Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Feb. 1976, p. 619-626Copyright © 1976 American Society for Microbiology
Vol. 13, No. 2Printed in U.SA.
Activity of Two Streptococcus mutans Bacteriocins in thePresence of Saliva, Levan, and Dextran
ALLAN L. DELISLEDepartment of Microbiology, University of Maryland School of Dentistry, Baltimore, Maryland 21201
Received for publication 18 August 1975
The extracellular dextrans produced from sucrose by Streptococcus mutansstrains BHT and GS-5 did not prevent the synthesis or release of active bacterio-cins by these two strains. In addition, several streptococci that were geneticallysensitive to these bacteriocins, and that could synthesize a variety of extracellu-lar dextrans and levans from sucrose, remained phenotypically sensitive whengrown in the presence of sucrose. Bacteriocin activity was not altered by treat-ment with high-molecular-weight dextran or by human saliva. The bacteriocinsproduced by, and active against, S. mutans thus appear to be capable of acting invivo and may play a role in regulating the bacterial ecology ofthe oral cavity.
Bacteriocinogenic strains of cariogenic strep-tococci are known to exist in the human oralcavity (7), but whether their bacteriocins play arole in regulating the bacterial ecology of thisenvironment is not known. It is not yet possibleto quantitate, or even detect, active bacterio-cins in dental plaque and saliva, so in vivoproduction of bacteriocins cannot be unequivo-cally demonstrated; however, there are no com-pelling a priori reasons why bacteriocins shouldnot be produced in the oral cavity. Currentresearch in this area has therefore focused onthe question of whether bacteriocins, particu-larly those of Streptococcus mutans, can exerttheir lethal effects under conditions simulatingthose of the oral environment. Thus, it hasbeen reported that some S. mutans bacteriocinsappear to be inactivated by proteolytic enzymesin saliva (8) and, more recently, that bacterio-cin-sensitive strains of S. mutans and S. sali-varius become insensitive to bacteriocins whengrown in the presence of sucrose, due to theirelaboration of dextran and levan polysaccha-rides (11).
This investigation was undertaken to deter-mine whether two S. mutans bacteriocins pres-ently under study in this laboratory are inacti-vated by enzymes in saliva and whether theycan kill sensitive cells that are coated withextracellular glucan and fructan polysaccha-rides synthesized from sucrose.
MATERIALS AND METHODS
Bacteriocin-producing strains. S. mutans BHTwas obtained from D. D. Zinner, University ofMiami Institute of Oral Biology, Miami, Fla. Thisstrain, which is a cariogenic human isolate (14) be-
longing to Bratthall's serological group (b) (1), pro-duces a highly insoluble extracellular dextran fromsucrose. S. mutans GS-5 was obtained from R. J.Gibbons, Forsyth Dental Center, Boston, Mass. Thisstrain, which belongs to serological group c, is also adextran-producing, cariogenic human isolate (5).The bacteriocins produced by these strains, whichhave similar physical and chemical properties (7; D.Paul and H. D. Slade, Abstr. Annu. Meet. Am. Soc.Microbiol. 1974, M267, p. 110; A. L. Delisle, J. Dent.Res. 54A:L76, 1975), are lethal to all of the indicatorstrains given below.
Bacteriocin-sensitive indicator strains. S. mu-tans FA-1 was obtained from H. V. Jordon, NationalInstitute of Dental Research, Bethesda, Md.; it is acariogenic rat isolate (3) that produces no detectablebacteriocins but is otherwise similar to strain BHT.S. salivarius 13419 was obtained from the AmericanType Culture Collection (ATCC); it produces mainlya water-soluble levan from sucrose (10) and is onlyweakly cariogenic (12). S. sanguis 10556 (ATCC) is anoncariogenic organism (9), isolated from a patientwith subacute bacterial endocarditis, that producesboth dextran and levan from sucrose (13). S. bovis9809 (ATCC) is a bovine rumen isolate that wasincluded for comparative purposes because, al-though it is noncariogenic (9), it produces copiousamounts of a water-soluble dextran from a sucrose.A streptomycin-resistant mutant (9809strr) of thisstrain was isolated after ultraviolet light mutagene-sis and selection on agar containing 1 g of strepto-mycin per liter. The S. pyogenes culture used is agroup A strain from this department collection; itproduces no unique polysaccharides from sucrose.
