Embed Size (px)
Citation preview

fmars-06-00087 March 29 2019 Time 1853 1
REVIEWpublished 02 April 2019
doi 103389fmars201900087
Edited byRob Harcourt
Macquarie University Australia
Reviewed byHeidi Dewar
Southwest Fisheries Science Center(NOAA) United States
Melinda Grace ConnersOld Dominion University
United States
CorrespondenceJena E Edwards
edwar11nuwindsorca
Specialty sectionThis article was submitted to
Marine Megafaunaa section of the journal
Frontiers in Marine Science
Received 14 August 2018Accepted 14 February 2019
Published 02 April 2019
CitationEdwards JE Hiltz E Broell FBushnell PG Campana SE
Christiansen JS Devine BMGallant JJ Hedges KJ MacNeil MAMcMeans BC Nielsen J Praeligbel K
Skomal GB Steffensen JF Walter RPWatanabe YY VanderZwaag DL and
Hussey NE (2019) AdvancingResearch for the Management
of Long-Lived Species A Case Studyon the Greenland Shark
Front Mar Sci 687doi 103389fmars201900087
Advancing Research for theManagement of Long-Lived SpeciesA Case Study on the GreenlandSharkJena E Edwards1 Elizabeth Hiltz2 Franziska Broell1 Peter G Bushnell3Steven E Campana4 Joslashrgen S Christiansen56 Brynn M Devine7 Jeffrey J Gallant8Kevin J Hedges9 M Aaron MacNeil10 Bailey C McMeans11 Julius Nielsen12Kim Praeligbel13 Gregory B Skomal14 John F Steffensen15 Ryan P Walter16Yuuki Y Watanabe17 David L VanderZwaag18 and Nigel E Hussey1
1 Department of Biological Sciences University of Windsor Windsor ON Canada 2 Fisheries Management Fisheriesand Oceans Canada Winnipeg MB Canada 3 Department of Biological Sciences Indiana University South Bend SouthBend IN United States 4 Faculty of Life and Environmental Sciences University of Iceland Reykjaviacutek Iceland 5 Faculty ofBiosciences Fisheries and Economics (BFE) Department of Arctic and Marine Biology (AMB) UiT The Arctic University ofNorway Tromsoslash Norway 6 Environmental and Marine Biology Aringbo Akademi University Turku Finland 7 Centre for FisheriesEcosystems Research Fisheries and Marine Institute of Memorial University of Newfoundland St Johnrsquos NL Canada8 Greenland Shark and Elasmobranch Education and Research Group Drummondville QC Canada 9 Arctic AquaticResearch Division Fisheries and Oceans Canada Winnipeg MB Canada 10 Department of Biology Ocean FrontierInstitute Dalhousie University Halifax NS Canada 11 Department of Biology University of Toronto MississaugaMississauga ON Canada 12 Greenland Institute of Natural Resources Nuuk Greenland 13 Norwegian College of FisheryScience UiT The Arctic University of Norway Tromsoslash Norway 14 Massachusetts Division of Marine Fisheries New BedfordMA United States 15 Department of Biology University of Copenhagen Copenhagen Denmark 16 Department of BiologicalSciences California State University Fullerton Fullerton CA United States 17 National Institute of Polar ResearchTachikawa Japan 18 Marine amp Environmental Law Institute Dalhousie University Halifax NS Canada
Long-lived species share life history traits such as slow growth late maturity and lowfecundity which lead to slow recovery rates and increase a populationrsquos vulnerabilityto disturbance The Greenland shark (Somniosus microcephalus) has recently beenrecognized as the worldrsquos longest-lived vertebrate but many questions regarding itsbiology physiology and ecology remain unanswered Here we review how current andfuture research will fill knowledge gaps about the Greenland shark and provide an overallframework to guide research and management priorities for this species Key advancesinclude the potential for specialized aging techniques and demographic studies toshed light on the distribution and age-class structure of Greenland shark populationsAdvances in population genetics and genomics will reveal key factors contributing tothe Greenland sharkrsquos extreme longevity range and population size and susceptibilityto environmental change New tagging technologies and improvements in experimentaland analytical design will allow detailed monitoring of movement behaviors andinteractions among Greenland sharks and other marine species while shedding light onhabitat use and susceptibility to fisheries interactions Interdisciplinary approaches suchas the combined use of stable isotope analysis and high-tech data-logging devices (ieaccelerometers and acoustic hydrophones) have the potential to improve knowledgeof feeding strategies predatory capabilities and the trophic role of Greenland sharksMeasures of physiology including estimation of metabolic rate as well as heart rate
Frontiers in Marine Science | wwwfrontiersinorg 1 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 2
Edwards et al Advancing Research for Long-Lived Species
and function will advance our understanding of the causes and consequences of longlifespans Determining the extent and effects of current threats (as well as potentialmitigation measures) will assist the development of policies recommendations andactions relevant for the management of this potentially vulnerable species Through aninterdisciplinary lens we propose innovative approaches to direct the future study ofGreenland sharks and promote the consideration of longevity as an important factor inresearch on aquatic and terrestrial predators
Keywords future directions longevity management Somniosus microcephalus Arctic ecosystem
INTRODUCTION
The classification of life history traits along a continuum betweenr- and K-selection is regarded as a foundational tool for predictingthe ability of animal populations to effectively respond toenvironmental andor anthropogenic disturbance (Pianka 1970)In higher order vertebrates extreme longevity is commonlyassociated with the possession of K-selected life history traits thatinfluence maximum intrinsic population growth rates (rmax) ndashsuch as slow growth late maturity and relatively low recruitmentrates (McCann and Shuter 1997) Such traits are thought tolimit the ability of animal populations to sustain high levelsof mortality (Adams 1980 Musick 1999a Simpfendorfer andKyne 2009 Norse et al 2012) In chondrichthyans the frequencyof these K-selected traits becomes more pronounced withincreasing depth (Rigby and Simpfendorfer 2015) resultingin significantly lower rates of population increase in deep-water species and a heightened vulnerability to exploitation andincidental mortality relative to their shallow-water counterparts(Garciacutea et al 2008 Simpfendorfer and Kyne 2009) Manydeep-water species are currently targeted in commercial fisheries(Barker and Schluessel 2005) and the number of deep-waterhabitats affected by fishing is expected to increase (Halpern et al2008) In addition impacts from other human activities (egseismic surveys) on deep-sea biota are virtually unknown asare the chronic and cumulative impacts from multiple stressorsincluding climate change and pollution
The importance of longevity in influencing the vulnerability ofanimal populations to overexploitation has long been overlookedin conservation and management planning It is only relativelyrecently following well-known examples of overfishing (egNew Zealand orange roughly Hoplostethus atlanticus andNorth Atlantic cod Gadus morhua Walters and Maguire1996 Clark 2001) and declines in many long-lived marinespecies (Myers and Worm 2003 Heppell et al 2005) thatthere has beenx increased consideration of K-selected traits inmarine management policy (Musick 1999b Musick et al 2000Baum et al 2003) This is especially true for the protectionof species targeted by commercial or recreational fisheriesincluding some deep-water chondrichthyan and teleost fishes(Stevens et al 2000 Simpfendorfer and Kyne 2009 Norse et al2012 Rigby and Simpfendorfer 2015) While current literatureasserts the importance of longevity for predicting the effects ofanthropogenic disturbance on wild populations the mechanisms
driving extreme longevity and its influences on animal behaviorand population dynamics are not well understood
The Greenland shark (Somniosus microcephalus) is a newlyrecognized example of an extremely long-lived deep-waterchondrichthyan (estimated lifespan of at least 272 years Nielsenet al 2016) that is both data deficient and vulnerable to humanthreats such as fishery-related mortality (Davis et al 2013)Greenland sharks are primarily known to inhabit deep-waterand coastal regions of the Arctic and North Atlantic (Yanoet al 2007 see Figure 1 for capture locations from the reviewedstudies) but have the potential to range globally where deep-water temperatures are lt5C (MacNeil et al 2012) and havebeen observed both at the surface and at confirmed depthsup to 1816 m (Campana et al 2015a Somniosid spp havealso been observed at 2200 and 2992 m Herdendorf andBerra 1995 Porteiro et al 2017 respectively) While historicallyfished in the waters off Iceland Greenland and Norway tosupport a liver oil industry decreased demand in the mid 20thcentury prompted the closure of targeted fisheries currentlyGreenland sharks are primarily caught as bycatch in commerciallongline gillnet and bottom trawl fisheries (Davis et al 2013)Despite a recent spike in public interest due to its reportedlongevity logistical challenges continue to impede the detailedstudy of this species Consequently relatively little is knownabout several important aspects of Greenland shark physiology(eg metabolism reproduction) ecology (eg age class structuredistribution ecological role genetic variability) and behavior(eg predatory capabilities horizontal movement patterns)The primary objective of this paper is to highlight importantknowledge gaps that limit the development of a managementframework for the Greenland shark In the following text wediscuss a variety of forward-thinking experimental approachesthat will enable researchers to answer key questions aboutGreenland shark biology and ecology (Table 1) Additionally wepresent these current and future advances in Greenland sharkresearch within the context of broader issues regarding the studyof long-lived species (Table 2)
METHODS
Experts in the study of Greenland sharks from severalresearch and management institutions were consulted Guidedby the previous scientific contributions of this expert team
Frontiers in Marine Science | wwwfrontiersinorg 2 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 3
Edwards et al Advancing Research for Long-Lived Species
FIGURE 1 | Geographic coverage of Greenland shark studies by sampling location and research topic (A) locations of Greenland shark capturesampling (pointsrepresent sampling regions reported by each study multiple points used for studies with several disparate sampling sites) (B) study count by country of Greenlandshark capturesampling and research focus (studies counted more than once where multiple sampled countries are reported)
eight topics were deemed to be of high priority to assistfuture Greenland shark research and management The chosentopics are outlined in the text as follows (1) Demographicsand life history (2) Population genetics and genomics (3)Movement ecology (4) Behavior (5) Physiological adaptations(6) Diet and trophic ecology (7) Threats to Greenland sharkpopulations and (8) Management (see Figure 1 for study
count by research topic) Each topic was then organizedinto four key sections (i) current knowledge in the chosenfield (ii) identified knowledge gaps and limitations (iii)proposed strategies to address identified data deficiencies andfinally (iv) potential recommendations for Greenland sharkmanagement While some topic overlap among sections wasunavoidable due to the interconnected nature of much of
Frontiers in Marine Science | wwwfrontiersinorg 3 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 4
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|An
over
view
ofcu
rren
tkno
wle
dge
and
data
defic
ienc
ies
inei
ghtfi
elds
ofG
reen
land
shar
k(S
omni
osus
mic
roce
phal
us)r
esea
rch
stra
tegi
esto
addr
ess
data
defic
ienc
ies
and
pote
ntia
lact
ions
tosu
ppor
tm
anag
emen
torg
aniz
atio
nre
com
men
datio
nsan
dG
reen
land
shar
kco
nser
vatio
n
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Dem
ogra
phic
san
dlif
ehi
stor
yLi
fesp
anan
dag
ean
dsi
zeat
mat
urity
estim
ated
Lo
cala
bund
ance
sat
seve
rals
ites
inth
eC
anad
ian
Art
ices
timat
ed
Pot
entia
ljuv
enile
habi
tats
iden
tified
(Can
adia
nan
dR
ussi
anA
rctic
)
Sto
ckas
sess
men
tP
opul
atio
nsi
zes
grow
thra
tes
mor
talit
yra
tes
sex
ratio
san
dsu
stai
nabl
eha
rves
tlev
els
unkn
own
Are
adily
appl
icab
lean
dva
lidat
edag
ede
term
inat
ion
met
hod
nota
vaila
ble
Rep
rod
ucti
on
Rep
rodu
ctiv
ebi
olog
yan
dec
olog
yla
rgel
yun
know
n
Sto
ckas
sess
men
t(i)
Dev
elop
rout
ine
agin
gm
etho
d(ii
)Agi
ngof
indi
vidu
als
tode
term
ine
age-
at-le
ngth
and
info
rmpo
pula
tion
age
stru
ctur
e(ii
i)U
seB
RU
Vs
surv
eys
tobu
ilda
time
serie
son
whi
chto
estim
ate
biom
ass
and
abun
danc
ein
dice
s(v
iaa
rand
omsa
mpl
ing
desi
gna
cons
iste
ntan
nual
sam
plin
gre
gim
ean
da
broa
dne
twor
kof
sam
plin
gsi
tes)
(iv
)Req
uest
deta
iled
repo
rtin
gof
biol
ogic
alm
etric
s(e
g
leng
ths
exa
nddi
spos
ition
)for
byca
ught
shar
ksby
fishe
ries
obse
rver
sR
epro
duc
tio
n(i)
Use
ultr
asou
ndte
chni
ques
toid
entif
ypr
egna
ntfe
mal
es
(ii)E
lect
roni
cta
gssu
ture
din
tocl
oaca
ofpr
egna
ntfe
mal
esco
uld
help
dete
rmin
ebi
rthi
ngtim
ean
dlo
catio
n(a
spe
rBin
der
etal
20
14)
Sto
ckas
sess
men
t(i)
Dev
elop
Har
vest
Str
ateg
yin
clud
ing
Har
vest
Con
trol
Rul
es(e
g
ICE
SA
ppro
ach
toD
ata-
Lim
ited
Sto
cks)
(ii
)Pla
cevi
deo
mon
itorin
gon
fishi
ngve
ssel
sto
reco
rdpr
esen
cea
bsen
ceof
clas
pers
toes
timat
ese
xra
tio
(iii)
Com
pare
shar
kle
ngth
mea
sure
men
tsto
age-
at-le
ngth
toes
timat
eag
ecl
ass
abun
danc
ere
lativ
eca
ptur
era
tes
Rep
rod
ucti
on
Pro
tect
impo
rtan
tmat
ing
nurs
ery
area
san
dag
greg
atio
nsi
tes
(upo
nid
entifi
catio
n)
Pop
ulat
ion
gene
tics
and
geno
mic
sM
itoch
ondr
ialg
enom
eha
sbe
ense
quen
ced
Spe
cies
phyl
ogen
yha
sbe
ende
term
ined
E
volu
tiona
ryhi
stor
yha
sbe
enth
eoriz
ed
Loca
tion
ofhy
brid
izat
ion
with
Som
nios
uspa
cific
usun
know
nG
ener
atio
ntim
esan
dge
netic
varia
bilit
yam
ong
indi
vidu
als
and
popu
latio
nsun
know
nG
enet
ical
lyde
term
ined
susc
eptib
ility
toen
viro
nmen
talc
hang
eun
clea
r
Dat
aco
llect
ion
Req
uest
at-s
eaob
serv
ers
toco
llect
non-
leth
altis
sue
orpa
rasi
te(iD
NA
)sa
mpl
es
Defi
nep
op
ulat
ions
(i)
Use
gene
tics
toes
timat
ege
nera
tion
times
and
effe
ctiv
epo
pula
tion
size
(eg
us
ing
half
sibl
ing
appr
oach
)(ii
)Defi
nege
netic
ally
dist
inct
popu
latio
nss
tock
sD
efine
rang
eE
xam
inat
ion
ofhy
brid
izat
ion
inso
uthe
rnre
gion
s(e
g
Gul
fofM
exic
o)co
uld
exte
ndth
ekn
own
rang
eof
Gre
enla
ndsh
arks
beyo
ndth
epo
lar
regi
ons
Defi
nep
op
ulat
ions
and
rang
eR
ecom
men
dap
prop
riate
size
and
loca
tion
ofm
anag
emen
tbou
ndar
ies
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 4 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 5
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Mov
emen
teco
logy
Sea
sona
lmo
vem
ents
(i)
Hor
izon
talt
rack
ing
show
slo
ng-d
ista
nce
timed
dire
cted
mig
ratio
ns
(ii)O
bser
ved
occu
rren
cein
shal
low
erw
ater
sw
ithin
crea
sing
latit
ude
(iii)
Vis
ualr
ecor
dsof
philo
patr
yob
serv
edin
St
Law
renc
eE
stua
ry(G
alla
ntpe
rson
alco
mm
unic
atio
n)
Vert
ical
mo
vem
ents
(i)
Rec
ords
ofm
ovem
enti
nsh
allo
ww
ater
and
mid
wat
erco
lum
nun
der
ice
and
ines
tuar
ine
wat
ers
and
(aco
ustic
and
pop-
offs
atel
lite
trac
king
)(ii
)Ver
tical
mov
emen
tsap
pear
tobe
asso
ciat
edw
ithlig
htcy
cle
insh
allo
ww
ater
Sea
sona
lmo
vem
ents
(i)
Lack
oflo
ng-t
erm
(ie
mul
ti-ye
ar)
reco
rds
show
ing
seas
onal
ann
ual
mov
emen
ts
Vert
ical
mo
vem
ents
Li
mite
dre
cord
sof
deta
iled
vert
ical
mov
emen
tbeh
avio
rsav
aila
ble
(eg
ea
rlyde
pth
and
tem
pera
ture
reco
rds
binn
edat
60sndash
6h
inte
rval
s)
Dis
trib
utio
nS
hort
-ter
mm
onito
ring
insu
ffici
entt
oca
ptur
eco
mpl
ete
rang
eof
mov
emen
ts
Sea
sona
lmo
vem
ents
In
crea
sem
onito
ring
dura
tion
usin
glo
ng-t
erm
pass
ive
acou
stic
arc
hiva
lor
sate
llite
met
hods
and
impr
oved
tech
nolo
gies
(eg
ex
tend
edba
tter
ylif
ede
pth
ratin
gse
tc)
Vert
ical
mo
vem
ents
(i)
Use
high
-res
olut
ion
time-
serie
sda
tato
crea
teco
ntin
uous
mov
emen
ttra
cks
(ii)C
reat
est
ate-
spac
em
odel
sto
deco
mpo
sedi
ves
and
asso
ciat
eos
cilla
tory
patt
erns
with
disc
rete
beha
vior
alty
pes
(eg
po
st-r
elea
sere
cove
ry)
(iii)
Com
pare
over
lap
inve
rtic
aldi
strib
utio
nw
ithth
atof
fishe
ries
targ
etsp
ecie
sD
istr
ibut
ion
Use
larg
e-sc
ale
pass
ive
acou
stic
infra
stru
ctur
ean
dlo
ng-t
erm
sate
llite
tele
met
ryap
proa
ches
tode
fine
rang
ean
did
entif
ym
atin
gpu
ppin
ggr
ound
s
Sea
sona
lmo
vem
ents
(i)
Pot
entia
lsea
sona
lclo
sure
sfo
rhu
man
activ
ities
inan
nual
high
use
area
sto
min
imiz
ene
gativ
eim
pact
sto
Gre
enla
ndsh
ark
popu
latio
ns
(ii)D
ynam
icA
rea
Man
agem
entt
opr
otec
tun
expe
cted
aggr
egat
ions
Ve
rtic
alm
ove
men
ts
Info
rmfis
hha
rves
ters
ofsh
ark
beha
vior
patt
erns
tofa
cilit
ate
effo
rts
tom
inim
ize
shar
kca
tch
(eg
se
tgea
rat
dept
hsan
dtim
esof
day
whe
nsh
arks
tend
not
tobe
pres
ent)
Dis
trib
utio
nB
ilate
ralo
rm
ulti-
late
rala
gree
men
tsbe
twee
nna
tions
orvi
aR
egio
nal
Fish
erie
sM
anag
emen
tOrg
aniz
atio
ns(R
FMO
s)to
cond
ucts
cien
cean
dm
anag
esp
ecie
son
apo
pula
tion
leve
l
Beh
avio
ran
din
tera
ctio
nsC
aptu
rean
dre
leas
eP
relim
inar
yta
ggin
gst
udie
sha
veex
amin
edpo
st-r
elea
sebe
havi
oran
dsu
rviv
alra
tes
follo
win
gex
perim
enta
lca
ptur
eFo
rag
ing
beh
avio
r(i)
Indi
rect
evid
ence
ofliv
epr
edat
ion
onse
als
(ii)S
low
swim
spee
dsre
lativ
eto
body
size
buth
igh
roll
acce
lera
tion
(iii)
Dire
ctob
serv
atio
nsof
scav
engi
ng
(iv)A
ggre
gatio
nbe
havi
orob
serv
ed
Cap
ture
and
rele
ase
Pos
t-re
leas
esu
rviv
alra
tes
requ
irefu
rthe
rst
udy
(cur
rent
lyin
suffi
cien
tfor
adju
stin
gm
orta
lity
fact
ors
ina
popu
latio
nm
odel
)Fo
rag
ing
beh
avio
r(i)
Tact
ics
ofpr
eyca
ptur
eun
know
n(a
cryp
ticpr
edat
ion
appr
oach
hypo
thes
ized
)(ii
)Nat
ure
and
frequ
ency
ofin
tra-
and
inte
rspe
cific
inte
ract
ions
unkn
own
Fora
gin
gb
ehav
ior
(i)U
seca
mer
as(s
tilla
ndvi
deo)
depl
oyed
onva
rious
plat
form
s(e
g
anim
als
Rem
ote
Ope
rate
dVe
hicl
esfi
shin
gge
arc
able
s)to
obse
rve
fora
ging
beha
vior
dire
ctly
(ii)U
sean
imal
-bor
nehy
drop
hone
s(s
ampl
eso
unds
cape
)to
asse
ssin
tera
ctio
nsw
ithvo
caliz
ing
anim
als
(eg
m
arin
em
amm
als)
(ii
i)U
seof
VM
Ts(a
cous
tictr
ansp
onde
rs)t
oex
amin
ein
tra-
and
inte
r-sp
ecifi
cin
tera
ctio
ns
Cap
ture
and
rele
ase
Dev
elop
and
prom
ote
safe
hand
ling
prac
tices
for
the
rele
ase
ofsh
arks
caug
htas
byca
tch
Fora
gin
gb
ehav
ior
Use
unde
rsta
ndin
gof
feed
ing
mod
es(e
g
suct
ion
feed
ing)
and
pref
eren
ces
toin
form
gear
mod
ifica
tions
gea
rde
ploy
men
tan
dba
itus
e
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 5 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 6
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Phy
siol
ogic
alad
apta
tions
Die
tand
trop
hic
ecol
ogy
Phy
sio
log
y(i)
Exh
ibit
extr
eme
long
evity
(est
imat
edbe
twee
n27
2an
d51
2ye
ars)
(ii
)Hig
han
tioxi
dant
enzy
me
leve
lsan
dlo
wbl
ood
O2
affin
ityre
cord
ed
Sen
sory
abili
ties
(i)
Olfa
ctor
yor
gans
high
lyde
velo
ped
cons
iste
ntw
ithbe
nthi
cpr
edat
ors
cave
nger
lifes
tyle
(ii
)Hig
hin
cide
nce
ofbl
indn
ess
due
toey
epa
rasi
tism
(exc
epti
nth
eS
tLa
wre
nce
Riv
er)
(iii)
Obs
erva
tions
ofvi
sion
-dep
enda
ntbe
havi
ors
sugg
esta
bilit
yto
unde
rtak
eac
tive
pred
atio
nD
iet
Ben
thic
and
pela
gic
feed
ing
from
pred
omin
antly
offs
hore
tele
osts
ourc
essu
gges
ted
Eco
log
ical
role
G
reen
land
shar
ksar
elik
ely
impo
rtan
tpr
edat
ors
and
likel
yoc
cupy
ahi
ghtr
ophi
cpo
sitio
n
Phy
sio
log
y(i)
Max
imum
rese
rve
dens
itysu
gges
tslif
espa
nm
aycu
rren
tlybe
unde
rest
imat
ed
(ii)S
ensi
tivity
toen
viro
nmen
tals
hifts
unkn
own
(iii)
Det
ails
ofhe
artf
unct
ion
unkn
own
(may
play
aro
lein
long
evity
)M
etab
olic
rate
(i)
Met
abol
icra
tean
dra
teof
O2
cons
umpt
ion
unkn
own
(ii)P
oten
tiall
ink
betw
een
hem
oglo
bin
prop
ertie
san
dco
ld-t
oler
ance
(ii
i)P
oten
tiall
inks
betw
een
high
antio
xida
nten
zym
ele
vels
and
cold
tole
ranc
ede
epdi
ving
beha
vior
and
long
evity
S
enso
ryab
iliti
es
Impo
rtan
ceof
visi
onre
lativ
eto
othe
rse
nses
unkn
own
Die
tP
ossi
bilit
yfo
rsu
b-po
pula
tion
diet
ary
spec
ializ
atio
nun
know
nE
colo
gic
alro
le
(i)E
ffect
ofco
nsum
ptio
nby
Gre
enla
ndsh
arks
onpr
eypo
pula
tions
unkn
own
(ii)R
ole
info
odw
ebst
abiliz
atio
nan
dco
nnec
tivity
unkn
own
(iii)
Rol
eof
long
evity
nutr
ient
ene
rgy
tran
sfer
info
odw
ebs
over
time
and
spac
eun
cert
ain
Met
abo
licra
te
(i)M
easu
reox
ygen
cons
umpt
ion
and
dete
rmin
em
etab
olic
rate
usin
gco
ntai
nmen
tapp
roac
hes
(ii)U
sem
etab
olic
rate
tode
term
ine
ener
getic
requ
irem
ents
pos
sibl
eex
trap
olat
ion
tone
cess
ary
frequ
ency
offe
edin
gus
ing
abi
oene
rget
ics
appr
oach
S
enso
ryab
iliti
es
Furt
her
exam
inat
ion
ofse
nsor
ysy
stem
san
dbr
ain
mor
phol
ogy
requ
ired
(eg
vi
asa
mpl
ing
ofex
perim
enta
land
or
com
mer
cial
byca
tch
mor
talit
ies)
D
iet
(i)C
olle
ctsa
mpl
etis
sues
(mus
cle
bloo
dpl
asm
a)by
non-
leth
alm
etho
dsto
exam
ine
diet
E
colo
gic
alro
le
(i)M
easu
rem
etab
olic
and
cons
umpt
ion
rate
sin
situ
(ii
)Fur
ther
refin
eA
rctic
mar
ine
food
web
stru
ctur
ean
dqu
antif
yen
ergy
flow
and
trop
hic
tran
sfer
effic
ienc
ies
(iii)
Use
Eco
Sim
mod
els
toex
amin
eth
epo
tent
ialt
roph
icef
fect
sof
Gre
enla
ndsh
ark
popu
latio
nin
crea
ses
and
decr
ease
s
Met
abo
licra
te
Indi
vidu
alen
ergy
requ
irem
ents
coul
din
form
popu
latio
n-le
velr
equi
rem
ents
(ie
bio
mas
sre
quire
dto
sust
ain
popu
latio
n)ndash
coul
dpr
ovid
eco
nsid
erat
ions
for
Tota
lAllo
wab
leC
atch
ofpr
eysp
ecie
sS
enso
ryab
iliti
es
Info
rmde
velo
pmen
tofs
hark
dete
rren
tsfo
rfis
hing
gear
mak
ege
arm
ore
notic
eabl
eto
shar
ks(e
g
ping
ers
scen
tel
ectr
onic
wav
es)
Eco
log
ical
role
(i)
Enh
ance
unde
rsta
ndin
gof
Gre
enla
ndsh
arks
and
thei
rec
olog
ical
role
byst
akeh
olde
rsan
dge
nera
lpub
lic
(ii)D
eter
min
esu
stai
nabl
eha
rves
tlev
els
whi
char
epr
edic
ted
tom
aint
ain
ecos
yste
mfu
nctio
n
Frontiers in Marine Science | wwwfrontiersinorg 6 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 7
Edwards et al Advancing Research for Long-Lived Species
TABLE 2 | Research goals and considerations for advancing the study of long-lived species
Research goal Potential methods and considerations for long-lived species
Visual identification of individuals fordemographic studies
Scar patterns and external markings may be used to identify individuals over short time periods (Devine et al 2018)but these may also change or disappear with age (Robbins and Fox 2012) External markers (eg Floy tags) facilitateidentification (Section ldquoDemographics and Life Historyrdquo) however studies should determine the average duration of tagretention by target species
Determining population conservationparameters
While genetic markers allow estimation of effective population sizes (Ne) the application of this approach forconservation purposes requires generation time and a species-specific mutation rate to be accurately defined (seesection ldquoPopulation Genetics and Genomicsrdquo) For species that cannot easily be aged it is difficult to accuratelydetermine generation times and mutation rates The use of evolutionarily significant units (sensu Fraser and Bernatchez2001) should be explored as an alternative
Monitoring behavior over time For species whose movements cover large distances over extended time periods it is important that the duration ofmonitoring (limited by tag lifespan and duration of retention) reflects the temporal scale of the observed behaviorsImprovements to electronic tag design (eg increased battery life) or the use of multi-tag approaches (Hussey et al2018) will extend monitoring durations and can record behaviors occurring over extended timespans (see sectionldquoMovement Ecologyrdquo)
Understanding ontogenetic shifts insensory capabilities and behavior
In fishes ontogenetic changes in the brain and neuroanatomy may affect the relative importance of senses such asvision and olfaction and may in turn be associated with age-related alterations in diet habitat use and activity patterns(Lisney et al 2017) Examinations of brain morphology and behavioral studies should target animals from a broadrange of age-classes to understand the influence of ontogeny on shifts in the ecological role of individuals over centuriesof longevity
Determining the physiology of extremelongevity
Measurements of metabolic rate (ie using oxygen consumption as proxy) blood oxygen consumption and thecardiovascular system (see section ldquoPhysiological Adaptationsrdquo) could provide insight into the physiological mechanismsof longevity
Examining trophic relationships andconnectivity
Extreme longevity may play a role in the ability of mobile predators to connect and stabilize food webs (eg byextending the number of migrations and feeding events that can occur over an animalrsquos lifespan) Similarly anunderstanding of the ecological footprints of long-lived individuals may improve the accuracy of predictive ecosystemmodels (eg EcoSim)
the examined and proposed research attempts were made tominimize this
Demographics and Life HistoryRecent radiocarbon dating of Greenland shark eye lensessupports the long-standing prediction of exceptional longevityin this species (Nielsen et al 2016 Figure 2A) These data alsoprovide the first age-at-length data for Greenland sharks
Current knowledge of Greenland shark distribution andpopulation abundance are based on exploratory fisheries surveysstock assessments of commercial species (eg Greenland halibutReinhardtius hippoglossoides and Northern shrimp Pandalusborealis) and bycatch reports from commercial and historicalfisheries Contemporary catch is restricted to small-scale targetedfisheries in Greenland and Iceland as well as incidental bycatchin a variety of North Atlantic and Arctic fisheries (ICES 2017)In the Barents Sea annual Greenland shark bycatch is estimatedto be 140ndash150 tons (Rusyaev and Orlov 2013) and in northernCanada 105 tons between 1996 and 2015 (Department ofFisheries and Oceans [DFO] 2016) However annual bycatchis likely to be much higher with an estimated bycatch of 1000tonsyr in the Uummannaq district in northwestern Greenlandalone (Gunnarsdottir and Joslashrgensen 2008 ICES 2017)
Scientific catch data suggest regional differences in the relativeabundance of Greenland sharks by life stagesize and sex Forexample no adult females [gt4 m total length (LT) Yanoet al 2007] have been reported throughout the Canadian Arcticand in Svalbard (Norway) despite intense sampling (N gt 300individuals Skomal and Benz 2004 Fisk et al 2002 2012Leclerc et al 2012 Hussey et al 2015a 2018 Devine et al 2018)
In contrast adult females appear to be more frequent offsouthwestern Greenland Iceland and Newfoundland (Canada)(Yano et al 2007 McMeans et al 2010 Nielsen et al 20142016 Campana et al 2015a) and several females gt 5 mhave been measured off Atlantic Canada (Campana personalcommunication) Juvenile sharks defined as individuals le200 cmLT have been observed in both inshore and offshore waters(Hussey et al 2015a) including animals within the birth sizerange (40ndash100 cm MacNeil et al 2012) Specifically juvenileswere observed in longline trawl and camera surveys within ScottInlet Baffin Island and in offshore waters gt1000 m depth (Fisket al 2002 Yano et al 2007 Nielsen et al 2014 Hussey et al2015a Devine et al 2018)
Baited remote underwater video (BRUV) surveys haveprovided the first estimates of the relative abundance ofGreenland sharks in the Canadian Arctic (independent ofbycatch data Devine et al 2018 Figure 2C) By identifyingindividuals using scar patterns and coloration the number ofsharks observed in each camera deployment was quantifiedallowing estimations of observation rate across sampled regions(Devine et al 2018 Figure 2D) BRUV surveys and exploratoryfisheries in the Canadian Arctic Archipelago have also expandedthe speciesrsquo known northern and western ranges in inshoreCanadian waters highlighting the extent to which their range washistorically defined by the spatial extent of commercial fisheries
While radiocarbon dating of eye lenses has provided baselineage estimates for this species these estimates still need tobe verified with an independent accurate and precise agedetermination technique However the absence of hard tissuescontaining growth bands (such as fin spines calcified vertebra
Frontiers in Marine Science | wwwfrontiersinorg 7 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 2
Edwards et al Advancing Research for Long-Lived Species
and function will advance our understanding of the causes and consequences of longlifespans Determining the extent and effects of current threats (as well as potentialmitigation measures) will assist the development of policies recommendations andactions relevant for the management of this potentially vulnerable species Through aninterdisciplinary lens we propose innovative approaches to direct the future study ofGreenland sharks and promote the consideration of longevity as an important factor inresearch on aquatic and terrestrial predators
Keywords future directions longevity management Somniosus microcephalus Arctic ecosystem
INTRODUCTION
