Embed Size (px)
Citation preview

Brain Research 920 (2001) 32–40www.elsevier.com/ locate /bres
Research report
Advanced glycation endproducts co-localize with inducible nitric oxidesynthase in Alzheimer’s disease
a b a a¨Amanda Wong , Hans-Joachim Luth , Winnie Deuther-Conrad , Sladjana Dukic-Stefanovic ,a b a ,*¨Jovana Gasic-Milenkovic , Thomas Arendt , Gerald Munch
aNeuroimmunological Cell Biology, IZKF Leipzig, Johannisallee 30a, 04103 Leipzig, GermanybPaul Flechsig Institute for Brain Research, Department of Neuroanatomy, University of Leipzig, Jahnallee 59, 04109 Leipzig, Germany
Accepted 17 July 2001
Abstract
Advanced glycation endproducts (AGEs), protein-bound oxidation products of sugars, have been shown to be involved in thepathophysiological processes of Alzheimer’s disease (AD). AGEs induce the expression of various pro-inflammatory cytokines and theinducible nitric oxide synthase (iNOS) leading to a state of oxidative stress. AGE modification and resulting crosslinking of proteindeposits such as amyloid plaques may contribute to the oxidative stress occurring in AD. The aim of this study was to immuno-histochemically compare the localization of AGEs and b-amyloid (Ab) with iNOS in the temporal cortex (Area 22) of normal and ADbrains. In aged normal individuals as well as early stage AD brains (i.e. no pathological findings in isocortical areas), a few astrocytesshowed co-localization of AGE and iNOS in the upper neuronal layers, compared with no astrocytes detected in young controls. In lateAD brains, there was a much denser accumulation of astrocytes co-localized with AGE and iNOS in the deeper and particularly upperneuronal layers. Also, numerous neurons with diffuse AGE but not iNOS reactivity and some AGE and iNOS-positive microglia weredemonstrated, compared with only a few AGE-reactive neurons and no microglia in controls. Finally, astrocytes co-localized with AGEand iNOS as well as AGE and were found surrounding mature but not diffuse amyloid plaques in the AD brain. Our results show thatAGE-positive astrocytes and microglia in the AD brain express iNOS and support the evidence of an AGE-induced oxidative stressoccurring in the vicinity of the characteristic lesions of AD. Hence activation of microglia and astrocytes by AGEs with subsequentoxidative stress and cytokine release may be an important progression factor in AD. 2001 Elsevier Science B.V. All rights reserved.
Theme: Disorders of the nervous system
Topic: Degenerative disease: Alzheimer’s — other
Keywords: Astroglia; Advanced glycation endproduct; Alzheimer’s disease; b-Amyloid; Microglia; Nitric oxide synthase
1. Introduction characteristic manner in affected regions of the brainduring the different stages of the disease [5,14]. One
Alzheimer’s disease (AD) is a neurodegenerative disease characteristic modification of these long-lived proteins iswhich affects a growing proportion of the elderly popula- advanced glycation endproducts (AGEs), sugar-derivedtion. The clinical manifestation of this age-related neurode- protein adducts [46]. AGEs have not only been foundgenerative disease is a gradual onset and progression of closely associated with the SP of the AD brain but alsomemory loss and other cognitive functions. Characteristic with activated microglia. Widespread signs of oxidativehistopathological hallmarks of AD are neurofibrillary damage including nitration of tyrosine residues in proteinstangles and senile plaques (SP), composed of highly cross- are also evident in affected regions in AD suggesting that,linked and oxidized protein polymers, which develop in a among other radical species, NO production is upregulated
[15,40].A further characteristic feature of the disease is a*Corresponding author. Tel.: 149-341-9715-945; fax: 149-341-9715-
chronic, but silent, acute phase reaction (which is common-979.¨E-mail address: [email protected] (G. Munch). ly referred to as ‘neuroinflammation’) in affected brain
0006-8993/01/$ – see front matter 2001 Elsevier Science B.V. All rights reserved.PI I : S0006-8993( 01 )02872-4

A. Wong et al. / Brain Research 920 (2001) 32 –40 33
regions, which is best described as an innate immuno- pathological signs were detected by neuropathologicalreaction and does not involve B- and T-cells [1,30]. This examination. The clinical diagnosis of AD was based onacute phase reaction is evident by both the presence of the occurrence of significant intellectual dysfunction, i.e.activated microglia and astroglia, releasing pro-inflamma- the presence of deficits in at least four aspects of cognitivetory cytokines and chemokines including interleukin (IL)- and social behavior. Other causes of dementia were1, IL-6 and tumour necrosis factor-a as well as free excluded by medical, psychiatric and paraclinical examina-oxygen and nitrogen radicals [41]. tion (Diagnostic and Statistical Manual of Mental Dis-
The source of this inflammatory reaction is most likely orders, DSM-IIIR, American Psychiatric Association).the SP itself. Most of the published studies suggest that Cases with a history of diabetes mellitus were excluded.fibrillar amyloid alone can act as a pro-inflammatory Each case met the National Institute of Neurologic andligand. Other studies with b2-microglobulin, a major Communicative Disorders and Stroke (NINCDS) andconstituent of the amyloid deposits in b2-microglobulin Alzheimer’s Disease and Related Disorders Associationamyloidosis, indicate that AGEs induce or potentiate the (ADRDA) criteria for definite diagnosis of Alzheimer’sinflammatory response [10,34]. disease [31], based on the presence of NFTs and neuritic
