Embed Size (px)
Citation preview

ACTA BIOLOGICA CRACOVIENSIA Series Botanica 41: 143-150,1999
KARYOTYPE STRUCTURE AND INTERPHASE CHROMATIN ORGANIZATION IN RHOEO SPATHACEA (SW.) STEARN
(COMMELINACEAE)
HIERONIM GOLCZYK1 AND ANDRZEJ JOACHIMIAK1,2*
Department of Plant Cytology and Embryology, Jagiellonian University, ul. Grodzka 52, 31-044 Cracow, Poland
2Cytogenetics Group in the Department of Plant Breeding and Seed Science, The Agricultural University of Cracow, ul. Lobzowska 24, 31-140 Cracow, Poland
Received December 8, 1998; revision accepted January 25, 1999
Karyotype of conventionally stained and C-banded mitotic chromosomes of Rhoeo spathacea was analyzed. Chromo-somes of this diploid species (2n = 12) were classified according to their meiotic sequence described by earlier authors. The chromosome complex of Rhoeo is comparatively rich in heterochromatin (~ 18% of karyotype length), and the main differences between C-banded chromosomes concern the amount of pericentromeric heterochromatin and the presence of distally located segments. It is suggested that the karyotype of R. spathacea presented here fulfills the criteria of Bennett's "natural karyotype." C-banding of somatic interphase nuclei and mitotic prophases revealed a highly polarized arrangement of chromosomes (Rabl orientation). The centromeres have a distinct tendency to form rings within nuclei, and to gather into 1-3 large collective chromocenters. The presumptive role of non-random distribution of chromosomes at interphase is discussed.
Key words: Rhoeo spathacea (Sw.) Stearn, Rhoeo discolor (L'Heritier) Hance, karyotype, C-band-ing, heterochromatin, chromosomes, interphase nucleus.
INTRODUCTION
Rhoeo spathacea (Swartz) Stearn [synon. Rhoeo dis-color (L' Heritier) Hance] is one of the rare diploid species that form ring or chain chromosome configu-rations instead of bivalents at meiosis (Belling, 1927; Darlington, 1929a; Sax and Anderson, 1933). As in other diploid forms, its karyotype is considered to consist of two genomes, which in the case of Rhoeo have been designated α and β complexes (Flagg, 1958). These complexes are separated to the oppo-site poles during the first meiotic division. As a consequence, the gametes that arise further on con-tain only one of them (α or β). The unusual chromo-some catenation at meiosis is accomplished by
limitation of chiasma formation, confined only to the distal segments of chromosomes (Stebbins, 1950). A permanent heterozygous state is maintained within Rhoeo populations through the elimination of ho-mozygotes (α-α, or β-β), presumably because such individuals are homozygous for lethal factors (Sat-terfield and Mertens, 1972; Lin, 1979). It is generally accepted that the basis of these fundamental fea-tures of the Rhoeo genetic breeding system is a nontypical karyotype structure resulting from a series of reciprocal translocations between nonho-mologous chromosomes (Belling, 1927; Darlington, 1929b). Such a type of genetic system has also been described in the genus Oenothera (Darlington, 1929a, 1931).
* e-mail: [email protected]
PL ISSN 0001-5296 © Polish Academy of Sciences, Cracow 1999

144 Golczyk and Joachimiak
From the cytogenetic point of view, detailed data on karyotype structure in plants such as Rhoeo and Oenothera are of special interest, but only a few studies of this kind have been done so far. Data on the Rhoeo karyotype have been based mainly on analysis of the chromosome arrangement within meiotic rings (Kato, 1930; Sax, 1931; Anderson and Sax, 1936; Darlington, 1938; Coleman, 1941; Lin and Paddock, 1973). The majority of authors found a stable arrangement of chromosomes within rings but proposed different chromosome classifications, and it is troublesome to compare their results. On the other hand, it has been proposed to divide Rhoeo chromosomes into two major morphological groups: metacentrics and submetacentrics, or "heterobra-chial" and "isobrachial" chromosomes. However, the various authors differ with respect to the number of these chromosomes within karyotype (Sax, 1931; Bhaduri, 1942; Lin and Paddock, 1973). Rhoeo chro-mosomes have been C-banded by some previous authors (Natarajan and Natarajan, 1972; Pettenati, 1987; Pattankar and Ranjekar, 1988) but the C-band measurements were not presented.
This paper presents the results of karyotype analysis performed on classically stained and C-banded mitotic chromosomes of R. spathacea. The chromosomes within karyotype are classified here according to their meiotic sequence described by earlier authors (l.c.). The specific banding of Rhoeo chromosomes with prominent pericentromeric seg-ments of heterochromatin offer an opportunity for centromere arrangement studies in interphase nu-clei. In this report we describe some observations on the localization of these structures at interphase, and the spatial distribution of chromosomes within nuclei.