The classification of life history traits along a continuum betweenr- and K-selection is regarded as a foundational tool for predictingthe ability of animal populations to effectively respond toenvironmental andor anthropogenic disturbance (Pianka 1970)In higher order vertebrates extreme longevity is commonlyassociated with the possession of K-selected life history traits thatinfluence maximum intrinsic population growth rates (rmax) ndashsuch as slow growth late maturity and relatively low recruitmentrates (McCann and Shuter 1997) Such traits are thought tolimit the ability of animal populations to sustain high levelsof mortality (Adams 1980 Musick 1999a Simpfendorfer andKyne 2009 Norse et al 2012) In chondrichthyans the frequencyof these K-selected traits becomes more pronounced withincreasing depth (Rigby and Simpfendorfer 2015) resultingin significantly lower rates of population increase in deep-water species and a heightened vulnerability to exploitation andincidental mortality relative to their shallow-water counterparts(Garciacutea et al 2008 Simpfendorfer and Kyne 2009) Manydeep-water species are currently targeted in commercial fisheries(Barker and Schluessel 2005) and the number of deep-waterhabitats affected by fishing is expected to increase (Halpern et al2008) In addition impacts from other human activities (egseismic surveys) on deep-sea biota are virtually unknown asare the chronic and cumulative impacts from multiple stressorsincluding climate change and pollution
The importance of longevity in influencing the vulnerability ofanimal populations to overexploitation has long been overlookedin conservation and management planning It is only relativelyrecently following well-known examples of overfishing (egNew Zealand orange roughly Hoplostethus atlanticus andNorth Atlantic cod Gadus morhua Walters and Maguire1996 Clark 2001) and declines in many long-lived marinespecies (Myers and Worm 2003 Heppell et al 2005) thatthere has beenx increased consideration of K-selected traits inmarine management policy (Musick 1999b Musick et al 2000Baum et al 2003) This is especially true for the protectionof species targeted by commercial or recreational fisheriesincluding some deep-water chondrichthyan and teleost fishes(Stevens et al 2000 Simpfendorfer and Kyne 2009 Norse et al2012 Rigby and Simpfendorfer 2015) While current literatureasserts the importance of longevity for predicting the effects ofanthropogenic disturbance on wild populations the mechanisms
driving extreme longevity and its influences on animal behaviorand population dynamics are not well understood
The Greenland shark (Somniosus microcephalus) is a newlyrecognized example of an extremely long-lived deep-waterchondrichthyan (estimated lifespan of at least 272 years Nielsenet al 2016) that is both data deficient and vulnerable to humanthreats such as fishery-related mortality (Davis et al 2013)Greenland sharks are primarily known to inhabit deep-waterand coastal regions of the Arctic and North Atlantic (Yanoet al 2007 see Figure 1 for capture locations from the reviewedstudies) but have the potential to range globally where deep-water temperatures are lt5C (MacNeil et al 2012) and havebeen observed both at the surface and at confirmed depthsup to 1816 m (Campana et al 2015a Somniosid spp havealso been observed at 2200 and 2992 m Herdendorf andBerra 1995 Porteiro et al 2017 respectively) While historicallyfished in the waters off Iceland Greenland and Norway tosupport a liver oil industry decreased demand in the mid 20thcentury prompted the closure of targeted fisheries currentlyGreenland sharks are primarily caught as bycatch in commerciallongline gillnet and bottom trawl fisheries (Davis et al 2013)Despite a recent spike in public interest due to its reportedlongevity logistical challenges continue to impede the detailedstudy of this species Consequently relatively little is knownabout several important aspects of Greenland shark physiology(eg metabolism reproduction) ecology (eg age class structuredistribution ecological role genetic variability) and behavior(eg predatory capabilities horizontal movement patterns)The primary objective of this paper is to highlight importantknowledge gaps that limit the development of a managementframework for the Greenland shark In the following text wediscuss a variety of forward-thinking experimental approachesthat will enable researchers to answer key questions aboutGreenland shark biology and ecology (Table 1) Additionally wepresent these current and future advances in Greenland sharkresearch within the context of broader issues regarding the studyof long-lived species (Table 2)
METHODS
Experts in the study of Greenland sharks from severalresearch and management institutions were consulted Guidedby the previous scientific contributions of this expert team
Frontiers in Marine Science | wwwfrontiersinorg 2 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 3
Edwards et al Advancing Research for Long-Lived Species
FIGURE 1 | Geographic coverage of Greenland shark studies by sampling location and research topic (A) locations of Greenland shark capturesampling (pointsrepresent sampling regions reported by each study multiple points used for studies with several disparate sampling sites) (B) study count by country of Greenlandshark capturesampling and research focus (studies counted more than once where multiple sampled countries are reported)
eight topics were deemed to be of high priority to assistfuture Greenland shark research and management The chosentopics are outlined in the text as follows (1) Demographicsand life history (2) Population genetics and genomics (3)Movement ecology (4) Behavior (5) Physiological adaptations(6) Diet and trophic ecology (7) Threats to Greenland sharkpopulations and (8) Management (see Figure 1 for study
count by research topic) Each topic was then organizedinto four key sections (i) current knowledge in the chosenfield (ii) identified knowledge gaps and limitations (iii)proposed strategies to address identified data deficiencies andfinally (iv) potential recommendations for Greenland sharkmanagement While some topic overlap among sections wasunavoidable due to the interconnected nature of much of
Frontiers in Marine Science | wwwfrontiersinorg 3 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 4
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|An
over
view
ofcu
rren
tkno
wle
dge
and
data
defic
ienc
ies
inei
ghtfi
elds
ofG
reen
land
shar
k(S
omni
osus
mic
roce
phal
us)r
esea
rch
stra
tegi
esto
addr
ess
data
defic
ienc
ies
and
pote
ntia
lact
ions
tosu
ppor
tm
anag
emen
torg
aniz
atio
nre
com
men
datio
nsan
dG
reen
land
shar
kco
nser
vatio
n
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Dem
ogra
phic
san
dlif
ehi
stor
yLi
fesp
anan
dag
ean
dsi
zeat
mat
urity
estim
ated
Lo
cala
bund
ance
sat
seve
rals
ites
inth
eC
anad
ian
Art
ices
timat
ed
Pot
entia
ljuv
enile
habi
tats
iden
tified
(Can
adia
nan
dR
ussi
anA
rctic
)
Sto
ckas
sess
men
tP
opul
atio
nsi
zes
grow
thra
tes
mor
talit
yra
tes
sex
ratio
san
dsu
stai
nabl
eha
rves
tlev
els
unkn
own
Are
adily
appl
icab
lean
dva
lidat
edag
ede
term
inat
ion
met
hod
nota
vaila
ble
Rep
rod
ucti
on
Rep
rodu
ctiv
ebi
olog
yan
dec
olog
yla
rgel
yun
know
n
Sto
ckas
sess
men
t(i)
Dev
elop
rout
ine
agin
gm
etho
d(ii
)Agi
ngof
indi
vidu
als
tode
term
ine
age-
at-le
ngth
and
info
rmpo
pula
tion
age
stru
ctur
e(ii
i)U
seB
RU
Vs
surv
eys
tobu
ilda
time
serie
son
whi
chto
estim
ate
biom
ass
and
abun
danc
ein
dice
s(v
iaa
rand
omsa
mpl
ing
desi
gna
cons
iste
ntan
nual
sam
plin
gre
gim
ean
da
broa
dne
twor
kof
sam
plin
gsi
tes)
(iv
)Req
uest
deta
iled
repo
rtin
gof
biol
ogic
alm
etric
s(e
g
leng
ths
exa
nddi
spos
ition
)for
byca
ught
shar
ksby
fishe
ries
obse
rver
sR
epro
duc
tio
n(i)
Use
ultr
asou
ndte
chni
ques
toid
entif
ypr
egna
ntfe
mal
es
(ii)E
lect
roni
cta
gssu
ture
din
tocl
oaca
ofpr
egna
ntfe
mal
esco
uld
help
dete
rmin
ebi
rthi
ngtim
ean
dlo
catio
n(a
spe
rBin
der
etal
20
14)
Sto
ckas
sess
men
t(i)
Dev
elop
Har
vest
Str
ateg
yin
clud
ing
Har
vest
Con
trol
Rul
es(e
g
ICE
SA
ppro
ach
toD
ata-
Lim
ited
Sto
cks)
(ii
)Pla
cevi
deo
mon
itorin
gon
fishi
ngve
ssel
sto
reco
rdpr
esen
cea
bsen
ceof
clas
pers
toes
timat
ese
xra
tio
(iii)
Com
pare
shar
kle
ngth
mea
sure
men
tsto
age-
at-le
ngth
toes
timat
eag
ecl
ass
abun
danc
ere
lativ
eca
ptur
era
tes
Rep
rod
ucti
on
Pro
tect
impo
rtan
tmat
ing
nurs
ery
area
san
dag
greg
atio
nsi
tes
(upo
nid
entifi
catio
n)
Pop
ulat
ion
gene
tics
and
geno
mic
sM
itoch
ondr
ialg
enom
eha
sbe
ense
quen
ced
Spe
cies
phyl
ogen
yha
sbe
ende
term
ined
E
volu
tiona
ryhi
stor
yha
sbe
enth
eoriz
ed
Loca
tion
ofhy
brid
izat
ion
with
Som
nios
uspa
cific
usun
know
nG
ener
atio
ntim
esan
dge
netic
varia
bilit
yam
ong
indi
vidu
als
and
popu
latio
nsun
know
nG
enet
ical
lyde
term
ined
susc
eptib
ility
toen
viro
nmen
talc
hang
eun
clea
r
Dat
aco
llect
ion
Req
uest
at-s
eaob
serv
ers
toco
llect
non-
leth
altis
sue
orpa
rasi
te(iD
NA
)sa
mpl
es
Defi
nep
op
ulat
ions
(i)
Use
gene
tics
toes
timat
ege
nera
tion
times
and
effe
ctiv
epo
pula
tion
size
(eg
us
ing
half
sibl
ing
appr
oach
)(ii
)Defi
nege
netic
ally
dist
inct
popu
latio
nss
tock
sD
efine
rang
eE
xam
inat
ion
ofhy
brid
izat
ion
inso
uthe
rnre
gion
s(e
g
Gul
fofM
exic
o)co
uld
exte
ndth
ekn
own
rang
eof
Gre
enla
ndsh
arks
beyo
ndth
epo
lar
regi
ons
Defi
nep
op
ulat
ions
and
rang
eR
ecom
men
dap
prop
riate
size
and
loca
tion
ofm
anag
emen
tbou
ndar
ies
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 4 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 5
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Mov
emen
teco
logy
Sea
sona
lmo
vem
ents
(i)
Hor
izon
talt
rack
ing
show
slo
ng-d
ista
nce
timed
dire
cted
mig
ratio
ns
(ii)O
bser
ved
occu
rren
cein
shal
low
erw
ater
sw
ithin
crea
sing
latit
ude
(iii)
Vis
ualr
ecor
dsof
philo
patr
yob
serv
edin
St
Law
renc
eE
stua
ry(G
alla
ntpe
rson
alco
mm
unic
atio
n)
Vert
ical
mo
vem
ents
(i)
Rec
ords
ofm
ovem
enti
nsh
allo
ww
ater
and
mid
wat
erco
lum
nun
der
ice
and
ines
tuar
ine
wat
ers
and
(aco
ustic
and
pop-
offs
atel
lite
trac
king
)(ii
)Ver
tical
mov
emen
tsap
pear
tobe
asso
ciat
edw
ithlig
htcy
cle
insh
allo
ww
ater
Sea
sona
lmo
vem
ents
(i)
Lack
oflo
ng-t
erm
(ie
mul
ti-ye
ar)
reco
rds
show
ing
seas
onal
ann
ual
mov
emen
ts
Vert
ical
mo
vem
ents
Li
mite
dre
cord
sof
deta
iled
vert
ical
mov
emen
tbeh
avio
rsav
aila
ble
(eg
ea
rlyde
pth
and
tem
pera
ture
reco
rds
binn
edat
60sndash
6h
inte
rval
s)
Dis
trib
utio
nS
hort
-ter
mm
onito
ring
insu
ffici
entt
oca
ptur
eco
mpl
ete
rang
eof
mov
emen
ts
Sea
sona
lmo
vem
ents
In
crea
sem
onito
ring
dura
tion
usin
glo
ng-t
erm
pass
ive
acou
stic
arc
hiva
lor
sate
llite
met
hods
and
impr
oved
tech
nolo
gies
(eg
ex
tend
edba
tter
ylif
ede
pth
ratin
gse
tc)
Vert
ical
mo
vem
ents
(i)
Use
high
-res
olut
ion
time-
serie
sda
tato
crea
teco
ntin
uous
mov
emen
ttra
cks
(ii)C
reat
est
ate-
spac
em
odel
sto
deco
mpo
sedi
ves
and
asso
ciat
eos
cilla
tory
patt
erns
with
disc
rete
beha
vior
alty
pes
(eg
po
st-r
elea
sere
cove
ry)
(iii)
Com
pare
over
lap
inve
rtic
aldi
strib
utio
nw
ithth
atof
fishe
ries
targ
etsp
ecie
sD
istr
ibut
ion
Use
larg
e-sc
ale
pass
ive
acou
stic
infra
stru
ctur
ean
dlo
ng-t
erm
sate
llite
tele
met
ryap
proa
ches
tode
fine
rang
ean
did
entif
ym
atin
gpu
ppin
ggr
ound
s
Sea
sona
lmo
vem
ents
(i)
Pot
entia
lsea
sona
lclo
sure
sfo
rhu
man
activ
ities
inan
nual
high
use
area
sto
min
imiz
ene
gativ
eim
pact
sto
Gre
enla
ndsh
ark
popu
latio
ns
(ii)D
ynam
icA
rea
Man
agem
entt
opr
otec
tun
expe
cted
aggr
egat
ions
Ve
rtic
alm
ove
men
ts
Info
rmfis
hha
rves
ters
ofsh
ark
beha
vior
patt
erns
tofa
cilit
ate
effo
rts
tom
inim
ize
shar
kca
tch
(eg
se
tgea
rat
dept
hsan
dtim
esof
day
whe
nsh
arks
tend
not
tobe
pres
ent)
Dis
trib
utio
nB
ilate
ralo
rm
ulti-
late
rala
gree
men
tsbe
twee
nna
tions
orvi
aR
egio
nal
Fish
erie
sM
anag
emen
tOrg
aniz
atio
ns(R
FMO
s)to
cond
ucts
cien
cean
dm
anag
esp
ecie
son
apo
pula
tion
leve
l
Beh
avio
ran
din
tera
ctio
nsC
aptu
rean
dre
leas
eP
relim
inar
yta
ggin
gst
udie
sha
veex
amin
edpo
st-r
elea
sebe
havi
oran
dsu
rviv
alra
tes
follo
win
gex
perim
enta
lca
ptur
eFo
rag
ing
beh
avio
r(i)
Indi
rect
evid
ence
ofliv
epr
edat
ion
onse
als
(ii)S
low
swim
spee
dsre
lativ
eto
body
size
buth
igh
roll
acce
lera
tion
(iii)
Dire
ctob
serv
atio
nsof
scav
engi
ng
(iv)A
ggre
gatio
nbe
havi
orob
serv
ed
Cap
ture
and
rele
ase
Pos
t-re
leas
esu
rviv
alra
tes
requ
irefu
rthe
rst
udy
(cur
rent
lyin
suffi
cien
tfor
adju
stin
gm
orta
lity
fact
ors
ina
popu
latio
nm
odel
)Fo
rag
ing
beh
avio
r(i)
Tact
ics
ofpr
eyca
ptur
eun
know
n(a
cryp
ticpr
edat
ion
appr
oach
hypo
thes
ized
)(ii
)Nat
ure
and
frequ
ency
ofin
tra-
and
inte
rspe
cific
inte
ract
ions
unkn
own
Fora
gin
gb
ehav
ior
(i)U
seca
mer
as(s
tilla
ndvi
deo)
depl
oyed
onva
rious
plat
form
s(e
g
anim
als
Rem
ote
Ope
rate
dVe
hicl
esfi
shin
gge
arc
able
s)to
obse
rve
fora
ging
beha
vior
dire
ctly
(ii)U
sean
imal
-bor
nehy
drop
hone
s(s
ampl
eso
unds
cape
)to
asse
ssin
tera
ctio
nsw
ithvo
caliz
ing
anim
als
(eg
m
arin
em
amm
als)
(ii
i)U
seof
VM
Ts(a
cous
tictr
ansp
onde
rs)t
oex
amin
ein
tra-
and
inte
r-sp
ecifi
cin
tera
ctio
ns
Cap
ture
and
rele
ase
Dev
elop
and
prom
ote
safe
hand
ling
prac
tices
for
the
rele
ase
ofsh
arks
caug
htas
byca
tch
Fora
gin
gb
ehav
ior
Use
unde
rsta
ndin
gof
feed
ing
mod
es(e
g
suct
ion
feed
ing)
and
pref
eren
ces
toin
form
gear
mod
ifica
tions
gea
rde
ploy
men
tan
dba
itus
e
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 5 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 6
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Phy
siol
ogic
alad
apta
tions
Die
tand
trop
hic
ecol
ogy
Phy
sio
log
y(i)
Exh
ibit
extr
eme
long
evity
(est
imat
edbe
twee
n27
2an
d51
2ye
ars)
(ii
)Hig
han
tioxi
dant
enzy
me
leve
lsan
dlo
wbl
ood
O2
affin
ityre
cord
ed
Sen
sory
abili
ties
(i)
Olfa
ctor
yor
gans
high
lyde
velo
ped
cons
iste
ntw
ithbe
nthi
cpr
edat
ors
cave
nger
lifes
tyle
(ii
)Hig
hin
cide
nce
ofbl
indn
ess
due
toey
epa
rasi
tism
(exc
epti
nth
eS
tLa
wre
nce
Riv
er)
(iii)
Obs
erva
tions
ofvi
sion
-dep
enda
ntbe
havi
ors
sugg
esta
bilit
yto
unde
rtak
eac
tive
pred
atio
nD
iet
Ben
thic
and
pela
gic
feed
ing
from
pred
omin
antly
offs
hore
tele
osts
ourc
essu
gges
ted
Eco
log
ical
role
G
reen
land
shar
ksar
elik
ely
impo
rtan
tpr
edat
ors
and
likel
yoc
cupy
ahi
ghtr
ophi
cpo
sitio
n
Phy
sio
log
y(i)
Max
imum
rese
rve
dens
itysu
gges
tslif
espa
nm
aycu
rren
tlybe
unde
rest
imat
ed
(ii)S
ensi
tivity
toen
viro
nmen
tals
hifts
unkn
own
(iii)
Det
ails
ofhe
artf
unct
ion
unkn
own
(may
play
aro
lein
long
evity
)M
etab
olic
rate
(i)
Met
abol
icra
tean
dra
teof
O2
cons
umpt
ion
unkn
own
(ii)P
oten
tiall
ink
betw
een
hem
oglo
bin
prop
ertie
san
dco
ld-t
oler
ance
(ii
i)P
oten
tiall
inks
betw
een
high
antio
xida
nten
zym
ele
vels
and
cold
tole
ranc
ede
epdi
ving
beha
vior
and
long
evity
S
enso
ryab
iliti
es
Impo
rtan
ceof
visi
onre
lativ
eto
othe
rse
nses
unkn
own
Die
tP
ossi
bilit
yfo
rsu
b-po
pula
tion
diet
ary
spec
ializ
atio
nun
know
nE
colo
gic
alro
le
(i)E
ffect
ofco
nsum
ptio
nby
Gre
enla
ndsh
arks
onpr
eypo
pula
tions
unkn
own
(ii)R
ole
info
odw
ebst
abiliz
atio
nan
dco
nnec
tivity
unkn
own
(iii)
Rol
eof
long
evity
nutr
ient
ene
rgy
tran
sfer
info
odw
ebs
over
time
and
spac
eun
cert
ain
Met
abo
licra
te
(i)M
easu
reox
ygen
cons
umpt
ion
and
dete
rmin
em
etab
olic
rate
usin
gco
ntai
nmen
tapp
roac
hes
(ii)U
sem
etab
olic
rate
tode
term
ine
ener
getic
requ
irem
ents
pos
sibl
eex
trap
olat
ion
tone
cess
ary
frequ
ency
offe
edin
gus
ing
abi
oene
rget
ics
appr
oach
S
enso
ryab
iliti
es
Furt
her
exam
inat
ion
ofse
nsor
ysy
stem
san
dbr
ain
mor
phol
ogy
requ
ired
(eg
vi
asa
mpl
ing
ofex
perim
enta
land
or
com
mer
cial
byca
tch
mor
talit
ies)
D
iet
(i)C
olle
ctsa
mpl
etis
sues
(mus
cle
bloo
dpl
asm
a)by
non-
leth
alm
etho
dsto
exam
ine
diet
E
colo
gic
alro
le
(i)M
easu
rem
etab
olic
and
cons
umpt
ion
rate
sin
situ
(ii
)Fur
ther
refin
eA
rctic
mar
ine
food
web
stru
ctur
ean
dqu
antif
yen
ergy
flow
and
trop
hic
tran
sfer
effic
ienc
ies
(iii)
Use
Eco
Sim
mod
els
toex
amin
eth
epo
tent
ialt
roph
icef
fect
sof
Gre
enla
ndsh
ark
popu
latio
nin
crea
ses
and
decr
ease
s
Met
abo
licra
te
Indi
vidu
alen
ergy
requ
irem
ents
coul
din
form
popu
latio
n-le
velr
equi
rem
ents
(ie
bio
mas
sre
quire
dto
sust
ain
popu
latio
n)ndash
coul
dpr
ovid
eco
nsid
erat
ions
for
Tota
lAllo
wab
leC
atch
ofpr
eysp
ecie
sS
enso
ryab
iliti
es
Info
rmde
velo
pmen
tofs
hark
dete
rren
tsfo
rfis
hing
gear
mak
ege
arm
ore
notic
eabl
eto
shar
ks(e
g
ping
ers
scen
tel
ectr
onic
wav
es)
Eco
log
ical
role
(i)
Enh
ance
unde
rsta
ndin
gof
Gre
enla
ndsh
arks
and
thei
rec
olog
ical
role
byst
akeh
olde
rsan
dge
nera
lpub
lic
(ii)D
eter
min
esu
stai
nabl
eha
rves
tlev
els
whi
char
epr
edic
ted
tom
aint
ain
ecos
yste
mfu
nctio
n
Frontiers in Marine Science | wwwfrontiersinorg 6 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 7
Edwards et al Advancing Research for Long-Lived Species
TABLE 2 | Research goals and considerations for advancing the study of long-lived species
Research goal Potential methods and considerations for long-lived species
Visual identification of individuals fordemographic studies
Scar patterns and external markings may be used to identify individuals over short time periods (Devine et al 2018)but these may also change or disappear with age (Robbins and Fox 2012) External markers (eg Floy tags) facilitateidentification (Section ldquoDemographics and Life Historyrdquo) however studies should determine the average duration of tagretention by target species
Determining population conservationparameters
While genetic markers allow estimation of effective population sizes (Ne) the application of this approach forconservation purposes requires generation time and a species-specific mutation rate to be accurately defined (seesection ldquoPopulation Genetics and Genomicsrdquo) For species that cannot easily be aged it is difficult to accuratelydetermine generation times and mutation rates The use of evolutionarily significant units (sensu Fraser and Bernatchez2001) should be explored as an alternative
Monitoring behavior over time For species whose movements cover large distances over extended time periods it is important that the duration ofmonitoring (limited by tag lifespan and duration of retention) reflects the temporal scale of the observed behaviorsImprovements to electronic tag design (eg increased battery life) or the use of multi-tag approaches (Hussey et al2018) will extend monitoring durations and can record behaviors occurring over extended timespans (see sectionldquoMovement Ecologyrdquo)
Understanding ontogenetic shifts insensory capabilities and behavior
In fishes ontogenetic changes in the brain and neuroanatomy may affect the relative importance of senses such asvision and olfaction and may in turn be associated with age-related alterations in diet habitat use and activity patterns(Lisney et al 2017) Examinations of brain morphology and behavioral studies should target animals from a broadrange of age-classes to understand the influence of ontogeny on shifts in the ecological role of individuals over centuriesof longevity
Determining the physiology of extremelongevity
Measurements of metabolic rate (ie using oxygen consumption as proxy) blood oxygen consumption and thecardiovascular system (see section ldquoPhysiological Adaptationsrdquo) could provide insight into the physiological mechanismsof longevity
Examining trophic relationships andconnectivity
Extreme longevity may play a role in the ability of mobile predators to connect and stabilize food webs (eg byextending the number of migrations and feeding events that can occur over an animalrsquos lifespan) Similarly anunderstanding of the ecological footprints of long-lived individuals may improve the accuracy of predictive ecosystemmodels (eg EcoSim)
the examined and proposed research attempts were made tominimize this
Demographics and Life HistoryRecent radiocarbon dating of Greenland shark eye lensessupports the long-standing prediction of exceptional longevityin this species (Nielsen et al 2016 Figure 2A) These data alsoprovide the first age-at-length data for Greenland sharks
Current knowledge of Greenland shark distribution andpopulation abundance are based on exploratory fisheries surveysstock assessments of commercial species (eg Greenland halibutReinhardtius hippoglossoides and Northern shrimp Pandalusborealis) and bycatch reports from commercial and historicalfisheries Contemporary catch is restricted to small-scale targetedfisheries in Greenland and Iceland as well as incidental bycatchin a variety of North Atlantic and Arctic fisheries (ICES 2017)In the Barents Sea annual Greenland shark bycatch is estimatedto be 140ndash150 tons (Rusyaev and Orlov 2013) and in northernCanada 105 tons between 1996 and 2015 (Department ofFisheries and Oceans [DFO] 2016) However annual bycatchis likely to be much higher with an estimated bycatch of 1000tonsyr in the Uummannaq district in northwestern Greenlandalone (Gunnarsdottir and Joslashrgensen 2008 ICES 2017)
Scientific catch data suggest regional differences in the relativeabundance of Greenland sharks by life stagesize and sex Forexample no adult females [gt4 m total length (LT) Yanoet al 2007] have been reported throughout the Canadian Arcticand in Svalbard (Norway) despite intense sampling (N gt 300individuals Skomal and Benz 2004 Fisk et al 2002 2012Leclerc et al 2012 Hussey et al 2015a 2018 Devine et al 2018)
In contrast adult females appear to be more frequent offsouthwestern Greenland Iceland and Newfoundland (Canada)(Yano et al 2007 McMeans et al 2010 Nielsen et al 20142016 Campana et al 2015a) and several females gt 5 mhave been measured off Atlantic Canada (Campana personalcommunication) Juvenile sharks defined as individuals le200 cmLT have been observed in both inshore and offshore waters(Hussey et al 2015a) including animals within the birth sizerange (40ndash100 cm MacNeil et al 2012) Specifically juvenileswere observed in longline trawl and camera surveys within ScottInlet Baffin Island and in offshore waters gt1000 m depth (Fisket al 2002 Yano et al 2007 Nielsen et al 2014 Hussey et al2015a Devine et al 2018)
Baited remote underwater video (BRUV) surveys haveprovided the first estimates of the relative abundance ofGreenland sharks in the Canadian Arctic (independent ofbycatch data Devine et al 2018 Figure 2C) By identifyingindividuals using scar patterns and coloration the number ofsharks observed in each camera deployment was quantifiedallowing estimations of observation rate across sampled regions(Devine et al 2018 Figure 2D) BRUV surveys and exploratoryfisheries in the Canadian Arctic Archipelago have also expandedthe speciesrsquo known northern and western ranges in inshoreCanadian waters highlighting the extent to which their range washistorically defined by the spatial extent of commercial fisheries
While radiocarbon dating of eye lenses has provided baselineage estimates for this species these estimates still need tobe verified with an independent accurate and precise agedetermination technique However the absence of hard tissuescontaining growth bands (such as fin spines calcified vertebra
Frontiers in Marine Science | wwwfrontiersinorg 7 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 3
Edwards et al Advancing Research for Long-Lived Species
FIGURE 1 | Geographic coverage of Greenland shark studies by sampling location and research topic (A) locations of Greenland shark capturesampling (pointsrepresent sampling regions reported by each study multiple points used for studies with several disparate sampling sites) (B) study count by country of Greenlandshark capturesampling and research focus (studies counted more than once where multiple sampled countries are reported)
eight topics were deemed to be of high priority to assistfuture Greenland shark research and management The chosentopics are outlined in the text as follows (1) Demographicsand life history (2) Population genetics and genomics (3)Movement ecology (4) Behavior (5) Physiological adaptations(6) Diet and trophic ecology (7) Threats to Greenland sharkpopulations and (8) Management (see Figure 1 for study
count by research topic) Each topic was then organizedinto four key sections (i) current knowledge in the chosenfield (ii) identified knowledge gaps and limitations (iii)proposed strategies to address identified data deficiencies andfinally (iv) potential recommendations for Greenland sharkmanagement While some topic overlap among sections wasunavoidable due to the interconnected nature of much of
Frontiers in Marine Science | wwwfrontiersinorg 3 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 4
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|An
over
view
ofcu
rren
tkno
wle
dge
and
data
defic
ienc
ies
inei
ghtfi
elds
ofG
reen
land
shar
k(S
omni
osus
mic
roce
phal
us)r
esea
rch
stra
tegi
esto
addr
ess
data
defic
ienc
ies
and
pote
ntia
lact
ions
tosu
ppor
tm
anag
emen
torg
aniz
atio
nre
com
men
datio
nsan
dG
reen
land
shar
kco
nser
vatio
n
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Dem
ogra
phic
san
dlif
ehi
stor
yLi
fesp
anan
dag
ean
dsi
zeat
mat
urity
estim
ated
Lo
cala
bund
ance
sat
seve
rals
ites
inth
eC
anad
ian
Art
ices
timat
ed
Pot
entia
ljuv
enile
habi
tats
iden
tified
(Can
adia
nan
dR
ussi
anA
rctic
)
Sto
ckas
sess
men
tP
opul
atio
nsi
zes
grow
thra
tes
mor
talit
yra
tes
sex
ratio
san
dsu
stai
nabl
eha
rves
tlev
els
unkn
own
Are
adily
appl
icab
lean
dva
lidat
edag
ede
term
inat
ion
met
hod
nota
vaila
ble
Rep
rod
ucti
on
Rep
rodu
ctiv
ebi
olog
yan
dec
olog
yla
rgel
yun
know
n
Sto
ckas
sess
men
t(i)
Dev
elop
rout
ine
agin
gm
etho
d(ii
)Agi
ngof
indi
vidu
als
tode
term
ine
age-
at-le
ngth
and
info
rmpo
pula
tion
age
stru
ctur
e(ii
i)U
seB
RU
Vs
surv
eys
tobu
ilda
time
serie
son
whi
chto
estim
ate
biom
ass
and
abun
danc
ein
dice
s(v
iaa
rand
omsa
mpl
ing
desi
gna
cons
iste
ntan
nual
sam
plin
gre
gim
ean
da
broa
dne
twor
kof
sam
plin
gsi
tes)
(iv
)Req
uest
deta
iled
repo
rtin
gof
biol
ogic
alm
etric
s(e
g
leng
ths
exa
nddi
spos
ition
)for
byca
ught
shar
ksby
fishe
ries
obse
rver
sR
epro
duc
tio
n(i)
Use
ultr
asou
ndte
chni
ques
toid
entif
ypr
egna
ntfe
mal
es
(ii)E
lect
roni
cta
gssu
ture
din
tocl
oaca
ofpr
egna
ntfe
mal
esco
uld
help
dete
rmin
ebi
rthi
ngtim
ean
dlo
catio
n(a
spe
rBin
der
etal
20
14)
Sto
ckas
sess
men
t(i)
Dev
elop
Har
vest
Str
ateg
yin
clud
ing
Har
vest
Con
trol
Rul
es(e
g
ICE
SA
ppro
ach
toD
ata-
Lim
ited
Sto
cks)
(ii
)Pla
cevi
deo
mon
itorin
gon
fishi
ngve
ssel
sto
reco
rdpr
esen
cea
bsen
ceof
clas
pers
toes
timat
ese
xra
tio
(iii)
Com
pare
shar
kle
ngth
mea
sure
men
tsto
age-
at-le
ngth
toes
timat
eag
ecl
ass
abun
danc
ere
lativ
eca
ptur
era
tes
Rep
rod
ucti
on
Pro
tect
impo
rtan
tmat
ing
nurs
ery
area
san
dag
greg
atio
nsi
tes
(upo
nid
entifi
catio
n)
Pop
ulat
ion
gene
tics
and
geno
mic
sM
itoch
ondr
ialg
enom
eha
sbe
ense
quen
ced
Spe
cies
phyl
ogen
yha
sbe
ende
term
ined
E
volu
tiona
ryhi
stor
yha
sbe
enth
eoriz
ed
Loca
tion
ofhy
brid
izat
ion
with
Som
nios
uspa
cific
usun
know
nG
ener
atio
ntim
esan
dge
netic
varia
bilit
yam
ong
indi
vidu
als
and
popu
latio
nsun
know
nG
enet
ical
lyde
term
ined
susc
eptib
ility
toen
viro
nmen
talc
hang
eun
clea
r
Dat
aco
llect
ion
Req
uest
at-s
eaob
serv
ers
toco
llect
non-
leth
altis
sue
orpa
rasi
te(iD
NA
)sa
mpl
es
Defi
nep
op
ulat
ions
(i)
Use
gene
tics
toes
timat
ege
nera
tion
times
and
effe
ctiv
epo
pula
tion
size
(eg
us
ing
half
sibl
ing
appr
oach
)(ii
)Defi
nege
netic
ally
dist
inct
popu
latio
nss
tock
sD
efine
rang
eE
xam
inat
ion
ofhy
brid
izat
ion
inso
uthe
rnre
gion
s(e
g
Gul
fofM
exic
o)co
uld
exte
ndth
ekn
own
rang
eof
Gre
enla
ndsh
arks
beyo
ndth
epo
lar
regi
ons
Defi
nep
op
ulat
ions
and
rang
eR
ecom
men
dap
prop
riate
size
and
loca
tion
ofm
anag
emen
tbou
ndar
ies
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 4 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 5
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Mov
emen
teco
logy
Sea
sona
lmo
vem
ents
(i)
Hor
izon
talt
rack
ing
show
slo
ng-d
ista
nce
timed
dire
cted
mig
ratio
ns
(ii)O
bser
ved
occu
rren
cein
shal
low
erw
ater
sw
ithin
crea
sing
latit
ude
(iii)
Vis
ualr
ecor
dsof
philo
patr
yob
serv
edin
St
Law
renc
eE
stua
ry(G
alla
ntpe
rson
alco
mm
unic
atio
n)
Vert
ical
mo
vem
ents
(i)
Rec
ords
ofm
ovem
enti
nsh
allo
ww
ater
and
mid
wat
erco
lum
nun
der
ice
and
ines
tuar
ine
wat
ers
and
(aco
ustic
and
pop-
offs
atel
lite
trac
king
)(ii
)Ver
tical
mov
emen
tsap
pear
tobe
asso
ciat
edw
ithlig
htcy
cle
insh
allo
ww
ater
Sea
sona
lmo
vem
ents
(i)
Lack
oflo
ng-t
erm
(ie
mul
ti-ye
ar)
reco
rds
show
ing
seas
onal
ann
ual
mov
emen
ts
Vert
ical
mo
vem
ents
Li
mite
dre
cord
sof
deta
iled
vert
ical
mov
emen
tbeh
avio
rsav
aila
ble
(eg
ea
rlyde
pth
and
tem
pera
ture
reco
rds
binn
edat
60sndash
6h
inte
rval
s)
Dis
trib
utio
nS
hort
-ter
mm
onito
ring
insu
ffici
entt
oca
ptur
eco
mpl
ete
rang
eof
mov
emen
ts
Sea
sona
lmo
vem
ents
In
crea
sem
onito
ring
dura
tion
usin
glo
ng-t
erm
pass
ive
acou
stic
arc
hiva
lor
sate
llite
met
hods
and
impr
oved
tech
nolo
gies
(eg
ex
tend
edba
tter
ylif
ede
pth
ratin
gse
tc)
Vert
ical
mo
vem
ents
(i)
Use
high
-res
olut
ion
time-
serie
sda
tato
crea
teco
ntin
uous
mov
emen
ttra
cks
(ii)C
reat
est
ate-
spac
em
odel
sto
deco
mpo
sedi
ves
and
asso
ciat
eos
cilla
tory
patt
erns
with
disc
rete
beha
vior
alty
pes
(eg
po
st-r
elea
sere
cove
ry)
(iii)
Com
pare
over
lap
inve
rtic
aldi
strib
utio
nw
ithth
atof
fishe
ries
targ
etsp
ecie
sD
istr
ibut
ion
Use
larg
e-sc
ale
pass
ive
acou
stic
infra
stru
ctur
ean
dlo
ng-t
erm
sate
llite
tele
met
ryap
proa
ches
tode
fine
rang
ean
did
entif
ym
atin
gpu
ppin
ggr
ound
s
Sea
sona
lmo
vem
ents
(i)
Pot
entia
lsea
sona
lclo
sure
sfo
rhu
man
activ
ities
inan
nual
high
use
area
sto
min
imiz
ene
gativ
eim
pact
sto
Gre
enla
ndsh
ark
popu
latio
ns
(ii)D
ynam
icA
rea
Man
agem
entt
opr
otec
tun
expe
cted
aggr
egat
ions
Ve
rtic
alm
ove
men
ts
Info
rmfis
hha
rves
ters
ofsh
ark
beha
vior
patt
erns
tofa
cilit
ate
effo
rts
tom
inim
ize
shar
kca
tch
(eg
se
tgea
rat
dept
hsan
dtim
esof
day
whe
nsh
arks
tend
not
tobe
pres
ent)
Dis
trib
utio
nB
