functional component of Africal trypanosomes and suggest that it plays an important role in mediating the host-parasite relationship.
Introduction
Trypanosomes of the brucei group are flagel- lated protozoa which produce lethal infections in humans and domestic mammals throughout much of subsaharan Africa. T. brucei are cyclically trans- mitted by tsetse flies. During their developmental cycle, they undergo major changes in cell structure and biochemical composition, including: (1) a
Abbreviations: PMSF, phenylmethylsulfonyl fluoride; TLCK, N-a-p-tosyl-L-lysine chloromethyl ketone; EGTA, ethylenegly- col bis(fl-aminoethyl ether)-N, N' tetraacetic acid.
series of morphological and biochemical altera- tions in organelle composition to accomodate utili- zation of different metabolic pathways, and (2) antigenic variation of surface coat glycoproteins [1]. Continuous association of parasite with host apparently generates signals responsible for the observed changes in parasite populations. In spite of the critical nature of these changes for parasite survival, little is known about the physiological signals and biochemical pathways responsible for their mediation [2].
In general, activation of lower eukaryotes by extracellular signals is a poorly understood process
0304-4165/83/$03.00 © 1983 Elsevier Science Publishers B.V.

while the analogous situation in higher eukaryotic cells, including responsiveness to hormones and neurotransmitters is fairly well defined. In higher eukaryotes Ca 2÷ in conjunction with calmodulin, along with cyclic nucleotides and protein kinases generally functions to activate otherwise quiescent cells [3]. The Ca2+-calmodulin complex is of par- ticular interest with regards to the study of trypanosomes, since in higher eukaryotes this com- plex activates a number of cell processes whose analogous functions in trypanosomes are poten- tially critical for parasite survival. Such functions include cell secretion [4-6], microtubule related motility [7-9], cyclic nucleotide metabolism [10], activity of protein kinase [10], myosin light chain kinase [10], NAD-kinase [10], (Ca2++Mg2+) - ATPase [10], phospholipase A 2 [10] and methyl transferase [11]. At present Ca 2÷ has been im- plicated in trypanosomes as a factor responsible for release of variant coat glycoproteins [12,13] and activation of membrane-bound adenylate cyclase [14]. However, the physiological signifi- cance of these observations has not been ade- quately assessed. Nonetheless, in both cases, Ca 2÷ activation occurred without any apparent calmodulin requirement. By contrast, cAMP phos- phodiesterase, which is known to have a calmodu- lin-regulated component in mammalian cells was shown to be Ca2+-independent in Trypanosoma cruzi [15]. These observations are surprising since calmodulin or calmodulin-like proteins have been isolated from a number of lower eukaryotes, in- cluding: Tetrahymena [ 16], Paramecium [ 17], yeast [ 18], Dictyostelium [ 19], Neurospora [20], and Limulus amoebocyte [21 ].
We report here that trypanosomes contain calmodulin, identifiable as a low molecular weight, heat-stable protein which undergoes a well-defined conformational change upon binding Ca 2+. Trypanosome calmodulin is inhibited by pheno- thiazines and activates both bovine brain phos- phodiesterase and human erythrocyte (Ca2++ Mg2+)-ATPase. Despite the similarities between trypanosome and host calmodulins, these proteins are structurally and functionally distinct. By con- trast, calmodulins isolated from trypanosomes and Tetrahymena appear to be more closely related. Portions of this work have previously been pre- sented in abstract form [22].
105
Materials and Methods
Materials Penfluridol was obtained as a gift from M.F.
Ralston of Mc Neil Laboratories. Trifluoperazine and trifluoperazine sulfoxide were kindly provided by E. Gallagher of Smith, Kline and French Laboratories. Phenyl-Sepharose is a product of Pharmacia.
Methods Trypanosomes. A triple c loned Yale
Trypanozoan antigenic type 1.1 (YTat 1.1) variant of Trypanosoma brucei rhodesiense, derived from U G A N D A / 6 0 / T R E U 164 (ETat 3), was used throughout this study [23]. This clone was initially classified as T.b. brucei [24] and referred to as such in several of our recent publications [23,25]. Re- classification occurred when a clone from the parent stock of ETat 3 was implicated in a human infection [26] and was reported resistant to normal human serum [27]. Maintenance of stabilates, cell growth and harvesting were as described previ- ously [23]. Trypanosomes which eluted from a DE-52 column were washed once in 47 mM Na2HPO 4, 2.5 mM NaH2PO4, 100 mM NaC1, 5.4 mM glucose (pH 8.0) before use.
Isolation of calmodulin. Washed trypanosomes were resuspended in 20 mM Tris-HC1, 1 mM EDTA, pH 7.4 containing 0.5 mM PMSF and 1 mM TLCK (buffer A) and lysed by sonic disrup- tion with 6 cycles at 20% power on a Branson Sonifier. The homogenate volume was adjusted to the equivalent of 3. 10 9 cells/ml with buffer A and centrifuged at 100000×g. The pellet was extracted with 8 M urea for 2 h then dialyzed against 3 changes of 20 mM Tris-HC1, pH 7.4. The dialyzed extract was centrifuged at 10000 × g for 15 min, combined with the 100000Xg super- natant and layered onto a DE-52 column (50 ml per 30- 10 I° original cells). The column was washed with 5 column volumes of 0.15 mM NaC1, 20 mM Tris-HC1 (pH 7.4) and eluted with 5 volumes of 0.4 M NaC1, 20 mM Tris-HC1, pH 7.4. The col- umn eluant was concentrated with an Amicon YM-2 filter, and dialyzed against one change of 20 mM Tris-HC1, pH 7.4. The eluant was then made 5 mM with Ca 2+, and layered onto a 10 ml phenyl-Sepharose column, pre-equilibrated with

106
200 /~M Ca z+, 20 mM Tris-HCl (pH 7.4) as de- scribed by Gopalakrishna and Anderson [28]. The column was washed with 50 ml of 0.5 M NaC1, 200 /~M Ca 2+, 20 mM Tris-HC1 (pH 7.4) and eluted with the same buffer containing 5 mM E G T A instead of Ca 2+. The t rypanosome calmodulin thus obtained was dialyzed against deionized H20 and lyophylized.
Bovine brain calmodulin was isolated as de- scribed by others [29], except that following DE-52 chromatography, the crude calmodulin was sub- jected to phenyl-Sepharose chromatography [28].