Media and chemicals. All bacteriological mediawere purchased from BBL, Division of Becton, Dick-inson & Co., Cockeysville, Md. Trypticase soy broth(TSB) was commonly used, with 15 g of agar addedper liter for bottom agar (TSA) or 7.5 g/liter addedfor top agar overlays. To provide a substrate forsynthesis of extracellular levans and dextrans, su-
619
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

620 DELISLE
crose was added to a final concentration of 5% (wt/vol) to TSB, TSA, or TSB top agar. Strain BHT wasadditionally tested on sucrose-containing mediasupplemented with 1.5% (wt/vol) L-arginine (SigmaChemical Co.). One milliliter of a 1% aqueous solu-tion of bromocresol purple (Fisher Scientific Co.)was added per liter of agar media in some experi-ments as a pH indicator. S. bovis 9809strr was al-ways grown in media containing 1 g of streptomycinsulfate per liter.
Bacteriological-grade sucrose was obtained fromBBL. Dextran type 2000 (molecular weight, 2 x 106)was obtained from Sigma Chemical Co. Dialysistubing with an average molecular weight cut-offpore size of 3,500 (Spectropor type 3) was obtainedfrom Spectrum Medical Industries, Los Angeles,Calif. Phosphate buffer (0.05 M, pH 7.0) was used fordialyzing and diluting purified bacteriocin.
Bacteriocin testing procedures. A modification ofthe classical stab overlay method was used to testthe activity of and susceptibility to bacteriocins pro-duced on agar media. Producer cultures werestreaked across the surface of an agar plate with astraight inoculating needle, using TSA slant cul-tures as the inocula. The plates were incubated for48 h at 37 C in Brewer Jars containing an atmos-phere of 95% H2 and 5% CO2 (GasPak, BBL). Indica-tor cultures were grown overnight (16 to 18 h) at37 C in TSB containing arginine and/or sucrose, and2 to 3 drops was added to the appropriate top agar(melted and held at 42 C). The seeded top agar tubeswere then immediately poured onto producer streakplates. After solidifying, the overlayed plates wereincubated aerobically overnight at 37 C. Inhibitionzones were measured and recorded the followingmorning. In each experiment the compositions ofthemedia used for bottom agar, top agar, and indicatorbroth cultures were identical except for agar concen-tration and the addition of streptomycin.The above method was used to determine whether
producer cells synthesize and release active bacteri-ocins when grown in sucrose and whether sucrose-grown indicator cells are sensitive to these bacterio-cins. The latter was also tested in an aqueous, agar-free system by using a partially purified BHT bacte-riocin preparation (described below). In this method,serial dilutions of the bacteriocin suspension werespotted onto TSB-sucrose top agar overlays seededwith TSB-sucrose-grown indicator cells. Bacteriocindestruction was revealed by a reduction in the dilu-tion of bacteriocin that showed visible inhibition ofthe indicator lawn after 24 h incubation at 37 C.The effects of saliva and purified dextran on bac-
teriocin activity were also tested in two ways. In thefirst method, 0.1 ml of 1% (wt/vol) dextran 2000 orfreshly collected saliva (unstimulated saliva sam-ples from two individuals with average dentalhealth were pooled) was spotted directly on producerstreaks, either 4 h before (and held at 37 C) or imme-diately after overlaying with indicator cells in TSBtop agar. Both untreated and sterile filtered salivawere used. After an additional 24-h incubation pe-riod, any reduction in size of the inhibition zones inthe area of the spots was noted. In the secondmethod, the cell-free BHT bacteriocin preparation
was again used. The preparation was mixed with anequal volume of dextran 2000 (1% wt/vol) or unstim-ulated, freshly collected saliva (either untreated orsterile filtered) and incubated for 1 h in a 37 C waterbath, and then serial dilutions were spotted ontoTSB top agar lawns seeded with S. pyogenes. After 4h of incubation at 37 C, destruction of bacteriocinactivity was evidenced by a reduction in the highestdilution that showed visible inhibition of the indica-tor lawn.
