Embed Size (px)
Citation preview

ACTION OF CMF FROM ARENICOLA MARINA 65THE JOURNAL OF EXPERIMENTAL ZOOLOGY 281:65–71 (1998)
© 1998 WILEY-LISS, INC.
Action of CMF (Coelomic Maturation Factor) onOocytes of the Polychaete Arenicola marina (L.)
G.J. WATSON* AND M.G. BENTLEYGatty Marine Laboratory, School of Environmental and EvolutionaryBiology, University of St Andrews, St Andrews, Fife, KY16 8LB, Scotland,United Kingdom
ABSTRACT Oocyte maturation in the lugworm Arenicola marina is controlled by two hor-monal steps: a prostomial maturation hormone and a coelomic maturation factor (CMF). Oocytematuration and spawning follows release of the maturation hormone from the prostomium andthe appearance of CMF activity within the coelomic fluid.
In this study, injection of female prostomial homogenate into gravid females induced the produc-tion of CMF after approximately 1 to 2 h. Once induced, CMF activity remained for 1 to 4 h. Titres ofCMF assayed in vitro between females also varied. CMF in some females induced 100% of oocytes tomature, whereas CMF removed from others induced only about 30% maturation. Oocytes requiredan exposure time of approximately 20 min to CMF before maturation is able to proceed indepen-dently. Optimal CMF activity in vitro was observed at temperatures at or below 10°C.
CMF has already been shown to be thermolabile and have a molecular mass greater than 10kDa. CMF is shown here to be trypsin sensitive and to have a molecular mass of greater than 30kDa, suggesting a proteinaceous nature. J. Exp. Zool. 281:65�71, 1998. © 1998 Wiley-Liss, Inc.
In comparison to other invertebrate groups, suchas molluscs and echinoderms, the control of re-production of polychaetes is still poorly under-stood. Relatively little progress has been made inthe isolation and purification of oocyte matura-tion-inducing substances from polychaetes. To dateno substance has been completely characterised,either in terms of its structure or action. Consid-erable advances have been made in recent yearsin the control of sperm maturation and activa-tion in Arenicola marina (see Bentley and Pacey,’89; Bentley et al., ’90; Pacey and Bentley, ’92a,b;Bentley and Hardege, ’96). Since the pioneeringwork by Howie (’61), Meijer and Durchon (’77) andMeijer (’79a,b; ’80), however, the control of oocytematuration has not been investigated further indetail until recently (Watson and Bentley, ’97). InA. marina, CMF production can only be inducedby injection of prostomial homogenate. Simple invitro incubation of oocytes with prostomial homog-enate does not induce maturation (Watson andBentley, ’97).
Oocytes of A. marina are arrested at prophaseof meiosis I, and maturation involves the progres-sion to metaphase I, at which stage oocytes arespawned and fertilized (Howie, ’61). The develop-ment of an in vitro assay for maturation byWatson and Bentley (’97) has permitted the as-sessment of the presence of maturation inducing
activity of coelomic fluid. Work has shown thatoocytes require two steps to induce maturation. Asubstance from the prostomium induces theproduction of a thermolabile substance with a mo-lecular mass of greater than 10 kDa in the coelo-mic fluid, termed the Coelomic Maturation Factor(CMF), which acts on the oocyte (Watson andBentley, ’97). This bears some similarities to thecontrol of oocyte maturation in asteroid echino-derms by Gonad Stimulating Substance (GSS) andMeiosis-Inducing Substance (MIS) (1-methyl ad-enine) (for a review, see Meijer and Mordet, ’94).Studies on the control of oocyte maturation inechinoderms have progressed much further thanthose in polychaetes. The MIS in starfish, possi-bly analogous in function to CMF in A. marina,was first identified as 1-MeAde in the late 1960s(Kanatani et al., ’69), and its mode of action isnow well understood. The identification of CMFand the use of the fluorescence assay for oocytematuration in A. marina has enabled us to inves-tigate further oocyte maturation in this species,
Grant sponsor: Biotechnology and Biology Sciences Research Coun-cil; Grant sponsor: Natural Environment Research Council; Grantnumber: GR3/8628.
*Correspondence to: Dr. Gordon J. Watson, Gatty Marine Labora-tory, School of Environmental and Evolutionary Biology, Universityof St Andrews, St Andrews, Fife, KY16 8LB, Scotland, U.K.
Received 28 July 1997; Accepted 15 December 1997

66 G.J. WATSON AND M.G. BENTLEY
and to compare and contrast the processes ofmaturation in A. marina with those of, for ex-ample, echinoderms.
