Embed Size (px)
Citation preview

1
Periplasmic Nicotine Dehydrogenase NdhAB Utilizes Pseudoazurin as Its 1
Physiological Electron Acceptor in Agrobacterium tumefaciens S33 2
3
Wenjun Yu,a Rongshui Wang,
a Haiyan Huang,
b Huijun Xie,
c and Shuning Wang
a# 4
5
State Key Laboratory of Microbial Technology, School of Life Science, Shandong 6
University, Jinan, People’s Republic of China,a Institute of Basic Medicine, Shandong 7
Academy of Medical Science, Jinan, People’s Republic of China,b and Environment 8
Research Institute, Shandong University, Jinan, People’s Republic of Chinac 9
10
# Address correspondence to Shuning Wang, [email protected] 11
12
RUNNING TITLE: Pseudoazurin involved in nicotine hydroxylation 13
14
KEYWORDS: hydroxylase, electron acceptor, nicotine dehydrogenase, pseudoazurin, 15
periplasm, nicotine degradation, Agrobacterium tumefaciens 16
17
18
AEM Accepted Manuscript Posted Online 16 June 2017Appl. Environ. Microbiol. doi:10.1128/AEM.01050-17Copyright © 2017 American Society for Microbiology. All Rights Reserved.
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 19
Agrobacterium tumefaciens S33 can grow with nicotine as the sole source of carbon, 20
nitrogen, and energy via a novel hybrid of the pyridine pathway and the pyrollidine 21
pathway. Characterization of the enzymes involved in the hybrid pathway is important 22
for understanding its biochemical mechanism. Here we report that the 23
molybdenum-containing nicotine dehydrogenase (NdhAB), which catalyzes the initial 24
step of nicotine degradation, is located in the periplasm of strain S33, while the 25
6-hydroxynicotine oxidase and 6-hydroxypseudooxynicoine oxidase are in the 26
cytoplasm. This is consistent with the fact that NdhA has a Tat signal peptide. 27
Interestingly, an ORF adjacent to the ndhAB gene was verified to encode a 28
copper-containing electron carrier, pseudoazurin (Paz), that has a typical signal 29
peptide of bacterial Paz. Both were transported into the periplasm after being 30
produced in the cytoplasm. We purified NdhAB from the periplasmic fraction of 31
strain S33 and found that, with Paz as the physiological electron acceptor, NdhAB 32
catalyzed the hydroxylation of nicotine at a specific rate of 110.52 ± 8.09 μmol min−1
33
mg protein−1
, where the oxygen atom in the hydroxyl group of the product 34
6-hydroxynicotine was derived from H2O. The apparent Km values for nicotine and 35
Paz were 1.64 ± 0.07 μM and 3.61 ± 0.23 μM, respectively. NAD(P)+, O2, and 36
ferredoxin could not serve as electron acceptors. Disruption of the paz gene disabled 37
the strain for nicotine degradation, indicating that Paz is required for nicotine 38
catabolism in the strain. These findings help our understanding of electron transfer 39
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

3
during nicotine degradation in bacteria. 40
41
IMPORTANCE 42
Nicotine is a toxic and addictive N-heterocyclic aromatic alkaloid produced in tobacco. 43
Its catabolism in organisms and degradation in tobacco wastes have become the major 44
concerns of health and the environment. Bacteria usually decompose nicotine using 45
the classical strategy of hydroxylating the pyridine ring with the help of activated 46
oxygen by nicotine dehydrogenase, which binds one molybdopterin, two [2Fe2S] 47
clusters, and usually one FAD as well. However, the physiological electron acceptor 48
for the reaction is still unknown. In this study, we found the two-component nicotine 49
dehydrogenase from Agrobacterium tumefaciens S33, naturally lacking FAD-binding 50
domain, is located in the periplasmic space and uses a copper-containing electron 51
carrier, pseudoazurin, as its physiological electron acceptor. We report here the role of 52
pseudoazurin in a reaction catalyzed by a molybdopterin-containing hydroxylase 53
occurring in the periplasmic space. These results provide new biochemical knowledge 54
in microbial degradation of N-heterocyclic aromatic compound. 55
56
57
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

4
INTRODUCTION 58
Nicotine is a toxic N-heterocyclic aromatic compound and is the main addictive 59
alkaloid produced in tobacco. Its catabolism in organisms and degradation in tobacco 60
wastes have become the major concerns of health and the environment (1-4). 61
Microbial degradation and transformation of nicotine are thought to be an 62
environmentally friendly method for disposing of tobacco wastes and a promising 63
technology for producing highly valuable drug and insecticide precursors from 64
nicotine (5-9). Various bacteria capable of degrading nicotine have been isolated and 65
characterized (5). They degrade nicotine mainly via two pathways: the pyridine 66
pathway, found in the Gram-positive bacterium Arthrobacter sp., and the pyrrolidine 67
pathway, identified in the Gram-negative bacterium Pseudomonas sp. (10-13). The 68
biochemical and molecular mechanisms involved in the two types of pathways have 69
been well elucidated. Recently, a hybrid form of the pyridine and pyrrolidine 70
pathways (Fig. 1) was discovered in Agrobacterium tumefaciens S33 (8), Shinella sp. 71
HZN7 (14, 15), and Ochrobactrum sp. SJY1 (16, 17). In the hybrid pathway, nicotine 72
is first degraded into 6-hydroxypseudooxynicotine via the pyridine pathway through 73
6-hydroxynicotine and 6-hydroxy-N-methylmyosmine and then produces 74
6-hydroxy-3-succinoylpyridine and 2,5-dihydroxypyridine along the pyrrolidine 75
pathway. These findings provide new insight for the microbial metabolic and 76
molecular diversity of nicotine catabolism in organisms. However, the biochemical 77
mechanism involved in the hybrid pathway still is not completely clear. 78
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

5
FIG 1 79
We previously purified nicotine dehydrogenase (NdhAB) and 80
6-hydroxy-3-succinoylpyridine hydroxylase (Hsh), two of the key enzymes involved 81
in the hybrid pathway, from the crude cell extracts of A. tumefaciens S33 (18, 19). 82
These enzymes are functionally similar to those in the pyridine pathway in 83
Arthrobacter nicotinovorans (20), and the pyrrolidine pathway in Pseudomonas 84
putida S16 (21), respectively. Hsh from A. tumefaciens S33 is an NADH-dependent 85
FAD-containing homodimeric monoxygenase with 62% identity to the enzyme from P. 86
putida S16 (21). It catalyzes the oxidative decarboxylation of 87
6-hydroxy-3-succinoylpyridine (HSP) to 2,5-dihydroxypyridine and succinic acid in 88
the presence of NADH and O2 (Fig. 1) (18). NdhAB from A. tumefaciens S33 is a 89
molybdenum-containing hydroxylase that was co-purified with a novel 90
6-hydroxypseudooxynicotine oxidase (Pno). They catalyzes the initial step of nicotine 91
oxidation to 6-hydroxynicotine, and the fourth step of the oxidative deamination of 92
6-hydroxypseudooxynicotine to 6-hydroxy-3-succinoylsemialdehyde pyridine, 93
respectively (Fig. 1) (19). In the complex, NdhA (82.4 kDa) harbors a molybdopterin 94
cofactor that has 27% and 14% identity to the large subunits of the heterodimeric 95
isoquinoline 1-oxidoreductase from Brevundimonas diminuta 7 (formerly 96
Pseudomonas diminuta 7) (22, 23) and the heterotrimeric NdhLMS from A. 97
nicotinovorans (20, 24), respectively. NdhB (17.1 kDa) harbors two [2Fe2S] clusters 98
and is 27% and 36.4% identical to the small subunits of isoquinoline 1-oxidoreductase 99
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

6
from B. diminuta 7 (22, 23) and NdhLMS from A. nicotinovorans (20, 24), 100
respectively. Pno (73.3 kDa) harbors an FMN and a [4Fe4S] cluster and is 48% 101
identical to histamine dehydrogenase from Pimelobacter simplex (formerly 102
Nocardioides simplex) (25). Compared with the three-component NdhLMS from A. 103
nicotinovorans, the middle-size subunit harboring an FAD is missing in the NdhAB 104
from strain S33. Interestingly, the encoding genes of NdhAB, Pno, and Hsh form a 105
gene cluster in the genome of strain S33, together with an ORF encoding a 106
flavoprotein almost identical (99%) to the 6-hydroxynicotine oxidase (Hno) from 107
Shinella sp. strain HZN7 (15) and VppB from Ochrobactrum sp. SJY1 (17). The 108
amine oxidase is predicted to catalyze the second step of the 6-hydroxynicotine 109
oxidation with O2 into 6-hydroxy-N-methylmyosmine forming H2O2 like its 110
orthologous enzymes, in strain S33. These results help greatly to understand the 111
biochemical processes behind the oxidative degradation of nicotine; however, the 112
reaction catalyzed by the NdhAB complex was determined only using an artificial dye 113
2,6-dichlorophenolindophenol (DCPIP); the actual physiological electron acceptor for 114
this enzymatic reaction remained unknown. This is an important question because the 115
electron transfer during nicotine oxidation plays a critical role in energy metabolism, 116
as strain S33 grows with nicotine as the sole source of carbon, nitrogen, and energy. 117
In this study, further efforts were put forth to discover the biochemical 118
mechanism involved in nicotine degradation by A. tumefaciens S33. We investigated 119
the activities of NdhAB, Hno, and Pno in periplasmic and cytoplasmic fractions, and 120
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

