Embed Size (px)
Citation preview
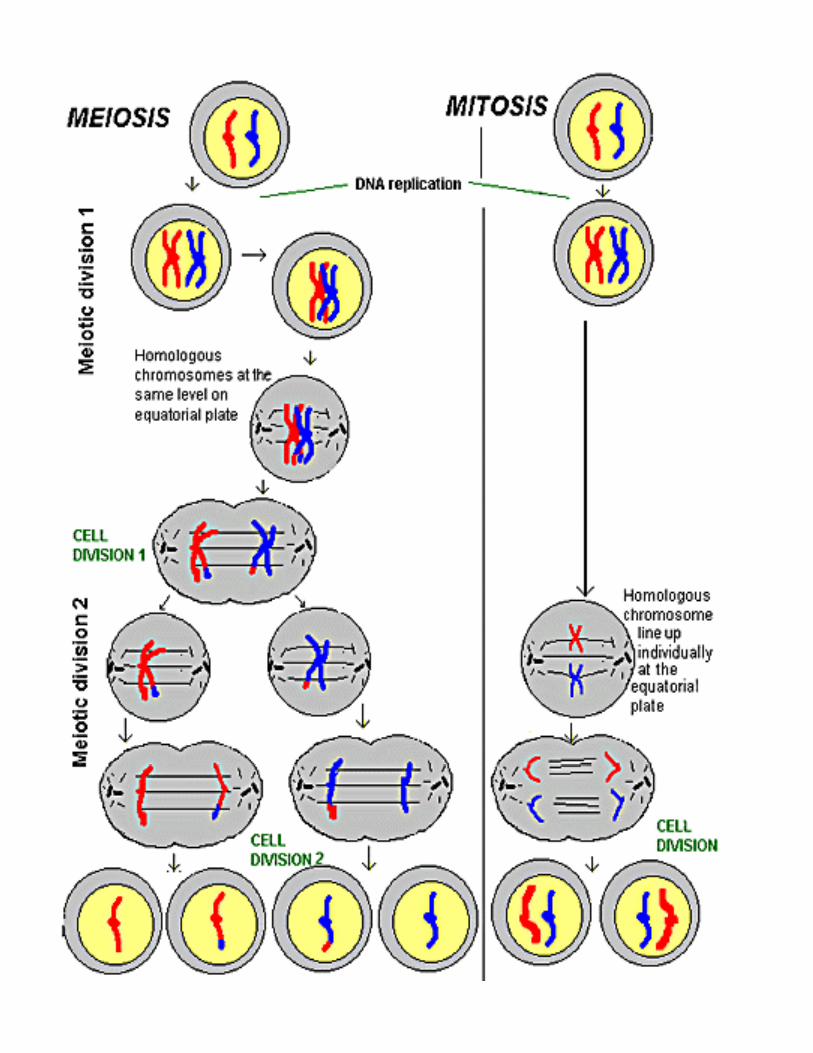

Activities of meiosis that differ from it imitosis
Pairing of homologous chromosomes
C i b t h lCrossing over between homologues
Reduction of chromosome number
Slow pace of meiotic prophase
Requirement of two cell divisions instead of one to complete the process
Lack of an S-period between the two di i idivisions