Rat erythrocyte calmodulin was isolated from citrated rat blood, following sonic disruption of the cells. The 15 000 × g, 20 min supernatant was precipitated with 50% saturating ammonium sulfate followed by isoelectric precipitation at pH 4.2. The pellet obtained was dialyzed against 20 mM Tris-HC1 (pH 7.4) heated at 70°C for 2 min and layered onto a DE-52 column. The column was washed as described for t rypanosome calmodulin and the eluant was layered onto a phenyl-Sepharose column.
Cultures of Tetrahymena thermophila strain B III were grown in 2% proteose peptone, 0.1% yeast extract. Cultures were harvested and extracted for calmodulin as described above for trypanosome calmodulin.
Phosphodiesterase. Bovine bra in phos- phodiesterase was partially purified by ammonium sulfate precipitation, DE-52 chromatography, and Blue dextran affinity chromatography, as de- scribed by others [30]. Phosphodiesterase activity was assayed at 37°C for 40 min using the one-step procedure [31]. Samples were incubated in 36 mM Tris-HC1, 36 mM imidazole, 2.7 mM magnesium acetate (pH 7.5) containing 0.15 U / m l 5'- nucleotidase (Sigma) and either 500 ~tM Ca 2+ or 1 mM EGTA. Phosphate released during this time was monitored by the procedure of Fiske and SubbaRow [32]. The reaction was linear under these conditions for all concentrat ions of calmodulin tested.
Erythrocyte (Ca e+ + Mge+)-ATPase. Human erythrocytes were washed in isotonic phosphate buffer containing 0.9% NaCI and lysed in hypo- tonic phosphate. Erythrocyte membranes were iso- lated as described [33]. Activity of the (Ca2++ Mg2+)-ATPase was measured at 37°C in a medium
containing 0.1 mM ouabain, 40 mM NaC1, 3 mM MgC12, 7.5 mM KC1, 20 mM glycylglycine, pH 7.1. Where appropriate, 0.5 mM EGTA, 150 #M Ca 2+ or 1 /~g/ml calmodulin were added. The reaction was started by the addition of 2 mM ATP. At appropriate times, triplicate aliquots were removed and stopped by the addition of 5% tri- chloroacetic acid final concentration. Inorganic phosphate was measured by the method of Fiske and SubbaRow [32].
Quantitation of cellular calmodulin. Trypano- somes were sonicated in buffer A, PMSF and 1 mM TLCK and were separated by centrifugation into a supernatant and pellet at 15000 x g for 10 min. The pellet, containing particulate material such as nuclei, flagella and membranes, was ex- tracted with 8 M urea, dialyzed and assayed for its ability to activate brain phosphodiesterase. The supernatant was dialyzed and assayed directly for its ability to activate brain phosphodiesterase. Standard curves were prepared with known amounts of trypanosome calmodulin and the amount of calmodulin in each cell fraction was quantitated by comparison with the standard curve.
CNBr cleavage. Calmodulin (200 #g) from vari- ous sources was dialyzed, lyophilyzed and sus- pended in 0.1 ml 77% formic acid containing 20 m g / m l CNBr for 24 h at room temperature. The lyophilyzed sample was washed once and resus- pended in electrophoresis sample buffer with 1.0 mM EGTA as described below.
Polyacrylamide gel electrophoresis. Polyacryla- mide gel electrophoresis was performed in dis- continuous gels containing SDS and the buffer system of Laemmli [34]. Low molecular weight CNBr-cleavage fragments were separated on 15% gels containing 8 M urea and 1.15% bisacrylamide with the buffers of Laemmli [34].
Cell motility. Washed trypanosomes at a final cell density of 2.5 • 1 0 6 cells/ml were suspended in 47 mM Na2HPO4, 2.5 mM NaHzPO 4, 100 mM NaC1, 5.0 mM glucose (pH 8.0) containing 1 mM Ca 2+ and 1 m g /m l bovine serum albumin. Tri- fluoperazine or trifluoperazine sulfoxide, diluted with the same buffer, was added and following a 15 min incubation at 4°C, motile cells were counted with an Improved Neubauer haemocytometer. Motile cells in the presence or absence of trifluo- perazine were compared to give percent motile cells.
Protein. Protein determinations were by the method of Lowry et al. [35] or by the modified Bradford [36] procedure of Sedmak and Grossberg [37].
Results
Physical characteristics of trypanosome calmodulin Isolation of calmodulin from small quantities of
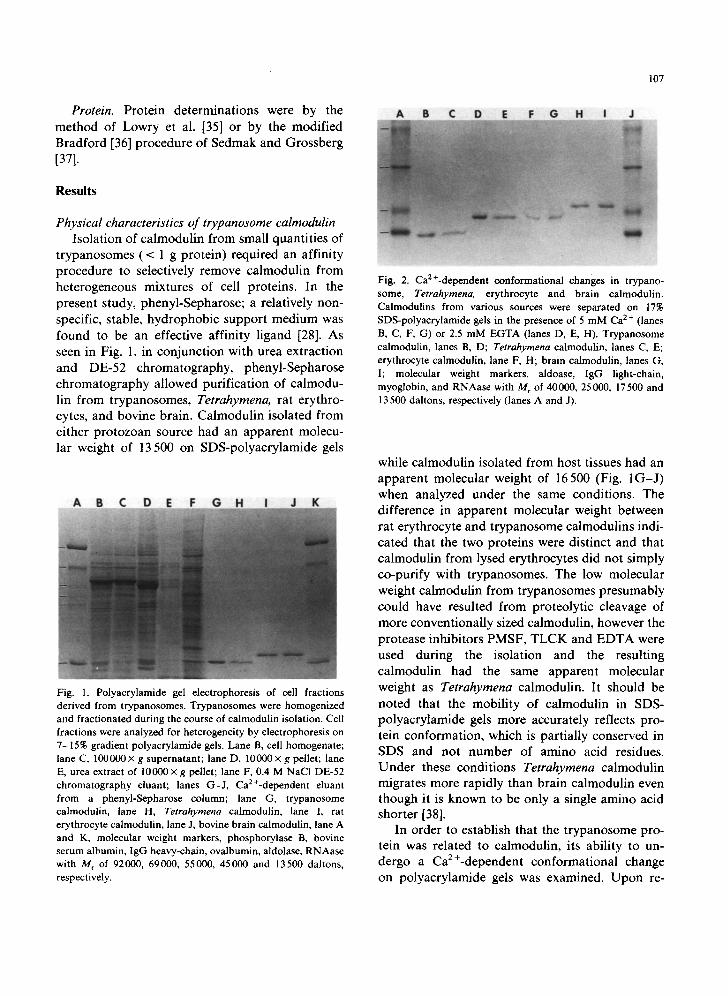
t rypanosomes ( < 1 g protein) required an affinity procedure to selectively remove calmodulin from heterogeneous mixtures of cell proteins. In the present study, phenyl-Sepharose; a relatively non- specific, stable, hydrophobic support medium was found to be an effective affinity ligand [28]. As seen in Fig. 1, in conjunct ion with urea extraction and DE-52 chromatography, phenyl-Sepharose ch romatography allowed purification of calmodu- lin f rom trypanosomes, Tetrahymena, rat erythro- cytes, and bovine brain. Calmodul in isolated from either p ro tozoan source had an apparent molecu- lar weight of 13 500 on SDS-polyacrylamide gels
A
Fig. 1. Polyacrylamide gel electrophoresis of cell fractions derived from trypanosomes. Trypanosomes were homogenized and fractionated during the course of calmodulin isolation. Cell fractions were analyzed for heterogeneity by electrophoresis on 7-15% gradient polyacrylamide gels. Lane B, cell homogenate; lane C, 100000× g supernatant; lane D, 10000× g pellet; lane E, urea extract of 10000x g pellet; lane F, 0.4 M NaCl DE-52 chromatography eluant; lanes G-J, CaE+-dependent eluant from a phenyl-Sepharose column; lane G, trypanosome calmodulin, lane H, Tetrahymena calmodulin, lane l, rat erythrocyte calmodulin, lane J, bovine brain calmodulin, lane A and K, molecular weight markers, phosphorylase B, bovine serum albumin, IgG heavy-chain, ovalbumin, aldolase, RNAase with M r of 92000, 69000, 55000, 45000 and 13500 daltons, respectively.