MATERIALS AND METHODSSexually mature worms were collected and
maintained as described previously (Watson andBentley, ’97). All techniques for the induction ofCMF by injection of female prostomial homoge-nate and subsequent harvesting, and the in vitroassessment of the presence of CMF in coelomicfluid using oocytes from other females (oocyte do-nors) in conjunction with the fluorescence assaywere as described previously (Watson and Bentley,’97). Oocytes were examined for maturation us-ing either unfixed Hoechst 33342 dyed material,or fixed Hoechst 33258 dyed material whensamples could not all be examined within a 12-hperiod. All chemicals were purchased from SigmaChemical Co. (St. Louis, MO).
The timing of production of CMF afterinjection of prostomial homogenate
Animals were collected in 1994 and 1995 for twoseparate experiments that were identical except thatthe number of females injected with prostomial ho-mogenate and assayed for CMF activity in 1995 wasincreased from five to ten. On each occasion, threefemales were used to provide oocytes for the assayof CMF (maturation inducing) activity.
Prior to injection with prostomial homogenate(to initiate CMF production), 150 µl of coelomicfluid was removed from each female. Additional150 µl samples of coelomic fluid were removedfrom each female at 0.5, 1, 2, 3, 4, and 5 h post-injection. All coelomic fluid samples removed wereassessed for CMF activity independently with oo-cytes from the three oocyte donors. The oocyteswere then fixed in Hoechst 33258 for subsequentexamination.
On each occasion that coelomic fluid was har-vested from a female, a sample of the female’sown oocytes, taken from the coelomic fluid, wasfixed immediately and then assessed for theirmaturational state.
Exposure time required for CMF to induceoocyte maturation
To determine the in vitro exposure time to CMFrequired by oocytes before maturation can proceedindependently, oocytes were incubated in CMF for5, 10, 20, 30, 40, 50, 60, 80, and 100 min. In eachcase, 5 µl of oocytes from each of five oocyte do-nors were incubated in 50 µl of coelomic fluid with
CMF. After incubation at the times stated aboveat 10°C under ambient illumination, oocytes wereremoved and washed three times in twice filtered(0.2 µm) seawater (TFSW). Oocytes were then in-cubated for 3 h in TFSW to complete any matu-rational processes that had been initiated.
Oocytes were incubated in coelomic fluid withCMF activity and also TFSW for 3 h at 10°C un-der ambient illumination, for positive and nega-tive controls respectively. Unfixed oocytes from thetreatments and the positive and negative controlswere then assessed for maturation.
Effects of temperature on theactivity of CMF
Oocytes (3 µl) from each of six donors were eachincubated in 50-µl aliquots of coelomic fluid withCMF activity (coelomic fluid was assayed for CMFactivity immediately after harvesting). Remain-ing coelomic fluid samples were incubated at dif-ferent temperatures (0.5, 5, 10, 15, 22, 37°C) for1 h under ambient illumination. Each was thenassayed for CMF activity. Oocytes were incubateddirectly in TFSW as a control for spontaneousmaturation.
Assessment of CMF activity afterincubation with trypsin
Oocytes (3 µl oocytes) from five donors were usedto assess CMF activity of the coelomic fluid aftereach treatment. Oocytes were also incubated di-rectly in TFSW as a control. The coelomic fluid frac-tions were incubated with the treatments at 22°Cunder ambient illumination. To ensure completemixing, each coelomic fluid sample was shaken for10 sec after the addition of each solution.
Treatment 1Twenty-five percent of the coelomic fluid with
CMF activity (250 µl) was incubated for 30 minunder ambient illumination and 22°C (positivecontrol).
Treatment 2Twenty-five percent of the coelomic fluid with
CMF activity (250 µl) was incubated with 10 µlof trypsin solution (100 mg · ml–1 trypsin in TFSW(Type: II-S Porcine Pancreas 1,000–2,000 BAEEunits · mg solid) for 15 min at 22°C. To this wasadded 10 µl of trypsin inhibitor solution (100 mg· ml–1 trypsin inhibitor in TFSW (Type II-S: Soy-bean, 1 mg will inhibit 1.6 mg of trypsin with10,000 BAEE units) and incubated for a further15 min at 22°C.

ACTION OF CMF FROM ARENICOLA MARINA 67
Treatment 3Twenty-five percent of the coelomic fluid (250
µl) with CMF activity was incubated with 10 µlof trypsin inhibitor solution for 30 min at 22°Cunder ambient illumination.
Treatment 4Twenty-five percent of the active coelomic fluid
(250 µl) with CMF activity was incubated with10 µl of trypsin/trypsin inhibitor solution (100 mgtrypsin, 100 mg trypsin inhibitor in 1 ml of TFSWmade up 5 min before use) for 30 min at 22°Cunder ambient illumination.