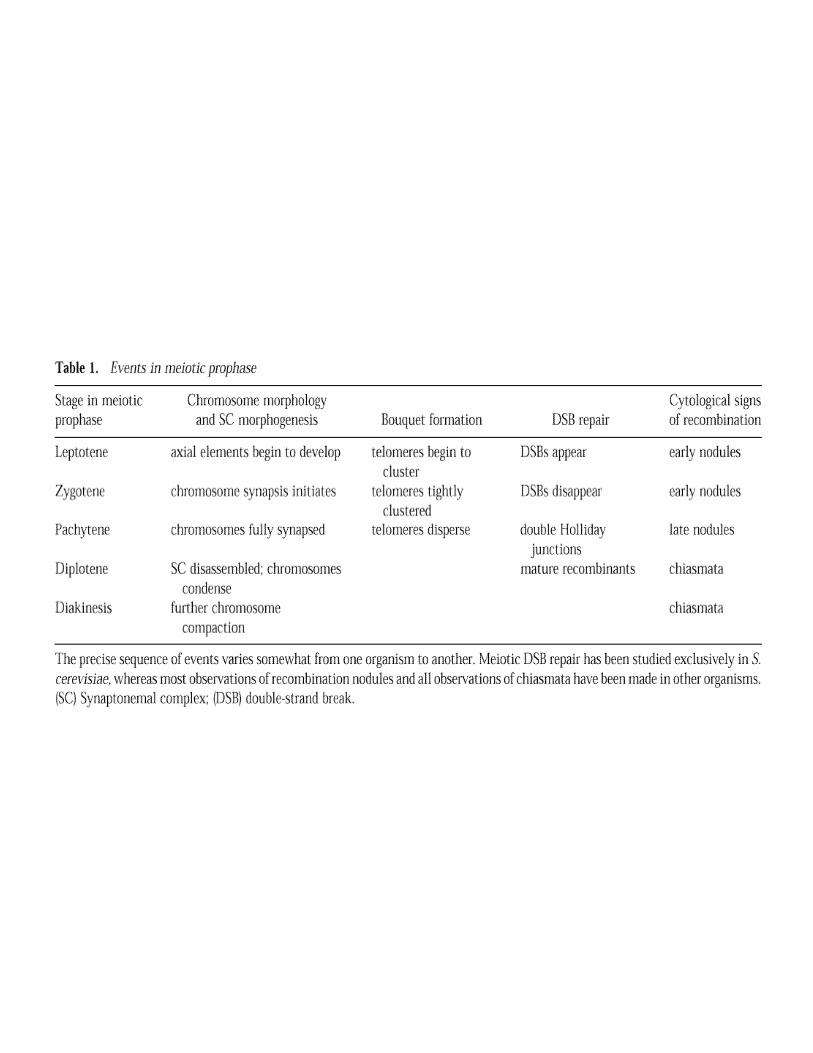
Meiotic chromosomes: it takes two to tango
G.S. Roeder
Second division of meiosis resembles mitosis
First division (reductional division) is unique
Sister chromatids remain associated Homologs behave in a coordinated fashiong