107
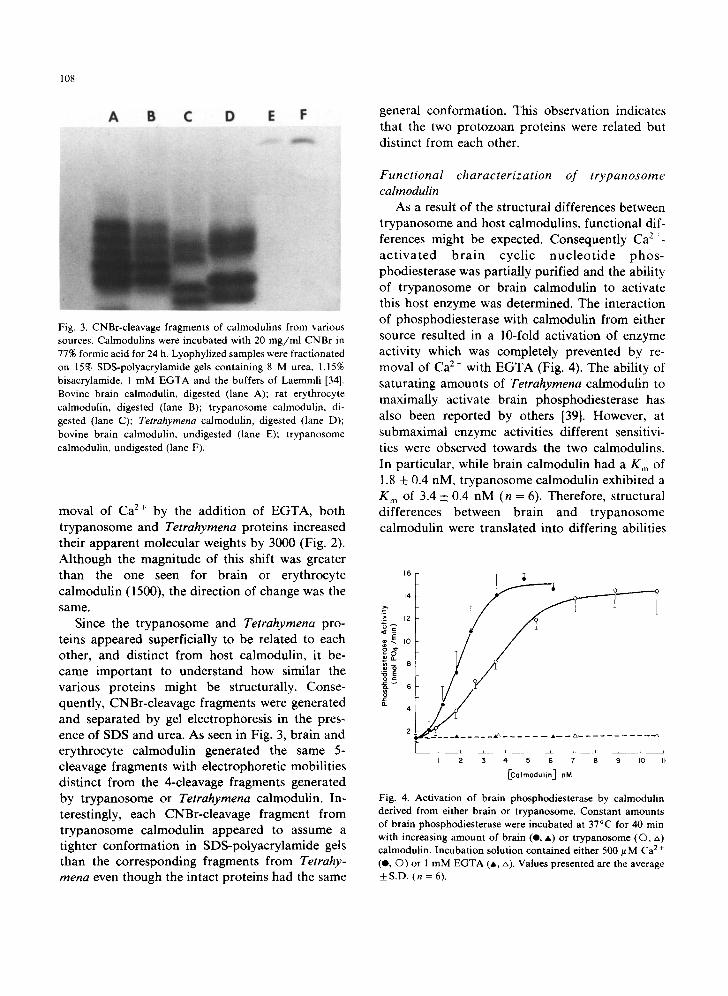
Fig. 2. Ca2+-dependent conformational changes in trypano- some, Tetrahymena, erythrocyte and brain calmodulin. Calmodulins from various sources were separated on 17% SDS-polyacrylamide gels in the presence of 5 mM Ca 2 + (lanes B, C, F, G) or 2.5 mM EGTA (lanes D, E, H). Trypanosome calmodulin, lanes B, D; Tetrahymena calmodulin, lanes C, E; erythrocyte calmodulin, lane F, H; brain calmodulin, lanes G, I; molecular weight markers, aldoase, IgG light-chain, myoglobin, and RNAase with M r of 40000, 25000, 17500 and 13 500 daltons, respectively (lanes A and J).
while calmodulin isolated from host tissues had an apparent molecular weight of 16500 (Fig. 1 G - J ) when analyzed under the same conditions. The difference in apparent molecular weight between rat erythrocyte and t rypanosome calmodulins indi- cated that the two proteins were distinct and that calmodulin from lysed erythrocytes did not simply co-purify with trypanosomes. The low molecular weight calmodulin from t rypanosomes presumably could have resulted from proteolytic cleavage of more conventionally sized calmodulin, however the protease inhibitors PMSF, T L C K and E D T A were used during the isolation and the resulting calmodulin had the same apparent molecular weight as Tetrahymena calmodulin. It should be noted that the mobili ty of calmodulin in SDS- polyacrylamide gels more accurately reflects pro- tein conformation, which is partially conserved in SDS and not number of amino acid residues. Unde r these conditions Tetrahymena calmodulin migrates more rapidly than brain calmodulin even though it is known to be only a single amino acid shorter [38].