MATERIALS AND METHODS
Roots were taken from cuttings of different R. spa-thacea plants incubated in glass jars filled with tap water and wrapped with aluminum foil. They were maintained at 25-28°C with a 16 h photoperiod provided by cool-white fluorescent light. Plants were obtained from different sources.
CYTOLOGICAL TECHNIQUES
The root tips were treated with α-bromonaphtalene (3 h) at room temperature, refrigerated overnight (about 5°C) and then fixed in AA (glacial acetic acid and absolute alcohol, 1:3) for 2 h. Conventional chro-
mosome studies were conducted on Feulgen-stained squashes. For the sequential staining procedure, fixed root tips were squashed in acetocarmine, the cover slips were removed by the dry-ice method, and then the preparations were C-banded according to the schedule described by Schwarzacher et al. (1980).
CHROMOSOME MEASUREMENTS AND KARYOTYPE ANALYSIS
Complete metaphase plates were photographed and scanned, or directly transferred to the Lucia G com-puter system via video camera. For calculations, statistical analysis and chromosome drawing, MR Karyo ver. 3.10 (by Tokarski and Joachimiak) was used.
The chromosome lengths and banding amounts were calculated as a percentage of total karyotype length (expressed as 100%). Conventional karyotype analysis was performed on eleven complete Feulgen-stained metaphases obtained from five plants. To construct C-band karyograms, chromosomes from ten complete metaphase plates with maximum banding response were measured.
For karyotype construction, chromosomes were classified and numbered (1-12) according to their relative position within meiotic end-to-end associ-ations (rings or chains). The sequence of chromo-some types proposed here was deduced from observations of meiotic chromosome associations published by previous authors (Sax, 1931, 1935; Anderson and Sax, 1936; Coleman, 1941; Flagg, 1958; Lin and Paddock, 1973; Lin, 1979; Pettenati, 1987). Photos of chromosome rings or chains and drawings from those papers were scanned, scaled and measured to determine the relative position of each chromosome type in the meiotic superstruc-ture. Although the authors arbitrarily placed differ-ent chromosomes at the first place in the meiotic superstructure, comparison of the published data confirmed the stable positions of particular chromo-some types within rings/chains.
RESULTS AND DISCUSSION
