Embed Size (px)
Citation preview

Review
ISSN 1743-588910.2217/NNM.10.80 © 2010 Future Medicine Ltd Nanomedicine (2010) 5(7), 1103–1127 1103
Advancing nonviral gene delivery: lipid- and surfactant-based nanoparticle design strategies
BackgroundGene therapy is a medical technique used to repair genetic defects by inserting a normal or functional gene to replace or correct a faulty gene, to turn off or repress an overexpressing gene, or to introduce a desirable function in a host. Successful transfer of genes into cells must first take place in order to achieve these goals. To deliver genes, which often involves transfec-tion with plasmid DNA, the DNA must first be packaged in a viral [1–4] or nonviral [5–10] deliv-ery system. Virus-based delivery is currently more effective and has resulted in the world’s first commercially available gene therapy treat-ment, Gendicine® (Shenzhen Sibiono Bentech, China), an adenoviral gene therapy approved in 2003 by regulatory agencies in China as a treatment for head and neck cancer [11,12]. This treatment uses an injectable form of an adenovi-ral vector for the p53 tumor suppressor gene [13]. A recent success in viral gene delivery, reported in 2009 by Cartier et al. [14], demonstrated the treatment of X-linked adrenoleukodystrophy (ALD), which is caused by an ALD protein deficiency encoded by the ABCD1 gene. In this trial, autologous CD34+ cells, removed from the patient and genetically corrected ex vivo with a lentiviral vector encoding the ALD protein, were re-infused into the patients, who also had myeloablative treatment. This led to restored ALD protein expression, a successful therapeutic outcome.
Prior to a study conducted by Felgner and colleagues [15], the first in which a gene was delivered by a nonviral cationic lipid vector, research on transfection focused mainly on the use of viral vectors. Although the majority of gene therapy clinical trials use viral vectors [301], there is serious concern that their use can elicit a strong immune response [1,16,17]. They are also costly to produce and have the potential for large-scale contamination during produc-tion [5]. Tragic events have occurred, including the death of a clinical trial patient in 1999 who was treated with an engineered adenovirus vec-tor that expressed the human ornithine carbam-oyltransferase (OTC) gene [17]. The death was linked to an unexpected severe immune response triggered by viral peptides and proteins [18]. In another case, a French team administered a ret-rovirus to treat 11 children with severe immu-nodeficiency in an ex vivo clinical trial. This led to a major therapeutic setback as the treatment resulted in a blood disorder similar to leuke-mia in two of the children [18]. Due to these risks, emphasis is shifting to the development of nonviral vectors. Nonviral delivery systems have significant advantages over viral delivery systems, as they are safer to use [19], easier to prepare and can transfect larger sized plasmids into cells. However, nonviral delivery systems with high transfection efficiency must be devel-oped in order to realize their application in gene therapy [20,21].
Gene therapy is a technique utilized to treat diseases caused by missing, defective or overexpressing genes. Although viral vectors transfect cells efficiently, risks associated with their use limit their clinical applications. Nonviral delivery systems are safer, easier to manufacture, more versatile and cost effective. However, their transfection efficiency lags behind that of viral vectors. Many groups have dedicated considerable effort to improve the efficiency of nonviral gene delivery systems and are investigating complexes composed of DNA and soft materials such as lipids, polymers, peptides, dendrimers and gemini surfactants. The bottom-up approach in the design of these nanoparticles combines components essential for high levels of transfection, biocompatibility and tissue-targeting ability. This article provides an overview of the strategies employed to improve in vitro and in vivo transfection, focusing on the use of cationic lipids and surfactants as building blocks for nonviral gene delivery systems.
KEYWORDS: active targeting n DNA n gemini surfactant n gene therapy n lipoplexes n nonviral delivery systems n soft nanoparticles
McDonald Donkuru1, Ildiko Badea1, Shawn Wettig2, Ronald Verrall3, Mahmoud Elsabahy2 & Marianna Foldvari†1,2
1College of Pharmacy & Nutrition, University of Saskatchewan, Saskatoon, SK, S7N 5C9, Canada 2School of Pharmacy, Faculty of Science, University of Waterloo, Waterloo, ON, N2L 3G1, Canada 3Department of Chemistry, University of Saskatchewan, Saskatoon, SK, S7N 5C9, Canada†Author for correspondence: [email protected]
For reprint orders, please contact: [email protected]

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1104
Considerable research into nonviral gene deliv-ery has been conducted in the last two decades on synthetic soft materials such as cationic lipids, polymers, dendrimers and surfactants, and efforts are intensifying [6,9,20,22–25]. An effective nonviral gene delivery system must provide a means of binding and condensing DNA [26,27], protecting DNA from degradation by serum and extracel-lular nucleases, a mechanism for localizing DNA at the cell membrane [28,29], followed by translo-cation through the membrane [30,31], endosome disruption, and subsequent nuclear localization before transcription to mRNA and, ultimately, translation into the therapeutic protein. A sche-matic illustration of a hypothetical nanoparticle with a potentially effective c ombination of these components is shown in Figure 1.
Currently recognized barriers to gene deliveryThe mechanism of gene delivery by both viral and nonviral delivery systems is being investi-gated rigorously to gain a better understanding
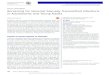
of the requirements for high transfection effi-ciency. Figure 2 outlines the steps required for suc-cessful nonviral gene delivery and expression. The initial step is binding of the DNA/lipid complex to the cell. Both specific and nonspe-cific binding can trigger internalization of the lipoplex [32]. This is followed by cellular internal-ization of lipoplexes by clathrin- and/or cavolae/lipid raft-mediated endocytosis, with temporal bagging of lipoplexes in endosomes [33,34]. The endocytotic pathway is believed to account for most of the DNA entering cells [35–37]. Endocytosis is mediated by proteoglycan inter-actions [38,39] and can be increased several-fold by ligand–receptor interaction [40–42]. Endocytosis of nontargeted cationic lipoplexes is relatively weak. Fusion between cells and lipoplexes, in which the contents of a lipoplex become part of a cell to which it is fused, is a secondary path-way of lipoplex internalization [43,44]. However, reports suggest that cell–lipoplex fusion accounts for a small fraction of o verall lipoplex internalization [45].
Figure 1. Building blocks of nonviral DNA delivery systems. The bottom-up approach combines components essential for efficient cellular transfection. A variety of cationic lipids can be used to construct lipid-based systems, such as liposomes, lipoplexes and solid lipid particles. Use of pH-sensitive lipids, surfactants or polymers can enhance endosomal escape. Use of PEG lipids augments biological and colloidal stability of nanoparticles from steric hindrance due to the inert, nonionic, hydrophilic and flexible polymer coating. Specific ligands to cell surface receptors increase the specificity of binding of nanoparticles to cells. Placing the ligand at the far end of the polymer chain can extend the ligand’s reach to nearly the full extended length of the polymer, thus an approaching lipoplex can be sensed by a receptor from afar. A nuclear-targeting ligand can enhance the nuclear localization of DNA. PEG: Polyethylene glycol.
Ligands targeted to cell surface receptors to increase binding specificity
Nuclear localization signalsto increase nuclear targeting
Stealth groups (e.g., PEG chains)to reduce binding of soft DNAnanoparticles to nontargeted cells and prolong circulation time
++ +
+
+
+
+
+
+
++
+
+
+
+
+
Cationic lipids as basic building blocks to neutralize, condense and encapsulateDNA into soft nanoparticles
+ pH-sensitive lipids, surfactants or polymers to enhance endosomal escape
+ other lipids to augmentbiological and colloidal stability and/or to impart fusogenic properties
www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1105
DNA complexes internalized into endosomes must be released into the cytoplasm for transfer into the nucleus. DNA escape before the endo-somes fully mature into acidified lysosomes, leading to DNA destruction, can be facilitated by pH-sensitive or fusogenic components of the nanoparticle. Once in the cytoplasm, the free DNA or the lipoplex must be imported into the nucleus [46], where the therapeutic gene is tran-scribed to mRNA. Translation of mRNA into therapeutic proteins occurs in the cytoplasm. An ideal nanoparticle would have a specific function for each step of the transfection process. Figure 3
shows the schematic for such a nanoparticle. In the next section, the characteristics of nanopar-ticle components that can confer these functions are described.
Building blocks for lipid-based DNA nanoparticles�n Lipids
A wide variety of cationic lipids have been described as nucleic acid delivery agents (see recent reviews [47–50]). Cationic lipids spontane-ously form vesicles, aggregates or specific types of complex supramolecular structures, as well as condensing and encapsulating DNA into par-ticles readily internalized by cells. Entrapment of DNA presents an interface for interactions with a host cell and its organelles and can protect DNA from degradation by endonucleases [51]. Cationic lipids also contribute an overall net positive charge to the DNA complex, which appears to be important to the electrostatic interaction with the negatively charged cell sur-face. The fine tuning of the cationic to anionic
charge ratio (i.e., +/- ratio) has an important role in transfection efficiency [52,53]. Interaction of DNA complexes with cells occurs in a series of steps and creates a pathway for DNA to reach the nucleus. The chemical structure of the cationic lipids has been the main focus of extensive modi-fications. Rational design principles and quan-titative structure–activity relationship model-ing has been performed to elucidate important structural characteristics of cationic lipids that facilitate more effective transfection [50].
The first cationic lipid system, DOTMA/dioleylphosphatidylethanolamine (DOPE), shown to deliver DNA in vitro [15], is commer-cially available as Lipofectin®. It contains the fundamental features common to such delivery systems, namely a monocationic head and two lipidic tails that enable neutralization, condensa-tion and encapsulation of DNA into lipoplexes. The DOTMA/DOPE lipid system has inspired the development of several lipid combinations with structural features that introduce advanced characteristics such as pH-sensitivity [54], poly-morphism, targeted delivery due to ligand–recep-tor interaction, and high-affinity lipid–DNA interaction and condensation due to polycationic heads [55]. About 115 lipid/surfactant-based and polymer-based transfection reagents are commer-cially available from approximately a dozen ven-dors, although the molecular identities of many are protected by patents, making it difficult to determine structural differences and obtain a total figure of chemically different varieties.
The most widely examined cationic lipids belong to one of the following groups: mono-cationic lipids with ammonium head groups
Figure 2. Intracellular delivery of DNA lipoplexes. The process is initiated with the binding of the lipoplex to the cell surface (A), followed by internalization into endosomes (B). Endosomal escape of the lipoplex (C) is necessary for the subsequent release of the plasmid with dissociation of the carrier molecules and uptake into the nucleus (D). Transcription of the therapeutic gene in the nucleus (E) is followed by its translation in the cytoplasm (F) and expression of therapeutic protein (G).
5´5´3´
3´
Nanomedicine © Future Science Group (2010)

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1106
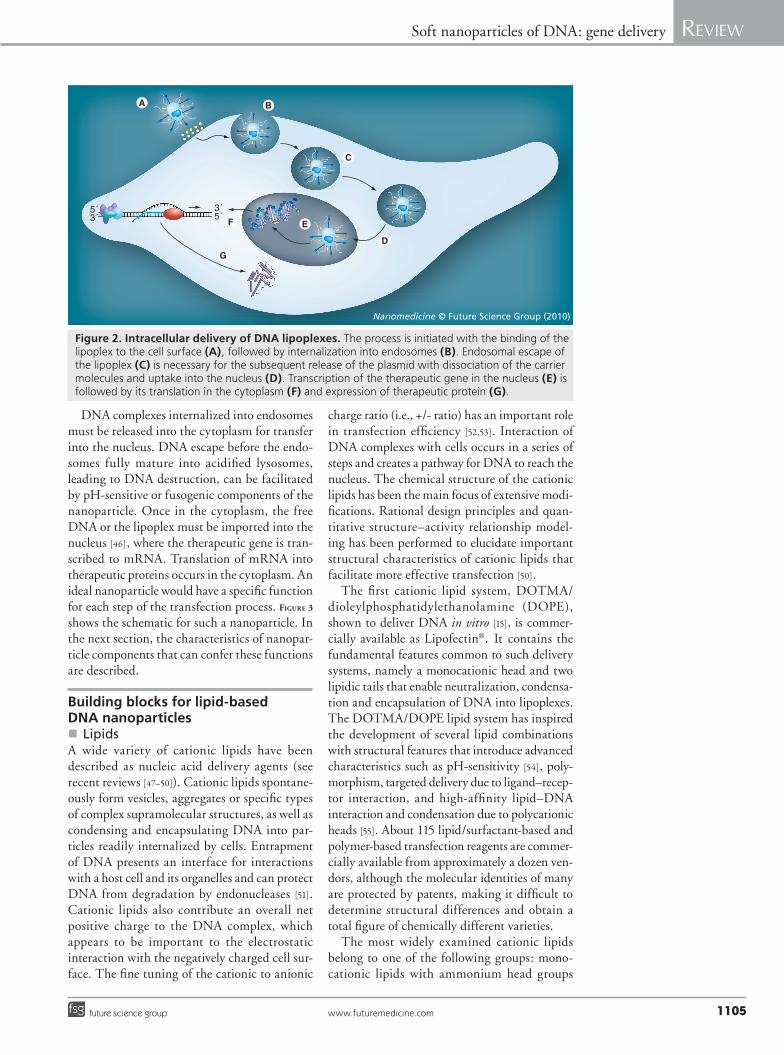
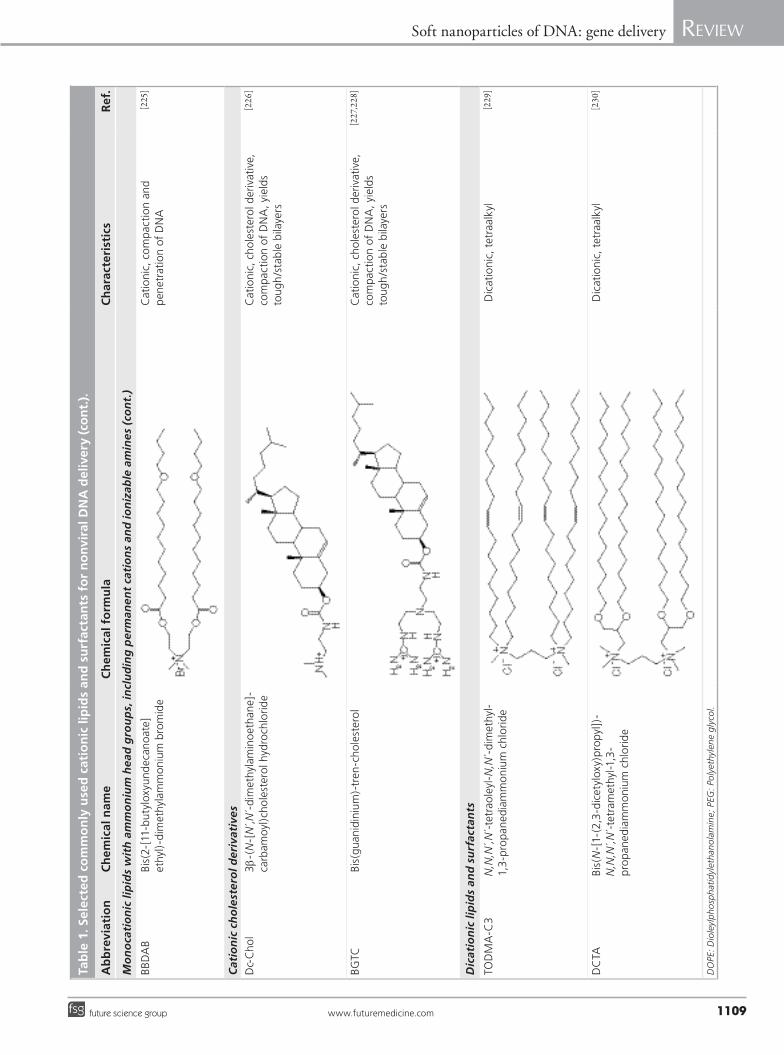
(including permanent cations and ionizable amines); cationic cholesterol derivatives; dica-tionic lipids and surfactants; polycationic lipids; and cationic lipid hybrids with sugar, peptide and polymer residues (Table 1).
Lipofectamine (DOSPA/DOPE) and Cell-fectin (TM-TPS (N, N´, N´́ , N´́ -́tetramethyl-N, N´, N´´, N´́ -́tetra palmitylspermine)/DOPE) are commercially available DNA transfection lipid formulae that possess pH-sensitivity due to the polyamine head of the cationic lipid components. They also show potential poly-morphic behavior based on the ability of the neutral lipid DOPE (replaced by cholesterol in some cases) to induce lamellar-to-inverted-hex-agonal phase transitions [56]. Recent molecular design approaches have led to the development of better transfection molecules with efficient DNA condensation and pH-sensitivity effects, such as the RPR lipid family [57,58] and the MVL/MVLBG lipid family [59,60]. Additional effects such as lamellar-to-noninverted micel-lar phase transition produced by sugar-based gemini surfactants [61], cell targeting and nuclear localization achieved with peptide-based gemini surfactants [62], bilayer-to-micelle transition achieved with carbohydrate-based gemini surfactant [61,63], and reduced toxic-ity to cells achieved with amino acid- and
carbohydrate-conjugated gemini surfactants [62] also mark recent progress in the search for more efficient and b iocompatible transfection agents.
Several synthetic, positively charged 2,3-dial-kyloxypropyl quaternary ammonium com-pounds have been found to increase transfec-tion efficiency of plasmid DNA in vivo [64–66]. Plasmid DNA–alkylphospholipid lipoplexes containing dioctadecyldimethylammonium bromide (DODAB) injected intratumorally significantly reduced tumor size compared with injection of naked DNA and Lipofectin–DNA complexes [67–69]. In addition to the classi-cal approach of using monovalent quaternary ammonium compounds, such as DODAB and 1,2-dioleoyl-3-(trimethylammonium) propane (DOTAP), and cationic derivatives of choles-terol, new compounds such as reducible cationic lipids, disulfide-bond bearing lipids, acid-labile cationic lipids and glycine betaine derivatives have also been developed to overcome barriers to DNA delivery [70]. Reducible cationic lipids with disulfide bonds improve the release of DNA in the cytoplasm, where the bond is cleaved. This cleavage is hypothesized to aid DNA release, and up to 1000-fold increase in transfection effi-ciency has been reported for disulfide-contain-ing delivery agents [57]. Cationic lipids with acid-labile vinyl ether linkages are stable at neutral pH, but degrade in acidic endosomes, promot-ing the escape of DNA into the cytoplasm and increasing transfection two- to three-fold com-pared with the DOTMA–cholesterol system. The cationic cholesterol-derivative (Dc-Chol), formulated in liposomes or emulsions, was used to transfer DNA into cells. The advantage of this compound lies in its high transfection efficiency and low cellular toxicity in vitro. The presence of serum reduced the transfection efficiency of the liposomal formulation, but had no effect on the emulsion-type formulations [71]. The transfec-tion ability of the Dc-Chol emulsion in the pres-ence of serum makes these types of formulations suitable for parenteral administration. Another cationic cholesterol-derivative, dexamethasone–spermine, increased DNA delivery in vitro ten-fold, compared with Lipofectamine, at a +/- charge ratio of 6. The advantage of the system in vivo was the intrinsic anti-inflammatory prop-erty of the corticosteroid [72]. Ultradeformable cationic liposomes prepared with DOTAP had an in vitro transfection efficiency comparable with Lipofectamine, and were able to deliver plasmid DNA to the liver, kidneys and lungs after transdermal application [73]. Some cationic lipids, especially monovalent lipids, are relatively
Figure 3. Physicochemical characteristics of nanoparticles deemed important in DNA delivery and transfection. Several of these properties play a role in the extent of transfection efficiency of lipoplexes. Structural flexibility and polymorphic transformation of bilayer arrangements in lipoplexes are important at several stages of lipoplex uptake and intracellular processing. Various degrees of lamellar-to-hexagonal-to-cubic transformations may take place.
++ +
+
+
+
+
+
+
++
+
+
+
+
+
Internal structure andstructural flexibility
Surface charge andcharge density
Particle shape andaspect ratio
Size and sizedistribution