In order to establish that the t rypanosome pro- tein was related to calmodulin, its ability to un- dergo a Ca2+-dependent conformat ional change on polyacrylamide gels was examined. Upon re-
108
A B C D E F
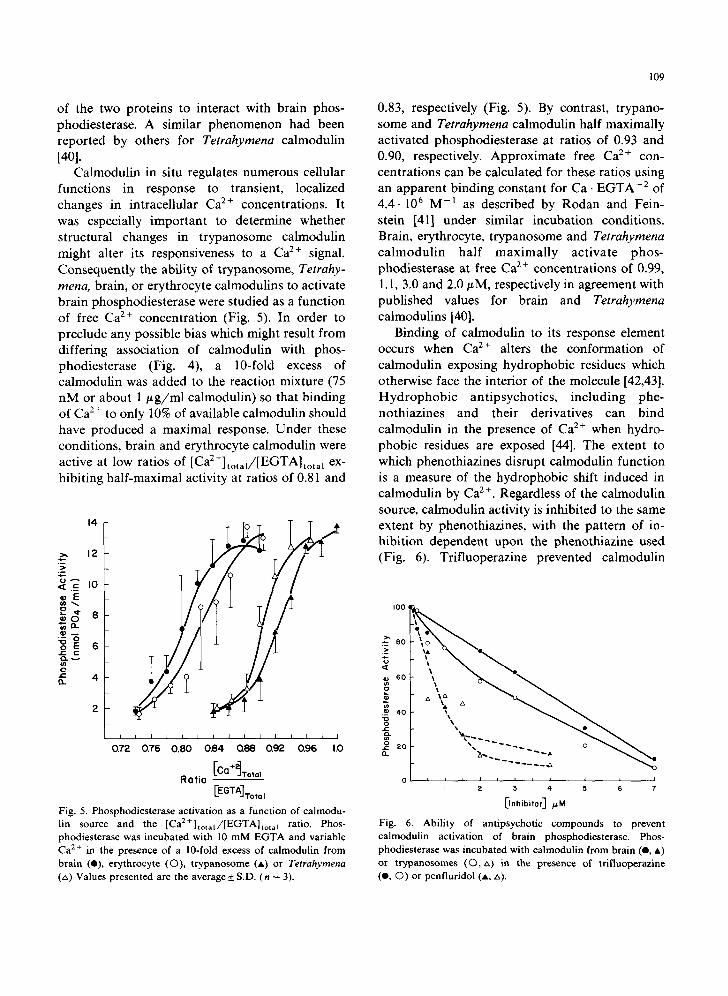
Fig. 3. CNBr-cleavage fragments of calmodulins from various sources. Calmodulins were incubated with 20 mg/ml CNBr in 77% formic acid for 24 h. Lyophylized samples were fractionated on 15% SDS-polyacrylamide gels containing 8 M urea, 1.15% bisacrylamide, 1 mM EGTA and the buffers of Laemmli [34]. Bovine brain calmodulin, digested (lane A); rat erythrocyte calmodulin, digested (lane B); trypanosome calmodulin, di- gested (lane C); Tetrahymena calmodulin, digested (lane D); bovine brain calmodulin, undigested (lane E); trypanosome calmodulin, undigested (lane F).
moval of Ca z+ by the addition of EGTA, both trypanosome and Tetrahymena proteins increased their apparent molecular weights by 3000 (Fig. 2). Although the magnitude of this shift was greater than the one seen for brain or erythrocyte calmodulin (1500), the direction of change was the same.
Since the trypanosome and Tetrahymena pro- teins appeared superficially to be related to each other, and distinct from host calmodulin, it be- came important to understand how similar the various proteins might be structurally. Conse- quently, CNBr-cleavage fragments were generated and separated by gel electrophoresis in the pres- ence of SDS and urea. As seen in Fig. 3, brain and erythrocyte calmodulin generated the same 5- cleavage fragments with electrophoretic mobilities distinct from the 4-cleavage fragments generated by trypanosome or Tetrahymena calmodulin. In- terestingly, each CNBr-cleavage fragment from trypanosome calmodulin appeared to assume a tighter conformation in SDS-polyacrylamide gels than the corresponding fragments from Tetrahy- mena even though the intact proteins had the same
general conformation. This observation indicates that the two protozoan proteins were related but distinct from each other.
Functional characterization of trypanosome calrnodulin
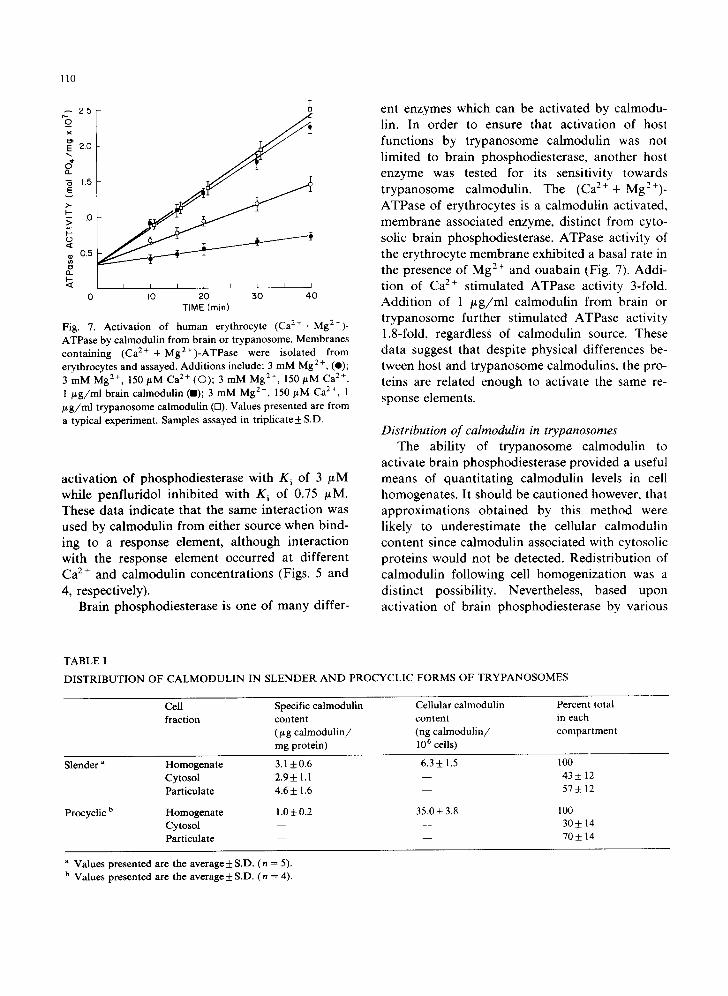
As a result of the structural differences between trypanosome and host calmodulins, functional dif- ferences might be expected. Consequently Ca :+- ac t iva ted bra in cyclic nuc l eo t ide phos- phodiesterase was partially purified and the ability of trypanosome or brain calmodulin to activate this host enzyme was determined. The interaction of phosphodiesterase with calmodulin from either source resulted in a 10-fold activation of enzyme activity which was completely prevented by re- moval of Ca 2 + with EGTA (Fig. 4). The ability of saturating amounts of Tetrahymena calmodulin to maximally activate brain phosphodiesterase has also been reported by others [39]. However, at submaximal enzyme activities different sensitivi- ties were observed towards the two calmodulins. In particular, while brain calmodulin had a K m of 1.8 _+ 0.4 nM, trypanosome calmodulin exhibited a K m of 3.4 + 0.4 nM (n = 6). Therefore, structural differences between brain and trypanosome calmodulin were translated into differing abilities
~" i2
4
t z 3 4 5 6 7 e 9 to Ib
~Colmoclu iin] nM
Fig. 4. Activation of brain phosphodiesterase by calmodulin derived from either brain or trypanosome. Constant amounts of brain phosphodiesterase were incubated at 37°C for 40 min with increasing amount of brain (O, A) or trypanosome (O, zx) calmodulin. Incubation solution contained either 500/t M Ca 2 + (O, O) or 1 mM EGTA (A, r,). Values presented are the average + S.D. (n = 6).