MITOTIC CHROMOSOMES
All analyzed plants are diploids with somatic chro-mosome number 2n = 12 (Figs. 1-3). This observa-tion agrees with the results obtained by previous authors. Additionally, cells with aneuploid chromo-
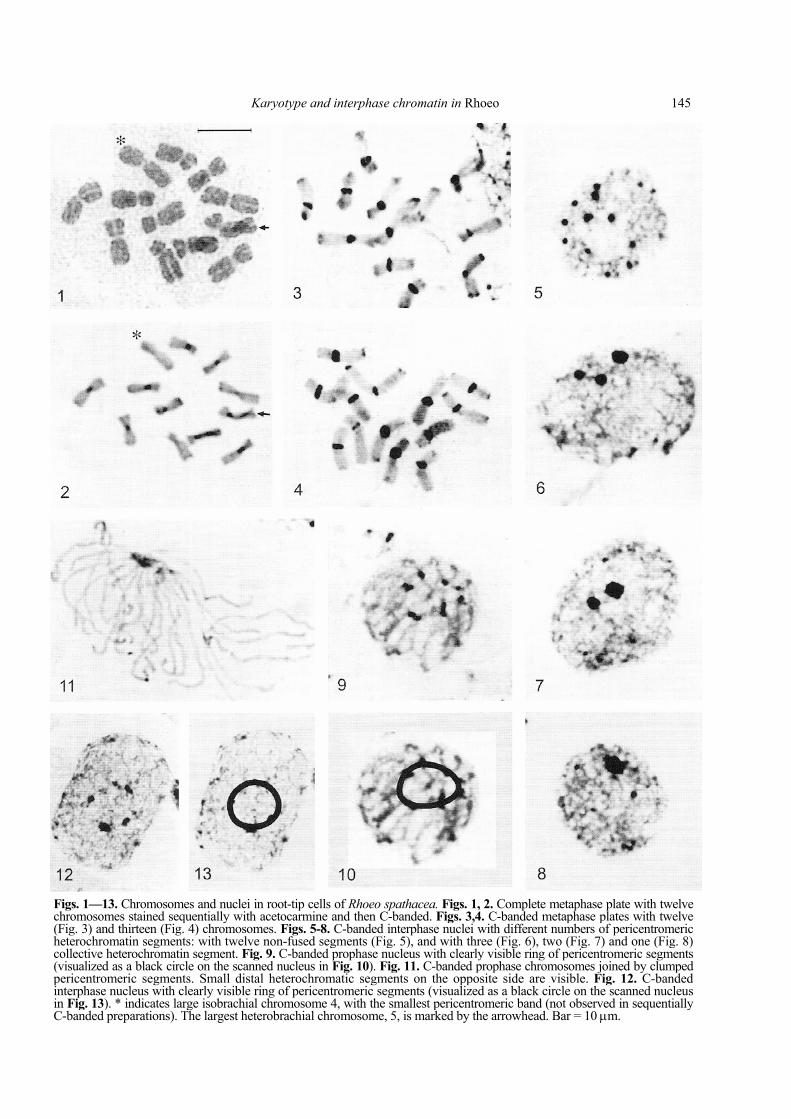
Karyotype and interphase chromatin in Rhoeo 145
Figs. 1—13. Chromosomes and nuclei in root-tip cells of Rhoeo spathacea. Figs. 1, 2. Complete metaphase plate with twelve chromosomes stained sequentially with acetocarmine and then C-banded. Figs. 3,4. C-banded metaphase plates with twelve (Fig. 3) and thirteen (Fig. 4) chromosomes. Figs. 5-8. C-banded interphase nuclei with different numbers of pericentromeric heterochromatin segments: with twelve non-fused segments (Fig. 5), and with three (Fig. 6), two (Fig. 7) and one (Fig. 8) collective heterochromatin segment. Fig. 9. C-banded prophase nucleus with clearly visible ring of pericentromeric segments (visualized as a black circle on the scanned nucleus in Fig. 10). Fig. 11. C-banded prophase chromosomes joined by clumped pericentromeric segments. Small distal heterochromatic segments on the opposite side are visible. Fig. 12. C-banded interphase nucleus with clearly visible ring of pericentromeric segments (visualized as a black circle on the scanned nucleus in Fig. 13). * indicates large isobrachial chromosome 4, with the smallest pericentromeric band (not observed in sequentially C-banded preparations). The largest heterobrachial chromosome, 5, is marked by the arrowhead. Bar = 10 µm.
146 Golczyk and Joachimiak
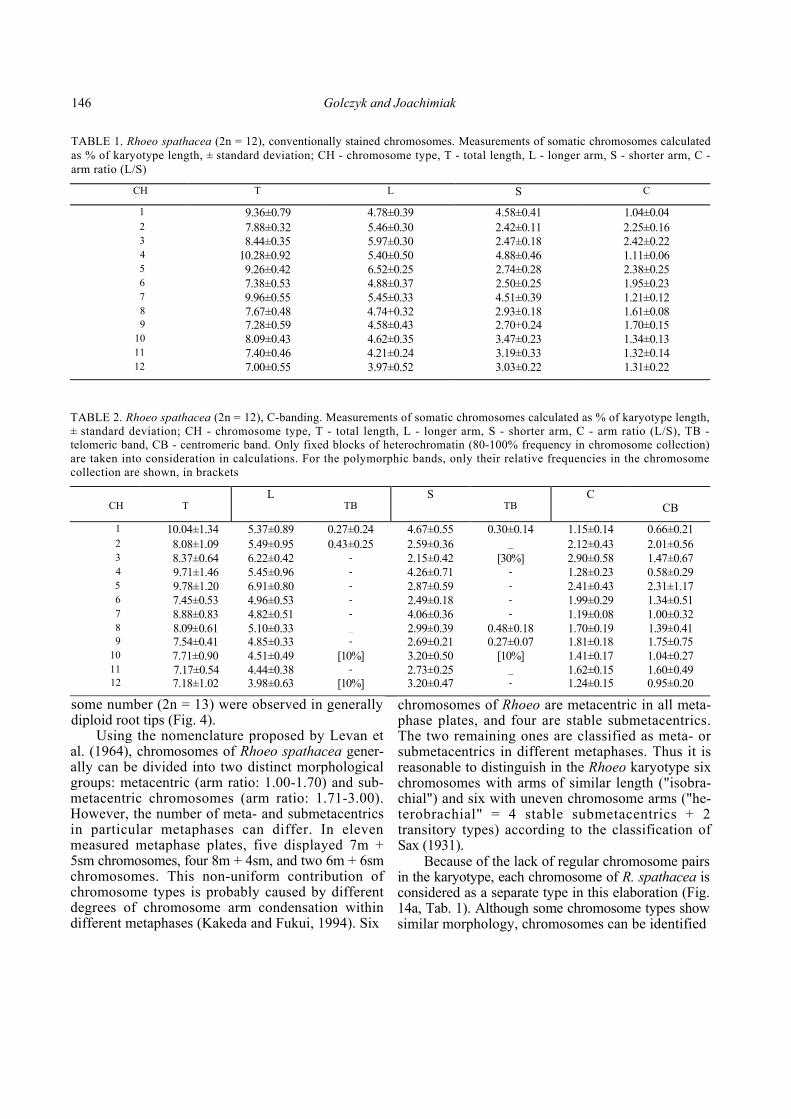
TABLE 1. Rhoeo spathacea (2n = 12), conventionally stained chromosomes. Measurements of somatic chromosomes calculated as % of karyotype length, ± standard deviation; CH - chromosome type, T - total length, L - longer arm, S - shorter arm, C -arm ratio (L/S)
CH T L S C 1 9.36±0.79 4.78±0.39 4.58±0.41 1.04±0.04 2 7.88±0.32 5.46±0.30 2.42±0.11 2.25±0.16 3 8.44±0.35 5.97±0.30 2.47±0.18 2.42±0.22 4 10.28±0.92 5.40±0.50 4.88±0.46 1.11±0.06 5 9.26±0.42 6.52±0.25 2.74±0.28 2.38±0.25 6 7.38±0.53 4.88±0.37 2.50±0.25 1.95±0.23 7 9.96±0.55 5.45±0.33 4.51±0.39 1.21±0.12 8 7.67±0.48 4.74+0.32 2.93±0.18 1.61±0.089 7.28±0.59 4.58±0.43 2.70+0.24 1.70±0.15
10 8.09±0.43 4.62±0.35 3.47±0.23 1.34±0.13 11 7.40±0.46 4.21±0.24 3.19±0.33 1.32±0.14 12 7.00±0.55 3.97±0.52 3.03±0.22 1.31±0.22
