Embed Size (px)
Citation preview

Comp. Biochem. Physiol. Vol. 92B, No. 1, pp. 175-180, 1989 0305-0491/89 $3.00 + 0.00 Printed in Great Britain © 1989 Pergamon Press pie
ADENYLATE METABOLIZING ENZYMES IN INVERTEBRATE TISSUES
A. LAZOU Laboratory of Animal Physiology, Department of Zoology, Science School, University of Thessaloniki,
Thessaloniki 54006, Greece
(Received 9 March 1988)
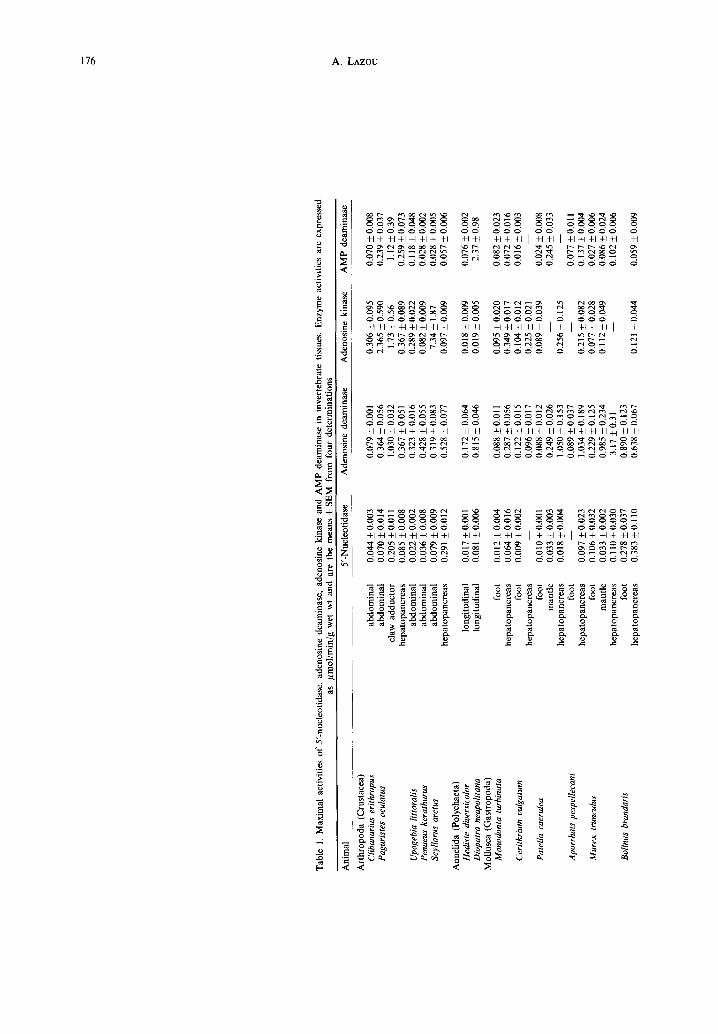
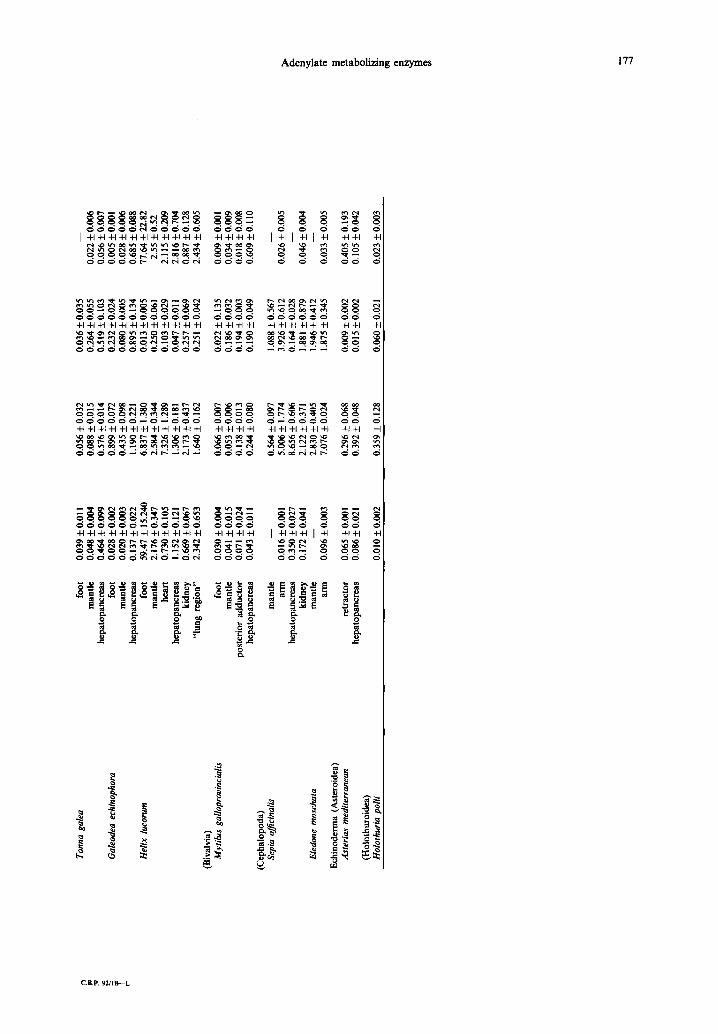
Abstract--1. Maximal activities of 5'-nucleotidase, adenosine deaminase, adenosine kinase and AMP deaminase were measured in various tissues from invertebrates.
2. In general, enzyme activities were very low as compared to vertebrates. 3. The highest activities of AMP deaminase were found in anaerobic muscles. 5'-nucleotidase, adenosine
deaminase and adenosine kinase activities were higher in more aerobic tissues. 4. Cephalopod tissues showed very active adenosine deaminase and adenosine kinase with a lower level
of AMP deaminase activity. 5. The activities of all enzymes measured in the tissues of the snail, Helix lucorum, were considerably
higher as compared to the other animals investigated. 6. On the basis of the enzyme levels, different pathways of adenylate degradation and the role of
adenosine in invertebrate tissues are discussed.
INTRODUCTION
Relatively rapid changes can occur in the total ade- nine nucleotide pool size as a result of perturbations of energy metabolism. Regulation of the adenylate pool size can occur by degradation of AMP which has the effect of maintaining the ATP/AMP ratio at relatively constant levels. Degradation of AMP fol- lows two different pathways, both regulated by ATP. The first involves deamination of AMP followed by IMP dephosphorylation (the "direct" pathway). The second involves dephosphorylation of AMP followed by adenosine deamination (the "indirect" pathway).