of the two proteins to interact with brain phos- phodiesterase. A similar phenomenon had been reported by others for Tetrahymena calmodulin [40].
Calmodulin in situ regulates numerous cellular functions in response to transient, localized changes in intracellular Ca z+ concentrations. It was especially important to determine whether structural changes in trypanosome calmodulin might alter its responsiveness to a Ca 2÷ signal. Consequently the ability of trypanosome, Tetrahy- mena, brain, or erythrocyte calmodulins to activate brain phosphodiesterase were studied as a function of free Ca 2÷ concentration (Fig. 5). In order to preclude any possible bias which might result from differing association of calmodulin with phos- phodiesterase (Fig. 4), a 10-fold excess of calmodulin was added to the reaction mixture (75 nM or about 1 /~g/ml calmodulin) so that binding of Ca 2÷ to only 10% of available calmodulin should have produced a maximal response. Under these conditions, brain and erythrocyte calmodulin were active at low ratios of [Ca 2+]total//[EGTA]total ex- hibiting half-maximal activity at ratios of 0.81 and
14
>., 12
,e-
E
'~ 8
0
/
I I I I I
0.72 0.76 0.80
/
/
I I l I I I I I I I
0.84 0.88 0.92 0.96 1.0
[Co+~xotol Ratio
[EGTA] Tot* I
Fig. 5. Phosphodiesterase activation as a function of calmodu- lin source and the [Ca2+]total/[EGTA]total ratio. Phos- phodiesterase was incubated with 10 m M EGTA and variable Ca 2+ in the presence of a 10-fold excess of calmodulin from brain (O), erythrocyte (O), t rypanosome (A) or Tetrahymena (zx) Values presented are the average + S.D. (n = 3).
109
0.83, respectively (Fig. 5). By contrast, trypano- some and Tetrahymena calmodulin half maximally activated phosphodiesterase at ratios of 0.93 and 0.90, respectively. Approximate free Ca 2÷ con- centrations can be calculated for these ratios using an apparent binding constant for Ca. EGTA-2 of 4.4-106 M - l as described by Rodan and Fein- stein [41] under similar incubation conditions. Brain, erythrocyte, trypanosome and Tetrahymena ca lmodul in half maximally activate phos- phodiesterase at free Ca 2+ concentrations of 0.99, 1.1, 3.0 and 2.0/~M, respectively in agreement with published values for brain and Tetrahymena calmodulins [40].
Binding of calmodulin to its response element occurs when Ca 2÷ alters the conformation of calmodulin exposing hydrophobic residues which otherwise face the interior of the molecule [42,43]. Hydrophobic antipsychotics, including phe- nothiazines and their derivatives can bind calmodulin in the presence of Ca 2 ÷ when hydro- phobic residues are exposed [44]. The extent to which phenothiazines disrupt calmodulin function is a measure of the hydrophobic shift induced in calmodulin by Ca 2 +. Regardless of the calmodulin source, calmodulin activity is inhibited to the same extent by phenothiazines, with the pattern of in- hibition dependent upon the phenothiazine used (Fig. 6). Trifluoperazine prevented calmodulin
I00
.~8o \o
I I I I I I I I I I I
I 2 3 ,4 5 6 7
13."ib"orl ~M
Fig. 6. Ability of antipsychotic compounds to prevent calmodulin activation of brain phosphodiesterase. Phos- phodiesterase was incubated with calmodulin from brain (O, &) or t rypanosomes (©, zx) in the presence of trifluoperazine (e, ©) or penfluridol (A, a).
110
2 5 %
2.0
ta
1 .5 E
>-
c.)
0.5
I--
o t I I l I J I 410
I0 20 30 TIME (rain)
Fig. 7. Activation of human erythrocyte ( C a 2 + + M g 2 + ) -
ATPase by calmodulin from brain or trypanosome. Membranes containing (Ca 2+ + Mg2+)-ATPase were isolated from erythrocytes and assayed. Additions include: 3 mM Mg 2+, (e); 3 mM Mg 2+, 150/~M Ca 2+ (O); 3 mM Mg 2+, 150 #M Ca 2+, 1 /~g/ml brain calmodulin (m); 3 mM Mg 2+, 150 #M Ca 2+, 1 #g/ml trypanosome calmodulin (n). Values presented are from a typical experiment. Samples assayed in triplicate + S.D.
activation of phosphodiesterase with K i of 3 #M while penfluridol inhibited with Ki of 0.75 #M. These data indicate that the same interaction was used by calmodulin from either source when bind- ing to a response element, although interaction with the response element occurred at different Ca 2+ and calmodulin concentrations (Figs. 5 and 4, respectively).
Brain phosphodiesterase is one of many differ-
ent enzymes which can be activated by calmodu- lin. In order to ensure that activation of host functions by trypanosome calmodulin was not limited to brain phosphodiesterase, another host enzyme was tested for its sensitivity towards trypanosome calmodulin. The (Ca2++ Mg2+) - ATPase of erythrocytes is a calmodulin activated, membrane associated enzyme, distinct from cyto- solic brain phosphodiesterase. ATPase activity of the erythrocyte membrane exhibited a basal rate in the presence of Mg 2+ and ouabain (Fig. 7). Addi- tion of Ca 2÷ stimulated ATPase activity 3-fold. Addition of 1 /~g/ml calmodulin from brain or trypanosome further stimulated ATPase activity 1.8-fold, regardless of calmodulin source. These data suggest that despite physical differences be- tween host and trypanosome calmodulins, the pro- teins are related enough to activate the same re- sponse elements.
