Embed Size (px)
Citation preview

296 FRANCO ET AL.
© 2001 Wiley-Liss, Inc.
DRUG DEVELOPMENT RESEARCH 52:296–302 (2001) DD
RResearcher Overview
Adenosine/Dopamine Receptor-Receptor Interactionsin the Central Nervous System
Rafael Franco,1* Sergi Ferré,2 Maria Torvinen,2 Silvia Ginés,1 Joëlle Hillion,2
Francisco Ciruela,1 Enric I. Canela,1 Josefa Mallol,1 Vicent Casadó,1 Carmen Lluis,1
and Kjell Fuxe2
1Department of Biochemistry and Molecular Biology of the University of Barcelona, Barcelona, Spain2Department of Neuroscience, Division of Cellular and Molecular Neurochemistry, Karolinska Institutet,
Stockholm, Sweden
ABSTRACT There is evidence that adenosine/dopamine antagonism in the central nervous system mayinvolve an adenosine/dopamine receptor-receptor interaction. There also is an anatomical basis for theexistence of functional interactions between adenosine A1 receptors (A1Rs) and dopamine D1 receptors(D1Rs) and between adenosine A2A (A2ARs) and dopamine D2 (D2Rs) receptors in the same neurons. Selec-tive A1R agonists negatively affect the high-affinity binding of D1Rs. Activation of A2ARs leads to a decreasein receptor affinity for dopamine agonists acting on D2Rs, especially with regard to the high-affinity state. Allthese findings suggest receptor subtype–specific interactions between adenosine and dopamine receptorsthat may be achieved by molecular interactions, for example, receptor heterodimers. There exists evidencethat A1Rs form homodimers in membranes and also form high-order molecular structures containingheterotrimeric G proteins and adenosine deaminase. In view of the occurrence of homodimers of adenos-ine and of dopamine receptors, it was speculated that heteromers between these receptors, belonging totwo different families of G protein–coupled receptors, can be formed. Evidence that A1/D1 receptors canform heterodimers in co-transfected cells and in primary cultures of neurons, in fact, has been obtained.These intramembrane interactions may constitute a major molecular basis to explain the behavioral effectsof adenosine-related drugs in health and disease. There is also evidence that intramembrane dopamine/adenosine receptor antagonism not only is mediated by direct receptor/receptor interaction (heteromers)but also involves other interacting proteins. In fact, adenosine deaminase absolutely is required for adenos-ine/dopamine antagonism. In the central nervous system, functional modules in which receptors for neu-rotransmitters or neuromodulators interact with each other and with other interacting proteins are crucialto understanding the physiology of the neuron and higher neural functions, such as learning and memory.Refined knowledge of the molecular mechanism of these interactions would be beneficial, to improve ourtherapeutic arsenal against neurological and neuropsychiatric diseases. Drug Dev. Res. 52:296–302,2001. © 2001 Wiley-Liss, Inc.
Key words: G protein–coupled receptors; heteromerization; neurological diseases; signal transduction; adenosinedeaminase
*Correspondence to: Rafael Franco, Department of Biochem-istry and Molecular Biology of the University of Barcelona, MartiFranques 1, Barcelona 08028, Spain.
Strategy, Management and Health Policy
Venture CapitalEnablingTechnology
PreclinicalResearch
Preclinical DevelopmentToxicology, FormulationDrug Delivery,Pharmacokinetics
Clinical DevelopmentPhases I-IIIRegulatory, Quality,Manufacturing
PostmarketingPhase IV
DOPAMINE/ADENOSINE ANTAGONISM
Adenosine is an endogenous nucleoside acting asa neuromodulator in the central nervous system. Its ac-tions are mediated by adenosine receptors, four of whichhave been cloned and pharmacologically characterized:

ADENOSINE/DOPAMINE RECEPTOR-RECEPTOR HETEROMERIZATION 297
A1, A2A, A2B, and A3 [Fredholm et al., 1994]. Among thesefour subtypes, A1 (A1R) and A2A (A2AR) receptors are themain target of the behavioral effects in animals treatedwith adenosine agonists and antagonists [Ferré et al.,1992, 1993, 1997; Fredholm, 1995]. Caffeine, as an ex-ample of an antagonist acting at adenosine receptors, istoday the most consumed psychostimulant drug in theworld.