ilate
ralo
rm
ulti-
late
rala
gree
men
tsbe
twee
nna
tions
orvi
aR
egio
nal
Fish
erie
sM
anag
emen
tOrg
aniz
atio
ns(R
FMO
s)to
cond
ucts
cien
cean
dm
anag
esp
ecie
son
apo
pula
tion
leve
l
Beh
avio
ran
din
tera
ctio
nsC
aptu
rean
dre
leas
eP
relim
inar
yta
ggin
gst
udie
sha
veex
amin
edpo
st-r
elea
sebe
havi
oran
dsu
rviv
alra
tes
follo
win
gex
perim
enta
lca
ptur
eFo
rag
ing
beh
avio
r(i)
Indi
rect
evid
ence
ofliv
epr
edat
ion
onse
als
(ii)S
low
swim
spee
dsre
lativ
eto
body
size
buth
igh
roll
acce
lera
tion
(iii)
Dire
ctob
serv
atio
nsof
scav
engi
ng
(iv)A
ggre
gatio
nbe
havi
orob
serv
ed
Cap
ture
and
rele
ase
Pos
t-re
leas
esu
rviv
alra
tes
requ
irefu
rthe
rst
udy
(cur
rent
lyin
suffi
cien
tfor
adju
stin
gm
orta
lity
fact
ors
ina
popu
latio
nm
odel
)Fo
rag
ing
beh
avio
r(i)
Tact
ics
ofpr
eyca
ptur
eun
know
n(a
cryp
ticpr
edat
ion
appr
oach
hypo
thes
ized
)(ii
)Nat
ure
and
frequ
ency
ofin
tra-
and
inte
rspe
cific
inte
ract
ions
unkn
own
Fora
gin
gb
ehav
ior
(i)U
seca
mer
as(s
tilla
ndvi
deo)
depl
oyed
onva
rious
plat
form
s(e
g
anim
als
Rem
ote
Ope
rate
dVe
hicl
esfi
shin
gge
arc
able
s)to
obse
rve
fora
ging
beha
vior
dire
ctly
(ii)U
sean
imal
-bor
nehy
drop
hone
s(s
ampl
eso
unds
cape
)to
asse
ssin
tera
ctio
nsw
ithvo
caliz
ing
anim
als
(eg
m
arin
em
amm
als)
(ii
i)U
seof
VM
Ts(a
cous
tictr
ansp
onde
rs)t
oex
amin
ein
tra-
and
inte
r-sp
ecifi
cin
tera
ctio
ns
Cap
ture
and
rele
ase
Dev
elop
and
prom
ote
safe
hand
ling
prac
tices
for
the
rele
ase
ofsh
arks
caug
htas
byca
tch
Fora
gin
gb
ehav
ior
Use
unde
rsta
ndin
gof
feed
ing
mod
es(e
g
suct
ion
feed
ing)
and
pref
eren
ces
toin
form
gear
mod
ifica
tions
gea
rde
ploy
men
tan
dba
itus
e
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 5 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 6
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Phy
siol
ogic
alad
apta
tions
Die
tand
trop
hic
ecol
ogy
Phy
sio
log
y(i)
Exh
ibit
extr
eme
long
evity
(est
imat
edbe
twee
n27
2an
d51
2ye
ars)
(ii
)Hig
han
tioxi
dant
enzy
me
leve
lsan
dlo
wbl
ood
O2
affin
ityre
cord
ed
Sen
sory
abili
ties
(i)
Olfa
ctor
yor
gans
high
lyde
velo
ped
cons
iste
ntw
ithbe
nthi
cpr
edat
ors
cave
nger
lifes
tyle
(ii
)Hig
hin
cide
nce
ofbl
indn
ess
due
toey
epa
rasi
tism
(exc
epti
nth
eS
tLa
wre
nce
Riv
er)
(iii)
Obs
erva
tions
ofvi
sion
-dep
enda
ntbe
havi
ors
sugg
esta
bilit
yto
unde
rtak
eac
tive
pred
atio
nD
iet
Ben
thic
and
pela
gic
feed
ing
from
pred
omin
antly
offs
hore
tele
osts
ourc
essu
gges
ted
Eco
log
ical
role
G
reen
land
shar
ksar
elik
ely
impo
rtan
tpr
edat
ors
and
likel
yoc
cupy
ahi
ghtr
ophi
cpo
sitio
n
Phy
sio
log
y(i)
Max
imum
rese
rve
dens
itysu
gges
tslif
espa
nm
aycu
rren
tlybe
unde
rest
imat
ed
(ii)S
ensi
tivity
toen
viro
nmen
tals
hifts
unkn
own
(iii)
Det
ails
ofhe
artf
unct
ion
unkn
own
(may
play
aro
lein
long
evity
)M
etab
olic
rate
(i)
Met
abol
icra
tean
dra
teof
O2
cons
umpt
ion
unkn
own
(ii)P
oten
tiall
ink
betw
een
hem
oglo
bin
prop
ertie
san
dco
ld-t
oler
ance
(ii
i)P
oten
tiall
inks
betw
een
high
antio
xida
nten
zym
ele
vels
and
cold
tole
ranc
ede
epdi
ving
beha
vior
and
long
evity
S
enso
ryab
iliti
es
Impo
rtan
ceof
visi
onre
lativ
eto
othe
rse
nses
unkn
own
Die
tP
ossi
bilit
yfo
rsu
b-po
pula
tion
diet
ary
spec
ializ
atio
nun
know
nE
colo
gic
alro
le
(i)E
ffect
ofco
nsum
ptio
nby
Gre
enla
ndsh
arks
onpr
eypo
pula
tions
unkn
own
(ii)R
ole
info
odw
ebst
abiliz
atio
nan
dco
nnec
tivity
unkn
own
(iii)
Rol
eof
long
evity
nutr
ient
ene
rgy
tran
sfer
info
odw
ebs
over
time
and
spac
eun
cert
ain
Met
abo
licra
te
(i)M
easu
reox
ygen
cons
umpt
ion
and
dete
rmin
em
etab
olic
rate
usin
gco
ntai
nmen
tapp
roac
hes
(ii)U
sem
etab
olic
rate
tode
term
ine
ener
getic
requ
irem
ents
pos
sibl
eex
trap
olat
ion
tone
cess
ary
frequ
ency
offe
edin
gus
ing
abi
oene
rget
ics
appr
oach
S
enso
ryab
iliti
es
Furt
her
exam
inat
ion
ofse
nsor
ysy
stem
san
dbr
ain
mor
phol
ogy
requ
ired
(eg
vi
asa
mpl
ing
ofex
perim
enta
land
or
com
mer
cial
byca
tch
mor
talit
ies)
D
iet
(i)C
olle
ctsa
mpl
etis
sues
(mus
cle
bloo
dpl
asm
a)by
non-
leth
alm
etho
dsto
exam
ine
diet
E
colo
gic
alro
le
(i)M
easu
rem
etab
olic
and
cons
umpt
ion
rate
sin
situ
(ii
)Fur
ther
refin
eA
rctic
mar
ine
food
web
stru
ctur
ean
dqu
antif
yen
ergy
flow
and
trop
hic
tran
sfer
effic
ienc
ies
(iii)
Use
Eco
Sim
mod
els
toex
amin
eth
epo
tent
ialt
roph
icef
fect
sof
Gre
enla
ndsh
ark
popu
latio
nin
crea
ses
and
decr
ease
s
Met
abo
licra
te
Indi
vidu
alen
ergy
requ
irem
ents
coul
din
form
popu
latio
n-le
velr
equi
rem
ents
(ie
bio
mas
sre
quire
dto
sust
ain
popu
latio
n)ndash
coul
dpr
ovid
eco
nsid
erat
ions
for
Tota
lAllo
wab
leC
atch
ofpr
eysp
ecie
sS
enso
ryab
iliti
es
Info
rmde
velo
pmen
tofs
hark
dete
rren
tsfo
rfis
hing
gear
mak
ege
arm
ore
notic
eabl
eto
shar
ks(e
g
ping
ers
scen
tel
ectr
onic
wav
es)
Eco
log
ical
role
(i)
Enh
ance
unde
rsta
ndin
gof
Gre
enla
ndsh
arks
and
thei
rec
olog
ical
role
byst
akeh
olde
rsan
dge
nera
lpub
lic
(ii)D
eter
min
esu
stai
nabl
eha
rves
tlev
els
whi
char
epr
edic
ted
tom
aint
ain
ecos
yste
mfu
nctio
n
Frontiers in Marine Science | wwwfrontiersinorg 6 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 7
Edwards et al Advancing Research for Long-Lived Species
TABLE 2 | Research goals and considerations for advancing the study of long-lived species
Research goal Potential methods and considerations for long-lived species
Visual identification of individuals fordemographic studies
Scar patterns and external markings may be used to identify individuals over short time periods (Devine et al 2018)but these may also change or disappear with age (Robbins and Fox 2012) External markers (eg Floy tags) facilitateidentification (Section ldquoDemographics and Life Historyrdquo) however studies should determine the average duration of tagretention by target species
Determining population conservationparameters
While genetic markers allow estimation of effective population sizes (Ne) the application of this approach forconservation purposes requires generation time and a species-specific mutation rate to be accurately defined (seesection ldquoPopulation Genetics and Genomicsrdquo) For species that cannot easily be aged it is difficult to accuratelydetermine generation times and mutation rates The use of evolutionarily significant units (sensu Fraser and Bernatchez2001) should be explored as an alternative
Monitoring behavior over time For species whose movements cover large distances over extended time periods it is important that the duration ofmonitoring (limited by tag lifespan and duration of retention) reflects the temporal scale of the observed behaviorsImprovements to electronic tag design (eg increased battery life) or the use of multi-tag approaches (Hussey et al2018) will extend monitoring durations and can record behaviors occurring over extended timespans (see sectionldquoMovement Ecologyrdquo)
Understanding ontogenetic shifts insensory capabilities and behavior
In fishes ontogenetic changes in the brain and neuroanatomy may affect the relative importance of senses such asvision and olfaction and may in turn be associated with age-related alterations in diet habitat use and activity patterns(Lisney et al 2017) Examinations of brain morphology and behavioral studies should target animals from a broadrange of age-classes to understand the influence of ontogeny on shifts in the ecological role of individuals over centuriesof longevity
Determining the physiology of extremelongevity
Measurements of metabolic rate (ie using oxygen consumption as proxy) blood oxygen consumption and thecardiovascular system (see section ldquoPhysiological Adaptationsrdquo) could provide insight into the physiological mechanismsof longevity
Examining trophic relationships andconnectivity
Extreme longevity may play a role in the ability of mobile predators to connect and stabilize food webs (eg byextending the number of migrations and feeding events that can occur over an animalrsquos lifespan) Similarly anunderstanding of the ecological footprints of long-lived individuals may improve the accuracy of predictive ecosystemmodels (eg EcoSim)
the examined and proposed research attempts were made tominimize this
Demographics and Life HistoryRecent radiocarbon dating of Greenland shark eye lensessupports the long-standing prediction of exceptional longevityin this species (Nielsen et al 2016 Figure 2A) These data alsoprovide the first age-at-length data for Greenland sharks
Current knowledge of Greenland shark distribution andpopulation abundance are based on exploratory fisheries surveysstock assessments of commercial species (eg Greenland halibutReinhardtius hippoglossoides and Northern shrimp Pandalusborealis) and bycatch reports from commercial and historicalfisheries Contemporary catch is restricted to small-scale targetedfisheries in Greenland and Iceland as well as incidental bycatchin a variety of North Atlantic and Arctic fisheries (ICES 2017)In the Barents Sea annual Greenland shark bycatch is estimatedto be 140ndash150 tons (Rusyaev and Orlov 2013) and in northernCanada 105 tons between 1996 and 2015 (Department ofFisheries and Oceans [DFO] 2016) However annual bycatchis likely to be much higher with an estimated bycatch of 1000tonsyr in the Uummannaq district in northwestern Greenlandalone (Gunnarsdottir and Joslashrgensen 2008 ICES 2017)
Scientific catch data suggest regional differences in the relativeabundance of Greenland sharks by life stagesize and sex Forexample no adult females [gt4 m total length (LT) Yanoet al 2007] have been reported throughout the Canadian Arcticand in Svalbard (Norway) despite intense sampling (N gt 300individuals Skomal and Benz 2004 Fisk et al 2002 2012Leclerc et al 2012 Hussey et al 2015a 2018 Devine et al 2018)
In contrast adult females appear to be more frequent offsouthwestern Greenland Iceland and Newfoundland (Canada)(Yano et al 2007 McMeans et al 2010 Nielsen et al 20142016 Campana et al 2015a) and several females gt 5 mhave been measured off Atlantic Canada (Campana personalcommunication) Juvenile sharks defined as individuals le200 cmLT have been observed in both inshore and offshore waters(Hussey et al 2015a) including animals within the birth sizerange (40ndash100 cm MacNeil et al 2012) Specifically juvenileswere observed in longline trawl and camera surveys within ScottInlet Baffin Island and in offshore waters gt1000 m depth (Fisket al 2002 Yano et al 2007 Nielsen et al 2014 Hussey et al2015a Devine et al 2018)
Baited remote underwater video (BRUV) surveys haveprovided the first estimates of the relative abundance ofGreenland sharks in the Canadian Arctic (independent ofbycatch data Devine et al 2018 Figure 2C) By identifyingindividuals using scar patterns and coloration the number ofsharks observed in each camera deployment was quantifiedallowing estimations of observation rate across sampled regions(Devine et al 2018 Figure 2D) BRUV surveys and exploratoryfisheries in the Canadian Arctic Archipelago have also expandedthe speciesrsquo known northern and western ranges in inshoreCanadian waters highlighting the extent to which their range washistorically defined by the spatial extent of commercial fisheries
While radiocarbon dating of eye lenses has provided baselineage estimates for this species these estimates still need tobe verified with an independent accurate and precise agedetermination technique However the absence of hard tissuescontaining growth bands (such as fin spines calcified vertebra
Frontiers in Marine Science | wwwfrontiersinorg 7 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 4
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|An
over
view
ofcu
rren
tkno
wle
dge
and
data
defic
ienc
ies
inei
ghtfi
elds
ofG
reen
land
shar
k(S
omni
osus
mic
roce
phal
us)r
esea
rch
stra
tegi
esto
addr
ess
data
defic
ienc
ies
and
pote
ntia
lact
ions
tosu
ppor
tm
anag
emen
torg
aniz
atio
nre
com
men
datio
nsan
dG
reen
land
shar
kco
nser
vatio
n
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Dem
ogra
phic
san
dlif
ehi
stor
yLi
fesp
anan
dag
ean
dsi
zeat
mat
urity
estim
ated
Lo
cala
bund
ance
sat
seve
rals
ites
inth
eC
anad
ian
Art
ices
timat
ed
Pot
entia
ljuv
enile
habi
tats
iden
tified
(Can
adia
nan
dR
ussi
anA
rctic
)
Sto
ckas
sess
men
tP
opul
atio
nsi
zes
grow
thra
tes
mor
talit
yra
tes
sex
ratio
san
dsu
stai
nabl
eha
rves
tlev
els
unkn
own
Are
adily
appl
icab
lean
dva
lidat
edag
ede
term
inat
ion
met
hod
nota
vaila
ble
Rep
rod
ucti
on
Rep
rodu
ctiv
ebi
olog
yan
dec
olog
yla
rgel
yun
know
n
Sto
ckas
sess
men
t(i)
Dev
elop
rout
ine
agin
gm
etho
d(ii
)Agi
ngof
indi
vidu
als
tode
term
ine
age-
at-le
ngth
and
info
rmpo
pula
tion
age
stru
ctur
e(ii
i)U
seB
RU
Vs
surv
eys
tobu
ilda
time
serie
son
whi
chto
estim
ate
biom
ass
and
abun
danc
ein
dice
s(v
iaa
rand
omsa
mpl
ing
desi
gna
cons
iste
ntan
nual
sam
plin
gre
gim
ean
da
broa
dne
twor
kof
sam
plin
gsi
tes)
(iv
)Req
uest
deta
iled
repo
rtin
gof
biol
ogic
alm
etric
s(e
g
leng
ths
exa
nddi
spos
ition
)for
byca
ught
shar
ksby
fishe
ries
obse
rver
sR
epro
duc
tio
n(i)
Use
ultr
asou
ndte
chni
ques
toid
entif
ypr
egna
ntfe
mal
es
(ii)E
lect
roni
cta
gssu
ture
din
tocl
oaca
ofpr
egna
ntfe
mal
esco
uld
help
dete
rmin
ebi
rthi
ngtim
ean
dlo
catio
n(a
spe
rBin
der
etal
20
14)
Sto
ckas
sess
men
t(i)
Dev
elop
Har
vest
Str
ateg
yin
clud
ing
Har
vest
Con
trol
Rul
es(e
g
ICE
SA
ppro
ach
toD
ata-
Lim
ited
Sto
cks)
(ii
)Pla
cevi
deo
mon
itorin
gon
fishi
ngve
ssel
sto
reco
rdpr
esen
cea
bsen
ceof
clas
pers
toes
timat
ese
xra
tio
(iii)
Com
pare
shar
kle
ngth
mea
sure
men
tsto
age-
at-le
ngth
toes
timat
eag
ecl
ass
abun
danc
ere
lativ
eca
ptur
era
tes
Rep
rod
ucti
on
Pro
tect
impo
rtan
tmat
ing
nurs
ery
area
san
dag
greg
atio
nsi
tes
(upo
nid
entifi
catio
n)
Pop
ulat
ion
gene
tics
and
geno
mic
sM
itoch
ondr
ialg
enom
eha
sbe
ense
quen
ced
Spe
cies
phyl
ogen
yha
sbe
ende
term
ined
E
volu
tiona
ryhi
stor
yha
sbe
enth
eoriz
ed
Loca
tion
ofhy
brid
izat
ion
with
Som
nios
uspa
cific
usun
know
nG
ener
atio
ntim
esan
dge
netic
varia
bilit
yam
ong
indi
vidu
als
and
popu
latio
nsun
know
nG
enet
ical
lyde
term
ined
susc
eptib
ility
toen
viro
nmen
talc
hang
eun
clea
r
Dat
aco
llect
ion
Req
uest
at-s
eaob
serv
ers
toco
llect
non-
leth
altis
sue
orpa
rasi
te(iD
NA
)sa
mpl
es
Defi
nep
op
ulat
ions
(i)
Use
gene
tics
toes
timat
ege
nera
tion
times
and
effe
ctiv
epo
pula
tion
size
(eg
us
ing
half
sibl
ing
appr
oach
)(ii
)Defi
nege
netic
ally
dist
inct
popu
latio
nss
tock
sD
efine
rang
eE
xam
inat
ion
ofhy
brid
izat
ion
inso
uthe
rnre
gion
s(e
g
Gul
fofM
exic
o)co
uld
exte
ndth
ekn
own
rang
eof
Gre
enla
ndsh
arks
beyo
ndth
epo
lar
regi
ons
Defi
nep
op
ulat
ions
and
rang
eR
ecom
men
dap
prop
riate
size
and
loca
tion
ofm
anag
emen
tbou
ndar
ies
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 4 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 5
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Mov
emen
teco
logy
Sea
sona
lmo
vem
ents
(i)
Hor
izon
talt
rack
ing
show
slo
ng-d
ista
nce
timed
dire
cted
mig
ratio
ns
(ii)O
bser
ved
occu
rren
cein
shal
low
erw
ater
sw
ithin
crea
sing
latit
ude
(iii)
Vis
ualr
ecor
dsof
philo
patr
yob
serv
edin
St
Law
renc
eE
stua
ry(G
alla
ntpe
rson
alco
mm
unic
atio
n)
Vert
ical
mo
vem
ents
(i)
Rec
ords
ofm
ovem
enti
nsh
allo
ww
ater
and
mid
wat
erco
lum
nun
der
ice
and
ines
tuar
ine
wat
ers
and
(aco
ustic
and
pop-
offs
atel
lite
trac
king
)(ii
)Ver
tical
mov
emen
tsap
pear
tobe
asso
ciat
edw
ithlig
htcy
cle
insh
allo
ww
ater
Sea
sona
lmo
vem
ents
(i)
Lack
oflo
ng-t
erm
(ie
mul
ti-ye
ar)
reco
rds
show
ing
seas
onal
ann
ual
mov
emen
ts
Vert
ical
mo
vem
ents
Li
mite
dre
cord
sof
deta
iled
vert
ical
mov
emen
tbeh
avio
rsav
aila
ble
(eg
ea
rlyde
pth
and
tem
pera
ture
reco
rds
binn
edat
60sndash
6h
inte
rval
s)
Dis
trib
utio
nS
hort
-ter
mm
onito
ring
insu
ffici
entt
oca
ptur
eco
mpl
ete
rang
eof
mov
emen
ts
Sea
sona
lmo
vem
ents
In
crea
sem
onito
ring
dura
tion
usin
glo
ng-t
erm
pass
ive
acou
stic
arc
hiva
lor
sate
llite
met
hods
and
impr
oved
tech
nolo
gies
(eg
ex
tend
edba
tter
ylif
ede
pth
ratin
gse
tc)
Vert
ical
mo
vem
ents
(i)
Use
high
-res
olut
ion
time-
serie
sda
tato
crea
teco
ntin
uous
mov
emen
ttra
cks
(ii)C
reat
est
ate-
spac
em
odel
sto
deco
mpo
sedi
ves
and
asso
ciat
eos
cilla
tory
patt
erns
with
disc
rete
beha
vior
alty
pes
(eg
po
st-r
elea
sere
cove
ry)
(iii)
Com
pare
over
lap
inve
rtic
aldi
strib
utio
nw
ithth
atof
fishe
ries
targ
etsp
ecie
sD
istr
ibut
ion
Use
larg
e-sc
ale
pass
ive
acou
stic
infra
stru
ctur
ean
dlo
ng-t
erm
sate
llite
tele
met
ryap
proa
ches
tode
fine
rang
ean
did
entif
ym
atin
gpu
ppin
ggr
ound
s
Sea
sona
lmo
vem
ents
(i)
Pot
entia
lsea
sona
lclo
sure
sfo
rhu
man
activ
ities
inan
nual
high
use
area
sto
min
imiz
ene
gativ
eim
pact
sto
Gre
enla
ndsh
ark
popu
latio
ns
(ii)D
ynam
icA
rea
Man
agem
entt
opr
otec
tun
expe
cted
aggr
egat
ions
Ve
rtic
alm
ove
men
ts
Info
rmfis
hha
rves
ters
ofsh
ark
beha
vior
patt
erns
tofa
cilit
ate
effo
rts
tom
inim
ize
shar
kca
tch
(eg
se
tgea
rat
dept
hsan
dtim
esof
day
whe
nsh
arks
tend
not
tobe
pres
ent)
Dis
trib
utio
nB
ilate
ralo
rm
ulti-
late
rala
gree
men
tsbe
twee
nna
tions
orvi
aR
egio
nal
Fish
erie
sM
anag
emen
tOrg
aniz
atio
ns(R
FMO
s)to
cond
ucts
cien
cean
dm
anag
esp
ecie
son
apo
pula
tion
leve
l
Beh
avio
ran
din
tera
ctio
nsC
aptu
rean
dre
leas
eP
relim
inar
yta
ggin
gst
udie
sha
veex
amin
edpo
st-r
elea
sebe
havi
oran
dsu
rviv
alra
tes
follo
win
gex
perim
enta
lca
ptur
eFo
rag
ing
beh
avio
r(i)
Indi
rect
evid
ence
ofliv
epr
edat
ion
onse
als
(ii)S
low
swim
spee
dsre
lativ
eto
body
size
buth
igh
roll
acce
lera
tion
(iii)
Dire
ctob
serv
atio
nsof
scav
engi
ng
(iv)A
ggre
gatio
nbe
havi
orob
serv
ed
Cap
ture
and
rele
ase
Pos
t-re
leas
esu
rviv
alra
tes
requ
irefu
rthe
rst
udy
(cur
rent
lyin
suffi
cien
tfor
adju
stin
gm
orta
lity
fact
ors
ina
popu
latio
nm
odel
)Fo
rag
ing
beh
avio
r(i)
Tact
ics
ofpr
eyca
ptur
eun
know
n(a
cryp
ticpr
edat
ion
appr
oach
hypo
thes
ized
)(ii
)Nat
ure
and
frequ
ency
ofin
tra-
and
inte
rspe
cific
inte
ract
ions
unkn
own
Fora
gin
gb
ehav
ior
(i)U
seca
mer
as(s
tilla
ndvi
deo)
depl
oyed
onva
rious
plat
form
s(e
g
anim
als
Rem
ote
Ope
rate
dVe
hicl
esfi
shin
gge
arc
able
s)to
obse
rve
fora
ging
beha
vior
dire
ctly
(ii)U
sean
imal
-bor
nehy
drop
hone
s(s
ampl
eso
unds
cape
)to
asse
ssin
tera
ctio
nsw
ithvo
caliz
ing
anim
als
(eg
m
arin
em
amm
als)
(ii
i)U
seof
VM
Ts(a
cous
tictr
ansp
onde
rs)t
oex
amin
ein
tra-
and
inte
r-sp
ecifi
cin
tera
ctio
ns
Cap
ture
and
rele
ase
Dev
elop
and
prom
ote
safe
hand
ling
prac
tices
for
the
rele
ase
ofsh
arks
caug
htas
byca
tch
Fora
gin
gb
ehav
ior
Use
unde
rsta
ndin
gof
feed
ing
mod
es(e
g
suct
ion
feed
ing)
and
pref
eren
ces
toin
form
gear
mod
ifica
tions
gea
rde
ploy
men
tan
dba
itus
e
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 5 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 6
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Phy
siol
ogic
alad
apta
tions
Die
tand
trop
hic
ecol
ogy
Phy
sio
log
y(i)
Exh
ibit
extr
eme
long
evity
(est
imat
edbe
twee
n27
2an
d51
2ye
ars)
(ii
)Hig
han
tioxi
dant
enzy
me
leve
lsan
dlo
wbl
ood
O2
affin
ityre
cord
ed
Sen
sory
abili
ties
(i)
Olfa
ctor
yor
gans
high
lyde
velo
ped
cons
iste
ntw
ithbe
nthi
cpr
edat
ors
cave
nger
lifes
tyle
(ii
)Hig
hin
cide
nce
ofbl
indn
ess
due
toey
epa
rasi
tism
(exc
epti
nth
eS
tLa
wre
nce
Riv
er)
(iii)
Obs
erva
tions
ofvi
sion
-dep
enda
ntbe
havi
ors
sugg
esta
bilit
yto
unde
rtak
eac
tive
pred
atio
nD
iet
Ben
thic
and
pela
gic
feed
ing
from
pred
omin
antly
offs
hore
tele
osts
ourc
essu
gges
ted
Eco
log
ical
role
G
reen
land
shar
ksar
elik
ely
impo
rtan
tpr
edat
ors
and
likel
yoc
cupy
ahi
ghtr
ophi
cpo
sitio
n
Phy
sio
log
y(i)
Max
imum
rese
rve
dens
itysu
gges
tslif
espa
nm
aycu
rren
tlybe
unde
rest
imat
ed
(ii)S
ensi
tivity
toen
viro
nmen
tals
hifts
unkn
own
(iii)
Det
ails
ofhe
artf
unct
ion
unkn
own
(may
play
aro
lein
long
evity
)M
etab
olic
rate
(i)
Met
abol
icra
tean
dra
teof
O2
cons
umpt
ion
unkn
own
(ii)P
oten
tiall
ink
betw
een
hem
oglo
bin
prop
ertie
san
dco
ld-t
oler
ance
(ii
i)P
oten
tiall
inks
betw
een
high
antio
xida
nten
zym
ele
vels
and
cold
tole
ranc
ede
epdi
ving
beha
vior
and
long
evity
S
enso
ryab
iliti
es
Impo
rtan
ceof
visi
onre
lativ
eto
othe
rse
nses
unkn
own
Die
tP
ossi
bilit
yfo
rsu
b-po
pula
tion
diet
ary
spec
ializ
atio
nun
know
nE
colo
gic
alro
le
(i)E
ffect
ofco
nsum
ptio
nby
Gre
enla
ndsh
arks
onpr
eypo
pula
tions
unkn
own
(ii)R
ole
info
odw
ebst
abiliz
atio
nan
dco
nnec
tivity
unkn
own
(iii)
Rol
eof
long
evity
nutr
ient
ene
rgy
tran
sfer
info
odw
ebs
over
time
and
spac
eun
cert
ain
Met
abo
licra
te
(i)M
easu
reox
ygen
cons
umpt
ion
and
dete
rmin
em
etab
olic
rate
usin
gco
ntai
nmen
tapp
roac
hes
(ii)U
sem
etab
olic
rate
tode
term
ine
ener
getic
requ
irem
ents
pos
sibl
eex
trap
olat
ion
tone
cess
ary
frequ
ency
offe
edin
gus
ing
abi
oene
rget
ics
appr
oach
S
enso
ryab
iliti
es
Furt
her
exam
inat
ion
ofse
nsor
ysy
stem
san
dbr
ain
mor
phol
ogy
requ
ired
(eg
vi
asa
mpl
ing
ofex
perim
enta
land
or
com
mer
cial
byca
tch
mor
talit
ies)
D
iet
(i)C
olle
ctsa
mpl
etis
sues
(mus
cle
bloo
dpl
asm
a)by
non-
leth
alm
etho
dsto
exam
ine
diet
E
colo
gic
alro
le
(i)M
easu
rem
etab
olic
and
cons
umpt
ion
rate
sin
situ
(ii
)Fur
ther
refin
eA
rctic
mar
ine
food
web
stru
ctur
ean
dqu
antif
yen
ergy
flow
and
trop
hic
tran
sfer
effic
ienc
ies
(iii)
Use
Eco
Sim
mod
els
toex
amin
eth
epo
tent
ialt
roph
icef
fect
sof
Gre
enla
ndsh
ark
popu
latio
nin
crea
ses
and
decr
ease
s
Met
abo
licra
te
Indi
vidu
alen
ergy
requ
irem
ents
coul
din
form
popu
latio
n-le
velr
equi
rem
ents
(ie
bio
mas
sre
quire
dto
sust
ain
popu
latio
n)ndash
coul
dpr
ovid
eco
nsid
erat
ions
for
Tota
lAllo
wab
leC
atch
ofpr
eysp
ecie
sS
enso
ryab
iliti
es
Info
rmde
velo
pmen
tofs
hark
dete
rren
tsfo
rfis
hing
gear
mak
ege
arm
ore
notic
eabl
eto
shar
ks(e
g
ping
ers
scen
tel
ectr
onic
wav
es)
Eco
log
ical
role
(i)
Enh
ance
unde
rsta
ndin
gof
Gre
enla
ndsh
arks
and
thei
rec
olog
ical
role
byst
akeh
olde
rsan
dge
nera
lpub
lic
(ii)D
eter
min
esu
stai
nabl
eha
rves
tlev
els
whi
char
epr
edic
ted
tom
aint
ain
ecos
yste
mfu
nctio
n
Frontiers in Marine Science | wwwfrontiersinorg 6 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 7
Edwards et al Advancing Research for Long-Lived Species
TABLE 2 | Research goals and considerations for advancing the study of long-lived species
Research goal Potential methods and considerations for long-lived species
Visual identification of individuals fordemographic studies
Scar patterns and external markings may be used to identify individuals over short time periods (Devine et al 2018)but these may also change or disappear with age (Robbins and Fox 2012) External markers (eg Floy tags) facilitateidentification (Section ldquoDemographics and Life Historyrdquo) however studies should determine the average duration of tagretention by target species
Determining population conservationparameters
While genetic markers allow estimation of effective population sizes (Ne) the application of this approach forconservation purposes requires generation time and a species-specific mutation rate to be accurately defined (seesection ldquoPopulation Genetics and Genomicsrdquo) For species that cannot easily be aged it is difficult to accuratelydetermine generation times and mutation rates The use of evolutionarily significant units (sensu Fraser and Bernatchez2001) should be explored as an alternative
Monitoring behavior over time For species whose movements cover large distances over extended time periods it is important that the duration ofmonitoring (limited by tag lifespan and duration of retention) reflects the temporal scale of the observed behaviorsImprovements to electronic tag design (eg increased battery life) or the use of multi-tag approaches (Hussey et al2018) will extend monitoring durations and can record behaviors occurring over extended timespans (see sectionldquoMovement Ecologyrdquo)
Understanding ontogenetic shifts insensory capabilities and behavior
In fishes ontogenetic changes in the brain and neuroanatomy may affect the relative importance of senses such asvision and olfaction and may in turn be associated with age-related alterations in diet habitat use and activity patterns(Lisney et al 2017) Examinations of brain morphology and behavioral studies should target animals from a broadrange of age-classes to understand the influence of ontogeny on shifts in the ecological role of individuals over centuriesof longevity
Determining the physiology of extremelongevity
Measurements of metabolic rate (ie using oxygen consumption as proxy) blood oxygen consumption and thecardiovascular system (see section ldquoPhysiological Adaptationsrdquo) could provide insight into the physiological mechanismsof longevity
Examining trophic relationships andconnectivity
Extreme longevity may play a role in the ability of mobile predators to connect and stabilize food webs (eg byextending the number of migrations and feeding events that can occur over an animalrsquos lifespan) Similarly anunderstanding of the ecological footprints of long-lived individuals may improve the accuracy of predictive ecosystemmodels (eg EcoSim)
the examined and proposed research attempts were made tominimize this
Demographics and Life HistoryRecent radiocarbon dating of Greenland shark eye lensessupports the long-standing prediction of exceptional longevityin this species (Nielsen et al 2016 Figure 2A) These data alsoprovide the first age-at-length data for Greenland sharks
Current knowledge of Greenland shark distribution andpopulation abundance are based on exploratory fisheries surveysstock assessments of commercial species (eg Greenland halibutReinhardtius hippoglossoides and Northern shrimp Pandalusborealis) and bycatch reports from commercial and historicalfisheries Contemporary catch is restricted to small-scale targetedfisheries in Greenland and Iceland as well as incidental bycatchin a variety of North Atlantic and Arctic fisheries (ICES 2017)In the Barents Sea annual Greenland shark bycatch is estimatedto be 140ndash150 tons (Rusyaev and Orlov 2013) and in northernCanada 105 tons between 1996 and 2015 (Department ofFisheries and Oceans [DFO] 2016) However annual bycatchis likely to be much higher with an estimated bycatch of 1000tonsyr in the Uummannaq district in northwestern Greenlandalone (Gunnarsdottir and Joslashrgensen 2008 ICES 2017)
Scientific catch data suggest regional differences in the relativeabundance of Greenland sharks by life stagesize and sex Forexample no adult females [gt4 m total length (LT) Yanoet al 2007] have been reported throughout the Canadian Arcticand in Svalbard (Norway) despite intense sampling (N gt 300individuals Skomal and Benz 2004 Fisk et al 2002 2012Leclerc et al 2012 Hussey et al 2015a 2018 Devine et al 2018)
In contrast adult females appear to be more frequent offsouthwestern Greenland Iceland and Newfoundland (Canada)(Yano et al 2007 McMeans et al 2010 Nielsen et al 20142016 Campana et al 2015a) and several females gt 5 mhave been measured off Atlantic Canada (Campana personalcommunication) Juvenile sharks defined as individuals le200 cmLT have been observed in both inshore and offshore waters(Hussey et al 2015a) including animals within the birth sizerange (40ndash100 cm MacNeil et al 2012) Specifically juvenileswere observed in longline trawl and camera surveys within ScottInlet Baffin Island and in offshore waters gt1000 m depth (Fisket al 2002 Yano et al 2007 Nielsen et al 2014 Hussey et al2015a Devine et al 2018)
Baited remote underwater video (BRUV) surveys haveprovided the first estimates of the relative abundance ofGreenland sharks in the Canadian Arctic (independent ofbycatch data Devine et al 2018 Figure 2C) By identifyingindividuals using scar patterns and coloration the number ofsharks observed in each camera deployment was quantifiedallowing estimations of observation rate across sampled regions(Devine et al 2018 Figure 2D) BRUV surveys and exploratoryfisheries in the Canadian Arctic Archipelago have also expandedthe speciesrsquo known northern and western ranges in inshoreCanadian waters highlighting the extent to which their range washistorically defined by the spatial extent of commercial fisheries
While radiocarbon dating of eye lenses has provided baselineage estimates for this species these estimates still need tobe verified with an independent accurate and precise agedetermination technique However the absence of hard tissuescontaining growth bands (such as fin spines calcified vertebra
Frontiers in Marine Science | wwwfrontiersinorg 7 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 5
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Mov
emen
teco
logy
Sea
sona
lmo
vem
ents
(i)
Hor
izon
talt
rack
ing
show
slo
ng-d
ista
nce
timed
dire
cted
mig
ratio
ns
(ii)O
bser
ved
occu
rren
cein
shal
low
erw
ater
sw
ithin
crea
sing
latit
ude
(iii)
Vis
ualr
ecor
dsof
philo
patr
yob
serv
edin
St
Law
renc
eE
stua
ry(G
alla
ntpe
rson
alco
mm
unic
atio
n)
Vert
ical
mo
vem
ents
(i)
Rec
ords
ofm
ovem
enti
nsh
allo
ww
ater
and
mid
wat
erco
lum
nun
der
ice
and
ines
tuar
ine
wat
ers
and
(aco
ustic
and
pop-
offs
atel
lite
trac
king
)(ii
)Ver
tical
mov
emen
tsap
pear
tobe
asso
ciat
edw
ithlig
htcy
cle
insh
allo
ww
ater
Sea
sona
lmo
vem
ents
(i)
Lack
oflo
ng-t
erm
(ie
mul
ti-ye
ar)
reco
rds
show
ing
seas
onal
ann
ual
mov
emen
ts
Vert
ical
mo
vem
ents
Li
mite
dre
cord
sof
deta
iled
vert
ical
mov
emen
tbeh
avio
rsav
aila
ble
(eg
ea
rlyde
pth
and
tem
pera
ture
reco
rds
binn
edat
60sndash
6h
inte
rval
s)
Dis
trib
utio
nS
hort
-ter
mm
onito
ring
insu
ffici
entt
oca
ptur
eco
mpl
ete
rang
eof
mov
emen
ts
Sea
sona
lmo
vem
ents
In
crea
sem
onito
ring
dura
tion
usin
glo
ng-t
erm
pass
ive
acou
stic
arc
hiva
lor
sate
llite
met
hods
and
impr
oved
tech
nolo
gies
(eg
ex
tend
edba
tter
ylif
ede
pth
ratin
gse
tc)
Vert
ical
mo
vem