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1107
cytotoxic so their use in vivo is limited. Natural glycine betaine compounds covalently linked to acyl chains by enzymatically hydrolyzable pep-tide and ester bonds showed reduced cytotoxic-ity and improved 40% transfection efficiency compared with commercial agents in cultured hepatocytes [74].
�n Helper lipidsThe transfection efficiency of cationic lipids can be enhanced by incorporating helper lipids into the delivery system. Helper lipids bear a neutral charge and help build liposomes that are stable, fusogenic, enhance tissue penetrantion, and help initiate breakout of DNA from endosomes following uptake of liposome-encapsulated DNA into a cell. Therefore, when added to cat-ionic lipid–DNA complexes, a helper lipid can increase the stability of the delivery system and increase its in vitro and in vivo transfection effi-ciency. DOPE is by far the most extensively used helper lipid and its usefulness is attributed to membrane-active properties, namely the ability to induce hexagonal lipid conformation [75–79]. DOPE has been combined with a variety of cationic lipids, such as DOTAP, DOTMA, DODAC, DOGS, DMRIE [53,80–87], cholesterol derivatives [88,89] and poly-l-lysine (PLL) [90,91] to increase transfection. Injected intramuscu-larly, DOTAP-containing liposomes entrap-ping plasmid DNA encoding for the hepatitis B surface antigen triggered antigen-specific IgG production at significantly higher levels than did intramuscular injection of naked DNA. The magnitude of the immune response depended on the nature of the helper lipid, the largest being with DOPE–DOTAP vesicles with a component of phosphatidylcholine, compared with the same vesicles having a component of d ipalmitoylphosphatidylcholine (DPPC) or c holesterol [92].
A comparison of several helper lipids such as dilauroylphosphatidylethanolamine (C12:0), dimiristoylphosphatidylethanolamine (C14:0), dipalmitoylphosphatidylethanolamine (C16:0), diphytanoylphosphatidylethanolamine (C16:0 branched), distearoylphosphatidylethanolamine (DSPE, C18:0) and DOPE (C18:1) showed that the branched and unsaturated species combined with cationic lipids act in physical synergism to increase transfection efficiency [93].
�n Stealth groupsSteric stabilization is usually required to avoid the rapid clearance of the DNA–lipid complexes by the mononuclear phagocyte system and is usually
achieved by grafting hydrophilic polymers (e.g., polyethylene glycol [PEG]) to the surface of the liposomes. Coating with nonionic hydrophilic polymers prevents the adsorption of opsonins, thereby limiting the rapid uptake by the mono-nuclear phagocyte system and prolonging the circulation half-life of the encapsulated genetic material. Liposomes bearing PEG chains on their surface, the so-called stealth liposomes, were first used for gene delivery in the early 1990s. A sig-nificant increase in the circulation time of the intravenously injected stealth liposome-encap-sulated genetic material (2.3 days), compared with unencapsulated oligonucleotides (1 h), was observed [94]. Although the liposomes could protect the DNA from nucleases in the blood and tissues [94], the transfection efficiency of the stealth particles appeared to be compromised by the lack of direct exposure of the cationic lipids. Since cationic lipid interaction with the proteo-glycans on the target cell [95] is necessary for cel-lular uptake, the long PEG chains on the surface of the cationic lipoplexes reduced their ability to bind to cells [96]. To promote the release of DNA from cationic complexes, PEG was replaced with an acid-labile PEG diorthoester conjugate of monomethyl-PEG 2000 and distearoylglycerol (PODS2000), which improved transfection effi-ciency without i nterfering with the stealth nature of the particle [97].
�n Components to enhance endosomal escapeEndosomal escape of lipoplexes must occur before the endosomes become hydrolytic (low pH) or ultimately fuse with lysosomes. Though DNA is protected in lipoplexes, its degradation is inevitable unless it escapes from the endo-some [35]. In reality, only a fraction of lipoplexes reach the nucleus, the rest being destroyed in the destructive, hydrolytic phase of endosomes and lysosomes [98]. As such, assisted endosomal escape by using lipid systems capable of bilayer-to-micelle or lamellar-to-inverted hexagonal (H
II) transition is helpful [61,99–101].
Additional design involves introduc-ing sensitive and cleavable chemical bonds such as disulf ide bonds, occuring in the –CH
2–S–S–CH
2– unit [102], or pH-sensitive
groups such as imidazole or pyridine [103,104], which are protonated in the endosomal pH range (pH 4.0–7.4) [105,106]. A disulfide bond placed between the head group and tail of a lipid, between a hydrophilic PEG chain and the hydrophobic lipid domain or at another position, allows for cleavage in an acidic environment,

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1108
Tab
le 1
. Sel
ecte
d c
om
mo
nly
use
d c
atio
nic
lip
ids
and
su
rfac
tan
ts f
or
no
nvi
ral D
NA
del
iver
y.
Ab
bre
viat
ion
Ch
emic
al n
ame
Ch
emic
al f
orm
ula
Ch
arac
teri
stic
sR
ef.
Mo
no
cati
on
ic li
pid
s w
ith
am
mo
niu
m h
ead
gro
up
s, in
clu
din
g p
erm
anen
t ca
tio
ns
and
ion
izab
le a
min
es
CTA
B H
exad
ecyl
trim
ethy
lam
mon
ium
br
omid
eC
atio
nic,
sin
gle
chai
ned
[220]
DO
DA
B,D
OD
AC
Dim
ethy
ldio
ctad
ecyl
amm
oniu
m
brom
ide
or c
hlor
ide
Cat
ioni
c, d
oubl
e ch
ain
ed[221,222]
DO
TMA
N
-{1-
(2,3
-dio
leyl
oxy)
pro
pyl}
-N,N
,N-
trim
ethy
lam
mon
ium
chl
orid
eC
atio
nic,
dou
ble
chai
ned
, eth
er
links
, com
pact
ion
and
pen
etra
tion
of
DN
A
[15]
DO
TAP
N-{
1-(2
,3-d
iole
oylo
xy)p
ropy
l}-N
,N,N
-tr
imet
hyla
mm
oniu
m c
hlor
ide
Cat
ioni
c, d
oubl
e ch
ain
ed, e
ster
lin
ks, c
ompa
ctio
n an
d p
enet
rati
on
of D
NA
[223]
DM
RIE
Dim
yris
tylo
xypr
opy
l-3-d
imet
hyl-
hydr
oxye
thyl
am
mon
ium
bro
mid
eC
atio
nic,
dou
ble
chai
ned
, co
mpa
ctio
n an
d p
enet
rati
on
of D
NA
[224
]
BPD
AB
Bis(
2-[1
1-ph
enox
yund
ecan
oat
e]et
hyl)
dim
ethy
lam
mon
ium
bro
mid
eC
atio
nic,
com
pact
ion
and
pen
etra
tion
of
DN
A
[225]
HA
DA
BN
-hex
adec
yl-N
-(10
-[O
-(4
-ace
toxy
) ph
enyl
-und
ecan
oat
e]et
hyl)
di
met
hyla
mm
oniu
m b
rom
ide
Cat
ioni
c, c
ompa
ctio
n an
d p
enet
rati
on o
f D
NA
[225]
DO
PE: D
iole
ylp
hosp
hati
dyl
etha
nola
min
e; P
EG: P
oly
ethy
lene
gly
col.
www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1109
Tab
le 1
. Sel
ecte
d c
om
mo
nly
use
d c
atio
nic
lip
ids
and
su
rfac
tan
ts f
or
no
nvi
ral D
NA
del
iver
y (c
on
t.).
Ab
bre
viat
ion
Ch
emic
al n
ame
Ch
emic
al f
orm
ula
Ch
arac
teri
stic
sR
ef.
Mo
no
cati
on
ic li
pid
s w
ith
am
mo
niu
m h
ead
gro
up
s, in
clu
din
g p
erm
anen
t ca
tio
ns
and
ion
izab
le a
min
es (
con
t.)
BB
DA
BBi
s(2-
[11-
buty
loxy
und
ecan
oat
e]et
hyl)
-dim
ethy
lam
mon
ium
bro
mid
eC
atio
nic,
com
pact
ion
and
pen
etra
tion
of
DN
A
[225]
Cat
ion
ic c
ho
lest
ero
l der
ivat
ives
Dc-
Cho
l3b-
(N-[
N,́N
´-di
met
hyla
min
oet
han
e]-
carb
amoy
l)ch
ole
ster
ol h
ydro
chlo
rid
eC
atio
nic,
cho
lest
ero
l der
ivat
ive,
co
mpa
ctio
n of
DN
A, y
ield
s
toug
h/s
tabl
e bi
laye
rs
[226
]
BGTC
Bis(
guan
idin
ium
)-tr
en-c
hole
ster
ol
Cat
ioni
c, c
hole
ster
ol d
eriv
ativ
e,
com
pact
ion
of D
NA
, yie
lds
to
ugh
/sta
ble
bila
yers
[227,228]
Dic
atio
nic
lip
ids
and
su
rfac
tan
ts
TOD
MA
-C3
N,N
,N´,N
´-tet
rao
leyl
-N,N
´-di
met
hyl-
1,3
-pro
pan
edia
mm
oniu
m c
hlor
ide
Dic
atio
nic,
tet
raal
kyl
[229]
DC
TABi
s(N
-[1-
(2,3
-dic
etyl
oxy)
pro
pyl]
)-N
,N,N
´,N´-t
etra
met
hyl-1
,3-
pro
pan
edia
mm
oniu
m c
hlor
ide
Dic
atio
nic,
tet
raal
kyl
[230]
DO
PE: D
iole
ylp
hosp
hati
dyl
etha
nola
min
e; P
EG: P
oly
ethy
lene
gly
col.

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1110
Tab
le 1
. Sel
ecte
d c
om
mo
nly
use
d c
atio
nic
lip
ids
and
su
rfac
tan
ts f
or
no
nvi
ral D
NA
del
iver
y (c
on
t.).
Ab
bre
viat
ion
Ch
emic
al n
ame
Ch
emic
al f
orm
ula
Ch
arac
teri
stic
sR
ef.
Dic
atio
nic
lip
ids
and
su
rfac
tan
ts (
con
t.)
BCA
TO
-(2R
-1,2
-di-
O-[
1’Z,
9
’Z-o
ctad
ecad
ieny
l]-g
lyce
rol)
-3-N
-(bi
s-2-
amin
oet
hyl)
-car
bam
ate
Cat
ioni
c, a
cid
-lab
ile, i
ncre
ase
end
oso
mal
esc
ape
[231]
GA
P-D
LRIE
N-(
3-a
min
opr
opy
l)-N
,N-d
imet
hyl-2
,3-
bis(
do
dec
ylox
y)-1
-pro
pani
mid
ium
br
omid
e
Cat
ioni
c, c
ompa
ctio
n an
d p
enet
rati
on o
f D
NA
[232]
Poly
cati
on
ic li
pid
s
DPP
ES1,
2-di
palm
itoyl
-sn
-gly
cero
-3-
pho
spho
etha
nola
mid
osp
erm
ine
Poly
cati
onic
, com
pact
ion
and
pen
etra
tion
of
DN
A
[233]
DO
SPA
2,3
-dio
leyl
oxy-
N-(
2-[s
per
min
ecar
box
yam
ido
]eth
yl)-
N,N
-di
met
hyl-1
-pro
pani
min
ium
bro
mid
e
Poly
cati
onic
, com
pact
ion
and
pen
etra
tion
of
DN
A
[222
,234]
RPR
sRe
duci
ble
lipid
sPo
lyca
tion
ic, r
educ
tion
-sen
siti
ve
disu
lfid
e b
ond,
impr
oved
DN
A
rele
ase
in t
he c
yto
plas
m
[57]
DO
PE: D
iole
ylp
hosp
hati
dyl
etha
nola
min
e; P
EG: P
oly
ethy
lene
gly
col.