7
we purified the key enzyme NdhAB from the periplasmic fraction of A. tumefaciens 121
S33. We found an ORF adjacent to ndhAB that encodes a small blue copper protein, 122
pseudoazurin (Paz), which functions as the physiological electron acceptor in the 123
hydroxylation of nicotine, catalyzed by NdhAB in the periplasmic space. We report 124
here the role of Paz in a reaction catalyzed by a molybdopterin-containing 125
hydroxylase. These results provide new biochemical evidences for understanding 126
electron transfer during the oxidative degradation of nicotine via the special hybrid 127
pathway in A. tumefaciens S33. 128
129
RESULTS 130
Bioinformatics analysis of NdhAB, Pno, Hno, and Paz from A. tumefaciens S33. 131
In the genome of strain S33 (26), the encoding genes of NdhAB (locus tags in 132
GenBank: AWN88_01355 and 01360) and Pno (AWN88_01220) form a big cluster, 133
together with the genes of Hsh (AWN88_01205) and Hno (AWN88_01345). 134
Interestingly, one ORF next to ndhAB is predicted to encode Paz (AWN88_01350, 135
66-bp apart from ndhA; Fig. 2A). This kind of organization suggests that Paz might be 136
involved in nicotine degradation although ndhAB-paz has a different transcription 137
direction than others. We performed bioinformatics analysis on the protein sequences 138
of these enzymes for the transmembrane helix and signal peptide. Based on the 139
prediction using SignalP 4.1 Server (Technical University of Denmark, 140
http://www.cbs.dtu.dk/services/SignalP/), TatP 1.0 Server 141
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

8
(http://www.cbs.dtu.dk/services/TatP/), Signal-BLAST 142
(http://sigpep.services.came.sbg.ac.at/signalblast.html), and TMHMM Server v. 2.0 143
(http://www.cbs.dtu.dk/services/TMHMM/), none of the enzymes has a 144
transmembrane helix, indicating that they are not membrane bound proteins. This was 145
confirmed by the successful purification of NdhAB, Pno, and Hsh from the soluble 146
fractions of strain S33 (18, 19), and Hno as well (Yu et al. unpublished results). Signal 147
peptide prediction showed that NdhA (749 amino acids) has a typical Tat (twin 148
arginines translocation system) signal peptide at the N-terminus, similar to that in the 149
periplasmic nitrous oxide reductase (NosZ) from denitrifiers, such as Pseudomonas 150
fluorescens (Fig. 2B) (27). The Tat signal peptide is cleaved most likely at the site 151
between the 18th and 19th amino acid residues. Moreover, when the ORF is annotated 152
to encode Paz, it actually produces a Paz precursor (148 amino acid residues) that has 153
a signal peptide highly similar to those of the periplasmic Paz from Rhizobium 154
leguminosarum bv. viciae (28) and Paracoccus pantotrophus (formerly Thiosphaera 155
pantotropha) (29) (Fig. 2C). It is predicted that the bonds cleave between the 22nd 156
and 23rd amino acid residues. None of the enzymes, NdhB, Pno, or Hno, has a signal 157
peptide. Based on these results, we predicted that both NdhAB and Paz might be 158
transported into the periplasm after synthesis in the cytoplasm. 159
FIG 2 160
It is known that Paz is a small blue copper protein, and it plays an important role 161
in electron transfer in the periplasmic space. In the periplasm of P. pantotrophus it 162
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

9
functions as an electron donor for several enzymes of the denitrification pathway, 163
such as nitrite reductase (30), nitrous oxide reductase (31), and nitric oxide reductase 164
(32). In Hyphomicrobium denitrificans, the protein (named HdBCP) can pick up 165
electrons from cytochrome cL in the periplasmic space and subsequently transfer them 166
to the nitrite reductase (33). The Paz from A. tumefaciens S33 has 63% and 38% 167
identity to the proteins from Sinorhizobium meliloti (Paz2) (34) and P. pantotrophus 168
(29), respectively. It shares the common type I copper binding motif 169
[Cys-(X)n-His-(X)n-Met] of Pazs (Fig. 3A) (35, 36) and has the same rare initial 170
codon, TTG, as HdBCP, according to the sequence alignment. The qRT-PCR analysis 171
(Fig. 3B) showed that the transcriptional level of the paz gene was significantly 172
upregulated when strain S33 was cultured with nicotine as the sole source of carbon 173
and nitrogen. This was also demonstrated by the transcriptomic analysis, where a 174
Log2 Ratio (FPKM of Nic/FPKM of Glu) of 4.9 for the paz gene was determined in 175
nicotine medium (Nic) and glucose-ammonium medium (Glu) (Huang et al., 176
unpublished results). Thus, we predicted that Paz is involved potentially in the 177
oxidative degradation of nicotine, and that it most likely functions as the physiological 178
electron acceptor of NdhAB. 179
FIG 3 180
Location of NdhAB, Pno, and Hno in the cells of A. tumefaciens S33. To 181
identify the location of the NdhAB, Pno, and Hno enyzmes, we prepared the 182
periplasmic fraction and cytoplasmic fraction by treating fresh cells from strain S33 183
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

10
with lysozyme and EDTA and then determined their activities in these fractions. As 184
shown in Table 1, around 65% of the total activities of NdhAB were in the 185
periplasmic fraction, while around 90% of the Pno and Hno activities were in the 186
cytoplasmic fraction, indicating that only NdhAB was translocated into the 187
periplasmic space after being produced in the cytoplasm. These results were 188
consistent with the results of the signal peptide prediction. 189
TABLE 1 190
Purification of NdhAB from the periplasmic fractions of A. tumefaciens S33. 191
Compared with the previous procedure for purification of NdhAB from the crude cell 192
extracts of strain S33 (19), the procedure to purify NdhAB from the periplasmic 193
fractions of strain S33 was much simpler and included only an anion exchange step. 194
The periplasmic fraction (Table 1) was directly applied to a DEAE column, and 195
NdhAB activity was eluted at 0.4 M NaCl concentration in brown-colored fractions 196
that were already pure, as detected by SDS-PAGE. The activity of NdhAB with 197
DCPIP as an artificial electron acceptor was enriched ~18-fold with a yield and 198
specific activity of ~35.60% and 16.70 ± 1.28 U/mg, respectively. The obtained total 199
protein was 0.39 ± 0.02 mg and had a total activity of ~6.51 U. As shown by 200
SDS-PAGE analysis (Fig. 4A and B), only two bands could be detected, and they 201
were consistent in their sizes with NdhA and NdhB, which were reported previously 202
as 80 and 15 kDa, respectively (19). Results of the LC–MS analysis for the big band 203
cut from the gel (Fig. 4A) also confirmed that NdhA exists in the enzyme and was 204
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

11
purified from the periplasmic space (Fig. S1 in the supplemental material). However, 205
a small amount of Pno was sometimes co-purified together with NdhAB from the 206
periplasmic fraction of strain S33 (Fig. S2 in the supplemental material) due to a leak 207
from the cytoplasmic fraction when the membrane was broken during extractions 208
(Table 1). We reported previously that NdhAB and Pno could be co-purified from the 209
crude cell extracts of strain S33 that were prepared by sonication and were not easily 210
separated during the purification. From all these results together, we can conclude that 211
NdhAB is transported into the periplasm via the Tat system after synthesis in the 212
cytoplasm (37, 38), while Pno and Hno remain in the cytoplasm. Determination of the 213
N-terminal amino acid sequence of NdhA, purified from the periplasmic fraction of 214
strain S33, failed. We speculate that the N-terminus of NdhA might be modified 215
additionally after the signal peptide is cut off. Thus, the cleavage site of the signal 216
peptide in NdhA could not be defined. UV-visible absorption spectrum (Fig. 4C) of 217
the purified brown NdhAB as isolated showed a broad double peak at 340 and 467 nm, 218
and two shoulders in the regions of 420 and 553 nm, similar to those of the 219
heterodimeric isoquinoline 1-oxidoreductase, IorAB, from B. diminuta 7 (23). This is 220
consistent in that both enzymes bind a molybdenum cofactor and two [2Fe2S] clusters, 221
where 1.1 ± 0.2 mol of molybdenum (Mo) and 3.8 ± 0.3 mol of iron (Fe) were 222
measured per mol of NdhAB. The peaks and shoulders decreased upon treatment of 223
the enzyme with nicotine, indicating that the cofactors were reduced in the presence 224
of nicotine. A flavin cofactor was not detectable in the supernatant of the enzyme 225
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

12
sample after heat treatment. These data are consistent with the absence of typical 226
peaks of flavin in the UV-visible absorption spectrum. 227
FIG 4 228
Heterologous expression of paz in E. coli BL21 (DE3) and properties of 229
recombinant Paz. The complete paz gene, including the sequence encoding its signal 230
peptide, was cloned into the expression vector pET24a(+) and then subsequently 231
transformed into E. coli BL21 (DE3), which produced a fusion Paz with a His-tag at 232
the C-terminus in the periplasmic space. The recombinant cells were grown in LB at 233
16C for 36 h without induction, where the medium was supplemented with 2 mM 234
CuSO4 to enhance the copper incorporation. The target protein was extracted by 235
osmotic shock and then purified by first passing through a HisTrap HP column and 236
then a HiTrap Q HP column. The purified His-tagged protein from the periplasmic 237
fraction was blue with a molecular mass of 17 kDa, as determined by SDS-PAGE in 238
11.25% polyacrylamide gel (Fig. 3C). The UV-visible spectrum of the recombinant 239
Paz (Fig. 3D) revealed three broad peaks at 450, 595, and 760 nm; and the maximum 240
peak was close to 595 nm, which is typical for a type I copper protein (29, 39). The 241
protein had an A595nm/A280nm ratio of 0.24 ± 0.04, which was consistent with the Paz 242
purified from a mutant strain of P. denitrificans lacking cytochrome c550 (40). Its 243
extinction coefficient was determined to be 3.58 ± 0.09 mM−1
cm−1
at 595 nm and pH 244
7.0 when completely oxidized, which is similar to that of Paz from P. pantotrophus 245
(41). The determination of copper showed that 1 mM protein contained 0.88 ± 0.12 246
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