ents
(i)
Use
high
-res
olut
ion
time-
serie
sda
tato
crea
teco
ntin
uous
mov
emen
ttra
cks
(ii)C
reat
est
ate-
spac
em
odel
sto
deco
mpo
sedi
ves
and
asso
ciat
eos
cilla
tory
patt
erns
with
disc
rete
beha
vior
alty
pes
(eg
po
st-r
elea
sere
cove
ry)
(iii)
Com
pare
over
lap
inve
rtic
aldi
strib
utio
nw
ithth
atof
fishe
ries
targ
etsp
ecie
sD
istr
ibut
ion
Use
larg
e-sc
ale
pass
ive
acou
stic
infra
stru
ctur
ean
dlo
ng-t
erm
sate
llite
tele
met
ryap
proa
ches
tode
fine
rang
ean
did
entif
ym
atin
gpu
ppin
ggr
ound
s
Sea
sona
lmo
vem
ents
(i)
Pot
entia
lsea
sona
lclo
sure
sfo
rhu
man
activ
ities
inan
nual
high
use
area
sto
min
imiz
ene
gativ
eim
pact
sto
Gre
enla
ndsh
ark
popu
latio
ns
(ii)D
ynam
icA
rea
Man
agem
entt
opr
otec
tun
expe
cted
aggr
egat
ions
Ve
rtic
alm
ove
men
ts
Info
rmfis
hha
rves
ters
ofsh
ark
beha
vior
patt
erns
tofa
cilit
ate
effo
rts
tom
inim
ize
shar
kca
tch
(eg
se
tgea
rat
dept
hsan
dtim
esof
day
whe
nsh
arks
tend
not
tobe
pres
ent)
Dis
trib
utio
nB
ilate
ralo
rm
ulti-
late
rala
gree
men
tsbe
twee
nna
tions
orvi
aR
egio
nal
Fish
erie
sM
anag
emen
tOrg
aniz
atio
ns(R
FMO
s)to
cond
ucts
cien
cean
dm
anag
esp
ecie
son
apo
pula
tion
leve
l
Beh
avio
ran
din
tera
ctio
nsC
aptu
rean
dre
leas
eP
relim
inar
yta
ggin
gst
udie
sha
veex
amin
edpo
st-r
elea
sebe
havi
oran
dsu
rviv
alra
tes
follo
win
gex
perim
enta
lca
ptur
eFo
rag
ing
beh
avio
r(i)
Indi
rect
evid
ence
ofliv
epr
edat
ion
onse
als
(ii)S
low
swim
spee
dsre
lativ
eto
body
size
buth
igh
roll
acce
lera
tion
(iii)
Dire
ctob
serv
atio
nsof
scav
engi
ng
(iv)A
ggre
gatio
nbe
havi
orob
serv
ed
Cap
ture
and
rele
ase
Pos
t-re
leas
esu
rviv
alra
tes
requ
irefu
rthe
rst
udy
(cur
rent
lyin
suffi
cien
tfor
adju
stin
gm
orta
lity
fact
ors
ina
popu
latio
nm
odel
)Fo
rag
ing
beh
avio
r(i)
Tact
ics
ofpr
eyca
ptur
eun
know
n(a
cryp
ticpr
edat
ion
appr
oach
hypo
thes
ized
)(ii
)Nat
ure
and
frequ
ency
ofin
tra-
and
inte
rspe
cific
inte
ract
ions
unkn
own
Fora
gin
gb
ehav
ior
(i)U
seca
mer
as(s
tilla
ndvi
deo)
depl
oyed
onva
rious
plat
form
s(e
g
anim
als
Rem
ote
Ope
rate
dVe
hicl
esfi
shin
gge
arc
able
s)to
obse
rve
fora
ging
beha
vior
dire
ctly
(ii)U
sean
imal
-bor
nehy
drop
hone
s(s
ampl
eso
unds
cape
)to
asse
ssin
tera
ctio
nsw
ithvo
caliz
ing
anim
als
(eg
m
arin
em
amm
als)
(ii
i)U
seof
VM
Ts(a
cous
tictr
ansp
onde
rs)t
oex
amin
ein
tra-
and
inte
r-sp
ecifi
cin
tera
ctio
ns
Cap
ture
and
rele
ase
Dev
elop
and
prom
ote
safe
hand
ling
prac
tices
for
the
rele
ase
ofsh
arks
caug
htas
byca
tch
Fora
gin
gb
ehav
ior
Use
unde
rsta
ndin
gof
feed
ing
mod
es(e
g
suct
ion
feed
ing)
and
pref
eren
ces
toin
form
gear
mod
ifica
tions
gea
rde
ploy
men
tan
dba
itus
e
(Con
tinue
d)
Frontiers in Marine Science | wwwfrontiersinorg 5 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 6
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Phy
siol
ogic
alad
apta
tions
Die
tand
trop
hic
ecol
ogy
Phy
sio
log
y(i)
Exh
ibit
extr
eme
long
evity
(est
imat
edbe
twee
n27
2an
d51
2ye
ars)
(ii
)Hig
han
tioxi
dant
enzy
me
leve
lsan
dlo
wbl
ood
O2
affin
ityre
cord
ed
Sen
sory
abili
ties
(i)
Olfa
ctor
yor
gans
high
lyde
velo
ped
cons
iste
ntw
ithbe
nthi
cpr
edat
ors
cave
nger
lifes
tyle
(ii
)Hig
hin
cide
nce
ofbl
indn
ess
due
toey
epa
rasi
tism
(exc
epti
nth
eS
tLa
wre
nce
Riv
er)
(iii)
Obs
erva
tions
ofvi
sion
-dep
enda
ntbe
havi
ors
sugg
esta
bilit
yto
unde
rtak
eac
tive
pred
atio
nD
iet
Ben
thic
and
pela
gic
feed
ing
from
pred
omin
antly
offs
hore
tele
osts
ourc
essu
gges
ted
Eco
log
ical
role
G
reen
land
shar
ksar
elik
ely
impo
rtan
tpr
edat
ors
and
likel
yoc
cupy
ahi
ghtr
ophi
cpo
sitio
n
Phy
sio
log
y(i)
Max
imum
rese
rve
dens
itysu
gges
tslif
espa
nm
aycu
rren
tlybe
unde
rest
imat
ed
(ii)S
ensi
tivity
toen
viro
nmen
tals
hifts
unkn
own
(iii)
Det
ails
ofhe
artf
unct
ion
unkn
own
(may
play
aro
lein
long
evity
)M
etab
olic
rate
(i)
Met
abol
icra
tean
dra
teof
O2
cons
umpt
ion
unkn
own
(ii)P
oten
tiall
ink
betw
een
hem
oglo
bin
prop
ertie
san
dco
ld-t
oler
ance
(ii
i)P
oten
tiall
inks
betw
een
high
antio
xida
nten
zym
ele
vels
and
cold
tole
ranc
ede
epdi
ving
beha
vior
and
long
evity
S
enso
ryab
iliti
es
Impo
rtan
ceof
visi
onre
lativ
eto
othe
rse
nses
unkn
own
Die
tP
ossi
bilit
yfo
rsu
b-po
pula
tion
diet
ary
spec
ializ
atio
nun
know
nE
colo
gic
alro
le
(i)E
ffect
ofco
nsum
ptio
nby
Gre
enla
ndsh
arks
onpr
eypo
pula
tions
unkn
own
(ii)R
ole
info
odw
ebst
abiliz
atio
nan
dco
nnec
tivity
unkn
own
(iii)
Rol
eof
long
evity
nutr
ient
ene
rgy
tran
sfer
info
odw
ebs
over
time
and
spac
eun
cert
ain
Met
abo
licra
te
(i)M
easu
reox
ygen
cons
umpt
ion
and
dete
rmin
em
etab
olic
rate
usin
gco
ntai
nmen
tapp
roac
hes
(ii)U
sem
etab
olic
rate
tode
term
ine
ener
getic
requ
irem
ents
pos
sibl
eex
trap
olat
ion
tone
cess
ary
frequ
ency
offe
edin
gus
ing
abi
oene
rget
ics
appr
oach
S
enso
ryab
iliti
es
Furt
her
exam
inat
ion
ofse
nsor
ysy
stem
san
dbr
ain
mor
phol
ogy
requ
ired
(eg
vi
asa
mpl
ing
ofex
perim
enta
land
or
com
mer
cial
byca
tch
mor
talit
ies)
D
iet
(i)C
olle
ctsa
mpl
etis
sues
(mus
cle
bloo
dpl
asm
a)by
non-
leth
alm
etho
dsto
exam
ine
diet
E
colo
gic
alro
le
(i)M
easu
rem
etab
olic
and
cons
umpt
ion
rate
sin
situ
(ii
)Fur
ther
refin
eA
rctic
mar
ine
food
web
stru
ctur
ean
dqu
antif
yen
ergy
flow
and
trop
hic
tran
sfer
effic
ienc
ies
(iii)
Use
Eco
Sim
mod
els
toex
amin
eth
epo
tent
ialt
roph
icef
fect
sof
Gre
enla
ndsh
ark
popu
latio
nin
crea
ses
and
decr
ease
s
Met
abo
licra
te
Indi
vidu
alen
ergy
requ
irem
ents
coul
din
form
popu
latio
n-le
velr
equi
rem
ents
(ie
bio
mas
sre
quire
dto
sust
ain
popu
latio
n)ndash
coul
dpr
ovid
eco
nsid
erat
ions
for
Tota
lAllo
wab
leC
atch
ofpr
eysp
ecie
sS
enso
ryab
iliti
es
Info
rmde
velo
pmen
tofs
hark
dete
rren
tsfo
rfis
hing
gear
mak
ege
arm
ore
notic
eabl
eto
shar
ks(e
g
ping
ers
scen
tel
ectr
onic
wav
es)
Eco
log
ical
role
(i)
Enh
ance
unde
rsta
ndin
gof
Gre
enla
ndsh
arks
and
thei
rec
olog
ical
role
byst
akeh
olde
rsan
dge
nera
lpub
lic
(ii)D
eter
min
esu
stai
nabl
eha
rves
tlev
els
whi
char
epr
edic
ted
tom
aint
ain
ecos
yste
mfu
nctio
n
Frontiers in Marine Science | wwwfrontiersinorg 6 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 7
Edwards et al Advancing Research for Long-Lived Species
TABLE 2 | Research goals and considerations for advancing the study of long-lived species
Research goal Potential methods and considerations for long-lived species
Visual identification of individuals fordemographic studies
Scar patterns and external markings may be used to identify individuals over short time periods (Devine et al 2018)but these may also change or disappear with age (Robbins and Fox 2012) External markers (eg Floy tags) facilitateidentification (Section ldquoDemographics and Life Historyrdquo) however studies should determine the average duration of tagretention by target species
Determining population conservationparameters
While genetic markers allow estimation of effective population sizes (Ne) the application of this approach forconservation purposes requires generation time and a species-specific mutation rate to be accurately defined (seesection ldquoPopulation Genetics and Genomicsrdquo) For species that cannot easily be aged it is difficult to accuratelydetermine generation times and mutation rates The use of evolutionarily significant units (sensu Fraser and Bernatchez2001) should be explored as an alternative
Monitoring behavior over time For species whose movements cover large distances over extended time periods it is important that the duration ofmonitoring (limited by tag lifespan and duration of retention) reflects the temporal scale of the observed behaviorsImprovements to electronic tag design (eg increased battery life) or the use of multi-tag approaches (Hussey et al2018) will extend monitoring durations and can record behaviors occurring over extended timespans (see sectionldquoMovement Ecologyrdquo)
Understanding ontogenetic shifts insensory capabilities and behavior
In fishes ontogenetic changes in the brain and neuroanatomy may affect the relative importance of senses such asvision and olfaction and may in turn be associated with age-related alterations in diet habitat use and activity patterns(Lisney et al 2017) Examinations of brain morphology and behavioral studies should target animals from a broadrange of age-classes to understand the influence of ontogeny on shifts in the ecological role of individuals over centuriesof longevity
Determining the physiology of extremelongevity
Measurements of metabolic rate (ie using oxygen consumption as proxy) blood oxygen consumption and thecardiovascular system (see section ldquoPhysiological Adaptationsrdquo) could provide insight into the physiological mechanismsof longevity
Examining trophic relationships andconnectivity
Extreme longevity may play a role in the ability of mobile predators to connect and stabilize food webs (eg byextending the number of migrations and feeding events that can occur over an animalrsquos lifespan) Similarly anunderstanding of the ecological footprints of long-lived individuals may improve the accuracy of predictive ecosystemmodels (eg EcoSim)
the examined and proposed research attempts were made tominimize this
Demographics and Life HistoryRecent radiocarbon dating of Greenland shark eye lensessupports the long-standing prediction of exceptional longevityin this species (Nielsen et al 2016 Figure 2A) These data alsoprovide the first age-at-length data for Greenland sharks
Current knowledge of Greenland shark distribution andpopulation abundance are based on exploratory fisheries surveysstock assessments of commercial species (eg Greenland halibutReinhardtius hippoglossoides and Northern shrimp Pandalusborealis) and bycatch reports from commercial and historicalfisheries Contemporary catch is restricted to small-scale targetedfisheries in Greenland and Iceland as well as incidental bycatchin a variety of North Atlantic and Arctic fisheries (ICES 2017)In the Barents Sea annual Greenland shark bycatch is estimatedto be 140ndash150 tons (Rusyaev and Orlov 2013) and in northernCanada 105 tons between 1996 and 2015 (Department ofFisheries and Oceans [DFO] 2016) However annual bycatchis likely to be much higher with an estimated bycatch of 1000tonsyr in the Uummannaq district in northwestern Greenlandalone (Gunnarsdottir and Joslashrgensen 2008 ICES 2017)
Scientific catch data suggest regional differences in the relativeabundance of Greenland sharks by life stagesize and sex Forexample no adult females [gt4 m total length (LT) Yanoet al 2007] have been reported throughout the Canadian Arcticand in Svalbard (Norway) despite intense sampling (N gt 300individuals Skomal and Benz 2004 Fisk et al 2002 2012Leclerc et al 2012 Hussey et al 2015a 2018 Devine et al 2018)
In contrast adult females appear to be more frequent offsouthwestern Greenland Iceland and Newfoundland (Canada)(Yano et al 2007 McMeans et al 2010 Nielsen et al 20142016 Campana et al 2015a) and several females gt 5 mhave been measured off Atlantic Canada (Campana personalcommunication) Juvenile sharks defined as individuals le200 cmLT have been observed in both inshore and offshore waters(Hussey et al 2015a) including animals within the birth sizerange (40ndash100 cm MacNeil et al 2012) Specifically juvenileswere observed in longline trawl and camera surveys within ScottInlet Baffin Island and in offshore waters gt1000 m depth (Fisket al 2002 Yano et al 2007 Nielsen et al 2014 Hussey et al2015a Devine et al 2018)
Baited remote underwater video (BRUV) surveys haveprovided the first estimates of the relative abundance ofGreenland sharks in the Canadian Arctic (independent ofbycatch data Devine et al 2018 Figure 2C) By identifyingindividuals using scar patterns and coloration the number ofsharks observed in each camera deployment was quantifiedallowing estimations of observation rate across sampled regions(Devine et al 2018 Figure 2D) BRUV surveys and exploratoryfisheries in the Canadian Arctic Archipelago have also expandedthe speciesrsquo known northern and western ranges in inshoreCanadian waters highlighting the extent to which their range washistorically defined by the spatial extent of commercial fisheries
While radiocarbon dating of eye lenses has provided baselineage estimates for this species these estimates still need tobe verified with an independent accurate and precise agedetermination technique However the absence of hard tissuescontaining growth bands (such as fin spines calcified vertebra
Frontiers in Marine Science | wwwfrontiersinorg 7 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 6
Edwards et al Advancing Research for Long-Lived Species
TAB
LE1
|Con
tinue
d
Res
earc
hto
pic
Cur
rent
kno
wle
dg
eK
now
led
ge
gap
san
dlim
itat
ions
Str
ateg
ies
toad
dre
ssd
ata
defi
cien
cies
Po
tent
iala
ctio
nsto
sup
po
rtm
anag
emen
t
Phy
siol
ogic
alad
apta
tions
Die
tand
trop
hic
ecol
ogy
Phy
sio
log
y(i)
Exh
ibit
extr
eme
long
evity
(est
imat
edbe
twee
n27
2an
d51
2ye
ars)
(ii
)Hig
han
tioxi
dant
enzy
me
leve
lsan
dlo
wbl
ood
O2
affin
ityre
cord
ed
Sen
sory
abili
ties
(i)
Olfa
ctor
yor
gans
high
lyde
velo
ped
cons
iste
ntw
ithbe
nthi
cpr
edat
ors
cave
nger
lifes
tyle
(ii
)Hig
hin
cide
nce
ofbl
indn
ess
due
toey
epa
rasi
tism
(exc
epti
nth
eS
tLa
wre
nce
Riv
er)
(iii)
Obs
erva
tions
ofvi
sion
-dep
enda
ntbe
havi
ors
sugg
esta
bilit
yto
unde
rtak
eac
tive
pred
atio
nD
iet
Ben
thic
and
pela
gic
feed
ing
from
pred
omin
antly
offs
hore
tele
osts
ourc
essu
gges
ted
Eco
log
ical
role
G
reen
land
shar
ksar
elik
ely
impo
rtan
tpr
edat
ors
and
likel
yoc
cupy
ahi
ghtr
ophi
cpo
sitio
n
Phy
sio
log
y(i)
Max
imum
rese
rve
dens
itysu
gges
tslif
espa
nm
aycu
rren
tlybe
unde
rest
imat
ed
(ii)S
ensi
tivity
toen
viro
nmen
tals
hifts
unkn
own
(iii)
Det
ails
ofhe
artf
unct
ion
unkn
own
(may
play
aro
lein
long
evity
)M
etab
olic
rate
(i)
Met
abol
icra
tean
dra
teof
O2
cons
umpt
ion
unkn
own
(ii)P
oten
tiall
ink
betw
een
hem
oglo
bin
prop
ertie
san
dco
ld-t
oler
ance
(ii
i)P
oten
tiall
inks
betw
een
high
antio
xida
nten
zym
ele
vels
and
cold
tole
ranc
ede
epdi
ving
beha
vior
and
long
evity
S
enso
ryab
iliti
es
Impo
rtan
ceof
visi
onre
lativ
eto
othe
rse
nses
unkn
own
Die
tP
ossi
bilit
yfo
rsu
b-po
pula
tion
diet
ary
spec
ializ
atio
nun
know
nE
colo
gic
alro
le
(i)E
ffect
ofco
nsum
ptio
nby
Gre
enla
ndsh
arks
onpr
eypo
pula
tions
unkn
own
(ii)R
ole
info
odw
ebst
abiliz
atio
nan
dco
nnec
tivity
unkn
own
(iii)
Rol
eof
long
evity
nutr
ient
ene
rgy
tran
sfer
info
odw
ebs
over
time
and
spac
eun
cert
ain
Met
abo
licra
te
(i)M
easu
reox
ygen
cons
umpt
ion
and
dete
rmin
em
etab
olic
rate
usin
gco
ntai
nmen
tapp
roac
hes
(ii)U
sem
etab
olic
rate
tode
term
ine
ener
getic
requ
irem
ents
pos
sibl
eex
trap
olat
ion
tone
cess
ary
frequ
ency
offe
edin
gus
ing
abi
oene
rget
ics
appr
oach
S
enso
ryab
iliti
es
Furt
her
exam
inat
ion
ofse
nsor
ysy
stem
san
dbr
ain
mor
phol
ogy
requ
ired
(eg
vi
asa
mpl
ing
ofex
perim
enta
land
or
com
mer
cial
byca
tch
mor
talit
ies)
D
iet
(i)C
olle
ctsa
mpl
etis
sues
(mus
cle
bloo
dpl
asm
a)by
non-
leth
alm
etho
dsto
exam
ine
diet
E
colo
gic
alro
le
(i)M
easu
rem
etab
olic
and
cons
umpt
ion
rate
sin
situ
(ii
)Fur
ther
refin
eA
rctic
mar
ine
food
web
stru
ctur
ean
dqu
antif
yen
ergy
flow
and
trop
hic
tran
sfer
effic
ienc
ies
(iii)
Use
Eco
Sim
mod
els
toex
amin
eth
epo
tent
ialt
roph
icef
fect
sof
Gre
enla
ndsh
ark
popu
latio
nin
crea
ses
and
decr
ease
s
Met
abo
licra
te
Indi
vidu
alen
ergy
requ
irem
ents
coul
din
form
popu
latio
n-le
velr
equi
rem
ents
(ie
bio
mas
sre
quire
dto
sust
ain
popu
latio
n)ndash
coul
dpr
ovid
eco
nsid
erat
ions
for
Tota
lAllo
wab
leC
atch
ofpr
eysp
ecie
sS
enso
ryab
iliti
es
Info
rmde
velo
pmen
tofs
hark
dete
rren
tsfo
rfis
hing
gear
mak
ege
arm
ore
notic
eabl
eto
shar
ks(e
g
ping
ers
scen
tel
ectr
onic
wav
es)
Eco
log
ical
role
(i)
Enh
ance
unde
rsta
ndin
gof
Gre
enla
ndsh
arks
and
thei
rec
olog
ical
role
byst
akeh
olde
rsan
dge
nera
lpub
lic
(ii)D
eter
min
esu
stai
nabl
eha
rves
tlev
els
whi
char
epr
edic
ted
tom
aint
ain
ecos
yste
mfu
nctio
n
Frontiers in Marine Science | wwwfrontiersinorg 6 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 7
Edwards et al Advancing Research for Long-Lived Species
TABLE 2 | Research goals and considerations for advancing the study of long-lived species
Research goal Potential methods and considerations for long-lived species
Visual identification of individuals fordemographic studies
Scar patterns and external markings may be used to identify individuals over short time periods (Devine et al 2018)but these may also change or disappear with age (Robbins and Fox 2012) External markers (eg Floy tags) facilitateidentification (Section ldquoDemographics and Life Historyrdquo) however studies should determine the average duration of tagretention by target species
Determining population conservationparameters
While genetic markers allow estimation of effective population sizes (Ne) the application of this approach forconservation purposes requires generation time and a species-specific mutation rate to be accurately defined (seesection ldquoPopulation Genetics and Genomicsrdquo) For species that cannot easily be aged it is difficult to accuratelydetermine generation times and mutation rates The use of evolutionarily significant units (sensu Fraser and Bernatchez2001) should be explored as an alternative
Monitoring behavior over time For species whose movements cover large distances over extended time periods it is important that the duration ofmonitoring (limited by tag lifespan and duration of retention) reflects the temporal scale of the observed behaviorsImprovements to electronic tag design (eg increased battery life) or the use of multi-tag approaches (Hussey et al2018) will extend monitoring durations and can record behaviors occurring over extended timespans (see sectionldquoMovement Ecologyrdquo)
Understanding ontogenetic shifts insensory capabilities and behavior
In fishes ontogenetic changes in the brain and neuroanatomy may affect the relative importance of senses such asvision and olfaction and may in turn be associated with age-related alterations in diet habitat use and activity patterns(Lisney et al 2017) Examinations of brain morphology and behavioral studies should target animals from a broadrange of age-classes to understand the influence of ontogeny on shifts in the ecological role of individuals over centuriesof longevity
Determining the physiology of extremelongevity
Measurements of metabolic rate (ie using oxygen consumption as proxy) blood oxygen consumption and thecardiovascular system (see section ldquoPhysiological Adaptationsrdquo) could provide insight into the physiological mechanismsof longevity
Examining trophic relationships andconnectivity
Extreme longevity may play a role in the ability of mobile predators to connect and stabilize food webs (eg byextending the number of migrations and feeding events that can occur over an animalrsquos lifespan) Similarly anunderstanding of the ecological footprints of long-lived individuals may improve the accuracy of predictive ecosystemmodels (eg EcoSim)
the examined and proposed research attempts were made tominimize this
Demographics and Life HistoryRecent radiocarbon dating of Greenland shark eye lensessupports the long-standing prediction of exceptional longevityin this species (Nielsen et al 2016 Figure 2A) These data alsoprovide the first age-at-length data for Greenland sharks
Current knowledge of Greenland shark distribution andpopulation abundance are based on exploratory fisheries surveysstock assessments of commercial species (eg Greenland halibutReinhardtius hippoglossoides and Northern shrimp Pandalusborealis) and bycatch reports from commercial and historicalfisheries Contemporary catch is restricted to small-scale targetedfisheries in Greenland and Iceland as well as incidental bycatchin a variety of North Atlantic and Arctic fisheries (ICES 2017)In the Barents Sea annual Greenland shark bycatch is estimatedto be 140ndash150 tons (Rusyaev and Orlov 2013) and in northernCanada 105 tons between 1996 and 2015 (Department ofFisheries and Oceans [DFO] 2016) However annual bycatchis likely to be much higher with an estimated bycatch of 1000tonsyr in the Uummannaq district in northwestern Greenlandalone (Gunnarsdottir and Joslashrgensen 2008 ICES 2017)
Scientific catch data suggest regional differences in the relativeabundance of Greenland sharks by life stagesize and sex Forexample no adult females [gt4 m total length (LT) Yanoet al 2007] have been reported throughout the Canadian Arcticand in Svalbard (Norway) despite intense sampling (N gt 300individuals Skomal and Benz 2004 Fisk et al 2002 2012Leclerc et al 2012 Hussey et al 2015a 2018 Devine et al 2018)
In contrast adult females appear to be more frequent offsouthwestern Greenland Iceland and Newfoundland (Canada)(Yano et al 2007 McMeans et al 2010 Nielsen et al 20142016 Campana et al 2015a) and several females gt 5 mhave been measured off Atlantic Canada (Campana personalcommunication) Juvenile sharks defined as individuals le200 cmLT have been observed in both inshore and offshore waters(Hussey et al 2015a) including animals within the birth sizerange (40ndash100 cm MacNeil et al 2012) Specifically juvenileswere observed in longline trawl and camera surveys within ScottInlet Baffin Island and in offshore waters gt1000 m depth (Fisket al 2002 Yano et al 2007 Nielsen et al 2014 Hussey et al2015a Devine et al 2018)
Baited remote underwater video (BRUV) surveys haveprovided the first estimates of the relative abundance ofGreenland sharks in the Canadian Arctic (independent ofbycatch data Devine et al 2018 Figure 2C) By identifyingindividuals using scar patterns and coloration the number ofsharks observed in each camera deployment was quantifiedallowing estimations of observation rate across sampled regions(Devine et al 2018 Figure 2D) BRUV surveys and exploratoryfisheries in the Canadian Arctic Archipelago have also expandedthe speciesrsquo known northern and western ranges in inshoreCanadian waters highlighting the extent to which their range washistorically defined by the spatial extent of commercial fisheries
While radiocarbon dating of eye lenses has provided baselineage estimates for this species these estimates still need tobe verified with an independent accurate and precise agedetermination technique However the absence of hard tissuescontaining growth bands (such as fin spines calcified vertebra
Frontiers in Marine Science | wwwfrontiersinorg 7 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 7
Edwards et al Advancing Research for Long-Lived Species
TABLE 2 | Research goals and considerations for advancing the study of long-lived species
Research goal Potential methods and considerations for long-lived species
Visual identification of individuals fordemographic studies
Scar patterns and external markings may be used to identify individuals over short time periods (Devine et al 2018)but these may also change or disappear with age (Robbins and Fox 2012) External markers (eg Floy tags) facilitateidentification (Section ldquoDemographics and Life Historyrdquo) however studies should determine the average duration of tagretention by target species
Determining population conservationparameters
While genetic markers allow estimation of effective population sizes (Ne) the application of this approach forconservation purposes requires generation time and a species-specific mutation rate to be accurately defined (seesection ldquoPopulation Genetics and Genomicsrdquo) For species that cannot easily be aged it is difficult to accuratelydetermine generation times and mutation rates The use of evolutionarily significant units (sensu Fraser and Bernatchez2001) should be explored as an alternative
Monitoring behavior over time For species whose movements cover large distances over extended time periods it is important that the duration ofmonitoring (limited by tag lifespan and duration of retention) reflects the temporal scale of the observed behaviorsImprovements to electronic tag design (eg increased battery life) or the use of multi-tag approaches (Hussey et al2018) will extend monitoring durations and can record behaviors occurring over extended timespans (see sectionldquoMovement Ecologyrdquo)
Understanding ontogenetic shifts insensory capabilities and behavior
In fishes ontogenetic changes in the brain and neuroanatomy may affect the relative importance of senses such asvision and olfaction and may in turn be associated with age-related alterations in diet habitat use and activity patterns(Lisney et al 2017) Examinations of brain morphology and behavioral studies should target animals from a broadrange of age-classes to understand the influence of ontogeny on shifts in the ecological role of individuals over centuriesof longevity
Determining the physiology of extremelongevity
Measurements of metabolic rate (ie using oxygen consumption as proxy) blood oxygen consumption and thecardiovascular system (see section ldquoPhysiological Adaptationsrdquo) could provide insight into the physiological mechanismsof longevity
Examining trophic relationships andconnectivity
Extreme longevity may play a role in the ability of mobile predators to connect and stabilize food webs (eg byextending the number of migrations and feeding events that can occur over an animalrsquos lifespan) Similarly anunderstanding of the ecological footprints of long-lived individuals may improve the accuracy of predictive ecosystemmodels (eg EcoSim)
the examined and proposed research attempts were made tominimize this
Demographics and Life HistoryRecent radiocarbon dating of Greenland shark eye lensessupports the long-standing prediction of exceptional longevityin this species (Nielsen et al 2016 Figure 2A) These data alsoprovide the first age-at-length data for Greenland sharks
Current knowledge of Greenland shark distribution andpopulation abundance are based on exploratory fisheries surveysstock assessments of commercial species (eg Greenland halibutReinhardtius hippoglossoides and Northern shrimp Pandalusborealis) and bycatch reports from commercial and historicalfisheries Contemporary catch is restricted to small-scale targetedfisheries in Greenland and Iceland as well as incidental bycatchin a variety of North Atlantic and Arctic fisheries (ICES 2017)In the Barents Sea annual Greenland shark bycatch is estimatedto be 140ndash150 tons (Rusyaev and Orlov 2013) and in northernCanada 105 tons between 1996 and 2015 (Department ofFisheries and Oceans [DFO] 2016) However annual bycatchis likely to be much higher with an estimated bycatch of 1000tonsyr in the Uummannaq district in northwestern Greenlandalone (Gunnarsdottir and Joslashrgensen 2008 ICES 2017)
Scientific catch data suggest regional differences in the relativeabundance of Greenland sharks by life stagesize and sex Forexample no adult females [gt4 m total length (LT) Yanoet al 2007] have been reported throughout the Canadian Arcticand in Svalbard (Norway) despite intense sampling (N gt 300individuals Skomal and Benz 2004 Fisk et al 2002 2012Leclerc et al 2012 Hussey et al 2015a 2018 Devine et al 2018)
In contrast adult females appear to be more frequent offsouthwestern Greenland Iceland and Newfoundland (Canada)(Yano et al 2007 McMeans et al 2010 Nielsen et al 20142016 Campana et al 2015a) and several females gt 5 mhave been measured off Atlantic Canada (Campana personalcommunication) Juvenile sharks defined as individuals le200 cmLT have been observed in both inshore and offshore waters(Hussey et al 2015a) including animals within the birth sizerange (40ndash100 cm MacNeil et al 2012) Specifically juvenileswere observed in longline trawl and camera surveys within ScottInlet Baffin Island and in offshore waters gt1000 m depth (Fisket al 2002 Yano et al 2007 Nielsen et al 2014 Hussey et al2015a Devine et al 2018)
Baited remote underwater video (BRUV) surveys haveprovided the first estimates of the relative abundance ofGreenland sharks in the Canadian Arctic (independent ofbycatch data Devine et al 2018 Figure 2C) By identifyingindividuals using scar patterns and coloration the number ofsharks observed in each camera deployment was quantifiedallowing estimations of observation rate across sampled regions(Devine et al 2018 Figure 2D) BRUV surveys and exploratoryfisheries in the Canadian Arctic Archipelago have also expandedthe speciesrsquo known northern and western ranges in inshoreCanadian waters highlighting the extent to which their range washistorically defined by the spatial extent of commercial fisheries
While radiocarbon dating of eye lenses has provided baselineage estimates for this species these estimates still need tobe verified with an independent accurate and precise agedetermination technique However the absence of hard tissuescontaining growth bands (such as fin spines calcified vertebra
Frontiers in Marine Science | wwwfrontiersinorg 7 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 8
Edwards et al Advancing Research for Long-Lived Species
FIGURE 2 | Examples of Greenland shark demographics research (A) Greenland shark eye lens for use in radiocarbon dating (photo Julius Nielsen used withpermission) (B) Measuring the length of a Greenland shark captured through ice (photo Gregory Skomal used with permission) (C) Multiple Greenland sharkscaptured on BRUV footage (photo Brynn Devine used with permission) (D) BRUV image showing scar patterns used for identification of individual Greenland sharks(photo Brynn Devine used with permission) (E) Greenland sharks caught as bycatch in a fisheries trawl (photo Julius Nielsen used with permission)