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1111
Tab
le 1
. Sel
ecte
d c
om
mo
nly
use
d c
atio
nic
lip
ids
and
su
rfac
tan
ts f
or
no
nvi
ral D
NA
del
iver
y (c
on
t.).
Ab
bre
viat
ion
Ch
emic
al n
ame
Ch
emic
al f
orm
ula
Ch
arac
teri
stic
sR
ef.
Poly
cati
on
ic li
pid
s (c
on
t.)
DO
GS
Dio
ctad
ecyl
amid
ogl
ycyl
sper
min
ePo
lyca
tion
ic, c
ompa
ctio
n an
d p
enet
rati
on o
f D
NA
[235]
Gua
nidi
n-y
late
d d
eriv
ativ
esN
,N-d
i-n
-alk
yl-N
-(2-
guan
idin
yl)
ethy
l-N
-met
hyl a
mm
oniu
m c
hlor
ide;
alky
l = h
ydro
carb
on c
hain
, ≥ C
14
Poly
cati
onic
, com
pact
ion
and
pen
etra
tion
of
DN
A
[236
]
Cat
ion
ic li
pid
hyb
rid
s w
ith
su
gar
, pep
tid
e an
d p
oly
mer
res
idu
es
Mon
osa
ccha
rid
e-co
njug
ated
gem
ini
surf
acta
nts
Car
bo
hydr
ate-
base
d
gem
ini s
urfa
ctan
tsD
NA
com
plex
atio
n, a
ble
to in
duce
bi
laye
r-to
-mic
elle
tra
nsit
ion
at
phys
iolo
gica
l pH
, low
er t
oxic
ity
due
to n
atur
al m
anno
se o
r gl
uco
se u
nit
[237]
Pept
ide-
conj
ugat
ed g
emin
i su
rfac
tant
s
Pept
ide-
base
d g
emin
i sur
fact
ants
DN
A c
ompl
exat
ion,
cel
l tar
get
ing,
nu
clea
r lo
caliz
atio
n du
e to
p
epti
de
sequ
ence
[238
,239]
DO
PE: D
iole
ylp
hosp
hati
dyl
etha
nola
min
e; P
EG: P
oly
ethy
lene
gly
col.

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1112
causing bilayer destabilization/disruption [64,107] via bilayer-to-micelle transition, aiding release of lipoplexes from endosomal compartments. Similarly, a lamellar-to-inverted hexagonal (H
II)
phase transition occuring in DOPE-containing bilayers [61,101] promotes release of endosome contents into the cytoplasm [61,63].
Fusogenic peptides (composed of short sequences of amino acids containing glutamates) derived from the influenza virus, hemagglutin, increased the transfection efficiency of lipo-fectamine by three- to four-fold for the integrin receptor-targeted delivery of the PLL–mollos-sin–DNA vector. The mechanism of action probably relies on the more effective escape of the complexes from endocytic vesicles [108,109].
�n Cell-targeting ligands & improved cellular uptakeAdvances in restoring transfection efficacy of PEGylated lipoplexes have been reviewed in the literature [110]. One strategy involves using ligand–receptor interaction [111,112] to direct lipoplexes to specific recognition molecules (sites) such as glycoproteins embedded in cell membranes [113]. Incorporation of folate-coupled PEG-lipid into cationic liposomes (an RPR 120535:DOPE coformulation) has been used to restore gene transfer to M109 murine lung carcinoma cells, which express folate receptors, compared with 4T1 breast cancer control cells, that do not express folate receptors. In addition to this finding, targeted PEGylated lipoplexes showed restored transfection activity compared with their nontargeted counterparts [114].
The first step in efficient gene expression is efficient transport of the macromolecule through the cell membrane. Peptides, such as sequences of Tat protein from HIV and adenine nucleotide translocator protein, can promote cellular uptake of macromolecules. Poly(3-guanidinopropyl methacrylate), a polycationic agent that mimics the effect of these peptides, was able to deliver DNA into cells at higher efficiency than polyarginine-based agents. This compound can introduce genes into cells due to its cell-penetration and DNA-compaction prop-erties [115]. Another example of increased cellular uptake is observed with fusogenic liposomes for-mulated with anionic 1,2-dioleyl-phosphoeth-anolamine-N-dodecanoyl (N-C12-DOPE) and DOPC (1,2-dioleoyl-sn-glycero-3-phosphocho-line) encapsulating spermine-compacted DNA. These complexes efficiently transfected ovarian cancer cells (OVCAR-3) in vitro. In addition, they showed significantly lower toxicity than Ta
ble
1. S
elec
ted
co
mm
on
ly u
sed
cat
ion
ic li
pid
s an
d s
urf
acta
nts
fo
r n
on
vira
l DN
A d
eliv
ery
(co
nt.
).
Ab
bre
viat
ion
Ch
emic
al n
ame
Ch
emic
al f
orm
ula
Ch
arac
teri
stic
sR
ef.
Cat
ion
ic li
pid
hyb
rid
s w
ith
su
gar
, pep
tid
e an
d p
oly
mer
res
idu
es (
con
t.)
Pept
ide-
conj
ugat
ed g
emin
i su
rfac
tant
s
Pept
ide-
base
d g
emin
i sur
fact
ants
DN
A c
ompl
exat
ion,
cel
l tar
get
ing,
nu
clea
r lo
caliz
atio
n du
e to
p
epti
de
sequ
ence
[240
]
DO
PE-p
epti
de-
PEG
A li
pid
exam
ple,
wit
h D
OPE
, p
epti
de
and
PEG
com
pon
ents
in it
s cu
stom
ized
str
uctu
re
Ass
istin
g in
DN
A c
ompl
exat
ion,
ce
ll ta
rget
ing,
nuc
lear
loca
lizat
ion,
bi
laye
r ef
fect
s e.
g., l
amel
lar
to
inve
rted
hex
agon
al, e
xten
ded
ha
lf-liv
e in
viv
o du
e to
PEG
-lin
ked
stea
lth
bila
yers
[240
]
DO
PE: D
iole
ylp
hosp
hati
dyl
etha
nola
min
e; P
EG: P
oly
ethy
lene
gly
col.

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1113
cationic delivery agents [116]. Tissue-specific delivery can be achieved with terplex DNA, created by complexation of plasmid DNA with stearyl-PLL and low-density lipoprotein. Using this system, increased cellular delivery into the heart and elevated protein expression for up to 30 days were observed. In comparison, after treatment with naked DNA, levels of protein were significantly lower and expression lasted for only 5 days [117]. Similarly, after systemic administration in mice of terplex DNA, protein expression persisted in major organs for 35 days, versus only 7 days after administration of naked DNA [118]. Mannose receptor-mediated gene transfer, a macrophage targeting method, was carried out by intravenously injecting plasmid DNA complexed with a mannosylated choles-terol derivative. These complexes selectively accumulated in the liver, as measured by the amount of 32P-labelled DNA, and achieved the highest protein expression compared with nonmannosylated complexes [119]. Similarly, galactose-modified liposomes were also used to transfect DNA into HepG2 and HuH-6 cells, which express asialoglycoprotein receptors, resulting in a three- to four-fold rise in trans-fection efficiency in theses cells compared with the negative controls [120]. Lipid-linked target-ing peptides (e.g., arginine-glycine-aspartic acid containing peptide-DOPE), selected to deliver genes into epithelial cells, were able to increase transfection efficiency of the DOTMA–choles-terol–DNA complexes tenfold. This improve-ment is important as it demonstrates that spe-cifically designed peptides are able to target certain cell types (e.g., tumor cells) and increase the uptake of DNA, leading to higher transgene expression [121,122].
Estrogen receptor-targeting moieties, such as 17b estradiol [123], attached to the surface of stealth liposomes, could deliver killer genes into breast cancer cells at significantly higher levels than cells lacking estrogen receptors. Similarly, sigma receptor-targeting haloperidol grafted onto lipoplexes increased gene expression ten-fold in tumor cells compared with the nontar-geting lipoplexes [124]. A urokinase plasminogen activator receptor-targeting ligand attached to a lipid and grafted onto stealth liposomes suc-cessfully delivered its load to urokinase plas-minogen activator receptor-positive cells [125]. A combination of the targeting stealth particles with agents that promote endosomal escape and intracellular trafficking can elevate nonviral gene delivery to a level competitive with that of viral vectors.
�n Nuclear-targeting ligands Efficient intracellular traff icking of DNA released from endosomes, another explored approach, is necessary for increased gene expres-sion [126,127]. Intradermal injection of DNA, complexed with high-mobility group box 1 protein (having a DNA binding domain and a nuclear localization signal) and encapsulated into liposomes, increased gene expression tenfold due to enhanced nuclear targeting [128]. After endosomal escape, breakdown of lipoplexes should occur, allowing nuclear entry of DNA. Nuclear entry of unbound, decondensed DNA competes with its rapid degradation by cytoplas-mic nucleases. The timing of the nuclear entry step is critical in enhancing transfection. In both dividing and nondividing cells, translocation of naked DNA into the nucleus is inefficient. Molecules larger than 40 kDa require active transport through the nuclear pore complexes, which provide a shuttle function between the cytoplasm and nucleus [129]. As they lack a rec-ognition sequence, the amount of DNA success-fully entering the nucleus through nuclear pores is often further reduced [127].
Although gene expression occurs after deliv-ery of target-free DNA [35], enhanced gene expression from the use of targeted DNA is possible. Nuclear entry represents a signifi-cant barrier for gene expression in nondividing cells [130]. The first nuclear localization signal was identified in the simian virus 40, large-T antigen, having a sequence of PKKKRK [131]. Similarly, lysine-rich peptides were also iden-tified from Xenopus phosphoprotein nucleo-plasmin (KRPAATKKAGQAKKKK), proto-oncogene c-myc (PAAKRVKLD) and a yeast homeodomain-containing protein MATa2 (NKIPIKDLLNPQ) [132]. Unfortunately, these peptides are more efficient in linear DNA deliv-ery and electrostatic interaction between them results in shielding of the peptide by the DNA, hindering its function [133]. Grafting a PEG spacer onto the surface of the lipoplexes bypasses this deficiency, improving nuclear transport [134]. Another shuttle peptide is the 38-amino acid M9 sequence of the human mRNA-binding protein hn-RNPA1, a hydrophobic peptide hav-ing a transport mechanism distinct from that of nuclear localization signals [135,136]. Other exploits of directed DNA entry into the cell nucleus have been covered in recent reviews [137].
Glucocorticoids facilitate nuclear pore dilation when attached to their receptors, aiding entry of DNA from poly(amidoamine) particles contain-ing triamcinolone acetonide [138]. Cytoplasmic

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1114
receptors (importins) responsible for the trans-port of oligopeptides and other proteins to and from the nucleus (a process known as nuclear localization) are constantly shuttling across the nuclear membrane [139], offering an opportunity to tag recognition molecules, such as oligopep-tide moieties, to DNA. The tagged DNA can then be selected and expressly transported to the nucleus, allowing DNA to escape degrada-tion by cytoplasmic nucleases. Enhanced gene expression via nuclear localization signals has been reported [140,141]. A recent design scheme was reported that exploits NFkB transcription factor, which shuttles between the cytoplasm and the nucleus, to promote an efficient nuclear import of DNA. A designed nucleotide sequence termed 3NF when attached to plasmids allowed the plasmids to be recognized and imported into the nucleus by NFkB, leading to higher gene expression [142]. Nuclear entry can also be increased in dividing cells, as such cells have tran-sient opening of their nuclei. However, this may not be easily employed for routine DNA delivery.
�n Overall remarks on nanoparticle design The design of successful nonviral gene deliv-ery systems follows a bottom-up approach. Starting with basic building blocks such as cationic lipids, polymers, polypeptides or den-drimers, multifunctional complex systems can be engineered. These systems bear molecular modules for cell binding, uptake, endosomal escape, nuclear uptake and, for in vivo applica-tions, mobility within the body. The practical feasibility of the modular design approach will be crucial in the development of nonviral gene delivery systems [143]. For example, using lipids such as DOPE to facilitate DNA uptake can be associated with targeting moieties such as trans-ferrin to promote endocytosis [144]. Fusogenic peptides (e.g., influenza hemagglutin peptide) increase the escape of DNA from endosomes/lysosomes [106,145,146]. Combination with an inte-grin-targeted nonviral DNA vector (PLL–molos-sin) can further improve gene expression [147]. These examples illustrate that multicomponent systems have the potential for efficiency that might approach that of viral delivery systems, but without their high toxicity.
Nanoparticle properties important in transfection The formation of DNA–soft material complexes is usually achieved by simply mixing the com-ponents in a specific order. The DNA plasmid
then undergoes compaction, forming particles of various sizes and morphologies. Figure 4 shows an example of DNA plasmid compaction with the 16-3-16 gemini surfactant [148]. Particle formation is a relatively slow process. In this example, the initial 30 s result in the binding of gemini surfac-tant to DNA, condensing the plasmid into a ‘spa-ghetti and meatballs’ structure, followed by for-mation of mostly round particles within 15 min of initiating mixing. The resulting particles appear to have a rough surface and h eterogeneous size.
In general, many of the physical and physi-cochemical properties of lipoplexes, such as particle size, surface charge density, morphol-ogy or shape and polymorphic structure affect gene delivery both in vitro and in vivo. Evidence of this influence and possible reasons for it are discussed below.
�n Size & size distributionComplex size can vary widely and complexes as large as 2000 nm have been noted [96]. Published studies indicate that small complexes efficiently transfect cells in some cases and large ones in other cases. Lipoplex size depends mainly on fac-tors such as the size of the transgene, efficiency of DNA condensation (determined by +/- ratio and the valency of the transfection vector) and size-controlling measures, including vesicle extrusion, ultrasonication and nanoparticle fractionation through density gradient centrifugation [149–151]. Based on diameter, particles can be categorized as small (≤100 nm), medium (100–200 nm), large (200–1000 nm) or giant (≥1000 nm) [152,153]. The observed differences in transfection efficiency between nanoparticles of different size distribu-tions result not only from how well these par-ticles overcome the membrane barrier, but also how well they overcome external barriers, such as interaction with serum proteins, as well as barri-ers within the cell once they become internalized. Since overall transfection efficiency is assessed by eventual protein expression, the impact of l ipoplex size alone seems to be moderate.
Most transfection complexes fall within an average size range of 100–300 nm. A DNA/ dimethyldioctadecyl ammonium bromide/DOPE system, which embodies complexes within this size range, has transfected cells with high efficiency [154]. Larger complexes formed from a DNA/a-galacto-W-spermine bolaamphiphile/DOPE system, having a size of 1100 nm, also transfected HepG2 cells effi-ciently. However, exclusion of the DOPE com-ponent from this transfection system resulted in a heterogeneous mixture of varying complex

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1115
sizes, 100 to 800 nm, which were inefficient in transfecting HepG2 cells [155]. Despite limited correlation with transfection efficiency, size does play a role in the mode of cellular internaliza-tion of transfection complexes. Effective endo-cytosis has been reported for complexes 100 nm or slightly larger [156–159], whereas significantly larger complexes can pass their contents into cells more easily through cell–lipoplex fusion, suggesting that overall cellular uptake could occur through both pathways.
In most cases transgene delivery and expres-sion differ markedly for nanoparticles of differ-ent size ranges [160–163]. Lack of effect of size on gene transfection has been reported only in a few studies [164]. Such observations are balanced by the fact that although several studies have noted a relationship between nanoparticle size and transfection efficiency, the relationship is not always consistent. For instance, depending on the study, either small [160,161,165] or large [153,166] transfection nanoparticles can be more efficient.
The impact that size has on transfection efficiency is affected by factors such as mode of cellular uptake, endocytotic ability of cells and interaction characteristics along the route of gene delivery encountered in vitro and in vivo. Using cell cultures, numerous studies have found efficient transfection for complexes with sizes falling within the medium-to-large range, but not greater than 400 nm [161,167]. This particle size range is better suited for cell attachment, spontaneous uptake and inducing endosomal disruption to release DNA in actively endocy-tosing cells. Detectable transfection has also been reported for particles up to 1000 nm, although such examples are rare. Particles therefore have an upper size limit, and cells in cell cultures [162,168,169] would not be predicted to take up complexes as large as or larger than themselves. However, there have been reports of rare occurrence of giant lipoplexes with high transfection rates [163,170,171].
Compared with in vitro delivery, small par-ticles tend to have high transfection efficiency in vivo because of the ability of small particles to traverse narrow capillary networks (e.g., in the lungs, spleen, kidney and liver). Large particles typically have low in vivo transfection efficien-cies, thus lipoplex size is a major determinant of lipofection efficiency, although the effect of size may be dominated by other factors such as the cell type.
An additional feature common among many DNA lipoplex systems is a definite heteroge-neity in the size(s) of the nanoparticles, with
two or more distinct populations of particles forming depending on the composition of the system. This is well demonstrated in Figure 4, which shows the cationic gemini surfactant 16–3–16-DOPE condensing plasmid DNA into nanoparticles of two primary size populations – smaller (approximately 148 nm) and larger (approximately 468 nm) particles – as observed by atomic force microscopy and dynamic light scattering studies [148,172].
While few reports are available that specifi-cally address the effect of DNA plasmid size, Pitard’s group [173] and others [174,175] have examined the effect of DNA size on the physi-cochemical properties of lipoplexes and trans-fection efficiency. Nanoparticles formed from RPR lipids and plasmids ranging from 900 to 52,000 base pairs were examined, and transfec-tion efficiencies were observed to decrease with increasing plasmid size. The bilayer morphology
Naked plasmid
30 s after adding 16-3-16
15 min afteradding16-3-16
Co
mp
acti
ng
pla
smid
DN
A in
to n
ano
par
ticl
es
Figure 4. Formation of DNA nanoparticles from the 16-3-16 gemini surfactant. AFM images illustrating the compaction of pGTmCMV.IFN-GFP plasmid DNA (prepared by our group) by gemini surfactant 16-3-16 as a function of time. Bar represents 400 nm. Adapted with permission from [204].