The aim of this study was to immunohistochemically plaques observed in the hippocampal formation andcompare the localization of AGEs and Ab with iNOS in neocortical areas, as recommended [17].the temporal cortex of young and aged controls and inearly- and late-stage AD brains. This would allow us to 2.2. Preparation of tissueclarify the possible role of AGEs or Ab in the induction ofiNOS in vivo, and the cell types, which might be involved Tissue blocks were taken from the temporal cortexin the pathological process of oxidative stress. (Brodmann Area 22) and immersed in 4%
paraformaldehyde/0.5% glutaraldehyde in phosphate buf-fer (0.1 M; pH 7.4) for 4 days at 48C. Blocks were
2. Material and methods subsequently immersed in 15% sucrose in phosphate-buf-fered saline (PBS) for 24 h, followed by 30% sucrose in
2.1. Cases PBS for 48 h. Coronal sections, 30 mm thick, were cut on afreezing microtome and processed for the immunohistoch-
Brains used in the present study were obtained from emical detection of AGE, iNOS, b-amyloid and microglia.eight controls, subdivided into an older (65–90 years, four The Gallyas staining method for demonstration of neuriticmales) and a younger (30–50 years, four males) group and plaques, neurofibrillary tangles and neuropil threads wasfrom 12 patients with AD, subdivided into an advanced performed following the original description [13].(Braak stage III–VI; one male, seven females) and early(Braak stage I–II; three males) stage of the disease [4]. 2.3. ImmunohistochemistryThe AD cases were matched with respect to age (meanage: aged controls; 79.368.1 years; AD, 83.866.6 years; Free floating sections were briefly boiled in sodiumP.0.20, Student’s t-test), post-mortem interval (aged citrate buffer (NaCl 150 mM, Na-citrate 100 mM, pH 6)controls: 64630 h; AD: 47633 h; P.0.20) and the ‘Pre- and pre-incubated with 1% H O for 30 min, followed by2 2
mortem Severity Index’ (PMSI) by Monfort et al. [36] blocking of unspecific binding sites with 5% normal goat(P.0.20) to minimize the likelihood of an artificial serum (Sigma), 0.5% Triton X-100 (Ferak, Berlin) in 0.1 Ninfluence by pre-mortem hypoxia and hypovolemia. The Tris-buffered saline (TBS, pH 7.4) for 1 h. Next, sectionsyoung control group (mean age: 39 years) was used to were incubated with one of the following primary anti-evaluate age-related changes as well as to compare with bodies overnight at 48C: polyclonal anti-AGE (1:750,changes found in early stage AD. The whole procedure of RNAse-AGE K1936, Roche Diagnostics), polyclonal anti-case recruitment, acquisition of patients’ personal data, iNOS (1:500, Alexis, clone L05074), monoclonal anti-b-performing the autopsy, and handling the autoptic material amyloid (1:50, Dako, clone 6F/3D), monoclonal anti-was performed in accordance with the Helsinki Declaration monocyte (1:2, Biologo, MAC 387). In additions to thein its latest version and the convention of the council of manufacturers’ specifications, specificity of the iNOS andEurope on Human Rights and Biomedicine, and was AGE K1936 antibody was also checked by Western blot,approved by the responsible Ethical Committee of Leipzig where recognized bands of the appropriate molecularUniversity. weight were recognized. Primary antibodies were detected
Brains used for the group of controls were obtained at by a green IgG-Cy2 (1:100, Dianova) or an IgG-Cy3routine autopsy from patients who died without a history (1:1000, Dianova) by immunofluorescence, respectively.of neuropsychiatric disorder or mental impairment or For simultaneous detection AGE and iNOS, sections werediabetes mellitus. There had to be clear evidence that the first incubated with the anti-AGE antibody, then with apatient was alert, well oriented, and capable of functioning green CyDye (fluorescent)-labelled species-specific sec-relatively independently shortly before death. No ondary antibody, goat anti-rabbit IgG-Cy2 (1:100,

34 A. Wong et al. / Brain Research 920 (2001) 32 –40
Dianova) and reblocked. The sections were then incubated lysine- and arginine-specific anti-AGE antibodies werewith anti-iNOS antibody, followed by a red CyDye seen, the arginine-specific anti-AGE antibody K 1936(fluorescent)-labelled species-specific secondary antibody, worked best and was used for all further experiments. AGEgoat anti-rabbit IgG-Cy3 (1:1000, Dianova). For simulta- immunostaining could be eliminated by omission of theneous detection of AGE and b-amyloid /microglia or iNOS primary antibody and also by pre-adsorption with excessand b-amyloid /microglia, sections were incubated with a antigen (1 mg/ml BSA-AGE).cocktail containing anti-AGE and the corresponding anti-body. Immunodetection was carried out accordingly with a
3.2. Distribution of iNOS and AGE in young and agedcocktail of the CyDye-labelled species-specific secondarycontrol temporal cortexantibodies goat anti-rabbit IgG-Cy2 (1:100, Dianova) and
goat anti-mouse IgG-Cy3 (1:1000, Dianova). All incuba-In the four young control brains, AGE reactivity wastion steps were separated by intensive washing with TBS.