etc) complicates such a validation Other chemical datingtechniques such as aspartic acid racemization (which has beensuccessfully applied on several cetaceans George et al 1999Garde et al 2007) have proven to be unsuccessful in providingindependent age estimates for this species (Nielsen 2013)highlighting the need for innovative aging methodologies
In addition many questions concerning populationdemographics ndash particularly population size and productivityas well as reproductive biology fertility and natural mortalityremain unknown or poorly understood For example knowledge
of the reproductive biology of Greenland sharks is extremelylimited Only one pregnant female with ten near-term pups of37 cm LT has been reported (Koefoed 1957) Observations ofa high ovarian fecundity (gt400 goose-egg sized unfertilizedova Bjerkan 1957 Yano et al 2007 Nielsen et al 2014Campana personal communication) suggest that embryos maybe aplacental viviparous similar to those of spiny dogfish(Squalus acanthias) and Portuguese sharks (Centroscymnuscoelolepis) (Campana et al 2009 Castro 2011) The gestationperiod the locations of mating and pupping grounds and the
Frontiers in Marine Science | wwwfrontiersinorg 8 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 9
Edwards et al Advancing Research for Long-Lived Species
mating period all remain unknown Greenland sharks mayexhibit seasonal migrations for reproductive purposes (matingand pupping) but the existence or possible extent of connectivitybetween regions has yet to be documented Given its extendedlongevity and the low productivity of sharks relative to teleosts(Musick 1999a) the Greenland shark is expected to haveparticularly low productivity
There is also a general lack of knowledge on the size and sex ofGreenland sharks caught as bycatch in commercial fisheries (pastand present) (Section ldquoThreats to Greenland Shark PopulationsrdquoFigure 2E) Consequently data available to evaluate the spatialdistribution of shark life stages are sparse Fishery-derived dataare also problematic for estimating population abundance dueto the spatially targeted nature of fisheries which limits theability to extrapolate catch rates to larger areas Estimation ofthe spatial distribution of Greenland sharks using commercialbycatch records is further confounded by misidentification withother large (but unrelated) shark species for instance the baskingshark (Cetorhinus maximus Campana et al 2008b)
Improved or novel age determination methods need to bedeveloped for the routine aging of Greenland sharks (andsleeper sharks in general) With uncertainty surrounding ageestimates for Greenland sharks an improved understandingof the residence time and pathway of past bomb radiocarbonpulses into deep Arctic waters and in Greenland shark preyis required to improve the precision and accuracy of theradiocarbon dating method Data are available for the otolithsof some Arctic fishes beluga whale teeth and vertebraefrom several lamniform sharks (Stewart et al 2006 Campanaet al 2008a Hamady et al 2014) but the organic pathwayfor the eye lens is not well-documented Equally mark-recapture methods could provide accurate estimates of size-dependent growth which can be used to model growthrate (Francis 1988) and thus age with few assumptionsHowever acquiring precise lengths for such large sharks atboth tagging and recapture requires careful measurementin the field under logistically challenging conditions (asdescribed by Hansen 1963 see also Figure 2B) and areliance on fishers to provide accurate length estimates forbycaught sharks
The broad-scale use of BRUVs and modification ofestablished population assessment methodologies shouldaddress many remaining questions about Greenland sharkdistribution and abundance (Table 1) Systematic surveysof local traditional knowledge and accounts from fishermencould help fill gaps in distribution while the expansionof BRUV surveys could explore differences in relativeabundances among sites and seasonal BRUV surveys couldhelp determine cyclic patterns in distribution Althoughscar patterns and the coloration of individuals may changeover time (Robbins and Fox 2012) the use of coloredFloy tags or other external markers could provide reliablelong-term identification for both BRUVs and fishery-mediated surveys (Table 2) By modifying conventionalmark-recapture models to integrate auxiliary data derivedfrom acoustic or satellite telemetry the effects of low recapturerates can be offset resulting in more precise estimates of
demographic parameters such as apparent survival andabundance (Dudgeon et al 2015)
More data are needed to evaluate variation in life stagedistribution patterns among regions Such data can be collectedfrom sharks caught in commercial fisheries as bycatch (egimproving data collection on capture location LT and sexsee Table 1) as well as by expanding large-scale long-termtagging studies to include all life stages (see section ldquoMovementEcologyrdquo) Detailed reporting of Greenland shark bycatch willalso improve growth rate estimates as well as our understandingof exploitation rates and migration pathways and could highlightsensitive areas for specific life stages Tagging and release ofmature females with pop-up archival satellite tags (during theopen water season) could help identify critical habitats or areasthat are important for mating andor pupping while at-seasampling of bycatch mortalities will provide opportunities forthe biological examination of mature or maturing females andadditional data on ovarian and uterine fecundity
Biological reference points and conservation targets typicallyrequire either population productivity values (based on growthand reproduction) or stock-recruitment curves (based onspawning stock biomass and fecundity) In the absence ofthese data for Greenland sharks precautionary measures will beneeded to protect sensitive habitats (such as potential matinggrounds pupping grounds and nursery areas) and limit totalcatch Identification of these ecologically important habitats istherefore critical for mitigating bycatch (see section ldquoMovementEcologyrdquo) Another key difficulty in setting conservation limitsis uncertainty over whether historical catch rates of around50000 individualsyr in the northern European liver oil fishery(MacNeil et al 2012) were sustained by local populations oremigration from a wider meta-population Given the importanceof these population-level data for implementing precautionarymeasures assessments of current Greenland shark abundanceand distribution should be made a high priority (Table 1)
Population Genetics and GenomicsMolecular genetic data primarily based on mitochondrialDNA (mtDNA) has yielded important insight into thephylogeographic evolutionary history of the Greenland sharkand other members of the genus Somniosus Early work focusedon the Pacific sleeper shark (S pacificus) but also includeda number of Greenland sharks and revealed relatively shallowgenetic divergence between these two species (sim18 sequencedivergence Murray et al 2008) This study also documentedthe first evidence of haplotype sharing between two recognizedSomniosus species (S pacificus and the southern sleeper sharkS antarcticus Murray et al 2008) contrasting the earlierresurrection of S antarcticus as a distinct species (Yano et al2004) Further evidence of interspecies haplotype sharing hasbeen reported for several juvenile Greenland sharks from thehigh Arctic that were found to be carrying S pacificus mtDNAhaplotypes (Hussey et al 2015a) More recently a full mtDNAgenome sequenced with phylogenetic reconstruction placed theGreenland shark and Pacific sleeper shark as sister species withrespect to other major shark orders (Santaquiteria et al 2017)By calibrating the mitogenomic phylogenetic reconstruction
Frontiers in Marine Science | wwwfrontiersinorg 9 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 10
Edwards et al Advancing Research for Long-Lived Species
with fossil records and geological events the speciation timeof these two species was confirmed to be 35 million yearsago (mya) (Santaquiteria 2016 in agreement with Murrayet al 2008) A further phylogeographic study also confirmedmtDNA haplotype sharing between Greenland and Pacific sleepersharks at additional locations as well as nuclear admixtureimplicating interspecific hybridization as a phenomenon amongAtlantic Somniosids (Walter et al 2017) These data yielded amore recent speciation time for Greenland sharks than findingsobtained using whole mitogenomes (21 mya versus 35 mya)Reconciling the rates of intra- and inter-specific admixtureamong the Somniosids with (incipient) speciation therefore holdsmuch promise for clarifying the phylogeographic evolutionaryhistory of this genus
Reliance on mtDNA data to date paints an incomplete pictureof the evolutionary history of the Greenland shark Sharedgenetic signatures among extant species have led researchers toquestion the validity of current species descriptions suggestingthat a more comprehensive systematic revision of the genus isneeded In addition to signals of interspecific gene flow andincipient speciation the demographic history of the Greenlandshark across its range indicates that the Atlantic populationhas persisted at a low but stable size over the last 5 millionyears This was likely followed by a recent population increasearound 500000 years ago possibly linked to fluctuations in Arcticsea ice conditions during the Pleistocene As yet no clear orconspicuous regional geographic structure has emerged fromthe mtDNA data aside from indications of genetic admixture(Murray et al 2008 Walter et al 2017)
The potential for extreme longevity exhibited by Greenlandsharks (Nielsen et al 2016) presents unique challenges forestimating relevant conservation parameters such as effectivepopulation size (Ne) (Table 2) Firstly the value of Ne ishighly sensitive to generation time While estimates of Ne(both long-term and short-term estimates) are possible fromgenetic data the translation of these values to biologicallyrelevant terms is not straightforward For example coalescent-based estimates of long-term Ne were obtained from a 702 bpfragment of Cytochrome b using 20 and 150 years generationtimes (Walter et al 2017) the latter based on an estimatedage at first reproduction of 156 plusmn 22 years (Nielsen et al2016) These long generation times yielded Ne estimates 75times
smaller than if a 20 years generation time was used It isimportant to note that long-term Ne estimates are largely onlyapplicable for comparative rather than conservation purposesNevertheless these scalar problems will remain a challengeeven with genomic advances until firm generation times forGreenland sharks are determined
Estimates of mutation rates in elasmobranchs are consideredto be among the slowest in the animal kingdom (Martinet al 1992) To date mutation rates for Somniosus spp areunknown Consequently much of the dating for speciationtimes are hypothetical however the accumulation of genome-wide variation among multiple Greenland shark individualsholds promise for estimating Somniosus-specific mutation ratesObtaining approximate timing of speciation events populationexpansions and reductions and other genus-specific events using
genetic methods will continue to be problematic until accuratemutation rates can be estimated
Advances in population genomics including completemitogenomes nuclear genomes and genotype-by-sequencingapproaches appear promising for detecting population structureand evolutionary relationships for the Greenland shark andSomniosids in general The most desirable nuclear molecularmarkers are those that permit detection and spatial delineationof populations (Ahonen et al 2009) Such resolution is criticalfor determining evolutionarily significant units (ESUs Table 2)or management units for conservation and managementplanning (Moritz 1994) Genomic and genetic resources suchas numerous single nucleotide polymorphism (SNP) loci andhighly polymorphic nuclear microsatellites may provide thetools for determining the number of extant populations andthe extent of connectivity among them (Milano et al 2014)Novel sampling methodologies such as the collection of externalparasites to obtain host mitochondrial DNA sequences (Meekanet al 2017) have proven effective for obtaining genetic samplesfor such analyses These invertebrate DNA (iDNA) sequenceshave helped resolve the genetic structure and connectivity ofglobal whale shark populations (Rhincodon typus Meekan et al2017) and could be similarly applied using copepods sampledfrom parasitized Greenland sharks While the spatial scale ofelasmobranch populations varies widely from species to speciesthe geographical range of Greenland shark populations is likelyfar-reaching The coupling of population genomic structure forS microcephalus in the Atlantic and beyond with movement datafor analyzed individuals will yield the most accurate distributiondata for this species
Despite growing conservation concerns for many sharkspecies the assignment of conservation status and the effectivemanagement of at-risk populations is commonly hindered by alack of knowledge of stock structure and estimates of absolutepopulation size Detailed examination of the genomic structureof Greenland shark populations (using non-lethal biopsies oriDNA Meekan et al 2017) might allow researchers to definegenetically distinct stocks thereby supporting the delineationof appropriate management boundaries and multinationalagreements Furthermore the examination of genetic relatednessamong conspecifics is becoming an increasingly popular tool toestimate total population abundance (Table 1) For example arecent study used genetic analyses to identify half-sibling pairs(HSPs) and unrelated pairs (UPs) to estimate abundance andsurvival rates of adult white sharks in Eastern Australia andNew Zealand (Carcharodon carcharias) These data were furthersupplemented by acoustic tag data to estimate juvenile survivalrates Ultimately this method provided direct estimates of totalabundance across both spatial and temporal life-history gradients(Hillary et al 2018) and could similarly be used to assessGreenland shark populations
Movement EcologyData on Greenland shark movements have been primarilyrecorded using a variety of telemetry technologies Early workinvolved active short-term acoustic tracking of sharks under ice(Skomal and Benz 2004 Figure 3F) and in estuarine waters
Frontiers in Marine Science | wwwfrontiersinorg 10 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 11
Edwards et al Advancing Research for Long-Lived Species
FIGURE 3 | Examples of Greenland shark movement studies (A) A biologging package containing an accelerometer (Little Leonardo) mounted to the head (photo copyYuuki Watanabe used with permission) (B) A biologging package containing a spot 6 tag (Wildlife Computers) a VHF tag an accelerometer (Maritime Biologgers)and a CTD (Star Oddi) (photo copy HusseyLab used with permission) (C) An archival satellite tag and multiple mrPAT tags (Wildlife Comupters) mounted to the dorsalfin (photo copy HusseyLab used with permission) (D) A fin-mounted archival satellite tag (Wildlife Computers) and external marker tag (photo copy HusseyLab used withpermission) (E) Internal implantation of an acoustic tag (VEMCO) for static acoustic tracking (photo copy Kelsey Johnson used with permission) (F) A dorsalfin-mounted acoustic tag (VEMCO) for mobile acoustic tracking (photo copy Gregory Skomal used with permission)
(Stokesbury et al 2005 Gallant et al 2016) These tracksrecorded vertical movements and light-based depth preferencesindicating that the Greenland shark is not strictly a demersalspecies More recently acoustic tracking in the St LawrenceEstuary revealed that vertical movements into shallow andwarmer water coincided with the pre-dawn high tide (Gallantet al 2016) Archived depth records from pop-off archivalsatellite tags have also captured Greenland sharks swimming inthe water column in deep-water offshore regions of the Arcticand North Atlantic (Campana et al 2015a) These records
demonstrate a preference among Greenland sharks for greaterdepths at lower latitudes (mean time-weighted depth over 6 hwas 367 plusmn 4 m for Arctic sharks compared to 949 plusmn 10 mfor N Atlantic sharks) and revealed long-distance horizontalmovements including one shark that traveled 1015 km over125 days (Campana et al 2015a)
Past studies using biotelemetry to record Greenland sharkmovements have largely relied on two methods mobileacoustic tracking (Figure 3F) and archival satellite telemetry(Figures 3CD) two approaches that require researchers
Frontiers in Marine Science | wwwfrontiersinorg 11 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 12
Edwards et al Advancing Research for Long-Lived Species
to sacrifice either monitoring duration or data resolutionFor example active acoustic tracking can provide fine-scalehorizontal movement data but over limited timespans (hours ordays Skomal and Benz 2004 Stokesbury et al 2005 Gallantet al 2016) In contrast satellite tags can remain on animalsfor periods of up to several months (Stokesbury et al 2005Campana et al 2015a) but produce only straight-line trajectoriesof tagged animals (via extrapolation between the location ofanimal capture and the ARGOS location transmitted by satellitetags following their pre-programmed release) Knowledge ofthe locations and timing of movement pathways aggregationsites and areas of ecological importance for Greenlandsharks (eg feeding grounds and nurseries) therefore remainslimited Understanding the biological significance of movementbehaviors is consequently difficult to assess and will requirefurther study using available technologies and experimentaldesign innovations
The application of extensive passive acoustic telemetryinfrastructure (fixed receivers that detect tagged individualswithin their detection range Heupel et al 2005 Husseyet al 2015b) and long lifespan transmitters (sim10 yearsFigure 3E) provides one solution to examine the long-termmovements of Greenland sharks over a range of spatialscales By combining multi-year records of animal detectionswith environmental data (ice cover temperature salinityand dissolved oxygen) it will be possible to determine thespatiotemporal scales of predictable horizontal movementsSpecifically these data will delineate migration routes andactivity hotspots as well as the environmental factors drivingobserved behaviors The continued growth of a large-scalenetwork telemetry approach in the Arctic (Hussey et al 2015b)will promote collaborative data storage and handling and willrapidly increase data collection For example a telemetry networkestablished in Baffin Bay was designed by the Ocean TrackingNetwork (OTN) in collaboration with the Canadian fishingindustry and the Department of Fisheries and Oceans Canada(DFO) and is maintained by the joint efforts of all threegroups (Cooke et al 2011)
Innovative experimental designs including the novelapplication or combination of existing telemetry technologiescan also allow researchers to overcome some of the limitationsof tag design For example the attachment of multiple timed-release mark-report satellite tags (mrPATs Wildlife ComputersFigure 3C) to individual sharks has allowed the detectionof a potential migration pathway and revealed synchronicityin Greenland shark movements (Hussey et al 2018) Theintermediate location estimates provided by this approach(typically not captured by satellite tag studies on non-surfacinganimals) increased the resolution of recorded movementtrajectories and provided data which are key for identifyingsuitable locations for the deployment of future acoustic telemetryinfrastructure (Hussey et al 2018) Furthermore by combininga variety of telemetry technologies on individuals (eg satellitetags acoustic transmitters and receivers and accelerometersFigures 3AB) researchers can compare movement behaviors atdifferent data resolutions and timescales informing habitat useenvironmental conditions and intra- or interspecies interactions
while maximizing the data collected for each tagged individual(Holland et al 2009 Meyer et al 2010 Hussey et al 2018)
Despite rapid advances in telemetry technology andapplications capturing the ranges and movements of extremelylong-lived species will require continued improvementsin equipment design to increase tag longevity and themaximum depth ratings of telemetry equipment Technologicaldevelopments such as piezoelectric energy harvesting whichpowers battery-free acoustic tags using the flexing motions ofswimming fish (Li et al 2016) will alleviate the limitationsimposed by tag battery life Additionally mobile monitoringusing animal-borne transceivers or autonomous vehicles(gliders Lennox et al 2017) will increase the coverage oftelemetry studies and maximize potential data retrievalAcoustic transceivers (VEMCO Mobile Transceiver VMT) aretransforming animals into mobile receivers capable of detectingother tagged individuals (Lidgard et al 2012 Baker et al 2014Figure 3B) and have thus far been used to detect conspecificinteractions in remote habitats (Holland et al 2009) andseasonal patterns in group dynamics (fissionndashfusion behaviorHaulsee et al 2016) However these methods currently requirethe animal to be recaptured to recover the tag and associateddata The development of Bluetooth technology linking VMTsand pop-off archival satellite tags (following the approach ofLidgard et al 2014) will allow the remote transfer of animaldetection data from VMTs to ARGOS satellites eliminatingthe need for tag retrieval This will thereby reveal interactionsbetween Greenland sharks and other tagged animals providinginsight into their feeding and predatory behaviors (see sectionldquoBehaviorrdquo) Further advancement of archival satellite tagsincluding the recording of high quality oceanographic data andfaster data transmission to ARGOS satellites (increasing thevolume of data transmitted and received) will also improve ourunderstanding of environmental factors influencing the spatialhabitat use of Greenland sharks
Understanding the seasonal horizontal movements andspatial habitat use of Greenland sharks is critical for marinespatial planning (eg ATLAS case study for Davis StraitEastern Arctic) Large-scale movements recorded by acousticandor satellite telemetry (tied with genomic data to identifypopulation units see section ldquoPopulation Genetics andGenomicsrdquo) will help to refine our understanding of broad-scaleGreenland shark distributions allowing for the establishmentof bilateral or multilateral agreements between nations or viaRegional Management Organizations (RFMOs see sectionldquoManagementrdquo) to manage at the population level (Table 1)At a finer scale telemetry approaches will allow high-useor hotspot areas to be identified and predictable movementpatterns to be defined These data along with the distributionsof potential threats (such as fisheries activity) can be integratedinto predator-prey models (McClellan et al 2009) which willindicate the spatial and temporal extent of regions that shouldbe avoided or where fishing should be time-restricted duringperiods of increased Greenland shark presence If necessary thisinformation would allow the implementation of precautionarymeasures such as Dynamic Area Management (DAM NOAA2002) to reduce the likelihood of excessive bycatch (Table 1)
Frontiers in Marine Science | wwwfrontiersinorg 12 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 13
Edwards et al Advancing Research for Long-Lived Species
Developing an understanding of the environmental driversaffecting Greenland shark movements will also improveour ability to predict how these patterns will be modifiedunder various climate change scenarios further directingthe implementation of precautionary measures Systematicconservation planning using spatial prioritization software toolssuch as Marxan (Ball et al 2009) will provide an approachto integrate these data to produce direct management advice(Metcalfe et al 2015 Table 1)
BehaviorTo date direct observations of Greenland shark behaviorare extremely limited Our understanding of their foragingmechanisms and predatory capabilities are largely based oncombined evidence from movement records (see sectionldquoMovement Ecologyrdquo) and dietary analyses (see section ldquoDietand Trophic Ecologyrdquo) A single study on the three-dimensionalmovements of Greenland sharks recorded by high-resolutiondata loggers reported the slowest observed mean swim speed(034 msminus1) and tailbeat frequency (015 Hz) relative to size forany fish species leading researchers to question how they mightcatch much faster-moving prey (Watanabe et al 2012) Whilescavenging behavior has been well documented (eg inertialsuction feeding from a demersal pot Grant et al 2018 photoevidence of Greenland sharks feeding on carrion at the surfaceMacNeil et al 2012 see section ldquoDiet and Trophic Ecologyrdquo)direct observations of active predation have yet to be recordedDespite this fact several lines of evidence including freshlyingested seal remains in Greenland shark stomachs and bitemarks observed on live seals and overwintering beluga whales(Delphinapterus leucas) suggest that active predation on fast-moving prey like marine mammals does occur (Fisk et al 2002Leclerc et al 2012 MacNeil et al 2012 Nielsen et al 2014)Members of the Inuit community of Pangnirtung (NunavutCanada) have also documented Greenland sharks scavengingseals and beluga from nets and have reported their belief in thesharkrsquos ability to capture and consume newborn seal pups (basedon whole specimens found in shark stomachs) and to attack adultseals visiting breathing holes in the ice (from observations ofbite wounds Idrobo and Berkes 2012) Given their observedslow swimming speed it is hypothesized that sharks may adopta stealth approach to target seals when they rest underwater or atthe surface (Skomal and Benz 2004 Watanabe et al 2012)
With the vision of Greenland sharks in the Arctic consideredto be impaired by copepod parasites (Ommatokoita elongataBerland 1961 Borucinska et al 1998 Skomal and Benz 2004Figure 4B) and their slow swim speeds (Watanabe et al2012) the mechanism of active predation remains unknownConsequently the primary role of this species as a predator orscavenger of marine mammals has not been established (Bigelowand Schroeder 1948 Ridoux et al 1998 Lucas and McAlpine2002 Horning and Mellish 2014) despite the importance of thisinformation to our understanding of their true ecological role
Another key question concerns the feeding frequency ofGreenland sharks particularly in relation to their metabolic rateand energy budgets Given their large body mass ectothermicphysiology and cold water habitat these sharks are likely to
have extremely low mass-specific metabolic rates as a result ofslow growth long life spans (Nielsen et al 2016) and slowswimming speeds (Watanabe et al 2012) Seals provide a highlyenergy-rich food due to their large body size and high fat content(Stirling and McEwan 1975 Addison and Stobo 1993 Lucas andNatanson 2010) It is possible therefore that Greenland sharkscould survive for several months without feeding following theconsumption of such energy-dense prey The consumption rateof teleost prey (eg Atlantic cod Gadus morhua or Greenlandhalibut Nielsen et al 2014) by Greenland sharks may also bemuch lower than that of other large sharks (time scale of daysrather than hours)
The frequent vertical movements exhibited by Greenlandsharks raise additional questions over the physiological andmetabolic costs of inhabiting and transitioning between shallowand deep-water environments and the factors influencing suchmovements Deep-sea sharks rely on a high liver mass to provideenergy storage for ovary production during maturation howeverat shallow depths this large proportion of fatty tissue (relativeto body mass) results in positive buoyancy potentially leading toa higher energetic requirement during descent (Nakamura et al2015) Further insight into the energetic costs of these movementsmay help to resolve questions regarding the frequency of feedingand quantity of prey intake required to maintain them
To address questions surrounding feeding mechanisms andfrequency direct observation of interactions and huntingbehavior using animal-borne video cameras (eg Shark Tagcamera Kukulya et al 2015) or other technologies (egactive sonar) are needed In recent years telemetry advanceshave increasingly allowed indirect observations of animals inremote environments (Hussey et al 2015b) For instanceactive predation of pinnipeds by Pacific sleeper sharks wasinferred from post-mortem temperature records collected bydual life history tags (LHX tags) implanted in Stellar sealions (Horning and Mellish 2014) Camera-bearing AUVssuch as Remote Environmental Monitoring UnitS (REMUS)have provided unique observations of the feeding behaviorof white sharks in situ (Skomal et al 2015) and could beeasily adapted to monitor Greenland sharks Several othertechnologies could provide indirect methods of recordingGreenland shark interactions with conspecifics and potentialprey For example through the deployment of hydrophonesthat record ambient noise on sharks (D-tag or AcousondeOleson et al 2010) it may be possible to assess sharkproximity to vocalizing marine mammals (sperm whale Physetermacrocephalus narwhal Monodon monoceros and ringed sealPusa hispida) In addition recoverable VMT packages orsatellite-linked units deployed on Greenland sharks (see sectionldquoMovement Ecologyrdquo) could provide data on co-occurrence andthe potential rate of interactions among tagged sharks andbetween sharks and potential prey species equipped with acoustictags (eg Greenland halibut Arctic skate Amblyraja hyperboreaand narwhal Broell et al personal communication)