Nanomedicine (2010) 5(7) future science group1116
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
was observed by small-angle x-ray scattering to be lamellar in all cases, irrespective of the size of the plasmid, and the particles consisted of ordered spherical or ellipsoidal microdomains (i.e., the so-called fingerprint pattern).
�n Surface charge & charge densityThe molar or +/- charge ratio and zeta potential of lipoplexes have been repeatedly demonstrated as being important in achieving high transfec-tion efficiencies, with complexes having a net positive charge being important for in vitro transfection. The effect of charge ratio on in vivo transfection efficiency is much less clear, partly because positively charged particles can often be complexed by serum components, thus reducing efficiency [73,176]. Reports have generally indi-cated that a +/- charge ratio of at least 1:1 is nec-essary to ensure complete neutralization of the negatively charged phosphate groups on DNA by the cationic lipid or surfactant; DNA con-densation also occurs and can differ depending on the particular structure of the cationic agent. Transfection efficiency generally increases when the proportion of the cationic agent is increased beyond the amount required for complete neu-tralization, and declines after going through a peak. In practice, the optimal charge ratio can differ depending on the type of cationic agent and the cell or tissue type. For instance, the +/- charge ratio at which optimal transfection was realized using a monocationic agent with a single head group (DPPC) was 5:1, although the agent was highly toxic to cells at this ratio. On the other hand, a dimeric (dicationic) surfactant agent yielded optimal transfection when pre-pared at a +/- charge of ratio of 3:1 [177]. Other studies have shown that optimal transfection of airway epithelial cells occurs at a +/- charge ratio of 1:1, while others have realized optimal transfection of these cells with a +/- ratio of 2.8:1 for a cationic cholesterol derivative, N,N-dimethylaminpropyl-amidosuccinyl-cholesteryl-formylhydrazide [52]. These two findings, espe-cially the latter, are in contrast with a +/- ratio of 1.25:1 that was found to optimize transfection of epithelial cells using DMRIE [52,177].
At constant pH, the +/- charge ratio of lipo-plexes is a key factor governing the first step of the transfection process. Lipoplex +/- charge ratios representing higher proportions of cat-ionic lipid/surfactant agents create an overall positive charge that ensures a favorable electro-static interaction with subsequent attachment of these nanoparticles to the negatively charged cell surface, an important early step in efficient
transfection. This is accompanied by a shift in the zeta potential of these particles to positive values. Anionic complexes, having a negative zeta potential, have been successfully applied in a few cases [178,179]. The use of anionic com-plexes has been observed to reduce cytotoxicity of the resulting nanoparticles, possibly due to increased biocompatibility of the anionic lipids. Condensation of DNA and formation of the lipoplex is usually mediated by Ca2+ ions, given the inherent i ncompatibility of the anionic l ipids and anionic DNA.
Membrane charge density has become recog-nized as a universal parameter [180] that governs the transfection efficiencies in nanoparticles formed from a binary mixture of lipids (i.e., one cationic and one neutral helper lipid) [181,182]. When extended to multicomponent lipid sys-tems, the transfection efficiency can be mod-eled as a function of membrane charge density fit to a Gaussian equation. It was demonstrated that the multicomponent systems exhibited transfection efficiencies of approximately an order of magnitude greater than binary lipid systems for the same membrane charge den-sity [180]. In this work, it was recognized that the interaction of nanoparticle lipids with endo-somal membrane lipids is a critical step related to efficient transfection; however, no link was found between membrane charge density and appearance of nonlamellar structures thought to facilitate DNA release (see later). Nevertheless, it is clear that membrane charge density, and therefore z and surface charge, play an impor-tant role, if not in determining transfection effi-ciency, then possibly in our understanding of the mechanism(s) involved.
�n Particle shapeThe shape of nanoparticles formed from nonvi-ral vectors and DNA depends on many factors; however, the most important are the structure(s) of the vector itself, the structure of the DNA and the ratio of vector to DNA (usually expressed as a +/- charge ratio, or as a nitrogen-to-phosphate ratio). Commonly observed structures include rods, globules, spheres and toroids, although fila-mentous or fibrous structures and netlike struc-tures are not uncommon. As with many aspects of such systems, no single shape has proven to correlate consistently with high transfection e fficiencies [176,183].
Nanoparticles formed from lipids or surfac-tants are generally spherical with sizes less than approximately 300–500 nm (see later), irrespec-tive of the choice of cationic lipid. The choice of
www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1117
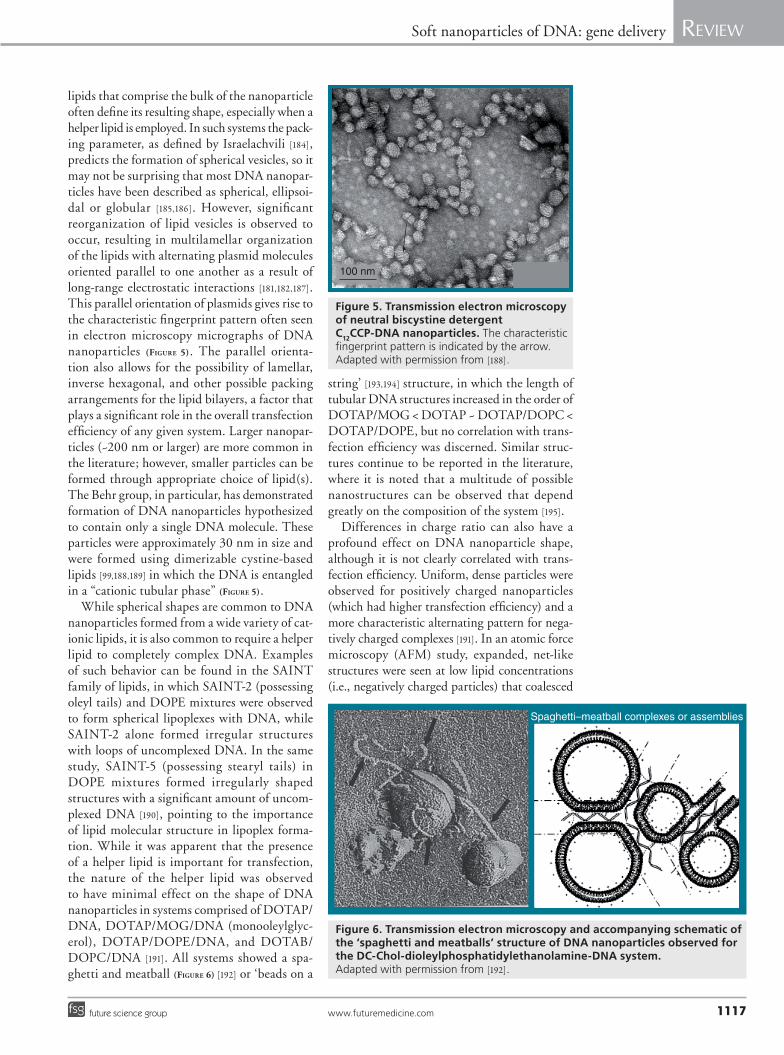
lipids that comprise the bulk of the nanoparticle often define its resulting shape, especially when a helper lipid is employed. In such systems the pack-ing parameter, as defined by Israelachvili [184], predicts the formation of spherical vesicles, so it may not be surprising that most DNA nanopar-ticles have been described as spherical, ellipsoi-dal or globular [185,186]. However, significant reorganization of lipid vesicles is observed to occur, resulting in multilamellar organization of the lipids with alternating plasmid molecules oriented parallel to one another as a result of long-range electrostatic interactions [181,182,187]. This parallel orientation of plasmids gives rise to the characteristic fingerprint pattern often seen in electron microscopy micrographs of DNA nanoparticles (Figure 5). The parallel orienta-tion also allows for the possibility of lamellar, inverse hexagonal, and other possible packing arrangements for the lipid bilayers, a factor that plays a significant role in the overall transfection efficiency of any given system. Larger nanopar-ticles (~200 nm or larger) are more common in the literature; however, smaller particles can be formed through appropriate choice of lipid(s). The Behr group, in particular, has demonstrated formation of DNA nanoparticles hypothesized to contain only a single DNA molecule. These particles were approximately 30 nm in size and were formed using dimerizable cystine-based lipids [99,188,189] in which the DNA is entangled in a “cationic tubular phase” (Figure 5).
While spherical shapes are common to DNA nanoparticles formed from a wide variety of cat-ionic lipids, it is also common to require a helper lipid to completely complex DNA. Examples of such behavior can be found in the SAINT family of lipids, in which SAINT-2 (possessing oleyl tails) and DOPE mixtures were observed to form spherical lipoplexes with DNA, while SAINT-2 alone formed irregular structures with loops of uncomplexed DNA. In the same study, SAINT-5 (possessing stearyl tails) in DOPE mixtures formed irregularly shaped structures with a significant amount of uncom-plexed DNA [190], pointing to the importance of lipid molecular structure in lipoplex forma-tion. While it was apparent that the presence of a helper lipid is important for transfection, the nature of the helper lipid was observed to have minimal effect on the shape of DNA nanoparticles in systems comprised of DOTAP/DNA, DOTAP/MOG/DNA (monooleylglyc-erol), DOTAP/DOPE/DNA, and DOTAB/DOPC/DNA [191]. All systems showed a spa-ghetti and meatball (Figure 6) [192] or ‘beads on a
string’ [193,194] structure, in which the length of tubular DNA structures increased in the order of DOTAP/MOG < DOTAP ~ DOTAP/DOPC < DOTAP/DOPE, but no correlation with trans-fection efficiency was discerned. Similar struc-tures continue to be reported in the literature, where it is noted that a multitude of possible nanostructures can be observed that depend greatly on the composition of the system [195].
Differences in charge ratio can also have a profound effect on DNA nanoparticle shape, although it is not clearly correlated with trans-fection efficiency. Uniform, dense particles were observed for positively charged nanoparticles (which had higher transfection efficiency) and a more characteristic alternating pattern for nega-tively charged complexes [191]. In an atomic force microscopy (AFM) study, expanded, net-like structures were seen at low lipid concentrations (i.e., negatively charged particles) that coalesced
Spaghetti–meatball complexes or assemblies
Figure 6. Transmission electron microscopy and accompanying schematic of the ‘spaghetti and meatballs’ structure of DNA nanoparticles observed for the DC-Chol-dioleylphosphatidylethanolamine-DNA system. Adapted with permission from [192].
Figure 5. Transmission electron microscopy of neutral biscystine detergent C12CCP-DNA nanoparticles. The characteristic fingerprint pattern is indicated by the arrow. Adapted with permission from [188].
100 nm