Assessment of CMF activity afterultrafiltration
Three treatment groups were used. In each, oo-cytes from six oocyte donors were incubated in 50µl of the treatments at 10°C under natural illu-mination. Oocytes were also incubated in TFSWfor 3 h for a negative control. Unfixed oocytes fromthe treatments and the positive and negative con-trols were then assessed for maturation.
Treatment 1Fifty percent of the coelomic fluid with CMF
activity was incubated with prophase oocytes.
Treatment 2Coelomic fluid with CMF activity was filtered
through a 30-kDa membrane filter at 5°C for 20to 30 min. The filter system used was a StirredUltrafiltration Cell 8050 on a magnetic stirrerwith a 30 kDa Diaflo® membrane Ultrafilter(Amicon Inc., Beverly, MA) using dry nitrogen ata pressure of 3.7 atmospheres. Oocytes were in-cubated in the filtrate.
Treatment 3Oocytes were incubated in the remaining coelo-
mic fluid that did not pass through the filter (ap-proximately 25% of the total amount of coelomicfluid). Unfixed oocytes from the treatments, andpositive and negative controls were then assessedfor maturation.
Statistical analysisTwo-way analysis of variance on arcsine-trans-
formed data using the statistical package Minitabver. 8.2 was used for data on effects of tempera-ture on CMF activity. A multiple-comparisonTukey test was used for pairwise comparisons.
The data from the exposure time, trypsin treat-ment, and ultrafiltration experiments required
statistical analysis using Generalised Linear Inter-active Modelling (GLIM Version 3.77). This allowsthe distribution of non-normal data to be specifiedfor analysis of variance. Interaction effects were ex-cluded from the analyses, and further analysis ofthe data using Student t-tests were performed forexposure time only because the nature of the otherdata resulted in overparameterization within themodel. Comparisons of differences between indi-vidual treatments for trypsin treatment and ultra-filtration experiments were therefore carried out bygraphical inspection only.
RESULTSThe production of CMF following injection of
prostomial homogenate in female Arenicola ma-rina is shown in Figure 1. Of 15 individuals thatwere injected, 12 showed CMF activity and matu-ration of their own oocytes in vivo (a–l). The re-maining three individuals from 1995 did notgenerate CMF, their oocytes did not undergomaturation in vivo and they did not spawn. It canbe concluded, therefore, that these individualswere not at a state of spawning readiness. Figure1 shows that in three individuals (a–c), CMF ac-tivity could be detected in coelomic fluid 0.5 h af-ter injection of prostomial homogenate into theworms. After 1 h, an additional four animals (d–g) had generated CMF, and by 2 h, all 12 indi-
Fig. 1. Number of females (a–l) with CMF activity presentin their coelomic fluid prior to the initiation of spawning attime 0, and at 0.5, 1, 2, 3, 4, and 5 h after spawning hadbeen initiated in each female by injection of one homogenisedprostomial equivalent. Females have been ranked for onsetof CMF activity.
68 G.J. WATSON AND M.G. BENTLEY
viduals had CMF activity present in their coelo-mic fluid. After this time, CMF activity declinedsuch that after 3 h it was no longer present infive individuals (c, g, j, k, and l) and after 4 h ineight individuals (c, f–l). CMF activity was lostfrom all females after 5 h.
The persistence of CMF activity varies consid-erably between females. Figure 1 shows that intwo individuals (a and b), it could be harvestedfrom the coelom for a period of 3.5 h. In contrastit could be harvested from three of the females(j–k) only for a short time around 2 h after injec-tion of prostomial homogenate. Although the co-elomic fluid of a given female induced maturationof all oocytes deriving from that female, not alloocytes from other females incubated in the samecoelomic fluid underwent maturation.