The combined use of accelerometers and animal-bornecameras on individual sharks provides one possible approach toexamine the cost of vertical movement and determine the feedingfrequency of Greenland sharks For example following the use
Frontiers in Marine Science | wwwfrontiersinorg 13 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87
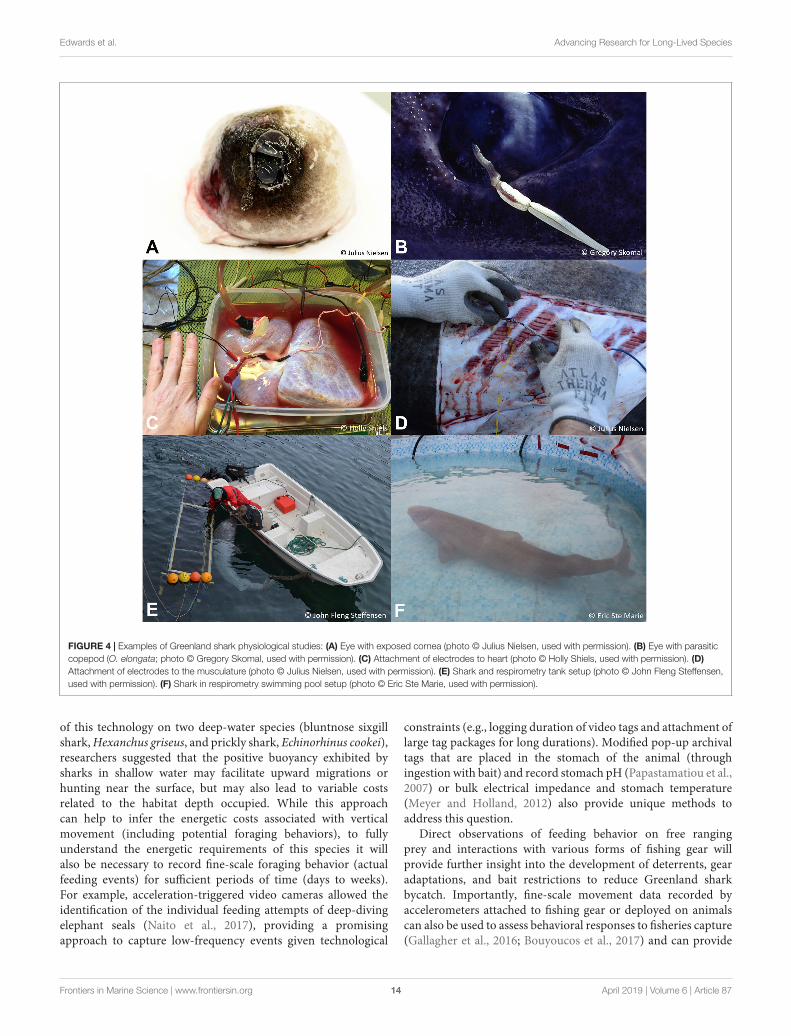
fmars-06-00087 March 29 2019 Time 1853 14
Edwards et al Advancing Research for Long-Lived Species
FIGURE 4 | Examples of Greenland shark physiological studies (A) Eye with exposed cornea (photo copy Julius Nielsen used with permission) (B) Eye with parasiticcopepod (O elongata photo copy Gregory Skomal used with permission) (C) Attachment of electrodes to heart (photo copy Holly Shiels used with permission) (D)Attachment of electrodes to the musculature (photo copy Julius Nielsen used with permission) (E) Shark and respirometry tank setup (photo copy John Fleng Steffensenused with permission) (F) Shark in respirometry swimming pool setup (photo copy Eric Ste Marie used with permission)
of this technology on two deep-water species (bluntnose sixgillshark Hexanchus griseus and prickly shark Echinorhinus cookei)researchers suggested that the positive buoyancy exhibited bysharks in shallow water may facilitate upward migrations orhunting near the surface but may also lead to variable costsrelated to the habitat depth occupied While this approachcan help to infer the energetic costs associated with verticalmovement (including potential foraging behaviors) to fullyunderstand the energetic requirements of this species it willalso be necessary to record fine-scale foraging behavior (actualfeeding events) for sufficient periods of time (days to weeks)For example acceleration-triggered video cameras allowed theidentification of the individual feeding attempts of deep-divingelephant seals (Naito et al 2017) providing a promisingapproach to capture low-frequency events given technological
constraints (eg logging duration of video tags and attachment oflarge tag packages for long durations) Modified pop-up archivaltags that are placed in the stomach of the animal (throughingestion with bait) and record stomach pH (Papastamatiou et al2007) or bulk electrical impedance and stomach temperature(Meyer and Holland 2012) also provide unique methods toaddress this question
Direct observations of feeding behavior on free rangingprey and interactions with various forms of fishing gear willprovide further insight into the development of deterrents gearadaptations and bait restrictions to reduce Greenland sharkbycatch Importantly fine-scale movement data recorded byaccelerometers attached to fishing gear or deployed on animalscan also be used to assess behavioral responses to fisheries capture(Gallagher et al 2016 Bouyoucos et al 2017) and can provide
Frontiers in Marine Science | wwwfrontiersinorg 14 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 15
Edwards et al Advancing Research for Long-Lived Species
evidence of post-release recovery and survival following captureand release (Brownscombe et al 2013 Table 1) This method(using time-series vertical dive data recorded by animal-borneaccelerometers) is currently being used to determine survivalrates and the time required for Greenland sharks to returnto normal behaviors following release from capture by bottomlonglines (Watanabe et al personal communication)
Physiological AdaptationsDue to the Greenland sharkrsquos large size and remote habitat whichhave precluded sophisticated in vivo experimentation and captivestudies many studies are conducted under field conditions thatfavor experiments on isolated tissues rather than whole animalsAs a result the basic physiology of the Greenland shark remainslargely a mystery (MacNeil et al 2012 Herbert et al 2017)
While the cardiovascular system underlies every facet of ananimalrsquos life relatively few studies have examined this in theGreenland shark A recent study on the whole blood of Greenlandsharks noted that their blood properties (high oxygen affinity andlow Bohr effect) were typical of sluggish elasmobranchs (Herbertet al 2017) However using isolated haemoglobins the bloodwas found to have a relatively low affinity and consist of threehemoglobin isoforms with no functional differences (Russo et al2017) This analysis suggested that cellular changes in allostericeffectors may be important for controlling the O2 transportproperties of the blood Additionally in an investigation of thestress response of Greenland sharks caught on bottom longlinescapture-induced changes in blood glucose and lactate values werefound to be variable and were weakly related to capture depth(lactate) and body length (glucose Barkley et al 2017)
Characteristics of the Greenland sharkrsquos heart and bloodvessels are also beginning to emerge Greenland sharks werefound to have a low intrinsic heart rate paired with a highventricular volume (measured ex vivo Shiels et al 2018) (Figure4C) Histological analysis of heart samples showed significantfibrosis that increased with age a preliminary analysis revealedno evidence of atherosclerosis Efforts to quantify details of heartperformance using isolated cardiac heart strips (Larsen et al2017) have proved to be largely unsuccessful as the tissue doesnot remain viable for long periods In heart strips that appearedto be contracting normally a single contractionrelaxation cycle(ie a twitch) required 3ndash5 s to complete at 5C suggestingthat maximal heart rate ranges from 12 to 20 beatsmin (inagreement with Shiels et al 2018) While blood pressure inGreenland sharks has never been directly measured it hasbeen estimated by analyzing the relative amounts of elastinand collagen in the wall of the ventral aorta and measuring itscompliance characteristics over a range of pressures (Shadwicket al 2018) These data suggest that the Greenland sharkrsquosaverage blood pressure is approximately 23ndash28 kPa muchlower than other slow swimming sharks such as the epauletshark Hemiscyllium ocellatum (39 kPa) or catshark Scyliorhinuscanicula (53 kPa) where it has been directly measured (Tayloret al 1977 Speers-Roesch et al 2012)
Further physiological studies have largely focused on systemswhich are likely to play an important role in foraging behaviorsOlfactory cues are typically important for locating prey especially
where visual cues are limited An anatomical study of theolfactory rosette in the Greenland shark indicated that while thearrangement of the olfactory lamellae and epithelium are similarto those found in benthicslow swimming animals the relativelylarge olfactory epithelium surface area is more reflective of abentho-pelagic animal (Ferrando et al 2016)
Preliminary biochemical assays and work loops (force vslength curves used to determine the mechanical work of musclefibers) indicate that both red and white skeletal muscle fibershave very low metabolic capacities and contraction (twitch) rates(Bernal personal communication) This suggests that they areunable to sustain high levels of either anaerobic or aerobic musclecontraction for extended periods These data consequently raisequestions over how Greenland sharks actively forage in the watercolumn for teleosts and marine mammals (Fisk et al 2012MacNeil et al 2012 Nielsen et al 2014 see section ldquoBehaviorrdquo)
While sharks in general have good vision little is knownabout sharksrsquo visual adaptations to their environment andhow this is impacted by their ecology and habitat (McCombet al 2010) Greenland sharks are thought to have poorvision potentially as a result of corneal damage producedby the attachment of the parasitic copepod O elongata(Berland 1961 Borucinska et al 1998 Figure 4B) as wellas the dark environment they inhabit (300ndash500 m Nielsenet al 2014) In the absence of active parasitism sharksmay still possess scar tissue on the cornea from previousinfections however no evidence of a correlation betweenshark sizeage and degree of corneal damage has yet beenrecorded (Nielsen personal communication) Additionally whilea high incidence of copepod parasitism has been reported athigh latitudes in the Eastern Canadian Arctic and off easternand western Greenland (sim90 of sharks showing parasitismSteffensen personal communication) a much lower incidenceof active parasitism has been recorded in the St LawrenceEstuary and in waters off southern Atlantic Canada (lt10of observed sharks Gallant personal communication and noparasitized sharks observed Campana personal communicationrespectively) Despite this observed spatial disparity in copepodpresenceabsence the degree of corneal damage present in sharksfrom these discrete locations has yet to be examined thoroughlySystematic investigations of this disparity and of the cumulativeeffects of multiple parasitic infections on corneal structureand vision are thereby required Furthermore novel recordsof vision-associated behaviors (defensive posturing) performedby non-parasitized sharks in shallow waters (St LawrenceEstuary Harvey-Clark et al 2005) and evidence to suggestthat Greenland sharks are potential active predators (Watanabeet al 2012 Nielsen et al 2014) further warrant a thoroughunderstanding of their visual system
Despite the limitations precluding in vivo experimentationfuture anatomical studies and in vitro experiments will leadto important insights with regards to whole animal functionFor example studies directed at defining skeletal musclecharacteristics (fiber type distribution contraction rates andpH buffering properties Figure 4D) and cardiorespiratorycharacteristics (eg heart rate stroke volume blood pressureventilation rate ventilation volume extraction efficiency and
Frontiers in Marine Science | wwwfrontiersinorg 15 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 16
Edwards et al Advancing Research for Long-Lived Species
blood buffering capacity Figure 4C) will be important in buildinga complete picture of the Greenland shark as a top predatorin the Arctic ecosystem Understanding these physiologicalattributes is also key for improving knowledge of the mechanismsdriving longevity
Established in vitro techniques will also facilitate futureexamination of the visual system in Greenland sharks(Figure 4A) Approaches such as electroretinography could beused to determine spectralluminous sensitivities and temporalresolution (Kalinoski et al 2014) while microspectrophotometrycan elucidate the type and distribution of visual pigments (Hartet al 2005) Of particular interest given the long-livednature of this species is the study of ontogenetic changesin brain organization that might occur over 200 yearspotentially altering sensory acuity and therefore the relativeimportance of senses such as vision and olfaction over individuallifespans (Lisney et al 2017)
Further study of the importance of olfaction andelectroreception for prey location will provide insight intothe foraging behavior of Greenland sharks including factorsaffecting incidental capture by fishing gear Specifically age-related changes in the olfactory epithelium could be examinedgiven documented changes in other species (Ferrando et al2016) Greenland sharks also appear to have a well-developednetwork of electroreceptors (Ampullae of Lorenzini) that allowthem to detect the bio-electrical activity of animals at closerange (Kalmijn 1971) but their arrangement has yet to bedescribed Given the lack of response exhibited by Greenlandsharks presented with electropositive metals (Grant et al 2018)further study of the sensitivity of their electroreceptors may bebeneficial for the development of effective hook deterrents tomitigate bycatch
Understanding the role of the Greenland shark as a potentialapex predator also requires accurate measurement of itsmetabolic rate and scope (aerobic and anaerobic) during restingswimming and digestion cycles (Figures 4EF) In additionto establishing rates of energy expenditure for these activitiesmetabolic rate has also been shown as a determinant of lifespan based on temperature and body size (Gillooly et al 2001)Preliminary studies on sim250 kg Greenland sharks found that theydid not have an unusual metabolic rate (Steffensen et al personalcommunication Figure 4E) compared to what could be expectedgiven its large size and body temperature More metabolic studiesare warranted particularly comparing the resting metabolic rateof starved vs fed sharks to evaluate the time course and energyexpenditure associated with digestion (Specific Dynamic ActionSDA Sims and Davies 1994 Ferry-Graham and Gibb 2001)
A greater understanding of these physiological traits andenergetic requirements will ultimately lead to improved speciesmanagement for both Greenland sharks and their commerciallytargeted prey species Specifically there is a fundamentalneed for baseline (unstressed) values for virtually all bloodparameters from Greenland sharks and for the accuratequantification of capture stress Furthermore understanding thebioenergetic requirements of individual Greenland sharks (viafield measurements of metabolic rate and direct observations ofconsumption rate see section ldquoBehaviorrdquo) will help to estimate
the energetic needs of sharks at the population level This couldbe crucial for informing harvest levels (eg Total AllowableCatches TAC) of targeted Greenland shark prey species toensure that the remaining biomass is resilient to sustainedfishing effort (Table 1)
Diet and Trophic EcologyThe diet of Greenland sharks includes an impressive diversity ofprey (Yano et al 2007 McMeans et al 2010 Leclerc et al 2012Nielsen et al 2014 Nielsen 2017) For example stomachs of39 Greenland sharks from western Greenland waters contained25 different fish species at least 3 pinniped species and severalgroups of invertebrates (molluscs echinoderms decapods Yanoet al 2007) Live prey capture is possible (see section ldquoBehaviorrdquo)however scavenging of marine mammal carrion (Williamson1963 Beck and Mansfield 1969 Leclerc et al 2011) andcannibalism of conspecifics captured in fishing gear (Jensen1948 Nielsen et al 2014) are known to occur Several reportsindicate that Greenland sharks consume both benthic and pelagicprey (MacNeil et al 2012) and the simultaneous occurrence ofboth prey types in the diet of Greenland sharks is consistent withdiurnal behavior observed using biotelemetry (Campana et al2015a) Spatial variations in the relative importance of pelagicprey (ie a higher proportion of pelagic fishes in shark stomachsfrom Iceland vs Canadian Arctic) have also been reportedhowever this may reflect differences in sampling method (gillnetand trawl vs bottom longline) or LT (mean plusmn SE 416 plusmn 25284 plusmn 44 cm respectively Fisk et al 2002) rather than truedietary variations (MacNeil et al 2012) Stable isotopes fattyacids and biomagnifying contaminants confirm a high trophicposition and consumption of diverse prey types (Fisk et al 2002McMeans et al 2010 2015 Hussey et al 2014) Geographicalvariation in diet further suggests that Greenland sharks exhibit aflexible response to variation in prey availability (McMeans et al2013a Nielsen et al 2014) However some prey selectivity mayoccur because some abundant fish species (eg redfish Sebastesspp) are rarely present in shark stomachs from Greenland waters(Nielsen et al personal communication) Stomach contents andbiochemical markers also provide evidence that larger sharksconsume more teleost and marine mammal prey comparedto smaller sharks (less than 200 cm LT Yano et al 2007McMeans et al 2013a Nielsen 2017) which have been foundto feed predominantly on lower trophic level prey such as squid(Nielsen 2017)
Greenland sharks are mobile opportunistic top predatorsthat obtain and couple energy from multiple habitats within anecosystem (McMeans et al 2013b) and across geographicallydistant ecosystems (see section ldquoMovement Ecologyrdquo) Althoughtheir effect on food web dynamics has yet to be quantifiedexisting food web theory suggests that mobile opportunistictop predators are important for food web stability (McCannet al 2005 Rooney et al 2006) Empirical examples for thistheory include Atlantic cod (G morhua) which are thoughtto stabilize the ecosystem by preventing oscillatory overshootdynamics in their prey (Frank et al 2011) Given their broaddistribution and potential capacity for active predation (seesection ldquoBehaviorrdquo) Greenland shark populations could play
Frontiers in Marine Science | wwwfrontiersinorg 16 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 17
Edwards et al Advancing Research for Long-Lived Species
a similar role in marine food webs despite expected slowmetabolism and low consumption rates (see section ldquoBehaviorrdquo)While it has been suggested that Greenland sharks may be asignificant source of mortality for some seal populations (Leclercet al 2012) the effects of consumption by Greenland sharks onprey populations must be examined further This is of particularimportance as facultative scavenging may amplify the top-downtrophic effects exerted by Greenland sharks on their food webbecause high-quality carrion can inflate predator biomass andincrease a predatorrsquos capacity for prey control (eg in wolvesWilson and Wolkovich 2011) As both potential predators andscavengers of many prey taxa Greenland sharks could thereforehave widespread effects on food webs throughout the Arcticand North Atlantic A lack of evidence to define potential sub-population dietary specialization as well as further uncertaintysurrounding the role of extreme longevity in nutrient and energytransfer in marine food webs also indicate the potential value ofmore detailed study of the trophic role of Greenland sharks inArctic marine environments
Future work will need to quantify the type (active predationvs scavenging) and frequency of trophic interactions betweenGreenland sharks and their prey (Table 1) Video observationsand tracking technology (see section ldquoBehaviorrdquo) combinedwith non-lethal sampling of tissues (blood and muscle)for dietary analysis (stable isotopes amino acid compoundspecific isotope analysis and fatty acids) will allow a morecomprehensive understanding of Greenland shark diet andtheir functional role within food webs without the needto sacrifice individuals Through a combination of accurateestimates of diet field measurements of consumption andmetabolic rates (see sections ldquoBehaviorrdquo and ldquoPhysiologicalAdaptationsrdquo respectively) detailed movement behaviors (seesections ldquoMovement Ecologyrdquo and ldquoBehaviorrdquo) and abundanceestimates (see section ldquoDemographics and Life Historyrdquo) itwill be possible to quantify energy flow and trophic transferefficiencies shedding light on Arctic food web dynamics (egbiomass structure ndash pyramidinverted pyramid and interactionstrengths) that may prove important for understanding past andfuture food web stability (McCann et al 2005) Geographicalcomparisons of biomass structure (the distribution of biomassacross trophic levels) could be undertaken to study howvariation in Greenland shark abundance regulates these foodwebs A particularly exciting possibility is to synthesize foodweb data (ie predator prey relationships and abundance datathrough time) which can then be used to estimate indices ofstability (eg the coefficient of variation of prey populationdynamics Korpimaumlki 1984) Such an effort would providequantitative data on how Greenland shark density and foragingbehavior (eg the extent of cross habitat foraging) influencesfood web dynamics
Ecosystem models (EcoPath and EcoSim) can help predict theeffects of various fisheries management decisions (and associatedchanges in fishing pressure) on marine food webs (Travers et al2010) Through these models diet and abundance metrics canbe used to predict the potential effects of changes in Greenlandshark abundance on prey species (including those affected bycommercially harvest eg Greenland halibut) Bycatch limits
for Greenland sharks could then be set at levels to ensure thatthe remaining shark biomass is sufficient to maintain ecosystemfunction while also maintaining adequate prey biomass (Table 1)
Threats to Greenland Shark PopulationsDespite occupying great depths and remote polar regionsGreenland sharks experience many of the same anthropogenicand climatic stressors as other long-lived marine species (egfisheries vessel traffic seismic surveys pollution and climatechange Clarke and Harris 2003 Halpern et al 2008 Daviset al 2013) This may be particularly true for sharks inhabitingcoastal regions of the North and Norwegian seas and theNorth American eastern seaboard areas that are predicted tobe cumulatively impacted by numerous anthropogenic driversof ecological change (Halpern et al 2008) Importantly giventheir K-selected nature impacted Greenland shark populationswill likely be slow to recover
Of these threats fisheries have the greatest impact onGreenland shark populations Directed fisheries for Greenlandsharks existed in Norway Iceland and Greenland from the 1600sto the mid-1990s (Davis et al 2013) Catch rates grew to 15000Greenland sharks per year by the end of the 18th century witha peak catch of 32000 individuals in Greenland in 1910 (Daviset al 2013) Greenland shark catches continue at much lowerlevels today (see section ldquoDemographics and Life Historyrdquo) Thisincludes Greenland shark bycatch in fisheries targeting otherspecies particularly Greenland halibut or shrimp which occurredduring the period of directed fishing and have continued since(Davis et al 2013) Bycatch is typically returned to the oceanbut the proportion of animals that survive varies with fishinggear type fishing duration and handling method A recentexamination of Greenland shark bycatch in Canada found that36 of sharks caught in bottom trawls (Greenland halibutand northern shrimp fisheries) and 16 of sharks caught onbenthic longlines (Greenland halibut fisheries) were already deadwhen released (North Atlantic Fisheries Organization [NAFO]2018bc) and the percentage of dead sharks increased with trawlduration (North Atlantic Fisheries Organization [NAFO] 2018c)
The threats posed by incidental capture are not limited topost-release mortality Like many elasmobranchs Greenlandsharks are often viewed as competitors for commercially valuablespecies by harvesters For example Greenland sharks that feedfrom and are incidentally captured by bottom longlines canbecome entangled in the fishing gear and rolling behavior canresult in the mainline becoming tightly wrapped around thecaudal peduncle complicating release efforts (Edwards personalobservation Figures 5AB) For this reason Greenland sharksare often considered a nuisance species in commercial fisheriesand entangled individuals may be removed by severing the caudalfin (Davis et al 2013) Historically Greenland sharks in theSaguenay Fjord were hauled out as trophies but now they must bereleased while still in the water This law has been strictly enforcedby the Department of Fisheries and Oceans Canada since 2006(Gallant personal communication)
Even in the absence of directed fisheries the rate ofinteractions among Greenland sharks and commercial fishinggear can be expected to increase in coming years Ongoing
Frontiers in Marine Science | wwwfrontiersinorg 17 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 18
Edwards et al Advancing Research for Long-Lived Species
FIGURE 5 | Incidental capture of Greenland sharks via scientific bottom longline (A) Shark entangled in mainline around caudle peduncle (B) Shark hooked bymouth (photos Jena Edwards)
reductions in sea ice extent and duration in the Arcticand changes in the composition and productivity of Arcticmarine communities (Clarke and Harris 2003) are allowingexisting fisheries (bottom longline trawl and gillnet) to expandnorthward into previously unfinished habitats and extend theirfishing season while also creating the potential for new fisheriesto develop (Christiansen et al 2014)
In addition to fisheries pressures Greenland sharks can alsobe affected by chronic and acute changes in their environmentfrom pollution increased noise and climate change factorswhich are likely to affect most fish and marine mammals in theArctic (Clarke and Harris 2003 Huntington 2009) Pollutionfrom global atmospheric deposition coastal communities andvessels can affect Greenland sharks directly by impacting theirphysiological balances (eg heavy metals hormones toxins Stridet al 2007 2010 Corsolini et al 2014) or indirectly by alteringecosystem productivity which can lead to eutrophication andreduced oxygen availability (eg St Lawrence hypoxic dead zoneBelley et al 2010 Howarth et al 2011) Alongside naturalsounds from animals tides and currents anthropogenic noisefrom vessels and industrial development are increasing in theArctic with unknown effects (Ivanova et al 2018)
Mirroring this extensive list of potential threats to Greenlandshark populations is a series of questions that must be addressedIn the absence of population size or demographic data it iscurrently not possible to estimate the impacts of historicalfisheries on Greenland shark populations (size age sex ormaturity trends) or to examine changes in population metrics(eg recovery rate) following the cessation of directed fishing Inaddition when examining the available commercial bycatch datait is important to consider potential inaccuracies Large animals
such as Greenland sharks are typically not weighed Insteadbycatch records are generally limited to counts and may includesupplementary data such as individual lengths sex and status(alive or dead) When weights are recorded they are often a bestguess based on the experience of the observer at estimating theweight of fish in a net
There is also insufficient data to accurately estimate post-release mortality rates for bycaught Greenland sharks and todetermine the effects of key factors such as gear type Whilepost-release mortality rates of up to 50 have been recordedin other sharks (as measured with PSATs Campana et al2015b) this is known to vary widely across species Additionaldata will therefore be required to accurately estimate the effectsof incidental capture on Greenland shark populations anddetermine handling practices that will ensure optimum survivalrates (see section ldquoBehaviorrdquo)
While the precise impacts of indirect threats such as noisepollution and seismic surveys on Greenland shark behaviorand physiology have not been quantified sharks may need toadapt to changes in prey availability in areas where seismicsurveys are conducted Vessel noise can alter both short andlong-term habitat use decisions by animals affecting overallspecies distributions (Saragrave et al 2007 Slabbekoorn et al 2010McCarthy et al 2011 Ivanova et al 2018) For exampleresearch in the Barents Sea has shown that seismic shootingcan significantly affect fish distribution local abundance andcatch rates (Engarings et al 1996) High intensity sounds fromseismic surveys or pile driving could also have more directimpacts on individuals causing sensory damage (and potentiallydeath) leading to avoidance behaviors and changes in habitat use(Popper and Hastings 2009)
Frontiers in Marine Science | wwwfrontiersinorg 18 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 19
Edwards et al Advancing Research for Long-Lived Species
Scientific efforts to increase our understanding of themany threats faced by Greenland shark populations and theirsurrounding ecosystems are underway In an effort to improvethe quality of data obtained from Greenland shark bycatch DFOrequested on a trial basis that at-sea observers use a statisticallength-weight key to estimate Greenland shark weight based on ameasured length along with criteria to assess condition
Changes to fishing gear (Grant et al 2018) and practicessuch as limiting set duration (Cosandey-Godin et al 2014)have been examined as methods to reduce Greenland sharkbycatch mortality However testing of additional bycatchmitigation strategies (eg gear modifications such as modifiedmono-filament gangions) as well as the data required tomonitor the status of Greenland shark populations (abundanceandor biomass indices size and sex ratios see sectionldquoDemographics and Life Historyrdquo) will also be necessary formaintaining sustainable mortality levels Bycatch records andvideo observations of Greenland sharks interacting with fishinggear (Grant et al 2018) are invaluable for adapting gear tominimize the probability of shark capture or entanglementand to allow monitoring of the frequency of interactionsSimilarly data from telemetry and archival tags are revealing thehorizontal and vertical movement patterns of Greenland sharks(see section ldquoMovement Ecologyrdquo) which can be used to adaptfishing practices (eg harvester avoidance of high use areasDAM plans NOAA 2002)
Continuing to articulate the link between threats andGreenland shark population(s) status in the scientific literatureis useful for fishery management and marine spatial planningFurthermore regular communication of Greenland sharkresearch results to managers stakeholders listing organizationsand the general public is crucial for raising awareness ofthe role of Greenland sharks in Arctic ecosystems and fordrawing attention to the need for their effective managementThis can be achieved through various forms of outreachincluding publicationsreports presentations media (socialradio print and television) community posters studentengagement and music videos