13
mM copper atoms, indicating that per mol of bound recombinant Paz there was 1 mol 247
of Cu, which is in agreement with previous reports (42-45). 248
The determination of N-terminal amino acid sequences from the recombinant Paz 249
purified from the periplasmic fraction of the recombinant E. coli BL21(DE3) cells 250
showed that the first 10 amino acids were ADHQVEMTNK, which were identical to 251
those of the deduced protein sequence from the paz gene of A. tumefaciens S33. The 252
expected signal peptide, composed of the first 25 amino acids of the recombinant Paz 253
with 3 amino acids from the vector pET24a(+), was cut off after the protein matured, 254
which was consistent with the signal peptide prediction from the SignalP 4.1 Server 255
(Fig. 2C). The first amino acid of the matured Paz, Ala, was the same as the protein 256
from H. denitrificans (33). These results also confirmed that the protein in strain S33 257
is located in the periplasmic space. 258
259
Activity of NdhAB with Paz as the electron acceptor. We previously used 260
DCPIP as an artificial electron acceptor to monitor the oxidation of nicotine (19). 261
Based on the bioinformatics analysis, the transcriptional analysis, and the knowledge 262
about the role of Paz in other bacteria, we predicted that Paz most likely functions as 263
the electron acceptor for NdhAB. Thus, we replaced phenazine methosulfate (PMS) 264
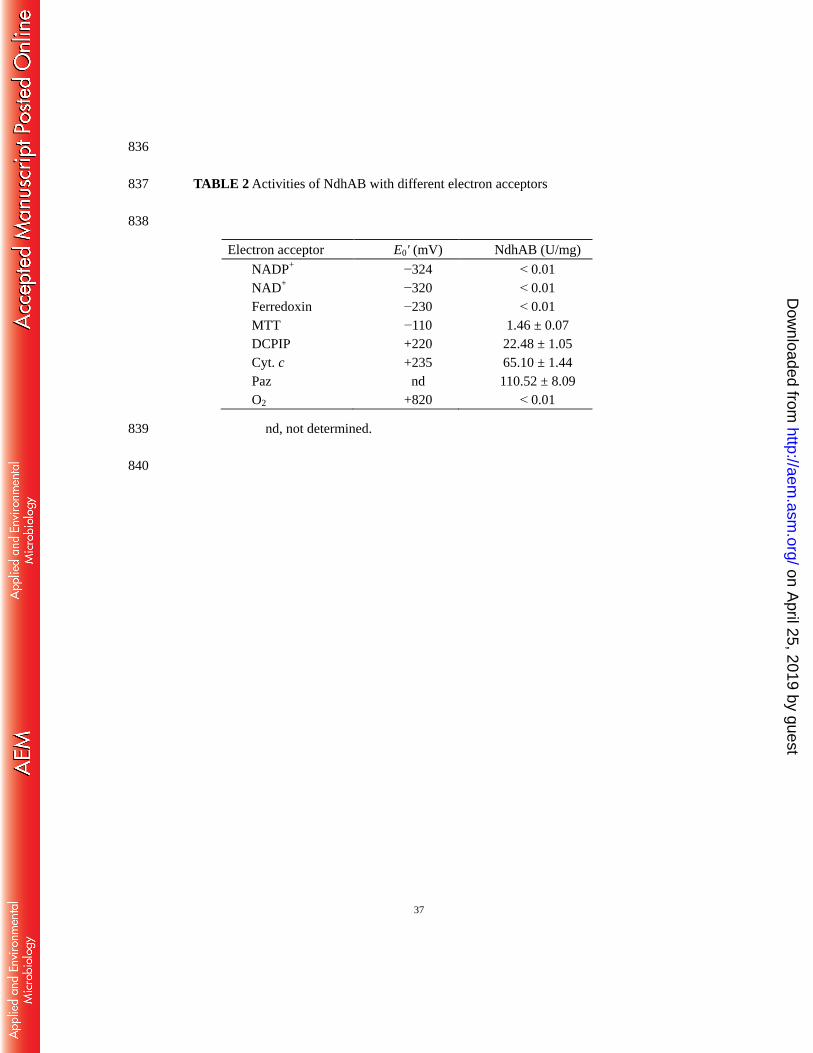
and DCPIP with Paz to test the activity of NdhAB. The results showed that Paz was 265
reduced significantly when NdhAB was added (Fig. 5A and B), with its activity of 266
110.52 ± 8.09 U/mg being similar to the previously reported blue copper protein, 267
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

14
HdBCP, that could accept electrons from cytochrome cL (33). Stoichiometric analysis 268
of the reaction showed that 1 mol nicotine oxidized can reduce 2.2 ± 0.067 mol Paz 269
(Fig. 5C), indicating that Paz is a single electron carrier and is in consistent with its 270
ability to bind a single copper atom while accepting or discharging one electron, 271
depending on its change in oxidation state (+2 or +1) (44). With Paz as the electron 272
acceptor, NdhAB showed the highest activity around pH 7.0 (Fig. S3 in the 273
supplemental material). The enzyme presented poor thermal stability, as in our 274
previous report (19). At pH 7.0 and the growth temperature of the strain, 30C, the 275
apparent Km values for nicotine and Paz were determined to be around 1.64 ± 0.07 μM 276
and 3.61 ± 0.23 μM (Fig. S4AB in the supplemental material), respectively. When 277
DCPIP was used as electron acceptor of NdhAB (Fig. S4CD in the supplemental 278
material), the apparent Km value for nicotine was 0.87 ± 0.03 μM, similar to our 279
previous report (19), while the apparent Km value for DCPIP was 1.24 ± 0.14 μM. For 280
both nicotine and the electron acceptor, Km values appear to be in the same order. The 281
results of LC−MS analysis (Fig. S5 in the supplemental material and Fig. 6) 282
demonstrated that NdhAB could catalyze the oxidation of nicotine (C10H14N2, 283
calculated molecular: 162.1157) into 6-hydroxynicotine (C10H14ON2, calculated 284
molecular: 178.1106) when Paz was used as the sole electron acceptor, indicating that 285
Paz could function as a physiological electron acceptor for NdhAB in strain S33. 286
FIG 5 287
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

15
Moreover, we also tested several other electron acceptors for the NdhAB activity. 288
Each assay mixture contained 50 mM phosphate buffer (pH 7.0), 1 mM nicotine, and 289
one of the following electron acceptors: O2 (feeding oxygen into the reaction system, 290
constantly), 0.05 mM each of DCPIP, 291
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide (MTT), Cyt. c, or 292
ferredoxin (harboring one [2Fe2S] cluster), or 0.1 mM each of NADP+ or NAD
+. As 293
shown in Table 2, NdhAB could not catalyze nicotine oxidation in the presence of 294
often used physiological electron acceptors such as O2, NAD+, NADP
+, or ferredoxin, 295
while the artificial electron acceptors DCPIP, MTT, and Cyt. c could be easily reduced 296
by nicotine. In comparison, Paz demonstrated the highest activity. Considering the 297
redox potentials of the tested electron acceptors, we speculate that the reaction 298
requires an electron acceptor with a relatively positive redox potential (e.g., higher 299
than +200 mV). This is consistent with the fact that Paz normally has a redox 300
potential between +200 mV and +277 mV (34). Protein sequence alignments showed 301
that Paz from strain S33 has 63% identity to Paz2 from S. meliloti, which has a redox 302
potential of +200 mV (34). Thus, the redox potential of Paz from strain S33 also 303
should be in this range. 304
TABLE 2 and FIG 6 305
To identify the source of the oxygen atom in the product of the reaction, 306
6-hydroxynicotine, as catalyzed by NdhAB, we performed the reaction assay with 307
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

16
18O-labeled water as one of the substrates and Paz as the electron acceptor. The 308
LC−MS analysis of the end reaction mixture showed that 6-hydroxynicotine was 309
labeled with 18
O (Fig. 6), indicating that the hydroxylation of nicotine incorporates the 310
oxygen atom derived from a water molecule into the product 6-hydroxynicotine. This 311
is the same as other typical molybdenum-containing hydroxylases (46). 312
313
Disruption of the paz gene. To further confirm the role of Paz in nicotine 314
degradation by strain S33, we disrupted the paz gene in strain S33 with the suicide 315
plasmid vector, pJQ200SK, harboring a truncated paz gene (Fig. S6 in the 316
supplemental material), and then determined whether nicotine degradation was 317
affected. The mutant strain was cultured in media with nicotine, 6-hydroxynicotine, or 318
HSP as the sole source of carbon and nitrogen. The results showed that A. tumefaciens 319
S33-∆paz could not utilize nicotine (Fig. 7), while it grew weakly on 320
6-hydroxynicotine or HSP, indicating that the Paz is required for the first step of 321
nicotine catabolism in strain S33 (Fig. 1). The function of Paz as an electron acceptor 322
during nicotine degradation in strain S33 could not be replaced by other electron 323
acceptors in cells, such as cytochrom c552 as in Marinobacter hydrocarbonclasticus 324
(47). Testing the nicotine transformation activity showed that the resting cells of the 325
mutant strain S33-∆paz prepared with glucose-ammonium sulfate medium 326
supplemented with nicotine or HSP could not transform nicotine, further confirming 327
the function of Paz in nicotine degradation. 328
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