A1Rs and A2ARs localized in the basal ganglia, moreprecisely, in the striatum, are responsible for the motor-depressant effects of adenosine agonists and for the mo-tor-stimulatory effects of adenosine antagonists [Ferréet al., 1992, 1997]. Large amounts of striatal adenosinereceptors are located in the medium-sized spinyGABAergic neurons, the efferent neurons that consti-tute more than 90% of the neuronal population in thestriatum [Schiffmann et al., 1991; Rivkees et al., 1995].One subtype of GABA efferent neurons, the striopallidalones, contains mainly D2 dopamine receptors (D2Rs). Asecond subtype, the strionigro-strioentopeduncularGABA neurons, contains mainly D1 dopamine receptors(D1Rs) [Ferré et al., 1997]. The two subtypes of neuronsboth contain A1 adenosine receptors, but only thestriopallidal neurons contain A2ARs. These anatomicallocalizations indicate that in the basal ganglia A2ARs ex-ist only in the D2R-containing neurons, whereas A1Rsexist in both D1R- and D2R-containing nerve cells. Thelocalization of dopamine and adenosine receptor sub-types has provided the anatomical basis for the exist-ence of functional interactions between A1 and D1 orbetween A2A and D2 receptors in the strionigro-strio-entopeduncular GABA neurons and the striopallidalGABA neurons, respectively. These functional interac-tions have been investigated using a variety of tech-niques, from ligand binding and second messengerdeterminations to behavioral studies.
At the behavioral level, adenosine receptor antago-nist–induced motor activation is counteracted largely bytreatments that cause acute dopamine depletion or block-ade of D1Rs or D2Rs [Ferré et al., 1992, 1997]. Further-more, adenosine receptor agonists inhibit and adenosinereceptor antagonists potentiate the motor-activating ef-fects of dopamine agonists [Ferré et al., 1992, 1997]. Spe-cifically, low doses of A1R and A2AR agonists selectivelycounteract the motor-activating effects induced by D1Rand D2R agonists, respectively. On the other hand, A1Rantagonists preferentially potentiate D1R agonist–in-duced motor activation, whereas A2AR antagonists pref-erentially potentiate D2R agonist–mediated motor effects[Ferré et al., 1997; Kull et al., 1999; Salim et al., 2000].On the other hand, there is still controversy concerningthe exact role of A2AR/D2R interactions in the neurophysi-ological defects found in D2 knockout mice [Aoyama etal., 2000; Zahniser et al., 2000].
ADENOSINE/DOPAMINE RECEPTOR-RECEPTORINTERACTIONS
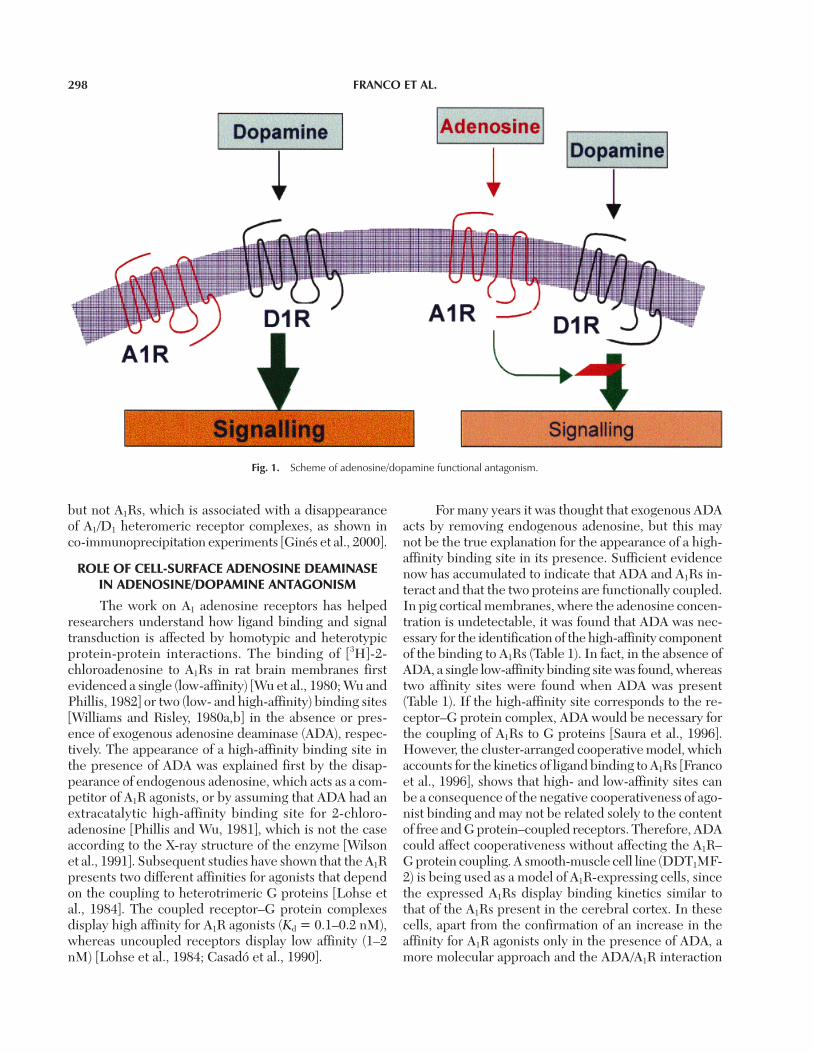