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1118
into globular structures at high lipid concentra-tions (positively charged particles); in this case, multivalent cationic lipids produced smaller-sized globules than did monovalent lipids [99,196,197]. For systems with charge ratios close to one (i.e., near stochiometry), aggregation and precipitation are common and large heterogeneously shaped struc-tures are usually formed. Such aggregated systems generally exhibit good in vitro transfection effi-ciency, in part because they undergo significant sedimentation that results in good contact with cell surfaces in culture. The same aggregates show poor in vivo transfection because of poor d iffusion as a result of their large size [91,198].
In any discussion of DNA nanoparticle shape, it is important to keep in mind potential limita-tions and/or artifacts of the imaging technique. electron microscopy and, more recently, AFM are commonly used to examine the shape of DNA nanoparticles. Freeze-fracture, cryo-transmission electron microscopy, and nega-tive staining methods are used to characterize DNA nanoparticulate systems, but all of these techniques require some form of sample prepa-ration or manipulation that may alter the shape of the system. Negative staining and AFM both require deposition of the sample onto a sub-strate, followed by drying, and can potentially alter the shape of particles either as a result of dehydration or interaction of the particles with the substrate itself.
Unlike the case of bilayer packing (i.e., lamel-lar, inverse hexagonal), it seems clear that the role nanoparticle shape plays in transfection effi-ciency is poorly understood. However, it is likely that particle shape is of less significance than other factors such as bilayer packing, particle size, surface charge, toxicity and complexation by serum protein.
�n Effect of internal structure of lipoplexes The structure of lipoplexes has been the focus of both theoretical and experimental studies that examine the relationship between the morpho-logical characteristics of lipoplexes and their func-tional activity [96]. Mixing cationic liposomes and DNA yields not only lipoplexes of various sizes, but also lipoplexes with different lipid-packing arrangements or morphologies, with lamellar, hexagonal and cubic phase structures having been reported [96,185,186]. These structural forms have been revealed in numerous studies using diffrac-tion or magnetic resonance methods; however, the lack of understanding of the relationship between transfection and the nature of complexes has also
necessitated determining other physical charac-teristics using various methods. Recent studies suggest that for lamellar lipoplexes to yield effi-cient transfection, their membrane charge density needed to be greater than a critical minimum of 1.04 × 10–2 e/Å2 [199]. In comparison, a fusogenic hexagonal complex yielded efficient transfection with a membrane charge below this critical mini-mum [180,199,200]. These results combined suggest that the ability of a given lipoplex to transfect DNA efficiently may depend very much on its ability to adopt various packing arrangements (i.e., to adopt a polymorphic structure).
A unique characteristic of transfection com-plexes that has attracted significant interest is the ability to adopt a combination of phases simul-taneously. Small-angle x-ray scattering studies have indicated that for systems involving DNA and a group of dimeric (dicationic) surfactants based on a tetramethylene spacer, the complexes mainly showed 2D hexagonal packing [201]. The complexes effectively revealed additional but unidentified structures after an increase in either the surfactant:DNA mole ratio or in tempera-ture, with stronger response observed for longer-tail surfactants. Introduction of dilauroylphos-phatidycholine to the binary surfactant/DNA system resulted in the formation of a condensed lamellar phase [202,203].
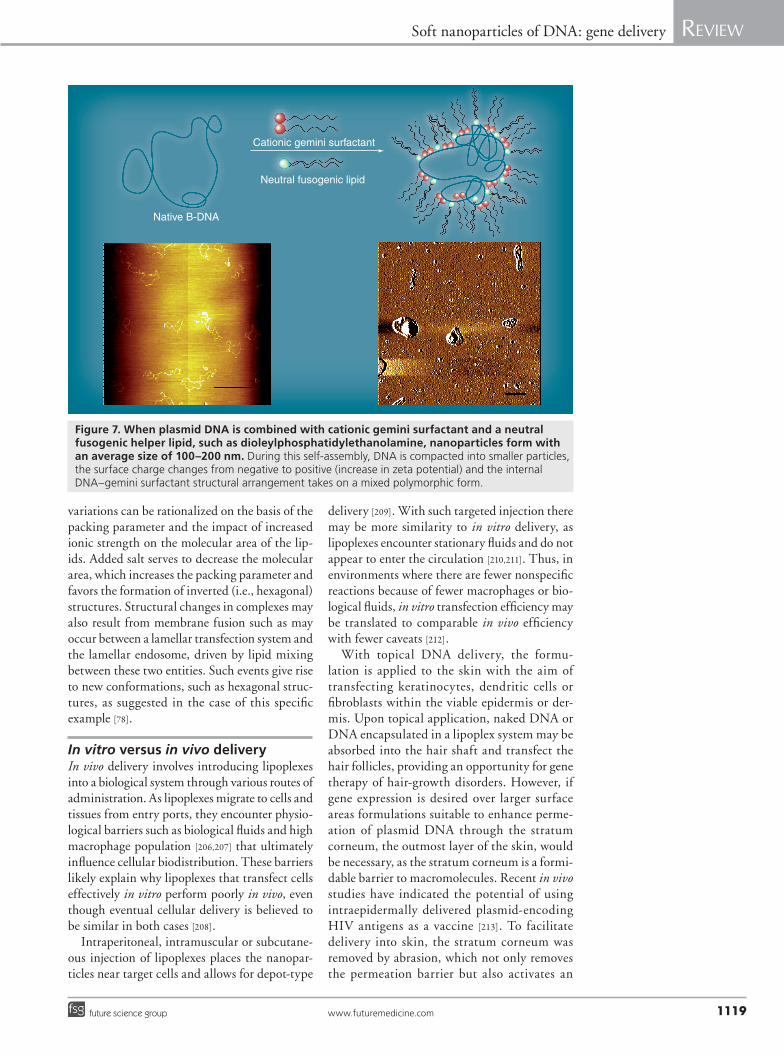
The importance of structural polymorphism has been demonstrated by using a group of dimeric surfactants with different lengths of hydrocarbon tail [204,205]. Transfection efficien-cies were found to improve with increasing alkyl tail length, but the strongest transfection was demonstrated by systems displaying a mixed polymorphic structure. Complexes that showed the commonly encountered lamellar structures in addition to Pn3m cubic structures had the greatest transfection efficiency. In a subsequent study [9], a similar strong enhancement of trans-fection was demonstrated using amine-substi-tuted spacer analogues of the gemini surfactants involved in earlier work. These surfactants adopt a wide range of structural morphologies, unlike the less efficient unsubstituted spacer analogues (Figure 7). Other important observa-tions have been made with regards to structural p olymorphism of transfection complexes.
The potential impact of environmental factors such as pH and ionic strength is exemplified by a transfection system consisting of a pyridinium derivative, DOPE and DNA. In aqueous solu-tion this system formed lamellar complexes, but when it was prepared in HEPES buffered saline a hexagonal conformation was observed. These
www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
1119
variations can be rationalized on the basis of the packing parameter and the impact of increased ionic strength on the molecular area of the lip-ids. Added salt serves to decrease the molecular area, which increases the packing parameter and favors the formation of inverted (i.e., hexagonal) structures. Structural changes in complexes may also result from membrane fusion such as may occur between a lamellar transfection system and the lamellar endosome, driven by lipid mixing between these two entities. Such events give rise to new conformations, such as hexagonal struc-tures, as suggested in the case of this specific example [78].
In vitro versus in vivo deliveryIn vivo delivery involves introducing lipoplexes into a biological system through various routes of administration. As lipoplexes migrate to cells and tissues from entry ports, they encounter physio-logical barriers such as biological fluids and high macrophage population [206,207] that ultimately influence cellular biodistribution. These barriers likely explain why lipoplexes that transfect cells effectively in vitro perform poorly in vivo, even though eventual cellular delivery is believed to be similar in both cases [208].
Intraperitoneal, intramuscular or subcutane-ous injection of lipoplexes places the nanopar-ticles near target cells and allows for depot-type
delivery [209]. With such targeted injection there may be more similarity to in vitro delivery, as lipoplexes encounter stationary fluids and do not appear to enter the circulation [210,211]. Thus, in environments where there are fewer nonspecific reactions because of fewer macrophages or bio-logical fluids, in vitro transfection efficiency may be translated to comparable in vivo efficiency with fewer caveats [212].
With topical DNA delivery, the formu-lation is applied to the skin with the aim of transfecting keratinocytes, dendritic cells or fibroblasts within the viable epidermis or der-mis. Upon topical application, naked DNA or DNA encapsulated in a lipoplex system may be absorbed into the hair shaft and transfect the hair follicles, providing an opportunity for gene therapy of hair-growth disorders. However, if gene expression is desired over larger surface areas formulations suitable to enhance perme-ation of plasmid DNA through the stratum corneum, the outmost layer of the skin, would be necessary, as the stratum corneum is a formi-dable barrier to macromolecules. Recent in vivo studies have indicated the potential of using intraepidermally delivered plasmid-encoding HIV antigens as a vaccine [213]. To facilitate delivery into skin, the stratum corneum was removed by abrasion, which not only removes the permeation barrier but also activates an
Figure 7. When plasmid DNA is combined with cationic gemini surfactant and a neutral fusogenic helper lipid, such as dioleylphosphatidylethanolamine, nanoparticles form with an average size of 100–200 nm. During this self-assembly, DNA is compacted into smaller particles, the surface charge changes from negative to positive (increase in zeta potential) and the internal DNA–gemini surfactant structural arrangement takes on a mixed polymorphic form.
Native B-DNA
Cationic gemini surfactant
Neutral fusogenic lipid

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1120
irritation response to summon dendritic cells to the application site for enhanced response to the antigen. In other applications, causing irritation or physical injury to the skin is coun-terproductive and an effective noninvasive topi-cal delivery system would be needed. Biphasic vesicles [214–216] and gemini nanoparticles (Figure 7) [9,172] were shown to effectively deliver plasmid DNA into intact human or mouse skin in vitro and in vivo.
Nonviral gene delivery systems move toward the clinicFour plasmid DNA products are currently licensed for veterinary applications for the treatment of viral infections (West Nile virus in horses [Ft Dodge Animal Health] and infec-tious haematopoietic necrosis virus in salmon [Novartis] and cancer [melanoma cancer vac-cine for dogs; Merial] and a growth hormone-releasing factor therapy for pigs [VGX Animal Health]) [217]. In addition, 72 Phase I, 20 Phase II and 2 Phase III human DNA vaccines are in clinical trials [218]. The vast majority of these trials are for cancer treatment (melanoma, lung, head and neck, ovarian, pancreatic, breast and prostate cancer), but they also include treat-ment for viral infections, cystic fibrosis and cor-onary artery disease. The latest updates indicate that 109 out of 1537 (7.1%) of all gene therapy clinical trials use lipid-based gene delivery/transfection agents [301].
Intense efforts are currently underway to develop efficient and safe nonviral vectors for transporting macromolecules such as oligonucle-otides and polynucleotides. Recently, a Phase I trial to assess the safety and tolerability of human interleukin-12 plasmid formulated with a syn-thetic lipopolymer, PEG-polyethyleneimine-cholesterol, was conducted for the treatment of chemotherapy-resistant recurrent ovarian cancer, and showed promising results. Indeed, the administration of these lipopolymer/DNA complexes could increase the mean survival time of the patients [219]. These data demonstrate that the rational and controlled design of multifunc-tional nonviral vectors for gene delivery is feasible for the treatment of various genetic diseases. In view of the conflicting structural requirements for the lipoplex-targeting properties and efficacy, future work should focus on the development and clinical application of multifunctional, yet easy to formulate, lipoplexes capable of accu-mulating at the target sites and elucidating the desired response. Although some new delivery technologies show promise in this regard, a
significant amount of work is still required to develop new products.
Conclusion & future perspectiveThe ultimate usefulness of nonviral gene delivery systems will depend on their efficiency, specificity and controllability; that is, the rate, extent and duration of transfection, the ability of the gene delivery systems to specifically interact with tar-get cells and the ability to negotiate in vivo and cellular barriers. Notwithstanding the consider-able effort, the establishment of strong correlation between transfection efficiency and many mea-sured characteristics of the complexes still needs significantly more data. However, it is true that there has been so much diversity amongst cell lines, vectors of different structures and co-lipids used so far in the reported studies, that the lack of clear correlations perhaps is not too surprising. In addition, the diversity of formulations from different laboratories has also contributed to this outcome. Nonviral vector development may still be considered in its infancy, and significantly more work is required to better understand the mechanism of the multiple steps in the transfec-tion pathway. Perhaps then, the results of ‘one-off’ experiments can lead to the development of more systematic studies that will yield firmer ideas about correlations between the chemical nature of the transfection agent, the structure of the delivery system, cell types and transfection efficiency. The bottom-up approach to construction described herein allows component-by-component design, testing, development and optimization of safer and more effective nonviral gene delivery systems. Additionally, better understanding of the effect of the resulting complexity of the internal structure of gene delivery complexes (not just external mor-phology) will also yield information about design factors contributing to the development of specific ‘intelligent’ functions capable of negotiating the barriers in gene delivery such as cellular, endocy-totic and nuclear membranes. Future improve-ments are expected from the application of novel nanomaterials and composites, for example car-bon nanotubes to construct delivery systems that do not necessarily rely on endocytotic uptake. Conjugation of ultraspecific targeting ligands, novel nonviral vectors/transgenes and combina-tion of chemical and physical on/off switching controls will provide the next generation of gene delivery methods.
AcknowledgementThe authors would like to thank Joe Petrik for editing the manuscript.

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
www.futuremedicine.com 1121
Bibliography1 Douglas KL: Toward development of
artificial viruses for gene therapy: a comparative evaluation of viral and non-viral transfection. Biotechnol. Prog. 24, 871–883 (2008).
2 Peterson CY, Shaterian A, Borboa AK et al.: The noninvasive, quantitative, in vivo assessment of adenoviral-mediated gene delivery in skin wound biomaterials. Biomaterials 30, 6788–6793 (2009).
3 Waehler R, Russell SJ, Curiel DT: Engineering targeted viral vectors for gene therapy. Nat. Rev. Genet. 8, 573–587 (2007).
4 Eto Y, Gao JQ, Sekiguchi F et al.: PEGylated adenovirus vectors containing RGD peptides on the tip of PEG show high
transduction efficiency and antibody evasion ability. J. Gene Med. 7, 604–612 (2005).
5 Wolff J, Lewis DL, Herweijer H, Hegge J, Hagstrom J: Non-viral approaches for gene transfer. Acta Myologica 24, 202–208 (2005).
6 Wettig DS, Verrall ER, Foldvari M: Gemini surfactants: a new family of building blocks for non-viral gene delivery systems. Curr. Gene Ther. 8, 9–23 (2008).
7 Oba M, Aoyagi K, Miyata K et al.: Polyplex micelles with cyclic RGD peptide ligands and disulfide cross-links directing to the enhanced transfection via controlled intracellular trafficking. Mol. Pharm. 5, 1080–1092 (2008).
8 Nemoto Y, Zhou YM, Tatsumi E, Nakayama Y: Photoinduced cross-linking of star vector for improvement of gene transfer efficiency. Bioconjug. Chem. 19, 2513–2519 (2008).
9 Wettig SD, Badea I, Donkuru M, Verrall RE, Foldvari M: Structural and transfection properties of amine-substituted gemini surfactant-based nanoparticles. J. Gene Med. 9, 649–658 (2007).
10 Elsabahy M, Wazen N, Bayó-Puxan N et al.: Delivery of nucleic acids through the controlled disassembly of multifunctional nanocomplexes. Adv. Funct. Mater. 19, 3862–3867 (2009).
11 Räty JK, Lesch HP, Wirth T, Ylä-Herttuala S: Improving safety of gene therapy. Curr. Drug Saf. 3, 46–53 (2008).
Executive summary
Currently recognized barriers to gene delivery � There are many obstacles in the development of successful gene therapy, such as, low cellular uptake, endosomal degradation and
inefficient nuclear localization of the genetic materials. � Steps required for successful nonviral gene delivery and expression include; binding of the DNA/lipid complex to the cell, internalization
into endosomes and subsequent release into the cytoplasm, translocation into the nucleus followed by transcription to mRNA and translation into the therapeutic protein that occurs in the cytoplasm.
Building blocks for lipid-based nanoparticles of DNA � The design of successful nonviral gene delivery systems follows a bottom-up approach. � Starting with basic building blocks such as cationic lipids, polymers, polypeptides or dendrimers, multifunctional complex systems can
be engineered. � Ideal systems bear molecular modules for cell binding, uptake, endosomal escape, nuclear localization, and for in vivo applications,
mobility within the body. � Cationic lipids are commonly used as nucleic acid delivery agents as they are efficient at condensing and encapsulating the DNA into
particles that can be internalized by the cells. They also contribute an overall net positive charge to the DNA complex, which appears to be important in the electrostatic interaction with the negatively charged cell surface.
� The fine-tuning of the cationic-to-anionic charge ratio (i.e., +/- ratio) has an important role in transfection efficiency. A high +/- ratio is more toxic to cells but results in higher transfection efficiency.
� Rational design principles and quantitative structure–activity relationship modeling elucidate important structural characteristics of cationic lipids that facilitate more effective transfection.
� The transfection efficiency of cationic lipids can be augmented by incorporating neutral lipids (e.g., DOPE) into the delivery system. � Steric stabilization is usually required to avoid the rapid clearance of the DNA–lipid complexes by the mononuclear phagocyte system
and is usually achieved by grafting hydrophilic polymers (e.g., polyethylene glycol) to the surface of the liposomes.
Nanoparticle properties important in transfection � Many of the physical and physicochemical properties of lipoplexes, such as particle size, surface charge density, morphology or shape,
internal (polymorphic) structure, and compactness affect gene delivery both in vitro and in vivo.
Conclusions � The ultimate usefulness of nonviral gene delivery systems depends on their efficiency, specificity and controllability. � Intense efforts are currently given to develop efficient and safe nonviral vectors for transporting macromolecules such as DNA, RNA
and proteins. � The establishment of a strong correlation between transfection efficiency and various measured characteristics of the complexes requires
significantly more research. � The bottom-up approach to construction described herein allows for a component-by-component design, testing and optimization of
safer nonviral gene delivery systems to meet these goals.
Financial & competing interests disclosureThe research is supported by grants from the Canadian Institutes of Health Research and the Natural Sciences and Engineering Research Council of Canada. The authors have no other relevant affiliations or financial involvement with
any organization or entity with a financial interest in or financial conflict with the subject matter or materials d iscussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1122
12 Harrington KJ, Nutting CM, Pandha HS: Gene therapy for head and neck cancer. Cancer Metastasis Rev. 24, 147–164 (2005).
13 Abadie J, Blouin V, Guigand L, Wyers M, Cherel Y: Recombinant adeno-associated virus type 2 mediates highly efficient gene transfer in regenerating rat skeletal muscle. Gene Ther. 9, 1037–1043 (2002).
14 Cartier N, Hacein-Bey-Abina S, Bartholomae CC et al.: Hematopoietic stem cell gene therapy with a lentiviral vector in X-linked adrenoleukodystrophy. Science 326, 818–823 (2009).
15 Felgner PL, Gadek TR, Holm M et al.: Lipofection: a highly efficient, lipid-mediated DNA-transfection procedure. Proc. Natl Acad. Sci. USA 84, 7413–7417 (1987).
16 Verma IM, Somia N: Gene therapy-promises, problems and prospects. Nature 389, 239–242 (1997).
17 Lehrman S: Virus treatment questioned after gene therapy death. Nature 401, 517–518 (1999).
18 Marshall E: Gene therapy: What to do when clear success comes with an unclear risk? Science 298, 510–511 (2002).
19 Li SD, Huang L: Non-viral is superior to viral gene delivery. J. Control Release 123, 181–183 (2007).
20 Li SD, Huang L: Gene therapy progress and prospects: non-viral gene therapy by systemic delivery. Gene Therapy 13, 1313–1319 (2006).
21 Domashenko A, Gupta S, Cotsarelis G: Efficient delivery of transgenes to human hair follicle progenitor cells using topical lipoplex. Nat. Biotechnol. 18, 420–423 (2000).
22 Wolff JA, Rozema DB: Breaking the bonds: non-viral vectors become chemically dynamic. Mol. Ther. 16, 8–15 (2008).
23 Park TG, Jeong JH, Kim SW: Current status of polymeric gene delivery systems. Adv. Drug Deliv. Rev. 58, 467–486 (2006).
24 Meyer M, Wagner E: pH-responsive shielding of non-viral gene vectors. Expert Opin. Drug Deliv. 3, 563–571 (2006).
25 Wettig SD, Wang C, Verrall RE, Foldvari M: Thermodynamic and aggregation properties of aza- and imino-substituted gemini surfactants designed for gene delivery. Phys. Chem. Chem. Phys. 9, 871–877 (2007).
26 Saunders M, Taylor KM, Craig DQ, Palin K, Robson H: High sensitivity differential scanning calorimetry study of DNA-cationic liposome complexes. Pharm. Res. 24, 1954–1961 (2007).
27 Zhang HW, Zhang L, Sun X, Diao S, Zhang ZR: Assembly of plasmid DNA into liposomes after condensation by cationic lipid in anionic detergent solution. Biotechnol. Lett. 27, 1701–1705 (2005).
28 Kim WJ, Yockman JW, Lee M, Jeong JH, Kim YH, Kim SW: Soluble Flt-1 gene delivery using PEI-g-PEG-RGD conjugate for anti-angiogenesis. J. Control Release 106, 224–234 (2005).
29 Elfinger M, Maucksch C, Rudolph C: Characterization of lactoferrin as a targeting ligand for nonviral gene delivery to airway epithelial cells. Biomaterials 28, 3448–3455 (2007).
30 Dinh AT, Pangarkar C, Theofanous T, Mitragotri S: Understanding intracellular transport processes pertinent to synthetic gene delivery via stochastic simulations and sensitivity analyses. Biophys. J. 92, 831–846 (2007).
31 Nishikawa M, Huang L: Nonviral vectors in the new millennium: delivery barriers in gene transfer. Hum. Gene Ther. 12, 861–870 (2001).
32 Elouahabi A, Ruysschaert JM: Formation and intracellular trafficking of lipoplexes and polyplexes. Mol. Ther. 11, 336–347 (2005).
33 Sigismund S, Woelk T, Puri C et al.: Clathrin-independent endocytosis of ubiquitinated cargos. Proc. Natl Acad. Sci. USA 102, 2760–2765 (2005).
34 Mayor S, Pagano RE: Pathways of clathrin-independent endocytosis. Nat. Rev. Mol. Cell. Biol. 8, 603–612 (2007).
35 Zabner J, Fasbender AJ, Moninger T, Poellinger KA, Welsh MJ: Cellular and molecular barriers to gene transfer by a cationic lipid. J. Biol. Chem. 270, 18997–19007 (1995).
36 Ruponen M, Arkko S, Urtti A, Reinisalo M, Ranta VP: Intracellular DNA release and elimination correlate poorly with transgene expression after non-viral transfection. J. Control Release 136, 226–231 (2009).
37 Doyle SR, Chan CK: Differential intracellular distribution of DNA complexed with polyethylenimine (PEI) and PEI-polyarginine PTD influences exogenous gene expression within live COS-7 cells. Genet. Vaccines Ther. 5, 11 (2007).
38 Friend DS, Papahadjopoulos D, Debs RJ: Endocytosis and intracellular processing accompanying transfection mediated by cationic liposomes. Biochim. Biophys. Acta 1278, 41–50 (1996).
39 Wrobel I, Collins D: Fusion of cationic liposomes with mammalian cells occurs after endocytosis. Biochim. Biophys. Acta 1235, 296–304 (1995).
40 Jeong JH, Lee M, Kim WJ et al.: Anti-GAD antibody targeted non-viral gene delivery to islet beta cells. J. Control Release 107, 562–570 (2005).
41 Huang R-Q, Qu Y-H, Ke W-L, Zhu J-H, Pei Y-Y, Jiang C: Efficient gene delivery targeted to the brain using a transferrin-conjugated polyethyleneglycol-modified polyamidoamine dendrimer. FASEB J. 21, 1117–1125 (2007).
42 Torchilin VP: Recent approaches to intracellular delivery of drugs and DNA and organelle targeting. Annu. Rev. Biomed. Eng. 8, 343–375 (2006).
43 Goryacheva YA, Vekshina OM, Yashin VA, Kim YA: Fusion and endocytosis of anionic liposomes with Ehrlich ascitic carcinoma cells. Bull. Exp. Biol. Med. 140, 733–735 (2005).
44 Savva M, Chen P, Aljaberi A, Selvi B, Spelios M: In vitro lipofection with novel asymmetric series of 1,2-dialkoylamidopropane-based cytofectins containing single symmetric bis-(2-dimethylaminoethane) polar headgroups. Bioconjug. Chem. 16, 1411–1422 (2005).
45 Smith JG, Walzemb RL, Germana BJ: Liposomes as agents of DNA transfer. Biochim. Biophys. Acta 1154, 327–340 (1993).
46 Moore NM, Sheppard CL, Sakiyama-Elbert SE: Characterization of a multifunctional PEG-based gene delivery system containing nuclear localization signals and endosomal escape peptides. Acta Biomater 5, 854–864 (2009).
47 Bhattacharya S, Bajaj A: Advances in gene delivery through molecular design of cationic lipids. Chem. Commun. 31, 4632–4656 (2009).
48 Martin B, Sainlos M, Aissaoui A et al.: The design of cationic lipids for gene delivery. Curr. Pharm. Des. 11, 375–394 (2005).
49 Midoux P, Pichon C, Yaouanc JJ, Jaffres PA: Chemical vectors for gene delivery: a current review on polymers, peptides and lipids containing histidine or imidazole as nucleic acids carriers. Br. J. Pharmacol. 157, 166–178 (2009).
50 Horobin RW, Weissig V: A QSAR-modeling perspective on cationic transfection lipids. 1. Predicting efficiency and understanding mechanisms. J. Gene Med. 7, 1023–1034 (2005).
51 Barry ME, Pinto-Gonzalez D, Orson FM, McKenzie GJ, Petry GR, Barry MA: Role of endogenous endonucleases and tissue site in transfection and CpG-mediated immune activation after naked DNA injection. Hum. Gene Ther. 10, 2461–2480 (1999).
52 Singh M, Ariatti M: A cationic cytofectin with long spacer mediates favourable transfection in transformed human epithelial cells. Int. J. Pharm. 309, 189–198 (2006).

