Embed Size (px)
Citation preview

Aw
DMB
a
ARRAA
KDRRSV
1
i(tpDsrse[oi
1d
DNA Repair 8 (2009) 813–821
Contents lists available at ScienceDirect
DNA Repair
journa l homepage: www.e lsev ier .com/ locate /dnarepai r
ccessibility of chromosomal recombination breaks in nuclei ofild-type and DNA-PKcs-deficient cells
aniel Franco, Yung Chang ∗
olecular and Cellular Biology Program, School of Life Sciences, Center for Infectious Diseases and Vaccinology, The Biodesign Institute at Arizona State University,DA220, 1001 S. McAllister Avenue, Tempe, AZ 85287-5501, United States
r t i c l e i n f o
rticle history:eceived 16 October 2008eceived in revised form 24 February 2009ccepted 23 March 2009vailable online 22 April 2009
eywords:NA-PKcsAG1AG2cid(D)J recombination
a b s t r a c t
V(D)J recombination is a highly regulated process, proceeding from a site-specific cleavage to an impreciseend joining. After the DNA excision catalyzed by the recombinase encoded by recombination activatinggenes 1 and 2 (RAG1/2), newly generated recombination ends are believed held by a post-cleavage complex(PC) consisting of RAG1/2 proteins, and are subsequently resolved by non-homologous end joining (NHEJ)machinery. The relay of these ends from PC to NHEJ remains elusive. It has been speculated that NHEJfactors modify the RAG1/2-PC to gain access to the ends or act on free ends after the disassembly of the PC.Thus, recombination ends may either be retained in a complex throughout the recombination process orleft as unprotected free ends after cleavage, a condition that may permit an alternative, non-classical NHEJend joining pathway. To directly test these scenarios on recombination induced chromosomal breaks, wehave developed a recombination end protection assay to monitor the accessibility of recombination endsto exonuclease-V in intact nuclei. We demonstrate that these ends are well protected in the nuclei ofwild-type cells, suggesting a seamless cleavage-joining reaction. However, divergent end protection ofcoding versus signal ends was found in cells derived from severe combined immunodeficient (scid) mice
that are defective in the catalytic subunit of DNA-dependent protein kinase (DNA-PKcs). While signal endsare resistant, opened coding ends are susceptible to enzymatic modification. Our data suggests a role ofDNA-PKcs in protecting chromosomal coding ends. Furthermore, using recombination inducible scid celllines, we demonstrate that conditional protection of coding ends is inversely correlated with the levelof their resolution, i.e., the greater the accessibility of the coding ends, the higher level of coding jointsformed. Taken together, our findings provide important insights into the resolution of recombinationnativ
ends by error-prone alter. Introduction
The variable regions of antigen-receptor genes, coding formmunoglobulins and T cell receptors, are assembled from variableV), diversity (D) and joining (J) gene segments by a recombina-ion reaction, known as V(D)J recombination. This recombinationrocess occurs in two distinct, but coupled steps, a site-specificNA excision followed by imperfect DNA end joining [1,2]. The
ite-specific cleavage is dictated by the presence of conservedecombination signal sequences (RSS) flanking each V, D or J geneegment, which are targeted by lymphoid-specific recombinases
ncoded by the recombination activation genes, RAG1 and RAG21]. RAG1/2 proteins initiate recombination by recognizing a pairf RSS, catalyzing excision at the border between RSS and cod-ng segments, generating hairpinned coding ends (CEs) and blunt∗ Corresponding author. Tel.: +1 480 965 8672; fax: +1 480 727 0599.E-mail address: [email protected] (Y. Chang).
568-7864/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.dnarep.2009.03.007
e NHEJ pathways.© 2009 Elsevier B.V. All rights reserved.
signal ends (SEs) [3–5]. After cleavage, these ends are believed toremain associated with RAG1/2 proteins, constituting the transitorystructure referred to as the post-cleavage complex (PC) [6,7]. Subse-quently, SEs and CEs are resolved into signal joints (SJs) and codingjoints (CJs), respectively, by non-homologous end joining (NHEJ)proteins, including the catalytic subunit of DNA-dependent proteinkinase (DNA-PKcs), Artemis, Ku70/80, XRCC4, Cernunnos/XLF andLigase IV [8–12].
Although inactivation of any of these NHEJ genes results indefective V(D)J recombination and aborted lymphocyte develop-ment in mutant mice, the cells derived from these individualmutants are still capable of, albeit at very low levels, resolvingsome double-stranded DNA breaks (DSBs), presumably by alterna-tive NHEJ pathways [13]. Interestingly, this alternative pathway has
also been observed in cells that express certain mutated forms ofRAG1 or RAG2 [14,15]. These findings have led to a model in whichfunctional RAG1/2 direct recombination ends to the classical NHEJpathway for efficient resolution. According to this model, eitherdefective RAG1/2 proteins that destabilize the PC or NHEJ mutants
8 NA Re
teiei
otpfpllbpjntbwprastTpasifo
endDocfsavfaftc
2
2
pbmma1BB3rti
14 D. Franco, Y. Chang / D
hat fail to resolve the ends efficiently may expose recombinationnds to alternative pathways or factors for aberrant resolution,ncluding homologous recombination (HR) factors [16–18]. How-ver, the molecular nature of alternative factors and how theynteract with a functional RAG-PC is largely unknown.
To address these issues, we focus on severe combined immun-deficient (scid) mice that are defective in DNA-PKcs and are knowno have leaky recombination activity via a DNA-PK-independentathway [19,20]. We derived recombination inducible cell linesrom scid mice, designated as scid-ts, by transformation of scidrogenitor B cells with temperature sensitive Ableson Murine
eukemia virus (Abl-MuLV). These cells display conditional reso-ution of newly induced recombination ends. Specifically, they cane induced by incubation at 39 ◦C, i.e., the non-permissive tem-erature, to initiate recombination cleavage, but form very few
oining products. A significant amount of coding joints and sig-al joints is made after these cells are returned to the permissiveemperature, 33 ◦C [21]. Thus, the alternative NHEJ pathway coulde readily induced in these cells via experimental manipulation,hich offers a model to further explore molecular components androcesses of the alternative NHEJ pathway. Our observation of a cor-elation between the level of CJs with the level of opened CEs, andn inverse correlation between end resolution and RAG1/2 expres-ion (AppendixBsupplementary Fig. 1A) agrees with the idea thathe RAG1/2-mediated PC may impede final end resolution [22–24].hen, the disassembly of the PC complex, probably occurring at theermissive temperature in our scid-ts cells, may render the endsccessible to resolution machinery independent of DNA-PKcs. Asuch, we hypothesize that the accessibility of recombination endsn scid cells exerts an influence on the ability of non-classical NHEJactors to recognize and execute joining of these ends in the absencef DNA-PKcs.
To directly test this hypothesis, we developed the recombinationnd protection assay (REPA) to monitor the accessibility of recombi-ation ends to a processing enzyme, exonuclease-V (Exo-V), whichegrades double-strand DNA (ds-DNA) and single-strand DNA (ss-NA) with free DNA ends [25]. Here, we focused on the analysisf opened coding ends and signal ends, because of their techni-al feasibility. In particular, we examined whether these ends areree DNA breaks or protected by protein complexes. By analyzingusceptibility of recombination ends to Exo-V, we compared theccessibility of these ends in scid and wild-type cells, and pro-ide evidence for a correlation of opened CE accessibility with CJormation in scid, but not wild-type cells. Thus, our novel assayllows us to monitor recombination induced chromosomal breaksor their availability to end resolution machinery, offering impor-ant insight into the alternative pathway for end resolution in scidells.