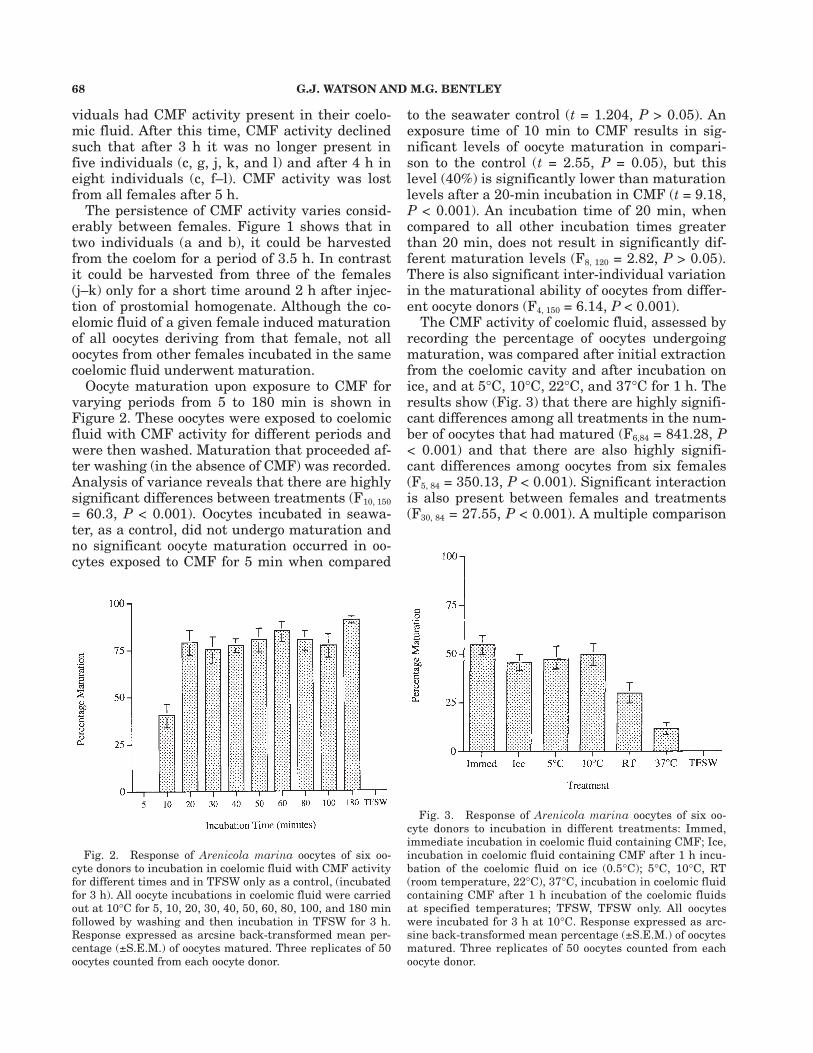
Oocyte maturation upon exposure to CMF forvarying periods from 5 to 180 min is shown inFigure 2. These oocytes were exposed to coelomicfluid with CMF activity for different periods andwere then washed. Maturation that proceeded af-ter washing (in the absence of CMF) was recorded.Analysis of variance reveals that there are highlysignificant differences between treatments (F10, 150= 60.3, P < 0.001). Oocytes incubated in seawa-ter, as a control, did not undergo maturation andno significant oocyte maturation occurred in oo-cytes exposed to CMF for 5 min when compared
to the seawater control (t = 1.204, P > 0.05). Anexposure time of 10 min to CMF results in sig-nificant levels of oocyte maturation in compari-son to the control (t = 2.55, P = 0.05), but thislevel (40%) is significantly lower than maturationlevels after a 20-min incubation in CMF (t = 9.18,P < 0.001). An incubation time of 20 min, whencompared to all other incubation times greaterthan 20 min, does not result in significantly dif-ferent maturation levels (F8, 120 = 2.82, P > 0.05).There is also significant inter-individual variationin the maturational ability of oocytes from differ-ent oocyte donors (F4, 150 = 6.14, P < 0.001).
The CMF activity of coelomic fluid, assessed byrecording the percentage of oocytes undergoingmaturation, was compared after initial extractionfrom the coelomic cavity and after incubation onice, and at 5°C, 10°C, 22°C, and 37°C for 1 h. Theresults show (Fig. 3) that there are highly signifi-cant differences among all treatments in the num-ber of oocytes that had matured (F6,84 = 841.28, P< 0.001) and that there are also highly signifi-cant differences among oocytes from six females(F5, 84 = 350.13, P < 0.001). Significant interactionis also present between females and treatments(F30, 84 = 27.55, P < 0.001). A multiple comparison
Fig. 2. Response of Arenicola marina oocytes of six oo-cyte donors to incubation in coelomic fluid with CMF activityfor different times and in TFSW only as a control, (incubatedfor 3 h). All oocyte incubations in coelomic fluid were carriedout at 10°C for 5, 10, 20, 30, 40, 50, 60, 80, 100, and 180 minfollowed by washing and then incubation in TFSW for 3 h.Response expressed as arcsine back-transformed mean per-centage (±S.E.M.) of oocytes matured. Three replicates of 50oocytes counted from each oocyte donor.