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
www.futuremedicine.com 1123
53 Kawakami S, Harada A, Sakanaka K et al.: In vivo gene transfection via intravitreal injection of cationic liposome/plasmid DNA complexes in rabbits. Int. J. Pharm. 278, 255–262 (2004).
54 Simoes S, Slepushkin V, Pires P, Gaspar R, de Lima MP, Duzgunes N: Mechanisms of gene transfer mediated by lipoplexes associated with targeting ligands or pH-sensitive peptides. Gene Ther. 6, 1798–1807 (1999).
55 Zuidam NJ, Hirsch-Lerner D, Margulies S, Barenholz Y: Lamellarity of cationic liposomes and mode of preparation of lipoplexes affect transfection efficiency. Biochim. Biophys. Acta 1419, 207–220 (1999).
56 Zuhorn IS, Bakowsky U, Polushkin E et al.: Nonbilayer phase of lipoplex-membrane mixture determines endosomal escape of genetic cargo and transfection efficiency. Mol. Ther. 11, 801–810 (2005).
57 Wetzer B, Byk G, Frederic M et al.: Reducible cationic lipids for gene transfer. Biochem. J. 356, 747–756 (2001).
58 Byk G, Wetzer B, Frederic M et al.: Reduction-sensitive lipopolyamines as a novel nonviral gene delivery system for modulated release of DNA with improved transgene expression. J. Med. Chem. 43, 4377–4387 (2000).
59 Ewert KK, Evans HM, Zidovska A, Bouxsein NF, Ahmad A, Safinya CR: A columnar phase of dendritic lipid-based cationic liposome-DNA complexes for gene delivery: hexagonally ordered cylindrical micelles embedded in a DNA honeycomb lattice. J. Am. Chem. Soc. 128, 3998–4006 (2006).
60 Ewert K, Ahmad A, Evans HM, Schmidt HW, Safinya CR: Efficient synthesis and cell-transfection properties of a new multivalent cationic lipid for nonviral gene delivery. J. Med. Chem. 45, 5023–5029 (2002).
61 Wasungu L, Stuart MC, Scarzello M, Engberts JB, Hoekstra D: Lipoplexes formed from sugar-based gemini surfactants undergo a lamellar-to-micellar phase transition at acidic pH. Evidence for a non-inverted membrane-destabilizing hexagonal phase of lipoplexes. Biochim. Biophys. Acta 1758, 1677–1684 (2006).
62 Bombelli C, Giansanti L, Luciani P, Mancini G: Gemini surfactant based carriers in gene and drug delivery. Curr. Med. Chem. 16, 171–183 (2009).
63 Wasungu L, Scarzello M, van Dam G et al.: Transfection mediated by pH-sensitive sugar-based gemini surfactants; potential for in vivo gene therapy applications. J. Mol. Med. 84, 774–784 (2006).
64 Yoshioka T, Yoshida S, Kurosaki T et al.: Cationic liposomes-mediated plasmid DNA delivery in murine hepatitis induced by carbon tetrachloride. J. Liposome Res. 19, 1–7 (2009).
65 Wu Q, Mahendran R, Esuvaranathan K: Nonviral cytokine gene therapy on an orthotopic bladder cancer model. Clin. Cancer Res. 9, 4522–4528 (2003).
66 Hu Y, Li K, Wang L, Yin S, Zhang Z, Zhang Y: Pegylated immuno-lipopolyplexes: a novel non-viral gene delivery system for liver cancer therapy. J. Control Release 144(1), 75–81 (2010).
67 Kurosaki T, Kitahara T, Teshima M et al.: Exploitation of de novo helper-lipids for effective gene delivery. J Pharm Pharm Sci 11, 56–67 (2008).
68 Prata CA, Li Y, Luo D, McIntosh TJ, Barthelemy P, Grinstaff MW: A new helper phospholipid for gene delivery. Chem. Commun. (Camb.) 1566–1568 (2008).
69 Zhao YZ, Luo YK, Liang HD et al.: Comparing transfection efficiency and safety for antisense oligodeoxyribonucleotide between phospholipids-based microbubbles and liposomes. J. Drug Target 14, 687–693 (2006).
70 Tranchant I, Thompson B, Nicolazzi C, Mignet N, Scherman D: Physicochemical optimisation of plasmid delivery by cationic lipids. J. Gene Med. 6, S24–S35 (2004).
71 Choi WJ, Kim JK, Choi SH, Park JS, Ahn WS, Kim CK: Low toxicity of cationic lipid-based emulsion for gene transfer. Biomaterials 25, 5893–5903 (2004).
72 Gruneich JA, Price A, Zhu J, Diamond SL: Cationic corticosteroid for nonviral gene delivery. Gene Ther. 11, 668–674 (2004).
73 Kim A, Lee EH, Choi SH, Kim CK: In vitro and in vivo transfection efficiency of a novel ultradeformable cationic liposome. Biomaterials 25, 305–313 (2004).
74 Gilot D, Miramon ML, Benvegnu T et al.: Cationic lipids derived from glycine betaine promote efficient and non-toxic gene transfection in cultured hepatocytes. J. Gene Med. 4, 415–427 (2002).
75 Koynova R, Wang L, MacDonald RC: An intracellular lamellar-nonlamellar phase transition rationalizes the superior performance of some cationic lipid transfection agents. Proc. Natl Acad. Sci. USA 103, 14373–14378 (2006).
76 Dobbs W, Heinrich B, Bourgogne C et al.: Mesomorphic imidazolium salts: new vectors for efficient siRNA transfection. J. Am. Chem. Soc. 131, 13338–13346 (2009).
77 Cui L, Chen D, Zhu L: Conformation transformation determined by different self-assembled phases in a DNA complex
with cationic polyhedral oligomeric silsesquioxane lipid. ACS Nano 2, 921–927 (2008).
78 Hafez IM, Maurer N, Cullis PR: On the mechanism whereby cationic lipids promote intracellular delivery of polynucleic acids. Gene Ther. 8, 1188–1196 (2001).
79 Hara T, Liu F, Liu D, Huang L: Emulsion formulations as a vector for gene delivery in vitro and in vivo. Adv. Drug Deliv. Rev. 24, 265–271 (1997).
80 Ciani L, Ristori S, Salvati A, Calamai L, Martini G: DOTAP/DOPE and DC-Chol/DOPE lipoplexes for gene delivery: zeta potential measurements and electron spin resonance spectra. Biochim. Biophys. Acta 1664, 70–79 (2004).
81 Zhi D, Zhang S, Wang B, Zhao Y, Yang B, Yu S: Transfection efficiency of cationic lipids with different hydrophobic domains in gene delivery. Bioconjug. Chem. 21(4), 563–577 (2010).
82 Kikuchi A, Aoki Y, Sugaya S et al.: Development of novel cationic liposomes for efficient gene transfer into peritoneal disseminated tumor. Hum. Gene Ther. 10, 947–955 (1999).
83 Kerner M, Meyuhas O, Hirsch-Lerner D, Rosen LJ, Min Z, Barenholz Y: Interplay in lipoplexes between type of pDNA promoter and lipid composition determines transfection efficiency of human growth hormone in NIH3T3 cells in culture. Biochim. Biophys. Acta 1532, 128–136 (2001).
84 Zuidam NJ, Barenholz Y: Characterization of DNA-lipid complexes commonly used for gene delivery. Int. J. Pharm. 183, 43–46 (1999).
85 Kim TW, Chung H, Kwon IC, Sung HC, Jeong SY: Optimization of lipid composition in cationic emulsion as in vitro and in vivo transfection agents. Pharm. Res. 18, 54–60 (2001).
86 Sheikh M, Feig J, Gee B, Li S, Savva M: In vitro lipofection with novel series of symmetric 1,3-dialkoylamidopropane-based cationic surfactants containing single primary and tertiary amine polar head groups. Chem. Phys. Lipids 124, 49–61 (2003).
87 Aljaberi A, Chen P, Savva M: Synthesis, in vitro transfection activity and physicochemical characterization of novel N,N ’-diacyl-1,2-diaminopropyl-3-carbamoyl-(dimethylaminoethane) amphiphilic derivatives. Chem Phys Lipids 133, 135–149 (2005).
88 Kearns MD, Donkor AM, Savva M: Structure-transfection activity studies of novel cationic cholesterol-based amphiphiles. Mol. Pharm. 5, 128–139 (2008).