17
FIG 7 329
DISCUSSION 330
We previously purified NdhAB from the crude cell extracts of A. tumefaciens S33 331
prepared by sonication and found that it was co-purified with Pno (19). Since they 332
were bound tightly to each other, we thought they might form a complex and function 333
by quickly detoxifying nicotine in cells, even though their encoding genes are far 334
away from each other. In this study, we verified that NdhAB was transported into the 335
periplasm of the cells after being produced in the cytoplasm, based on (1) finding a 336
Tat signal peptide in NdhA, (2) determination of ~65% of the total Ndh activities in 337
the periplasmic fraction, (3) purification of NdhAB, free of Pno, from the periplasmic 338
fractions of strain S33, and (4) identification of periplasmic Paz as the physiological 339
electron acceptor of NdhAB. By contrast, Pno and Hno were located in the cytoplasm 340
of the cells. These results changed our views on the mechanism of bacterial nicotine 341
degradation. However, when leaks occurred in the cytoplasmic fraction due to 342
spheroplast disprution during extracting the periplasmic fraction, very small amounts 343
of Pno could be still found to sticking to NdhAB (Fig. S2 in the supplemental 344
material), indicating that NdhAB and Pno have a strong interaction with each other. 345
Whether they could form a complex in the cytoplasm is unknown. The fact that Pno 346
could not utilize Paz as its electron acceptor (data not shown) suggests that Pno has no 347
direct connection with NdhAB in respect to their electron acceptors. 348
NdhAB catalyzes the initial step of nicotine degradation in A. tumefaciens S33 349
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

18
and hydroxylates nicotine at the 6-position carbon atom in the pyridine ring. This is a 350
crucial step for overcoming the resonance energy that stabilizes the ring structure 351
using the classical strategy of attacking the ring with the help of activated oxygen by 352
hydroxylation (48). Hydroxylases of N-heterocyclic aromatic compounds usually 353
belong to the xanthine oxidase family. They typically bind one redox center, called 354
molybdopterin, two [2Fe2S] clusters, and usually one FAD as well, which transport 355
electrons from the reducing substrate (N-heterocyclic aromatic compound) to the 356
oxidizing substrate (the electron acceptor) (49). The most common physiological 357
electron acceptors are NAD(P)+ and O2, which react at the FAD site; thus, the 358
intramolecular electron transfer between molybdenum and FAD via the intervening 359
iron−sulfur centers (46). Examples of these types of enzymes include 360
NADP-dependent nicotinate dehydrogenase from anaerobic bacterium Eubacterium 361
barkeri (50), NAD-dependent xanthine dehydrogenase from Rhodobacter capsulatus 362
(51), and O2-dependent quinaldine 4-oxidase from Arthrobacter sp. Rü61a (52). The 363
periplasmic aldehyde oxidoreductase (named YagTSR or PaoABC) from E. coli is an 364
exception. It uses ferredoxin, instead of NAD+ or molecular oxygen, as electron 365
acceptor (53). Nicotine dehydrogenase NdhLMS catalyzing the nicotine 366
hydroxylation in A. nicotinovorans (20) is also a typical heterotrimeric hydroxylase; 367
however, its physiological electron acceptor has not been known until now. Since it 368
was found to be partially associated with the membrane, it was speculated that the 369
enzyme, in its reduced form, associates strongly with the membrane and delivers its 370
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

19
electrons to the respiratory chain after the oxidized enzyme is reduced by its substrate 371
nicotine in the cytoplasm. NdhAB from A. tumefaciens S33 is a novel two-component 372
molydbedum-containing hydroxylase, which lacks the classical FAD-containing 373
subunit, just like the heterodimeric isoquinoline 1-oxidoreductase IorAB from B. 374
diminuta 7 (22, 23), aldehyde oxidoreductase (named Mop or Aor) from Desulfovibrio 375
gigas (54, 55), and nicotinate hydroxylase NicAB from P. putida KT2440 (49). 376
Interestingly, none of these could use the traditional electron acceptors, NAD(P)+ and 377
O2. Reportedly, Mop was able to use flavodoxin as electron acceptor (54). NicAB was 378
proposed to transfer electrons to a CytC oxidase via its CtyC domain on NicB (49). 379
Thus, the physiological electron acceptors for NdhAB and IorAB have remained 380
unknown. Both enzymes have a signal peptide, however, the signal peptide of IorAB 381
does not have a typical Tat motif, and its function is not known (22). 382
In this study, we identify that the small copper protein Paz functions as the 383
physiological electron acceptor of nicotine hydroxylation by NdhAB based on the 384
bioinformatics analysis, determination of the paz gene transcriptional level, 385
measurement of the Paz biochemical reaction, and disruption of the paz gene. The 386
experiments clearly showed that Paz is necessary for nicotine degradation and 387
functions efficiently as the electron acceptor in nicotine hydroxylation catalyzed by 388
NdhAB in the periplasm of A. tumefaciens S33. Like other hydroxylases of 389
N-heterocyclic aromatic compounds, the oxygen atom in the hydroxyl group of the 390
product, 6-hydroxynicotine, is derived from a water molecule in the Paz-dependent 391
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

20
reaction. Paz has been reported as the electron donor to the periplasmic cytochrome 392
cd1 nitrite reductase in P. pantotrophus (39), nitrous oxide reductase (31), 393
membrane-bound nitric oxide reductase in P. denitrificans (56), and copper containing 394
nitrite reductase in Achromobacter cycloclastes (57). Moreover, azurin, a blue 395
electron carrier very similar to Paz, can be used as an electron acceptor for the 396
molybdenum-containing and flavin-free arsenite oxidase AioAB, which is membrane 397
associated but is located in the periplasm of Alcaligenes faecalis, and belongs to the 398
DMSO reductase family (46, 58). This is very similar to the case reported in this study. 399
Surprisingly, cytochrome c can replace Paz or azurin in the reactions catalyzed by 400
AioAB and the denitrifying enzymes mentioned above (31, 56, 58). Paz can also 401
replace cytochrome c as the electron donor of cytochrome c peroxidase in 402
P. pantotrophus due to the so-called pseudospecificity in interactions with a range of 403
redox partners (41, 45). In this study, the Cyt. c from the horse heart also showed a 404
high activity when tested as an electron acceptor for NdhAB (Table 2), suggesting that 405
a Cyt. c in strain S33 might also be able to replace Paz in the nicotine-hydroxylating 406
reaction. However, we did not find a Cyt. c gene with high expression levels or that 407
had been upregulated, in nicotine medium in the transcriptomic analysis (Huang et al., 408
unpublished results), indicating that there is not a specific Cyt. c directly involved in 409
nicotine catabolism. Furthermore, we also tested the [2Fe2S]-type ferredoxin from 410
strain S33 as an electron acceptor, which did not show any activity for NdhAB (Table 411
2). These are consistent with the fact that strain S33 lost the ability to degrade nicotine 412
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

21
when the paz gene was disrupted, confirming that there is no intrinsic electron carrier 413
that can replace Paz (Fig. 7). 414
There is still an open question regarding the electron transfer in nicotine 415
catabolism: what is the destination of the electron carried on Paz? Since strain S33 416
grows on nicotine aerobically, the electron must be transferred ultimately to O2 via the 417
respiratory chain. For the electron transfer pathway, we speculate that the reduced Paz 418
might be re-oxidized by transferring its electron to the cytochromes in the respiratory 419
chain (e.g., transferred to Cyt. c, then to O2, catalyzed by a cytochrome oxidase). 420
Alternatively, the electron carried on Paz could be transferred directly to O2, catalyzed 421
by the cytochrome oxidase based on the “pseudospecificity” between Paz and Cyt. c. 422
Whether a Cyt. c participates in the pathway is dependent on the redox potentials of 423
Paz and Cyt. c. This is similar to two other cases of the electron transfer pathway: (1) 424
in the periplasmic methylamine dehydrogenase-amicyanin-cytochrome c551i complex 425
from P. denitrificans (59) and (2) in the electron transfer pathway between the 426
periplasmic sulfite dehydrogenase SorT and the respiratory chain via Cyt. c and azurin 427
in S. meliloti (60). Thus, determination of their redox potentials and interactions will 428
help to answer these questions in the future. 429
430
MATERIALS AND METHODS 431
Biochemicals, bacterial strains, plasmids, and culture condition. A. tumefaciens 432
S33 was isolated from tobacco plant rhizosphere and deposited at the China Center for 433
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

22
Type Culture Collection (CCTCC) under accession number CCTCC AB 2016054 434
(originally CCTCC M 206131) (61). It was grown aerobically at 30C in nicotine 435
medium plus 1.0 g l−1
glucose, 0.2 g l−1
(NH4)2SO4, and 1.0 g l−1
yeast extract (18). 436
Nicotine (99% purity, Fluka) was added to the medium before inoculation, with the 437
final concentration of 1.0 g l−1
. NADP+, NAD
+, DCPIP, MTT, and PMS were 438
purchased from Sigma-Aldrich (St. Louis, MO). Cyt. c from horse heart and H218
O 439
(97% purity) were purchased from Aladdin (Shanghai, China). [2Fe2S]-type 440
ferredoxin from strain S33 was heterologously produced and purified as described 441
(62). 6-Hydroxynicotine oxidase from A. nicotinovorans was a gift from Professor 442
Roderich Brandsch (University of Freiburg, Germany). 6-Hydroxynicotine was 443
obtained from Professor Brandsch or bought from Toronto Research Chemicals, Inc. 444
(Toronto, Canada). HSP was purified from the culture broth of the nicotine-degrading 445
P. putida S16 (9). E. coli BL21(DE3) was used for heterologous production of Paz, 446
and E. coli S17-1 was used for disruption of the paz gene. Both were cultivated with 447
LB medium at 37C. Kanamycin was used with a final concentration of 50 mg/L. 448
Analysis of real-time quantitative reverse transcription PCR (qRT-PCR). To 449
study the connection between the paz gene and nicotine degradation in strain S33, 450
qRT-PCR was performed. Strain S33 was cultured in nicotine medium (Nic) and 451
glucose-ammonium medium (Glu), where nicotine or glucose-ammonium sulfate as 452
the sources of carbon and nitrogen, respectively. Cells were harvested 453
at the prophase of logarithmic growth and total RNA was isolated using Ezol total 454
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