observed in blood vessels only. Neither AGE nor iNOSSections were mounted, covered with Entellan (Merck)was detected in nerve or glial cells. In the four agedand scanned with a laser scanning microscope (LSM 510,control brains, AGE but not iNOS reactivity was observedZeiss). The Cy2 channel collected the anti-AGE immuno-in blood vessels and in the soma and proximal dendrites oflabelled structures, while the Cy3 channel collected thea few neurons (data not shown). In one of these cases,anti-b-amyloid, anti-microglia or anti-iNOS structures. Forthere was some AGE and iNOS immunoreactivity ob-the green fluorescence (Cy2) the argon laser (excitationserved in the soma and processes of a few astroglia (data488 nm) and the emission filter 3P (505–530 nm) werenot shown).applied. For Cy3, an excitation of 543 nm and emission
filter LP 560 nm were used. Sections were also examinedand digitized by the use of an Axiophot fluorescence 3.3. Distribution of iNOS and AGE in early ADmicroscope equipped with a SL XRS CCD camera and temporal cortexappropriate Zeiss filter combinations for red Cy3 (no. 15)and green Cy2 (no. 09) fluorescence. In brains diagnosed with early AD (Braak stage I–II), a
few pyramidal neurons were diffuse AGE but no iNOSreactive. Scattered astrocytes in the upper and deeper
3. Results neuronal layers showed AGE and iNOS immunoreactivityin the cell processes (data not shown).
3.1. Comparison of AGE antibodies
Three polyclonal anti-AGE antibodies (Roche Diagnos- 3.4. Distribution of iNOS and AGE in advanced ADtics, Tutzing) were tested, two lysine-specific and one temporal cortexarginine-specific (unpublished observations), and comparedfor the structures which they recognized in both AD and An abundance of astrocytes was found immunoreactivenormal brain sections. The results are listed in Table 1. In for AGEs (Fig. 1A,B) and iNOS (Fig. 1A9,B9) in thesummary, all three anti-AGE antibodies stained blood temporal cortex of medium to severely affected (Braakvessels and neurons in both AD and control brain sections. stages III–VI) AD brains. Whereas most of the reactiveIn addition, plaques and astroglial processes, confirmed astrocytes were located in the upper (I and II) layers andmorphologically, were detected in AD tissue. Although grouped in clusters, some single astrocytes were alsominimal differences in the structures stained between the distributed in the deeper neuronal layers. AGE staining
Table 1Structures stained by various anti-AGE antibodies in aged control and AD brain (Area 22)
Antibody Immunogen Age specificity Case type Structures stained
K1935 RNAse-AGE Lysine Aged Blood vessels, punctate clusters(polyclonal) control in WM and GM, neurons
AD Blood vessels, punctate clusters in WM and GM,neurons, some astroglia, diffuse plaques
K1936 RNAse-AGE Arginine Aged Blood vessels, punctate clusters(polyclonal) control in WM and GM, neurons
AD Blood vessels, punctate clusters in WM and GM,neurons, some astroglia, diffuse plaques
K1937 RNAse-AGE Lysine Aged Blood vessels, punctate clusters(polyclonal) control in WM and GM, neurons
AD Blood vessels, punctate clusters in WM and GM,neurons, some astroglia, diffuse plaques

A. Wong et al. / Brain Research 920 (2001) 32 –40 35
appeared more diffuse in the processes compared with 4. DiscussioniNOS staining, which was observed in the cell soma aswell as processes of astrocytes. Some co-localization was In this study, major differences were observed in theobserved in astrocyte processes (Fig. 1A0,B0). Plaques distribution of AGE and iNOS between control and ADthroughout the neuronal layers were also stained AGE- temporal cortices. In the aged control and early AD but notpositive. The soma, apical and proximal basal dendrites of young control brains, some astrocytes were AGE andnumerous and selective pyramidal neurons in the deeper iNOS reactive. In comparison, in late AD cases, a largerlayers of the temporal cortex showed diffuse AGE- but not accumulation of AGE and iNOS-staining astrocytes, par-iNOS-reactivity (Fig. 2A). Whether the AGE deposits are ticularly in the upper neuronal layers was detected. Neu-located in vacuoles (as previously described [43]) or are rons with diffuse AGE but not iNOS reactivity were foundfreely distributed in the cytoplasm is unclear. most numerously in late AD, compared with early AD and
control brains. Furthermore, punctate AGE and iNOSstaining was detected in some reactive microglia in late
3.5. Distribution of iNOS and AGE in microglia inAD but not in aged control temporal cortices. Finally, we
advanced AD temporal cortexdemonstrated co-localization of AGE and iNOS as well asAGE and b-amyloid in astrocytes adjacent to mature but
Since iNOS expression and AGE-mediated activation isnot diffuse b-amyloid plaques in the AD brain.
known to occur in microglia as well as astrocytes, amonoclonal antibody against reactive macrophages was
4.1. iNOS expression in astrocytes and microglia in ADused to verify microglial involvement. Using this antibody,we observed the presence of numerous microglia in the
From our results, it is suggested that iNOS is expressedmiddle to deeper neuronal layers of the AD temporal
by astrocytes and, to a lesser extent, by some microglia incortex (Fig. 1C9,D9). In some of these microglia, punctate
the AD brain. Roles proposed for NO in CNS patho-AGE reactivity was detected (Fig. 1C). In addition, we
physiology are increasingly diverse and range from inter-were able to demonstrate that some reactive microglia
cellular signaling, through necrotic killing of cells andcontain iNOS (Fig. 1D). AGE and iNOS staining in the
invading pathogens, to the involvement of NO in apoptosissoma and processes of reactive microglia appeared granu-
[6,12,20]. High levels of NO, synthesized by iNOS may belar and sparse, compared with the more diffuse staining in
cytotoxic, due to peroxynitrite formation and nitration ofthe astrocytes. Reactive microglia not co-localized with
tyrosine residues in proteins [21,24,42] or the generation ofAGE were also located adjacent to AGE-positive pyrami-
superoxides [21,24,38]. It is well established that as-dal neurons (Fig. 2A0). No reactive microglia were de-
trocytes and microglia can be induced by various cytokinestected in the older control group.
or (when stimulated) by b-amyloid to express iNOS invitro [2,19,33]. Although astrocytes have been suggested
3.6. Differential pattern of AGE /iNOS staining to be a major source of NO in the human brain, theaccording to plaque development expression of iNOS in human microglia is controversial.