Fig. 3. Response of Arenicola marina oocytes of six oo-cyte donors to incubation in different treatments: Immed,immediate incubation in coelomic fluid containing CMF; Ice,incubation in coelomic fluid containing CMF after 1 h incu-bation of the coelomic fluid on ice (0.5°C); 5°C, 10°C, RT(room temperature, 22°C), 37°C, incubation in coelomic fluidcontaining CMF after 1 h incubation of the coelomic fluidsat specified temperatures; TFSW, TFSW only. All oocyteswere incubated for 3 h at 10°C. Response expressed as arc-sine back-transformed mean percentage (±S.E.M.) of oocytesmatured. Three replicates of 50 oocytes counted from eachoocyte donor.
ACTION OF CMF FROM ARENICOLA MARINA 69
(Tukey Test) was performed, revealing significantdifferences (P < 0.001) between CMF activity ofcoelomic fluid incubated at 22°C and 37°C. Thereare also significant differences between these twotreatments and all other treatments. CMF activ-ity is, therefore, lost progressively following incu-bation at 22°C and 37°C.
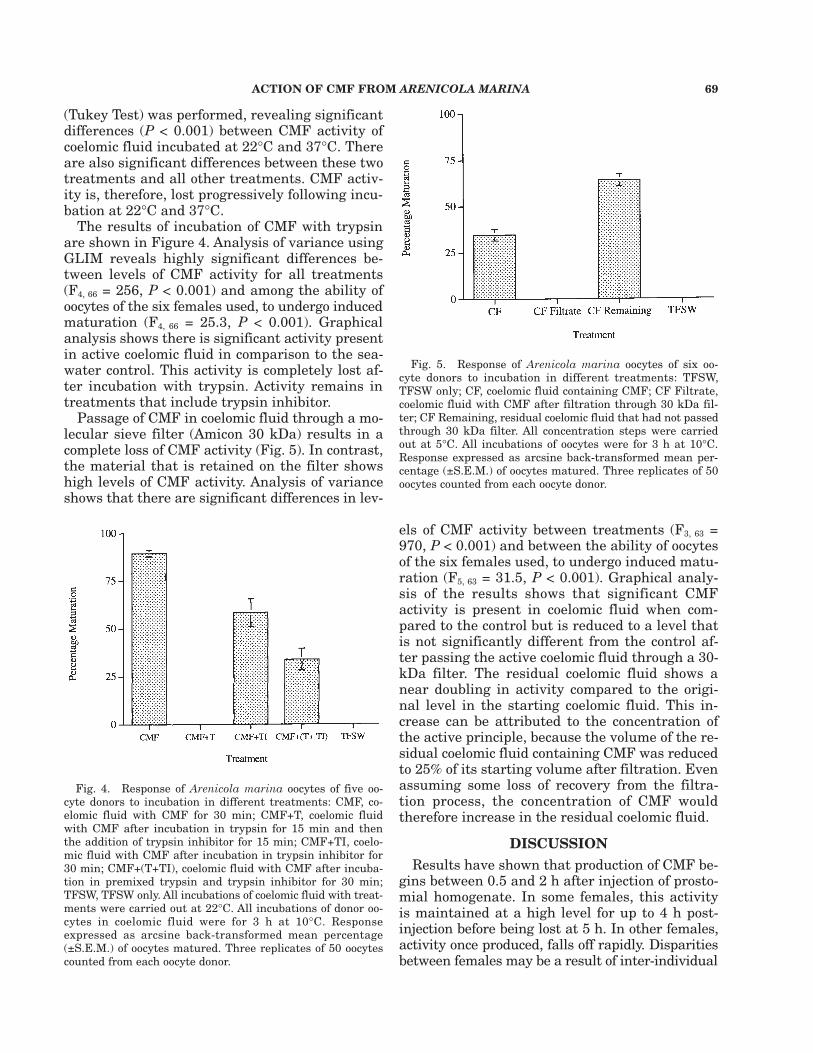
The results of incubation of CMF with trypsinare shown in Figure 4. Analysis of variance usingGLIM reveals highly significant differences be-tween levels of CMF activity for all treatments(F4, 66 = 256, P < 0.001) and among the ability ofoocytes of the six females used, to undergo inducedmaturation (F4, 66 = 25.3, P < 0.001). Graphicalanalysis shows there is significant activity presentin active coelomic fluid in comparison to the sea-water control. This activity is completely lost af-ter incubation with trypsin. Activity remains intreatments that include trypsin inhibitor.