TABLE 2. Rhoeo spathacea (2n = 12), C-banding. Measurements of somatic chromosomes calculated as % of karyotype length, ± standard deviation; CH - chromosome type, T - total length, L - longer arm, S - shorter arm, C - arm ratio (L/S), TB -telomeric band, CB - centromeric band. Only fixed blocks of heterochromatin (80-100% frequency in chromosome collection) are taken into consideration in calculations. For the polymorphic bands, only their relative frequencies in the chromosome collection are shown, in brackets
CH
T
L TB
S TB
C CB
1 10.04±1.34 5.37±0.89 0.27±0.24 4.67±0.55 0.30±0.14 1.15±0.14 0.66±0.21 2 8.08±1.09 5.49±0.95 0.43±0.25 2.59±0.36 _ 2.12±0.43 2.01±0.56 3 8.37±0.64 6.22±0.42 - 2.15±0.42 [30%] 2.90±0.58 1.47±0.67 4 9.71±1.46 5.45±0.96 - 4.26±0.71 - 1.28±0.23 0.58±0.29 5 9.78±1.20 6.91±0.80 - 2.87±0.59 - 2.41±0.43 2.31±1.17 6 7.45±0.53 4.96±0.53 - 2.49±0.18 - 1.99±0.29 1.34±0.51 7 8.88±0.83 4.82±0.51 - 4.06±0.36 - 1.19±0.08 1.00±0.32 8 8.09±0.61 5.10±0.33 _ 2.99±0.39 0.48±0.18 1.70±0.19 1.39±0.419 7.54±0.41 4.85±0.33 - 2.69±0.21 0.27±0.07 1.81±0.18 1.75±0.75
10 7.71±0.90 4.51±0.49 [10%] 3.20±0.50 [10%] 1.41±0.17 1.04±0.27 11 7.17±0.54 4.44±0.38 - 2.73±0.25 _ 1.62±0.15 1.60±0.4912 7.18±1.02 3.98±0.63 [10%] 3.20±0.47 - 1.24±0.15 0.95±0.20
some number (2n = 13) were observed in generally diploid root tips (Fig. 4).
Using the nomenclature proposed by Levan et al. (1964), chromosomes of Rhoeo spathacea gener-ally can be divided into two distinct morphological groups: metacentric (arm ratio: 1.00-1.70) and sub-metacentric chromosomes (arm ratio: 1.71-3.00). However, the number of meta- and submetacentrics in particular metaphases can differ. In eleven measured metaphase plates, five displayed 7m + 5sm chromosomes, four 8m + 4sm, and two 6m + 6sm chromosomes. This non-uniform contribution of chromosome types is probably caused by different degrees of chromosome arm condensation within different metaphases (Kakeda and Fukui, 1994). Six
chromosomes of Rhoeo are metacentric in all meta-phase plates, and four are stable submetacentrics. The two remaining ones are classified as meta- or submetacentrics in different metaphases. Thus it is reasonable to distinguish in the Rhoeo karyotype six chromosomes with arms of similar length ("isobra-chial") and six with uneven chromosome arms ("he-terobrachial" = 4 stable submetacentrics + 2 transitory types) according to the classification of Sax (1931).
