Embed Size (px)
Citation preview

0003–
ANIMAL BEHAVIOUR, 2005, 70, 579–587doi:10.1016/j.anbehav.2004.11.015
African elephant vocal communication I: antiphonal
calling behaviour among affiliated females
JOSEPH SOLTIS, KIRSTEN LEONG & ANNE SAVAGE
Education & Science, Animal Programs Administration, Disney’s Animal Kingdom
(Received 24 July 2004; initial acceptance 28 September 2004;
final acceptance 12 November 2004; published online 6 July 2005; MS. number: A9946)
African elephants, Loxodonta africana, are well known for their use of a low-frequency ‘rumble’vocalization, which is thought to function in long-distance communication. Less work, however, hasbeen conducted on short-distance communication within groups, and on spontaneously occurring vocalexchanges among identified individuals in particular. This is due in part to the fact that low-frequencyrumbles are difficult to assign to individual callers. We collected vocal data on a group of six female Africanelephants housed at Disney’s Animal Kingdom to determine whether they exchange rumbles inalternating sequences (also known as antiphonal calling). Subjects wore collars outfitted with microphonesand radiotransmitters that allowed identification of individual callers, and behavioural and endocrine datawere collected so that vocal activity could be examined in the context of social behaviour and reproductivestate. First, we found that females did not produce rumbles at random, but were nearly twice as likely toproduce rumbles shortly after rumbles from other group members. Second, the relative dominance rankand reproductive state of callers did not affect the probability of vocal response, but affiliative relationshipwith the caller had a strong influence on rumble response. Females were most likely to respond in kind tothe rumbles of their most affiliated partners compared to less affiliated group members. Third, videoanalysis showed that rumble exchanges occurred in variable contexts, including when animals were out ofcontact, during reunions, and while in close proximity. Also, affiliated partners often vocalized in sequencewhen approached by dominant individuals. The results of these analyses show that affiliated femaleAfrican elephants exchange rumbles antiphonally, and imply multiple functions for such vocal exchanges.
� 2005 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Temporally associated calling behaviour among adultanimals, including antiphonal calling, duets and cho-ruses, can mediate a wide range of both cooperative andcompetitive interactions among individuals (e.g. insectsand anurans: Greenfield 1994; Bailey 2003; birds: Hall2004). In social species, cooperative vocal exchangesamong adults are common, most notably the separationor contact call, which may signal identity and locationwhen out of visual contact (e.g. birds: Brown et al. 1988;Aubin et al. 2000; mammals: Janik & Slater 1998; Sugiura1998; Jordan et al. 2004). Antiphonal exchanges amongaffiliated partners also occur during ongoing social in-teraction (e.g. nonhuman primates: Smith et al. 1982;humans: Smoski & Bachorowski 2003). Additionally,avian duetting among breeding pairs may sometimes
Correspondence and present address: J. Soltis, Animal ProgramsAdministration, Disney’s Animal Kingdom, PO BOX 10000, Lake BuenaVista, FL 32830, U.S.A. (email: [email protected]). K. Leong isnow at the Department of Natural Resources, Cornell University, Ithaca,NY 14853, U.S.A.
573472/04/$30.00/0 � 2005 The Association for the
serve cooperative functions, including joint territorialdefence (e.g. Hall 2000; Mulder et al. 2003) or ensuringreproductive synchrony (see Hall 2004).Competitive interactions can also result in temporally
associated calling behaviour. For example, territorial malegrasshoppers engage in ‘acoustic duelling’ wherein theyalternate ‘shuck’ calls that signal vigour, and most disputesare settled via this acoustic ritual (Greenfield & Minckley1993). Similarly,male reddeer produce roar vocalizations inthe early assessment stage of rutting season contests, andthese acoustic signals of competitive ability also influencethe termination of contests before escalation (Clutton-Brock & Albon 1979; Reby & McComb 2003). Temporallyassociated vocalizations from different individuals can alsoresult from competitive signal jamming. In many anuransand acoustic insects, for example,males congregate in largeaggregations and produce acoustic choruses, and femalesoften choose individual males based on the acousticqualities of calls. In some cases, males time their calls suchthat they occur before the onsets of the calls of neighbour-ingmales. Because late-onset calls aremaskedbyearly-onset
9Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.