The first pathway is part of the so-called "purine nucleotide cycle", the metabolic role of which is supposed to be important in amino acid degradation, energy charge stabilization and ammonia produc- tion, especially in kidney and liver (Lowenstein and Tornheim, 1971; Lowenstein, 1972; Chapman and Atkinson, 1973; Bogusky et al., 1976). The second pathway involves the formation of adenosine, which plays a more important role than simply being an intermediate in the degradation pathway of adenine nucleotides. Adenosine is considered to be a local hormone and changes in its concentration appear to affect metabolism in general and to regulate a number of important physiological processes in vertebrates including the rate of blood flow in the heart, skeletal muscle, liver, kidney and brain (Berne, 1963; Scott et al., 1965; Berne et al., 1974; Bockman et aL, 1975; Arch and Newsholme, 1978a). The role of adenosine in invertebrate tissues though is not yet clear. Apart from a few reports on the activities of adenosine metabolizing enzymes, data on the physiological role of adenosine in invertebrate tissues is lacking.
Invertebrates are found in very different environ- ments which have necessitated the development of muscles with different physiological functions. Vari- ations of purine nucleotide degradation appear to
exist among muscles from various animals depending on their physiological function. In view of the im- portant role that these nucleotides play in muscular contraction, their degradation pathway, as well as comparison with other tissues, would be of interest.
It was considered that the levels of enzymes in- volved in AMP metabolism would provide useful information about the AMP degrading pathways in this group of animals. Consequently, in this paper we measured and compared the activities of 5'-nucleotidase (EC 3.1.3.5), AMP deaminase (EC 3.5.4.6), adenosine deaminase (EC 3.5.4.4) and ade- nosine kinase (EC 2.7.1.20) in tissues of different invertebrate species.