Preparation of cell-free BHT bacteriocin. StrainBHT was grown overnight at 37 C in a sterile me-dium composed of half-strength APT broth (BBL)and 4% (wt/vol) yeast extract. The culture was soni-cated for 1 min (Branson LS 75 Sonifier, 6A) andcentrifuged for 10 min at 8,000 x g, and the superna-tant fluid was removed and adjusted to pH 7.4 with0.1 N NaOH. Solid (NH4)2SO4 (Sigma, enzymegrade) was then slowly added to 75% saturation, andthe mixture was stirred gently for 4 h at 4 C. Theprecipitate from 100 ml of culture was collected bycentrifuging for 10 min at 10,000 x g, redissolved in5 ml of phosphate buffer (0.05 M, pH 7.0), dialyzedfor 18 h against three 1,000-volume changes ofbufferat 4 C, adjusted to pH 7.0, and then sterile filtered(Nalgene Corp., disposable membrane filters, 0.45-,um pore size). The preparation used for all the ex-periments reported here showed detectable bacterio-cin activity in spot tests at a dilution of 1:100.
RESULTSBHT bacteriocin activity at neutral pH.
To eliminate the possibility that inhibitionzones might be due to the relatively high levelsof lactic acid produced by S. mutans rather thatto bacteriocins per se, L-arginine was added tobottom and top agar media to control the pH.Strain BHT, along with S. pyogenes and S.mutans strains belonging to Bratthall's sero-logical group b, has a potent arginine deimi-nase (4; L-arginine iminohydrolase, EC 3.5.3.6);the production ofammonia from added argininethus neutralizes the acid(s) produced duringmetabolism. The amount of arginine needed tomaintain the pH near 7.0 was determined em-pirically in preliminary experiments. When1.5% arginine was added to TSB agar, the finalpH, after 24 to 48 h ofgrowth at 37 C, was in therange of 6.8 to 7.2. Under these conditions thesensitive indicator S. pyogenes was stronglyinhibited by BHT bacteriocin; the inhibitionzone was, in fact, significantly larger than onunsupplemented TSB agar, where the pH coulddrop as low as 5.2 (Fig. 1). The larger zone athigher pH may have been due to a higher cellyield under these conditions, to stimulation ofbacteriocin production, or to enhancement ofbacteriocin activity. In any case, inhibition ofthe sensitive indicator strains by BHT was notdue to acid.
Bacteriocin production by sucrose-grown
INFECT. IMMUN.