ANIMAL BEHAVIOUR, 70, 3580
calls, females are more likely to choose early-onset callers(e.g. insects: Greenfield et al. 2004; anurans: Grafe 1996;Greenfield & Rand 2000).The social core of African elephant, Loxodonta africana,
society is the female family, which consists of adult femalekin and immature offspring of both sexes, and multiple,related families form larger social units that are oftenseparated in space (Laws et al. 1975; Moss & Poole 1983;Poole et al. 1988). Field experiments show that femalesrespond antiphonally to the playback rumble vocaliza-tions of related females at distances of up to 2.5 km(McComb et al. 2003; also see Poole et al. 1988; Langbaueret al. 1991), evidence that female rumble exchangesfunction as long-distance contact calls. Additionally, ob-servations of spontaneous vocal behaviour within groupsshow that rumbles are often produced in temporal clusterscontaining overlapping vocalizations (Payne et al. 1986;Poole et al. 1988; Leong et al. 2003a).Taken together, these findings suggest that antiphonal
calling is a functionally significant aspect of femaleAfrican elephant vocal communication. However, low-frequency rumble vocalizations are difficult to detect andlocalize to particular callers (e.g. Payne et al. 1986). In thispaper, we present data derived from a radiocollar audio-recording system that allowed collection of 1094 individ-ually identified rumbles from a captive social group of sixfemales, and we use these data to determine whethersocially living female African elephants exchange rumblevocalizations in antiphonal sequences.Given the highly cooperative nature of female African
elephant social interaction (e.g. Dublin 1983; Lee 1987,1989; McComb et al. 2001), it may be expected thatantiphonal calling behaviour, should it exist, would beaffiliative in nature. Even in groups with high kinshipdensity, however, individual interests will sometimes be inconflict (for African elephants, see Dublin 1983), so weexplored both cooperative and competitive interactionsthat may give rise to antiphonal calling behaviour. Wechose three variables of general biological significancethat may influence the probability of vocal response:dominance rank relative to the caller, reproductive stateof the caller, and affiliative relationship with the caller.First, we evaluated relative dominance rank of antiph-
onal callers to explore the possibility that overlappingvocalizations constitute competitive interactions, namely,acoustic interference. Berg (1983) noted that subordinatefemale African elephants sometimes produce rumblesduring aggressive encounters with dominant animals,and our unpublished data confirm these findings. Whilesuch vocalizations could serve many functions, onepossibility is the recruitment of allies who are nearbybut out of visual contact (e.g. Gouzoules & Gouzoules1990). If such recruitment required individual recognitionby voice, then overlapping rumbles by dominant femalescould acoustically mask the subordinate’s call (Greenfield1994).Second, we explored the effect of reproductive state of
the caller on vocal response. In the wild, one commonlyproposed function of the rumble is a long-distance mateattraction call by females that may signal impendingovulation to males, who are largely solitary and distributed
widely in space. Field experiments show that males re-spond to playbacks of female rumbles as part of their matesearch strategy (Langbauer et al. 1991; Poole 1999), anda captive study showed that females increase their rate ofrumble production before ovulation (Leong et al. 2003b).The same captive study also showed the vocal activity ofthe group as a whole increased when there was more thanone female approaching ovulation. The reproductive phaseof callers could influence vocal response if group membersact cooperatively to enhance long-distance mate attractioncalls, by producing rumble choruses that manipulate malemate-searching behaviour. If so, then vocal responseshould increase as callers approach ovulation.
Third, we examined the effect of affiliative relationshipwith the caller on vocal response, regardless of reproduc-tive phase. If antiphonal calling behaviour is largelycooperative or affiliative in nature, then females shouldexchange rumbles preferentially with their most closelybonded social partners. Finally, after determining theeffects of the above three variables on antiphonal calling,we examined concomitant social behaviour to furtherexplore the potential functions of antiphonal vocalactivity.
METHODS
Study Subjects and Housing
Disney’s Animal Kingdom (DAK; Lake Buena Vista,Florida, U.S.A.) maintained three adult male and sixadult female African elephants. Study subjects were sixadult nonpregnant, nulliparous females (age range 19–30 years). All subjects were wild born but reared incaptivity since an early age (3–5 years) and are presumedto be unrelated. The six females arrived in pairs at DAK2–3 years prior to the study. Each of the three pairsenjoyed long histories together prior to arrival at DAK,ranging from 17–25 years. Subjects were housed inindoor stalls overnight and released into one of twonaturalistic outdoor enclosures (main yard: 2.29 ha;small yard: 0.55 ha), or a smaller yard attached to theindoor barn (barn yard: 0.04 ha). Regardless of yard,subjects were in vocal and sometimes visual contactwith one another. Subjects were in outdoor yards fromapproximately 0800 to 1700 hours. Data analysed herewere collected from December 1999 to November 2000.Group compositions varied according to managementobjectives, and the data presented here are derived fromconfigurations in which all six females were housedtogether in the main yard without males, which totalled60 observation days (60 h).
Vocal Data Collection and Analysis
Elephant vocalizations were recorded from transmittersmounted in collars worn by the elephants (Leong et al.2003a, b). Collars were designed and built by Walt DisneyWorld Company Instrumentation Support Division ofRide and Show Engineering, modifying an original design

SOLTIS ET AL.: ANTIPHONAL CALLING IN ELEPHANTS 581
by William Langbauer, Jr and Steven Powell. Each collarcontained a condenser microphone element (Radio Shack270-090C with a flat frequency response down to 30 Hzand a 10-dB loss at 20 Hz), a radiotransmitter (Jobcom100, 150 to 160 MHz Ritron Handheld Radio with a cus-tom antenna), and a battery pack (2EA Gel Cell, RoyalBattery BC-632, 6 V/3.2 amp-hour).Each collar transmitted sound signal data on a unique
radio frequency to an antenna-receiver component in theelephant barn (antenna: Antenna Specialties ASP-655 Tem-porary Base Station half-wave antenna, 136–174 MHz;receiver: Hamtronics R-144/R226 VHF/FM receiver, 150–169 MHz). Each of these receivers was in turn connectedto a separate channel of an eight-channel digital audiorecorder (TASCAM DA-38) where they were recorded at44.1-kHz sample rate to separate tracks on digital audio-tape (Sony DARS-60MP). On recording days, collars wereput on elephants in the morning and removed at night torecharge batteries. A 1-h audiotape was recorded for eachelephant each day the collars were worn, always between1000 and 1400 hours.Data tapes were played on a DAT recorder (TASCAM DA-
38) through an audio mixer (Mackie 1202-VLZ) and anantialiasing filter (Tunable Active Filter Instrument 900C/9L8B 8-pole Buttorworth low-pass filter). Vocalizationswere located on each channel of the tape using Real TimeSpectrogram (RTS; version 2.0, Engineering Design, Berkeley,California, U.S.A.). RTS sample rate was set at 1200 Hz tovisually locate vocalizations that contained low-frequencycomponents (i.e. rumble vocalizations). Collar micro-phones were sensitive enough to pick up vocalizationsof nearby elephants. Identification of callers was deter-mined by relative strength and timing of signals usinga customized eight-channel Real Time Event Detector ata 1200-Hz sample rate (customized RTED; EngineeringDesign) and observation of video (see below). We recordedthe vocal identity, date and time of each rumble vocaliza-tion for later comparison with ongoing social behaviourfrom videotape.