Distribution of calmodulin in trypanosomes T h e abi l i ty of t r y p a n o s o m e c a l m o d u l i n to
ac t iva t e b r a i n p h o s p h o d i e s t e r a s e p r o v i d e d a useful
m e a n s of q u a n t i t a t i n g c a l m o d u l i n levels in cell
h o m o g e n a t e s . It s h o u l d b e c a u t i o n e d h o w ev e r , tha t
a p p r o x i m a t i o n s o b t a i n e d by this m e t h o d were
l ikely to u n d e r e s t i m a t e the ce l lu la r c a l m o d u l i n
c o n t e n t s ince c a l m o d u l i n a s s o c i a t e d wi th cy toso l i c
p r o t e i n s w o u l d n o t be d e t e c t e d . R e d i s t r i b u t i o n o f
c a l m o d u l i n f o l l o w i n g cell h o m o g e n i z a t i o n was a
d i s t i n c t poss ib i l i ty . N ev e r t h e l e s s , b a s e d u p o n
a c t i v a t i o n o f b r a i n p h o s p h o d i e s t e r a s e by va r ious
TABLE I
DISTRIBUTION OF CALMODULIN IN SLENDER AND PROCYCLIC FORMS OF TRYPANOSOMES
Cell Specific calmodulin Cellular calmodulin Percent total fraction content content in each
(# g calmodulin/ (ng calmodulin/ compartment mg protein) 106 cells)
Slender a
Procyclic b
Homogenate 3.1 __. 0.6 6.3 ± 1.5 100 Cytosol 2.9+ 1.1 - - 43 _ 12 Particulate 4.6 _+ 1.6 - - 57 _+ 12
Homogenate 1.0 + 0.2 35.0 + 3.8 100 Cytosol - - - - 30 _+ 14 Particulate - - - - 70 + 14
a Values presented are the average + S.D. (n = 5). b Values presented are the average _+ S.D. (n = 4).
cell fractions, calmodulin was found to represent a large proportion of cell protein throughout the trypanosome life cycle (Table I). Approximately 0.3% of total cell protein in slender bloodstream forms was present as calmodulin. This value was especially large considering that approx. 7% of remaining cell protein was comprised of a single species of variable surface glycoprotein [45]. In blood stream trypomastigotes, calmodulin was dis- tributed nearly evenly between particulate and cy- ,tosolic compartments (Table I).
The change in environment from mammalian host to insect vector is associated with dramatic alterations in trypanosome morphology and bio- chemistry. It is assumed that axenically grown procyclic trypomastigotes typify the cell structure and physiology common among cells growing in the tsetse fly mid-gut. Associated with cell mor- phogenesis was a 6-fold increase in cellular calmodulin (Table I). However, since procyclic cells are larger than blood stream forms, the specific content of calmodulin actually decreased to 0.1% of total cell protein. The distribution of calmodulin also shifted with 70% of the protein associated with particulate material and 30% asso- ciated with cytosol. These data indicate that calmodulin is important throughout the trypano- some life cycle. However, membrane-associated calmodulin functions appears to predominate in procyclics.
Calmodulin as a target for trypanocidal agents Phenothiazines and their derivatives are hydro-
phobic antipsychotic compounds which bind to hydrophobic cellular compartments, including: membranes [46], mitochondria [47] and calmodu- lin [44]. Although these drugs are not specific ligands for calmodulin, the sum of their interac- tions with cellular components should be cytotoxic to rapidly growing cells. Consequently, trifluo- perazine was tested to determine whether or not this compound had any general applicability as a therapeutic agent against protozoan parasites. This view was supported by recent work demonstrating the selective cytotoxic effects of phenothiazines on Leishmania grown either free in suspension, or intracellularly in macrophages [48].
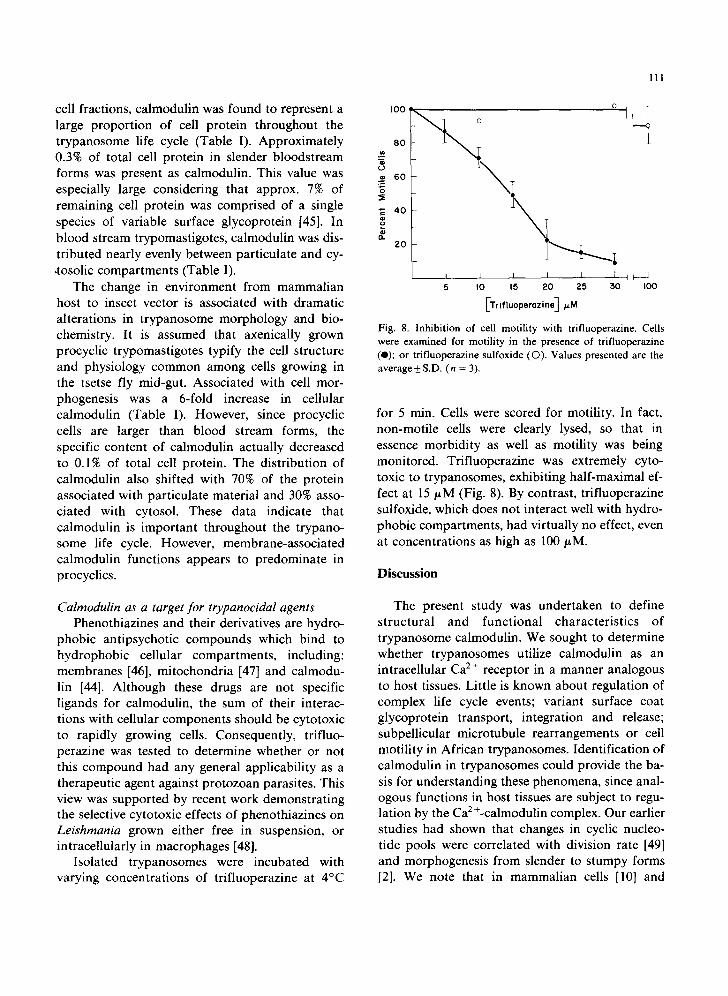
Isolated trypanosomes were incubated with varying concentrations of trifluoperazine at 4°C
I11
~ 6 o
~ zo
5 I0 15 20 25 50 I00
[Trifluoperazine] p.M
Fig. 8. Inhibition of cell motility with trifluoperazme. Cells were examined for motility in the presence of trifluoperazine (e); or trifluoperazine sulfoxide (©). Values presented are the average_+ S.D. (n = 3).
for 5 rain. Cells were scored for motility. In fact, non-motile cells were clearly lysed, so that in essence morbidity as well as motility was being monitored. Trifluoperazine was extremely cyto- toxic to trypanosomes, exhibiting half-maximal ef- fect at 15/~M (Fig. 8). By contrast, trifluoperazine sulfoxide, which does not interact well with hydro- phobic compartments, had virtually no effect, even at concentrations as high as 100 ~M.