23
RNA extraction reagent (Shanghai GenePharma, China). DNA was digested using 455
DNaseI. The cDNA template was synthesized by reverse transcription using 456
TransScript First-Strand cDNA Synthesis SuperMix (Beijing TransGen Biotech, 457
China). qRT-PCR was performed using SYBR Real Time PCR Master Mix (Shanghai 458
GenePharma, China) on the MX3000P real-time PCR detection system (Stratagene) 459
with 16S rDNA as a reference gene. The following primers were used for qRT-PCR: 460
5'-ACTCTGGAACTGCCTTTGATA-3' (16S-F), 5'-CGTTTACGGCGTGGACTA-3' 461
(16S-R), 5'-GCACAACAGCGCCTCTATCAA-3' (paz-F), and 462
5'-CACGCCAGGGACATCAATCT-3' (paz-R). 463
Preparation of the periplasmic and cytoplasmic fractions of A. tumefaciens 464
S33. The periplasmic and cytoplasmic proteins were fractionated as previously 465
described (63, 64), with modification. The cells grown on nicotine medium plus 466
glucose, ammonium sulfate, and yeast extract were harvested late in the exponential 467
growth phase. To improve the efficiency of subsequent treatments by lysozyme, 468
EDTA was supplemented to the medium during growth to a final concentration of 1 469
mM. Fresh wet cells (2.6 g) were re-suspended in 130 ml of 200 mM Tris-HCl buffer 470
(pH 8.0) containing 2 mM EDTA, 20% sucrose, 1 mM phenylmethylsulfonyl fluoride, 471
and 1 mM MgCl2. After adding ~26 mg lysozyme from chicken egg whites (Sigma), 472
the mixture was incubated at 35°C for 40 min. The periplasmic fraction was obtained 473
after removing the spheroplasts and intact cells by centrifugation at 16,000 g and 474
4°C for 20 min and was concentrated using an Amicon Filter (30-kDa cutoff) before 475
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

24
performing enzyme assays and enzyme purification. To isolate the cytoplasmic 476
proteins, the resulting pellet was re-suspended in 130 ml chilled Tris-HCl buffer (20 477
mM, pH 8.0), followed by brief sonication (Vibra-Cell VCX 500, amplitude 39%, 1 478
min, pulse on 6 s, and pulse off 6 s) in an ice–water bath to break the spheroplasts. 479
The intact cells were removed by centrifugation at 16,000 g and 4°C for 20 min. The 480
resulting supernatant contained the cytoplasmic fraction and the membrane fraction. 481
The cytoplasmic fraction was obtained after removing the membrane fraction by 482
centrifugation at 190,000 g and 4°C for 1 h. 483
Enzyme assays. All the assays were carried out in quartz cuvettes (1-cm light 484
path) filled with 1 ml of reaction mixture at 30°C using a UV-visible Ultrospec 2100 485
pro Spectrophotometer (GE Healthcare, USA). Reactions were initiated by the 486
addition of enzyme. 487
NdhAB and Pno activities were routinely determined at 600 nm by monitoring 488
the reduction of DCPIP (ε = 21 mM−1
cm−1
) at 30°C, as described previously with 489
modifications (19). One unit was defined as the reduction of 1 µmol DCPIP per 490
minute. For NdhAB activity, the reaction mixture consisted of 50 mM phosphate 491
buffer (pH 7.0), 1 mM nicotine, 0.5 mM PMS, and 0.05 mM DCPIP. For Pno activity, 492
the reaction was coupled with the oxidation of 6-hydroxynicotine using Hno because 493
6-hydroxypseudooxynicotine was not commercially available. The reaction mixture 494
contained 50 mM Tris-HCl (pH 8.5), 100 mM NaCl, 0.56 mM 6-hydroxynicotine, 2 495
units of Hno from A. nicotinovorans, 0.5 mM PMS, and 0.1 mM DCPIP. 496
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

25
Hno activity was measured at 30°C by detecting the formation of 497
6-hydroxypseudooxynicotine as previously reported (65). The assay mixture 498
contained 0.56 mM 6-hydroxynicotine, 100 mM NaCl, and 100 mM Glycine-NaOH 499
buffer (pH 9.2). The formation of 6-hydroxypseudooxynicotine was followed at 334 500
nm (ε = 20.7 mM−1
cm−1
). One unit was defined as the production of 1-µmol 501
6-hydroxypseudooxynicotine. 502
Purification of NdhAB from the periplasmic fraction of A. tumefaciens S33. 503
The chromatography step was carried out at 18C using an ÄKTA Basic 10 504
chromatography system (GE Healthcare). The periplasmic fraction was applied to a 505
DEAE Sepharose Fast Flow (16 mm by 10 cm, 20 ml), equilibrated with 50 mM 506
phosphate buffer (pH 7.0). The column was eluted at 4 ml min−1
with five column 507
volumes of NaCl in a stepwise gradient at 0.1, 0.2, 0.25, 0.4, and 0.5 M 508
concentrations in the same buffer. NdhAB activity was eluted at 0.4 M NaCl 509
concentration. The purity was detected by SDS-PAGE. After concentrating and 510
desalting using an Amicon Filter (30-kDa cutoff), the protein was used for enzyme 511
assays. 512
Heterologous production and purification of the periplasmic Paz. The paz 513
gene, containing the signal peptide, was amplified by PCR with TransStart FastPfu 514
DNA Ploymerase (TransGen Biotech, Inc., Beijing, China) using A. tumefaciens S33 515
genomic DNA as a template. The following primers were used: 516
5′-CCGGCTAGCATGAATTTTCACACCATAGTATTTTC-3′ (forward, NheI 517
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

26
recognition site is underlined) and 5′-CCGCTCGAGTTTCTCTGCCTCGATCG-3′ 518
(reverse, XhoI recognition site is underlined). The target fragment was digested with 519
NheI and XhoI and ligated into the expression vector pET24a(+), which had been 520
digested at the same sites. The recombinant plasmids were sequenced and transferred 521
into E. coli BL21 (DE3). E. coli BL21 (DE3) strains carrying the recombinant 522
plasmids were grown at 16C for 36 h in LB medium supplemented with 50 mg/L 523
kanamycin and 2 mM CuSO4, without induction of 524
isopropyl-β-D-thiogalactopyranoside (IPTG) because of the damage to the bacterial 525
outer membrane (41). Cells were harvested by centrifugation at 6,000× g for 10 min 526
and then washed twice with 50 mM sodium phosphate buffer (pH 7.0). 527
The periplasmic Paz was extracted from E. coli BL21 (DE3) by osmotic shock 528
(29, 66). Cells of 3.72 g wet weight were suspended in 186 ml of 200 mM Tris-HCl 529
(pH 8.0) containing 20% sucrose (w/v), 2 mM EDTA, and 1 mM PMSF, and were 530
shaken for 10 min at room temperature before being centrifuged for 20 min at 531
16,000 g, 4C. The cell pellet was re-suspended in 186 ml of ice-cold 20 mM 532
Tris-HCl (pH 8.0). After shaking at 4C for 10 min and centrifuging again at 16,000 533
g and 4C for 20 min, the supernatant was used as the periplasmic fraction for 534
purifying Paz. 535
To obtain the pure Paz, the periplasmic fraction was loaded onto a HisTrap HP 536
column (GE Healthcare), which was equilibrated with 20 mM sodium phosphate 537
containing 500 mM NaCl (pH 7.4). The recombinant protein was then eluted with a 538
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

27
linear gradient imidazole (50–75 mM) in 20 mM sodium phosphate containing 500 539
mM NaCl (pH 7.4). The purified protein was concentrated by centrifugal 540
ultrafiltration (10-kDa cutoff). After being washed with 20 mM Tris-HCl (pH 8.0), the 541
protein was applied to a HiTrap Q HP Column (GE Healthcare) that was equilibrated 542
with 20 mM Tris-HCl (pH 8.0). The recombinant protein was eluted with 150 mM 543
NaCl in 20 mM Tris-HCl (pH 8.0), concentrated by centrifugal ultrafiltration (10-kDa 544
cutoff) and washed with 50 mM sodium phosphate buffer (pH 7.0). 545
Measurement of NdhAB activity with Paz as the electron acceptor. To 546
determine the activity of Paz in relation to NdhAB, the reaction mixture contained 50 547
mM sodium phosphate buffer (pH 7.0), 1 mM nicotine, and around 0.1 mM Paz. The 548
reaction was performed at 30°C and was initiated by the addition of NdhAB, purified 549
from the periplasmic fraction of A. tumefaciens S33. Paz reduction was recorded 550
spectrophotometrically at 595 nm. One unit was defined as the reduction of 1 µmol 551
Paz per minute. To test other electron acceptors, O2 (feeding oxygen in reaction 552
system, constantly) or 0.05 mM each of MTT, Cyt. c, or [2Fe2S]-ferredoxin, and 0.1 553
mM of NADP+ or NAD
+, were added. The detection wavelengths and molar 554
extinction coefficients were as follows: MTT, 578 nm, ε = 16.2 mM–1
cm–1
; Cyt. c, 555
550 nm, ε = 21.0 mM–1
cm–1
; [2Fe2S]-ferredoxin, 456 nm, ε∆OX-Red = 9.3 mM–1
cm–1
556
(62); NADP+ and NAD
+, 340 nm, ε = 6.2 mM
–1 cm
–1. One unit was defined as the 557
reduction of 1 µmol of electron acceptor per minute. 558
Analytical methods. Protein concentration was estimated using the Bradford 559
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