Passage of CMF in coelomic fluid through a mo-lecular sieve filter (Amicon 30 kDa) results in acomplete loss of CMF activity (Fig. 5). In contrast,the material that is retained on the filter showshigh levels of CMF activity. Analysis of varianceshows that there are significant differences in lev-
els of CMF activity between treatments (F3, 63 =970, P < 0.001) and between the ability of oocytesof the six females used, to undergo induced matu-ration (F5, 63 = 31.5, P < 0.001). Graphical analy-sis of the results shows that significant CMFactivity is present in coelomic fluid when com-pared to the control but is reduced to a level thatis not significantly different from the control af-ter passing the active coelomic fluid through a 30-kDa filter. The residual coelomic fluid shows anear doubling in activity compared to the origi-nal level in the starting coelomic fluid. This in-crease can be attributed to the concentration ofthe active principle, because the volume of the re-sidual coelomic fluid containing CMF was reducedto 25% of its starting volume after filtration. Evenassuming some loss of recovery from the filtra-tion process, the concentration of CMF wouldtherefore increase in the residual coelomic fluid.
DISCUSSIONResults have shown that production of CMF be-
gins between 0.5 and 2 h after injection of prosto-mial homogenate. In some females, this activityis maintained at a high level for up to 4 h post-injection before being lost at 5 h. In other females,activity once produced, falls off rapidly. Disparitiesbetween females may be a result of inter-individual
Fig. 4. Response of Arenicola marina oocytes of five oo-cyte donors to incubation in different treatments: CMF, co-elomic fluid with CMF for 30 min; CMF+T, coelomic fluidwith CMF after incubation in trypsin for 15 min and thenthe addition of trypsin inhibitor for 15 min; CMF+TI, coelo-mic fluid with CMF after incubation in trypsin inhibitor for30 min; CMF+(T+TI), coelomic fluid with CMF after incuba-tion in premixed trypsin and trypsin inhibitor for 30 min;TFSW, TFSW only. All incubations of coelomic fluid with treat-ments were carried out at 22°C. All incubations of donor oo-cytes in coelomic fluid were for 3 h at 10°C. Responseexpressed as arcsine back-transformed mean percentage(±S.E.M.) of oocytes matured. Three replicates of 50 oocytescounted from each oocyte donor.
Fig. 5. Response of Arenicola marina oocytes of six oo-cyte donors to incubation in different treatments: TFSW,TFSW only; CF, coelomic fluid containing CMF; CF Filtrate,coelomic fluid with CMF after filtration through 30 kDa fil-ter; CF Remaining, residual coelomic fluid that had not passedthrough 30 kDa filter. All concentration steps were carriedout at 5°C. All incubations of oocytes were for 3 h at 10°C.Response expressed as arcsine back-transformed mean per-centage (±S.E.M.) of oocytes matured. Three replicates of 50oocytes counted from each oocyte donor.

70 G.J. WATSON AND M.G. BENTLEY
differences but could also be from other sources. Thepresence of blood in the coelomic fluid that has re-sulted from injury could interfere with CMF. Thecoelomic fluid cavity of Arenicola marina containsvariable numbers of coelomocytes (Bentley andPacey, ’89) that are especially prevalent post-spawning. These cells are amoeboid, phagocyticcells associated with removal of material from thecoelomic fluid. An increase in coelomocytes or theiractivation by tissue damage may affect CMF lev-els. It is possible that as worms are increasinglydamaged by the sequential sampling, cellular re-sponses may result in CMF being broken down,resulting in a rapid loss of activity.
One of the most interesting observations re-ported here, is that CMF activity in vitro doesnot produce an all-or-nothing response. Rather, aproportion of the available oocytes undergo matu-ration, and this may vary from 30% to 100%. Thissuggests that either the threshold concentrationvaries between oocytes from a single donor or theavailable amount of CMF may be limiting. Onepossible reason for the reduction in levels of CMFis that its production induces maturation in thefemales’ own oocytes and this may deplete exist-ing coelomic CMF.
Although the possible sources of variation in thestarting levels of CMF activity are numerous, in agiven experiment, initial CMF levels and oocytesused to assay maturation are from the samesources. Variation in CMF activity, therefore, hadno effect on the variability of the observed responsebetween treatments. Clearly an understanding ofthe precise mechanism by which CMF inducesmaturation and the fate of CMF will enable a fullinterpretation of CMF titres and action.
In vitro maturation of an oocyte in response toCMF, as with in vivo maturation, results in itsprogression to full metaphase I arrest (Watson andBentley, ’97). Recent work (Watson and Bentley,’98) has shown that in vitro maturation producesa metaphase spindle of equivalent size to in vivomatured oocytes and these oocytes are also fertil-izable. Post-fertilization development to blastulaappears normal in embryos produced from in vitroor in vivo maturation (pers. obs.). However, thedevelopment of in vitro matured oocytes appearsto proceed slower during the first 5 h (Watson andBentley, ’98).