Because of the lack of regular chromosome pairs in the karyotype, each chromosome of R. spathacea is considered as a separate type in this elaboration (Fig. 14a, Tab. 1). Although some chromosome types show similar morphology, chromosomes can be identified

Karyotype and interphase chromatin in Rhoeo 147
Fig. 14. Conventional karyotype of Rhoeo spahacea: (a) Idiogram, chromosomes ordered according to their relative position within the meiotic superstructure, (b) Diploid set of chromosomes (from metaphase presented in Fig. 1) arranged according to Bennett's model of interphase chromosome disposition.
after measurement in well-spread Feulgen-stained metaphases. Among six isobrachial chromosomes (1, 4, 7, 10, 11, 12), two chromosomes (1, 4) are the largest ones. Four heterobrachial chromosomes (2, 3,5,6) have the highest arm ratios (1.95-2.42) in the karyotype. Two other heterobrachials (8, 9) with average arm ratios about 1.7 (1.61 and 1.7) seem to represent transitory chromosome types. The longest heterobrachial chromosome (5) is the only chromo-some marker that is easy to identify (without exact measurements of chromosomes in the metaphase plate) in the conventionally analyzed Rhoeo ka-ryotype.
The chromosome sequence in the karyotype presented here (Fig. 14a, Tab. 1) is similar to the idiogram created by Lin and Paddock (1973). The chromosome labelling systems used by Lin and Pad-dock (1973) and Sax (1931) are included (Fig. 14a; letters used by Sax that are inconsistent with Lin and Paddock's terminology are displayed within brackets). Here we propose simple number labels (1-12) instead of this complicated nomenclature, which those authors used only in order to be in accordance with the segmental interchange hypo-thesis (Belling, 1927). For example, their labeling of one of the twelve Rhoeo chromosomes as Dd or DE was arbitrary. Up to now there is no real cytogenetic evidence supporting this particular chromosome la-belling system.
The C-banding karyotype of R. spathacea is heterochromatin-rich (~ 18% of karyotype length) and not very differentiated (Fig. 15). Because of alterations in chromosome morphology caused by the C-banding procedure (Joachimiak et al., 1994), exact identifica-tion of chromosome types (obtained in conventional karyotype analysis) in C-banded metaphases is diffi-cult. For correct karyotyping, each particular C-banded chromosome type was identified first in material by sequential staining (Figs. 1,2). For C-band measurements, however, only freshly C-banded prep-arations were selected. In these preparations the chro-mosomes show maximum banding response and excellent contrast between eu- and heterochromatin (Figs. 3,4). The majority of Rhoeo chromosomes reveal clearly visible blocks of pericentromeric heterochro-matin (16.1% of karyotype length). Heterobrachial chromosomes are especially rich in this type of hete-rochromatin (10.27%). Two large isobrachial chromo-somes (1, 4) possess only small pericentromeric segments (1.24% of karyotype length). In chromosome 4 the pericentromeric band is delicate, thus not ob-served in some preparations, especially sequentially stained ones (Fig. 2). Some chromosomes show a fixed telomeric block of heterochromatin: isobrachial chro-mosome 1 on both arms, chromosome 2 on the longer arm, and chromosomes 8 and 9 on the shorter arms (Fig. 15). Chromosomes 3, 10 and 12 reveal small polymorphic segments of heterochromatin (Tab. 2),

148 Golczyk and Joachimiak
Fig. 15. C-band karyotype of Rhoeo spathacea. Only fixed bands (80-100% frequency in chromosome collection) are shown.
observed only occasionally as two dots on the chromo-some termini. Fixed telomeric blocks of heterochro-matin occupy only 1.75% of the karyotype length. Thus the main differences between chromosomes concern the amount of pericentromeric heterochromatin and the presence of telomeric heterochromatin (Fig. 15, Tab. 2). The C-band distribution presented here is similar to the results obtained by Pettenati (1987), but some additional telomeric bands were revealed.
Some authors suggest that the mitotic/meiotic chromosome arrangement is conditioned by their spa-tial interphase arrangement (Ashley and Pocock, 1981; Bennett, 1982, 1984; Lavania and Sharma, 1984; Fussell, 1984). Based on a postulated regular interphase chromosome disposition, Bennett (1984) proposed a new system of plant karyotype construc-tion. In this system ("natural karyotype"), chromo-somes within genome should be ordered according to their position in the interphase structure (chain or ring). The chromosome position can be deduced (Ash-ley and Pocock, 1981; Bennett, 1984; Fussell, 1984) from the arm lengths: each chromosome in the "natu-ral karyotype" has constant neighbors with similar arm lengths.
The chromosomes in the karyotype presented here were ordered according to their relative posi-tion within the meiotic superstructures. There are three heterobrachial chromosome pairs (2-3, 5-6, 8-9) flanked by isobrachial chromosomes (Fig. 14a). According to Sax (1931), the most characteristic sequence of eight chromosomes (2-9) is invariably the same within different meiotic rings/chains. At meiosis, heterobrachial chromosomes are always conjoined by their short arms. An intuitive diagram of multichromosomal structure so organized was first depicted by Coleman (1941).