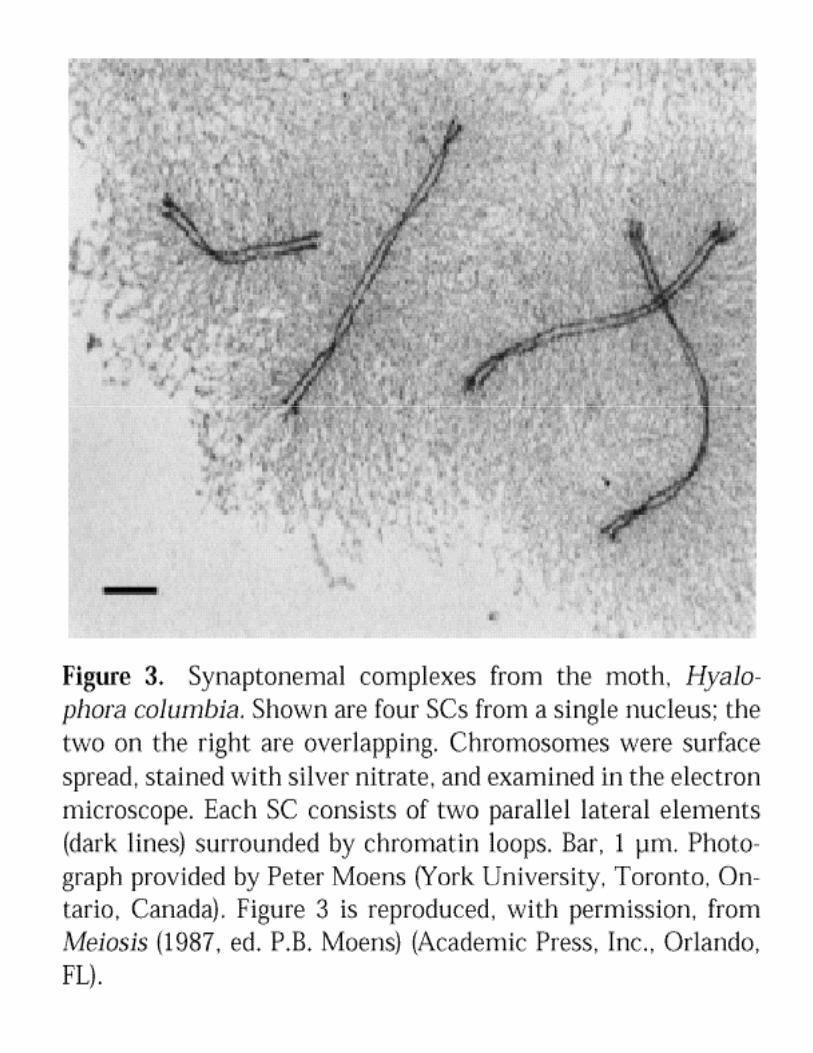
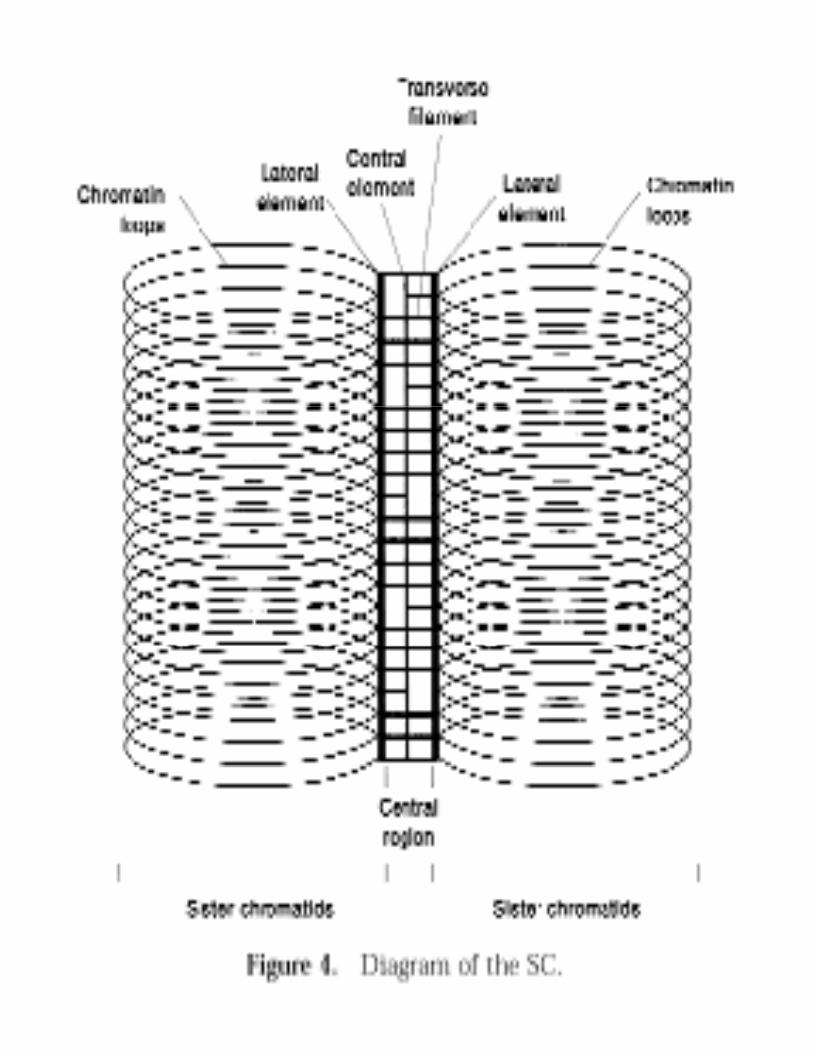
Alignment of homologsAssembly of Synaptonemal Complex (SC)Genetic RecombinationGenetic RecombinationFormation of Chiasmata