28
assay with bovine serum albumin as the standard (67). SDS-PAGE was performed 560
using a 12.5% or 6.0% gel with the Bio-Rad MiniProtean III cell. For N-terminal 561
amino acid sequence determination, the bands of target proteins in SDS-PAGE 562
analysis were transferred to a polyvinylidenedifluoride (PVDF) membrane by 563
electroblotting at 200 mV for 80 min, dyed by Coomassie Brilliant Blue R250, and 564
then degraded via Edman degradation. The N-terminal amino acid sequence was 565
analyzed using PPSQ-33A full-automatic protein polypeptide sequencer (Shimadzu, 566
Japan). 567
The UV-visible absorption spectra of NdhAB and Paz were determined using a 568
UV-visible Ultrospec 2100 Pro Spectrophotometer (GE Healthcare). For Paz, 569
complete oxidation of the protein was obtained by the addition of potassium 570
ferricyanide, which was then removed by loading the protein onto a Sephadex 200 571
column (10 mm by 30 cm, 24 ml). 572
The flavin in the protein was identified and determined by HPLC, as described 573
previously, after heat treatment of the protein samples (19). 574
The contents of Mo and Fe in NdhAB were determined with ICP-OES, as 575
described (19). The copper content in Paz was determined at 546 nm based on the 576
formation of a complex between Cu and 2,2-biquinoline in acetic acid (41). 577
The products of the reactions catalyzed by NdhAB were determined using a 578
Bruker’s Impact HD high-resolution mass spectrometry coupled to a Dionex’s 579
Ultimate 3000 UHPLC system (Thermo Scientific), according to the previous report 580
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

29
(19). 581
Disruption of the paz gene in A. tumefaciens S33. To confirm the key role of 582
paz in A. tumefaciens S33, the paz gene was inactivated by homologous 583
recombination using the suicide plasmid vector pJQ200SK, as described previously 584
(19) with modifications. Recombinant PCR was used to produce a 300-bp truncated 585
paz gene (∆paz) containing an upstream sequence of the paz gene (150 bp, using 586
primers A and B) and a downstream sequence of the paz gene (150 bp, using primers 587
C and D). The sequences of the primers used are as follows: 588
5′-CGCGGATCCTTGAATTTTCACACCATAGTATTTTC-3′ (primer A), 589
5′-GTGCGGGGTGCACTGGGGCTGGACGTGTAG-3′ (primer B), 590
5′-CTACACGTCCAGCCCCAGTGCACCCCGCAC-3′ (primer C), and 591
5′-CCGCTCGAGTTATTTCTCTGCCTCGATCG-3′ (primer D). The recombinant 592
plasmid pJQ-∆paz was constructed by ligating ∆paz gene into pJQ200SK with 593
BamHI/XhoI restriction sites. Then pJQ-∆paz was transformated into E. coli S17-1, 594
which was able to transfer the pJQ-∆paz plasmid into A. tumefaciens S33 by 595
conjugation, generating single-crossover mutants with Gm resistance as a selectable 596
marker. Double-crossover mutants, A. tumefaciens S33-∆paz, were selected after 597
growth in LB and HSP media with 20 % sucrose (w/v). 598
Nucleotide sequence accession number. The GenBank accession numbers of the 599
A. tumefaciens S33 genome are CP014259.1 and CP014260.1. 600
601
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

30
ACKNOWLEDGMENTS 602
This work was supported by the grant from the Fundamental Research Funds of 603
Shandong University (Grant No. 2014JC023). 604
We thank Professors Ping Xu from Shanghai Jiao Tong University and Luying 605
Xun from Washington State University for their valuable support and discussion. We 606
also acknowledge Professor Roderich Brandsch from University of Freiburg for the 607
gift of 6-hydroxynicotine and 6-hydroxynicotine oxidase. 608
We declare no conflicts of interest. 609
610
REFERENCES 611
612
1. Civilini M, Domenis C, Sebastianutto N, de Berfoldi M. 1997. Nicotine 613
decontamination of tobacco agro-industrial waste and its degradation by 614
microorganisms. Waste Manag Res 15:349-358. 615
2. Hecht SS, Hochalter JB, Villalta PW, Murphy SE. 2000. 2'-Hydroxylation of 616
nicotine by cytochrome P450 2A6 and human liver microsomes: formation of 617
a lung carcinogen precursor. Proc Natl Acad Sci U S A 97:12493-7. 618
3. Novotny TE, Zhao F. 1999. Consumption and production waste: another 619
externality of tobacco use. Tob Control 8:75-80. 620
4. Yildiz D. 2004. Nicotine, its metabolism and an overview of its biological 621
effects. Toxicon 43:619-32. 622
5. Liu J, Ma G, Chen T, Hou Y, Yang S, Zhang KQ, Yang J. 2015. 623
Nicotine-degrading microorganisms and their potential applications. Appl 624
Microbiol Biotechnol 99:3775-85. 625
6. Roduit JP, Wellig A, Kiener A. 1997. Renewable functionalized pyridines 626
derived from microbial metabolites of the alkaloid (S)-nicotine. Heterocycles 627
45:1687-1702. 628
7. Schmid A, Dordick JS, Hauer B, Kiener A, Wubbolts M, Witholt B. 2001. 629
Industrial biocatalysis today and tomorrow. Nature 409:258-68. 630
8. Wang S, Huang H, Xie K, Xu P. 2012. Identification of nicotine 631
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

31
biotransformation intermediates by Agrobacterium tumefaciens strain S33 632
suggests a novel nicotine degradation pathway. Appl Microbiol Biotechnol 633
95:1567-78. 634
9. Wang SN, Xu P, Tang HZ, Meng J, Liu XL, Qing C. 2005. "Green" route to 635
6-hydroxy-3-succinoyl-pyridine from (S)-nicotine of tobacco waste by whole 636
cells of a Pseudomonas sp. Environ Sci Technol 39:6877-80. 637
10. Brandsch R. 2006. Microbiology and biochemistry of nicotine degradation. 638
Appl Microbiol Biotechnol 69:493-8. 639
11. Li H, Li X, Duan Y, Zhang KQ, Yang J. 2010. Biotransformation of nicotine 640
by microorganism: the case of Pseudomonas spp. Appl Microbiol Biotechnol 641
86:11-7. 642
12. Tang H, Wang L, Wang W, Yu H, Zhang K, Yao Y, Xu P. 2013. Systematic 643
unraveling of the unsolved pathway of nicotine degradation in Pseudomonas. 644
PLoS Genet 9:e1003923. 645
13. Wang SN, Liu Z, Tang HZ, Meng J, Xu P. 2007. Characterization of 646
environmentally friendly nicotine degradation by Pseudomonas putida biotype 647
A strain S16. Microbiology 153:1556-65. 648
14. Ma Y, Wei Y, Qiu J, Wen R, Hong J, Liu W. 2014. Isolation, transposon 649
mutagenesis, and characterization of the novel nicotine-degrading strain 650
Shinella sp. HZN7. Appl Microbiol Biotechnol 98:2625-36. 651
15. Qiu J, Wei Y, Ma Y, Wen R, Wen Y, Liu W. 2014. A novel 652
(S)-6-hydroxynicotine oxidase gene from Shinella sp. strain HZN7. Appl 653
Environ Microbiol 80:5552-60. 654
16. Yu H, Tang H, Li Y, Xu P. 2015. Molybdenum-containing nicotine 655
hydroxylase genes in a nicotine degradation pathway that is a variant of the 656
pyridine and pyrrolidine pathways. Appl Environ Microbiol 81:8330-8. 657
17. Yu H, Tang H, Zhu X, Li Y, Xu P. 2015. Molecular mechanism of nicotine 658
degradation by a newly isolated strain, Ochrobactrum sp. strain SJY1. Appl 659
Environ Microbiol 81:272-81. 660
18. Li H, Xie K, Huang H, Wang S. 2014. 6-hydroxy-3-succinoylpyridine 661
hydroxylase catalyzes a central step of nicotine degradation in Agrobacterium 662
tumefaciens S33. PLoS One 9:e103324. 663
19. Li H, Xie K, Yu W, Hu L, Huang H, Xie H, Wang S. 2016. Nicotine 664
dehydrogenase complexed with 6-hydroxypseudooxynicotine oxidase 665
involved in the hybrid nicotine-degrading pathway in Agrobacterium 666
tumefaciens S33. Appl Environ Microbiol 82:1745-55. 667
20. Grether-Beck S, Igloi GL, Pust S, Schilz E, Decker K, Brandsch R. 1994. 668
Structural analysis and molybdenum-dependent expression of the 669
pAO1-encoded nicotine dehydrogenase genes of Arthrobacter nicotinovorans. 670
Mol Microbiol 13:929-36. 671
21. Tang H, Yao Y, Zhang D, Meng X, Wang L, Yu H, Ma L, Xu P. 2011. A novel 672
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