Whilst other studies provide evidence for the expression ofSenile plaques in the temporal cortex of AD brains in iNOS in human astrocytes but not in reactive microglia
the advanced Braak stages (III–VI) were stained with an [25,48,52], an increase in NO production in the CHME-5antibody against the b-amyloid peptide. Double immuno- human microglial cell line [9] and co-localization of iNOSfluorescence with anti-AGE and anti-b-amyloid antibodies and microglia in human immunodeficiency virus (HIV)-showed AGE-positive astrocytes surrounding mature but infected [39], ischaemic [28] and oedemic [27] brains hasnot diffuse amyloid plaques (Fig. 2B). In some of these been reported. Despite the functional differences for iNOSAGE-positive astrocytes, co-localization with b-amyloid between mice and human, it is possible that humanwas seen. Furthermore, double immunofluorescence with microglia can be induced to express iNOS and generateanti-AGE and anti-iNOS antibodies showed accumulation NO, particularly in light of their role in secreting freeof iNOS-positive astrocytes around mature amyloid radicals and inflammatory cytokines.plaques (Fig. 2C). No co-localization with b-amyloid and We did not observe any iNOS-positive neurons in ADiNOS was seen in these astrocytes. In fluorescent as well nor control temporal cortices. This is in agreement with
¨as DAB preparations, some light AGE- as well as iNOS- Wallace et al. [48] and Luth et al. [29], who demonstratedstaining of plaques was evident. Brain sections stained iNOS in astrocytes but not neurons in the AD brain.with AGE-DAB together with the fluorescent marker, Vodovotz et al. [47], however, reported the presence ofThioflavin S, to visualize plaques also showed AGE- iNOS in AD tangle-bearing neurons but not astrocytes.positive astrocytes in the vicinity of mature plaques (datanot shown). In summary, localization of iNOS and AGE 4.2. AGE cause (nitro)oxidative stress in ADwas evident in astroglia associated with mature but notdiffuse plaques. Our finding of AGE in astrocytes, microglia and neurons
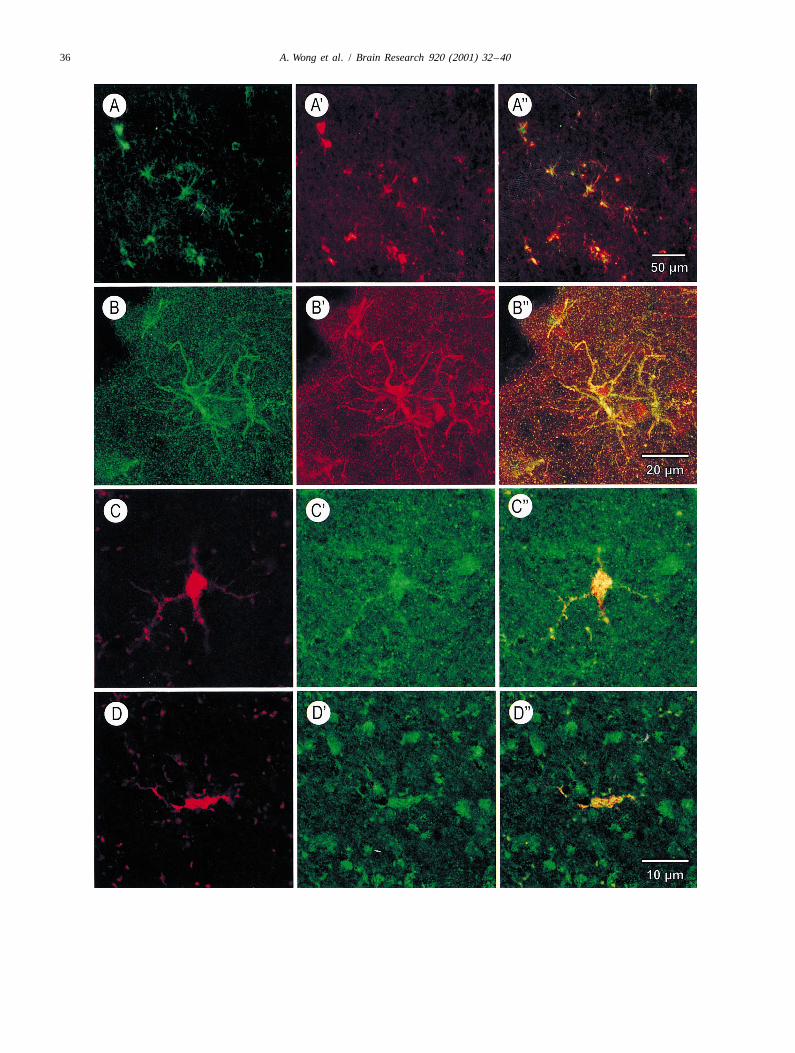
36 A. Wong et al. / Brain Research 920 (2001) 32 –40
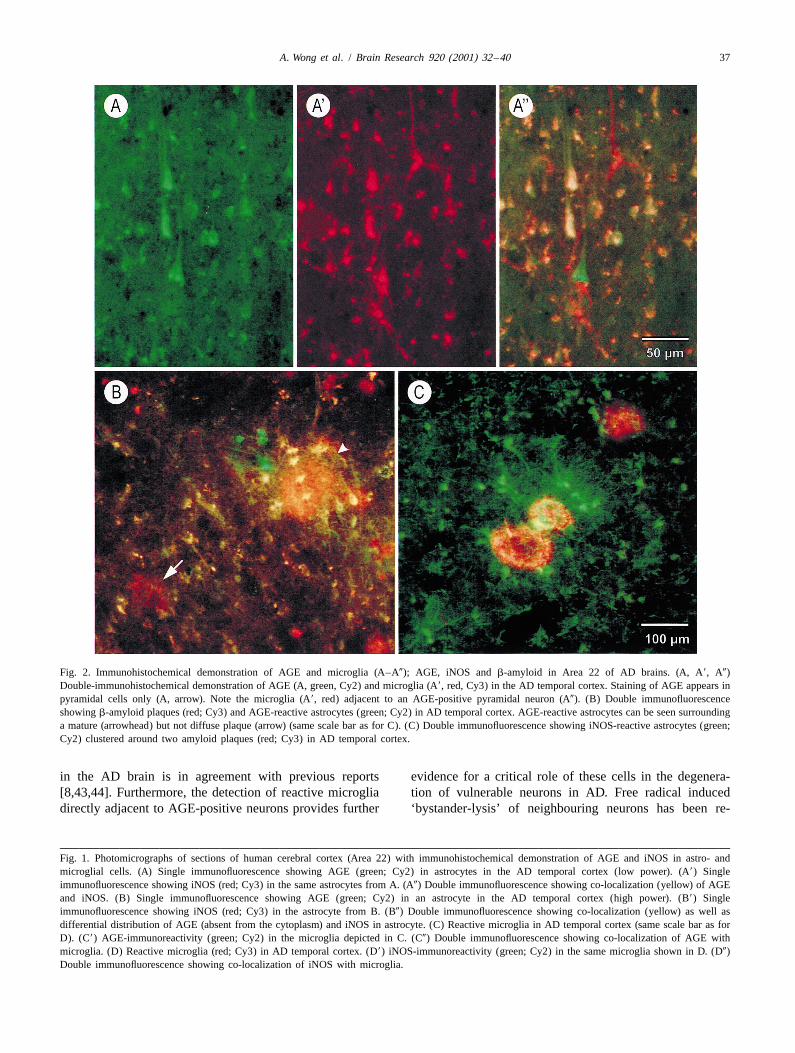
A. Wong et al. / Brain Research 920 (2001) 32 –40 37