Double-stranded Break
5’3’
3’5’
Exonucleotyic Digestion
3’5’
5’3’
5’3’
3’5’
3’5’
5’3’
Strand Invasion & Loop formation
5’3’
3’5’
3’5’
5’3’
DNA repair synthesis
5’3’
3’5’
3’5’
5’3’5 3
Branch Migration
5’3’
3’5’
i ii i
o o
3’5’
5’3’
i ii i
o o

5’ 3’
o o
3’ 5’
3’5’
5’3’
i ii i
o o
5’ 3’
o o
5’3’
3’5’
3’5’
5’3’
o o
o o
5’3’
3’5’
3’5’
5’3’
5’ 3’
5 3
o o
3’ 5’
3’5’
5’3’
o, o

5’3’
3’5’
o o3’5’
5’3’
o, o
The ends of duplex are the same as the original (i.e. no recombination of loci flanking
5’3’
3’5’
i i
the point of crossover)
3’5’
5’3’
i, i
5’3’
3’5’
i3’5’
5’3’
o, i
The ends of each duplex have recombined
5’3’
3’5’
The ends of each duplex have recombined (i.e. flanking loci may have crossed over)
3’5’
5’3’
i, o

Presynaptic alignmentHomolog recognitionGenome wide search
Repetitive sequencesBouquet formation & nuclear movementBouquet formation & nuclear movement
Formation of unstable side-by-side joints
Precedes and promotes
Initiation of recombination
Double Strand Break
Recombination nodulesEarly- Leptotene to ZygoteneLate- Pachytene (correspond to Cross over events)
SynapsisCOs promote proper chromosome segregation
Chiasmata
Proper chromosome segregation

Homology dependent DSB repair models can not explaingy p p p
Integration of extrachromosomal DNA into chromosomes via mitotic recombination
Repair of DSBs in mitotic cells
Single-strand annealing model (SSA)One-sided invasion model (OSI)



Requirements of the spindle checkpoint for proper chromosome segregation in buddingproper chromosome segregation in budding
yeast meiosis
M.A. Shonn, R. McCarroll and A.W. MurrayScience 298:300
Mad2 Spindle checkpoint
Pds1 APC-anaphase inhibitor
Spo11 Promoter of meiotic recombination-Linkage of homologous chromosomeLinkage of homologous chromosome
Spo13 Protein required to prevent sister kinetochores from separating at Anaphase IAnaphase I




In the absence of checkpoint frequency of meiosis IIn the absence of checkpoint frequency of meiosis I missegregation increased
Longer chromosomes missegregated more g g gfrequently than the smaller ones (19% for the longest chromosome of yeast)
N di j ti i i dl h k i t t tNondisjunction in spindle checkpoint mutants can be prevented by delaying the onset of anaphase
In recombination defective mutants the checkpointIn recombination defective mutants the checkpoint delays the biochemical events of anaphase I (i.e. attachment of microtubules and tension)
Spindle checkpoint mutants effect chromosome missegregation in meiosis I more than meiosis II


More than 50% of angiosperms are polyploid
Autopolyploid= Arising by multiplication of a basic set of chromosomes
Diploid ancestor Tetraploid descendent
Allopolyploid= Resulting from combining related, but not completely homologous genomes
p p
not completely homologous genomes
+
Related Diploid ancestors Tetraploid descendent

In allopolyploids pairing must become restricted to parental chromosomes related by ancestral
homology
Parental chromosomes are structurally distinct or can become distinct after hybridization by rapidly rearranging themselves (structural discrimination)discrimination)
There exists a process of chromosome assortment and alignment (chromosome recognition and g ( gassociation)

Structural discriminationStructural discrimination
The genomes of maize rice barley sorghum and The genomes of maize, rice, barley, sorghum and wheat now have markedly different genome sizes and number of chromosomes since their speciation 60 million years ago
Most of the difference in genome size is a result of an expansion of repeat sequences


Structural discriminationStructural discrimination
The genes in the rice map can be grouped into sets The genes in the rice map can be grouped into sets (rice linkage segments)
Rice linkage segments also describe the genetic maps of all the chromosomes in barley, foxtail millet, maize, sorghum, sugarcane and wheat



Chromosome recognition and association
Anther sections of up to three cell layers in thickness and confocal microscopy have been used to look at three dimensional meiotic pairing of homologue