MATERIALS AND METHODS
Chemicals
Enzymes and substrates were obtained from Sigma Chemical Co. (St Louis, Missouri). All other chemicals were obtained from Serva (Heidelberg, FRG).
Sources of animals
Bivalves, gastropods and echinoderms were collected locally and kept in recirculating sea-water at 17-18°C. Crustaceans, cephalopods and polychaetes were purchased fresh and frozen in liquid nitrogen until analysis. Snails, Helix lucorum, were a gift from Dr M. Lazaridou and were kept in glass containers.
Preparation of homogenates
All operations were conducted at 0-4°C. Tissues were removed from animals as soon as possible after death. The tissues were homogenized in ground glass homogenizers with 5-10 volumes of extraction medium which consisted of 20 mM potassium phosphate buffer, pH 7.1, and 0.I % 2-mercaptoethanol. For the determination of adenosine deaminase, adenosine kinase and AMP deaminase, the homogenates were centrifuged at 20,000 g for 5 min. For the determination of 5'-nucleotidase, the homogenates were filtered through gauze before use.
175
o~
Tab
le 1
. M
axim
al a
ctiv
itie
s o
f 5'
-nuc
leot
idas
e, a
deno
sine
dea
min
ase,
ade
nosi
ne k
inas
e an
d A
MP
dea
min
ase
in i
nver
tebr
ate
tissu
es.
Enz
yme
acti
viti
es a
re e
xpre
ssed
th
e m
ean
s ±
SE
M f
rom
fou
r de
term
inat
ions
Ani
mal
5'
-Nuc
leot
idas
e A
deno
sine
dea
min
ase
Ade
nosi
ne k
inas
e A
MP
dea
min
ase
as
/tm
ol/
min
/g w
et w
t an
d a
re
Art
hro
po
da
(Cru
stac
ea)
Clib
anar
ius
erith
ropu
s ab
do
min
al
0.04
4 +
0.0
03
0.07
9 +
0.0
01
0.30
6 +
0.0
95
0.07
0 +
0.0
08
Pag
uris
tes
ocul
atus
ab
do
min
al
0.07
0 ±
0.0
14
0.36
4 ±
0.0
56
2.36
5 +
0.5
90
0.23
9 ±
0.0
37
claw
ad
du
cto
r 0.
205
± 0
.011
1.
030
± 0
.032
1.
73 ±
0.5
6 1.
12 ±
0.3
9 h
epat
op
ancr
eas
0.08
5 ±
0.0
08
0.36
7 +
0.0
51
0.36
7 ±
0.0
89
0.25
9 +
0.0
73
Upo
gebi
a lit
tora
lis
abd
om
inal
0.
022
_+ 0
.002
0.
323
+ 0
.016
0.
289
± 0
.022
0.
118
+ 0
.048
P
enae
us k
erat
huru
s ab
do
min
al
0.03
6 ±
0.0
08
0.42
8 +
0.0
55
0.08
2 ±
0.0
09
0.02
8 +
0.0
02
Scyl
laru
s ar
ctus
ab
do
min
al
0.07
9 ±
0.0
09
0.31
9 +
0.0
83
7.34
± 1
.87
0.02
8 +
0.0
05
hep
ato
pan
crea
s 0.
291
± 0
.012
0.
528
± 0
.077
0.
097
± 0
.009
0.
057
± 0
.006
A
nnel
ida
(Pol
ycha
eta)
H
edis
te d
iver
s±co
lor
long
itud
inal
0.
017
± 0
.001
0.
172
± 0
.064
0.
018
± 0
.009
0.
076
± 0
.002
D
iopa
tra
neap
olit
ana
long
itud
inal
0.
081
± 0
.006
0.
815
± 0
.046
0.
019
± 0
.005
2.
37 ±
0.9
8 M
ollu
sca
(Gas
tro
po
da)
M
onod
onta
tur
bina
ta
foot
0.
012
± 0
.004
0.