To achieve maximum maturation levels in oo-cytes of A. marina, a minimum exposure time toCMF of 20 min at 10°C has been shown to be re-quired. This can be compared to other marine in-vertebrates, but only in starfish can the MIS,
1-MeAde, be shown to have a similar role as CMFin that it is a second endocrine step. Both A. ma-rina and starfish have a hormone that stimulatesthe production of CMF and 1-MeAde, respectively.In starfish, the exposure time for 1-MeAde re-quired to induce oocyte maturation has beentermed the hormone-dependent period (HDP)(Kanatani et al., ’69). The HDP is temperature-de-pendent; it increases as temperature decreases(Dorée and Guerrier, ’75). In Marthasterias glacialis,the maximum percentage maturation at 24°C isreached after 18 min (Guerrier and Dorée, ’75). Thistime is approximately the same for CMF; however,comparisons should be interpreted with caution.This is because of the probable differences in themechanisms of maturation and the differences inincubation temperature. There are also widevariations in the HDP between batches of oocytesin starfish that may also affect any comparisonsbetween A. marina and other systems (Chiba andHoshi, ’89).
The results shown above (Fig. 3) have indicatedthat maximal CMF activity remains after incu-bation at temperatures of 10°C or below. CMF ac-tivity is therefore most stable at a physiologicaltemperature for A. marina. Any increase in tem-perature results in loss of CMF activity and thisis reflected in the effects of higher temperatureson the metabolic processes of the whole worm. Inpopulations that spawn in autumn and early win-ter, maintaining worms at greater than 10°C inthe laboratory results in high mortality rates incomparison to worms maintained at 10°C or be-low (pers. obs.).
That CMF has a molecular mass of greater than30 kDa, and its thermolability coupled with itscomplete inactivation of CMF by trypsin suggesta proteinaceous nature. Although CMF activity ispartially inhibited by the addition of trypsin in-hibitor only, there is not a complete loss of activ-ity as is the case with just trypsin.
A two-step process, not dissimilar to that in A.marina, has been described for the control of oo-cyte maturation in starfish, where the matura-tion inducer is 1-methyl adenine (Kanatani et al.,’69). CMF, like 1-methyl adenine (1-MeAde) instarfish, is produced by the action of a differentsubstance from another source. In starfish, a 2-kDa peptide hormone, Gonad Stimulating Sub-stances (GSS) acts on the follicle cells surroundingthe oocytes which then produce 1-MeAde de novo(Kanatani, ’69, ’72; Kanatani et al., ’69; Kanataniand Shirai, ’70, ’72; Schuetz, ’72; Shirai andKanatani, ’72; Hirai et al., ’73). In species from

ACTION OF CMF FROM ARENICOLA MARINA 71
other phyla, such as the mollusc Spisula solidis-sima, the amine serotonin was confirmed as thematuration inducer (Hirai et al., ’84). In the hy-drozoan, Hydractinia echinata, a peptide isthought to be involved (Freeman, ’87) whilst inthe polychaete Sabellaria alveolata a number ofneutral serine proteases have been shown to in-duce maturation (Peaucellier, ’77). The chemicalstructure of substances shown to induce oocytematuration is clearly diverse. The control of oo-cyte maturation in the lugworm A. marina is aninteresting system for study both in terms of thebiochemical nature of CMF and the parallel in-vestigations of in vitro and in vivo matured oo-cytes all of which require further investigation.
ACKNOWLEDGMENTSThe authors thank those individuals who helped
collect and maintain the specimens. The authorsalso acknowledge the support of NERC grant GR3/8628 to M.G.B. and the award of a BBSRCstudentship to G.J.W.
LITERATURE CITEDBentley, M.G., and J.D. Hardege (1996) The role of a fatty
acid hormone in the reproduction of the polychaete Arenicolamarina (L.) Invert. Reprod. Dev., 30:159–165.
Bentley, M.G., and A.A. Pacey (1989) A scanning electron mi-croscope study of sperm development and activation inArenicola marina (L.) (Annelida: Polychaeta). Invert.Reprod. Dev., 15:211–219.
Bentley, M.G., S. Clark, and A.A. Pacey (1990) The role ofarachidonic acid and eicosatrienoic acids in the activationof spermatozoa in Arenicola marina (L.) (Annelida: Poly-chaeta). Biol. Bull., 178:1–9.
Chiba, K., and M. Hoshi (1989) Activation of starfish oo-cytes modifies their hormone dependent period for 1-methyladenine in meiosis reinitiation. Dev. Growth Differ.31:453–458.
Dorée, M., and P. Guerrier (1975) Site of action of 1-methyladenine in inducing oocyte maturation in starfish:kinetic evidence for receptors localized on the cell mem-brane. Exp. Cell Res. 96:296–300.