Some of the adenosine/dopamine antagonistic ef-fects can be explained by cross-talk at the level of, forinstance, adenylate cyclase (Fig. 1). A1R/D1R antagonismat the level of cAMP formation can be explained easilyby the fact that G proteins for A1Rs and D1Rs are coupleddifferently to adenylate cyclase. However, the modula-tion by adenosine agonists of the binding of dopamine todopamine receptors suggests that a direct receptor-re-ceptor interaction can indeed occur. A1R agonists nega-tively affect the high-affinity binding of dopamine to D1Rsby reducing the proportion of D1Rs in the high-affinitystate [Ferré et al., 1998]. These results have been obtainedin binding experiments with membranes from tissues orfrom cells co-transfected with the human A1R and D1RcDNAs. The adenosine A1 agonists shift high-affinitybinding to the low-affinity states of the D1R. On the otherhand, there is also an interaction at the adenylate cyclaselevel. A1Rs are coupled negatively, whereas D1Rs arecoupled positively to adenylate cyclase. Thus, the pres-ence of adenosine agonists leads to a decrease in dopam-ine D1-induced cAMP production, whereas, in turn, thepresence of antagonists acting on A1Rs leads to the po-tentiation of the cAMP responses occurring through ac-tivation of D1Rs [Ferré et al., 1998].
Owing to the specific antagonistic interaction be-tween adenosine and dopamine receptor subtypes [Ferréet al., 1991; Ferré et al., 1998] and the occurrence ofhomomers of adenosine [Ciruela et al., 1995] and ofdopamine [Ng et al., 1996; Zawarynski et al., 1998] recep-tors, heteromers between A1 and D1 receptors can beformed. In this article we focus on A1R/D1R molecularinteractions, but research is under way in our laborato-ries to prove a similar interaction for A2AR/D2R.
Although the hypothesis of heterodimerization hadbeen formulated [Agnati et al., 1982, 1983; Fuxe et al.,1983; Fuxe and Agnati, 1987; Zoli et al., 1993], it has notbeen possible until recently to study the possible di- orheteromerization of dopamine and adenosine receptors,owing to the lack of specific antibodies working in im-munoprecipitation, immunoblotting, and immunofluores-cence studies. Sufficient evidence now exists to indicatethat A1 and D1 form heteromers in both co-transfectedcells and primary cultures of neurons. A high level of co-localization was seen between A1Rs and D1Rs even inprimary cultures. Furthermore, antibodies against A1Rsare able to co-immunoprecipitate D1Rs (Fig. 2) [Ginés etal., 2000]. Proof of the functionality of the interaction hasbeen obtained by studying the effect of ligands on thedistribution of the receptors in the membrane. Thus,whereas agonists for A1Rs cluster the two receptors inco-transfected cells, the ligand for D1Rs cluster D1Rs only,
298 FRANCO ET AL.
but not A1Rs, which is associated with a disappearanceof A1/D1 heteromeric receptor complexes, as shown inco-immunoprecipitation experiments [Ginés et al., 2000].