088
± 0
.011
0.
095
± 0
.020
0.
082
± 0
.023
h
epat
op
ancr
eas
0.06
4 +
0.0
16
0.28
7 ±
0.0
56
0.34
9 ±
0.0
17
0.07
2 ±
0.0
16
Cer
ithri
um
vulg
atum
fo
ot
0.00
9 ±
0.0
02
0.12
2 ±
0.0
15
0.10
4 ±
0.0
12
0.01
6 ±
0.0
03
hep
ato
pan
crea
s --
0.
096
± 0
.017
0.
225
± 0
.021
--
P
atel
la c
aeru
lea
foot
0.
010
+ 0
.001
0.
088
± 0
.012
0.
089
± 0
.039
0.
024
± 0
.008
m
antl
e 0.
033
± 0
.003
0.
249
± 0
.026
--
0.
245
± 0
.033
h
epat
op
ancr
eas
0.01
8 ±
0.0
04
1.05
0 ±
0.1
53
0.25
6 +
0.1
25
--
Apo
rrha
is
pesp
elle
cani
fo
ot
--
0.08
9 ±
0.0
37
--
0.07
7 ±
0.0
11
hep
ato
pan
crea
s 0.
097
± 0
.023
1.
034
± 0
.189
0.
215
± 0
.082
0.
137
± 0
.004
M
urex
tru
ncul
us
foot
0.
106
± 0
.032
0.
229
± 0
.125
0.
077
+ 0
.028
0.
027
+_ 0
.006
m
antl
e 0.
033
+ 0
.002
0.
985
± 0
.234
0.
112
± 0
.049
0.
086
± 0
.024
h
epat
op
ancr
eas
0.11
0 ±
0.0
30
3.17
+ 0
.31
--
0.10
2 ±
0.0
06
Bol
inus
bra
ndar
is
foot
0.
278
± 0
.037
0.
890
± 0
.123
--
--
h
epat
op
ancr
eas
0.38
3 ±
0.1
10
0.63
8 ±
0.0
67
0.12
1 ±
0.0
44
0.05
9 ±
0.0
09
;>
t"
s ©
Tonn
a ga
lea
Gal
eode
a ec
hino
phor
a
Hel
ix l
ucor
um
(Biv
alvi
a)
Myt
ilus
gal
lopr
ovin
cial
is
(Cep
halo
poda
) Se
pia
offic
inal
is
Ele
done
mos
chat
a
Ech
inod
erm
a (A
ster
oide
a)
Ast
eria
s m
edite
rran
ean
(Hol
othu
roid
ea)
Hol
othu
ria
polii
foot
m
antl
e he
pato
panc
reas
fo
ot
man
tle
hepa
topa
ncre
as
foot
m
antl
e he
art
hepa
topa
ncre
as
kidn
ey
"lun
g re
gion
"
foot
m
antl
e po
ster
ior
addu
ctor
he
pato
panc
reas
man
tle
ann
he
pato
panc
reas
ki
dney
m
antl
e ar
la
retr
acto
r he
pato
panc
reas
0.03
9 ±
0.01
1 0.
048
+ 0.
004
0.46
4 ±
0.09
9 0.
028
± 0.
002
0.02
0 +
0.00
3 O
. 137
_+ 0
.022
59
.47
+_ 1
5.24
0 2
.17
6±
0.3
47
0.
730
+ 0.
105
1.15
2 +
0.12
1 0.
669
4- 0
.067
2.
342
± 0.
653
0.03
0 +
0.00
4 0.
041
± 0.
015
0.07
1 ±
0.02
4 0.
043
± 0.
011
0.01
6 ±
0.00
1 0.
350
± 0.
027
O. 1
72 ±
0.0
41
0.09
6 ±
0.00
3
0.06
5 _+
0.0
01
0.08
6 ±
0.02
i
0.01
0 ±
0.00
2
0.05
6 +
0.03
2 0.
088
+ 0.
015
0.57
6 ±
0.01
4 0.
899
± 0.
072
0.43
5 +
0.09
8 1.
190
+ 0.
221
6.83
7 ±
i.38
0 2.
584
± 0.