. Materials and methods
.1. Mice and cell lines
Scid and wild-type Balb/c mice were bred and maintained asreviously described [26]. Thymocytes from scid homozygous new-orn (1–3 days old) mice were pooled (6–8 mice) to obtain 10illion cells. Wild-type thymocytes were harvested from Balb/cice. The s/+−ts and scid-ts cell lines were derived from scid
nd s/+ mice, respectively [26]. RAG2−/− Abl-MuLV cell line, 63-2, was kindly provided by Dr. F. Alt (Harvard Medical School,oston, MA) via Dr. M. Schlissel (University of California, Berkeley,
erkeley, CA). Temperature sensitive cell lines were maintained at3 ◦C, and induced to undergo immunoglobulin light-chain geneearrangement by incubation at 39 ◦C for 48 h. Cells returnedo 33 ◦C after induction were incubated for 24 h before harvest-ng.pair 8 (2009) 813–821
2.2. Recombination end protection assay
This assay is designed to test the susceptibility of endogenouslyproduced broken DNA ends to exonulcease-V (Exo-V). While freeDNA ends are expected to be susceptible to the Exo-V mediateddigestion the ends bound by proteins or protein complex may beresistant to this enzyme treatment. Although hairpin CEs exist inlarge abundance in scid lymphocytes, it is technically difficult toexamine their accessibility in intact nuclei as the physiologicalenvironment of the isolated nuclei is not compatible with enzy-matic conditions necessary for hairpin processing enzymes, suchas mung bean nuclease (MBN). Therefore, we primarily focus onthe analysis of opened CEs and SEs. Nuclei were isolated frominducible cell lines or primary thymocytes by lysing cells in hypo-tonic buffer (50 mM Hepes, pH 7.5, 10 mM KCl, 10 mM MgCl2,1 mM dithiothreitol) containing 0.1% IGEPAL and protease inhibitors(Sigma–Aldrich, St. Louis, MO). The released nuclei were collectedby centrifugation at 3000 RPM for 1 min. 4–5 million scid nucleiand 10 million s/+ nuclei were resuspended in 100 �l and 200 �lof Exo-V digestion buffer, respectively. Isolated nuclei were dividedamong equivalent sample volumes of 50 �l, including buffer con-trol at 4 ◦C, buffer control at 37 ◦C (in the presence or absence ofdNTP), and buffer plus Exo-V (USB scientific, Cleveland, OH), at 7or 15 units/50 �l. For inhibition experiments, the nuclei were firstincubated with inhibitors [30 �M of the ATM inhibitor, KU 55933(Calbiochem CAS 587871-26-9) and 30 �M of the DNA-PK inhibitorNU 7026 (Sigma–Aldrich, CAS 154447-35-5), or 0.25 mM of ATP-�-S (Sigma–Aldrich, CAS 93839-89-5) on ice for 30 min before theirincubation at 37 ◦C. Stock solutions for the ATM and DNA-PKcsinhibitors were prepared in DMSO and a vehicle control samplewas prepared at 0.1% in the final reaction volume. ATP-�-S was pre-pared as a stock solution in H2O. In some experiments, purifiedDNA molecules embedded in agarose-plugs were also included forthe susceptibility of pre-existing DNA breaks to Exo-V-mediateddegradation. Digestions were carried out at 37 ◦C for 1 h followedby the addition of 50 �l of molten 1.2% low melt agarose (SeaKem)before deproteinization with proteinase K (Roche, Indianapolis,IN).
2.3. Ligation-mediated PCR (LM-PCR) and Southern blotting
Recombination ends were detected by modified LM-PCR fol-lowed by Southern blot analysis, as described previously [27,21]. Toexamine broken staggered ends, purified DNA molecules embed-ded in agarose-plugs were treated with T4 DNA polymerase andthen ligated to an artificial linker by T4 DNA ligase. To reveal hair-pin ends, the plugs were first treated with mung bean nuclease(MBN), followed by T4 DNA polymerase, and then ligated to thelinker. One 20th of ligated DNA plugs (equivalent to 200–250,000cells) were amplified for 31 cycles to reveal coding and signalends. Similar amounts of DNA were also amplified for CJs, SJs andgermline control (GL) or actin. Given the abundance of joined prod-ucts in the cells, CJs and SJs were amplified for 26 cycles whereasinput control, actin or GL were amplified for 21–22 cycles. A semi-quantitative PCR reaction was also performed, i.e., several dilutionsof ligated DNA were subjected to PCR to ensure relative linearity ofPCR amplification (AppendixBsupplementary Fig. 2A). The inten-sity of PCR bands representing recombination ends and productswas quantified using a Phosphorimager and analyzed by ImageQuant software (Molecular Dynamics). The quantification of eachband was adjusted to the input DNA control, i.e., actin (or GL), then
divided by the control-end (i.e., the ends present in the nuclei incu-bated at 4 ◦C or 37 ◦C) to its actin ratio. For example, the calculationfor CEs was sample-(CEs/actin)–control-(CEs/actin). For Fig. 1, theend input (i.e., C) was arbitrarily set at 100% as a reference for thesamples under various treatments. For the other figures, the inten-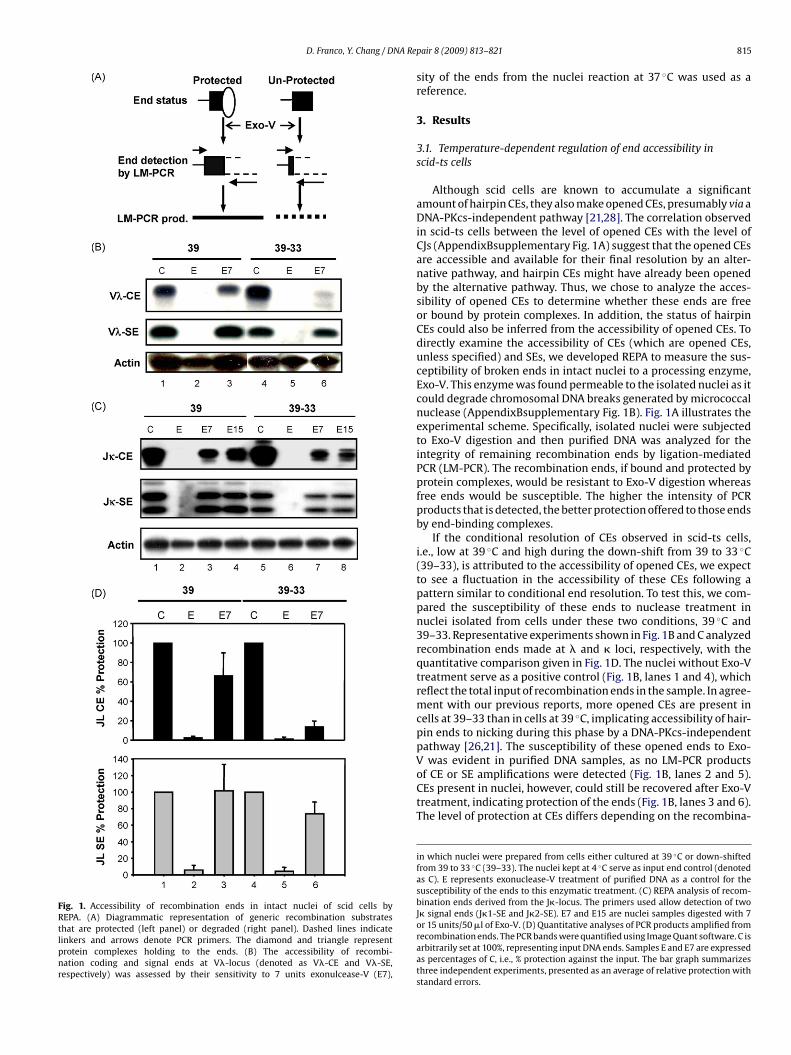
D. Franco, Y. Chang / DNA Re
Fig. 1. Accessibility of recombination ends in intact nuclei of scid cells byREPA. (A) Diagrammatic representation of generic recombination substratesthat are protected (left panel) or degraded (right panel). Dashed lines indicatelinkers and arrows denote PCR primers. The diamond and triangle representprotein complexes holding to the ends. (B) The accessibility of recombi-nation coding and signal ends at V�-locus (denoted as V�-CE and V�-SE,respectively) was assessed by their sensitivity to 7 units exonulcease-V (E7),
V
pair 8 (2009) 813–821 815
sity of the ends from the nuclei reaction at 37 ◦C was used as areference.