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
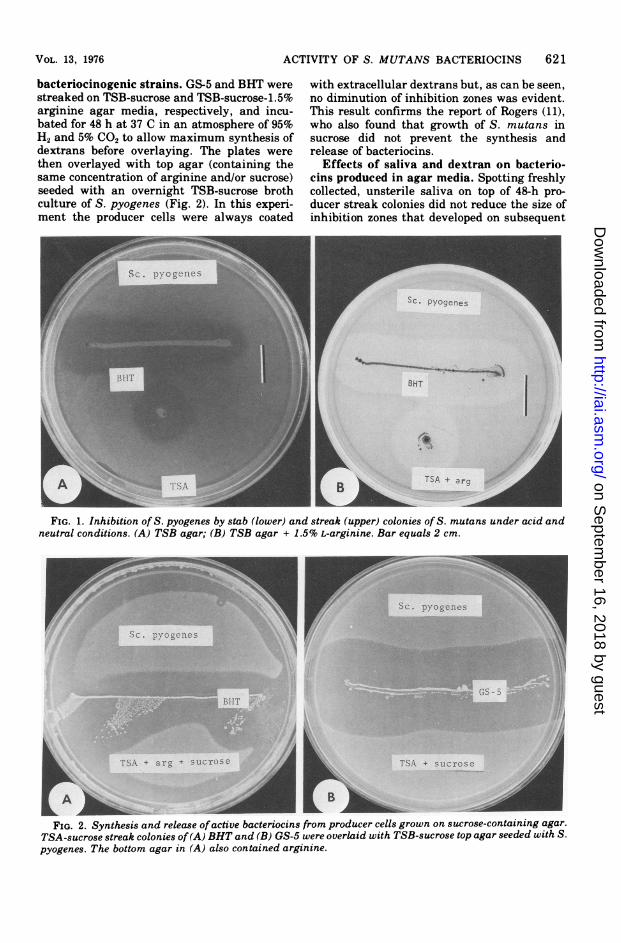
ACTIVITY OF S. MUTANS BACTERIOCINS 621
bacteriocinogenic strains. GS-5 and BHT werestreaked on TSB-sucrose and TSB-sucrose-1.5%arginine agar media, respectively, and incu-bated for 48 h at 37 C in an atmosphere of 95%H2 and 5% CO2 to allow maximum synthesis ofdextrans before overlaying. The plates werethen overlayed with top agar (containing thesame concentration of arginine and/or sucrose)seeded with an overnight TSB-sucrose brothculture of S. pyogenes (Fig. 2). In this experi-ment the producer cells were always coated
_~c7Lo~c eT_
with extracellular dextrans but, as can be seen,no diminution of inhibition zones was evident.This result confirms the report of Rogers (11),who also found that growth of S. mutans insucrose did not prevent the synthesis andrelease of bacteriocins.
Effects of saliva and dextran on bacterio-cins produced in agar media. Spotting freshlycollected, unsterile saliva on top of 48-h pro-ducer streak colonies did not reduce the size ofinhibition zones that developed on subsequent
r n mFIG. 1. Inhibition of S. pyogenes by stab (lower) and streak (upper) colonies of S. mutans under acid and
neutral conditions. (A) TSB agar; (B) TSB agar + 1.5% L-arginine. Bar equals 2 cm.
SC. pvOgc'c
SC pvCgcneS
C.S-ID I
L IA
L VBX + arg + Sucrs +
FIG. 2. Synthesis and release ofactive bacteriocins from producer cells grown on sucrose-containing agar.TSA-sucrose streak colonies of(A) BHT and (B) GS-5 were overlaid with TSB-sucrose top agar seeded with S.pyogenes. The bottom agar in (A) also contained arginine.
VOL. 13, 1976
-NJFl
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

622 DELISLE
incubation of plates overlayed with S. bovis9809strr (Fig. 3). Here streptomycin was incor-porated into the top agar to minimize growth ofcontaminants from the raw saliva. Identicalresults (not shown) were obtained by spottingwith sterile filtered saliva and dextran 2000.
Effects of saliva and dextran on cell-freeBHT bacteriocin activity. When the partiallypurified BHT bacteriocin was incubated withsaliva (either raw or sterile filtered), no reduc-tion in bacteriocin activity could be detected byspotting serial dilutions on lawns ofS. pyogenes(Fig. 4). Here the pH of the saliva-bacteriocinsystem could be controlled, and no differencewas observed when the pH was adjusted to 6.0,6.5, or 7.0. Again, identical results (not shown)were obtained by incubating the bacteriocinpreparation with dextran 2000.