Fig. 2. Immunohistochemical demonstration of AGE and microglia (A–A0); AGE, iNOS and b-amyloid in Area 22 of AD brains. (A, A9, A0)Double-immunohistochemical demonstration of AGE (A, green, Cy2) and microglia (A9, red, Cy3) in the AD temporal cortex. Staining of AGE appears inpyramidal cells only (A, arrow). Note the microglia (A9, red) adjacent to an AGE-positive pyramidal neuron (A0). (B) Double immunofluorescenceshowing b-amyloid plaques (red; Cy3) and AGE-reactive astrocytes (green; Cy2) in AD temporal cortex. AGE-reactive astrocytes can be seen surroundinga mature (arrowhead) but not diffuse plaque (arrow) (same scale bar as for C). (C) Double immunofluorescence showing iNOS-reactive astrocytes (green;Cy2) clustered around two amyloid plaques (red; Cy3) in AD temporal cortex.
in the AD brain is in agreement with previous reports evidence for a critical role of these cells in the degenera-[8,43,44]. Furthermore, the detection of reactive microglia tion of vulnerable neurons in AD. Free radical induceddirectly adjacent to AGE-positive neurons provides further ‘bystander-lysis’ of neighbouring neurons has been re-
Fig. 1. Photomicrographs of sections of human cerebral cortex (Area 22) with immunohistochemical demonstration of AGE and iNOS in astro- andmicroglial cells. (A) Single immunofluorescence showing AGE (green; Cy2) in astrocytes in the AD temporal cortex (low power). (A9) Singleimmunofluorescence showing iNOS (red; Cy3) in the same astrocytes from A. (A0) Double immunofluorescence showing co-localization (yellow) of AGEand iNOS. (B) Single immunofluorescence showing AGE (green; Cy2) in an astrocyte in the AD temporal cortex (high power). (B9) Singleimmunofluorescence showing iNOS (red; Cy3) in the astrocyte from B. (B0) Double immunofluorescence showing co-localization (yellow) as well asdifferential distribution of AGE (absent from the cytoplasm) and iNOS in astrocyte. (C) Reactive microglia in AD temporal cortex (same scale bar as forD). (C9) AGE-immunoreactivity (green; Cy2) in the microglia depicted in C. (C0) Double immunofluorescence showing co-localization of AGE withmicroglia. (D) Reactive microglia (red; Cy3) in AD temporal cortex. (D9) iNOS-immunoreactivity (green; Cy2) in the same microglia shown in D. (D0)Double immunofluorescence showing co-localization of iNOS with microglia.