Analysis of Antiphonal Calling
We defined rumble exchanges between females based onthe following rationale. Leong et al. (2003a) found thatmost vocalizations occurredwithin 30 s of one another andthat most overlapping bouts involved only two individu-als. Thus, dyadic rumble exchanges that occurred within30 s of each other were the focus of this study. Specifically,a second-in-sequence rumble was considered a potentialanswer to an initial rumble if its onset occurred after theonset of the initial rumble but before the termination of theinitial rumble (termed an ‘overlapping rumble’) or occurredwithin 30 s of the termination of the initial rumble (termedan ‘adjacent rumble’). In these analyses, the dyadic vocalexchange is the unit of analysis. For example, a sequence of‘adjacent’ rumbles from two females, ABA, is broken downinto two dyadic exchanges, AB and BA.Temporally associated vocalizations are not necessarily
communicative events. Even if females vocalize indepen-dently of one another, some proportion of rumbles will be
overlapping or adjacent by chance alone. Modifyingmethods developed by Wasserman (1977), we determinedwhether females were more, less, or equally likely toproduce a rumble after female conspecific rumbles, com-pared to periods of time in which no conspecific hadproduced a rumble. We defined an ‘answer’ rumble asbeginning anywhere after the onset of a conspecificrumble to 30 s after the end of the rumble. Forany given female, let a be the upper-bound amount oftime that conspecific rumbles were answerable, which isgiven by:
aZNoðdCxÞ
where No is the total number of rumbles by all otherfemales, d is the average duration of rumble vocalizations(4.6 s, NZ 270 rumbles), and x is the amount of time aftera rumble during which another rumble by a differentfemale is considered an answer (30 s). Let t be the totalamount of time the elephants were recorded(60 hZ 216000 s). Thus, the proportion of time thatrumbles were ‘answerable’ for any given female is a/t. IfNs is the number of rumbles produced by the female inquestion, then the expected number of her rumbles thatwould be ‘answers’ by chance alone is given by:
Nsða=tÞ:
By comparing the expected versus the observed number ofanswer rumbles for each female, we determined whetherthe probability of rumbling is increased, decreased orunaffected by conspecific rumbles.Note that this analysis is conservative in that it is biased
against rejecting the null hypothesis. The analysis as-sumes that none of the ‘answerable’ time intervals over-lap. Since in actuality many of these intervals do overlap,the result is an overestimate of the proportion of time thatrumbles were considered answerable. This, in turn, givesan overestimate of the number of rumbles that will beanswers by chance alone. Thus, this analysis is biasedagainst finding a significant increase in rumbling afterconspecific rumbles.
Behavioural Data Collection and Analysis
Three to four observers documented behaviour for each1-h observation session using Panasonic mini-digitalvideo cameras, as described previously (Leong et al.2003a, b). Briefly, field observers filmed elephants fromviewing locations surrounding the enclosure. Observersfilmed the elephants for 10-min focal periods, with anaverage of two 10-min periods recorded per observationper elephant. The video signals from all four field videocameras were routed through an analogue quad-splitterand copied to a VHS tape with four windows, eachcontaining the signal from one of the mini-digital videocameras, synchronized in time, with the accompanyingaudio information from radio transmissions.Scoring of behaviour during antiphonal exchanges was
conducted on the 60 h of quad-split videos. An approachwas scored when one animal moved within 8 m of

ANIMAL BEHAVIOUR, 70, 3582
another, after which the pair members were consideredin proximity. Distances between subjects that were inview on separate cameras were measured using an aerialmap of the main yard. ‘Reunions’ occurred if theantiphonal callers were greater than 20 m apart, andeither or both moved more than 8 m to within 20 m ofthe other.Dominance ranks were determined by displacements
(NZ 126). A displacement occurred when one animalmovedwithin 4 m of another, and the other animalmovedmore than 4 m away, within 5 s of the initial approach. For10 of the 15 possible dyads, there were no displacementreversals and relative dominance rank was unambiguous.In four of the remaining dyads, displacements occurred inboth directions but the member of the dyad with the mostdisplacements scored anywhere from 2 to 20 times asmany displacements as the other member. In these cases,the member of the dyad with the most displacements wasconsidered dominant. In the final dyad, the two animalshad equal numbers of displacements, and they wereconsidered codominant (the pair had been social compan-ions for 17 years at the time of the study).For statistical tests, we expressed dominance rank as the
proportion of the other five group members outranked,and each member of a codominant pair outranked oneanother by one-half. For example, a female who out-ranked two individuals in the group, and was codominantwith one individual, would have a dominance rank of2.5/5Z 0.5.
Endocrine Data Collection and Analysis
Blood samples were assayed for progestogen (P) andluteinizing hormone (LH) to determine phases of the cyclefor three cycling females, as described previously (Leonget al. 2003b). Briefly, blood samples were collected twiceper week during the luteal phase and daily during thefollicular period. Serum P and LH concentrations werequantified using enzyme-immunoassays (EIA) validatedfor the African elephant (Graham et al. 2001, 2002; Leonget al. 2003b). Intra- and interassay coefficients of variationwere 7.5% and 11.2%, respectively, for the progestogenEIA, and 8.7% and 13.6% for the LH EIA.The patterns of serum P and LH concentrations were
used to determine the phases of the oestrous cycle.Ovulation was assumed to take place within 24 h of theovulatory LH surge, and the luteal phase was defined asbeginning 48 h after the ovulatory LH surge and endingthe day serum P concentrations reached baseline values.The follicular period was divided into two distinct phasesaccording to the recent findings of Hermes et al. (2000)showing two waves of follicular growth. The anovulatoryfollicular phase was defined as beginning 20 days prior tothe anovulatory LH surge and ending on the day of theanovulatory LH surge. The ovulatory follicular phase wasdefined as beginning 24 h after the anovulatory LH surgeand ending 24 h after the ovulatory LH surge. Thus, theoestrous cycle was divided into luteal, anovulatory follic-ular and ovulatory follicular phases. There were threecycling and three noncycling females.
Statistical Analyses
Nonparametric tests were performed in SPSS (version11.0, Chicago, Illinois, U.S.A.) with two-tailed alpha set at0.05 for all tests.