3. Results
3.1. Temperature-dependent regulation of end accessibility inscid-ts cells
Although scid cells are known to accumulate a significantamount of hairpin CEs, they also make opened CEs, presumably via aDNA-PKcs-independent pathway [21,28]. The correlation observedin scid-ts cells between the level of opened CEs with the level ofCJs (AppendixBsupplementary Fig. 1A) suggest that the opened CEsare accessible and available for their final resolution by an alter-native pathway, and hairpin CEs might have already been openedby the alternative pathway. Thus, we chose to analyze the acces-sibility of opened CEs to determine whether these ends are freeor bound by protein complexes. In addition, the status of hairpinCEs could also be inferred from the accessibility of opened CEs. Todirectly examine the accessibility of CEs (which are opened CEs,unless specified) and SEs, we developed REPA to measure the sus-ceptibility of broken ends in intact nuclei to a processing enzyme,Exo-V. This enzyme was found permeable to the isolated nuclei as itcould degrade chromosomal DNA breaks generated by micrococcalnuclease (AppendixBsupplementary Fig. 1B). Fig. 1A illustrates theexperimental scheme. Specifically, isolated nuclei were subjectedto Exo-V digestion and then purified DNA was analyzed for theintegrity of remaining recombination ends by ligation-mediatedPCR (LM-PCR). The recombination ends, if bound and protected byprotein complexes, would be resistant to Exo-V digestion whereasfree ends would be susceptible. The higher the intensity of PCRproducts that is detected, the better protection offered to those endsby end-binding complexes.
If the conditional resolution of CEs observed in scid-ts cells,i.e., low at 39 ◦C and high during the down-shift from 39 to 33 ◦C(39–33), is attributed to the accessibility of opened CEs, we expectto see a fluctuation in the accessibility of these CEs following apattern similar to conditional end resolution. To test this, we com-pared the susceptibility of these ends to nuclease treatment innuclei isolated from cells under these two conditions, 39 ◦C and39–33. Representative experiments shown in Fig. 1B and C analyzedrecombination ends made at � and � loci, respectively, with thequantitative comparison given in Fig. 1D. The nuclei without Exo-Vtreatment serve as a positive control (Fig. 1B, lanes 1 and 4), whichreflect the total input of recombination ends in the sample. In agree-ment with our previous reports, more opened CEs are present incells at 39–33 than in cells at 39 ◦C, implicating accessibility of hair-pin ends to nicking during this phase by a DNA-PKcs-independentpathway [26,21]. The susceptibility of these opened ends to Exo-
was evident in purified DNA samples, as no LM-PCR productsof CE or SE amplifications were detected (Fig. 1B, lanes 2 and 5).CEs present in nuclei, however, could still be recovered after Exo-Vtreatment, indicating protection of the ends (Fig. 1B, lanes 3 and 6).The level of protection at CEs differs depending on the recombina-
in which nuclei were prepared from cells either cultured at 39 ◦C or down-shiftedfrom 39 to 33 ◦C (39–33). The nuclei kept at 4 ◦C serve as input end control (denotedas C). E represents exonuclease-V treatment of purified DNA as a control for thesusceptibility of the ends to this enzymatic treatment. (C) REPA analysis of recom-bination ends derived from the J�-locus. The primers used allow detection of twoJ� signal ends (J�1-SE and J�2-SE). E7 and E15 are nuclei samples digested with 7or 15 units/50 �l of Exo-V. (D) Quantitative analyses of PCR products amplified fromrecombination ends. The PCR bands were quantified using Image Quant software. C isarbitrarily set at 100%, representing input DNA ends. Samples E and E7 are expressedas percentages of C, i.e., % protection against the input. The bar graph summarizesthree independent experiments, presented as an average of relative protection withstandard errors.
8 NA Repair 8 (2009) 813–821
tFiiahttift
maipttew(oStIrcetcStdt
3
ntjewhaWaaviatfan
tv(DtoaioC
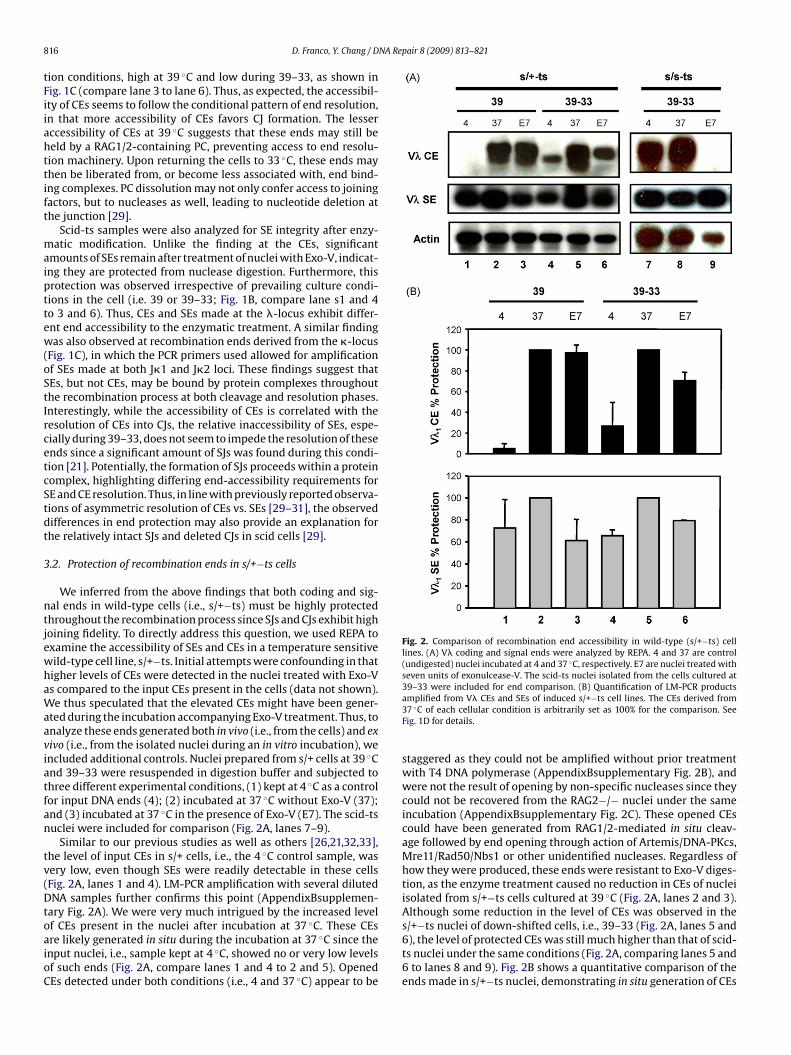
Fig. 2. Comparison of recombination end accessibility in wild-type (s/+−ts) celllines. (A) V� coding and signal ends were analyzed by REPA. 4 and 37 are control(undigested) nuclei incubated at 4 and 37 ◦C, respectively. E7 are nuclei treated withseven units of exonulcease-V. The scid-ts nuclei isolated from the cells cultured at39–33 were included for end comparison. (B) Quantification of LM-PCR products
16 D. Franco, Y. Chang / D
ion conditions, high at 39 ◦C and low during 39–33, as shown inig. 1C (compare lane 3 to lane 6). Thus, as expected, the accessibil-ty of CEs seems to follow the conditional pattern of end resolution,n that more accessibility of CEs favors CJ formation. The lesserccessibility of CEs at 39 ◦C suggests that these ends may still beeld by a RAG1/2-containing PC, preventing access to end resolu-ion machinery. Upon returning the cells to 33 ◦C, these ends mayhen be liberated from, or become less associated with, end bind-ng complexes. PC dissolution may not only confer access to joiningactors, but to nucleases as well, leading to nucleotide deletion athe junction [29].