38 A. Wong et al. / Brain Research 920 (2001) 32 –40
ported to occur in a co-culture of microglia and PC-12 and not unmodified b-amyloid alone, exert a strongcells [32]. Hence AGE accumulated in neurons could be influence in the inflammatory processes occurring in AD.one source of AGE-induced activation of microglia, re- From this study, it is proposed that an AGE-inducedsulting in the generation of free radicals and cytokines upregulation of iNOS expression could contribute to an[11,16,45] and further neuronal damage. increased level of oxidative stress, neuronal damage and
The presence of AGEs in neurons may also cause injury functional loss in the AD brain. Consequently, inhibitionvia a direct cytotoxic effect [26]. In addition, disturbances of this enzyme by antioxidant intervention could reduceare seen in lysosomal protein degradation in human AD free radical damage and inflammation and hence, may bepyramidal neurons, where endosomal enlargement contri- an important target in slowing the progression of AD.butes to a larger total endosomal volume per neuron [7],concomitantly with an increased AGE load in this cell type[23]. Acknowledgements
Co-localization of AGE and iNOS in astrocytes andmicroglia cells reported in our study not only supports a We thank Peter Stahl, Rosemarie Kientsch-Engel, Peterrole for AGE in nitro-oxidative stress in the AD brain Riederer, Glenda Halliday and Siegfried Hoyer for con-[3,11,37,45], but also a pathway for AGE-induced activa- ¨tinuous support. We also thank Mrs Barbara Bar, Dr Ulition via the upregulation of iNOS. In our study, AGEs ¨Gartner and Dr Jens Groscher for valuable technicalwere localized mainly in astrocyte processes, whereas assistance. This work was supported by the AlzheimeriNOS-immunoreactivity was found in both the processes Forschung International (AFI), the SKB Stiftung, Rocheand soma of astrocytes. This finding is in agreement with ¨Diagnostics GmbH and the Bundesministerium fur Bil-the notion that AGE receptor-mediated binding induces dung, Forschung und Technologie (BMBF), Interdiszip-oxidative stress [3,22,35,45,51], which, as our study ¨ ¨linares Zentrum fur Klinische Forschung (IZKF) at thesuggests, may involve the expression of of iNOS. The University of Leipzig (01KS9504, Project N1) (to G.M.)importance of AGEs in nitro-oxidative stress is also and the DAAD (to A.W.).confirmed by the lack of iNOS, in the absence of AGEs,-positive staining astroglia in our study.
References4.3. Do AGEs or b-amyloid induce iNOS expression?
[1] P.S. Aisen, Inflammation and Alzheimer disease, Mol. Chem.Our results show that astrocytes immunoreactive for Neuropathol. 28 (1996) 83–88.
[2] K.T. Akama, L.J. Van Eldik, Beta-amyloid stimulation of inducibleAGE and iNOS mainly appeared around mature but notnitric oxide synthase in astrocytes is interleukin-1beta- and tumordiffuse b-amyloid plaques. This supports the notion thatnecrosis factor-alpha (TNF alpha)-dependent and involves a TNFastrocytosis may be a secondary rather than primaryalpha receptor-associated factor- and NFkappaB-inducing kinase-
feature during plaque development [49]. In addition, co- dependent signalling mechanism, J. Biol. Chem. 275 (2000) 7918–localization of AGE and b-amyloid but not iNOS and 7924.
[3] A. Bierhaus, M.A. Hofmann, R. Ziegler, P.P. Nawroth, AGEs andb-amyloid in glia from our work suggests that AGEs maytheir interaction with AGE-receptors in vascular disease and dia-induce a more pronounced pro-inflammatory effect thanbetes mellitus. I. The AGE concept, Cardiovasc Res. 37 (1998)Ab alone. These data are supported by studies in non586–600.
pre-stimulated mouse microglial cells (N-11 cell line), [4] H. Braak, E. Braak, The human entorhinal cortex: normal morpholo-where LPS and AGEs induce a much higher NO pro- gy and lamina specific pathology in various diseases, Neurosci. Res.
15 (1992) 6–31.duction than Ab alone (unpublished observations). Ab has[5] H. Braak, E. Braak, J. Bohl, R. Reintjes, Age, neurofibrillarybeen reported to cause a direct pro-inflammatory effect via
changes, A-beta-amyloid and the onset of Alzheimer’s disease,the release of cytokines and free radicals [2,18,50]. Al-Neurosci. Lett. 210 (1996) 87–90.
though Ab activates some transduction factors such as [6] D. S Bredt, Endogenous nitric oxide synthesis: biological functionsNF-kB, it appears that AGEs are more potent inducers of and pathophysiology, Free Radic. Res. 6 (1999) 577–596.an intricately regulated immunorelevant gene such as [7] A.M. Cataldo, J.L. Barnett, C. Pieroni, R.A. Nixon, Increased
neuronal endocytosis and protease delivery to early endosomes iniNOS.sporadic Alzheimers disease — neuropathologic evidence for aOur results support the finding that an autodestructivemechanism of increased beta-amyloidogenesis, J. Neurosci. 17
process, involving overactive microglia and astroglia (1997) 6142–6151.producing high levels of free radicals occurs at the [8] D.W. Dickson, S. Sinicropi, S.H. Yen, L.W. Ko, L.A. Mattiace, R.characteristic lesions in AD [30]. Since AGEs accumulate Bucala, H. Vlassara, Glycation and microglial reaction in lesions of
Alzheimer’s disease, Neurobiol. Aging 17 (1996) 733–743.in astrocytes, microglia and on b-amyloid plaques in AD,[9] J.A. Edwards, F. Denis, P.J. Talbot, Activation of glial cells bythey may increasingly contribute to oxidative stress, evi-
human coronavirus OC43 infection, J. Neuroimmunol. 108 (2000)dent by the co-localization of AGE and iNOS in these glia 73–81.cells. Furthermore, co-localization of AGE and iNOS, but [10] G. Ehlerding, J. Schaeffer, W. Drommer, T. Miyata, K.M. Koch, J.not b-amyloid and iNOS in astroctyes suggests that AGEs, Floege, Alterations of synovial tissue and their potential role in the

A. Wong et al. / Brain Research 920 (2001) 32 –40 39
deposition of beta2-microglobulin-associated amyloid, Nephrol. the NINCDS-ADRDA work group under the auspices of DepartmentDial. Transplant. 13 (1998) 1465–1475. of Health and Human Services Task Force on Alzheimer’s disease,
Neurology 34 (1984) 934–944.[11] C.E. Finch, D.M. Cohen, Aging, metabolism, and Alzheimer disease— review and hypotheses, Exp. Neurol. 143 (1997) 82–102. [32] M. McMillian, L.Y. Kong, S.M. Sawin, B. Wilson, K. Das, P.