Discussion
The present study was undertaken to define structural and functional characteristics of trypanosome calmodulin. We sought to determine whether trypanosomes utilize calmodulin as an intracellular Ca 2÷ receptor in a manner analogous to host tissues. Little is known about regulation of complex life cycle events; variant surface coat glycoprotein transport, integration and release; subpellicular microtubule rearrangements or cell motility in African trypanosomes. Identification of calmodulin in trypanosomes could provide the ba- sis for understanding these phenomena, since anal- ogous functions in host tissues are subject to regu- lation by the Ca 2 +-calmodulin complex. Our earlier studies had shown that changes in cyclic nucleoo tide pools were correlated with division rate [49] and morphogenesis from slender to stumpy forms [2]. We note that in mammalian cells [10] and

112
Tetrahymena [38] cyclic nucleotide pools are, in part, regulated by caimodulin.
We report here that T.b. rhodesiense contain large quantities of calmodulin throughout their life cycle. The persistence of calmodulin contrasts with other trypanosomal proteins which are present only at specific times during the life cycle [1], indicating that reponsiveness to Ca 2÷ signals is important at all times of development. Neverthe- less, the specific calmodulin content decreases in slow growing procyclic cultures, suggesting that selective expression of the calmodulin gene(s) and presumably expression of calmodulin response ele- ments is also important for development. Similar observations are made on transformed mammalian cells where the increased growth rate is associated with a higher specific calmodulin content [50]. In both slender and procyclic cell types, calmodulin is localized in cytosolic and particulate fractions.
Trypanosome calmodulin is distinct from host calmodulin when compared using the criteria of: [1] apparent molecular weight in SDS-poly- acrylamide gels (Fig. 2); [2], Ca2+-induced confor- mational changes (Fig. 2); [3], CNBr-cleavage fragments (Fig. 3); [4], interaction with cyclic nucleotide phosphodiesterase (Fig. 4) and [5] Ca2+-dependency of phosphodiesterase activation (Fig. 5). Trypanosome calmodulin was similar to Tetrahymena calmodulin but differed in apparent size of the CNBr-cleavage fragments separated by SDS-urea polyacrylamide gels. Variability between the different calmodulins probably reflects early evolutionary divergence of trypanosome, Tetrahy- mena and mammalian cells. Calmodulin from other lower eukaryotes (e.g. Tetrahymena, Dictyostelium and Neurospora) are as distinct from each other as from mammalian calmodulin.
The structure of trypanosome calmodulin re- sults in a less efficient interaction with bovine brain cyclic nucleotide phosphodiesterase (Fig. 4) and an increased Ca 2+ requirement for phos- phodiesterase activation (Fig. 5). With Tetrahy- mena these same properties have been ascribed to amino acid substitutions at residues 143 and 146 and in CaZ+-binding domains III and IV [38]. An implication of these data is that lower eukaroytes require higher intracellular C a 2 + concentrations to activate calmodulin-dependent enzyme processes.
Distinct calmodulin may be a common feature
among other parasitic protozoa, such as T. cruzi, Leishmania and Plasmodia. This proposition sug- gests that calmodulin could function in general at a number of levels to modify the host-parasite complex. As an intracellular Ca 2+ receptor, calmodulin is a likely candidate to coordinate many of the complex life cycle changes in trypanosomes and other parasitic protozoa. Parasite calmodulin released into the bloodstream and tissues as a consequence of host responses and parasite killing might activate ectoenzymes [51] and disrupt the, usual functions of host ceils and tissues. Although we have no information concerning these events in vivo, we have shown that trypanosome calmodulin interacts in vitro with host enzymes. Although calmodulin is a highly conserved protein, parasite calmodulin may be sufficiently different structur- ally to function as an invariant antigen in mam- malian hosts. Tetrahymena calmodulin has already been reported by others to be a good antigen [52]. Finally, calmodulin and calmodulin activated pathways may provide suitable targets for chem- otherapy in parasitic infections. The recent dem- onstration of selective cytotoxic effects of pheno- thiazines, on Leishmania donovani grown either free in suspension or intracellularly within macro- phages [48] supports this contention. Trifluopera- zine is highly cytotoxic in vitro to 12.b. rhodesiense exhibiting half maximal effect at 15 #M (Fig. 8). Perhaps in conjunction with other compounds or following modification to prevent avid absorption by host tissues, the antipsychotic compounds might prove useful in the treatment of parasitic protozoa infections.
Addendum
The amino acid sequence for trypanosome calmodulin has been partially determined by Dr. James Strickler, Yale University. Of the amino acid residues that have been sequenced, trypano- some calmodulin differs from Tetrahymena calmodulin at residues 77, 79, 94 and 97 (Gin, Ser, Lys, and Asn, respectively). Trypanosome calmodulin differs from both Tetrahymena and bovine brain calmodulin at residues 99, 108 and 115 (Phe, lie and Lys, respectively). The variability in Ca2+-binding domain III and the absence of trimethyllysine are noteworthy.

113
Acknowledgements
We thank Dr. David Mitchell and Dr. Jeffrey Travis, Yale University for their assistance in pro- viding cultures of Tetrahymena thermophila strain B III. We also thank Dr. Eugene Baskerville for reading the manuscript. This work was supported by grants from N.I.A.I.D. No. A115742 and U.S. P.H.S. Training Grant No. AI07136.