32
NADH-dependent and FAD-containing hydroxylase is crucial for nicotine 673
degradation by Pseudomonas putida. J Biol Chem 286:39179-87. 674
22. Lehmann M, Tshisuaka B, Fetzner S, Lingens F. 1995. Molecular cloning of 675
the isoquinoline 1-oxidoreductase genes from Pseudomonas diminuta 7, 676
structural analysis of iorA and iorB, and sequence comparisons with other 677
molybdenum-containing hydroxylases. J Biol Chem 270:14420-9. 678
23. Lehmann M, Tshisuaka B, Fetzner S, Roger P, Lingens F. 1994. Purification 679
and characterization of isoquinoline 1-oxidoreductase from Pseudomonas 680
diminuta 7, a novel molybdenum-containing hydroxylase. J Biol Chem 681
269:11254-60. 682
24. Igloi GL, Brandsch R. 2003. Sequence of the 165-kilobase catabolic plasmid 683
pAO1 from Arthrobacter nicotinovorans and identification of a 684
pAO1-dependent nicotine uptake system. J Bacteriol 185:1976-86. 685
25. Reed T, Lushington GH, Xia Y, Hirakawa H, Travis DM, Mure M, Scott EE, 686
Limburg J. 2010. Crystal structure of histamine dehydrogenase from 687
Nocardioides simplex. J Biol Chem 285:25782-91. 688
26. Yu W, Li H, Xie K, Huang H, Xie H, Wang S. 2016. Genome sequence of the 689
nicotine-degrading Agrobacterium tumefaciens S33. J Biotechnol 228:1-2. 690
27. Philippot L, Mirleau P, Mazurier S, Siblot S, Hartmann A, Lemanceau P, 691
Germon JC. 2001. Characterization and transcriptional analysis of 692
Pseudomonas fluorescens denitrifying clusters containing the nar, nir, nor and 693
nos genes. Biochim Biophys Acta 1517:436-40. 694
28. Patschkowski T, Schluter A, Priefer UB. 1996. Rhizobium leguminosarum bv. 695
viciae contains a second fnr/fixK-like gene and an unusual fixL homologue. 696
Mol Microbiol 21:267-80. 697
29. Leung YC, Chan C, Reader JS, Willis AC, van Spanning RJ, Ferguson SJ, 698
Radford SE. 1997. The pseudoazurin gene from Thiosphaera pantotropha: 699
analysis of upstream putative regulatory sequences and overexpression in 700
Escherichia coli. Biochem J 321 ( Pt 3):699-705. 701
30. Sam KA, Fairhurst SA, Thorneley RN, Allen JW, Ferguson SJ. 2008. 702
Pseudoazurin dramatically enhances the reaction profile of nitrite reduction by 703
Paracoccus pantotrophus cytochrome cd1 and facilitates release of product 704
nitric oxide. J Biol Chem 283:12555-63. 705
31. Berks BC, Baratta D, Richardson J, Ferguson SJ. 1993. Purification and 706
characterization of a nitrous oxide reductase from Thiosphaera pantotropha. 707
Implications for the mechanism of aerobic nitrous oxide reduction. Eur J 708
Biochem 212:467-76. 709
32. Thorndycroft FH, Butland G, Richardson DJ, Watmough NJ. 2007. A new 710
assay for nitric oxide reductase reveals two conserved glutamate residues form 711
the entrance to a proton-conducting channel in the bacterial enzyme. Biochem 712
J 401:111-9. 713
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

33
33. Hira D, Nojiri M, Yamaguchi K, Suzuki S. 2007. Identification of a blue 714
copper protein from Hyphomicrobium denitrificans and its functions in the 715
periplasm. J Biochem 142:335-41. 716
34. Laming EM, McGrath AP, Guss JM, Kappler U, Maher MJ. 2012. The X-ray 717
crystal structure of a pseudoazurin from Sinorhizobium meliloti. J Inorg 718
Biochem 115:148-54. 719
35. Adman ET. 1991. Copper protein structures. Adv Protein Chem 42:145-97. 720
36. Guss JM, Bartunik HD, Freeman HC. 1992. Accuracy and precision in protein 721
structure analysis: restrained least-squares refinement of the structure of poplar 722
plastocyanin at 1.33 Å resolution. Acta Crystallogr B 48 ( Pt 6):790-811. 723
37. Berks BC, Sargent F, Palmer T. 2000. The Tat protein export pathway. Mol 724
Microbiol 35:260-74. 725
38. Palmer T, Berks BC. 2012. The twin-arginine translocation (Tat) protein 726
export pathway. Nat Rev Microbiol 10:483-96. 727
39. Moir JW, Baratta D, Richardson DJ, Ferguson SJ. 1993. The purification of a 728
cd1-type nitrite reductase from, and the absence of a copper-type nitrite 729
reductase from, the aerobic denitrifier Thiosphaera pantotropha; the role of 730
pseudoazurin as an electron donor. Eur J Biochem 212:377-85. 731
40. Koutný M, Kucera I, Tesarik R, Turanek J, Van Spanning RJ. 1999. 732
Pseudoazurin mediates periplasmic electron flow in a mutant strain of 733
Paracoccus denitrificans lacking cytochrome c550. FEBS Lett 448:157-9. 734
41. Pauleta SR, Guerlesquin F, Goodhew CF, Devreese B, Van Beeumen J, Pereira 735
AS, Moura I, Pettigrew GW. 2004. Paracoccus pantotrophus pseudoazurin is 736
an electron donor to cytochrome c peroxidase. Biochemistry 43:11214-25. 737
42. Ferroni FM, Marangon J, Neuman NI, Cristaldi JC, Brambilla SM, Guerrero 738
SA, Rivas MG, Rizzi AC, Brondino CD. 2014. Pseudoazurin from 739
Sinorhizobium meliloti as an electron donor to copper-containing nitrite 740
reductase: influence of the redox partner on the reduction potentials of the 741
enzyme copper centers. J Biol Inorg Chem 19:913-21. 742
43. Inoue T, Nishio N, Suzuki S, Kataoka K, Kohzuma T, Kai Y. 1999. Crystal 743
structure determinations of oxidized and reduced pseudoazurins from 744
Achromobacter cycloclastes. Concerted movement of copper site in redox 745
forms with the rearrangement of hydrogen bond at a remote histidine. J Biol 746
Chem 274:17845-52. 747
44. Najmudin S, Pauleta SR, Moura I, Romao MJ. 2010. The 1.4 Å resolution 748
structure of Paracoccus pantotrophus pseudoazurin. Acta Crystallogr Sect F 749
Struct Biol Cryst Commun 66:627-35. 750
45. Williams PA, Fulop V, Leung YC, Chan C, Moir JW, Howlett G, Ferguson SJ, 751
Radford SE, Hajdu J. 1995. Pseudospecific docking surfaces on electron 752
transfer proteins as illustrated by pseudoazurin, cytochrome c550 and 753
cytochrome cd1 nitrite reductase. Nat Struct Biol 2:975-82. 754
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

34
46. Hille R, Hall J, Basu P. 2014. The mononuclear molybdenum enzymes. Chem 755
Rev 114:3963-4038. 756
47. Dell'acqua S, Pauleta SR, Monzani E, Pereira AS, Casella L, Moura JJ, Moura 757
I. 2008. Electron transfer complex between nitrous oxide reductase and 758
cytochrome c552 from Pseudomonas nautica: kinetic, nuclear magnetic 759
resonance, and docking studies. Biochemistry 47:10852-62. 760
48. Fuchs G, Boll M, Heider J. 2011. Microbial degradation of aromatic 761
compounds - from one strategy to four. Nat Rev Microbiol 9:803-16. 762
49. Jiménez JI, Canales A, Jimenez-Barbero J, Ginalski K, Rychlewski L, Garcia 763
JL, Diaz E. 2008. Deciphering the genetic determinants for aerobic nicotinic 764
acid degradation: the nic cluster from Pseudomonas putida KT2440. Proc Natl 765
Acad Sci U S A 105:11329-34. 766
50. Alhapel A, Darley DJ, Wagener N, Eckel E, Elsner N, Pierik AJ. 2006. 767
Molecular and functional analysis of nicotinate catabolism in Eubacterium 768
barkeri. Proc Natl Acad Sci U S A 103:12341-6. 769
51. Truglio JJ, Theis K, Leimkuhler S, Rappa R, Rajagopalan KV, Kisker C. 2002. 770
Crystal structures of the active and alloxanthine-inhibited forms of xanthine 771
dehydrogenase from Rhodobacter capsulatus. Structure 10:115-25. 772
52. Stephan I, Tshisuaka B, Fetzner S, Lingens F. 1996. Quinaldine 4-oxidase 773
from Arthrobacter sp. Ru61a, a versatile procaryotic molybdenum-containing 774
hydroxylase active towards N-containing heterocyclic compounds and 775
aromatic aldehydes. Eur J Biochem 236:155-62. 776
53. Neumann M, Mittelstadt G, Iobbi-Nivol C, Saggu M, Lendzian F, Hildebrandt 777
P, Leimkuhler S. 2009. A periplasmic aldehyde oxidoreductase represents the 778
first molybdopterin cytosine dinucleotide cofactor containing 779
molybdo-flavoenzyme from Escherichia coli. FEBS J 276:2762-74. 780
54. Barata BA, LeGall J, Moura JJ. 1993. Aldehyde oxidoreductase activity in 781
Desulfovibrio gigas: in vitro reconstitution of an electron-transfer chain from 782
aldehydes to the production of molecular hydrogen. Biochemistry 783
32:11559-68. 784
55. Romão MJ, Archer M, Moura I, Moura JJ, LeGall J, Engh R, Schneider M, 785
Hof P, Huber R. 1995. Crystal structure of the xanthine oxidase-related 786
aldehyde oxido-reductase from D. gigas. Science 270:1170-6. 787
56. Moir JWB, Ferguson SJ. 1994. Properties of a Paracoccus denitrificans 788
mutant deleted in cytochrome c550 indicate that a copper protein can substitute 789
for this cytochrome in electron-transport to nitrite, nitric oxide and nitrous 790
oxide. Microbiology-SGM 140:389-397. 791
57. Kashem MA, Dunford HB, Liu MY, Payne WJ, LeGall J. 1987. Kinetic 792
studies of the copper nitrite reductase from Achromobacter cycloclastes and its 793
interaction with a blue copper protein. Biochem Biophys Res Commun 794
145:563-8. 795
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