RESULTS
Rumble Exchanges versus a Random Model
A total of 1094 rumbles attributable to identifiedindividuals were recorded from the six females. Of these,159 (14.5%) were followed by an overlapping rumble, and115 (10.5%) were followed by an adjacent rumble froma different female (within 30 s), for a total of 274 (25.1%)of rumbles potentially answered. Most rumbles (680/1094 Z 62.2%) were not followed by a rumble fromanother female within 30 s, and an additional 140(12.8%) were followed by a rumble from the same femaleor an unidentified individual.
A close temporal association between rumbles does notdemonstrate that the rumbles are causally related to eachother. If females were vocalizing independently of oneanother, some proportion of their rumbles would beoverlapping or adjacent by chance alone. Therefore, wetested each female to determine whether her rumbleproduction increased, decreased, or occurred at randomwith respect to the vocal activity of other females. Five ofsix females produced rumbles more often after anotherfemale’s rumble compared to periods of time in which nofemale had recently produced a rumble (Table 1). Four ofthese five were significantly more likely to rumble afterother females did so. Averaged across all six females,rumble production was 1.88 times higher after the pro-duction of a rumble by another female compared toperiods of time in which no other female produceda rumble.
Factors Influencing Antiphonal Response
To explore the possible communicative functions ofantiphonal calling behaviour, we examined three variablesthat may influence vocal interactions among females. First,we examined the relationship between relative dominancerank and the production of overlapping rumbles, whichmay constitute a form of vocal interference. Relativedominance rank did not have major influence on theproduction of overlapping rumbles, however. We deter-mined the dominance rank of the second-in-sequencecaller relative to the initial caller for all 159 overlappingrumble sequences. In 34 of 159 cases (21.4%), the vocaliz-ing pair was codominant. Overlapping callers were equallylikely to be dominant (61/159 Z 38.4% of cases) as theywere to be subordinate (64/159 Z 40.3% of cases) relativeto the initial callers (binomial test: NZ 125 cases,P Z 0.858; Fig. 1). Additionally, high-ranking females didnot produce proportionally more overlapping rumblesthan low-ranking females. Across females, there was nocorrelation between female dominance rank and thepercentage of her answer rumbles that were overlapping
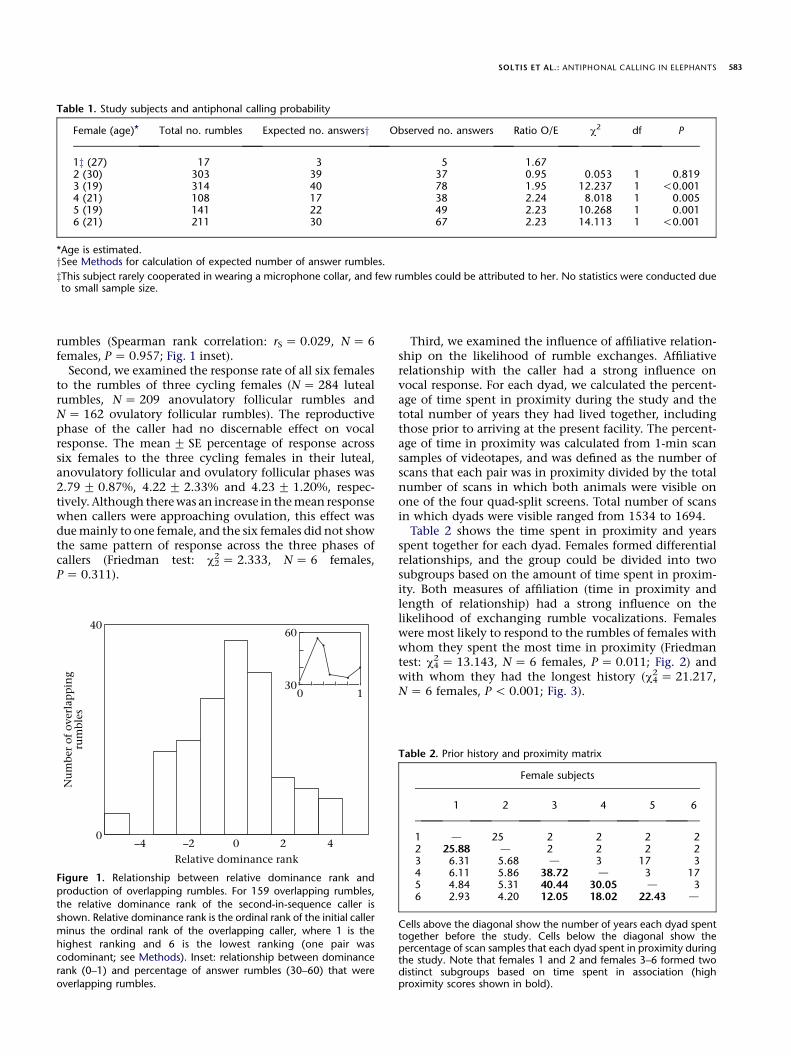
SOLTIS ET AL.: ANTIPHONAL CALLING IN ELEPHANTS 583
Table 1. Study subjects and antiphonal calling probability
Female (age)* Total no. rumbles Expected no. answersy Observed no. answers Ratio O/E c2 df P
1z (27) 17 3 5 1.672 (30) 303 39 37 0.95 0.053 1 0.8193 (19) 314 40 78 1.95 12.237 1 !0.0014 (21) 108 17 38 2.24 8.018 1 0.0055 (19) 141 22 49 2.23 10.268 1 0.0016 (21) 211 30 67 2.23 14.113 1 !0.001
*Age is estimated.ySee Methods for calculation of expected number of answer rumbles.
zThis subject rarely cooperated in wearing a microphone collar, and few rumbles could be attributed to her. No statistics were conducted dueto small sample size.