344
7.32
6 +
1.28
9 1.
306
± 0.
181
2.17
3 ±
0.43
7 1.
640_
+0.
162
0.06
6 +
0.00
7 0.
053
± 0.
006
O. 1
38 +
0.0
13
0.24
4 +
0.08
0
0.56
4 __. 0.0
97
5.00
6 ±
1.77
4 8.
656
_+ 0
.606
2.
122
+ 0.
371
2.83
0 ±
0.40
5 7.
076
_ 0.
024
0.29
6 _+
0.0
68
0.39
2 _+
0.0
48
0.35
9 +
0.12
8
0.03
6 _+
0.0
35
0.26
4 +
0.05
5 0.
519
± 0.
103
0.23
2 _+
0:0
24
0.08
0 __
0.0
05
0.89
5 +
0.13
4 0.
013
+_ 0
.005
0.
250
_+ 0
.061
O
. 103
± 0
.029
0.
047
+ 0.
011
0.25
7 ±
0.06
9 0.
251
± 0.
042
0.02
2 _+
O. 1
35
0.18
6 ±
0.03
2 0.
194
±0.
003
0.19
0 +
0.04
9
1.08
8 +
0.56
7 3.
926
+ 0.
612
0.16
4_+
0.02
8 1.
881
+ 0.
879
1.94
6 +
0.41
2 1.
875
__. 0
.345
0.00
9 +
0.00
2 0.
015 _+ 0
.002
0.06
0 ___ 0.0
21
0.02
2 +
0.00
6 0.
056
-t- 0
.007
0.
005
_+ 0
.001
0.
028
_+ 0
.006
0.
685
+ 0.
088
77.6
4 +
22.8
2 2.
55 +
0.
52
2.11
5 __. 0.2
09
2.81
6 __. 0.7
04
0.88
7 +_ O. 12
8 2.
434
_+ 0.6
05
0.00
9 _+
0.0
01
0.03
4 +
0.00
9 0.
018
+ 0.
008
0.6
09
+0
.11
0
0.02
6 +
0.00
5
0.04
6 _+
0.0
04
0.03
3 +
0.00
5
0.40
5 +
0.19
3 O
. 105
+ 0
.042
0.02
3 +
0.00
3
> o o"
o

178 A. LAZOU
Enzyme assays 5'-Nucleotidase, adenosine deaminase and AMP de-
aminase activities were measured by recording the decrease of optical density at 265 nm (Kalckar, 1947). The reaction mixture for 5'-nucleotidase contained 50 mM TRA, pH 7.0, 0.2 mM AMP, 18 mM MgCI, 180 mM KC1 and 0.5 units/ml adenosine deaminase. The reaction mixture for adenosine deaminase contained 50 mM potassium phosphate, pH 7.1, and 1 mM adenosine. The reaction mixture for AMP de- aminase contained 50 mM imidazole, pH 6.5, I00 mM KCI and 0.07 mM AMP. Adenosine kinase was assayed by following the oxidation of NADH at 340 nm. The reaction mixture contained 50mM TRA, pH7.0, 0.5mM MgC1, lmM ATP, 0.25mM PEP, 0.1mM adenosine, 0.1% NADH and excess amounts of lactate dehydrogenase, pyru- vate kinase and adenylate kinase.
In general, catch muscles showed low activities of AMP deaminase and 5'-nucleotidase. Cephalopod tissues showed markedly active adenosine deaminase and adenosine kinase with a lower level of AMP deaminase activity. The activities of all enzymes measured in the snail Helix lucorum tissues were considerably higher compared to the other animals investigated. Helix lucorum foot muscle exhibited a considerably active AMP deaminase. Furthermore, comparing the adenosine kinase activity in the tissues of the snail, the lowest value was found in the foot muscle whereas the highest values were found in more aerobic tissues such as the mantle, the kidney and the "lung" region.