ROLE OF CELL-SURFACE ADENOSINE DEAMINASEIN ADENOSINE/DOPAMINE ANTAGONISM
The work on A1 adenosine receptors has helpedresearchers understand how ligand binding and signaltransduction is affected by homotypic and heterotypicprotein-protein interactions. The binding of [3H]-2-chloroadenosine to A1Rs in rat brain membranes firstevidenced a single (low-affinity) [Wu et al., 1980; Wu andPhillis, 1982] or two (low- and high-affinity) binding sites[Williams and Risley, 1980a,b] in the absence or pres-ence of exogenous adenosine deaminase (ADA), respec-tively. The appearance of a high-affinity binding site inthe presence of ADA was explained first by the disap-pearance of endogenous adenosine, which acts as a com-petitor of A1R agonists, or by assuming that ADA had anextracatalytic high-affinity binding site for 2-chloro-adenosine [Phillis and Wu, 1981], which is not the caseaccording to the X-ray structure of the enzyme [Wilsonet al., 1991]. Subsequent studies have shown that the A1Rpresents two different affinities for agonists that dependon the coupling to heterotrimeric G proteins [Lohse etal., 1984]. The coupled receptor–G protein complexesdisplay high affinity for A1R agonists (Kd = 0.1–0.2 nM),whereas uncoupled receptors display low affinity (1–2nM) [Lohse et al., 1984; Casadó et al., 1990].
For many years it was thought that exogenous ADAacts by removing endogenous adenosine, but this maynot be the true explanation for the appearance of a high-affinity binding site in its presence. Sufficient evidencenow has accumulated to indicate that ADA and A1Rs in-teract and that the two proteins are functionally coupled.In pig cortical membranes, where the adenosine concen-tration is undetectable, it was found that ADA was nec-essary for the identification of the high-affinity componentof the binding to A1Rs (Table 1). In fact, in the absence ofADA, a single low-affinity binding site was found, whereastwo affinity sites were found when ADA was present(Table 1). If the high-affinity site corresponds to the re-ceptor–G protein complex, ADA would be necessary forthe coupling of A1Rs to G proteins [Saura et al., 1996].However, the cluster-arranged cooperative model, whichaccounts for the kinetics of ligand binding to A1Rs [Francoet al., 1996], shows that high- and low-affinity sites canbe a consequence of the negative cooperativeness of ago-nist binding and may not be related solely to the contentof free and G protein–coupled receptors. Therefore, ADAcould affect cooperativeness without affecting the A1R–G protein coupling. A smooth-muscle cell line (DDT1MF-2) is being used as a model of A1R-expressing cells, sincethe expressed A1Rs display binding kinetics similar tothat of the A1Rs present in the cerebral cortex. In thesecells, apart from the confirmation of an increase in theaffinity for A1R agonists only in the presence of ADA, amore molecular approach and the ADA/A1R interaction
Fig. 1. Scheme of adenosine/dopamine functional antagonism.

ADEN
OSIN
E/DO
PAMIN
E RECEPTO
R-RECEPTO
R HETERO
MERIZ
ATION
299
Fig. 2. Co-immunoprecipitation of A1 receptors (A1R) and D1 receptors (D1R) in co-transfected cells. Immunoblotting of cell lysates (positive control) and immunoprecipitates were done todetect A1R with anti-A1R antibody, D1R with anti-D1R antibody, or D2 receptors (D2R) with anti-D2R antibody. When indicated, A1/D1 co-transfected cells were incubated for 1 h with 10 µMSKF-38393 (agonist for D1R) in the absence or presence of 100 nM R. phenyl isopropyl adenosine (R-PIA) (agonist for A1R). The arrowhead indicates the band corresponding to D1Rs, and theasterisk the band for D2Rs. (From Ginés et al. [2000] with permission)

300 FRANCO ET AL.
was shown with confocal laser microscopy, affinity chro-matography, and co-immunoprecipitation [Ciruela et al.,1996; Saura et al., 1996].