Scid-ts samples were also analyzed for SE integrity after enzy-atic modification. Unlike the finding at the CEs, significant
mounts of SEs remain after treatment of nuclei with Exo-V, indicat-ng they are protected from nuclease digestion. Furthermore, thisrotection was observed irrespective of prevailing culture condi-ions in the cell (i.e. 39 or 39–33; Fig. 1B, compare lane s1 and 4o 3 and 6). Thus, CEs and SEs made at the �-locus exhibit differ-nt end accessibility to the enzymatic treatment. A similar findingas also observed at recombination ends derived from the �-locus
Fig. 1C), in which the PCR primers used allowed for amplificationf SEs made at both J�1 and J�2 loci. These findings suggest thatEs, but not CEs, may be bound by protein complexes throughouthe recombination process at both cleavage and resolution phases.nterestingly, while the accessibility of CEs is correlated with theesolution of CEs into CJs, the relative inaccessibility of SEs, espe-ially during 39–33, does not seem to impede the resolution of thesends since a significant amount of SJs was found during this condi-ion [21]. Potentially, the formation of SJs proceeds within a proteinomplex, highlighting differing end-accessibility requirements forE and CE resolution. Thus, in line with previously reported observa-ions of asymmetric resolution of CEs vs. SEs [29–31], the observedifferences in end protection may also provide an explanation forhe relatively intact SJs and deleted CJs in scid cells [29].
.2. Protection of recombination ends in s/+−ts cells
We inferred from the above findings that both coding and sig-al ends in wild-type cells (i.e., s/+−ts) must be highly protectedhroughout the recombination process since SJs and CJs exhibit highoining fidelity. To directly address this question, we used REPA toxamine the accessibility of SEs and CEs in a temperature sensitiveild-type cell line, s/+−ts. Initial attempts were confounding in thatigher levels of CEs were detected in the nuclei treated with Exo-Vs compared to the input CEs present in the cells (data not shown).e thus speculated that the elevated CEs might have been gener-
ted during the incubation accompanying Exo-V treatment. Thus, tonalyze these ends generated both in vivo (i.e., from the cells) and exivo (i.e., from the isolated nuclei during an in vitro incubation), wencluded additional controls. Nuclei prepared from s/+ cells at 39 ◦Cnd 39–33 were resuspended in digestion buffer and subjected tohree different experimental conditions, (1) kept at 4 ◦C as a controlor input DNA ends (4); (2) incubated at 37 ◦C without Exo-V (37);nd (3) incubated at 37 ◦C in the presence of Exo-V (E7). The scid-tsuclei were included for comparison (Fig. 2A, lanes 7–9).
Similar to our previous studies as well as others [26,21,32,33],he level of input CEs in s/+ cells, i.e., the 4 ◦C control sample, wasery low, even though SEs were readily detectable in these cellsFig. 2A, lanes 1 and 4). LM-PCR amplification with several dilutedNA samples further confirms this point (AppendixBsupplemen-
ary Fig. 2A). We were very much intrigued by the increased level
f CEs present in the nuclei after incubation at 37 ◦C. These CEsre likely generated in situ during the incubation at 37 ◦C since thenput nuclei, i.e., sample kept at 4 ◦C, showed no or very low levelsf such ends (Fig. 2A, compare lanes 1 and 4 to 2 and 5). OpenedEs detected under both conditions (i.e., 4 and 37 ◦C) appear to beamplified from V� CEs and SEs of induced s/+−ts cell lines. The CEs derived from37 ◦C of each cellular condition is arbitrarily set as 100% for the comparison. SeeFig. 1D for details.
staggered as they could not be amplified without prior treatmentwith T4 DNA polymerase (AppendixBsupplementary Fig. 2B), andwere not the result of opening by non-specific nucleases since theycould not be recovered from the RAG2−/− nuclei under the sameincubation (AppendixBsupplementary Fig. 2C). These opened CEscould have been generated from RAG1/2-mediated in situ cleav-age followed by end opening through action of Artemis/DNA-PKcs,Mre11/Rad50/Nbs1 or other unidentified nucleases. Regardless ofhow they were produced, these ends were resistant to Exo-V diges-tion, as the enzyme treatment caused no reduction in CEs of nucleiisolated from s/+−ts cells cultured at 39 ◦C (Fig. 2A, lanes 2 and 3).Although some reduction in the level of CEs was observed in thes/+−ts nuclei of down-shifted cells, i.e., 39–33 (Fig. 2A, lanes 5 and
6), the level of protected CEs was still much higher than that of scid-ts nuclei under the same conditions (Fig. 2A, comparing lanes 5 and6 to lanes 8 and 9). Fig. 2B shows a quantitative comparison of theends made in s/+−ts nuclei, demonstrating in situ generation of CEs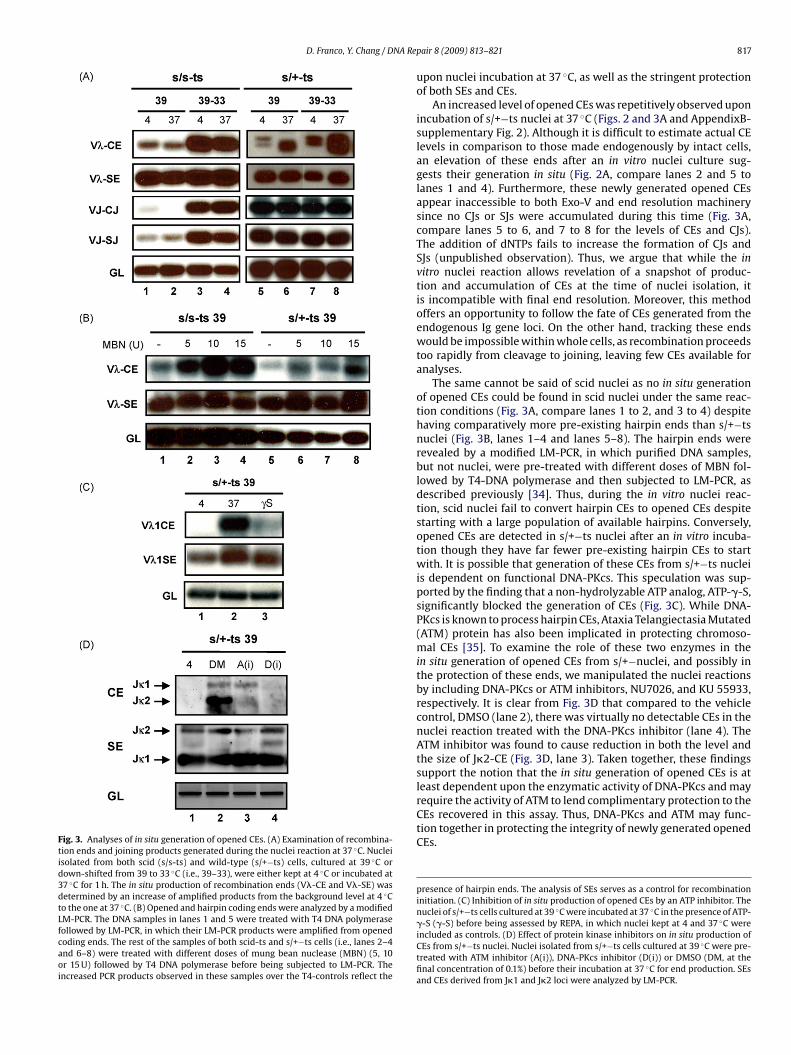
D. Franco, Y. Chang / DNA Re
Fig. 3. Analyses of in situ generation of opened CEs. (A) Examination of recombina-tion ends and joining products generated during the nuclei reaction at 37 ◦C. Nucleiisolated from both scid (s/s-ts) and wild-type (s/+−ts) cells, cultured at 39 ◦C ordown-shifted from 39 to 33 ◦C (i.e., 39–33), were either kept at 4 ◦C or incubated at37 ◦C for 1 h. The in situ production of recombination ends (V�-CE and V�-SE) wasdetermined by an increase of amplified products from the background level at 4 ◦Cto the one at 37 ◦C. (B) Opened and hairpin coding ends were analyzed by a modifiedLM-PCR. The DNA samples in lanes 1 and 5 were treated with T4 DNA polymerasefollowed by LM-PCR, in which their LM-PCR products were amplified from openedcoding ends. The rest of the samples of both scid-ts and s/+−ts cells (i.e., lanes 2–4and 6–8) were treated with different doses of mung bean nuclease (MBN) (5, 10or 15 U) followed by T4 DNA polymerase before being subjected to LM-PCR. Theincreased PCR products observed in these samples over the T4-controls reflect the