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1124
89 Maitani Y, Igarashi S, Sato M, Hattori Y: Cationic liposome (DC-Chol/DOPE=1:2) and a modified ethanol injection method to prepare liposomes, increased gene expression. Int. J. Pharm. 342, 33–39 (2007).
90 Kaneshiro TL, Wang X, Lu ZR: Synthesis, characterization, and gene delivery of poly-l-lysine octa(3-aminopropyl)silsesquioxane dendrimers: nanoglobular drug carriers with precisely defined molecular architectures. Mol. Pharm. 4, 759–768 (2007).
91 Tarwadi, Jazayeri JA, Prankerd RJ, Pouton CW: Preparation and in vitro evaluation of novel lipopeptide transfection agents for efficient gene delivery. Bioconjug. Chem. 19, 940–950 (2008).
92 Perrie Y, Gregoriadis G: Liposome-entrapped plasmid DNA: characterisation studies. Biochim. Biophys. Acta 1475, 125–132 (2000).
93 Heinze M, Brezesinski G, Dobner B, Langner A: Novel cationic lipids based on malonic acid amides backbone: transfection efficacy and cell toxicity properties. Bioconjug. Chem. 21, 696–708 (2010).
94 Yu RZ, Geary RS, Leeds JM et al.: Pharmacokinetics and tissue disposition in monkeys of an antisense oligonucleotide inhibitor of Ha-ras encapsulated in stealth liposomes. Pharm. Res. 16, 1309–1315 (1999).
95 Ruponen M, Honkakoski P, Tammi M, Urtti A: Cell-surface glycosaminoglycans inhibit cation-mediated gene transfer. J. Gene Med. 6, 405–414 (2004).
96 Martin-Herranz A, Ahmad A, Evans HM, Ewert K, Schulze U, Safinya CR: Surface functionalized cationic lipid-DNA complexes for gene delivery: PEGylated lamellar complexes exhibit distinct DNA-DNA interaction regimes. Biophys. J. 86, 1160–1168 (2004).
97 Li W, Huang Z, MacKay JA, Grube S, Szoka FC: Low-pH-sensitive poly(ethylene glycol) (PEG)-stabilized plasmid nanolipoparticles: effects of PEG chain length, lipid composition and assembly conditions on gene delivery. J. Gene Med. 7, 67–79 (2005).
98 Munkonge FM, Amin V, Hyde SC et al.: Identification and functional characterization of cytoplasmic determinants of plasmid DNA nuclear import. J. Biol. Chem. 284, 26978–26987 (2009).
99 Ma B, Zhang S, Jiang H, Zhao B, Lv H: Lipoplex morphologies and their influences on transfection efficiency in gene delivery. J. Control Release 123, 184–194 (2007).
100 Sakuragi M, Kusuki S, Hamada E et al.: Supramolecular structures of benzyl amine derivate/DNA complexes explored with synchrotron small angle X-ray scattering at SPring-8. J. Phys. Conf. Ser. 184, 1–6 (2009).
101 Koynova R, Wang L, Tarahovsky Y, MacDonald RC: Lipid phase control of DNA delivery. Bioconjug. Chem. 16, 1335–1339 (2005).
102 Xu Z, Chen L, Zhang Z, Gu W, Li Y: “Intelligent” nanoassembly for gene delivery: In vitro transfection and the possible mechanism. Int. J. Pharm. 383, 271–276 (2010).
103 Mevel M, Neveu C, Goncalves C et al.: Novel neutral imidazole-lipophosphoramides for transfection assays. Chem. Commun. (Camb.) 3124–3126 (2008).
104 Solodin I, Brown CS, Bruno MS et al.: A novel series of amphiphilic imidazolinium compounds for in vitro and in vivo gene delivery. Biochemistry 34, 13537–13544 (1995).
105 Chowdhury EH, Akaike T: High performance DNA nano-carriers of carbonate apatite: multiple factors in regulation of particle synthesis and transfection efficiency. Int. J. Nanomedicine 2, 101–106 (2007).
106 Moore NM, Sheppard CL, Barbour TR, Sakiyama-Elbert SE: The effect of endosomal escape peptides on in vitro gene delivery of polyethylene glycol-based vehicles. J. Gene Med. 10, 1134–1149 (2008).
107 Hoekstra D, Rejman J, Wasungu L, Shi F, Zuhorn I: Gene delivery by cationic lipids: in and out of an endosome. Biochem. Soc. Trans. 35, 68–71 (2007).
108 Tarahovsky YS, Koynova R, MacDonald RC: DNA release from lipoplexes by anionic lipids: correlation with lipid mesomorphism, interfacial curvature, and membrane fusion. Biophys. J. 87, 1054–1064 (2004).
109 Tarahovsky Y, Koynova R, MacDonald RC: DNA release from lipoplexes by anionic lipids: correlation with lipid mesomorphism, interfacial curvature, and membrane fusion. Biophys. J. 87, 1054–1064 (2004).
110 Oishi M, Kataoka K, Nagasaki Y: pH-responsive three-layered PEGylated polyplex micelle based on a lactosylated ABC triblock copolymer as a targetable and endosome-disruptive nonviral gene vector. Bioconjug. Chem. 17, 677–688 (2006).
111 Walker SA, Kennedy MT, Zasadzinski JA: Encapsulation of bilayer vesicles by self-assembly. Nature (London) 387, 61–64 (1997).
112 Kim TH, Jiang HL, Nah JW, Cho MH, Akaike T, Cho CS: Receptor-mediated gene delivery using chemically modified chitosan. Biomed. Mater. 2, S95–S100 (2007).
113 Chen CP, Kim JS, Liu D et al.: Synthetic PEGylated glycoproteins and their utility in gene delivery. Bioconjug. Chem. 18, 371–378 (2007).
114 Hofland HEJ, Masson C, Iginla S et al.: Folate-targeted gene transfer in vivo. Molecular Therapy 5, 739–744 (2002).
115 Funhoff AM, van Nostrum CF, Lok MC, Fretz MM, Crommelin DJ, Hennink WE: Poly(3-guanidinopropyl methacrylate): a novel cationic polymer for gene delivery. Bioconjug. Chem. 15, 1212–1220 (2004).
116 Shangguan T, Cabral-Lilly D, Purandare U et al.: A novel N-acyl phosphatidylethanolamine-containing delivery vehicle for spermine-condensed plasmid DNA. Gene Ther. 7, 769–783 (2000).
117 Affleck DG, Yu L, Bull DA, Bailey SH, Kim SW: Augmentation of myocardial transfection using TerplexDNA: a novel gene delivery system. Gene Ther. 8, 349–353 (2001).
118 Yu L, Suh H, Koh JJ, Kim SW: Systemic administration of TerplexDNA system: pharmacokinetics and gene expression. Pharm. Res. 18, 1277–1283 (2001).
119 Nakamura K, Kuramoto Y, Mukai H, Kawakami S, Higuchi Y, Hashida M: Enhanced gene transfection in macrophages by histidine-conjugated mannosylated cationic liposomes. Biol. Pharm. Bull. 32, 1628–1631 (2009).
120 You J, Kamihira M, Iijima S: Enhancement of transfection efficiency using ligand-modified lipid vesicles. J. Ferment. Bioeng. 85, 525–528 (1998).
121 Anwer K, Kao G, Rolland A, Driessen WH, Sullivan SM: Peptide-mediated gene transfer of cationic lipid/plasmid DNA complexes to endothelial cells. J. Drug Target 12, 215–221 (2004).
122 Sakae M, Ito T, Yoshihara C et al.: Highly efficient in vivo gene transfection by plasmid/PEI complexes coated by anionic PEG derivatives bearing carboxyl groups and RGD peptide. Biomed. Pharmacother. 62, 448–453 (2008).
123 Reddy BS, Banerjee R: 17b-estradiol-associated stealth-liposomal delivery of anticancer gene to breast cancer cells. Angew. Chem. Int. Ed. Engl. 44, 6723–6727 (2005).
124 Mukherjee A, Prasad TK, Rao NM, Banerjee R: Haloperidol-associated stealth liposomes: a potent carrier for delivering genes to human breast cancer cells. J. Biol. Chem. 280, 15619–15627 (2005).
125 Wang M, Lowik DW, Miller AD, Thanou M: Targeting the urokinase plasminogen activator receptor with synthetic self-assembly nanoparticles. Bioconjug. Chem. 20, 32–40 (2009).
126 Opanasopit P, Rojanarata T, Apirakaramwong A, Ngawhirunpat T, Ruktanonchai U: Nuclear localization signal peptides enhance transfection efficiency of chitosan/DNA complexes. Int. J. Pharm. 382, 291–295 (2009).

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
www.futuremedicine.com 1125
127 Nagasaki T, Kawazu T, Tachibana T, Tamagaki S, Shinkai S: Enhanced nuclear import and transfection efficiency of plasmid DNA using streptavidin-fused importin-beta. J. Control Release 103, 199–207 (2005).
128 Shen Y, Peng H, Pan S et al.: Interaction of DNA/nuclear protein/polycation and the terplexes for gene delivery. Nanotechnology 21, 045102 (2010).
129 Amiji MM: Polymeric gene delivery: principles and applications (1st Edition). Boca Raton FL (Ed.). Taylor & Francis, CRC Press (2007).
130 Miller AM, Dean DA: Tissue-specific and transcription factor-mediated nuclear entry of DNA. Adv. Drug Deliv. Rev. 61, 603–613 (2009).
131 Kalderon D, Richardson WD, Markham AF, Smith AE: Sequence requirements for nuclear location of simian virus 40 large-T antigen. Nature 311, 33–38 (1984).
132 Jans DA, Chan CK, Huebner S: Signals mediating nuclear targeting and their regulation: application in drug delivery. Med. Res. Rev. 18, 189–223 (1998).
133 Nagasaki T, Myohoji T, Tachibana T, Futaki S, Tamagaki S: Can nuclear localization signals enhance nuclear localization of plasmid DNA? Bioconjug. Chem. 14, 282–286 (2003).
134 Kurihara D, Akita H, Kudo A, Masuda T, Futaki S, Harashima H: Effect of polyethyleneglycol spacer on the binding properties of nuclear localization signal-modified liposomes to isolated nucleus. Biol. Pharm. Bull. 32, 1303–1306 (2009).
135 Michael WM, Choi M, Dreyfuss G: A nuclear export signal in hnRNP A1: a signal-mediated, temperature-dependent nuclear protein export pathway. Cell. Mol. Biol. Lett. 83, 415–422 (1995).
136 Ling Y, Smith AJ, Morgan GT: A sequence motif conserved in diverse nuclear proteins identifies a protein interaction domain utilised for nuclear targeting by human TFIIS. Nucleic Acids Res. 34, 2219–2229 (2006).
137 Wagstaff KM, Jans DA: Nucleocytoplasmic transport of DNA: enhancing non-viral gene transfer. Biochem. J. 406, 185–202 (2007).
138 MA K, Min-Xin H, QI Y et al.: PAMAM-triamcinolone acetonide conjugate as a nucleus-targeting gene carrier for enhanced transfer activity. Biomaterials 30, 6109–6118 (2009).
139 De Laporte L, Cruz Rea J, Shea LD: Design of modular non-viral gene therapy vectors. Biomaterials 27, 947–954 (2006).
140 Vaysse L, Harbottle R, Bigger B, Bergau A, Tolmachov O, Coutelle C: Development of a self-assembling nuclear targeting vector
system based on the tetracycline repressor protein. J. Biol. Chem. 279, 5555–5564 (2004).
141 Byrnes CK, Nass PH, Duncan MD, Harmon JW: A nuclear targeting peptide, m9, improves transfection efficiency in fibroblasts. J. Surg. Res. 108, 85–90 (2002).
142 Goncalves C, Ardourel MY, Decoville M et al.: An optimized extended DNA kappa B site that enhances plasmid DNA nuclear import and gene expression. J. Gene Med. 11, 401–411 (2009).
143 Shea LD, Houchin TL: Modular design of non-viral vectors with bioactive components. Trends Biotechnol. 22, 429–431 (2004).
144 Cruz MTd, Simões S, Lima MCd: Improving lipoplex-mediated gene transfer into C6 glioma cells and primary neurons. Exp. Neurol. 187, 65–75 (2004).
145 Hatakeyama H, Ito E, Akita H et al.: A pH-sensitive fusogenic peptide facilitates endosomal escape and greatly enhances the gene silencing of siRNA-containing nanoparticles in vitro and in vivo. J. Control Release 139, 127–132 (2009).
146 Sasaki K, Kogure K, Chaki S et al.: An artificial virus-like nano carrier system: enhanced endosomal escape of nanoparticles via synergistic action of pH-sensitive fusogenic peptide derivatives. Anal. Bioanal. Chem. 391, 2717–2727 (2008).
147 Zhang X, Collins L, Fabre JW: A powerful cooperative interaction between a fusogenic peptide and lipofectamine for the enhancement of receptor-targeted, non-viral gene delivery via integrin receptors. J. Gene Med. 3, 560–568 (2001).
148 Badea I, Verrall R, Baca-Estrada M et al.: In vivo cutaneous interferon-g gene delivery using novel dicationic (gemini) surfactant-plasmid complexes. J. Gene Med. 7, 1200–1214 (2005).
149 Mohammadabadi MR, El-Tamimy M, Gianello R, Mozafari MR: Supramolecular assemblies of zwitterionic nanoliposome-polynucleotide complexes as gene transfer vectors: Nanolipoplex formulation and in vitro characterisation. J. Liposome Res. 19(2), 105–115 (2009).
150 Narang AS, Thoma L, Miller DD, Mahato RI: Cationic lipids with increased DNA binding affinity for nonviral gene transfer in dividing and nondividing cells. Bioconjug. Chem. 16, 156–168 (2005).
151 Koumbi D, Clement JC, Sideratou Z, Yaouanc JJ, Loukopoulos D, Kollia P: Factors mediating lipofection potency of a series of cationic phosphonolipids in human cell lines. Biochim. Biophys. Acta 1760, 1151–1159 (2006).
152 Mortazavi SM, Mohammadabadi MR, Khosravi-Darani K, Mozafari MR: Preparation of liposomal gene therapy vectors by a scalable method without using volatile solvents or detergents. J. Biotechnol. 129, 604–613 (2007).
153 Staggs DR, Burton DW, Deftos LJ: Importance of liposome complexing volume in transfection optimization. Biotechniques 21, 792, 784, 796, 798 (1996).
154 Sternberg B, Hong K, Zheng W, Papahadjopoulos D: Ultrastructural characterization of cationic liposome-DNA complexes showing enhanced stability in serum and high transfection activity in vivo. Biochim. Biophys. Acta 1375, 23–35 (1998).
155 Gaucheron J, Santaella C, Vierling P: In vitro gene transfer with a novel galactosylated spermine bolaamphiphile. Bioconjug. Chem. 12, 569–575 (2001).
156 Caracciolo G, Caminiti R, Digman MA, Gratton E, Sanchez S: Efficient escape from endosomes determines the superior efficiency of multicomponent lipoplexes. J. Phys. Chem. B 113, 4995–4997 (2009).
157 Hufnagel H, Hakim P, Lima A, Hollfelder F: Fluid phase endocytosis contributes to transfection of DNA by PEI-25. Mol. Ther. 17, 1411–1417 (2009).
158 Rejman J, Conese M, Hoekstra D: Gene transfer by means of lipo- and polyplexes: role of clathrin and caveolae-mediated endocytosis. J. Liposome Res. 16, 237–247 (2006).
159 Stebelska K, Wyrozumska P, Gubernator J, Sikorski AF: Higly fusogenic cationic liposomes transiently permeabilize the plasma membrane of HeLa cells. Cell. Mol. Biol. Lett. 12, 39–50 (2007).
160 Rea JC, Barron AE, Shea LD: Peptide-mediated lipofection is governed by lipoplex physical properties and the density of surface-displayed amines. J. Pharm. Sci. 97, 4794–4806 (2008).
161 Kneuer C, Ehrhardt C, Bakowsky H et al.: The influence of physicochemical parameters on the efficacy of non-viral DNA transfection complexes: a comparative study. J. Nanosci. Nanotechnol. 6, 2776–2782 (2006).
162 Rejman J, Oberle V, Zuhorn IS, Hoekstra D: Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem. J. 377, 159–169 (2004).
163 Ross PC, Hui SW: Lipoplex size is a major determinant of in vitro lipofection efficiency. Gene Ther. 6, 651–659 (1999).
164 Hassani Z, Lemkine GF, Erbacher P et al.: Lipid-mediated siRNA delivery down-regulates exogenous gene expression in the mouse brain at picomolar levels. J. Gene Med. 7, 198–207 (2005).