Susceptibility of polysaccharide-synthesiz-ing, sucrose-grown indicators. When geneti-cally sensitive strains were pregrown in TSB-sucrose broth, inoculated into TSB-sucrose topagar, and overlayed on 2-day-old TSB-sucroseagar streak cultures of GS-5 and BHT, clearinhibition zones developed after incubation.Moreover, the zone sizes did not differ fromthose on sucrose-free media (data not shown).Figure 5 shows the results with GS-5, usingarginine-free media, and Fig. 6 shows the re-sults with BHT, using arginine-containing me-dia to control the pH. In all cases the sensitiveindicator strains remained sensitive to both
bacteriocins when grown in the presence of su-crose. Under these conditions strains 13419 and9809 synthesized copious amounts of extracellu-lar polysaccharides, which is clearly evidentfrom the figures. Strains FA-1 and 10556 pro-duced tenacious dextran lawns under these con-ditions, but these lawns did not have the highlymucoid, "photogenic" appearance obtained withthe other strains. The inhibition zones againstFA-1 were quite narrow because this strain wasthe least sensitive of all the indicator strains.Since the relative size of the inhibition zonesproduced against each indicator strain, by bothproducers, remained constant on a wide varietyof sucrose-free media (data not shown), the zonesizes reflect differences in sensitivity to the bac-teriocins rather than differences in the type orquantity of extracellular polysaccharides syn-thesized from sucrose. The possibility that zonesize differences may be due to multiple bacteri-ocins cannot be ruled out but is considered un-likely since single-step mutants of BHiT thathad lost bacteriocin activity against S. py-ogenes lost activity against all the other indica-tor strains at the same time (Delisle, unpub-lished data).When BHT cell-free bacteriocin was spotted
onto TSB-sucrose top agar lawns seeded withsucrose-grown cultures of strains 13419, 9809,FA-1, and 10556, definite inhibition zones de-veloped (after overnight incubation) at all dilu-tions that gave positive results in the complete
9809 StrI'
Bl.Ti
_ _~~~~~~~~- ~
FIG. 3. Effect ofsaliva on bacteriocin activity in agar. A streak colony ofBHT on TSA-arginine was spottedwith saliva (arrow), incubated for 4 h at 3 7 C, and then overlaid with TSB-streptomycin top agar seeded withS. bovis 9809strr. After 18 h the indicator lawn shows no discernible reduction in the size ofthe inhibition zonein the region of the saliva.
INFECT. IMMUN.
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

ACTIVITY OF S. MUTANS BACTERIOCINS
1
,- IitrC) I - .iI rI- ; c tcr.
, IC 11 a itOnl
i / 40
KI/1;'
1/ ,
S a I t -.'
'; _- t I's x<i,
/_ (
'1 4
I /Z(!
1 /_ AIi //D.)
F
jOictercin:/ i:'
1./>1
C
FIG. 4. Effect of untreated and sterile filtered saliva on the activity ofcell-free BHT bacteriocin. Bacteriocinwas mixed with an equal volume of(A) sterile TSB, (B) raw saliva, or (C) sterile filtered saliva and incubatedfor I h at 37 C. Serial dilutions (indicated on plates) were then spotted onto fresh overlays of S. pyogenes inTSB top agar. The plates were incubated at 37 C and photographed at 4 h.
absence of sucrose (data not shown), thus con-firming that the polysaccharides synthesized bythe indicators do not prevent bacteriocin ad-sorption and activity.
DISCUSSIONThe observation that inhibition zones pro-
duced by BHT in the presence of arginine arelarger than those in the absence of arginine(Fig. 1), for whatever reason, suggests that thefrequent, drastic changes that occur in the oralenvironment could greatly influence the pro-duction of bacteriocins in vivo. Information on
the effects of these changes is needed since it
might enable us to control specific species in themouth by treatments that stimulate (or re-
press) bacteriocin synthesis.Some workers have claimed that the coexist-
ence of producer and sensitive strains of S.mutans at the same site is common and thatthis implies that bacteriocins do not inhibitsensitive strains in vivo (8, 11), otherwise sensi-tive strains would be eliminated from the oralcavity; however, no data (or identification ofspecific strains) have been presented to supportthis view. These claims may therefore be basedon observations of acid sensitivity rather thanon sensitivity to bacteriocins. It has recently
/.
VOL. 13, 1976 623
I
J l -i
1.
B
Nftli
.,C-
1 /, I"I
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

INFECT. IMMUN.624 DELISLE
F¾
A.