The karyotype obtained in the above-described way fulfills Bennett's "natural karyotype" criteria (Bennett, 1984): particular chromosome types occupy stable positions in the superstructure, and each of
them has neighbors with similar arm lengths (Fig. 14b). It could be argued that the special meiotic behavior of Rhoeo chromosomes (formation of multiple end-to-end associations) is a reflection of the spatial interphase organization of the karyotype, not exclusively the segmental chromosome homology. In-terestingly, observations of meiotic synapsis and chi-asma formation in Rhoeo (Roller, 1932; McQuade and Wells, 1975; Stack and Soulliere, 1984) are not conclu-sive for the partial homology hypothesis. The postu-lated restriction of synapsis and crossing-over to small terminal segments of chromosomes has not been con-firmed in these studies. Observations of synaptonemal complexes by McQuade and Wells (1975) suggest that each chromosome of Rhoeo can be paired throughout its full length at early meiotic prophase. However, the relationships between crossover, synapsis and chiasma formation are not well recognized yet (for review: Maguire, 1995).
Williams and Heslop-Harrison (1975) observed the unusual peripheral membranous structures as-sociated with R. spathacea meiotic chromosomes. These structures have no obvious affinity with any other previously described chromosome-associated bodies, and they might form bridges between the ends of the adjoining chromosomes. As suggested by the authors, "the chromosome-associated mem-brane system might be related in some way with the chromosome catenation." Thus it is not obvious that the chromosome arrangement within meiotic rings is conditioned only by partial arm homology and terminal chiasmata formation.
INTERPHASE CHROMOSOMES
The specific banding of Rhoeo chromosomes (with clearly visible blocks of heterochromatin at the cen-tromeres) offers an opportunity for study of their interphase disposition. In the analyzed material, centromeric segments were located mainly on one

Karyotype and interphase chromatin in Rhoeo 149
side of the nucleus. They were observed as the num-ber of heterochromatic bodies (up to 12) or as 1-3 large collective segments (Figs. 5-8). In the gently squashed preparations, the centromeres form clear-ly visible rings within nuclei (Figs. 9, 10, 12, 13). Small telomeric segments are located on the oppo-site side. A highly polarized arrangement of chromo-somes is also observed at early prophase (Fig. 9). Thus, interphase chromosomes of Rhoeo maintain the telophase configuration, complying with Rabl's hypothesis (Rabl, 1885). Lately the Rabl orientation of chromosomes at interphase has been confirmed by many authors in mitotic (Fussell, 1975, 1977, 1983; Sperling and Ludtke, 1981; Cremer et al., 1982; Korf et al., 1982; Barnes et al., 1985; Joachimiak, 1987; Heslop-Harrison et al., 1988; Anamthawat-Jonsson and Heslop-Harrison, 1990; Rawlins et al., 1991; No-guchi and Fukui, 1995) and meiotic cells (Thermann and Denniston, 1984; Church and Moens, 1976; Suzu-ki et al., 1997). It is very probable that chromosome polarity is restricted to the undifferentiated mi-totic/meiotic nuclei. In some types of differentiated cells, chromosomes and chromatin can move within the nucleus, and this movement is cell-cycle dependent or corresponds to unknown regulatory mechanisms (for review: Noguchi and Fukui, 1995; Ceccarelli et al., 1998; Lamond and Earnshaw, 1998).
The individual nature of pericentromeric seg-ments is preserved within collective chromocenters of some interphase and prophase nuclei, but in other large heterochromatic segments can form homogen-ous, uniformly stained bodies. Even in very squashed, well-spread preparations these bodies maintain their structural homogeneity (Fig. 11). It is very probable that pericentromeric heterochro-matin in Rhoeo is capable of ectopic pairing, analo-gous to heterochromatic sequences in Drosophila (Mayfield and Ellison, 1975). Similar collective hete-rochromatic bodies including DNA domains from several chromosomes were observed in interphase nuclei of other plants (Ceccarelli et al., 1998).
The number and position of heteropycnotic seg-ments in Rhoeo nuclei have been studied by some authors (Heitz, 1932; Coleman, 1941; Huskins and Steinitz, 1948; Patankar and Ranjekar, 1988). Their observations were similar to those presented here. In mitotic and meiotic nuclei, heteropycnotic seg-ments were accumulated on one side of the nucleus as separate or collective chromocenters. In PMCs, Coleman (1941) observed distinct chromocenters, in number approximate to diploid (2n = 12) chromo-some number in premeiotic nuclei. These chro-mocenters are aggregated in one, two, or more rarely
three clumps in zygo- and pachyphase. Natarajan and Natarajan (1972) also observed centromere fu-sion at pachytene to form one or two large heteroch-romatin bodies. Meiotic centromere behavior in Rhoeo might be related to the unusual mode of chromosome segregation. In other plant species, the centromere associations occasionally formed before meiosis are usually resolved prior to zygotene pair-ing (Church and Moens, 1976; Suzuki et al., 1997).