RESULTS
The maximal activities of 5'-nucleotidase, ade- nosine deaminase, adenosine kinase and AMP de- aminase are given in Table 1. With few exceptions all the enzymes measured showed very low activity. The mean values of Vma x for 5'-nucleotidase in muscles ranged from 0.010 to 2.176 p mol/min/g fresh wt (Holothuria polii and Helix lucorum mantle respectively), with the exception of Helix lucorum foot muscle which showed a considerably higher activity of 5'-nucleotidase, 59#mol/min/g. The maximal activities of 5'-nucleotidase measured in hepatopancreas or kidney were in the same range (0.018-1.152 #mol/min/g fresh wt) but for each ani- mal studied the activity was higher in hepatopancreas than in muscles. The highest activity was observed in the snail Helix lucorum.
The mean values of Vm~ x for adenosine deaminase in muscles ranged from 0.053 to 7.326 #mol/min/g fresh wt (Mytilus mantle and Helix heart respectively) whereas in hcpatopancreas and kidney the range of activities was a little higher, 0.287-8.656 #mol/min/g (Monodonta turbinata and Sepia o~icinalis respect- ively). In addition, for animals of the same species, adenosine deaminase activity was considerably higher in hepatopancreas than in muscles.
The mean values of Vma x for adenosine kinase in muscles ranged from 0.013 to 1.946#mol/min/g (Helix lucorum heart and Eledone moschata mantle), with the highest values found in aerobic muscles such as cephalopod mantle. Exceptionally, very high adenosine kinase activity was found in the ab- dominal muscle of two crustaceans, Scyllarus arctus (7.34/~mol/min/g) and Paguristes oculatus (2.36 #tool/rain/g). For animals of the same species, adenosine kinase activities were higher in hepatopancreas and kidney than in muscles.
AMP deaminase maximal activities in muscles ranged from 0.005 to 77.64/tmol/min/g (Galeodea foot and Helix foot respectively). In hepatopancreas, AMP deaminase activity ranged from 0.056 to 2.816/~mol/min/g (Tonna galea and Helix lucorum respectively). AMP deaminase was more active than 5'-nucleotidase in muscles but still its activity was very low compared to those measured in vertebrate muscles (Fijisawa and Yoshino, 1987). The highest activity of AMP deaminase was found in anaer- obic muscles such as crustacean abdominal and polychaetes body wall muscles.
DISCUSSION
The low activities of 5'-nucleotidase, adenosine deaminase, adenosine kinase and AMP deaminase measured in the present study (Table 1) suggest that adenylates degrade at a very low rate in most in- vertebrate tissues studied. Comparing the maximal activities of the enzymes, it appears that both AMP degrading pathways, "direct" and "indirect" are operative. The highly active adenosine deaminase which is present in cephalopods indicates that the AMP-adenosine-ionosine pathway is operative. The very high activity of AMP deaminase found in the tissues of the snail suggests that AMP degradation occurs threugh the direct pathway. Even though AMP deaminase activity found in muscles was higher compared to that of 5'-nucleotidase, the presence of higher activities of adenosine deaminase and ade- nosine kinase suggest that a preferential de- phosphorylation of AMP to adenosine, which is further deaminated to inosine, might proceed in these tissues, though the deamination of AMP cannot be excluded. In a survey of the pathways of AMP degradation in rat tissues in the presence and absence of added ATP, Burger and Lowenstein (1967) found that in the absence of ATP the pathway involving AMP dephosphorylation is more active than the one involving deamination of AMP, whereas addition of ATP reversed this pattern. So if this is also the case in invertebrate tissues, then AMP degradation may depend on ATP tissue concentration and hence the metabolic state of the tissue.
The information on the activity of AMP deaminase in invertebrates is controversial. Early studies report the absence of AMP deaminase activity in crab, crayfish and the adductor muscle of pecten (Kitagawa and Tonomura, 1957; Umiastowski, 1964) while later on, low activity was described in lobster muscles (Dingle et al., 1968). From the results of the present study it can be seen that low AMP deaminase activity is found in all invertebrate tissues studied, with a few exceptions where AMP deaminase is very active (Table 1).