Common bread wheat, Triticum aestivum, is an allohexaploid consisting of three different genomes,
A, B, and D
T. aestivum behaves as a normal diploid with crossing over and chiasma formation restricted to ghomologous chromosomes within each genome even though sufficient homology exists to permit crossing over between homoeologous chromosome (th l t d b t )(those related by ancestary)
Suppression of paring/CO between homoeologous chromosomes is controlled by several genes (i.e.chromosomes is controlled by several genes (i.e. Ph= pairing homoeologous)

Chromosome recognition and associationg
Homologues come in contact at their centromertic repeats first ( Centromere dominance)
The process of centromere dominance is colchicine sensitive (i.e. involves centromeric microtubules of the spindle)p )
By pre-meiotic interphase virtually all the homologues in meiocytes and tapetum cells of
h t i t dwheat are associated
Wheat centromeres then migrate, the chromosomes decondense and DNA replication then occurs.decondense and DNA replication then occurs. Decondensed homologues appear to remain associated at the start of meiotic prophase.


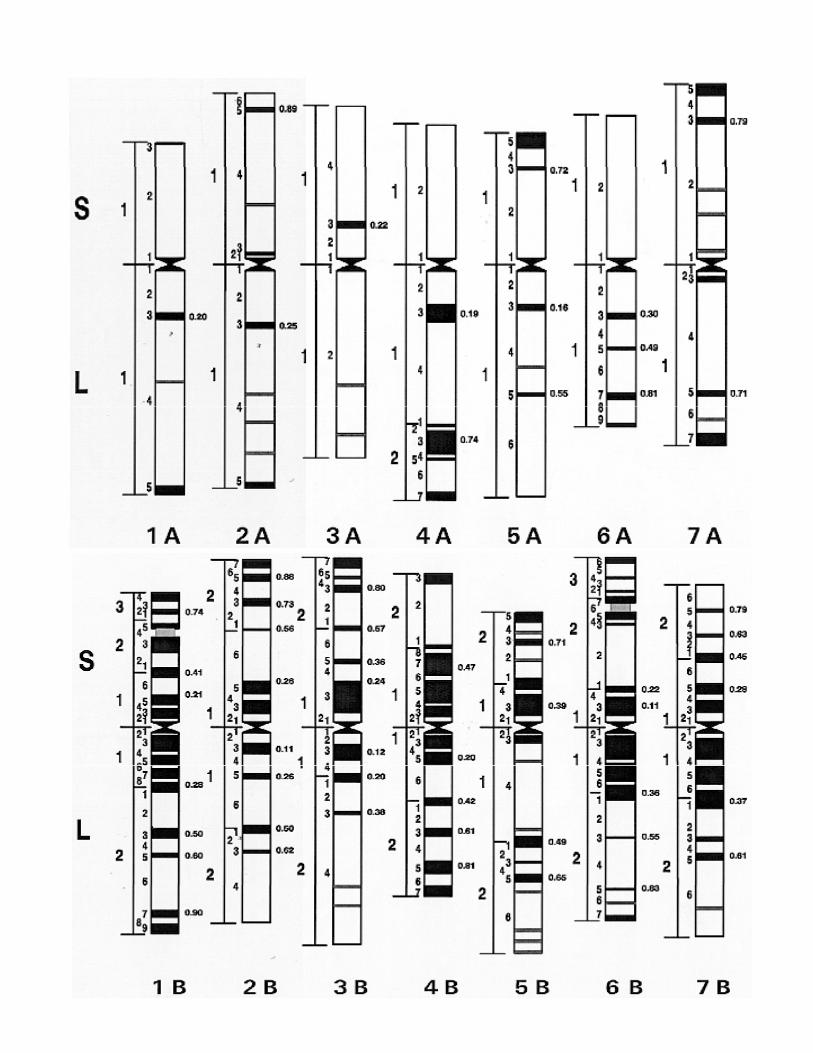
Distribution of recombination breakpointsp
Recombination does not occur randomly across the lengths of chromosomes
Gene density varies across chromosomal regions
Gene rich regions Recombination hotspotsg p


From Erayman et al. 2004Nucl. Acids Res. 32(12)