Hudson, J.S. Hong, G.Y. Bing, Selective killing of cholinergic[12] R.A. Floyd, Antioxidants, oxidative stress, and degenerative neuro-neurons by microglial activation in basal forebrain mixed neuronal /logical disorders, Proc. Soc. Exp. Biol. Med. 222 (1999) 236–245.glial cultures, Biochem. Biophys. Res. Commun. 215 (1995) 572–[13] F. Gallyas, Silver staining of Alzheimer’s neurofibrillary changes by577.means of physical development, Acta Morphol. Acad. Sci. Hung. 19
[33] L. Meda, M.A. Cassatella, G.I. Szendrei, L. Otvos Jr., P. Baron, M.(1971) 1–8.Villalba, D. Ferrari, F. Rossi, Activation of microglial cells by[14] S.G. Greenberg, P. Davies, J.D. Schein, L.I. Binder, Hydrofluoricbeta-amyloid protein and interferon-gamma, Nature 374 (1995)acid-treated tau PHF proteins display the same biochemical prop-647–650.erties as normal tau, J. Biol. Chem. 267 (1992) 564–569.
[34] T. Miyata, R. Inagi, Y. Iida, M. Sato M, N. Yamada, O. Oda, K.[15] K. Hensley, M.L. Maidt, Z.Q. Yu, H. Sang, W.R. Markesbery, R.A.Maeda, H. Seo, Involvement of beta 2-microglobulin modified withFloyd, Electrochemical analysis of protein nitrotyrosine andadvanced glycation end products in the pathogenesis ofdityrosine in the Alzheimer brain indicates region-specific accumu-hemodialysis-associated amyloidosis. Induction of human monocytelation, J. Neurosci. 18 (1998) 8126–8132.chemotaxis and macrophage secretion of tumor necrosis factor-alpha[16] Y. Iida, T. Miyata, R. Inagi, S. Sugiyama, K. Maeda, Beta(2)-and interleukin-1, J. Clin. Invest. 93 (2) (1994) 521–528.microglobulin modified with advanced glycation end products
[35] A.K. Mohamed, A. Bierhaus, S. Schiekofer, H. Tritschler, R.induces interleukin-6 from human macrophages — role in theZiegler, P.P. Nawroth, The role of oxidative stress and NF-kappaBpathogenesis of hemodialysis-associated amyloidosis, Biochem.activation in late diabetic complications, Biofactors 10 (1999) 157–Biophys. Res. Commun. 201 (1994) 1235–1241.167.[17] Z.S. Khatchaturian, Diagnosis of Alzheimer’s disease, Arch. Neurol.
[36] J.C. Monford, F. Javoy-Agid, J. Hauw, B. Dubois, Y. Agid, Brain42 (1985) 1097–1105.glutamate decarboxylase in Parkinson’s disease with particular[18] A. Klegeris, D.G. Walker, P.L. McGeer, Interaction of Alzheimerreference to premortem severity index, Brain 108 (1985) 301–313.beta-amyloid peptide with the human monocytic cell line THP-1
¨results in a protein kinase c-dependent secretion of tumor necrosis [37] G. Munch, R. Schinzel R, C. Loske, A. Wong, N. Durany, J.J. Li, H.factor-alpha, Brain Res. 747 (1997) 114–121. Vlassara, M.A. Smith, G. Perry, P. Riederer, Alzheimer’s disease —
synergistic effects of glucose deficit, oxidative stress and advanced[19] T. Komori, N. Shibata, M. Kobayashi, S. Sasaki, M. Iwata,glycation endproducts, J. Neural Transm. 105 (1998) 439–461.Inducible nitric oxide synthase (iNOS)-like immunoreactivity in
argyrophilic, tau-positive astrocytes in progressive supranuclear [38] S. Pou, W. Pou, D. Bredt, S. Snyder, G. Rosen, Generation ofpalsy, Acta Neuropathol. 95 (1998) 338–344. superoxide by purified brain nitric oxide synthase, J. Biol. Chem 267
(1992) 24173–24176.[20] U. Lamas, D. Perez-Sala, S. Moncada, Nitric oxide: from discoveryto the clinic, Trends Pharmacol. Sci. 19 (1998) 436–438. [39] K. Rostasy, L. Monti, C. Yiannoutsos, M. Kneissl, J. Bell, T.L.
Kemper, J.C. Hedreen, B.A. Navia, Human immunodeficiency virus[21] M. Leist, E. Fava, C. Montecucco, P. Nicotera, Peroxinitrite andinfection, inducible nitric oxide synthase expression, and microglialnitric oxide donors induce neuronal apoptosis by eliciting autocrineactivation: pathogenetic relationship to the acquired immuno-excitotoxicity, Eur. J. Neurosci. 9 (1997) 1488–1498.deficiency syndrome dementia complex, Ann. Neurol. 46 (1999)[22] J.J. Li, D. Dickson, P.R. Hof, H. Vlassara, Receptors for advanced207–216.glycosylation endproducts in human brain — role in brain homeo-
stasis, Mol. Med. 4 (1998) 46–60. [40] M.A. Smith, P.L.R. Harris, L.M. Sayre, J.S. Beckman, G. Perry,Widespread peroxynitrite-mediated damage in Alzheimer’s disease,[23] J.J. Li, D. Voisin, A.L. Quiquerez, C. Bouras, Differential expressionJ. Neurosci. 17 (1997) 2653–2657.of advanced glycosylation end-products in neurons of different
species, Brain Res. 641 (1994) 285–288. [41] M.A. Smith, L.M. Sayre, V.M. Monnier, G. Perry, Oxidativeposttranslational modifications in Alzheimer’s disease — a possible[24] S. Lipton, Y. Choi, Z. Pan, A red-based mechanism for thepathogenic role in the formation of senile plaques and neurofibrillaryneuroprotective and neurodestructive effects of nitric oxide andtangles, Mol. Chem. Neuropathol. 28 (1996) 41–48.related nitroso-compounds, Nature 364 (1993) 626–632.