rumbles (Spearman rank correlation: rS Z 0.029, NZ 6females, P Z 0.957; Fig. 1 inset).Second, we examined the response rate of all six females
to the rumbles of three cycling females (NZ 284 lutealrumbles, NZ 209 anovulatory follicular rumbles andNZ 162 ovulatory follicular rumbles). The reproductivephase of the caller had no discernable effect on vocalresponse. The mean G SE percentage of response acrosssix females to the three cycling females in their luteal,anovulatory follicular and ovulatory follicular phases was2.79 G 0.87%, 4.22G 2.33% and 4.23 G 1.20%, respec-tively. Although therewas an increase in themean responsewhen callers were approaching ovulation, this effect wasduemainly to one female, and the six females did not showthe same pattern of response across the three phases ofcallers (Friedman test: c2
2 Z 2.333, NZ 6 females,PZ 0.311).
40
0–4 –2 0 2 4
Relative dominance rank
60
300 1
Nu
mbe
r of
ove
rlap
pin
gru
mbl
es
Figure 1. Relationship between relative dominance rank and
production of overlapping rumbles. For 159 overlapping rumbles,the relative dominance rank of the second-in-sequence caller is
shown. Relative dominance rank is the ordinal rank of the initial caller
minus the ordinal rank of the overlapping caller, where 1 is the
highest ranking and 6 is the lowest ranking (one pair wascodominant; see Methods). Inset: relationship between dominance
rank (0–1) and percentage of answer rumbles (30–60) that were
overlapping rumbles.
Third, we examined the influence of affiliative relation-ship on the likelihood of rumble exchanges. Affiliativerelationship with the caller had a strong influence onvocal response. For each dyad, we calculated the percent-age of time spent in proximity during the study and thetotal number of years they had lived together, includingthose prior to arriving at the present facility. The percent-age of time in proximity was calculated from 1-min scansamples of videotapes, and was defined as the number ofscans that each pair was in proximity divided by the totalnumber of scans in which both animals were visible onone of the four quad-split screens. Total number of scansin which dyads were visible ranged from 1534 to 1694.Table 2 shows the time spent in proximity and years
spent together for each dyad. Females formed differentialrelationships, and the group could be divided into twosubgroups based on the amount of time spent in proxim-ity. Both measures of affiliation (time in proximity andlength of relationship) had a strong influence on thelikelihood of exchanging rumble vocalizations. Femaleswere most likely to respond to the rumbles of females withwhom they spent the most time in proximity (Friedmantest: c4
2 Z 13.143, NZ 6 females, P Z 0.011; Fig. 2) andwith whom they had the longest history (c4
2 Z 21.217,NZ 6 females, P! 0.001; Fig. 3).
Table 2. Prior history and proximity matrix
Female subjects
1 2 3 4 5 6
1 d 25 2 2 2 22 25.88 d 2 2 2 23 6.31 5.68 d 3 17 34 6.11 5.86 38.72 d 3 175 4.84 5.31 40.44 30.05 d 36 2.93 4.20 12.05 18.02 22.43 d
Cells above the diagonal show the number of years each dyad spenttogether before the study. Cells below the diagonal show thepercentage of scan samples that each dyad spent in proximity duringthe study. Note that females 1 and 2 and females 3–6 formed twodistinct subgroups based on time spent in association (highproximity scores shown in bold).

ANIMAL BEHAVIOUR, 70, 3584
% Time in proximity
% V
ocal
res
pon
se
25
0
0 10 20 30 40
Figure 2. Relationship between social affiliation and probability of
antiphonal response to the rumbles of other group members. Each
line represents one female, and the identical symbols on each linerepresent the female’s vocal response to, and time spent in proximity
with, the other females in her group. Each symbol marks the
coordinate indicating the percentage of time spent in proximity witha particular female, and the percentage of her rumbles that were
answered. Generally, females tended to respond in kind to the
rumbles of group members with whom they spent the most time in
proximity. Two females were somewhat of an exception ( ,). These females responded to each other the most, although
they only spent an intermediate amount of time in proximity with
each other (see peaks at 20% time in proximity).
Log10 years together25
% V
ocal
res
pon
se
3
25
0
2 17
Figure 3. Relationship between the number of years spent together
and the probability of antiphonal response to the rumbles of othergroup members. Each line represents one female, and the identical
symbols on each line represent the female’s vocal response to, and
years spent together with, the other females in her group. Each
symbol marks the coordinate indicating the number of years spenttogether with a particular female, and the percentage of her rumbles
that were answered. Years were log10 transformed for better
separation between 2 and 3 years. Females responded most to thefemales that they had known the longest.
The two measures of affiliation were highly correlatedwith one another, with one notable exception. One pair offemales that enjoyed a long history together did not spendthe most time with each other during the study, but theywere nevertheless most likely to exchange vocalizationswith one another compared to other females (see Fig. 2).Thus, the length of relationship better predicted theidentity of vocal responders than current affiliation.
The strength of affiliation (using proximity data) ex-plained a large part of the variance in the vocal responsedata set (Fig. 4). For all 274 antiphonal responses, 37.3%were answered by the most affiliated partner of the femalecaller and an additional 31.9% were answered by thesecond-most affiliated partner, for a cumulative total of69.2%. The top three affiliated partners accounted for90.1% of rumble responses, while the two least affiliatedpartners together accounted for only 9.9% of antiphonalrumble responses.
Description of Social Contexts DuringRumble Exchanges
Affiliative relationship had a strong influence on theidentity of rumble exchangers. We analysed videotapes toinvestigate the social contexts in which the rumbleexchanges took place. Animals were visible on screen for186 of 274 rumble exchanges (67.9%). Temporally associ-ated vocalizations by affiliated animals could be caused byexternal events that both members of the pair experiencesimultaneously, including interaction with third-partyindividuals. To test for this, we recorded whether or notother group members approached either of the pair within30 s of the onset of each vocal exchange. In 31.7% ofcases (59/186), rumble exchanges were associated with
100
50
0
Cu
mu
lati
ve p
erce
nta
geof
ru
mbl
es a
nsw
ered
Affiliation rank with initial caller
1st 2nd 3rd 4th 5th
Figure 4. Affiliative relationship with initial callers and vocal
response. Strongly affiliated partners accounted for the majority of
answer rumbles. Of 274 antiphonal answer rumbles, the percentage
answered by the first, second, third fourth and fifth most affiliatedpartner of the initial caller was 37.3, 31.9, 20.9, 2.8 and 7.1%,
respectively. Affiliation rank was based on percentage of time spent
in proximity.