The ADA/A1R interaction was needed for efficientsignaling via the adenosine A1R, and this was indepen-dent of the catalytic activity (Table 1). Therefore, ADA isacting not only catalytically but also in an extra-enzymaticway as a co-stimulatory molecule. The scenario is that atlow adenosine concentrations, ADA mainly facilitates sig-naling, whereas at high adenosine concentrations, bywhich the ADA/A1R interaction is disrupted, the low-af-finity state of the receptor predominates, and an autolo-
gous mechanism of desensitization is devised [see Francoet al., 1997, for review].
Can this ADA/A1R interaction have a role in ad-enosine/dopamine antagonism? This question was ad-dressed by studying the expression of cell-surface ADAin cells transfected with cDNA for the human D1R or co-transfected with the cDNAs for human D1Rs and A1Rs.Whereas D1R-transfected cells alone do not express ADAon the cell surface, A1R/D1R co-transfected cells do ex-press the enzyme, which co-localizes with D1Rs. Inexperiments on ligand-induced modulation of the cell-surface expression of D1Rs and A1Rs, it is found that cell-surface ADA always follows A1Rs. Thus, in co-transfectedcells, adenosine A1R agonists induce the clustering of bothADA/A1Rs and ADA/D1Rs, suggesting the clustering ofall three proteins. On the other hand, dopamine agonistsinduce the clustering of D1Rs alone and a reduction inthe degree of co-localization between ADA and D1Rs(unpublished data). These results are consistent with theexistence of modules composed of the three proteins inan undefined stoichiometry (Fig. 3).
The question remains whether these tripartite oli-gomers have any role in adenosine A1R-mediated loss ofthe high-affinity binding site for dopamine in D1Rs. Ex-periments on agonist binding to D1Rs in co-transfectedcells have been performed in the absence and presence
TABLE 1. Equilibrium parameters of [3H]R-PIA binding to pig braincortical membranes in the absence or presence of ADA*
Presence Bmax
of ADA Affinity state Kd (nM) (pmol/mg protein)
Non High affinity — —Low affinity 4.1 ± 0.5 0.80 ± 0.09
0.02 U/mL High affinity 0.22 ± 0.09 0.3 ± 0.1Low affinity 0.6 ± 0.9 0.5 ± 0.1
0.02 U/mL High affinity 0.25 ± 0.06 0.51 ± 0.06plus Hg2+ Low affinity 3.06 ± 0.5 0.55 ± 0.09
*Hg2+ (10 µM) was used to abolish the enzymatic activity of ADA with-out disrupting the interaction with the A1 adenosine receptor (modifiedfrom Saura et al. [1996]).
Fig. 3. Scheme of functional modules operating in the neuronal plasmamembrane. G proteins are not included in the scheme. For A1Rs, thereare two specific interactions with nonmembrane proteins, ADA [Ciruela
et al., 1996; Saura et al., 1996] and hsc73 [Sarrió et al., 2000]. No inter-acting nonmembrane proteins have been identified for D1R. The proteinhsc73 is the heat shock cognate protein of MW 73 kDa.

ADENOSINE/DOPAMINE RECEPTOR-RECEPTOR HETEROMERIZATION 301
of deoxycoformycin, which is an inhibitor of ADA butalso disrupts the ADA/A1R interaction. The presence ofdeoxycoformycin blocked the effect of adenosine A1Ragonists on the binding of dopamine analogs to D1Rs.This effect was not due to the increase in adenosine con-centration, because similar experiments performed in thepresence of exogenous adenosine failed to block the ef-fect of adenosine A1R agonists on dopamine binding toD1Rs. These results show that ADA is necessary to ob-tain adenosine/dopamine antagonism on D1R agonistbinding by making possible a high-affinity state of theA1R, which is required for the effect of A1R agonists ondopamine agonist binding to D1Rs.