As has been suggested, AMP deaminase is re- sponsible for the stimulation of glycolysis (Yoshino and Murakami, 1982, 1985) not only by affecting the AMP and ATP concentrations but by producing ammonia, an activator of phosphofructokinase as well (McComb and Yoshok, 1964). AMP deaminase activity is closely correlated with the activity of phosphofructokinase as a key enzyme of glycolysis in

Adenylate metabolizing enzymes 179
mammalian muscles (Winder et aL, 1974). The pres- ence of higher AMP deaminase activities in anaerobic muscles that depend on glycolysis for energy prod- uction such as the abdominal muscle of crustaceans and the longitudinal muscle of polychaetes (Table 1), is consistent with the above suggestion. The high activity found in Helix lucorum foot muscle can be compared with the very active AMP deaminase found in Helix pomatia foot muscle (Stankiewitz, 1982). In this muscle, as well as in other tissues of the snail where the enzyme activity is high, the main function of this enzyme may be the production of free ammo- nia. It is noteworthy that the activities of all enzymes measured were considerably higher in Helix lucorum tissues as compared to the rest of the invertebrates that were investigated. Helix lucorum was the only terrestrial invertebrate from the group that was stud- ied, so the great difference in the maximal activities of the enzymes may reflect the different environments that these animals are found in.
The results of the activities of adenosine metabo- lizing enzymes show that adenosine is metabolized by all the invertebrate tissues studied, though the phys- iological role of adenosine in invertebrate tissues remains to be further defined. It has been suggested (Arch and Newsholme, 1978b) that adenosine kinase is involved in the control of tissue adenosine concen- trations and so its activity is an indication of the measure of control which a tissue has over its concen- tration of adenosine. In support of the theory that adenosine regulates blood flow, it has been demon- strated that aerobic muscles have higher activities of adenosine kinase than anaerobic muscles, so that they can modulate their concentration of adenosine more rapidly. Furthermore, there is an inverse correlation between the activities of glycogen phosphorylase (which is an indication of the tissues' dependence on anaerobic rather than on aerobic metabolism for energy production) and adenosine kinase (Crabtree and Newsholme, 1972; Arch and Newsholme, 1978b). From the results of the present study it appears that very high adenosine kinase activities are present in aerobic muscles such as the mantle of cephalopods, while the activity measured in the mantle of gastro- pods is higher than the one measured in the foot muscle (Table 1). In general, the mantle is a highly aerobic muscle whereas foot muscles have low rates of energy expenditure (Hochachka, 1976). The very high adenosine kinase activities found in the abdom- inal muscles of some crustaceans which are anaerobic (Zammit and Newsholme, 1976) remain to be ex- plained. Nevertheless, this high activity indicates that in these muscles adenosine plays a regulatory role which is rather obscure at present. Adenosine may play a very different regulatory role in invertebrate muscles to that which it plays in vertebrates. The very high adenosine deaminase activities in hepa- topancreas suggest that in this tissue the enzyme functions to deaminate adenosine and together with adenosine kinase may play a role in the removal of adenosine.
Acknowledgements--The author wishes to thank Dr A. Koukouras and Dr E. Voultsiadou-Koukoura for identify- ing the animals.
REFERENCES
Arch J. R. S. and Newsholme E. A. (1978a) The control of the metabolism and the hormonal role of adenosine. In Essays in Biochemistry (Edited by Campbell P. N. and Aldridge W. N.), Vol. 14, pp. 82-123. Academic Press, New York.
Arch J. R. S. and Newsholme E. A. (1978b) Activities and some properties of 5'-nucleotidase, adenosine kinase and adenosine deaminasc in tissues from vertebrates and invertebrates in relation to the control of the concen- tration and the physiological role of adenosine. Biochem. J. 174, 965-977.
Berne R. M. (1963) Cardiac nucleotides in hypoxia: possible role of adenosine in the regulation of blood flow. Am. J. Physiol. 204, 317-322.
Berne R. M., Rubio R. and Curnish R. R. (1974) Release of adenosine from ischaemic brain. Effect of cerebral adenine nucleotides. Circ. Res. 35, 262-271.
Bockman E. L., Berne R. M. and Rubio R. 0975) Release of adenosine and lack of release of ATP from contracting skeletal muscle. Pflfigers Arch. ges. Physiol. 355, 229-241.