Fisheries managed by Arctic countries NAFO and ICESapply the precautionary approach to fishery management (Foodand Agricultural Organization [FAO] 1996 North AtlanticFisheries Organization [NAFO] 2004 Department of Fisheriesand Oceans [DFO] 2006 2009 ICES 2012) While full stockassessments are typically limited to species that are commerciallyharvested a similar level of analysis and management forkey bycatch species is being considered in some jurisdictions(Department of Fisheries and Oceans [DFO] 2013) andshould be considered for the Greenland shark Ecosystem-basedfishery management (EBFM) involves managing fisheries withconsideration of supporting ecosystem components and impactson marine habitats (eg physical disruption and soundscape) andcommunities (eg direct mortality competition with predators)in addition to commercial harvests and stock status (Pikitch et al2004 Trochta et al 2018) Incorporating Greenland sharks intoEBFM frameworks would require an accurate understanding ofthe sharkrsquos role in the ecosystem and a model of communitytrophic linkages data which are not currently available EBFM
also includes environmental drivers of ecosystem and animalproductivity (ie to rebuild and sustain populations speciesbiological communities and marine ecosystems at high levelsof productivity and biological diversity while providing foodrevenue and recreation for humans US National ResearchCouncil 1998) The full application of the EBFM approach tofisheries with Greenland shark bycatch will therefore require newinformation regarding the nature and magnitude of each driverrsquoseffects both in isolation and in combination Marine spatialplanning that accounts for potential impacts on Greenland sharkswill require the same data and analyses to be effective In themeantime multi-species stock assessment models provide oneoption to incorporate Greenland shark management actions intocurrent directed fisheries management plans (Moffitt et al 2016)Alternatively trophic pyramid modeling provides a method inwhich catch limits are applied to trophic levels rather thanindividual species in an effort to reduce a fisheryrsquos impacts onoverall community structure (Garcia et al 2012 2014 Froeseet al 2015 Burgess et al 2016) Results from such modelingexercises can inform management decisions and be incorporatedin fishery-level documents such as the Integrated FisheryManagement Plans that are developed for Canadian fisheries (seesection ldquoManagementrdquo) Fishery-level documents can then bereferenced by and linked to marine spatial planning initiativesat national and international levels to achieve integration amongmanagement sectors and activities
ManagementLaws and policies relevant to Greenland shark management occurat global regional and national levels
GlobalA fragmented array of global agreements and guidancedocuments have emerged to promote sustainable fisheriesand marine biodiversity conservation in general (Russell andVanderZwaag 2010 Techera and Klein 2017) but shark-specificlaw and policy developments are limited to four main avenues(Davis et al 2013) First various shark species have been listed asendangered or threatened under the Convention on InternationalTrade in Endangered Species of Wild Fauna and Flora (CITES)with resultant international trade restrictions imposed throughexport and import permit requirements (Hutton and Dickson2000 CITES nd) The Greenland shark has not been consideredfor CITES listing due to its conservation status being categorizedas near threatened under the IUCN Red List of ThreatenedSpecies (IUCN 2017) and lack of trade concerns
Second under the Convention on the Conservation ofMigratory Species of Wild Animals (CMS) a Memorandum ofUnderstanding on the Conservation of Migratory Sharks (SharksMOU) has been adopted that encourages signatories to applyprecautionary and ecosystem approaches to shark conservation(CMS 2016) A Conservation Plan included in Annex 3 to theSharks MOU sets out numerous implementation actions thatshould be prioritized by signatory states including improvingmigratory shark research and monitoring strengthening controlsover directed and non-directed shark fisheries ensuring theprotection of critical habitats and migratory corridors and
Frontiers in Marine Science | wwwfrontiersinorg 19 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 20
Edwards et al Advancing Research for Long-Lived Species
increasing public awareness of the threats to sharks (CMS 2016)However the MOU has limited application and is only applicableto 29 listed shark and shark-related species which does notinclude the Greenland shark (CMS 2018a) Additionally theMOU only has 48 signatories (CMS 2018b) which do not includekey Arctic states such as Canada Iceland Norway and theRussian Federation (CMS 2018b)
Third the Food and Agriculture Organization (FAO) hasadopted an International Plan of Action for the Conservationand Management of Sharks (Food and Agricultural Organization[FAO] 1999) which encourages states to adopt national plansof action for the conservation and management of sharks indirected or non-directed fisheries Such plans often seek tominimize the unutilized incidental catch of sharks contributeto the protection of biodiversity and ecosystem structureand function and facilitate improved species-specific catchand landings data States are further encouraged to assessimplementation of national plans at least every 4 years and toconsider where appropriate the development of sub-regional orregional shark plans
A fourth global avenue of specific shark-related law andpolicy development is through the UN General Assembly andits adoption of annual resolutions on sustainable fisherieswhich include paragraphs urging states and RFMOs (see sectionldquoMovement Ecologyrdquo) to better protect sharks For examplethe sustainable fisheries resolution (Res 7272) adopted inDecember 2017 urges states to take multiple science-basedmeasures to manage shark stocks including setting limits oncatch or fishing efforts requiring the reporting of species-specificdata on discards and landings undertaking comprehensivestock assessments of sharks and reducing shark bycatch andbycatch mortality Regional Fisheries Management Organizations(RFMOs) with competence to regulate highly migratory speciesare urged to strengthen or establish precautionary science-basedconservation and management measures for sharks
RegionalA major regional development relevant to Greenland sharkconservation was the conclusion in November 2017 of a draftAgreement to Prevent Unregulated High Seas Fisheries in theCentral Arctic Ocean (US Department of State 2017) TheAgreement expected to be adopted by the five Arctic Oceancoastal states along with China Iceland Japan South Koreaand the EU aims to apply a precautionary approach to possiblefuture commercial fisheries in the high seas of the central ArcticOcean (CAO) The Agreement pledges parties to not authorizetheir flagged vessels to conduct commercial fishing in the CAOuntil conservation and management measures for fish stocksare adopted by one or more regional or sub-regional fisheriesmanagement organizations Within 2 years of entry into forceparties agree to establish a Joint Program of Scientific Researchand Monitoring with the objectives of improving understandingsof ecosystems in the CAO and of determining whether fish stocksmight exist that could be harvested on a sustainable basis Partymeetings are to be held every 2 years to review implementationof the Agreement to review all available scientific informationand to consider whether to commence negotiations for one or
more additional regional or sub-regional fisheries managementorganizations or arrangements The Agreement holds promiseto substantially expand knowledge of the CAO and adjacentecosystems and to provide refuge for Greenland sharks frompotential unregulated fishing in the high Arctic
Three North Atlantic regional fisheries managementorganizations have also addressed shark conservation inlimited ways (Davis et al 2013) The Northwest AtlanticFisheries Organization (NAFO) has adopted general sharkconservation and enforcement measures (North AtlanticFisheries Organization [NAFO] 2018a) Shark finning isprohibited by requiring shark fins to remain attached tocarcasses and for non-directed shark catches parties arerequired to encourage the live release of sharks (especiallyjuveniles) that are not intended for use as food or subsistenceParties are urged to undertake research in relation to sharksincluding ways to make fishing gear more selective keybiological and ecological parameters life history behavioraltraits and migration patterns and the identification of potentialmating pupping and nursery grounds of key shark species Since2012 NAFO has required to the extent possible species-specificreporting of shark catches in the NAFO Regulatory Area withthe amount of Greenland sharks caught in 2013 reported as222 tons (North Atlantic Fisheries Organization [NAFO]2014) and rising to 2032 tons in 2016 (North Atlantic FisheriesOrganization [NAFO] 2017) Not all shark catches have beenreported to the species level and it is not known how manyshark species were simply lumped into the category of dogfishes(North Atlantic Fisheries Organization [NAFO] 2017)
NAFOrsquos Scientific Council upon a request from theCommission has been undertaking a review of the availableinformation on the life history population status and currentfishing mortality of Greenland sharks Notably at their mostrecent meeting held in June 2018 the Scientific Council (SC)made several suggestions which included the prohibitionof directed fishing and bycatch retention and the requiredrelease of live Greenland sharks captured within the NAFOConvention Area (North Atlantic Fisheries Organization[NAFO] 2018b) In addition the SC recommended thepromotion of safe handling practices by fishers and thatimprovements be made to the reporting of all shark species byfisheries observers including the collection of shark numbersmeasurements (when feasible) and the recording of sex dataand discard disposition (ie dead or alive North AtlanticFisheries Organization [NAFO] 2018b) To reduce bycatchthe application of gear restrictions and modifications andorspatial and temporal closures (where appropriate) was alsorecommended (North Atlantic Fisheries Organization [NAFO]2018b) Approval of these suggestions by the Commissionis pending and further discussion was scheduled to takeplace at the annual NAFO meeting in September 2018(North Atlantic Fisheries Organization [NAFO] 2018b)
The North-East Atlantic Fisheries Commission (NEAFC) hasadopted shark conservation measures similar to those of NAFONEAFC also prohibits shark finning encourages the live releaseof sharks caught in non-directed fisheries and requires annualreporting of shark catches (NEAFC 2015) NEAFC continues
Frontiers in Marine Science | wwwfrontiersinorg 20 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 21
Edwards et al Advancing Research for Long-Lived Species
to prohibit directed fishing for deep sea sharks including theGreenland shark in NEAFCrsquos Regulatory Area and encouragesparties to adopt prohibitions within national waters as well(NEAFC 2017)
The International Commission for the Conservation ofAtlantic Tunas (ICCAT) has adopted various recommendationsrelating to shark conservation over the years (Davis et al 2013)Some of the most recent include recommendations on biologicalsampling for shark species where retention is prohibited (ICCAT2014) and on shortfin mako (Isurus oxyrinchus) porbeagle(Lamna nasus) and blue sharks (Prionace glauca) caught inassociation with ICCAT fisheries (ICCAT 2013 2015 2016)No recommendations have been adopted specific to Greenlandsharks however these are needed
NationalBesides a broad array of general legislative regulatory andpolicy provisions in support of sustainable fisheries and marinebiodiversity protection (VanderZwaag et al 2012 Sybersma2015) Canada adopted a National Plan of Action for theConservation and Management of Sharks in 2007 whichdescribes the Canadian legislative and regulatory framework formanaging shark populations and fisheries noting the importanceof following precautionary and ecosystem approaches in futureimplementation (Fisheries and Ocean Canada 2007) Twolimited references are made to the Greenland shark notingthe problem of Greenland shark bycatch and discards at seain the Arctic region The Plan commits Canada to moveahead with measures that increase knowledge about the lifehistory and abundance of elasmobranchs in the Arctic includingthe Greenland shark Given the numerous knowledge gapsdiscussed in this paper and the suspected implications of extremelongevity to limit the recovery of Greenland shark populations todisturbance it is important that checks and balances are put inplace to ensure that these research goals are met
A Canadian progress report on implementation of theNational Plan of Action for Sharks (NPOA-sharks) issued in2012 provided little further information regarding Greenlandshark conservation issues and efforts (Fisheries and OceansCanada 2012) The progress report once again noted the bycatchand discarding reality in the Arctic and recognized the need forfurther research on the life history and abundance estimates Thereport mentioned the completion of satellite tagging of Arcticshark species with results forthcoming The report also noted thatCanada was considering developing future projects to evaluatehow changing Arctic conditions such as climate change andincreased shipping may affect shark species
An integrated fisheries management plan (IFMP) for oneof Canadarsquos main commercial fisheries in the Arctic hasaddressed Greenland shark bycatch in limited ways The IFMPfor Greenland halibut effective in 2014 noted the ongoingproblem of inaccurate bycatch reporting and the need to improvebycatch management (Fisheries and Oceans Canada 2014) TheIFMP listed Greenland shark as a species of concern notedthat the Committee on the Status of Endangered Wildlife inCanada (COSEWIC) had not yet assessed the conservationstatus of Greenland shark and indicated the main bycatch
management measure was to require release of incidental fishcatches other than groundfish and where alive in a mannercausing the least harm
CONCLUSION
Organisms characterized by low productivity are particularlysusceptible to overfishing and stock collapse and therefore areincapable of supporting more than nominal fishing mortalityrates (Stevens et al 2000) Despite historically high catch ratesin some regions Greenland sharks may be very sensitive tooverfishing even compared to other shark species With recentevidence to support extreme longevity and conservative lifehistory traits coupled with their vulnerability to incidentalcapture by commercial fisheries the sustainability of Greenlandshark populations is of growing concern among fisheriesmanagement organizations In spite of continued developmentsin the field of Greenland shark research many questionsstill remain unanswered To develop appropriate managementactions aimed at preventing population declines there is acurrent need to address these knowledge gaps through focusedstudy and continued innovations in technology and experimentaldesign In this paper we identify key biological questions fromseveral fields of Greenland shark research and by drawing fromrecent tools and techniques from the broader literature wepropose a number of strategies which could be used to addressthese key questions In particular knowledge of the generationtimes migration patterns stock status the locations of matinggrounds and detailed information on reproduction of Greenlandsharks will improve our ability to effectively manage this long-lived species Importantly vulnerability to population declineand the biological traits that influence it are common amonglong-lived species Consequently the critical parameters outlinedin this paper and proposed methodologies to quantify themare likely applicable to most long-lived aquatic species Thereis a need for researchers to use both traditional and creativeapproaches to engage stakeholders build trust and enhancecollaboration actions that will ultimately lead to improvedmanagement of this species and long-lived species in general
AUTHOR CONTRIBUTIONS
JE was responsible for contacting and coordinating thecollaborating authors and wrote the Abstract IntroductionMethods and Movement Ecology section (with NH) TheDemographics section was written by BD JN and SC Populationgenetics and genomics was written by RW and KP Behavior andinteractions was written by YW FB and GS JS PB and JC wrotethe Physiological adaptations section Diet and trophic ecologywas written by BM JN JC and JS The Threats section waswritten by KH the outline was created by EH and commentswere provided by EH YW and SC The Laws and Policies sectionwas written by DV Table 1 which formed the basis for thestructure of this paper was created by EH and NH and revised byJE Final edits were completed by MM JE and NH All authorscontributed to the writing and editing of the final manuscript
Frontiers in Marine Science | wwwfrontiersinorg 21 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 22
Edwards et al Advancing Research for Long-Lived Species
ACKNOWLEDGMENTS
We would like to sincerely thank S M Grant C J Harvey-Clark and J Bhak for their enthusiasm and feedback inthe early stages of this work and M Treble for yourinsightful review which helped to refine this manuscript
We would also like to personally thank E Davidsonand S Popov for their invaluable advice assistance andencouragement as well as our reviewers whose constructiveguidance fostered further improvement of this researchAdditional support for NH was provided by an NSERCDiscovery Grant
REFERENCESAdams P B (1980) Life history patterns in marine fishes and their consequences
for fisheries management Fish Bull 78 1ndash12Addison R F and Stobo W T (1993) Organochlorine residue concentrations
and burdens in grey seal (Halichoerus grypus) during the first year of life J ZoolLond 230 443ndash450 doi 101111j1469-79981993tb02696x
Ahonen H Harcourt R G and Stow A J (2009) Nuclear and mitochondrialDNA reveals isolation of imperilled grey nurse shark populations (Carchariastaurus) Mol Ecol 18 4409ndash4421 doi 101111j1365-294X200904377x
Baker L L Jonsen I D Mills Flemming JE Lidgard D C Bowen W D andIverson S J (2014) Probability of detecting marine predator-prey and speciesinteractions using novel hybrid acoustic transmitter-receiver tags PLoS One9e98117 doi 101371journalpone0098117
Ball I R Possingham H P and Watts M (2009) ldquoMarxan and relatives softwarefor spatial conservation prioritisationrdquo in Spatial Conservation PrioritisationQuantitative Methods And Computational Tools eds A Moilanen K A Wilsonand H P Possingham (Oxford Oxford University Press) 185ndash195
Barker M J and Schluessel V (2005) Managing global shark fisheriessuggestions for prioritizing management strategies Aquat Conserv MarFreshw Ecosyst 15 325ndash347 doi 101002aqc660
Barkley A N Cooke S J Fisk A T Hedges K and Hussey N E (2017)Capture-induced stress in deep-water Arctic fish species Polar Biol 40 213ndash220 doi 101007s00300-016-1928-8
Baum J K Myers R A Kehler D G Worm B Harley S J and DohertyP A (2003) Collapse and conservation of shark populations in the NorthwestAtlantic Science 299 389ndash392 doi 101126science1079777
Beck B and Mansfield A W (1969) Observations on the Greenland SharkSomniosus microcephalus in Northern Baffin Island J Fishe Res Board Can26 143ndash145
Belley R Archambault P Sundby B Gilbert F and Gagnon J M (2010)Effects of hypoxia on benthic macrofauna and bioturbation in the Estuaryand Gulf of St Lawrence Canada Continent Shelf Res 30 1302ndash1313doi 101016jcsr201004010
Berland B (1961) Copepod Ommatokoita elongata (Grant) in the eyes of theGreenland sharkmdasha possible cause of mutual dependence Nature 191 829ndash830doi 101038191829a0
Bigelow H B and Schroeder W C (1948) Fishes of the Western North AtlanticPart 1 Lancelets Cyclostomes Sharks Mem Sears Found Mar Res 1 29ndash58
Binder T R Holbrook C M Miehls S M Thompson H T and Krueger C C(2014) Use of oviduct-inserted acoustic transmitters and positional telemetryto estimate timing and location of spawning a feasibility study in lake troutSalvelinus namaycush Anim Biotelemetry 214 doi 1011862050-3385-2-14
Bjerkan P A U L (1957) Notes on the Greenland Shark Acanthorhinuscarcharias (Gunn) 1 The reproduction problem of the Greenland shark RepNorwegian Fish Mar Investigat 11 1ndash7
Borucinska J D Benz G W and Whiteley H E (1998) Ocular lesions associatedwith attachment of the parasitic copepod Ommatokoita elongata (Grant) tocorneas of Greenland sharks Somniosus microcephalus (Bloch amp Schneider)J Fish Dis 21 415ndash422 doi 101046j1365-2761199800122x
Bouyoucos I A Suski C D Mandelman J W and Brooks E J (2017) Theenergetic physiological and behavioral response of lemon sharks (Negaprionbrevirostris) to simulated longline capture Comp Biochem Physiol A 20765ndash72 doi 101016jcbpa201702023
Brownscombe J W Thiem J D Hatry C Cull F Haak C R Danylchuk A Jet al (2013) Recovery bags reduce post-release impairments in locomotoryactivity and behavior of bonefish (Albula spp) following exposure to angling-related stressors J Exp Mar Biol Ecol 440 207ndash215 doi 101016jjembe201212004
Burgess M G Diekert F K Jacobsen N S Andersen K H and Gaines S D(2016) Remaining questions in the case for balanced harvesting Fish Fish 171216ndash1226 doi 101111faf12123
Campana S E Casselman J M and Jones C M (2008a) Bomb radiocarbonchronologies in the Arctic with implications for the age validation of lake trout(Salvelinus namaycush) and other Arctic species Can J Fish Aquat Sci 65733ndash743 doi 101139f08-012
Campana S E Fisk A T and Klimley A P (2015a) Movements of Arctic andnorthwest Atlantic Greenland sharks (Somniosus microcephalus) monitoredwith archival satellite pop-up tags suggest long-range migrations Deep Sea ResII Top Stud Oceanogr 115 109ndash115 doi 101016jdsr2201311001
Campana S E Gibson J Brazner J Marks L Joyce W Gosselin J-Fet al (2008b) Status of Basking Sharks in Atlantic Canada Canadian StockAssessment Sec Res Doc 2008004 Dartmouth NS Fisheries and Oceansdoi 101371journalpone0082074
Campana S E Joyce W Fowler M and Showell M (2015b) Discards hookingand post-release mortality of porbeagle (Lamna nasus) shortfin mako (Isurusoxyrinchus) and blue shark (Prionace glauca) in the Canadian pelagic longlinefishery ICES J Mar Sci 73 520ndash528 doi 101093icesjmsfsv234
Campana S E Joyce W and Kulka D W (2009) ldquoGrowth and reproductionof spiny dogfish off the eastern coast of Canada including inferences on stockstructurerdquo in Biology and Management of Dogfish Sharks eds V F GallucciG A McFarlane and G G Bargmann (Bethesda American Fisheries Society)195ndash208
Castro J I (2011) The Sharks of North America Oxford Oxford University PressChristiansen J S Mecklenburg C W and Karamushko O V (2014) Arctic
marine fishes and their fisheries in light of global change Glob Change Biol20 352ndash359 doi 101111gcb12395
CITES (nd) How CITES Works Available at httpswwwcitesorgengdischowphp
Clark M (2001) Are Deepwater Fisheries Sustainable - the Example of OrangeRoughy (Hoplostethus atlanticus) in New Zealand Fish Res 51 123ndash135doi 101016S0165-7836(01)00240-5
Clarke A and Harris C M (2003) Polar marine ecosystems major threats andfuture change Environ Conserv 30 1ndash25 doi 101098rsbl20160796
CMS (2016) Memorandum of Understanding on the Conservation of MigratorySharks (as Amended by the Signatories at their Second Meeting Costa RicaFebruary 2016) Available at httpswwwcmsintsharksenpagesharks-mou-text
CMS (2018a) Memorandum of Understanding on the Conservation of MigratorySharks Signatories and Range Status Available at httpswwwcmsintsharksenmeetingsmeetings-of-signatories
CMS (2018b) Memorandum of Understanding on the Conservation of MigratorySharks Species Available at httpswwwcmsintsharksenspecies [accessedJune 22 2018]
Cooke S J Iverson S J Stokesbury M J Hinch S G Fisk A TVanderZwaag D L et al (2011) Ocean Tracking Network Canada a networkapproach to addressing critical issues in fisheries and resource managementwith implications for ocean governance Fisheries 36 583ndash592 doi 101080036324152011633464
Corsolini S Ancora S Bianchi N Mariotti G Leonzio C and ChristiansenJ S (2014) Organotropism of persistent organic pollutants and heavy metalsin the Greenland shark Somniosus microcephalus in NE Greenland Mar PollutBull 87 381ndash387 doi 101016jmarpolbul201407021
Cosandey-Godin A Krainski E T Worm B and Flemming J M (2014)Applying Bayesian spatiotemporal models to fisheries bycatch in the CanadianArctic Can J Fish Aquat Sci 72 186ndash197 doi 101139cjfas-2014-0159
Davis B VanderZwaag D L Cosandey-Godin A Hussey N E Kessel S Tand Worm B (2013) The conservation of the Greenland shark (Somniosus
Frontiers in Marine Science | wwwfrontiersinorg 22 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 23
Edwards et al Advancing Research for Long-Lived Species
microcephalus) setting scientific law and policy coordinates for avoiding aspecies at risk J Int Wildlife Law Policy 16 300ndash330 doi 101080138802922013805073
Department of Fisheries and Oceans [DFO] (2006) A Harvest Strategy Compliantwith the Precautionary (Approach) Canadian Science Advisory SecretariatScience Advisory Report Ottawa ON Fisheries and Oceans Canada 232006
Department of Fisheries and Oceans [DFO] (2009) A Fishery Decision-MakingFramework Incorporating the Precautionary Approach Sustainable FisheriesFramework [website] (23 March 2009) Available at httpwwwdfo-mpogccareports-rapportsregssff-cpdprecaution-back-fiche-enghtm
Department of Fisheries and Oceans [DFO] (2013) Policy on Managing BycatchSustainable Fisheries Framework [website] (8 January 2013) Available athttpwwwdfo-mpogccareports-rapportsregssff-cpdbycatch-policy-prise-access-enghtm
Department of Fisheries and Oceans [DFO] (2016) Greenland Shark BycatchEstimates in NAFO areas 0A and 0B Dartmouth NS Bedford Institute ofOceanography
Devine B M Wheeland L J and Fisher J A (2018) First estimates of Greenlandshark (Somniosus microcephalus) local abundances in Arctic waters Sci Rep8974 doi 101038s41598-017-19115-x
Dudgeon C L Pollock K H Braccini J M Semmens J M and Barnett A(2015) Integrating acoustic telemetry into markndashrecapture models to improvethe precision of apparent survival and abundance estimates Oecologia 178761ndash772 doi 101007s00442-015-3280-z
Engarings A Loslashkkeborg S Ona E and Soldal A V (1996) Effects of seismicshooting on local abundance and catch rates of cod ((Gadus morhua) andhaddock) (Melanogrammus aeglefinus) Can J Fish Aquat Sci 53 2238ndash2249doi 101139f96-177
Ferrando S Gallus L Ghigliotti L Vacchi M Nielsen J Christiansen J Set al (2016) Gross morphology and histology of the olfactory organ of theGreenland shark Somniosus microcephalus Polar Biol 39 1399ndash1409 doi 101007s00300-015-1862-1
Ferry-Graham L A and Gibb A C (2001) Comparison of fasting andpostfeeding metabolic rates in a sedentary shark Cephaloscyllium ventriosumCopeia 2001 1108ndash1113 doi 1016430045-8511(2001)001[1108COFAPM]20CO2
Fisheries and Ocean Canada (2007) National Plan of Action for the Conservationand Management of Sharks Ottawa ON Fish And Fisheries
Fisheries and Oceans Canada (2012) Canadarsquos Progress Report on theImplementation of Key Actions Taken Pursuant to the National Plan of Actionon the Conservation and Management of Sharks (March 2007) Ottawa ON FishAnd Fisheries
Fisheries and Oceans Canada (2014) Integrated Fishery Management PlanGreenland Halibut (Reinhardtius hippoglossoides) Northwest Atlantic FisheriesOrganization Subarea O Winnipeg MB Fish And Fisheries
Fisk A T Lydersen C and Kovacs K M (2012) Archival pop-off tag trackingof Greenland sharks Somniosus microcephalus in the High Arctic waters ofSvalbard Norway Mar Ecol Prog Ser 468 255ndash265 doi 103354meps09962
Fisk A T Tittlemier S A Pranschke J L and Norstrom R J (2002) Usinganthropogenic contaminants and stable isotopes to assess the feeding ecologyof Greenland sharks Ecology 83 2162ndash2172 doi 1018900012-9658(2002)083[2162UACASI]20CO2
Food and Agricultural Organization [FAO] (1996) Precautionary Approach toFisheries 1 Guidelines on the precautionary Approach to Capture Fisheries andSpecies Introductions Rome Food and Agricultural Organization
Food and Agricultural Organization [FAO] (1999) International Plan of Actionfor the Conservation and Management of Sharks Rome Food and AgriculturalOrganization
Francis R I C C (1988) Maximum likelihood estimation of growth andgrowth variability from tagging data N Z J Mar Freshwater Res 22 43ndash51doi 1010800028833019889516276
Frank K T Petrie B Fisher J A and Leggett W C (2011) Transient dynamicsof an altered large marine ecosystem Nature 47786 doi 101038nature10285
Fraser D J and Bernatchez L (2001) Adaptive evolutionary conservationtowards a unified concept for defining conservation units Mol Ecol 102741ndash2752 doi 101046j1365-294X2001t01-1-01411x
Froese R Walters C Pauly D Winker H Weyl O L Demirel N et al (2015)A critique of the balanced harvesting approach to fishing ICES J Mar Sci 731640ndash1650 doi 101093icesjmsfsv122
Gallagher A J Staaterman E R Cooke S J and Hammerschlag N(2016) Behavioural responses to fisheries capture among sharks caught usingexperimental fishery gear Can J Fish Aquat Sci 74 1ndash7 doi 101139cjfas-2016-0165
Gallant J J Rodriguez M A Stokesbury M J and Harvey-Clark C (2016)Influence of environmental variables on the diel movements of the greenlandshark (Somniosus microcephalus) in the St Lawrence Estuary Can Field Nat130 1ndash14 doi 1022621cfnv130i11784