ROLE OF FUNCTIONAL UNITS ON THENEURON CELL SURFACE
In view of the recent discovery of dimers orheteromers for G protein-coupled receptors (GPCRs), themolecular mechanism of dimerization and heteromer-ization is under investigation. The evidence so far indi-cates that certain transmembrane domains participate inthe interaction and that the interaction involves mainlynonpolar residues. The theoretical biology approachestaken by Reynolds and colleagues [Gouldson et al., 1997,1998] have predicted that the protein-protein interactionfor homodimer formation (and eventually for heterodimerformation) is mediated by domain swapping involvingtransmembrane regions 5 and 6 and probably other heli-cal transmembrane domains as well. The only proven ex-ception to this general trend is in the case of dimerizationof metabotropic glutamate 5 receptors, for which a disul-fide-linked structure has been shown for the dimer[Romano et al., 1996]. This may be unique to these recep-tors, since they show little homology with other membersof the GPCR family. On the other hand, the existence ofindirect interactions through bridging proteins cannot beruled out. The occurrence of clustering, as shown for ad-enosine/dopamine receptor-receptor interactions, clearlyreflects that GPCRs form high-molecular-order structuresin which multimers of the receptors and probably otherinteracting proteins form functional complexes whoseprecise role in the biochemistry and physiology of neu-rons will require more experimental effort in the future.
D1R and A1R, like other GPCRs [Maggio et al.,1993], are multifunctional proteins that are able to formhigh-order molecular structures containing at least tworeceptor molecules, heterotrimeric G proteins and, inall likelihood, other proteins. So far no proteins have beenidentified that interact with D1Rs, but for A1Rs it hasbeen possible to identify ADA and hsc73 as interactingproteins. As mentioned earlier, ADA interacts with theextracellular domains of A1Rs and has an important rolein ligand binding and in signaling. The protein hsc73has been shown to interact with the intracellular domains
of the receptor and, in contrast to what happens withADA, has a negative effect on ligand binding. This pro-tein affects the binding of adenosine deaminase to A1Rs,and the preliminary results indicate that A1Rs cannotbind to both proteins at the same time [Sarrió et al., 2000].It should be noted that all these interactions play a rolenot only in ligand binding and signaling but also in re-ceptor trafficking (Fig. 3).
Taken together, the existence of homodimers andheterodimers of various types indicates that the opera-tion of receptors for neurotransmitters or neuromodul-ators may involve homotypic and heterotypic interactionsthat are crucial for GPCR function and for ligand-gatedchannels. It is quite reasonable that the interactions mightassemble and disassemble, depending essentially uponthe types of receptors present and the presence of differ-ent transmitters/modulators in the extracellular medium.As an example, there is evidence that A1Rs interact withmetabotropic glutamate receptors in cerebellar neurons(see Ciruela et al., this issue). Therefore, the heterotypicinteractions established with A1Rs in a given cell woulddepend upon the presence of D1Rs, metabotropicglutamate receptors, or both. Furthermore, the geometryof the interactions in the clusters formed after activationof the receptors may depend upon the combination ofneuromodulators present in the extracellular medium andin the synapses.
In the central nervous system this scenario is at-tractive, since receptor-receptor interactions likely areinvolved in neurotransmission and neuromodulation aswell as in the development of higher neural functions,such learning and memory. At a given plasma membrane,there are many interactions between receptors and adap-tor proteins, such as ADA itself (Fig. 3). This leads to amosaic that is specific for a given neuron and is respon-sible for a certain action of a given neuromodulator. Onthe other hand, this mosaic is dynamic, since ligands af-fect the composition and geometry of the mosaic. Thismay constitute the basis to understand better how thecentral neuronal communication network is supportedby the multifunctional role of neuronal GPCRs.
REFERENCESAgnati L, Fuxe K, Zoli M, Rondanini C, Ogren SO. 1982. New vistas
on synaptic plasticity: mosaic hypothesis of the engram. Med Biol60:183–190.
Agnati L, Benfenati F, Solfrini V, Biagini G, Fuxe K, Guidolin D, CaraniC, Zini I. 1993. Intramembrane receptor-receptor interactions: in-tegration of signal transduction pathways in the nervous system.Neurochem Int 22:213–222.
Aoyama S, Kase H, Borrelli E. 2000. Rescue of locomotor impairmentin dopamine D2 receptor–deficient mice by an adenosine A2A re-ceptor antagonist. J Neurosci 20:5848–5858.
Casadó V, Cantí C, Mallol J, Canela EI, Lluis C, Franco R. 1990. Solu-bilization of A1 adenosine receptor from pig brain: characterization

302 FRANCO ET AL.
and evidence of the role of the cell membrane on the coexistence ofhigh- and low-affinity states. J Neurosci Res 26:461–473.