SOLTIS ET AL.: ANTIPHONAL CALLING IN ELEPHANTS 585
third-party approaches, usually an animal dominant to atleast one member of the vocalizing pair (51/59Z 86.4% ofcases), and the pair was nearly always within relativelyclose proximity (within 20m; see Fig. 5). In the remaining68.3% of cases (127/186), there was no approach by third-party individuals. In these cases, pairs usually, but notalways, were in relatively close proximity (Fig. 5), and in39 of these cases (39/127Z 30.7%), the vocalizing pairengaged in a reunion within 30 s of the vocal exchangeonset.In about one-third of cases, third-party individuals
approached the pair before either member of the pairvocalized. As such, these vocalizations may have beencaused by the third-party approach and may not consti-tute communicative events between members of thevocalizing pair. Therefore, we reanalysed our data exclud-ing these cases, and found that both measures of affiliativerelationship still strongly predicted vocal response (Fried-man test: proximity: c4
2 Z 14.226, NZ 6 females,PZ 0.007. history: c4
2 Z 21.217, NZ 6 females,P! 0.001).
DISCUSSION
Antiphonal Calling among Affiliated Females
The analyses presented here constitute strong evidencethat female African elephants exchange rumble vocaliza-tions in antiphonal sequences. Females were nearly twiceas likely to produce rumbles shortly after another female inthe group did so, compared to periods of time in which noother females were producing rumbles. This propensity torumble in response to the rumbles of other females resultsin bouts of rumble activity, including overlapping rumbles,
Nu
mbe
r of
rum
ble
exch
ange
s
30
20
10
80160
240
Approach
No approachDistance (m)
Figure 5. Histogram showing distances (in 4-m intervals) betweenantiphonal callers at the onset of the second vocalization. Data were
divided into those cases in which the antiphonal sequence was
preceded by an approach by a third-party individual, and those cases
in which there was no preceding approach. For visual clarity, twoextreme outliers were excluded from the histogram (both at 428 m
in the ‘no approach’ category).
which are separated by periods of relative vocal inactivity.Such temporal clustering of rumbles has been observed inboth wild and captive contexts (Payne et al. 1986; Pooleet al. 1988; Leong et al. 2003a), but in this analysis,identification of individual callers was possible because ofthe radiocollar system employed. In doing so, we were ableto demonstrate that all six females were most likely torespond to their most closely affiliated social partnerswithin the group, and were least likely to respond to theirleast affiliated partners, a pattern found in other socialspecies (e.g. Mitani & Nishida 1993; Soltis et al. 2002;Jordan et al. 2004).
Social Contexts of Antiphonal Callingby Affiliated Partners
The temporally related calling among affiliated femalescould be caused in two ways. The second-in-sequence callcould be an answer to the initial call (i.e. bona fideantiphonal calling constituting a communicative event).Alternatively, both calls could be in response to an eventexternal to the pair that impinges on the two callerssimultaneously. This may be especially applicable toaffiliated animals that spend time in close proximity. Wetested one obvious social event that may cause suchsimultaneous vocal activity, namely, approaches by othermembers of the social group. We found that in 32% ofcases, temporally associated rumbles by affiliated partnerswere preceded by approaches from another group mem-ber, usually a dominant animal, and almost always whilethe vocalizing pair was in relative close proximity (Fig. 5).Similarly, Berg (1983) noted that recipients of aggressionsometimes produce rumbles (‘growls’ in Berg’s terminology).In the case of the third-party approaches observed here,both antiphonal callers could be responding to theapproach of the dominant animal and not to each other.Several nonexclusive explanations could be proposed
for these cases. For example, such vocalizations could besimple expressions of negative emotional arousal coinci-dent with the approach of a dominant animal. On theother hand, they could be evolved signals of submissionthat affect the behaviour of dominant animals (e.g. reduceaggression), and they could be signals of distress thataffect the behaviour of potential allies (e.g. enlist support).Little work on vocal activity during dominance interac-tions has been conducted on elephants (but see Berg 1983;Langbauer 2000; and a companion paper, Soltis et al.2005), and this should prove to be a fruitful line ofinquiry. Vocal behaviour is known to mediate aggressiveinteractions in other species, signalling, for example,fighting ability or appeasement (e.g. Cheney et al. 1995;Harcourt & Stewart 1996; Reby & McComb 2003).In 68% of cases, other group members did not approach
affiliated partners during antiphonal calling, however,and these vocal exchanges are the most likely to beantiphonal communicative events. In the wild, there isevidence that antiphonal calling among females is a long-distance separation call used when animals are out ofvisual contact (Poole et al. 1988; Langbauer et al. 1991;McComb et al. 2000). In this captive study, antiphonal

ANIMAL BEHAVIOUR, 70, 3586
calling took place in a variety of close spatial contexts. Aspreviously reported by Leong et al. (2003a), such ex-changes occur mostly when animals are in relatively closeproximity, but also occur when animals are more distant(over 200 m; see Fig. 5), as well as during reunions. Vocalactivity during reunions has been observed in wildelephants (e.g. Poole et al. 1988). In other social species,affiliative vocalizations also occur in a wide variety ofcontexts, including during social separation (e.g. Janik &Slater 1998), reunions (e.g. Kalin et al. 1992), and duringongoing social activity (e.g. Smith et al. 1982). Thesevariable contexts suggest multiple within-group functionsfor the rumble, such as a short-distance contact call,a signal of submission, and reinforcement of social bonds.These results should act as guides for future work testingspecific functional hypotheses.