pair 8 (2009) 813–821 817
upon nuclei incubation at 37 ◦C, as well as the stringent protectionof both SEs and CEs.
An increased level of opened CEs was repetitively observed uponincubation of s/+−ts nuclei at 37 ◦C (Figs. 2 and 3A and AppendixB-supplementary Fig. 2). Although it is difficult to estimate actual CElevels in comparison to those made endogenously by intact cells,an elevation of these ends after an in vitro nuclei culture sug-gests their generation in situ (Fig. 2A, compare lanes 2 and 5 tolanes 1 and 4). Furthermore, these newly generated opened CEsappear inaccessible to both Exo-V and end resolution machinerysince no CJs or SJs were accumulated during this time (Fig. 3A,compare lanes 5 to 6, and 7 to 8 for the levels of CEs and CJs).The addition of dNTPs fails to increase the formation of CJs andSJs (unpublished observation). Thus, we argue that while the invitro nuclei reaction allows revelation of a snapshot of produc-tion and accumulation of CEs at the time of nuclei isolation, itis incompatible with final end resolution. Moreover, this methodoffers an opportunity to follow the fate of CEs generated from theendogenous Ig gene loci. On the other hand, tracking these endswould be impossible within whole cells, as recombination proceedstoo rapidly from cleavage to joining, leaving few CEs available foranalyses.
The same cannot be said of scid nuclei as no in situ generationof opened CEs could be found in scid nuclei under the same reac-tion conditions (Fig. 3A, compare lanes 1 to 2, and 3 to 4) despitehaving comparatively more pre-existing hairpin ends than s/+−tsnuclei (Fig. 3B, lanes 1–4 and lanes 5–8). The hairpin ends wererevealed by a modified LM-PCR, in which purified DNA samples,but not nuclei, were pre-treated with different doses of MBN fol-lowed by T4-DNA polymerase and then subjected to LM-PCR, asdescribed previously [34]. Thus, during the in vitro nuclei reac-tion, scid nuclei fail to convert hairpin CEs to opened CEs despitestarting with a large population of available hairpins. Conversely,opened CEs are detected in s/+−ts nuclei after an in vitro incuba-tion though they have far fewer pre-existing hairpin CEs to startwith. It is possible that generation of these CEs from s/+−ts nucleiis dependent on functional DNA-PKcs. This speculation was sup-ported by the finding that a non-hydrolyzable ATP analog, ATP-�-S,significantly blocked the generation of CEs (Fig. 3C). While DNA-PKcs is known to process hairpin CEs, Ataxia Telangiectasia Mutated(ATM) protein has also been implicated in protecting chromoso-mal CEs [35]. To examine the role of these two enzymes in thein situ generation of opened CEs from s/+−nuclei, and possibly inthe protection of these ends, we manipulated the nuclei reactionsby including DNA-PKcs or ATM inhibitors, NU7026, and KU 55933,respectively. It is clear from Fig. 3D that compared to the vehiclecontrol, DMSO (lane 2), there was virtually no detectable CEs in thenuclei reaction treated with the DNA-PKcs inhibitor (lane 4). TheATM inhibitor was found to cause reduction in both the level andthe size of J�2-CE (Fig. 3D, lane 3). Taken together, these findingssupport the notion that the in situ generation of opened CEs is atleast dependent upon the enzymatic activity of DNA-PKcs and may
require the activity of ATM to lend complimentary protection to theCEs recovered in this assay. Thus, DNA-PKcs and ATM may func-tion together in protecting the integrity of newly generated openedCEs.presence of hairpin ends. The analysis of SEs serves as a control for recombinationinitiation. (C) Inhibition of in situ production of opened CEs by an ATP inhibitor. Thenuclei of s/+−ts cells cultured at 39 ◦C were incubated at 37 ◦C in the presence of ATP-�-S (�-S) before being assessed by REPA, in which nuclei kept at 4 and 37 ◦C wereincluded as controls. (D) Effect of protein kinase inhibitors on in situ production ofCEs from s/+−ts nuclei. Nuclei isolated from s/+−ts cells cultured at 39 ◦C were pre-treated with ATM inhibitor (A(i)), DNA-PKcs inhibitor (D(i)) or DMSO (DM, at thefinal concentration of 0.1%) before their incubation at 37 ◦C for end production. SEsand CEs derived from J�1 and J�2 loci were analyzed by LM-PCR.

818 D. Franco, Y. Chang / DNA Repair 8 (2009) 813–821
Table 1Sequence analysis of opened coding ends recovered from nuclei of wild-type thymocytes.
Clone N/P-insertiona D�2-segment (N/Pb) J�1-segment
Germline ATCGGAGGGATACGAG CTACCGACAAACTCGTCJ�1-1c CTACCGACAAACTCGTCJ�1-2 GATGGCT CTACCGACAAACTCGTCJ�1-3 TGTCGGTAG CTACCGACAAACTCGTCD�2-1 GC ATCGGAGGGATACGAG CTACCGACAAACTCGTCD�2-2 CAT GGATACGAG CTACCGACAAACTCGTCD�2-3 GGATAGA TACCGACAAACTCGTCD�2-4 CATCCT ATCGGAGGGA TACCGACAAACTCGTCD�2-5 CATCAT ATCGGAGGGATAATTG CTACCGACAAACTCGTCD�2-6 GGCCCTCAGC CGGAGGGATACGAG CTACCGACAAACTCGTCD�2-7 TCTCTGCTATGAGTGGC ATCGGAGGGA TACCGACAAACTCGTCNS-1d 87BPS UPSTREAM OF J�1 CTACCGACAAACTCGTCNS-2d 164BPS UPSTREAM OF J�1 CTACCGACAAACTCGTCNS-3d TCR-V�-10 (256–334)e ATCGGAGGGATACGAGG CTACCGACAAACTCGTC
a Nucleotide insertion at the detected coding ends (J�1 or 5′D�2), of which the bold letters with italics are P-additions.b Letter in bold stands for nucleotide addition at the junction of DJ-coding joints between D�2 and J�1, in which the ones with italic are P-additions.