35
58. Anderson GL, Williams J, Hille R. 1992. The purification and characterization 796
of arsenite oxidase from Alcaligenes faecalis, a molybdenum-containing 797
hydroxylase. J Biol Chem 267:23674-82. 798
59. Choi M, Davidson VL. 2011. Cupredoxins--a study of how proteins may 799
evolve to use metals for bioenergetic processes. Metallomics 3:140-51. 800
60. Low L, Ryan Kilmartin J, Paul VB, Ulrike K. 2011. How are "atypical" sulfite 801
dehydrogenases linked to cell metabolism? Interactions between the SorT 802
sulfite dehydrogenase and small redox proteins. Front Microbiol 2:58. 803
61. Wang SN, Liu Z, Xu P. 2009. Biodegradation of nicotine by a newly isolated 804
Agrobacterium sp. strain S33. J Appl Microbiol 107:838-47. 805
62. Huang H, Hu L, Yu W, Li H, Tao F, Xie H, Wang S. 2016. Heterologous 806
overproduction of 2[4Fe4S]- and [2Fe2S]-type clostridial ferredoxins and 807
[2Fe2S]-type agrobacterial ferredoxin. Protein Expr Purif 121:1-8. 808
63. de Maagd RA, Lugtenberg B. 1986. Fractionation of Rhizobium 809
leguminosarum cells into outer membrane, cytoplasmic membrane, 810
periplasmic, and cytoplasmic components. J Bacteriol 167:1083-5. 811
64. Liu G, Liu M, Kim EH, Maaty WS, Bothner B, Lei B, Rensing C, Wang G, 812
McDermott TR. 2012. A periplasmic arsenite-binding protein involved in 813
regulating arsenite oxidation. Environ Microbiol 14:1624-34. 814
65. Brühmüller M, Mohler H, Decker K. 1972. Covalently bound flavin in 815
D-6-hydroxynicotine oxidase from Arthrobacter oxidans. Purification and 816
properties of D-6-hydroxynicotine oxidase. Eur J Biochem 29:143-51. 817
66. Neu HC, Heppel LA. 1965. The release of enzymes from Escherichia coli by 818
osmotic shock and during the formation of spheroplasts. J Biol Chem 819
240:3685-92. 820
67. Bradford MM. 1976. A rapid and sensitive method for the quantitation of 821
microgram quantities of protein utilizing the principle of protein-dye binding. 822
Anal Biochem 72:248-54. 823
824
825
826
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

36
TABLES 827
828
TABLE 1 Activities of NdhAB, Pno, and Hno in the periplasmic fraction (P), the 829
cytoplasmic fraction (C), and the membrane fraction (M) of A. tumefaciens S33. The 830
total protein in P, C+M, and C are 19.68 ± 1.38 mg, 92.62 ± 2.21 mg, and 82.34 ± 831
3.50 mg, respectively. Each value is the mean of three parallel replicates ± SD. 832
833
Enzyme Fraction Sp. act (U/mg) Activity in the
fraction (U) % in total activity
NdhAB P 0.93 ± 0.06 18.30 ± 1.18 66.40
C+M 0.10 ± 0.008 9.26 ± 0.74 33.60
C 0.11 ± 0.005 9.06 ± 0.41 32.87
Pno P 0.0052 ± 0.001 0.10 ± 0.02 3.37
C+M 0.031 ± 0.001 2.87 ± 0.09 96.63
C 0.032 ± 0.006 2.63 ± 0.49 88.55
Hno P 0.024 ± 0.002 0.47 ± 0.04 2.90
C+M 0.17 ± 0.02 15.75 ± 1.85 97.10
C 0.18 ± 0.004 14.82 ± 0.33 91.37
834
835
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from
37
836
TABLE 2 Activities of NdhAB with different electron acceptors 837
838
Electron acceptor E0' (mV) NdhAB (U/mg)
NADP+ −324 ˂ 0.01
NAD+ −320 ˂ 0.01
Ferredoxin −230 ˂ 0.01
MTT −110 1.46 ± 0.07
DCPIP +220 22.48 ± 1.05
Cyt. c +235 65.10 ± 1.44
Paz nd 110.52 ± 8.09
O2 +820 ˂ 0.01
nd, not determined. 839
840
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

38
FIGURE LEGENDS 841
842
FIG 1 The hybrid pathway of nicotine degradation by A. tumefaciens S33. NdhAB, 843
nicotine dehydrogenase; Paz, pseudoazurin; Hno, 6-hydroxynicotine oxidase; Pno, 844
6-hydroxypseudooxynicotine oxidase; Ald, aldehyde dehydrogenase; Hsh, 845
6-hydroxy-3-succinoylpyridine hydroxylase; Hpo, 2,5-dihydroxypyridine 846
dioxygenase; Nfo, N-formylmaleamate deformylase; Ami, maleamate amidohydrolase 847
(amidase); Iso, maleate cis/trans-isomerase. Ald is predicted by transcriptomic and 848
bioinformatics analysis and has not been biochemically identified. 849
850
FIG 2 The gene arrangement of NdhAB, Paz, Hno, and Pno in the genome of A. 851
tumefaciens S33 (A) and the possible signal peptides in NdhA (B) and Paz (C). NdhA 852
contains a Tat motif (RRAFL) in its signal peptide. NOSZ, nitrous oxide reductase 853
from P. fluorescens; AZUP, Paz from R. leguminosarum. The putative cleavage sites 854
were indicated as blue arrows. 855
856
FIG 3 Sequence analysis and heterologous production of Paz from A. tumefaciens 857
S33. A, protein sequence alignment with other bacterial Pazs whose crystal structures 858
have been determined. The boxed areas indicate four conserved amino acids for 859
copper binding. S. meliloti azu2 (PDB code, 3TU6; sequence identity to Paz from 860
strain S33, 63%), A. cycloclastes (1BQK, 50%), A. faecalis (1PAZ, 48%), H. 861
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

39
denitrificans (3EF4, 38%), P. pantotrophus (3ERX, 38%), and M. extorquens (1PMY, 862
38%). B, qRT-PCR analysis of the transcriptional level of the paz gene in A. 863
tumefaciens S33 cultured in glucose-ammonium sulfate medium (Glu) and nicotine 864
medium (Nic). The 16S rRNA gene was used as reference. Each value is the mean of 865
three parallel replicates ± SD. C, purification of recombinant Paz from A. tumefaciens 866
S33. Lane M, protein marker; lane 1, cell extract of E. coli BL21 (DE3); lane 2, 867
protein purified by HisTrap HP column; lane 3, protein purified by HiTrap Q HP 868
column. D, UV-visible absorption spectrum of recombinant Paz. The sample 869
contained 1.53 mg of purified protein/ml in 50 mM sodium phosphate buffer (pH 7.0). 870
An enlargement of the spectrum from 350 to 900 nm is shown in the inset. 871
872
FIG 4 SDS-PAGE analysis and UV-visible absorption spectra of NdhAB purified 873
from the periplasmic fraction of cell extracts from A. tumefaciens S33. A, 13% gel; B, 874
6% gel, where NdhB migrated outside the gel due to its small size. Lane M, protein 875
marker; lane 1, purified NdhAB. C, UV-visible absorption spectra of purified NdhAB 876
as isolated (solid line) and reduced by nicotine (dash line). The sample contained 0.84 877
mg of purified protein/ml in 50 mM sodium phosphate buffer (pH 7.0). The light path 878
of the cuvette is 3 mm. 879
880
FIG 5 The nicotine oxidation reaction catalyzed by periplasmic NdhAB from A. 881
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

40
tumefaciens S33 with Paz as the electron acceptor. A, spectrophotometric changes 882
from 350 to 900 nm during the transformation of nicotine. The reaction mixture 883
contained 50 mM sodium phosphate buffer (pH 7.0), 1 mM nicotine, 85 μM Paz, and 884
around 0.4 μg NdhAB purified from the periplasmic fraction of A. tumefaciens S33. B, 885
Paz reduction with nicotine catalyzed by NdhAB monitored at 595 nm. Solid line, the 886
reaction mixture containing all components mentioned above, where 73 μM Paz and 887
0.44 μg NdhAB were added; dashed line, the reaction mixture lacking nicotine was 888
used as a control, where 78 μM Paz and 0.44 μg NdhAB were added. C, stoichiometry 889
of Paz reduced and nicotine oxidized. Different amount of nicotine was added into the 890
reaction mixture, and the reduced Paz was determined according the decrease of 891
absorbance at 595 nm based on its extinction coefficient (3.58 mM−1
cm−1
). 892
893
FIG 6 LC−MS profiles of the reaction catalyzed by NdhAB in the presence of both 894
H218
O and H216
O. The reaction mixture contained 50 mM sodium phosphate buffer 895
(pH 7.0), around 0.1 mM Paz, 1 mM nicotine, and around 0.90 μg NdhAB purified 896
from the periplasmic fraction of A. tumefaciens S33; around 0.2 ml H218
O (97% purity) 897
was contained in the total mixture (0.5 ml). a, mass spectrum of the product 898
6-hydroxynicotine (m/z, 179.1172 for 16
O-labeled; m/z, 181.1215 for 18
O-labeled) 899
with retention time at 5.3 min. An enlargement of the spectrum with m/z ranging from 900
173 to 188 is shown in the inset, where the ratio of two peak areas is around 1.5, 901
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from

41
which is consistent with the ratio of H216
O/H218
O in the reaction mixture. b, mass 902
spectrum of the substrate nicotine (m/z, 163.1231) with retention time at 6.3 min. 903
904
FIG 7 Growth of A. tumefaciens S33 and A. tumefaciens S33-∆paz in the media with 905
nicotine (A), 6-hydroxynicotine (B), or HSP (C) as the sole source of carbon and 906
nitrogen. 1, wild-type strain; 2, S33-∆paz. 907
908
on April 25, 2019 by guest
http://aem.asm
.org/D
ownloaded from




