Alternative Explanations
We tested for the existence of cooperative vocal en-hancement of female rumbles that may function to attractmales to their group, but found that the reproductivephase of the caller did not affect vocal responses in otherfemales. In fact, African elephant female families may becharacterized by competition over reproduction ratherthan cooperation. Dublin (1983) showed for a wild pop-ulation that the pregnancies and births of lower-rankingfemales are delayed compared to those of higher-rankingfemales, and the offspring of lower-ranking femalesexperience higher mortality. Although the mechanismsthat give rise to delayed births are unknown, it suggeststhat dominant females are better able to optimally timebirths compared to lower-ranking females. Despite theoverwhelmingly cooperative nature of female Africanelephant society (e.g. Dublin 1983; Lee 1987, 1989;McComb et al. 2001), female reproduction may be oneimportant arena of competition, as it is in some othermammals with communal care of offspring (e.g. Abbottet al. 1993; Griffin et al. 2003).We also tested for the possible existence of competitive
vocalizations by examining the relative dominance rankof the overlapping callers, but found that relative rank didnot have a major influence on such vocal behaviour. Thisalone does not rule out the existence of competitivevocalizations, however. Indeed, about 40% of the over-lapping calls were produced by females that were domi-nant over the initial caller. None the less, the strength ofthe affiliative bond explained a great deal of the variancein this data set. Of all rumbles that were answered, nearly70% were answered by the two most affiliated partners ofthe initial caller, while the two least affiliated partnersaccounted for only 10% of the answers (see Fig. 4),suggesting that the majority of rumble exchanges werenot competitive in nature.
Acknowledgments
We thank Disney’s Animal Kingdom Animal Programs(Science Team) and Operations (Conservation StationOperations Team) for support of the project and assistance
in data collection. This project would not have beenpossible without the support and assistance of the keepersand managers of the Elephant Team. D. Rudolph andB. Walters of Walt Disney World Ride and Show Engineer-ing designed and maintained the elephant audiocollars,and K. Burks provided highly valued managerial andanalytical expertise to the project.
References
Abbott, D. H., Barret, J. & George, L. M. 1993. Comparativeaspects of the social suppression of reproduction in female
marmosets and tamarins. In: Marmosets and Tamarins: Systematics,
Behavior, and Ecology (Ed. by A. B. Rylands), pp. 152–163. Oxford:
Oxford University Press.
Aubin, T., Jouventin, P. & Hildebrand, C. 2000. Penguins use the
two-voiced system to recognize each other. Proceedings of theRoyal Society of London, Series B, 267, 1081–1087.
Bailey, W. J. 2003. Insect duets: underlying mechanisms and theirevolution. Physiological Entomology, 28, 157–174.
Berg, J. K. 1983. Vocalizations and associated behaviors of theAfrican elephant (Loxodonta africana) in captivity. Zeitschrift fur
Tierpsychologie, 63, 63–79.
Brown, S. D., Dooling, R. J. & O’Grady, K. 1988. Perceptual
organization of acoustic stimuli by budgerigars (Melopsittacus
undulatus): III. Contact calls. Journal of Comparative Psychology,
102, 236–247.
Cheney, L., Seyfarth, R. M. & Silk, J. B. 1995. The role of grunts in
reconciling opponents and facilitating interactions among adultfemale baboons. Animal Behaviour, 50, 249–257.
Clutton-Brock, T. H. & Albon, S. D. 1979. The roaring of red deerand the evolution of honest advertising. Behaviour, 69, 145–170.
Dublin, H. T. 1983. Cooperation and reproductive competitionamong female African elephants. In: Social Behavior of Female
Vertebrates (Ed. by S. K. Wasser), pp. 291–313. New York:
Academic Press.
Gouzoules, H. & Gouzoules, S. 1990. Matrilineal signatures in the
recruitment screams of pigtail macaques, Macaca nemestrina.
Behaviour, 115, 327–347.
Grafe, T. U. 1996. The function of call alternation in the African reed
frog (Hyperolius marmoratus): precise call timing prevents auditorymasking. Behavioral Ecology and Sociobiology, 38, 149–158.
Graham, L. H., Schwarzenberger, F., Mostl, E., Galama, W. &Savage, A. 2001. A versatile enzyme-immunoassay for thedetermination of progestogens in feces and serum. Zoo Biology,
20, 227–236.
Graham, L. H., Bolling, J., Miller, G., Pratt-Hawkes, N. & Joseph, S.2002. An enzyme-immunoassay for the determination of luteiniz-
ing hormone in the serum of African elephants. Zoo Biology, 21,403–408.
Greenfield, M. D. 1994. Cooperation and conflict in evolution of
signal interactions. Annual Review of Ecology and Systematics, 25,97–126.
Greenfield, M. D. & Minckley, R. L. 1993. Acoustic dueling intarbush grasshoppers: settlement of territorial contests via
alternation of reliable signals. Ethology, 95, 309–326.
Greenfield, M. D. & Rand, S. 2000. Frogs have rules: selective
attention algorithms regulating chorusing in Physalaemus pustu-
losus (Lepodactylidae). Ethology, 106, 331–347.
Greenfield, M. D., Siegfreid, E. & Snedden, W. A. 2004. Variation
and repeatability of female choice in a chorusing katydid,
Ephippiger ephippiger: an experimental exploration of the pre-cedence effect. Ethology, 110, 287–299.

SOLTIS ET AL.: ANTIPHONAL CALLING IN ELEPHANTS 587
Griffin, A. S., Pemberton, J. M., Brotherton, P. N. M., McIlrath, G.,Gaynor, D., Kansky, R., O’Rian, J. & Clutton-Brock, T. H. 2003. A
genetic analysis of breeding success in the cooperative meerkat(Suricata suricatta). Behavioral Ecology, 14, 472–480.
Hall, M. J. 2000. The function of duetting in magpie-larks: conflict,cooperation or commitment? Animal Behaviour, 60, 667–677.