3t
pd(soscoaWsD(
CalstafJgdATlttittpnitpttr
c Observed in two individual mice.d Non-standard (NS) ends recovered.e Part of TCR-V�-10 sequences derived from position 256 to 334.
.3. Comparison of end susceptibility between wild-type and scidhymocytes
By using the REPA assay, we have assessed the association ofroteins with SEs and opened CEs under various experimental con-itions. Three conclusions can be drawn from the above findings:1) opened CEs could be generated in situ from wild-type, but notcid nuclei during their incubation at 37 ◦C (Figs. 2 and 3); (2)pened CEs derived from wild-type cells are not readily acces-ible to Exo-V, and exhibit better protection than those in scidells (Figs. 1 and 2); and (3) SEs are much better protected thanpened CEs in the same scid cells (Fig. 1). To verify our approachnd conclusion, we extended our analyses to primary thymocytes.e selected the TCR�-DJ locus for analyzing the end accessibility of
cid and wild-type thymocyte nuclei, since both J�1coding and 3′
�2 signal ends can be revealed in the same LM-PCR amplificationFig. 4A).
Consistent with previous reports [28,36,37], very few openedEs were found in wild-type thymocytes, whereas a significantmount of these ends were detected in scid thymocytes (Fig. 4,anes 1 and 4). Like the observed in situ production of opened CEs in/+−ts nuclei, opened CEs were also generated from nuclei of wild-ype thymocytes during their incubation at 37 ◦C (Fig. 4A, lanes 1nd 2). The amplified LM-PCR products from these CEs were clonedor sequence analysis. As shown in Table 1, seven D�2-CEs and three�1-CEs display authentic CEs with relatively intact D�2 and/or J�1ene segments, in which the perfect J�1-CE (i.e., clone J�1-1) wasetected repetitively from thymocyte nuclei of two individual mice.dditionally, three non-standard (NS) ends were also recovered.wo were derived from the delta locus 87 or 164 bases 5′ of J�1ocus while the other contained a rearranged V�D�J�-joint, con-aining broken ends in the middle of the V�-region. It is possiblehat these ends resulted from non-RSS-dependent cleavage, reflect-ng the infidelity of end processing in isolated nuclei. Nonetheless,he majority of the CEs was authentic and appeared to be resistanto Exo-V digestion, implicating their association with protein com-lexes (Fig. 4C, lanes 2 and 3). In contrast, the opened CEs in theuclei of scid thymocytes were poorly protected. As demonstrated
n Fig. 4B, these CE ends were not only sensitive to Exo-V diges-
ion (lane 6), but also appear susceptible to endogenous nucleasesresent in the nuclei since the level of CEs at 37 ◦C was lower thanhat of the control, i.e., the nuclei at 4 ◦C (Fig. 4B, lane 4 and 5). Onhe other hand, a significant amount of intact 3′D�2-SEs was stilletained after Exo-V treatment of nuclei isolated from both scid andwild-type thymocytes (Fig. 4B, lanes 4–6 and lanes 1–3, top band).Thus, consistent with our finding in scid-ts cell lines, SEs generatedfrom both wild-type and scid thymocytes were largely protectedfrom enzymatic modification.
We can therefore conclude that opened CEs in primary scid thy-mocytes may be less, or unassociated with protective complexesmaking them available for alternative NHEJ resolution. CEs gener-ated in the DNA-PKcs-deficient background are more accessible toalternate end binding factors than their wild-type counterparts, butmay consequently be vulnerable to nucleases and aberrant joiningcommon to scid end resolution. Taken together with the ts cell linedata, we confirm that opened CEs are better protected in wild-typecells than in scid cells while SEs are more resistant to Exo-V thanCEs in both cell types.
4. Discussion
REPA offers a novel tool to examine the accessibility of newlygenerated chromosomal recombination breaks, and indirectlyassess the stability of protein complexes formed at these breaks,especially those that participate in the generation and resolution ofrecombination ends. While opening of hairpin CEs occurs en masseafter induction and before joining, there remain some open CEsat the transition from RAG to NHEJ, which can be influenced byDNA end management. We focused on the protection and accessi-bility of open CEs due to technical feasibility, and are less concernedwith the hairpin opening process. The presence and stability ofan end-binding complex relates to its ability to protect associatedDNA ends from digestion by exonucleases. The fact that recom-bination ends are relatively inaccessible in wild-type cells arguesthat these ends are likely held within protein complexes through-out the recombination reaction. Conversely, open CEs in scid-tscells display conditional accessibility, i.e., less accessible duringRAG1/2-mediated cleavage at 39 ◦C, and more accessible after cellsare returned from 39 to 33 ◦C (Fig. 1), where RAG proteins aredown-regulated and rearranged CJs are formed (AppendixBsupple-mentary Fig. 1A). Scid CE resolution in this system is presumablymediated by a DNA-PK-independent alternative pathway, a back-upsystem that is apparent in other NHEJ mutant cells [38]. Several well
executed investigations have identified RAG1/2 as a hub, linkingV(D)J recombination ends to their principle end joining program,NHEJ (14, 39). RAG mutations affect the stability of PCs, makingrecombination ends available to alternate repair [14,15]. These stud-ies argue that pre-mature release of recombination ends from the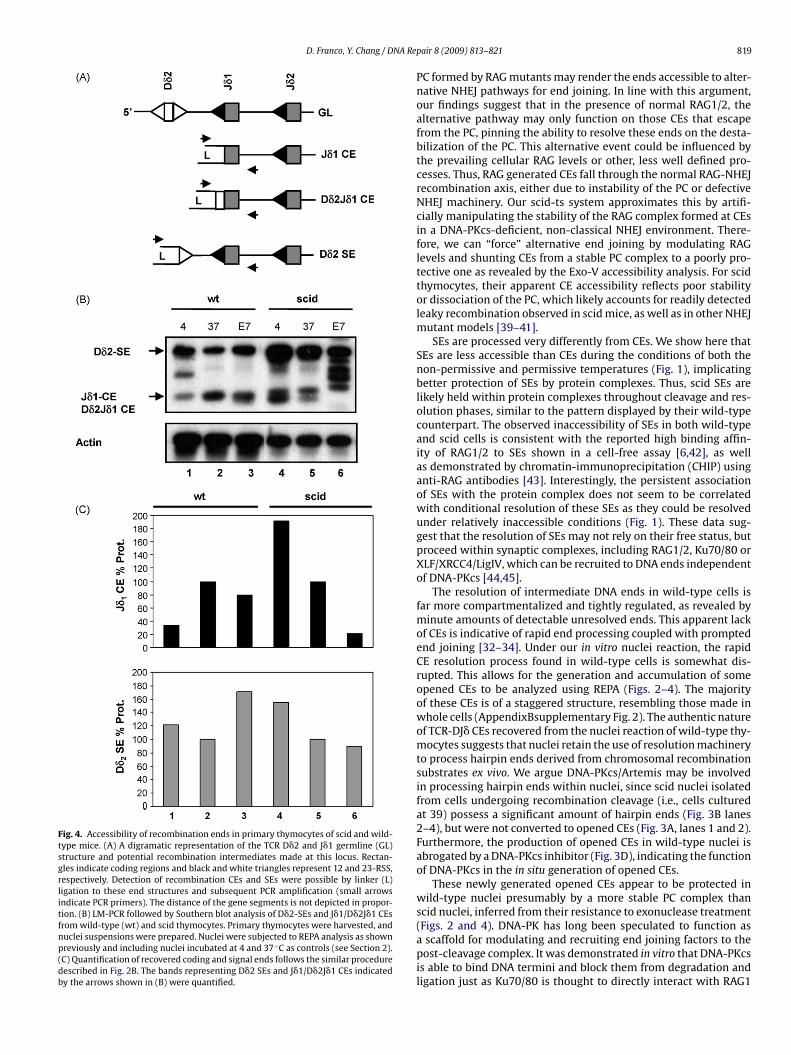
D. Franco, Y. Chang / DNA Re
Fig. 4. Accessibility of recombination ends in primary thymocytes of scid and wild-type mice. (A) A digramatic representation of the TCR D�2 and J�1 germline (GL)structure and potential recombination intermediates made at this locus. Rectan-gles indicate coding regions and black and white triangles represent 12 and 23-RSS,respectively. Detection of recombination CEs and SEs were possible by linker (L)ligation to these end structures and subsequent PCR amplification (small arrowsindicate PCR primers). The distance of the gene segments is not depicted in propor-tion. (B) LM-PCR followed by Southern blot analysis of D�2-SEs and J�1/D�2J�1 CEsfrom wild-type (wt) and scid thymocytes. Primary thymocytes were harvested, andnuclei suspensions were prepared. Nuclei were subjected to REPA analysis as shownpreviously and including nuclei incubated at 4 and 37 ◦C as controls (see Section 2).(C) Quantification of recovered coding and signal ends follows the similar proceduredescribed in Fig. 2B. The bands representing D�2 SEs and J�1/D�2J�1 CEs indicatedby the arrows shown in (B) were quantified.