Distribution of recombination breakpointsp
Gene density varies across chromosomal regions
Gene rich regions Recombination hotspots
Centromeres are gene poorLittle recombination was observed for 40%was observed for ~40% of the chromosome region around the centromerecentromere
Genes are organized in clustersGene clusters tend to localize in distal parts oflocalize in distal parts of the chromosomes

Ch M h l & N bChromosome Morphology & Number
Karyotype describes the chromosome complement of an individual or species in terms of number, size, and morphology of its p gychromosomes
Idiogram is a diagrammatic representation of the chromosomes based on observations andchromosomes, based on observations and measurements in a number of cells
Karyotypes can be based on mitotic or meioticchromosomes and are enhanced by chromosome b di h i h l i f hbanding techniques. The largest pair of chromosomes is usually designated as number 1 and the rest are numbered in order of their relative diminishing size, to the smallest pairto the smallest pair.

Chromosome landmarks
Primary constriction The position of centromere definesPrimary constriction The position of centromere defines chromosome arms(telocentric, acrocentric, subterminal, submedian, median, metacentric)metacentric)
Secondary constriction Satellites which define theSecondary constriction Satellites which define the locations of ribosomal RNA genes
Tertiary constriction Only in some species and often correspond to the sites of cold-sensitive hetrochromatic regions

Mitotic karyotype analysisNumber of chromosomesMorphology of chromosomesHeterochromatin vs. EuchromatinChromosome BandingLabeling (i.e. in situ hybridization)
Meiotic karyotype analysisNumber of chromosomesMorphology of chromosomes (more d t il d th it ti l i )detailed than mitotic analysis)Heterochromatin vs. Euchromatin (more detailed than mitotic analysis)Chromosome BandingLabeling (i e in situ hybridization)Labeling (i.e. in situ hybridization)Chromosome pairing behaviorGene expression (in certain situations)

Metacentric Submedian S b i l
10
MedianSubmedian Subterminal
Acrocentric
Telocentric
7.5
bitr
ary
unit
s
5
omos
ome
in a
rbL
engt
h of
chr
o
0
L
0 2.5 5.0 7.5 10.01 1.7 3.0 7.050 39.5 25.0 12.5 0
Distance in arm length (d=L-S)Arm ratio (L/S)Centromeric index (Sx100/total length)
L= Long arm; S=Short arm

Ch i l d b hChemicals used to observe chromosomes
Category Chemical
Stimulation of cell Phytohemagglutinin stimulatesStimulation of cell division
Phytohemagglutinin stimulates lymphocytes to divide
Cell synchronizationHydroxyurea; Amethopterin (folic acid analog); Cold treatmentg)
Spindle dispersion and spreading of chromosomes
Colchicine, -bromonaphthalene; 8-hydroxyquinoline; hypotonic treatment (20-30% isotonic)
Staining chromosomesFeulgen (DNA-specific staining); Giemsa, Carmine

Chromosome Banding Techniques
Name BasisOrganisms
Studied
Q b di B di f AT ifi R til bi dQ-banding Banding of AT-specific fluorochromes to DNA
Reptiles, birds, mammals
G-banding Giemsa staining after incubation in warm SSC or trypsin
Fish, amphibia, reptiles, birds, yp p , ,mammals
R-banding Giemsa staining after incubation in hot buffer
Mammals
C-banding Giemsa staining after alkali treatment
Most plants and animals
Replication Incorporation of BrdU during PlantsReplication banding
Incorporation of BrdU during either early or late S phase followed by Giemsa staining
Plants, vertebrates

Ch B di T h iChromosome Banding Techniques
C-Banding Giemsa stains the satellite-rich centromeric heterochromatin. Therefore C-bands reveal the location of constitutive heterochromatin
R-Banding R bands apparently contain GC rich DNA and are the interbands in G-bandedchromosomes.c o oso es.
G-Banding Dark bands (G-bands) alternating with light bands are produced in prophase or metaphase h h G b d i f hchromosomes. The G-bands are region of the
chromosome where the DNA is presumably free to stack with the dyes in the stain.
Q-Banding AT rich DNA reacts strongly with quinacrine and enhances its fluorescence in UV while GC rich DNA quenches the fluorescence.


Lampbrush chromsomes
In the arrested diplotene (dictyoene) chromosomes expand and transcription begins. In oocytes of certain vertebrates this transcription is evident under light microscopy.