Nanomedicine (2010) 5(7) future science group
Review Donkuru, Badea, Wettig, Verrall, Elsabahy & Foldvari
1126
165 Resina S, Prevot P, Thierry AR: Physico-chemical characteristics of lipoplexes influence cell uptake mechanisms and transfection efficacy. PLoS One 4, e6058 (2009).
166 Margineanu A, De Feyter S, Melnikov S et al.: Complexation of lipofectamine and cholesterol-modified DNA sequences studied by single-molecule fluorescence techniques. Biomacromolecules 8, 3382–3392 (2007).
167 Higuchi Y, Kawakami S, Fumoto S, Yamashita F, Hashida M: Effect of the particle size of galactosylated lipoplex on hepatocyte-selective gene transfection after intraportal administration. Biol. Pharm. Bull. 29, 1521–1523 (2006).
168 Mehier-Humbert S, Bettinger T, Yan F, Guy RH: Plasma membrane poration induced by ultrasound exposure: implication for drug delivery. J. Control Release 104, 213–222 (2005).
169 Ghonaim HM, Ahmed OAA, Pourzand C, Blagbrough IS: Varying the chain length in N4,N9-diacyl spermines: non-viral lipopolyamine vectors for efficient plasmid DNA formulation. Mol. Pharmaceutics 5, 1111–1121 (2008).
170 Berezhna S, Schaefer S, Heintzmann R et al.: New effects in polynucleotide release from cationic lipid carriers revealed by confocal imaging, fluorescence cross-correlation spectroscopy and single particle tracking. Biochim. Biophys. Acta 1669, 193–207 (2005).
171 Klein E, Ciobanu M, Klein J et al.: “HFP” fluorinated cationic lipids for enhanced lipoplex stability and gene delivery. Bioconjug. Chem. 21, 360–371 (2010).
172 Badea I, Wettig S, Verrall R, Foldvari M: Topical non-invasive gene delivery using gemini nanoparticles in interferon-g-deficient mice. Eur. J. Pharm. Biopharm. 65, 414–422 (2007).
173 Kreiss P, Cameron B, Rangara R et al.: Plasmid DNA size does not affect the physicochemical properties of lipoplexes but modulates gene transfer efficiency. Nucleic Acids Res. 27, 3792–3798 (1999).
174 Yin W, Xiang P, Li Q: Investigations of the effect of DNA size in transient transfection assay using dual luciferase system. Anal. Biochem. 346, 289–294 (2005).
175 McLenachan S, Sarsero JP, Ioannou PA: Flow-cytometric analysis of mouse embryonic stem cell lipofection using small and large DNA constructs. Genomics 89, 708–720 (2007).
176 Madeira C, Loura LM, Prieto M, Fedorov A, Aires-Barros MR: Effect of ionic strength and presence of serum on lipoplexes structure monitorized by FRET. BMC Biotechnol. 8, 20 (2008).
177 Fisicaro E, Compari C, Duce E, Donofrio G, Rozycka-Roszak B, Wozniak E: Biologically active bisquaternary ammonium chlorides: physico-chemical properties of long chain
amphiphiles and their evaluation as non-viral vectors for gene delivery. Biochim. Biophys. Acta 1722, 224–233 (2005).
178 Srinivasan C, Burgess DJ: Optimization and characterization of anionic lipoplexes for gene delivery. J. Control Release 136, 62–70 (2009).
179 Khiati S, Pierre N, Andriamanarivo S et al.: Anionic nucleotide-lipids for in vitro DNA transfection. Bioconjug. Chem. 20, 1765–1772 (2009).
180 Caracciolo G, Pozzi D, Caminiti R: Enhanced transfection efficiency of multicomponent lipoplexes in the regime of optimal membrane charge density. J. Phys. Chem. B 112, 11298–11304 (2008).
181 Caracciolo G, Pozzi D, Caminiti R et al.: Transfection efficiency boost by designer multicomponent lipoplexes. Biochim. Biophys. Acta 1768, 2280–2292 (2007).
182 Liang H, Harries D, Wong GC: Polymorphism of DNA-anionic liposome complexes reveals hierarchy of ion-mediated interactions. Proc. Natl Acad. Sci. USA 102, 11173–11178 (2005).
183 Audouy S, Hoekstra D: Cationic lipid-mediated transfection in vitro and in vivo (review). Mol. Membr. Biol. 18, 129–143 (2001).
184 Israelachvili JN: Intermolecular and Surface Forces: with Applications to Colloidal and Biological Systems. Academic Press, UK (1992).
185 Tresset G: The multiple faces of self-assembled lipidic systems: PMC Biophys 2, 3 (2009).
186 Gonzalez-Perez A, Schmutz M, Waton G, Romero MJ, Krafft MP: Isolated fluid polyhedral vesicles. J. Am. Chem. Soc. 129, 756–757 (2007).
187 Caracciolo G, Pozzi D, Amenitsch H, Caminiti R: Multicomponent cationic lipid-DNA complex formation: role of lipid mixing. Langmuir 21, 11582–11587 (2005).
188 Chittimalla C, Zammut-Italiano L, Zuber G, Behr JP: Monomolecular DNA nanoparticles for intravenous delivery of genes. J. Am. Chem. Soc. 127, 11436–11441 (2005).
189 Blessing T, Remy JS, Behr JP: Monomolecular collapse of plasmid DNA into stable virus-like particles. Proc. Natl Acad. Sci. USA 95, 1427–1431 (1998).
190 Zuhorn IS, Kalicharan R, Hoekstra D: Lipoplex-mediated transfection of mammalian cells occurs through the cholesterol-dependent clathrin-mediated pathway of endocytosis. J. Biol. Chem. 277, 18021–18028 (2002).
191 Xu Y, Hui SW, Frederik P, Szoka FC Jr: Physicochemical characterization and purification of cationic lipoplexes. Biophys. J. 77, 341–353 (1999).
192 Sternberg B, Sorgi FL, Huang L: New structures in complex formation between DNA and cationic liposomes visualized by
freeze-fracture electron microscopy. FEBS Lett. 356, 361–366 (1994).
193 Peng SF, Su CJ, Wei MC et al.: Effects of the nanostructure of dendrimer/DNA complexes on their endocytosis and gene expression. Biomaterials 21, 5660–5670 (2010).
194 Ayoubi MA, Zinchenko AA, Philippova OE, Khokhlov AR, Yoshikawa K: Visualization of different pathways of DNA release from interpolyelectrolyte complex. J. Phys. Chem. B 111, 8373–8378 (2007).
195 Rodriguez-Pulido A, Ortega F, Llorca O, Aicart E, Junquera E: A physicochemical characterization of the interaction between DC-Chol/DOPE cationic liposomes and DNA. J. Phys. Chem. B 112, 12555–12565 (2008).
196 Frerix A, Schonewald M, Geilenkirchen P, Muller M, Kula MR, Hubbuch J: Exploitation of the coil-globule plasmid DNA transition induced by small changes in temperature, pH salt, and poly(ethylene glycol) compositions for directed partitioning in aqueous two-phase systems. Langmuir 22, 4282–4290 (2006).
197 Elkady AS, Zhdanov RI: Dicationic DEGA-based lipid systems for gene transfer and delivery: supramolecular structure and activity. In: Nanocarrier Technologies Frontiers of Nanotherapy (Mozafari MR Ed.). Springer, The Netherlands 175–190 (2006).
198 Ito T, Iida-Tanaka N, Koyama Y: Efficient in vivo gene transfection by stable DNA/PEI complexes coated by hyaluronic acid. J. Drug Target 16, 276–281 (2008).
199 Ewert K, Slack NL, Ahmad A et al.: Cationic lipid-DNA complexes for gene therapy: Understanding the relationship between complex structure and gene delivery pathways at the molecular level. Curr. Med. Chem. 11, 133–149 (2004).
200 Lin AJ, Slack NL, Ahmad A, George CX, Samuel CE, Safinya CR: Three-dimensional imaging of lipid gene-carriers: membrane charge density controls universal transfection behavior in lamellar cationic liposome-DNA complexes. Biophys. J. 84, 3307–3316 (2003).
201 Uhríková D, Zajac I, Dubnicková M et al.: Interaction of gemini surfactants butane-1,4-diyl-bis(alkyldimethylammonium bromide) with DNA. Colloids Surf. B. Biointerfaces 42, 59–68 (2005).
202 Uhrikova D, Hanulova M, Funari SS, Lacko I, Devinsky F, Balgavy P: The structure of DNA-DLPC-cationic gemini surfactant aggregates: a small angle synchrotron X-ray diffraction study. Biophys Chem 111, 197–204 (2004).
203 Mirska D, Schirmer K, Funari SS, Langner A, Dobner B, Brezesinski G: Biophysical and biochemical properties of a binary lipid mixture for DNA transfection. Colloids Surf. B Biointerfaces 40, 51–59 (2005).

www.futuremedicine.comfuture science group
Soft nanoparticles of DNA: gene delivery Review
www.futuremedicine.com 1127
204 Foldvari M, Badea I, Wettig S, Verrall R, Bagonluri M: Structural characterization of novel gemini non-viral DNA delivery systems for cutaneous gene therapy. J. Exp. Nanosci. 1, 165–176 (2006).
205 Foldvari M, Wettig S, Badea I, Verrall R, Bagonluri M: Dicationic gemini surfactant gene delivery complexes contain cubic-lamellar mixed polymorphic phase. Presented at: NSTI Nanotech 2006, NSTI Nanotechnology Conference and Trade Show, Boston, MA, United States, 7–11 May, 2, 400–403 (2006).
206 Sun P, Zhong M, Shi X, Li Z: Anionic LPD complexes for gene delivery to macrophage: preparation, characterization and transfection in vitro. J. Drug Target 16, 668–678 (2008).
207 Burke RS, Pun SH: Extracellular barriers to in vivo PEI and PEGylated PEI polyplex-mediated gene delivery to the liver. Bioconjug. Chem. 19, 693–704 (2008).
208 McNeil SE, Vangala A, Bramwell VW, Hanson PJ, Perrie Y: Lipoplexes formulation and optimisation: in vitro transfection studies reveal no correlation with in vivo vaccination studies. Curr. Drug Deliv. 7, 175–187 (2010).
209 Huang YC, Riddle K, Rice KG, Mooney DJ: Long-term in vivo gene expression via delivery of PEI-DNA condensates from porous polymer scaffolds. Hum. Gene Ther. 16, 609–617 (2005).
210 Neves SS, Sarmento-Ribeiro AB, Simoes SP, Pedroso de Lima MC: Transfection of oral cancer cells mediated by transferrin-associated lipoplexes: mechanisms of cell death induced by herpes simplex virus thymidine kinase/ganciclovir therapy. Biochim. Biophys. Acta 1758, 1703–1712 (2006).
211 Pfeiffer T, Wallich M, Sandmann W, Schrader J, Godecke A: Lipoplex gene transfer of inducible nitric oxide synthase inhibits the reactive intimal hyperplasia after expanded polytetrafluoroethylene bypass grafting. J. Vasc. Surg. 43, 1021–1027 (2006).
212 Morton TJ, Furst W, Griensven Mv, Redl H: Controlled release of substances bound to fibrin-anchors or of DNA. Drug Delivery 16, 102–107 (2009).
213 Lisziewicz J, Calarota SA, Lori F: The potential of topical DNA vaccines adjuvanted by cytokines. Expert Opin. Biol. Ther. 7, 1563–1574 (2007).
214 Alcon VL, Baca-Estrada ME, Potter A, Babiuk LA, Kumar P, Foldvari M: Biphasic lipid vesicles as a subcutaneous delivery system for protein antigens and CpG oligonucleotides. Curr. Drug Deliv. 3, 129–135 (2006).
215 Babiuk S, Baca-Estrada ME, Middleton DM, Hecker R, Babiuk LA, Foldvari M: Biphasic lipid vesicles (Biphasix) enhance the adjuvanticity of CpG oligonucleotides following systemic and mucosal administration. Curr. Drug Deliv. 1, 9–15 (2004).
216 Babiuk S, Baca-Estrada ME, Pontarollo R, Foldvari M: Topical delivery of plasmid DNA using biphasic lipid vesicles (Biphasix). J. Pharm. Pharmacol. 54, 1609–1614 (2002).
217 Williams JA, Carnes AE, Hodgson CP: Plasmid DNA vaccine vector design: impact on efficacy, safety and upstream production. Biotechnol. Adv. 27, 353–370 (2009).
218 Kutzler MA, Weiner DB: DNA vaccines: ready for prime time? Nat. Rev. Genet. 9, 776–788 (2008).
219 Anwer K, Barnes MN, Fewell J, Lewis DH, Alvarez RD: Phase-I clinical trial of IL-12 plasmid/lipopolymer complexes for the treatment of recurrent ovarian cancer. Gene Ther. 17, 360–369 (2010).
220 Cui Z, Mumper RJ: Topical immunization using nanoengineered genetic vaccines. J. Control Release 81, 173–184 (2002).
221 Hafez IM, Ansell S, Cullis PR: Tunable pH-sensitive liposomes composed of mixtures of cationic and anionic lipids. Biophys. J. 79, 1438–1446 (2000).
222 Zhang S, Xu Y, Wang B, Qiao W, Liu D, Li Z: Cationic compounds used in lipoplexes and polyplexes for gene delivery. J. Control Release 100, 165–180 (2004).
223 Alexander MY, Akhurst RJ: Liposome-medicated gene transfer and expression via the skin. Hum. Mol. Genet. 4, 2279–2285 (1995).
224 Horiguchi Y, Larchian WA, Kaplinsky R, Fair WR, Heston WD: Intravesical liposome-mediated interleukin-2 gene therapy in orthotopic murine bladder cancer model. Gene Ther. 7, 844–851 (2000).
225 Subramanian M, Holopainen JM, Paukku T, Eriksson O, Huhtaniemi I, Kinnunen PKJ: Characterization of three novel cationic lipids as liposomal complexes with DNA. Biochim. Biophys. Acta 1466, 289–305 (2000).
226 Gao X, Huang L: A novel cationic liposome reagent for efficient transfection of mammalian cells. Biochem Biophys Res Commun 179, 280–285 (1991).
227 Castano S, Delord B, Fevrier A, Lehn J-M, Lehn P, Desbat B: Brewster angle microscopy and PMIRRAS study of DNA interactions with BGTC, a cationic lipid used for gene transfer. Langmuir 24, 9598–9606 (2008).
228 Vigneron JP, Oudrhiri N, Fauquet M et al.: Guanidinium-cholesterol cationic lipids: efficient vectors for the transfection of eukaryotic cells. Proc. Natl Acad. Sci. USA 93, 9682–9686 (1996).
229 Gaucheron J, Wong T, Wong KF, Maurer N, Cullis PR: Synthesis and properties of novel tetraalkyl cationic lipids. Bioconj. Chem. 13, 671–675 (2002).
230 Bhattacharya S, De S, George SK: Synthesis and vesicle formation from novel
pseudoglyceryl dimeric lipids. Evidence of formation of widely different membrane organizations with exceptional thermotropic properties. Chem. Comm. (Camb.) 2287–2288 (1997).
231 Boomer JA, Thompson DH, Sullivan SM: Formation of plasmid-based transfection complexes with an acid-labile cationic lipid: characterization of in vitro and in vivo gene transfer. Pharm. Res. 19, 1292–1301 (2002).
232 Baccaglini L, Shamsul Hoque AT, Wellner RB et al.: Cationic liposome-mediated gene transfer to rat salivary epithelial cells in vitro and in vivo. J. Gene Med. 3, 82–90 (2001).
233 Sola B, Staedel C, Remy JS, Bahr A, Behr JP: Lipospermine-mediated gene transfer technique into murine cultured cortical cells. J. Neurosci. Methods 71, 183–186 (1997).
234 Baca-Estrada ME, Foldvari M, Snider M, Van Drunen Littel-Van Den Hurk S, Babiuk LA: Effect of IL-4 and IL-12 liposomal formulations on the induction of immune response to bovine herpesvirus type-1 glycoprotein D. Vaccine 15, 1753–1760 (1997).
235 Remy JS, Sirlin C, Vierling P, Behr JP: Gene transfer with a series of lipophilic DNA-binding molecules. Bioconjug. Chem. 5, 647–654 (1994).
236 Sen J, Chaudhuri A: Design, syntheses, and transfection biology of novel non-cholesterol-based guanidinylated cationic lipids. J. Med. Chem. 48, 812–820 (2005).
237 Fielden ML, Perrin C, Kremer A et al.: Sugar-based tertiary amino gemini surfactants with a vesicle-to-micelle transition in the endosomal pH range mediate efficient transfection in vitro. Eur. J. Biochem. 268, 1269–1279 (2001).
238 Kirby AJ, Camilleri P, Engberts JB et al.: Gemini surfactants: new synthetic vectors for gene transfection. Angew. Chem. Int. Ed. Engl. 42, 1448–1457 (2003).
239 McGregor C, Perrin C, Monck M, Camilleri P, Kirby AJ: Rational approaches to the design of cationic gemini surfactants for gene delivery. J. Am. Chem. Soc. 123, 6215–6220 (2001).
240 Hatakeyama H, Akita H, Kogure K et al.: Development of a novel systemic gene delivery system for cancer therapy with a tumor-specific cleavable PEG-lipid. Gene Ther. 14, 68–77 (2007).
�n Website301 Vector-Statistics. Gene Therapy Clinical Trials
Worldwide. J. Gene Med. (2009, latest update). www.wiley.co.uk/genetherapy/clinical/