pair 8 (2009) 813–821 819
PC formed by RAG mutants may render the ends accessible to alter-native NHEJ pathways for end joining. In line with this argument,our findings suggest that in the presence of normal RAG1/2, thealternative pathway may only function on those CEs that escapefrom the PC, pinning the ability to resolve these ends on the desta-bilization of the PC. This alternative event could be influenced bythe prevailing cellular RAG levels or other, less well defined pro-cesses. Thus, RAG generated CEs fall through the normal RAG-NHEJrecombination axis, either due to instability of the PC or defectiveNHEJ machinery. Our scid-ts system approximates this by artifi-cially manipulating the stability of the RAG complex formed at CEsin a DNA-PKcs-deficient, non-classical NHEJ environment. There-fore, we can “force” alternative end joining by modulating RAGlevels and shunting CEs from a stable PC complex to a poorly pro-tective one as revealed by the Exo-V accessibility analysis. For scidthymocytes, their apparent CE accessibility reflects poor stabilityor dissociation of the PC, which likely accounts for readily detectedleaky recombination observed in scid mice, as well as in other NHEJmutant models [39–41].
SEs are processed very differently from CEs. We show here thatSEs are less accessible than CEs during the conditions of both thenon-permissive and permissive temperatures (Fig. 1), implicatingbetter protection of SEs by protein complexes. Thus, scid SEs arelikely held within protein complexes throughout cleavage and res-olution phases, similar to the pattern displayed by their wild-typecounterpart. The observed inaccessibility of SEs in both wild-typeand scid cells is consistent with the reported high binding affin-ity of RAG1/2 to SEs shown in a cell-free assay [6,42], as wellas demonstrated by chromatin-immunoprecipitation (CHIP) usinganti-RAG antibodies [43]. Interestingly, the persistent associationof SEs with the protein complex does not seem to be correlatedwith conditional resolution of these SEs as they could be resolvedunder relatively inaccessible conditions (Fig. 1). These data sug-gest that the resolution of SEs may not rely on their free status, butproceed within synaptic complexes, including RAG1/2, Ku70/80 orXLF/XRCC4/LigIV, which can be recruited to DNA ends independentof DNA-PKcs [44,45].
The resolution of intermediate DNA ends in wild-type cells isfar more compartmentalized and tightly regulated, as revealed byminute amounts of detectable unresolved ends. This apparent lackof CEs is indicative of rapid end processing coupled with promptedend joining [32–34]. Under our in vitro nuclei reaction, the rapidCE resolution process found in wild-type cells is somewhat dis-rupted. This allows for the generation and accumulation of someopened CEs to be analyzed using REPA (Figs. 2–4). The majorityof these CEs is of a staggered structure, resembling those made inwhole cells (AppendixBsupplementary Fig. 2). The authentic natureof TCR-DJ� CEs recovered from the nuclei reaction of wild-type thy-mocytes suggests that nuclei retain the use of resolution machineryto process hairpin ends derived from chromosomal recombinationsubstrates ex vivo. We argue DNA-PKcs/Artemis may be involvedin processing hairpin ends within nuclei, since scid nuclei isolatedfrom cells undergoing recombination cleavage (i.e., cells culturedat 39) possess a significant amount of hairpin ends (Fig. 3B lanes2–4), but were not converted to opened CEs (Fig. 3A, lanes 1 and 2).Furthermore, the production of opened CEs in wild-type nuclei isabrogated by a DNA-PKcs inhibitor (Fig. 3D), indicating the functionof DNA-PKcs in the in situ generation of opened CEs.
These newly generated opened CEs appear to be protected inwild-type nuclei presumably by a more stable PC complex thanscid nuclei, inferred from their resistance to exonuclease treatment
(Figs. 2 and 4). DNA-PK has long been speculated to function asa scaffold for modulating and recruiting end joining factors to thepost-cleavage complex. It was demonstrated in vitro that DNA-PKcsis able to bind DNA termini and block them from degradation andligation just as Ku70/80 is thought to directly interact with RAG1
8 NA Re
[PtsiDctdv
C
A
ycUDTDF
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
20 D. Franco, Y. Chang / D
46]. In addition, ATM has also been reported to maintain CEs inC complexes [47]. In support of this study, we found a reduc-ion in the amount and relative size of J�2-CEs recovered from the/+−nuclei reaction treated with an ATM inhibitor, thus confirm-ng the role of ATM in maintaining CE integrity. It is possible thatNA-PKcs may cooperate with ATM, forming a large scaffoldingomplex that functions to retain CEs for proper joining. It remainso be determined how DNA-PKcs protect these ends, either throughirect interaction or more indirectly by stimulating end protectionia ATM modification.
onflict of interest
None.
cknowledgements
We thank Mr. Jason Noble for his assistance in CE sequence anal-ses, Drs. F. Alt and M. Schlissel for providing us a RAG2−/− pro-Bell line. We greatly appreciate technical support from Arizona Stateniversity W. M. Keck Lab for Phospho-imaging analyses and theNA Laboratory at the School of Life Sciences for DNA sequencing.his work was partly supported by a NIH grant CA73857 (to Y.C., and. F. for supplemental support) and a Minority Graduate Educationellowship (to D. F.).
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.dnarep.2009.03.007.
eferences
[1] M. Gellert, V(D)J recombination: RAG proteins, repair factors, and regulation,Annu. Rev. Biochem. 71 (2002) 101–132.
[2] D. Jung, F.W. Alt, Unraveling V(D)J recombination; insights into gene regulation,Cell 116 (2004) 299–311.