[42] J. Stamler, Nitration and related target interactions of nitric oxide,[25] J. Liu, M.L. Zhao, C.F. Brosnan, S.C. Lee, Expression of type IICell 78 (1994) 931–936.nitric oxide synthase in primary human astrocytes and microglia:
role of IL-1beta and IL-1 receptor antagonist, J. Immunol. 157 [43] A. Takeda, T. Yasuda, T. Miyata, Y. Goto, M. Wakai, M. Watanabe,(1996) 3569–3576. Y. Yasuda, K. Horie, T. Inagaki, M. Doyu, K. Maeda, G. Sobue G,
[26] C. Loske, A. Neumann, A.M. Cunningham, K. Nichol, R. Schinzel, Advanced glycation end products co-localized with astrocytes and¨P. Riederer, G. Munch, Cytotoxicity of advanced glycation en- microglial cells in Alzheimer’s disease brain, Acta Neuropathol.
dproducts is mediated by oxidative stress, J. Neural Transm. 105 (Berl.) 95 (1998) 555–558.(1998) 1005–1015. [44] A. Takeda, T. Yasuda, T. Miyata, K. Mizuno, M. Li, S. Yoneyama,
[27] H.C. Ludwig, R. Ahkavan-Shigari, S. Rausch, K. Schallock, C. K. Horie, K. Maeda, G. Sobue, Immunohistochemical study ofQuentin, D. Ziegler,V. Bockermann, E. Markakis, Oedema extension advanced glycation end products in aging and Alzheimer’s diseasein cerebral metastasis and correlation with the expression of nitric brain, Neurosci Lett. 221 (1996) 17–20.oxide synthase isozymes (NOS I–III), Anticancer Res. 20 (2000) [45] P.J. Thornalley, Glutathione-dependent detoxification of alpha-ox-305–310. oaldehydes by the glyoxalase system — involvement in disease
[28] S. Love, Oxidative stress in brain ischemia, Brain Pathol. 9 (1999) mechanisms and antiproliferative activity of glyoxalase I inhibitors,119–131. Chem. Biol. Interact. 112 (1998) 137–151.
¨[29] H.J. Luth, M. Holzer, H.J. Gertz, Th. Arendt, Aberrant expression of [46] M.P.Vitek, K. Bhattacharya, J.M. Glendening, E. Stopa, H.Vlassara,nNOS in pyramidal neurons in Alzheimer’s disease is highly co- R. Bucala, K. Manogue, A. Cerami, Advanced glycation endlocalized with p21 ras and p16INK4a, Brain Res. 852 (2000) 45–55. products contribute to amyloidosis in Alzheimer disease, Proc. Natl.
[30] E.G. McGeer, P.L. McGeer, Aging, neurodegenerative disease and Acad. Sci. USA 91 (1994) 4766–4770.the brain, Can. J. Aging 16 (1997) 218–236. [47] Y. Vodovotz, M.S. Lucia, K.C. Flanders, L. Chesler, Q. Xie, T.W.
[31] G. McKhann, D. Drachmann, M. Folstein, R. Katzman, D. Price, Smith, F. Weidner, R. Mumford, R. Webber, C. Nathan, A.B.E.M. Stadlan, Clinical diagnosis of Alzheimer’s disease: report of Roberts, C.F. Lippa, M.B. Sporn, Inducible nitric oxide synthase in

40 A. Wong et al. / Brain Research 920 (2001) 32 –40
tangle-bearing neurons of patients with Alzheimer’s disease, J. Exp. amyloid induces microglial phagocytosis, expression of inducibleMed. 184 (1996) 1425–1433. nitric oxide synthase, and loss of a select population of neurons in
[48] M.N. Wallace, J.G. Geddes, D.A. Farquhar, M. R Masson, Nitric the rat CNS in vivo, J. Neurosci. 18 (1998) 2161–2173.oxide synthase in reactive astrocytes adjacent to b-amyloid plaques, [51] S.D. Yan, A.M. Schmidt, G.M. Anderson, J. Zhang, J. Brett, Y.S.Exp. Neurol. 144 (1997) 266–272. Zou, D. Pinsky, D. Stern, Enhanced cellular oxidant stress by the
[49] J. Wegiel, K.C. Wang, M. Tarnawski, B. Lach, Microglia cells are interaction of advanced glycation end products with their receptors /the driving force in fibrillar plaque formation, whereas astrocytes are binding proteins, J. Biol. Chem. 269 (1994) 9889–9897.a leading factor in plaque degradation, Acta Neuropathol. (Berl.) [52] M.L. Zhao, J.S. Liu, D. He, D.W. Dickson, S.C. Lee, Inducible nitric100 (2000) 356–364. oxide synthase expression is selectively induced in astrocytes
[50] D.T. Weldon, S.D. Rogers, J.R. Ghilardi, M.P. Finke, J.P. Cleary, E. isolated from adult human brain, Brain Res. 813 (1998) 402–405.O’Hare, W.P. Esler, J.E. Maggio, P. W Mantyh, Fibrillar beta-





