VFIG. 5. Activity of GS-5 bacteriocin against sucrose-grown indicator cells on sucrose agar. Two-day-old
TSA-sucrose streaks of GS-5 were overlaid with TSB-sucrose top agar seeded with overnight TSB-sucrosebroth cultures of (A) S. salivarius, (B) S. bovis, (C) S. sanguis, and (D) S. mutans FA-1.
been shown, for example, that the vast major-ity of bacteriocin-like inhibition zones producedby oral streptococci against each other are duesolely to acid (2).From the above, it could be argued that the
activity of and susceptibility to GS-5 bacterio-cin in the presence of saliva and polysaccha-rides may be reduced or even eliminated, andthe results reported here are really due to acidinhibition of the various indicator strains. Inthe case of strain BHT, the use of arginine-containing media precludes such an argumentsince the amount of ammonia released by thearginine deiminase of this organism is suffi-cient to neutralize all of the acid produced dur-ing growth.The results reported in this study do not an-
swer the question of whether S. mutans bacte-
riocins are produced in vivo but do clearly dem-onstrate that (i) saliva does not degrade all S.mutans bacteriocins, (ii) high-molecular-weight dextran does not inhibit all S. mutansbacteriocins, and (iii) synthesis of extracellularlevans and dextrans from sucrose by oral strep-tococci does not prevent bacteriocin adsorptionand action. Whether saliva from all individualsor all types of in vivo-synthesized dextrans andlevans behave similarly is not known, but thedata suggest that if, as is generally believed,the general matrix of dental plaque consists ofdextrans, bacteriocins can diffuse through thismatrix in vivo and exert their lethal effects. S.mutans bacteriocins may therefore play a sig-nificant role in regulating the ecology of theoral cavity. Furthermore, since S. mutans bac-teriocins are active against a rather wide spec-
AiKi. ........
S-I
;)4 --' 1-l)'-i
Ew..
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
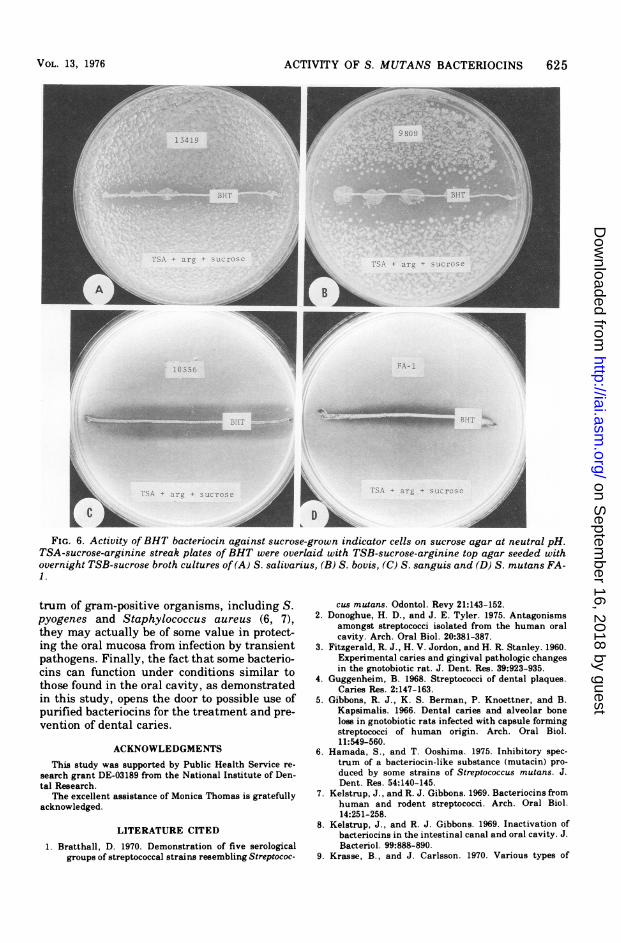
ACTIVITY OF S. MUTANS BACTERIOCINS 625
1.5 .1
A
FIG. 6. Activity ofBHT bacteriocin against sucrose-grown indicator cells on sucrose agar at neutral pH.TSA-sucrose-arginine streak plates of BHT were overlaid with TSB-sucrose-arginine top agar seeded withovernight TSB-sucrose broth cultures of (A) S. salivarius, (B) S. bovis, (C) S. sanguis and (D) S. mutans FA-1.