Freeman, G. (1987) The role of oocyte maturation in the on-togeny of the fertilization site in the hydrozoan Hydractiniaechinata. Roux’s Arch. Dev. Biol. 196:83–92.
Guerrier, P., and M. Dorée (1975) Hormonal control ofreinitiation of meiosis in starfish. The requirement of 1-methyladenine during nuclear maturation. Dev. Biol.47:341–348.
Hirai, S., K. Chiba, and H. Kanatani (1973) Role of folliclecells in maturation of starfish oocytes. Dev. Growth Differ.,15:21–31.
Hirai, S., T. Kishimoto, S.S. Koide, and H. Kanatani (1984)Serotonin induction of spawning and oocyte maturation inSpisula. Biol. Bull., 167:518.
Howie, D.I.D. (1961) The spawning of Arenicola marina (L.)III. Maturation and shedding of the ova. J. Mar. Biol. Assoc.U.K., 41:771–783.
Kanatani, H. (1969) Induction of spawning and oocyte matu-ration by 1-methyladenine in starfish. Exp. Cell Res.,57:333–337.
Kanatani, H. (1972) Adenine derivatives and oocyte matura-tion in starfishes. In: Oogenesis. J.D. Biggers and A.W.Schuetz, eds. University Park Press, Baltimore, Md., pp.459–478.
Kanatani, H., and H. Shirai (1970) Mechanism of starfishspawning. III. Properties and action of meiosis-inducing sub-stance produced in the gonad under influence of gonad-stimulating substance. Dev. Growth Differ., 12:119–140.
Kanatani, H. and H. Shirai (1972) On the maturation-induc-ing substance produced in starfish gonad by a neural sub-stance. Gen. Comp. Endocrinol., 3:571–579.
Kanatani, H., H. Shirai, K. Nakanishi, and T. Kurokawa(1969) Isolation and identification of meiosis-inducingsubstance in starfish Asterias amurensis. Nature,221:273–274.
Meijer, L. (1979a) Hormonal control of oocyte maturationin Arenicola marina L. (Annelida, Polychaeta). I. Mor-phological study of oocyte maturation. Dev. Growth Dif-fer., 21:303–314.
Meijer, L. (1979b) Hormonal control of oocyte maturation inArenicola marina L. (Annelida, Polychaeta). II. Maturationand Fertilization. Dev. Growth Differ., 21:315–329.
Meijer, L. (1980) Hormonal control of oocyte maturation inArenicola marina L. (Annelida, Polychaeta). III. Involve-ment of Ca2+ and -SH groups in meiosis reinitiation. Dev.Growth Differ., 22:33–38.
Meijer, L., and M. Durchon (1977) Contrôle neurohormonalde la maturation ovocytaire chez Arenicola marina(Annélide Polychète). Etude in vitro. C. R. Acad. Sci.Paris, 285:377–380.
Meijer, L., and G. Mordet (1994) Starfish oocyte maturation:from prophase to metaphase. Sem. Dev. Biol., 5:165–171.
Pacey, A.A., and M.G. Bentley (1992a) The fatty acid 8, 11,14-eicosatrienoic acid induces spawning in the male lug-worm Arenicola marina. J. Exp. Biol., 173:165–179.
Pacey, A.A., and M.G. Bentley (1992b) An ultrastructuralstudy of spermatogenesis and sperm morula breakdown inArenicola marina (L.) (Annelida: Polychaeta). Helgol.Meeresunters., 46:185–199.
Peaucellier, G. (1977) Initiation of meiotic maturation by spe-cific proteases in oocytes of the polychaete annelid, Sabel-laria alveolata. Exp. Cell Res., 106:1–14.
Schuetz, A.W. (1972) Activation of 1-methyladenosine by star-fish oocytes. Exp. Cell Res., 66:5–10.
Shirai, H., and H. Kanatani (1972) 1-methyladenosineribohydrolase in the starfish ovary and its relation to oo-cyte maturation. Exp. Cell Res., 75:79–88.
Watson, G.J. and M.G. Bentley (1997) Evidence for a coelo-mic maturation factor controlling oocyte maturation in thepolychaete Arenicola marina (L.). Invert. Reprod. Dev.,31:297–306.
Watson, G.J., and M.G. Bentley (1998) Oocyte matura-tion and post-fertilization development of Arenicola ma-rina (L.) (Annelida: Polychaeta). Invert. Reprod. Dev.,33:35–46.

















![Cmf access to_finance_rural_powerpoint_march14_final[1]](https://img.pdfslide.us/doc/110x75/5565c8c7d8b42a5b488b5041/cmf-access-tofinanceruralpowerpointmarch14final1.jpg)