Bogusky R. T., Lowenstein L. M. and Lowenstein J. M. (1976) The purine nucleotide cycle. A pathway for ammo- nia production in the rat kidney. J. clin. Invest. 58, 326-335.
Burger R. and Lowenstein J. M. (1967) Adenylate Deaminase--III. Regulation of deamination pathways in extracts of rat heart and lung. J. biol. Chem. 242, 5281-5288.
Chapman A. G. and Atkinson D. E. (1973) Stabilization of adenylate energy charge by the adenylate deaminase reaction J. bioL Chem. 248, 8309-8312.
Crabtree B. and Newsholme E. A. (1972) The activities of hexokinase, phosphorylase, phosphofructokinase, lactate dehydrogenase and the glycerol-3-phosphate dehy- drogenase in muscles from vertebrates and invertebrates. Biochem. J. 126, 49-58.
Dingle J. R., Hines J. A. and Fraser D. J. (1968) Post mortem degradation of adenine nucleotides in muscles of lobster Homarus americanus. J. Fd Sci. 33, 100-103.
Fijisawa K. and Yoshino M. (1987) Activities of adenylate degrading enzymes in muscles from vertebrates and in- vertebrates. Comp. Biochem. Physiol. 86B, 109-112.
Hochachka P. W. (1976) Design of metabolic and enzymic machinery to fit lifestyle and environment. Biochem. Soc. Symp. 41, 3-31.
Kalckar H. M. (1947) Differential spectrophotometry of purine compounds by means of specific enzymes--II. Determination of adenine compounds. J. biol. Chem. 167, 445-459.
Kitagawa S. and Tonomura Y. (1957) Contractile proteins from the adductor of pecten--III. Failure to detect adenylic acid deaminase activity. J. Biochem. Tokyo 44, 317-320.
Lowenstein J. M. (1972) The ammonia production in muscle and other tissues. The purine nucleotide cycle. Physiol. Rev. 52, 382-414.
Lowenstein J. M. and Tornheim K. (1971) Ammonia prod- uction in muscle. The purine nucleotide cycle. Science, N.Y. 171, 397-400.
McComb R. B. and Yoshok W. D. (1964) Metabolism of ascites tumor cells--IV. Enzymatic reactions involved in adenosine triphosphate degradation induced by 2-deoxy- glucose. Cancer Res. 24, 198-203.
Scott J. B., Daugherty R. M., Dabney J. M. and Haddy I. J. (1965) Role of chemical factors in regulation of blood flow through kidney, hindlimb and heart. Am. J. Physiol. 208, 813-824.
Stankiewicz A. (1982) Comparative studies on AMP- deaminase--VII. Purification and some properties of the enzyme from crayfish Orconectes limosus tail muscle. Comp. Biochem. Physiol. 72B, 127-132.

180 A. LAZOU
Umiastowski J. (1964) AMP-aminohydrolase in the muscles of some vertebrates and invertebrates. Acta Biochim. pol. XI, 459-464.
Winder W. W., Terjung R. L., Baldwin K. M. and Holloszy J. O. (1974) Effect of exercise on AMP deaminase and adenylosuccinate in rat skeletal muscle. Am. J. Physiol. 227, 1411-1414.
Yoshino M. and Murakami K. (1982) AMP deaminase reaction as a control system of glycolysis in yeast. Activa- tion of phosphofructokinase and pyruvate kinase by the AMP deaminase-ammonia system. J. biol. Chem. 257, 2822-2828.
Yoshino M. and Murakami K. (1985) AMP deaminase
reaction as a control system of glycolysis in yeast. Role of ammonium ion in the interaction of phosphofructo- kinase and pyruvate kinase activity with the adenylate energy charge. J. biol. Chem. 260, 4729-4732.
Zammit V. A. and Newsholme E. A. (1976) The maximum activities of hexokinase, phosphorylase, phosphofructo- kinase, glycerol phosphate dehydrogenase, octopine dehydrogenase, phosphoenolpyruvate carboxykinase, nu- cleoside diphosphate kinase, glutamate-oxaloacetate transaminase and arginine kinase in relation to carbo- hydrate utilization in muscles from marine invertebrates. Biochem. J. 160, 447-462.