Garcia S M Bianchi G Charles A Kolding J Rice J Rochet M-J et al(2014) Balanced Harvest in the Real World Scientific Policy and OperationalIssues in an Ecosystem Approach to Fisheries Report of an international scientificworkshop of the IUCN Fisheries Expert Group (IUCNCEMFEG) organized inclose cooperation with the Food and Agriculture Organization of the UnitedNations (FAO) Rome 2909-02102014 Available at httpslibrarywurnlWebQuerywurpubs492588
Garcia S M Kolding J Rice J Rochet M J Zhou S Arimoto T et al (2012)Reconsidering the consequences of selective fisheries Science 335 1045ndash1047doi 101126science1214594
Garciacutea V B Lucifora L O and Myers R A (2008) The importance of habitatand life history to extinction risk in sharks skates rays and chimaeras Proc RSoc Lond B Biol Sci 275 83ndash89 doi 101098rspb20071295
Garde E Heide-Joslashrgensen M P Hansen S H Nachman G and ForchhammerM C (2007) Age-specific growth and remarkable longevity in narwhals(Monodon monoceros) from West Greenland as estimated by aspartic acidracemization J Mammal 88 49ndash58 doi 10164406-MAMM-A-056R1
George J C Bada J Zeh J Scott L Brown S E Orsquohara T et al (1999) Ageand growth estimates of bowhead whales (Balaena mysticetus) via aspartic acidracemization Can J Zool 77 571ndash580 doi 101139z99-015
Gillooly J F Brown J H West G B Savage V M and Charnov E L (2001)Effects of size and temperature on metabolic rate Science 293 2248ndash2251doi 101126science1061967
Grant S M Sullivan R and Hedges K J (2018) Greenland shark(Somniosus microcephalus) feeding behavior on static fishing gear effect ofSMART (Selective Magnetic and Repellent-Treated) hook deterrent technologyand factors influencing entanglement in bottom longlines PeerJ 6e4751doi 107717peerj4751
Gunnarsdottir R and Joslashrgensen M W (2008) ldquoUtilization possibilites ofwaste products from fishing and hunting to biogas and bio-oil production inUummannaq Countyrdquo in Proceedings of the Sustainable Energy Supply in theArctic-sun wind water etc ARTEK Event (Copenhagen Arctic TechnologyCentre) 52ndash58
Halpern B S Walbridge S Selkoe K A Kappel C V Micheli F Drsquoagrosa Cet al (2008) A global map of human impact on marine ecosystems Science 319948ndash952 doi 101126science1149345
Hamady L L Natanson L J Skomal G B and Thorrold S R (2014) Vertebralbomb radiocarbon suggests extreme longevity in white sharks PLoS One9e84006 doi 101371journalpone0084006
Hansen P M (1963) Tagging experiments with the Greenland shark (Somniosusmicrocephalus (Bloch and Schneider)) in subarea 1 Int Comm Northwest AtlFish Spec Publ 4 172ndash175
Hart N S Lisney T J Marshall N J and Collin S P (2005) Multiple conevisual pigments and the potential for trichromatic colour vision in two speciesof elasmobranch J Exp Biol 207 4587ndash4594 doi 101242jeb01314
Harvey-Clark C J Gallant J J and Batt J H (2005) Vision and its relationshipto novel behaviour in St Lawrence River Greenland Sharks Somniosusmicrocephalus Can Field Nat 119 355ndash358 doi 1022621cfnv119i3145
Haulsee D E Fox D A Breece M W Brown L M Kneebone J Skomal G Bet al (2016) Social network analysis reveals potential fission-fusion behavior ina shark Sci Rep 634087 doi 101038srep34087
Heppell S S Heppell S A Read A J and Crowder L B (2005) Effects ofFishing on Long-Lived Marine Organisms Marine Conservation Biology theScience of Maintaining the searsquos Biodiversity Washington DC Island Press211ndash231
Herbert N A Skov P V Tirsgaard B Bushnell P G Brill R W Clark C Het al (2017) Blood O 2 affinity of a large polar elasmobranch the Greenland
Frontiers in Marine Science | wwwfrontiersinorg 23 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 24
Edwards et al Advancing Research for Long-Lived Species
shark Somniosus microcephalus Polar Biol 40 2297ndash2305 doi 101007s00300-017-2142-z
Herdendorf C E and Berra T M (1995) A Greenland shark from the wreckof the SS Central America at 2200 meters Trans Am Fish Soc 124 950ndash953doi 1015771548-8659(1995)124lt0950AGSFTWgt23CO2
Heupel M R Simpfendorfer C A and Lowe C (2005) Passive AcousticTelemetry Technology Current Applications and Future Directions TechnicalReport No 1066 Sarasota FL Mote Marine Laboratory
Hillary R M Bravington M V Patterson T A Grewe P Bradford RFeutry P et al (2018) Genetic relatedness reveals total population size of whitesharks in eastern Australia and New Zealand Sci Rep 82661 doi 101038s41598-018-20593-w
Holland K N Meyer C G and Dagorn L C (2009) Inter-animal telemetryresults from first deployment of acoustic lsquobusiness cardrsquotags Endanger SpeciesRes 10 287ndash293 doi 103354esr00226
Horning M and Mellish J A E (2014) In cold blood evidence of Pacific sleepershark (Somniosus pacificus) predation on Steller sea lions (Eumetopias jubatus)in the Gulf of Alaska Fish Bull 112 297ndash311 doi 107755FB11246
Howarth R Chan F Conley D J Garnier J Doney S C Marino Ret al (2011) Coupled biogeochemical cycles eutrophication and hypoxia intemperate estuaries and coastal marine ecosystems Front Ecol Environ 918ndash26 doi 10230741149673
Huntington H P (2009) A preliminary assessment of threats to arctic marinemammals and their conservation in the coming decades Mar Policy 33 77ndash82doi 101016jmarpol200804003
Hussey N E Cosandey-Godin A Walter R P Hedges K J VanGerwen-Toyne M Barkley A N et al (2015a) Juvenile Greenland sharks Somniosusmicrocephalus (Bloch amp Schneider 1801) in the canadian arctic Polar Biol 38493ndash504 doi 101007s00300-014-1610-y
Hussey N E Kessel S T Aarestrup K Cooke S J Cowley P D FiskA T et al (2015b) Aquatic animal telemetry a panoramic window into theunderwater world Science 3481255642 doi 101126science1255642
Hussey N E MacNeil M A McMeans B C Olin J A Dudley S F Cliff Get al (2014) Rescaling the trophic structure of marine food webs Ecol Lett 17239ndash250 doi 101111ele12226
Hussey N E Orr J Fisk A T Hedges K J Ferguson S H and BarkleyA N (2018) Mark report satellite tags (mrPATs) to detail large-scale horizontalmovements of deep water species First results for the Greenland shark(Somniosus microcephalus) Deep Sea Res I Oceanogr Res Pap 134 32ndash40doi 101016jdsr201803002
Hutton J M and Dickson B [eds] (2000) Endangered species threatenedconvention the past present and future of CITES the Convention onInternational Trade in Endangered Species of Wild Fauna and Flora LondonEarthscan
ICCAT (2013) Recommendation 13-10 on Biological Sampling of Prohibited SharkSpecies by Scientific Observers Available at httpswwwiccatintDocumentsRecscompendiopdf-e2013-10-epdf
ICCAT (2014) Recommendation 14-06 by ICCAT on Shortfin Mako Caughtin Association with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2014-06-epdf
ICCAT (2015) Recommendation 15-06 by ICCAT on Porbeagle Caught inAssociation with ICCAT Fisheries Available at httpswwwiccatintDocumentsRecscompendiopdf-e2015-06-epdf
ICCAT (2016) Recommendation 16-12 by ICCAT on Management Measures for theConservation of Atlantic Blue Shark Caught in Association with ICCAT FisheriesAvailable at httpswwwccsbtorgsitesdefaultfilesuserfilesfileother_rfmo_measuresiccatICCAT_2016-12-epdf
ICES (2012) ICES Implementation of Advice for Data-Limited Stocksin 2012 in its 2012 Advice ICES DLS Guidance Report 2012 ICESAdvisory Committee ICES CM 2012ACOM 68 Available at httpswwwicesdksitespubPublication20ReportsExpert20Group20Reportacom2012ADHOCDLS20Guidance20Report202012pdf
ICES (2017) Report of the North Western Working Group (NWWG) 27 April-4May 2017 Copenhagen ICES
Idrobo C J and Berkes F (2012) Pangnirtung inuit and the Greenland sharkco-producing knowledge of a little discussed species Hum Ecol 40 405ndash414doi 101007s10745-012-9490-7
IUCN (2017) The IUCN Red List of Threatened Species Available at httpwwwiucnredlistorgdetails602130
Ivanova S V Kessel S T Landry J OrsquoNeill C McLean M F Espinoza Met al (2018) Impact of vessel traffic on the home ranges and movement ofShorthorn Sculpin (Myoxocephalus scorpius) in the nearshore environment ofthe high Arctic Can J Fish Aquat Sci 75 2390ndash2400 doi 101139cjfas-2017-0418
Jensen A S (1948) Contribution to the ichthyofauna of Greenland 8ndash24 SpoliaZool MusHaun 9 1ndash84
Kalinoski M Hirons A Horodysky A and Brill R (2014) Spectral sensitivityluminous sensitivity and temporal resolution of the visual systems in threesympatric temperate coastal shark species J Comp Physiol A 200 997ndash1013doi 101007s00359-014-0950-y
Kalmijn A J (1971) The electric sense of sharks and rays J Exp Biol 55 371ndash383Koefoed E (1957) Notes on the Greenland shark Acanthorhinus carcharias
(Gunn) 2 A uterine foetus and the uterus from a Greenland shark RepNorwegian Fish Mar Invest 11 8ndash12
Korpimaumlki E (1984) Population dynamics of birds of prey in relation tofluctuations in small mammal populations in western Finland Ann ZoolFennici 21 287ndash293
Kukulya A L Stokey R Littlefield R Jaffre F Padilla E M H and Skomal G(2015) 3D Real-Time Tracking Following and Imaging of White Sharks withan Autonomous Underwater Vehicle OCEANS 2015-Genova Available at httpieeexploreieeeorgdocument7271546
Larsen J Bushnell P Steffensen J Pedersen M Qvortrup K and Brill R(2017) Characterization of the functional and anatomical differences in theatrial and ventricular myocardium from three species of elasmobranch fishessmooth dogfish (Mustelus canis) sandbar shark (Carcharhinus plumbeus) andclearnose skate (Raja eglanteria) J Comp Physiol B 187 291ndash313 doi 101007s00360-016-1034-9
Leclerc L M Lydersen C Haug T A Glover K T Fisk A and Kovacs K(2011) Greenland sharks (Somniosus microcephalus) scavenge offal from minke(Balaenoptera acutorostrata) whaling operations in Svalbard (Norway) PolarRes 307342 doi 103402polarv30i07342
Leclerc L M E Lydersen C Haug T Bachmann L Fisk A T and KovacsK M (2012) A missing piece in the Arctic food web puzzle Stomach contentsof Greenland sharks sampled in Svalbard Norway Polar Biol 35 1197ndash1208doi 101007s00300-012-1166-7
Lennox R J Aarestrup K Cooke S J Cowley P D Deng Z D Fisk A Tet al (2017) Envisioning the future of aquatic animal tracking technologyscience and application Bioscience 67 884ndash896 doi 101093bioscibix098
Li H Tian C Lu J Myjak M J Martinez J J Brown R S et al (2016) Anenergy harvesting underwater acoustic transmitter for aquatic animals Sci Rep633804 doi 101038srep33804
Lidgard D C Bowen W D Jonsen I D and Iverson S J (2012) Animal-borneacoustic transceivers reveal patterns of at-sea associations in an upper-trophiclevel predator PLoS One 7e48962 doi 101371journalpone0048962
Lidgard D C Bowen W D Jonsen I D McConnell B J Lovell P WebberD M et al (2014) Transmitting species-interaction data from animal-bornetransceivers through Service Argos using Bluetooth communication MethodsEcol Evol 5 864ndash871 doi 1011112041-210X12235
Lisney T J Yopak K E Camilieri-Asch V and Collin S P (2017) Ontogeneticshifts in brain organization in the bluespotted stingray Neotrygon kuhlii(Chondrichthyes Dasyatidae) Brain Behav Evol 89 68ndash83 doi 101159000455223
Lucas Z N and McAlpine D F (2002) Extralimital occurrences of ringed sealsPhoca hispida on Sable Island Nova Scotia Can Field Nat 116 607ndash610
Lucas Z N and Natanson L J (2010) ldquoTwo shark species involved in predationon seals at Sable Island Nova Scotia Canadardquo in Proceedings of the Nova ScotianInstitute of Science (NSIS) Nova Scotia
MacNeil M A McMeans B C Hussey N E Vecsei P Svavarsson J KovacsK M et al (2012) Biology of the Greenland shark Somniosus microcephalusJ Fish Biol 80 991ndash1018 doi 101111j1095-8649201203257x
Martin A P Naylor G J and Palumbi S R (1992) Rates of mitochondrialDNA evolution in sharks are slow compared with mammals Nature 357153doi 101038357153a0
Frontiers in Marine Science | wwwfrontiersinorg 24 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 25
Edwards et al Advancing Research for Long-Lived Species
McCann K and Shuter B (1997) Bioenergetics of life history strategies and thecomparative allometry of reproduction Can J Fishe Aquat Sci 54 1289ndash1298doi 101139f97-026
McCann K S Rasmussen J B and Umbanhowar J (2005) The dynamics ofspatially coupled food webs Ecol Lett 8 513ndash523 doi 101111j1461-0248200500742x
McCarthy E Moretti D Thomas L DiMarzio N Morrissey R Jarvis S et al(2011) Changes in spatial and temporal distribution and vocal behavior ofBlainvillersquos beaked whales (Mesoplodon densirostris) during multiship exerciseswith midndashfrequency sonar Mar Mammal Sci 27 206ndash226 doi 101111j1748-7692201000457x
McClellan C M Read A J Price B A Cluse W M and Godfrey M H (2009)Using telemetry to mitigate the bycatch of long-lived marine vertebrates EcolAppl 19 1660ndash1671 doi 10189008-10911
McComb M Frank T M Hueter R E and Kajiura S M (2010) Physiologicaltemporal resolution and spectral sensitivity of the visual system of three coastalshark species from different light environments Physiol Biochem Zool 83299ndash307 doi 101086648394
McMeans B C Arts M T and Fisk A T (2015) Impacts of food web structureand feeding behavior on mercury exposure in Greenland Sharks (Somniosusmicrocephalus) Sci Total Environ 509 216ndash225 doi 101016jscitotenv201401128
McMeans B C Arts M T Lydersen C Kovacs K M Hop H Falk-Petersen Set al (2013a) The role of Greenland sharks (Somniosus microcephalus) in anArctic ecosystem assessed via stable isotopes and fatty acids Mar Biol 1601223ndash1238 doi 101007s00227-013-2174-z
McMeans B C Rooney N Arts M T and Fisk A T (2013b) Food webstructure of a coastal Arctic marine ecosystem and implications for stabilityMar Ecol Prog Ser 482 17ndash28 doi 103354meps10278
McMeans B C Svavarsson J Dennard S and Fisk A T (2010) Diet andresource use among Greenland sharks (Somniosus microcephalus) and teleostssampled in Icelandic waters using δ13C δ15N and mercury Can J FishAquat Sci 67 1428ndash1438 doi 101139F10-072
Meekan M Austin C M Tan M H Wei N-W V Miller A Pierce S J et al(2017) iDNA at sea recovery of whale shark (Rhincodon typus) mitochondrialDNA sequences from the whale shark copepod (Pandarus rhincodonicus)confirms global population structure Front Marine Sci 4420 doi 103389fmars201700420
Metcalfe K Vaz S Engelhard G H Villanueva M C Smith R J andMackinson S (2015) Evaluating conservation and fisheries managementstrategies by linking spatial prioritization software and ecosystem and fisheriesmodelling tools J Appl Ecol 52 665ndash674 doi 1011111365-266412404
Meyer C G and Holland K N (2012) Autonomous measurement of ingestionand digestion processes in free-swimming sharks J Exp Biol 215 3681ndash3684doi 101242jeb075432
Meyer C G Papastamatiou Y P and Holland K N (2010) A multipleinstrument approach to quantifying the movement patterns and habitat use oftiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) atFrench Frigate Shoals Hawaii Mar Biol 157 1857ndash1868 doi 101007s00227-010-1457-x
Milano I Babbucci M Cariani A Atanassova M Bekkevold D CarvalhoG R et al (2014) Outlier SNP markers reveal fine-scale genetic structuringacross E uropean hake populations (Merluccius merluccius) Mol Ecol 23118ndash135 doi 101111mec12568
Moffitt E A Punt A E Holsman K Aydin K Y Ianelli J N and Ortiz I(2016) Moving towards ecosystem-based fisheries management options forparameterizing multi-species biological reference points Deep Sea Res II 134350ndash359 doi 101016jdsr2201508002
Moritz C (1994) Defining lsquoevolutionarily significant unitsrsquo for conservationTrends Ecol Evol 9 373ndash375 doi 1010160169-5347(94)90057-4
Murray B W Wang J Y Yang S C Stevens J D Fisk A and Svavarsson J(2008) Mitochondrial cytochrome b variation in sleeper sharks (SqualiformesSomniosidae) Mar Biol 153 1015ndash1022 doi 101007s00227-007-0871-1
Musick J A (1999a) Criteria to define extinction risk in marine fishes theAmerican Fisheries Society initiative Fisheries 24 6ndash14 doi 1015771548-8446(1999)024lt0006CTDERIgt20CO2
Musick J A (1999b) Ecology and conservation of long-lived marine animals AmFish Soc Symp 23 1ndash10
Musick J A Berkeley S A Cailliet G M Camhi M Huntsman GNammack M et al (2000) Protection of marine fish stocks at riskof extinction AFS policy statement Fisheries 25 6ndash8 doi 1015771548-8446(2000)025lt0006POMFSAgt20CO2
Myers R A and Worm B (2003) Rapid worldwide depletion ofpredatory fish communities Nature 423 280ndash283 doi 101038nature01610
Naito Y Costa D P Adachi T Robinson P W Peterson S H Mitani Yet al (2017) Oxygen minimum zone an important oceanographic habitatfor deep-diving northern elephant seals Mirounga angustirostris Ecol Evol 76259ndash6270 doi 101002ece33202
Nakamura I Meyer C G and Sato K (2015) Unexpected positive buoyancyin deep sea sharks Hexanchus griseus and a Echinorhinus cookei PLoS One10e0127667 doi 101371journalpone0127667
NEAFC (2015) Recommendation 102015 on Conservation of Sharks Caughtin Association with Fisheries Managed by the North-East Atlantic FisheriesCommission Available at httpwwwiotcorgcmmresolution-1705-E280A8-conservation-sharks-caught-association-fisheries-managed-iotc
NEAFC (2017) Recommendation 102017 on Conservation and ManagementMeasures for Deep Sea Sharks in the NEAFC Regulatory Area for 2017 to 2019London NEAFC
Nielsen J (2013) Age Estimation and Feeding Ecology of Greenland SharksSomniosus microcephalus in Greenland waters MSc thesis Aquatic Science andTechnology Danish Technical University Denmark
Nielsen J (2017) The Greenland shark (Somniosus microcephalus) Diet Trackingand Radiocarbon Age Estimates Reveal the Worldrsquos Oldest Vertebrate Doctoraldissertation Department of Biology Faculty of Science University ofCopenhagen Denmark
Nielsen J Hedeholm R B Heinemeier J Bushnell P G Christiansen J SOlsen J et al (2016) Eye lens radiocarbon reveals centuries of longevity inthe Greenland shark (Somniosus microcephalus) Science 353 702ndash704 doi101126scienceaaf1703
Nielsen J Hedeholm R B Simon M and Steffensen J F (2014) Distributionand feeding ecology of the Greenland shark (Somniosus microcephalus)in Greenland waters Polar Biol 37 37ndash46 doi 101007s00300-013-1408-3
NOAA (2002) Taking of marine mammals incidental to commercial fishingoperations atlantic large whale take reduction plan regulations US Departmentof Commerce Fed Regist 67 1133ndash1141
Norse E A Brooke S Cheung W W Clark M R Ekeland I Froese Ret al (2012) Sustainability of deep-sea fisheries Mar Policy 36 307ndash320doi 101016jmarpol201106008
North Atlantic Fisheries Organization [NAFO] (2004) NAFO (Precautionary)Approach Framework 2004 NAFO Fisheries Council Document 0418Available at httpswwwnafointScienceNAFO-FrameworksNAFO-Precautionary-Approach
North Atlantic Fisheries Organization [NAFO] (2014) Annual Compliance Review2014 (Compliance Report for Fishing Year 2013 Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2017) Annual Compliance Review2017 (Compliance Report for Fishing Year 2016) Available at httpswwwnafointFisheriesCompliance
North Atlantic Fisheries Organization [NAFO] (2018a) Conservation andEnforcement Measures 2018 Available at httpswwwnafointFisheriesConservation
North Atlantic Fisheries Organization [NAFO] (2018b) Report of the ScientificCouncil Meeting (1-14 June 2018 Halifax Nova Scotia) Dartmouth NS NAFOdoi 101051alr2008003
North Atlantic Fisheries Organization [NAFO] (2018c) Bryk JL KJ Hedgesand MA Treble Summary of Greenland Shark (Somniosus microcephalus)catch in Greenland Halibut (Reinhardtius hippoglossoides) fisheries and scientificsurveys conducted in NAFO Subarea 0 Dartmouth Northwest Atlantic FisheriesOrganization doi 101086282697
Oleson E M Calambokidis J McKenna M and Hidebrand J (2010) ldquoBluewhale behavioral response study amp field testing of the new bioacoustic proberdquo inProceedings of the National Oceanic and Atmospheric Administration (HonoluluHI International Tsunami Information Center) doi 101126science1098222
Frontiers in Marine Science | wwwfrontiersinorg 25 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 26
Edwards et al Advancing Research for Long-Lived Species
Papastamatiou Y P Meyer C G and Holland K N (2007) A new acoustic pHtransmitter for studying the feeding habits of free-ranging sharks Aquat LivingResour 20 287ndash290 doi 101111j1095-8649200902319x
Pianka E R (1970) On r-and K-selection Am Nat 104 592ndash597Pikitch E Santora C Babcock E A Bakun A Bonfil R Conover D O
et al (2004) Ecosystem-based fishery management Science 305 346ndash347doi 101371journalpone0022588
Popper A N and Hastings M C (2009) The effects of anthropogenic sources ofsound on fishes J Fish Biol 75 455ndash489 doi 101111j1095-8649200902319x
Porteiro F M Sutton T T Byrkjedal I Orlov A Heino M P MenezesG M M et al (2017) Fishes of the Northern Mid-Atlantic Ridge CollectedDuring the MAR-ECO Cruise in June-July 2004 An Annotated ChecklistAvailable at httpsnsuworksnovaeduocc_facreports102
Ridoux V Hall A J Steingrimsson G and Olafsson G (1998) An inadvertenthoming experiment with a young ringed seal Phoca hispida Mar Mammal Sci14 883ndash888 doi 101038nature04887
Rigby C and Simpfendorfer C A (2015) Patterns in life history traits ofdeep-water chondrichthyans Deep Sea Res II 115 30ndash40 doi 101163ej9789004174405i-5459
Robbins R and Fox A (2012) Further evidence of pigmentation change inwhite sharks Carcharodon carcharias Mar Freshwater Res 63 1215ndash1217doi 101371journalpone0186181
Rooney N McCann K Gellner G and Moore J C (2006) Structuralasymmetry and the stability of diverse food webs Nature 442265 doi 101134S0032945213010128
Russell D A and VanderZwaag D L (2010) ldquoThe International Law and PolicySeascape Governing Transboundary Fisheriesrdquo in Recasting TransboundaryFisheries Management Arrangements in Light of Sustainability PrinciplesCanadian and International Perspectives eds A Dawn Russell L David andVanderZwaag (Leiden Martinus Nijhoff) doi 101007s12686-016-0676-y
Russo R Giordano D Paredi G Marchesani F Milazzo L Altomonte Get al (2017) The Greenland shark Somniosus microcephalusmdashHemoglobinsand ligand-binding properties PLoS One 12e018618 doi 101371journalpone0186181
Rusyaev S M and Orlov A M (2013) Bycatches of the greenland sharkSomniosus microcephalus (Squaliformes Chondrichthyes) in the barents seaand the adjacent waters under bottom trawling data J Ichthyol 53 111ndash115doi 103354meps331243
Santaquiteria A Nielsen J Klemetsen T Willassen N P and Praeligbel K(2017) The complete mitochondrial genome of the long-lived Greenlandshark (Somniosus microcephalus) characterization and phylogenetic positionConserv Genet Resour 9 351ndash355 doi 101242jeb186957
Santaquiteria A G (2016) Mitogenomic Characterization and PhylogeneticPosition of the Oldest Living Vertebrate Species - the Greenland shark(Somniosus microcephalus) MSc thesis UiT The Arctic University of NorwayTromsoslash
Saragrave G Dean J M drsquoAmato D Buscaino G Oliveri A Genovese S et al(2007) Effect of boat noise on the behaviour of bluefin tuna Thunnus thynnusin the Mediterranean Sea Mar Ecol Prog Ser 331 243ndash253 doi 101017S0376892909990191
Shadwick R E Bernal D Bushnell P G and Steffensen J F (2018) Bloodpressure in the Greenland shark as estimated from ventral aortic elasticityJ Exp Biol 221jeb186957 doi 101242jeb186957
Shiels H A Kusu-Orkar T-E Ahmad S Reid B Gurney A Bernal Det al (2018) Cardiac structure of the worldrsquos oldest vertebrate the GreenlandShark Paper Presented at Society Experimental Biology meeting Florence doi101007s00227-004-1332-8
Simpfendorfer C A and Kyne P M (2009) Limited potential to recover fromoverfishing raises concerns for deep-sea sharks rays and chimaeras EnvironConserv 36 97ndash103 doi 101111jfb12828
Sims D W and Davies S J (1994) Does specific dynamic action (SDA) regulatereturn of appetite in the lesser spotted dogfish Scyliorhinus caniculcla J FishBiol 45 341ndash348 doi 101016jtree201004005
Skomal G B and Benz G W (2004) Ultrasonic Tracking of GreenlandSharks Somniosus Microcephalus under Arctic Ice Mar Biol 145 489ndash498doi 101242jeb059667
Skomal G B Hoyos-Padilla E M Kukulya A and Stokey R (2015) Subsurfaceobservations of white shark Carcharodon carcharias predatory behaviour using
an autonomous underwater vehicle J Fish Biol 87 1293ndash1312 doi 101111jfb12828
Slabbekoorn H Bouton N van Opzeeland I Coers A ten Cate C and PopperA N (2010) A noisy spring the impact of globally rising underwater soundlevels on fish Trends Ecol Evol 25 419ndash427 doi 101016jtree201004005
Speers-Roesch B Brauner C J Farrell A P Hickey A J RenshawG M Wang Y S et al (2012) Hypoxia tolerance in elasmobranchs IICardiovascular function and tissue metabolic responses during progressive andrelative hypoxia exposures J Exp Biol 215 103ndash114 doi 101242jeb059667
Stevens J D Bonfil R Dulvy N K and Walker P A (2000) The effects offishing on sharks rays and chimaeras (chondrichthyans) and the implicationsfor marine ecosystems ICES J Mar Sci 57 476ndash494 doi 101007s00227-005-0061-y
Stewart R E A Campana S E Jones C M and Stewart B E (2006) Bombradiocarbon dating calibrates beluga (Delphinapterus leucas) age estimates CanJ Zool 84 1840ndash1852 doi 101002etc330
Stirling I and McEwan E H (1975) The caloric value of whole ringed seals(Phoca hispida) in relation to polar bear (Ursus maritimus) ecology and huntingbehavior Can J Zool 8 1021ndash1027 doi 101016jmarpolbul200704018
Stokesbury M J Harvey-Clark C Gallant J Block B A and MyersR A (2005) Movement and environmental preferences of Greenland sharks(Somniosus microcephalus) electronically tagged in the St Lawrence EstuaryCanada Mar Biol 148 159ndash165 doi 101016jmarpol201507008
Strid A Athanassiadis I Athanasiadou M Svavarsson J Paumlpke O andBergman Aring (2010) Neutral and phenolic brominated organic compoundsof natural and anthropogenic origin in northeast Atlantic Greenlandshark (Somniosus microcephalus) Environ Toxicol Chem 29 2653ndash2659doi 101002etc330
Strid A Joumlrundsdoacutettir H Paumlpke O Svavarsson J and Bergman Aring (2007)Dioxins and PCBs in Greenland shark (Somniosus microcephalus) fromthe north-east Atlantic Mar Pollut Bull 54 1514ndash1522 doi 1011639789004345515
Sybersma S (2015) Review of shark legislation in Canada as a conservation toolMar Policy 61 121ndash126 doi 101016jjmarsys200907005
Taylor E W Short S T and Butler P J (1977) The role of the cardiac vagusin the response of the dogfish Scyliorhinus canicula to hypoxia J Exp Biol 7057ndash75 doi 101371journalpone0190467
Techera E J and Klein N (2017) International Law of Sharks Obstacles Optionsand Opportunities Netherlands Brill
Travers M Watermeyer K Shannon L J and Shin Y J (2010) Changes infood web structure under scenarios of overfishing in the southern Benguelacomparison of the Ecosim and OSMOSE modelling approaches J Mar Syst79 101ndash111
Trochta J T Pons M Rudd M B Krigbaum M Tanz A and Hilborn R(2018) Ecosystem-based fisheries management perception on definitionsimplementations and aspirations PLoS One 13e0190467 doi 101371journalpone0190467
US National Research Council (1998) A Report of the Committee on EcosystemManagement for Sustainable Fisheries Washington DC National AcademyPress doi 101002ece33325
US Department of State (2017) Meeting on High Seas Fisheries in the CentralArctic Ocean 28-30 November 2017 Chairmanrsquos Statement Available at httpswwwstategoveoesocnsoparls276136htm doi 101007BF00182340
VanderZwaag D L Hutchings J A Jennings S and Peterman R M(2012) Canadarsquos international and national commitments to sustain marinebiodiversity Environ Rev 20 312ndash352 doi 101016jjembe201204021
Walter R P Roy D Hussey N E Stelbrink B Kovacs K M Lydersen C et al(2017) Origins of the Greenland shark (Somniosus microcephalus) Impacts ofice-olation and introgression Ecol Evol 7 8113ndash8125 doi 101002ece33325
Walters C and Maguire J J (1996) Lessons for stock assessment from thenorthern cod collapse Rev Fish Biol Fish 6 125ndash137 doi 101016jtree201012011
Watanabe Y Y Lydersen C Fisk A T and Kovacs K M (2012) The slowestfish swim speed and tail-beat frequency of Greenland sharks J Exp Mar BiolEcol 426 5ndash11 doi 101007s10228-004-0244-4
Williamson G R (1963) Common porpoise from the stomach of a Greenlandshark J Fish Res Board Can 20 1085ndash1086 doi 101111j1095-8649200701308x
Frontiers in Marine Science | wwwfrontiersinorg 26 April 2019 | Volume 6 | Article 87
fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87

fmars-06-00087 March 29 2019 Time 1853 27
Edwards et al Advancing Research for Long-Lived Species
Wilson E E and Wolkovich E M (2011) Scavenging how carnivores andcarrion structure communities Trends Ecol Evol 26 129ndash135 doi 101016jtree201012011
Yano K Stevens J D and Compagno L J (2004) A review of thesystematics of the sleeper shark genus Somniosus with redescriptions ofSomniosus (Somniosus) antarcticus and Somniosus (Rhinoscymnus) longus(Squaliformes Somniosidae) Ichthyol Res 51 360ndash373
Yano K Stevens J D and Compagno L J V (2007) Distributionreproduction and feeding of the Greenland shark Somniosus (Somniosus)microcephalus with notes on two other sleeper sharks Somniosus(Somniosus) pacificus and Somniosus (Somniosus) antarcticus J fish Biol 70374ndash390
Conflict of Interest Statement The authors declare that the research wasconducted in the absence of any commercial or financial relationships that couldbe construed as a potential conflict of interest
Copyright copy 2019 Edwards Hiltz Broell Bushnell Campana Christiansen DevineGallant Hedges MacNeil McMeans Nielsen Praeligbel Skomal Steffensen WalterWatanabe VanderZwaag and Hussey This is an open-access article distributedunder the terms of the Creative Commons Attribution License (CC BY) The usedistribution or reproduction in other forums is permitted provided the originalauthor(s) and the copyright owner(s) are credited and that the original publicationin this journal is cited in accordance with accepted academic practice No usedistribution or reproduction is permitted which does not comply with these terms
Frontiers in Marine Science | wwwfrontiersinorg 27 April 2019 | Volume 6 | Article 87





