Polytene chromosomes
DNA replication takes place but mitosis is arrested at the G2 stage. The replicated chromatids of each chromosome remain attached to one centromere. This process of endoreduplicaton can lead to a p pfinal levels of 1024 DNA strands in the polytene chromosomes of Dorsophila melanogaster salivary glands.

2R-division 48
X-division 3

Telomere
Why
Complete DNA replication.
Broken chromosomes are unstable. Telomeres protect chromosomes from end to end fusion.
Features
The DNA strand running from 5’ to 3’ from the center toward the end of the chromosomes has more G residues than its complement. The G’s are clustered.
Telomeres are tandem arrays of (T/A)1-4 (G)1-8. For example (TTAGGG)n in human.
Th t f t l i DNA i ithi d b t The amount of telomeric DNA varies within and between species.

DNA synthesis is semi-discontinuous3’
continuous replication on
3’5’
leading strand
5’
Okazaki fragment synthesis on lagging strand
discontinuous replication on the lagging strand
C ti li ti l di t dContinuous replication on leading strand+
Discontinuous replication on lagging strandRNA primer formation and elongation by DNA polymerase III result in Okazaki fragment
3’synthesis on lagging strand
3’5’
Removed primer fragments
5’
Completion of an Okazaki fragment requires removal of RNA primer by DNA polymerase I. A final nick in the DNA backbone remains

DNA synthesis is semi discontinuousDNA synthesis is semi-discontinuous
DNA polymerase replicates DNA in 5’-3’ direction using 8-12 base stretch of RNA to prime DNA g psynthesis
One strand of a linear chromosome will be replicated to the very end, the other will have a 8-12 base gap generated by the removal of the primer.
Telomeres act as substrate for replication of the ends and prevent exonucleolytic degradation of chromosomes.
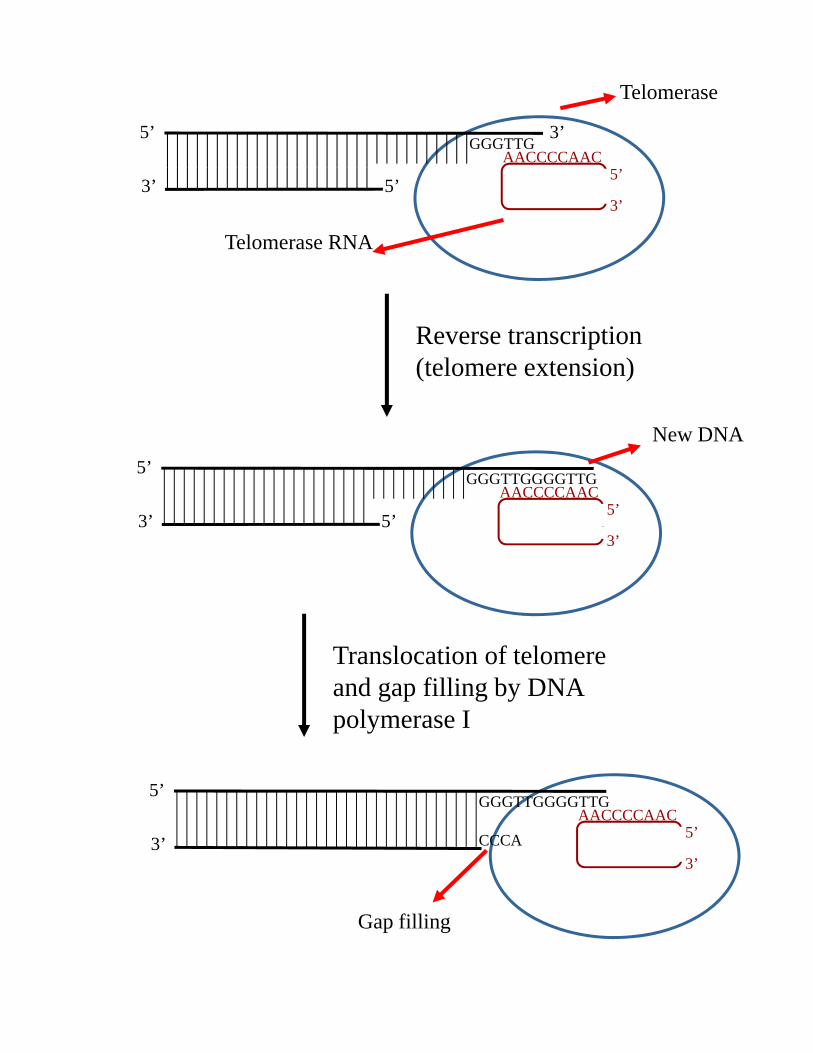
5’GGGTTG
AACCCCAAC
3’
Telomerase
3’3’
5’5’
Telomerase RNA
Reverse transcription(telomere extension)
3’
5’GGGTTGGGGTTG
AACCCCAAC5’
5’
New DNA
33’
5
l i f lTranslocation of telomere and gap filling by DNA polymerase I
3’
5’GGGTTGGGGTTG
AACCCCAAC
3’
5’CCCA
Gap filling