Ciruela F, Casadó V, Mallol J, Canela EI, Lluis C, Franco R. 1995.Immunological identification of A1 adenosine receptors in braincortex. J Neurosci Res 42:818–828.
Ciruela F, Saura C, Canela EI, Mallol J, Lluis C, Franco R. 1996. Ad-enosine deaminase affects ligand-induced signalling by interactingwith cell surface adenosine receptors. FEBS Lett 380:219–223.
Ferré S, von Euler G, Johansson B, Fredholm BB, Fuxe K. 1991. Stimu-lation of high affinity adenosine A-2 receptors decreases the affinityof dopamine D-2 receptors in rat striatal membranes. Proc Natl AcadSci U S A 88:7238–7241.
Ferré S, Fuxe K, von Euler G, Johansson B, Fredholm BB. 1992.Adenosine-dopamine interactions in the brain. Neuroscience 51:501–512.
Ferré S, O’Connor WT, Fuxe K, Ugerstedt U. 1993. The striopallidalneuron: a main locus for adenosine-dopamine interactions in thebrain. J Neurosci 13:5402–5406.
Ferré S, Fredholm BB, Morelli M, Popoli P, Fuxe K. 1997. Adenosine-dopmaine interactions as an integrative mechanism in the basal gan-glia. Trends Neurosci 20:482–487.
Ferré S, Torvinen M, Antoniou K, Irenius E, Civelli O, Arenas E,Fredholm BB, Fuxe K. 1998. Adenosine A1 receptor–mediatedmodulation of dopamine D1 receptors in stably cotransfected fibro-blasts cells. J Biol Chem 273:4718–4724.
Franco R, Casadó V, Ciruela F, Mallol J, Lluis C, Canela EI. 1996. Thecluster-arranged cooperative model: a model that accounts for thebinding kinetics to A1 adenosine receptors. Biochemistry 35:3007–3015.
Franco R, Casadó V, Ciruela F, Saura C, Mallol J, Canela EI, Lluis C.1997. Cell surface adenosine deaminase: much more than anectoenzyme. Prog Neurobiol 52:283–294.
Fredholm BB. 1995. Adenosine, adenosine receptors and the actionsof caffeine. Pharmacol Toxicol 76:93–101.
Fredholm BB, Abbracchio MP, Burnstock G, Daly JW, Harden TK,Jacobson KA, Leff P, Williams M. 1994. Nomenclature and classifi-cation of purinoceptors. Pharmacol Rev 46:143–156.
Fuxe K, Agnati L. 1987. Receptor-receptor interactions. Wenner-GrenCenter International Symposium Series. Southampton, UK: Mac-Millan Press.
Fuxe K, Agnati LF, Benfenati F, Celani M, Zini I, Zoli M, Mutt V. 1983.Evidence for the existence of receptor-receptor interactions in thecentral nervous system: studies on the regulation of monoaminereceptors by neuropeptides. J Neural Transm (Suppl.)18:165–179.
Ginés S, Hillion J, Torvinen M, Le Crom S, Casadó V, Canela EI, RondinS, Lew JY, Watson S, Zoli M, Agnati L, Vernier P, Lluis C, Ferré S,Fuxe K, Franco R. 2000. Dopamine D1 and adenosine A1 receptorsassemble into functionally interacting heteromeric complexes ProcNatl Acad Sci U S A 97:8606–8611.
Gouldson PR, Snell CR, Reynolds CA. 1997. A new approach to dock-ing in the beta 2-adrenergic receptor that exploits the domain struc-ture of G-protein-coupled receptors. J Med Chem 40:3871–3886.
Gouldson PR, Snell CR, Bywater RP, Higgs C, Reynolds CA. 1998.Domain swapping in G-protein coupled receptor dimers. ProteinEng 11:1181–1193.
Kull B, Ferré S, Arslan G, Svenningsson P, Fuxe K, Owman C, FredholmBB. 1999. Reciprocal interactions between adenosine A2A anddopamine D2 receptors in CHO cells co-transfected with the tworeceptors. Biochem Pharmacol 58:1035–1045.