trum of gram-positive organisms, including S.pyogenes and Staphylococcus aureus (6, 7),they may actually be of some value in protect-ing the oral mucosa from infection by transientpathogens. Finally, the fact that some bacterio-cins can function under conditions similar tothose found in the oral cavity, as demonstratedin this study, opens the door to possible use ofpurified bacteriocins for the treatment and pre-vention of dental caries.
ACKNOWLEDGMENTS
This study was supported by Public Health Service re-search grant DE-03189 from the National Institute of Den-tal Research.
The excellent assistance of Monica Thomas is gratefullyacknowledged.
LITERATURE CITED
1. Bratthall, D. 1970. Demonstration of five serologicalgroups of streptococcal strains resembling Streptococ-
cus mutans. Odontol. Revy 21:143-152.2. Donoghue, H. D., and J. E. Tyler. 1975. Antagonisms
amongst streptococci isolated from the human oralcavity. Arch. Oral Biol. 20:381-387.
3. Fitzgerald, R. J., H. V. Jordon, and H. R. Stanley. 1960.Experimental caries and gingival pathologic changesin the gnotobiotic rat. J. Dent. Res. 39:923-935.
4. Guggenheim, B. 1968. Streptococci of dental plaques.Caries Res. 2:147-163.
5. Gibbons, R. J., K. S. Berman, P. Knoettner, and B.Kapsimalis. 1966. Dental caries and alveolar boneloss in gnotobiotic rats infected with capsule formingstreptococci of human origin. Arch. Oral Biol.11:549-560.
6. Hamada, S., and T. Ooshima. 1975. Inhibitory spec-trum of a bacteriocin-like substance (mutacin) pro-duced by some strains of Streptococcus mutans. J.Dent. Res. 54:140-145.
7. Kelstrup, J., and R. J. Gibbons. 1969. Bacteriocins fromhuman and rodent streptococci. Arch. Oral Biol.14:251-258.
8. Kelstrup, J., and R. J. Gibbons. 1969. Inactivation ofbacteriocins in the intestinal canal and oral cavity. J.Bacteriol. 99:888-890.
9. Krasse, B., and J. Carlsson. 1970. Various types of
VOL. 13, 1976
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

626 DELISLE INFECT. IMMUN.
streptococci and experimental caries in hamsters.Arch. Oral Biol. 15:25-32.
10. Newbrun, E., and S. Baker. 1968. Physico-chemicalcharacteristics of the levan produced by Streptococcussalivarius. Carbohydr. Res. 6:165-170.
11. Rogers, A. H. 1974. Bacteriocin production and suscep-tibility among strains of Streptococcus mutans grownin the presence of sucrose. Antimicrob. Agents Chem-other. 6:547-550.
12. Rosen, S. 1969. Comparison of sucrose and glucose in
the causation of dental caries in gnotobiotic rats.Arch. Oral Biol. 14:445-450.
13. Sidebotham, R. L., H. Weigel, and W. H. Bowen. 1971.Studies on dextrans and dextranases. Part IX. Dex-trans elaborated by cariogenic organisms. Carbo-hydr. Res. 19:151-159.
14. Zinner, D. D., A. P. Aran, J. M. Jablon, M. S. Saslaw,and R. J. Fitzgerald. 1967. Induction of dental cariesin gnotobiotic rats by streptococci of human origin.Nature (London) 213:200-201.
on Septem
ber 16, 2018 by guesthttp://iai.asm
.org/D
ownloaded from





