Hall, M. J. 2004. A review of hypotheses for the functions of avianduetting. Behavioral Ecology and Sociobiology, 55, 415–430.
Harcourt, A. H. & Stewart, K. J. 1996. Function and meaning ofwild gorilla ‘close’ calls. 2. Correlations with rank and relatedness.
Behaviour, 133, 827–845.
Hermes, R., Olson, D., Goritz, F., Brown, J. B., Schmitt, D. L.,Hagan, D., Peterson, J., Fritsch, G. & Hildebrandt, T. B. 2000.
Ultrasonography of the estrous cycle in female African elephants
(Loxodonta africana). Zoo Biology, 19, 369–382.
Janik, V. M. & Slater, P. J. B. 1998. Context-specific use suggests
that bottlenose dolphin signature whistles are cohesion calls.Animal Behaviour, 56, 829–838.
Jordan, K., Weiss, D., Hauser, M. & McMurray, B. 2004.Antiphonal responses to loud contact calls produced by Saguinus
oedipus. International Journal of Primatology, 25, 465–475.
Kalin, N. H., Shelton, S. E. & Snowdon, C. T. 1992. Affiliative
vocalizations in infant rhesus macaques (Macaca mulatta). Journal
of Comparative Psychology, 106, 254–261.
Langbauer, W. R. 2000. Elephant communication. Zoo Biology, 19,
425–445.
Langbauer, W. R., Payne, K. B., Charif, R. A., Rapaport, L. &Osborn, F. 1991. African elephants respond to distant playbacks
of low-frequency conspecific calls. Journal of Experimental Biology,157, 35–46.
Laws, R. M., Parker, I. S. C. & Johnstone, R. C. B. 1975. Elephantsand Their Habitats: the Ecology of Elephants in North Bunyoro,
Uganda. Oxford: Clarendon.
Lee, P. C. 1987. Allomothering among African elephants. AnimalBehaviour, 35, 278–291.
Lee, P. C. 1989. Family structure, communal care and femalereproductive effort. In: Comparative Socioecology: Behavioural
Ecology of Humans and Other Mammals (Ed. by V. Standen & R.
Foley), pp. 323–340. Oxford: Blackwell Scientific.
Leong, K. M., Ortolani, A., Burks, K. D., Mellen, J. D. & Savage, A.2003a. Quantifying acoustic and temporal characteristics of
vocalizations for a group of captive African elephants, Loxodontaafricana. Bioacoustics, 13, 213–231.
Leong, K. M., Ortolani, A., Graham, L. H. & Savage, A. 2003b. Theuse of low-frequency vocalizations in African elephant (Loxodonta
africana) reproductive strategies. Hormones and Behavior, 43, 433–
443.
McComb, K., Moss, C., Sayialel, S. & Baker, L. 2000. Unusually
extensive networks of vocal recognition in African elephants.Animal Behaviour, 59, 1103–1109.
McComb, K., Moss, C., Durant, S. M., Baker, L. & Sayailel, S.2001. Matriarchs as repositories of social knowledge in African
elephants. Science, 292, 491–501.
McComb, K., Reby, D., Baker, L., Moss, C. & Sayialel, S. 2003.
Long-distance communication and acoustic cues to social identityin African elephants. Animal Behaviour, 65, 317–329.
Mitani, J. C. & Nishida, T. 1993. Contexts and social correlates oflong-distance calling by male chimpanzees. Animal Behaviour, 45,
735–746.
Moss, C. J. & Poole, J. H. 1983. Relationships and social structure
of African elephants. In: Primate Relationships: an Integrated
Approach (Ed. by R. A. Hinde), pp. 315–325. Oxford: Blackwell
Scientific.
Mulder, R. A., Bishop, H., Cooper, M., Dennis, S., Koetsveld, M.,Marshall, J., Saunders, B. L. & Langmore, N. E. 2003. Alternativefunctions for duet and solo songs in magpie-larks, Grallina
cyanoleuca. Australian Journal of Zoology, 51, 25–30.
Payne, K. B., Langbauer, W. R. & Thomas, E. M. 1986. Infrasonic
calls of the Asian elephant (Elephus maximus). Behavioral Ecology
and Sociobiology, 18, 297–301.
Poole, J. H. 1999. Signal assessment in African elephants: evidence
from playback experiments. Animal Behaviour, 58, 185–193.
Poole, J. H., Payne, K., Langbauer, W. R. & Moss, C. 1988. The
social contexts of some very low frequency calls of African
elephants. Behavioral Ecology and Sociobiology, 22, 385–392.
Reby, D. & McComb, K. 2003. Anatomical constraints generate
honesty: acoustic cues to age and weight in the roars of red deerstags. Animal Behaviour, 65, 519–530.
Smith, H. J., Newman, J. D. & Symmes, D. 1982. Vocalconcomitants of affiliative behavior in squirrel monkeys. In: Primate
Communication (Ed. by C. T. Snowdon), pp. 30–49. Cambridge:
Cambridge University Press.
Smoski, M. J. & Bachorowski, J. A. 2003. Antiphonal laughter
between friends and strangers. Cognition and Emotion, 17,
327–340.
Soltis, J., Bernhards, D., Donkin, H. & Newman, J. D. 2002.
Squirrel monkey chuck call: vocal response to playbackchucks based on acoustic structure and affiliative relationship
with the caller. American Journal of Primatology, 57,
119–130.
Soltis, J., Leong, K. & Savage, A. 2005. African elephant vocal
communication II: rumble variation reflects the individual
identity and emotional state of callers. Animal Behaviour, 70,589–599.
Sugiura, H. 1998. Matching of acoustic features during the vocalexchange of coo calls by Japanese macaques. Animal Behaviour,
55, 673–687.
Wasserman, F. E. 1977. Intraspecific acoustical interference in the
white-throated sparrow (Zonotrichia albicollis). Animal Behaviour,
25, 949–952.





