REFERENCES
ANAMTHAWAT-JONSSON K, and HESLOP-HARRISON JS. 1990. Cen-tromeres, telomeres and chromatin in the interphase nu-cleus of cereals. Caryologia 43: 205-213.
ANDERSON E, and SAX K. 1936. A cytological monograph of the American species of Tradescantia. Botanical Gazette 97: 433-476.
ASHLEY T, and POCOCK N. 1981. A proposed model of chromosomal organization in nuclei at fertilization. Genetica 55: 161-169.
BARNES SR, JAMES AM, and JAMIESON G. 1985. The organisation, nucleotide sequence, and chromosomal distribution of a satellite DNA from Allium cepa. Chromosoma 92: 185-192.
BELLING J. 1927. The attachment of chromosomes at the reduction division in flowering plants. Journal of Genetics 18: 177-205.
BHADURI PN. 1942. Cytological analysis of structural hybridity in Rhoeo discolor Hance. Journal of Genetics 44: 73-85.
BENNETT MD. 1982. Nucleotypic basis of the spatial ordering of chromosomes in eukaryotes and the implications of the order for genome evolution and phenotypic variation. In: Dover GA, Flavell RB [ed.], Genome evolution. Academic Press, New York.
BENNETT MD. 1984. The genome, the natural karyotype, and biosystematics. In: Grant WF [ed.], Plant Biosystematics, 41-66. Academic Press, Canada.
CECCARELLI M, MOROSI L, and CIONINI PG. 1998. Chromocenter association in plant cell nuclei: determinants, functional sig-nificance, and evolutionary implications. Genome 41: 96-103.
CHURCH K, and MOENS PB. 1976. Centromere behaviour during interphase and meiotic prophase in Allium fistulosum from 3-D E.M. reconstruction. Chromosoma 56: 249-263.
COLEMAN LC. 1941. The relation of chromocenters to the differen-tial segments in Rhoeo discolor Hance. American Journal of Botany 28: 742-748.
CREMER T, CREMER C, BAUMANN H, LUDTKE E-K, SPERLING K, TEUBER V, and ZERN C. 1982. Rabl's model of the interphase chromosome arrangement tested in Chinese hamster cells by premature chromosome condensation and laser-UV-microbeam experiments. Human Genetics 60: 46-56.
DARLINGTON CD. 1929a. Ring-formation in Oenothera and other genera. Journal of Genetics 20: 345-363.
DARLINGTON CD. 1929b. Chromosome behaviour and structural hybridity in the Tradescantiae. Journal of Genetics 21: 207-286.
DARLINGTON CD. 1931. The cytological theory of inheritance in Oenothera. Journal of Genetics 24: 405-474.

150 Golczyk and Joachimiak
DARLINGTON CD. 1938. Chromosome behaviour and structural hybridity in the Tradescantiae. II. Journal of Genetics 35: 259-280.
FLAGG RO. 1958. A mutation and an inversion in Rhoeo discolor. Journal of Heredity 49: 185-188.
FUSSELL CP. 1975. The position of interphase chromosomes and late replicating DNA in centromere and telomere regions of Allium cepa L. Chromosoma 50: 201-210.
FUSSELL CP. 1977. Telomere associations in interphase nuclei of Allium cepa demonstrated by C-banding. Experimental Cell Research 110: 111.
FUSSELL CP. 1983. Telomere arrangement in differentiated inter-phase cells of Allium cepa: a function of chromosome arm lengths at anaphase-telophase. Canadian Journal of Gene-tics and Cytology 25: 478-486.
FUSSELL CP. 1984. Interphase chromosome order: A proposal. Genetica 62: 193-201.
HEITZ E. 1932. Die Herkunft der Chromozentren. Dritter Beitrag zur Kenntnis der Beziehung zwischen Kernstruktur und qualitativer Verschiedenheit der Chromosomen in ihrer Langsrichtung. Planta 18: 571-636.
HESLOP-HARRISON JS, HUELSKAMP M, WENDROTH S, ATKINSON MD, LEITCH AR, and BENNETT MD. 1988. Chromatin and cen-tromeric structures in interphase nuclei. Kew Chromosome Conference III, HMSO, 209-217.
HUSKINS CL, and STEINITZ LM. 1948. The nucleus in differentia-tion and development. I. Heterochromatic bodies in energic nuclei of Rhoeo roots. Journal of Heredity 39: 35-43.
JOACHIMIAK A. 1987. Telomere arrangement in interphase nuclei of Allium cepa L. Acta Biologica Cracoviensia Series Botani-ca 29: 64-70.
JOACHIMIAK A, KORNAŚ A, and KRAWCZYK J. 1994. Alterations in chromosome morphology after sequentional aceto-orceine staining and C-banding. Acta Biologica Cracoviensia Series Botanica 36: 23-30.