Telomerase
Specialized reverse transcriptase
Ribonucleoprotein with activity depending on both RNA and protein components.
Uses RNA, not DNA to template synthesis of the telomeric DNA
The expected product of telomerase mediated replication is a duplex molecule with a single strand G tail
Telomerase RNA determines the sequence of telomeric DNA
Telomerase independent mechanisms exist for telomere i t i h ( t l ti it f d imaintenance in humans (no telomerase activity found in
somatic cells).


Centromere
CEN locus- DNA sequence that specifies the centromere location
Site of formation of the kinetochore
Final locus of sister chromatid pairing in mitosis and meiosis
Cell cycle checkpoint control
DNA contains repetitive sequences
Centromeric proteins have been identified which are associated with DNA in the centromere region (CENP-A, B, C and D).

Glossary of terms
Autopolyploid: Polyploid arising from multiplication of a basic set of chromosomes. Polyploid in which all the chromosomes come from h ithe same species.
Allopolyploid: Polyploid resulting from combining related, but not completely homologous genomes. Polyploid produced by the hybridization of two or more specieshybridization of two or more species.
homoeologous chromosome : Those chromosomes related by ancestary and sharing common segments (i.e. essentially identical in gene content)gene content).
Centromere: Constrictions in eukaryotic chromosomes on which the kinetochore lies. Also, the DNA sequence within the constriction, which is responsible for appropriate function.which is responsible for appropriate function.
Telomere: The ends of linear chromosomes that are required for replication and stability.

Karyotype: Describes the chromosome complement of an
Glossary of terms
Karyotype: Describes the chromosome complement of an individual or species in terms of number, size, and morphology of its chromosomes
Idiogram: A diagrammatic representation of the chromosomes, b d b ti d t i b f llbased on observations and measurements in a number of cells
2n : The somatic chromosome number for a species.
Satellite DNA: Highly repetitive eukaryotic DNA primarily located around centromeres Satellite DNA usually has a different buoyantaround centromeres. Satellite DNA usually has a different buoyant density than the rest of the cell’s DNA.
Endoreduplication: The process by which chromosomes replicate but the replicated chromatids of each chromosome remain attached to one centromere (i.e. polytene chromosomes).
AcentricCentromereDicentricHeterochromatinHigher-order structureHolocentricIsodicentricKinetochoreMajor satelliteMinor satelliteNeocentromereNucleosomePseudodicentricSatellite

ReferencesReferences
1. G. Moore (2000) Cereal chromosome structure, evolution, and pairing. Annu. Rev. Plant Physiol. Plant Mol. Biol.. 51:195–222
2. G. Moore (1998) To pair or not to pair: chromosome pairing and evolution. Curr. Opin. Plant Bio. 1:116-122
3. P. Schnable et al. (1998) Genetic recombination in plants. ( ) pCurr. Opin. Plant Bio. 1:123-129
4. K. Gill et al. (1996) Identification and high-density mapping of gene-rich regions in chromosome group 5 of wheat. Genetics 143:1001-1012
5. K. Gill et al. (1996) Identification and high-density mapping of gene-rich regions in chromosome group 1 of wheat. G ti 144 1883 1891Genetics 144:1883-1891





