References
1 Englund, P.T., Hajduk, S.L. and Marini, J.C. (1982) Annu. Rev. Biochem. 51,695-726
2 Mancini, P.E. and Patton, C.L. (1981) Mol. Biochem. Parasitol. 3, 19-31
31Rasmussen, H. and D. Waisman (1981) Biochem. Act. Horm. 3, 1-115
4 De Lorenzo, R.J., Freedman, S.D., Yohe, W.B. and Mauer, S.C. (1979) Proc. Natl. Acad. Sci. U.S.A. 76, 1838-1872
5 Schubart, U.K., Fleicher, N. and Erlichman, J. (1980) J. Biol. Chem. 255, 11063-11066
6 Steinhardt, R.L. and Alderton, J.M. (1982) Nature 295, 154-155
7 Burke, B.E. and De Lorenzo, R.J. (1981) Proc. Natl. Acad. Sci. U.S.A. 78, 991-885
8 Blum, J.J., Hayes, A., Jamieson, G.A. Jr. and Vanaman, T.C. (1980) J. Cell Biol. 87, 386-397
9 Welsh, M.J., Dedman, J.R., Brinkley, B.R. and Mean, A.R. (1979) J. Cell Biol. 81,624-634
10 Bromstrom, C.O. and Wolff, D.J. (1981) Biochem. Pharmacol. 30, 1395-1405
11 Alemany, S., Varela, I., Harper, J.F. and Mato, J.M. (1982) J. Biol. Chem. 257, 9249-9251
12 Voorheis, H.P., Bowles, D.J. and Smith, G.A. (1982) J. Biol. Chem. 257, 2300-2304
13 Bowles, D.J. and Voorheis, H.P. (1982) FEBS Lett. 139, 17-21
14 Voorheis, H.P. and Martin, B.R. (1981) Eur. J. Biochem. 116, 471-477
15 Goncalves, M.F., Zingales, B. and Colli, W. (1980) Mol. Biochem. Parasitol. 1, 107-118
16 Suzuki, Y., Hirabyashi, J. and Watanbe, Y. (1979) Biochem. Biophys. Res. Commun. 90, 253-260
17 Rauh, J.J. and Nelson, D.L. (1981) J. Cell Biol. 91,860-865 18 Hubbard, M., Bradley, J., Sullivan, P., Shepherd, J. and
Forrester, I. (1982) FEBS Lett. 137, 85-88 19 Bazari, W.L. and Clarke, M. (1981) J. Biol. Chem. 256,
3598-3603 20 Cox, J.A., Ferraz, L., Demaille, J.G., Perez, R.O., van
Tuinen, D. and Marine, D. (1982) J. Biol. Chem. 257, 10694-10700
21 Liang, S., Liang, C. and Liu, T. (1981) J. Biol. Chem. 256, 4968-4972
22 Ruben, L., Egwaugu, C. and Patton, C.L. (1982) J. Cell Biol. 95, 49a
23 Strickler, J.E. and Patton, C.L. (1982) Mol. Biochem. Parasitol. 5, 117-131
24 Lumsden, W.H.R. and Herbert, W.J. (1975) Trans R. Soc.
Trop. Med. Hyg. 69, 205-208 25 Mancini, P.E., Strickler, J.E. and Patton, C.L. (1982) Bio-
chim. Biophys. Acta 688, 399-410 26 Robertson, D.H.H. and Pickens, S. (1975) Commun. Dis.
Scotland Wk. Rpt. 9: Aug. 27 Van Meirvenne, N., Magnus, E. and Jansens, P.G. (1976)
Ann. Soc. Beige. Med. Trop. 56, 55-63 28 Gopalakrishna, R. and Anderson, W.B. (1982) Biochem.
Biophys. Res. Commun. 104, 830-836 29 Teo, T.S., Wang, T.H. and Wang, J.H. (1973) J. Biol. Chem.
248, 588-595 30 Morrill, M.E., Thompson, S.T. and Stellwagen, E. (1979) J.
Biol. Chem. 254, 4371-4374 31 Teo, T.S. and Wang, J.H. (1973) J. Biol. Chem. 248,
5950-5955 32 Fiske, C.H. and SubbaRow, Y., (1925) J. Biol. Chem. 66,
375-400 33 Steck, T.L. and Kant, J.A. (1974) in Methods in Enzymol-
ogy (Fleischer, S. and Packer, L., eds.), Vol. 31, pp. 172-180 34 Laemmli, U.K. (1970) Nature 227, 680-685 35 Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall,
R.J. (1951) J. Biol. Chem. 193, 265-275 36 Bradford, M. (1976)Anal. Biochem. 72, 248-254 37 Sedmak, J.J. and Grossberg, S.E. (1977) Anal. Biochem. 79,
544-552 38 Yazawa, M., Yagi, K., Toda, H., Kondo, K., Narita, K.,
Yamazaki, R., Sobue, K., Kakiuchi, S., Nagao, S. and Nozawa, Y. (1981) Biochem. Biophys. Res. Commun. 99, 1051-1057
39 Jamieson, G.A. Jr., Vanaman, T.C. and Blum, J.J. (1979) Proc. Natl. Acad. Sci. U.S.A. 76, 6471-6475
40 Kakiuchi, S., Sobue, K., Yamazaki, R., Nagao, S., Umeki, S., Nozawa, Y., Yazawa, M. and Yagi, K. (1981) J, Biol. Chem. 256, 19-22
41 Rodan, G.A. and Feinstein, M.B. (1976) Proc. Natl. Acad. Sci. USA 73, 1829-1833
42 Tanaka, T. and Hidaka, H. (1980) J. Biol. Chem. 255, 11078-11080
43 LaPorta, D.C., and Wierman, B.M. and Storm, D.R. (1980) Biochemistry 19, 3814-3819
44 Levin, R.M. and Weiss, B. (1977) Mol. Pharmacol. 13, 690-697
45 Borst, P. and Cross, G.A.M. (1982) Cell 29, 291-303 46 DiFrancesco, C. and Bickel, M.H. (1977) Chem. Biol. Inter-
act. 16, 335-346 47 Ruben, L. and Rasmussen, H. (1981) Biochim. Biophys.
Acta 637, 415-422 48 Pearson, R.D., Marian, A.A., Harcus, J.W., Hall, D. and
Hewlett, E.L. (1982) Science 217, 369-370 49 Strickler, J.E. and Patton, C.L. (1975) Science 190,
1110-1112 50 Chafouleas, J.G., Pardue, R.L., Brinkley, B.R., Dedman,
J.R. and Means, A.R. (1981) Proc. Natl. Acad. Sci. U.S.A. 78, 996-1000
51 Stanley, K.K., Newby, A.C. and Luzio, J.P. (1982) TIBS 7, 145-146
52 Suzuki, Y., Ohnishi, K., Hirabayaski, T. and Watanabee, Y. (1982) Exp. Cell. Res. 137, 1-14
53 Mansfield, John M. (1981) in Parasitic Diseases; The Im- munology (J.M. Mansfield, ed.), pp. 167-226, Marcel De- kker, Inc. New York





