Lohse MJ, Lenschow V, Schwabe U. 1984. Two affinity states of Riadenosine receptors in brain membranes. Mol Pharmacol 26:1–9.
Maggio R, Vogel Z, Wess J. 1993. Coexpression studies with mutantmuscarinic/adrenergic receptors provide evidence for intermolecular“cross-talk” between G-protein-linked receptors. Proc Natl AcadSci U S A 90:3103–3107.
Ng GY, O’Dowd BF, Lee SP, Chung HT, Brann S, Seeman P, GeorgeSR. 1996. Dopamine D2 receptor dimers and receptor-blockingpeptides. Biochim Biophys Res Commun 227:200–204.
Phillis JW, Wu PH. 1981. The role of adenosine and its nucleotides incentral synaptic transmission. Prog Neurobiol 16:187–239.
Rivkees SA, Price SL, Zhou FC. 1995. Immunohistochemical detec-tion of A1 adenosine receptors in rat brain with emphasis in local-ization in the hippocampal formation, cerebral cortex, cerebellum,and basal ganglia. Brain Res 677:193–203.
Romano C, Yang WL, O’Malley KL. 1996. Metabotropic glutamatereceptor 5 is a disulfide-linked dimer. J Biol Chem 271:28612–28616.
Salim H, Ferré S, Dalal A, Peterfreund RA, Fuxe K, Vincent D, LledóPM. 2000. Activation of adenosine A1 and A2A receptors modulatesdopamine D2 receptor–induced responses in stably transfectedhuman neuroblastoma cells. J Neurochem 74:432–439.
Sarrió S, Casadó V, Escriche M, Ciruela F, Mallol J, Canela EI, LluisC, Franco R. 2000. The heat shock cognate protein hsc73 assembleswith A1 adenosine receptors to form functional modules in the cellmembrane Mol Cell Biol 20:5164–5174.
Saura C, Ciruela F, Casadó V, Canela EI, Mallol J, Lluis C, Franco R.1996. Adenosine deaminase interacts with A1 adenosine receptorsin pig brain cortical membranes. J Neurochem 66:1675–1682.
Schiffmann SN, Jacobs O, Vanderhaeghen JJ. 1991. Striatal restrictedadenosine A2 receptor (RDC8) is expressed by enkephalin but notby substance P neurons: an in situ hybridization histochemistry study.J Neurochem 57:1062–1067.
Williams M, Risley E. 1980a. Biochemical characterization of putativecentral purinergic receptors using [3H]2-chloroadenosine, a stableanalog of adenosine. Proc Natl Acad Sci USA 77:6892–6896.
Williams M, Risley E. 1980b. High affinity binding of 2-chloro-adenosine to rat brain synaptic membranes. Eur J Pharmacol 64:369–370.
Wilson DK, Rudolph FB, Quiocho FA. 1991. Atomic structure of ad-enosine deaminase complexed with a transition-state analog: un-derstanding catalysis and immunodeficiency mutations. Science252:1278–1284.
Wu PH, Phillis JW. 1982. Adenosine receptors in rat brain membranes:characterization of high affinity binding of [3H]2-chloroadenosine.Int J Biochem 14:399–402.
Wu PH, Phillis JW, Balls K, Rinaldi B. 1980. Specific binding of [3H]2-chloroadenosine to rat brain cortical membranes. Can J PhysiolPharmacol 58:575–579.
Zahniser NR, Simosky JK, Mayfield RD, Negri CA, Hanania T, LarsonGA, Kelly MA, Grandy DK, Rubinstein M, Low MJ, Fredholm BB.2000. Functional uncoupling of adenosine A2A receptors and re-duced response to caffeine in mice lacking dopamine D2 receptors.J Neurosci 20:5949–5957.
Zawarynski P, Tallerico T, Seeman P, Lee SP, O’Dowd BE, George SR.1998. Dopamine D2 receptor dimers in human and rat brain. FEBSLett 441:383–386.
Zoli M, Agnati LF, Hedlund PB, Li XM, Ferré S, Fuxe K. 1993. Recep-tor-receptor interactions as an integrative mechanism in nerve cells.Mol Neurobiol 7:293–334.