KAKEDA K, and FUKUI K. 1994. Dynamic changes in the morpho-logy of barley chromosomes during the mitotic metaphase stage. Japanese Journal of Genetics 69: 545-554.
KATO K. 1930. Cytological studies of pollen mother cells of Rhoeo discolor Hance, with special reference to the question of the mode of syndesis. Memoirs of the Collegium of Science of the Kyoto Imperial University Series: B 5: 139-161.
ROLLER P. 1932. Further studies in Tradescantia virginiana var. humilis and Rhoeo discolor. Journal of Genetics 26: 81-96.
KORF BR, GERSHEY EL, and DIACUMACOS EG. 1982. Centromeres are arranged in clusters throughout the muntjac cell cycle. Experimental Cell Research 139: 393-396.
LAMOND AI, and EARNSHAW WC. 1998. Structure and function in the nucleus. Science 280: 547-553.
LAVANIA UC, and SHARMA AK. 1984. Mitotic spatial model: Ar-rangement of homologous chromosomes. Genetica 62, 203-208.
LEVAN A, FREDGA K, and SANDBERG AA. 1964. Nomenclature for the centromeric position on chromosomes. Hereditas 52: 201-220.
LIN YJ. 1979. Chromosome distribution and catenation in Rhoeo spathacea var. concolor. Chromosoma 71: 109-127.
LIN YJ, and PADDOCK EF. 1973. Ring-position and frequency of adjacent distribution of meiotic chromosomes in Rhoeo spa-thacea. American Journal of Botany 60: 685-690.
MAGUIRE MP. 1995. Is the synaptonemal complex a disjunction machine? Journal of Heredity. 86: 330-340.
MAYFIELD JE, and ELLISON JR. 1975. The organization of inter-phase chromatin in Drosophilidae. The self adhesion of chromatin containing the same DNA sequence. Chromoso-ma 52: 37-48.
MCQUADE HA, and WELLS B. 1975. The synaptinemal complex in Rhoeo spathacea. Journal of Cell Science 17: 349-369.
NATARAJAN AT, and NATARAJAN S. 1972. The heterochromatin of Rhoeo discolor. Hereditas 72: 323-330.
NOGUCHI J, and FUKUI K. 1995. Chromatin arrangements in intact interphase nuclei examined by laser confocal micro-scopy. Journal of Plant Research 108: 209-216.
PATANKAR S, and RANJEKAR PK. 1988. Constitutive heterochro-matin during meiosis and mitosis in Allium cepa and Rhoeo discolor. Cytologia 53: 275-281.
PETTENATI MJ. 1987. Giemsa C - banding of Rhoeo (Commelina-ceae). Genetica 74: 219-224.
RABL C. 1885. Uber Zellteilung. In: Gegenbaur C [ed.], Morpho-logisches Jahrbuch, Vol. 10: 214-330.
RAWLINS DJ, HIGHETT MI, and SHAW PJ. 1991. Localization of telomeres in plant interphase nuclei by in situ hybridization and 3D confocal microscopy. Chromosoma 100: 424-431.
SATTERFIELD SK, and MERTENSTR. 1972. Rhoeo spathacea: A tool for teaching meiosis and mitosis. Journal of Heredity 63: 375-378.
SAX K. 1931. Chromosome ring formation in Rhoeo discolor. Cytologia 3: 36-53.
SAX K. 1935. Chromosome structure in the meiotic chromosomes of Rhoeo discolor Hance. Journal of Arnold Arboretum 16: 216-224.
SAX K, and ANDERSON E. 1933. Segmental interchange in chromo-somes of Tradescantia. Genetics 18: 53-67.
SCHWARZACHER T, AMBROS P, and SCHWEIZER D. 1980. Application of Giemsa banding to orchid karyotype analysis. Plant Sys-tematics and Evolution 134: 293-297.
SPERLING K, and LUDTKE E-K. 1981. Arrangement of prematurely condensed chromosomes in cultured cells and lymphocytes of the Indian muntjac. Chromosoma 83: 541-553.
STACK SM, and SOULLIERE DL. 1984. The relation between synap-sis and chiasma formation in Rhoeo spathacea. Chromoso-ma 90: 72-83.
STEBBINS GL. 1950. Variation and evolution in plants. Columbia University Press, New York.
SUZUKI T, IDE N, and TANAKA I. 1997. Immunocytochemical vis-ualization of the centromeres during male and female meiosis in Lilium longiflorum. Chromosoma 106: 435-445.
THERMAN E, and DENNISTON C. 1984. Random arrangement of chromosomes in Uvularia (Liliaceae). Plant Systematics and Evolution 147: 289-297.
WILLIAMS E, and HESLOP-HARRISON J. 1975. Membrane-bounded cisternae associated with the chromosomes at first meiotic metaphase in microsporocytes of Rhoeo spathacea. Proto-plasma 86: 285-289.





