[3] Q.M. Eastman, T.M. Leu, D.G. Schatz, Initiation of V(D)J recombination in vitroobeying the 12/23 rule, Nature 380 (1996) 85–88.
[4] J.F. McBlane, D.C. van Gent, D.A. Ramsden, C. Romeo, C.A. Cuomo, M. Gellert,M.A. Oettinger, Cleavage at a V(D)J recombination signal requires only RAG1and RAG2 proteins and occurs in two steps, Cell 83 (1995) 387–395.
[5] D.C. van Gent, J.F. McBlane, D.A. Ramsden, M.J. Sadofsky, J.E. Hesse, M.Gellert, Initiation of V(D)J recombination in a cell-free system, Cell 81 (1995)925–934.
[6] A. Agrawal, D.G. Schatz, RAG1 and RAG2 form a stable postcleavage synapticcomplex with DNA containing signal ends in V(D)J recombination, Cell 89 (1997)43–53.
[7] K. Hiom, M. Gellert, A stable RAG1–RAG2–DNA complex that is active in V(D)Jcleavage, Cell 88 (1997) 65–72.
[8] T. Blunt, N.J. Finnie, G.E. Taccioli, G.C. Smith, J. Demengeot, T.M. Gottlieb, R.Mizuta, A.J. Varghese, F.W. Alt, P.A. Jeggo, et al., Defective DNA-dependent pro-tein kinase activity is linked to V(D)J recombination and DNA repair defectsassociated with the murine scid mutation, Cell 80 (1995) 813–823.
[9] U. Grawunder, M. Wilm, X. Wu, P. Kulesza, T.E. Wilson, M. Mann, M.R. Lieber,Activity of DNA ligase IV stimulated by complex formation with XRCC4 proteinin mammalian cells, Nature 388 (1997) 492–495.
10] Z. Li, T. Otevrel, Y. Gao, H.L. Cheng, B. Seed, T.D. Stamato, G.E. Taccioli, F.W. Alt,The XRCC4 gene encodes a novel protein involved in DNA double-strand breakrepair and V(D)J recombination, Cell 83 (1995) 1079–1089.
11] Y. Ma, U. Pannicke, H. Lu, D. Niewolik, K. Schwarz, M.R. Lieber, The DNA-dependent protein kinase catalytic subunit phosphorylation sites in humanArtemis, J. Biol. Chem. 280 (2005) 33839–33846.
12] Y. Ma, U. Pannicke, K. Schwarz, M.R. Lieber, Hairpin opening and overhangprocessing by an Artemis/DNA-dependent protein kinase complex in nonho-mologous end joining and V(D)J recombination, Cell 108 (2002) 781–794.
13] M.L. Brown, D. Franco, A. Burkle, Y. Chang, Role of poly(ADP-ribosyl)ation inDNA-PKcs-independent V(D)J recombination, Proc. Natl. Acad. Sci. U.S.A. 99(2002) 4532–4537.
14] B. Corneo, R.L. Wendland, L. Deriano, X. Cui, I.A. Klein, S.Y. Wong, S. Arnal,
A.J. Holub, G.R. Weller, B.A. Pancake, S. Shah, V.L. Brandt, K. Meek, D.B.Roth, Rag mutations reveal robust alternative end joining, Nature 449 (2007)483–486.15] G.S. Lee, M.B. Neiditch, S.S. Salus, D.B. Roth, RAG proteins shepherd double-strand breaks to a specific pathway, suppressing error-prone repair, but RAGnicking initiates homologous recombination, Cell 117 (2004) 171–184.
[
[
pair 8 (2009) 813–821
16] M. Audebert, B. Salles, P. Calsou, Effect of double-strand break DNA sequenceon the PARP-1 NHEJ pathway, Biochem. Biophys. Res. Commun. 369 (2008)982–988.
17] C. Couedel, K.D. Mills, M. Barchi, L. Shen, A. Olshen, R.D. Johnson, A. Nussen-zweig, J. Essers, R. Kanaar, G.C. Li, F.W. Alt, M. Jasin, Collaboration of homologousrecombination and nonhomologous end-joining factors for the survival andintegrity of mice and cells, Genes Dev. 18 (2004) 1293–1304.
18] H.C. Wang, W.C. Chou, S.Y. Shieh, C.Y. Shen, Ataxia telangiectasia mutated andcheckpoint kinase 2 regulate BRCA1 to promote the fidelity of DNA end-joining,Cancer Res. 66 (2006) 1391–1400.
19] W. Schuler, N.R. Ruetsch, M. Amsler, M.J. Bosma, Coding joint formation ofendogenous T cell receptor genes in lymphoid cells from scid mice: unusualP-nucleotide additions in VJ-coding joints, Eur. J. Immunol. 21 (1991) 589–596.
20] G.E. Taccioli, G. Rathbun, E. Oltz, T. Stamato, P.A. Jeggo, F.W. Alt, Impairment ofV(D)J recombination in double-strand break repair mutants, Science 260 (1993)207–210.
21] M.L. Brown, Y. Chang, Metabolism of recombination coding ends in scid cells, J.Immunol. 164 (2000) 4135–4142.
22] J.M. Jones, M. Gellert, Intermediates in V(D)J recombination: a stable RAG1/2complex sequesters cleaved RSS ends, Proc. Natl. Acad. Sci. U.S.A. 98 (2001)12926–12931.
23] T.M. Leu, Q.M. Eastman, D.G. Schatz, Coding joint formation in a cell-free V(D)Jrecombination system, Immunity 7 (1997) 303–314.
24] D.A. Ramsden, T.T. Paull, M. Gellert, Cell-free V(D)J recombination, Nature 388(1997) 488–491.
25] H. Yamagishi, T. Tsuda, S. Fujimoto, M. Toda, K. Kato, Y. Maekawa, M. Umeno, M.Anai, Purification of small polydisperse circular DNA of eukaryotic cells by useof ATP-dependent deoxyribonuclease, Gene 26 (1983) 317–321.
26] Y. Chang, M.L. Brown, Formation of coding joints in V(D)J recombination-inducible severe combined immune deficient pre-B cell lines, Proc. Natl. Acad.Sci. U.S.A. 96 (1999) 191–196.
27] D.B. Roth, C. Zhu, M. Gellert, Characterization of broken DNA moleculesassociated with V(D)J recombination, Proc. Natl. Acad. Sci. U.S.A. 90 (1993)10788–10792.
28] P.B. Nakajima, M.J. Bosma, Variable diversity joining recombination: nonhairpincoding ends in thymocytes of SCID and wild-type mice, J. Immunol. 169 (2002)3094–3104.
29] M.R. Lieber, J.E. Hesse, S. Lewis, G.C. Bosma, N. Rosenberg, K. Mizuuchi, M.J.Bosma, M. Gellert, The defect in murine severe combined immune deficiency:joining of signal sequences but not coding segments in V(D)J recombination,Cell 55 (1988) 7–16.
30] K.M. Sheehan, M.R. Lieber, V(D)J recombination: signal and coding joint resolu-tion are uncoupled and depend on parallel synapsis of the sites, Mol. Cell. Biol.13 (1993) 1363–1370.
31] S.M. Lewis, J.E. Hesse, K. Mizuuchi, M. Gellert, Novel strand exchanges in V(D)Jrecombination, Cell 55 (1988) 1099–1107.
32] D.A. Ramsden, M. Gellert, Formation and resolution of double-strand breakintermediates in V(D)J rearrangement, Genes Dev. 9 (1995) 2409–2420.
33] M. Schlissel, Structure of nonhairpin coding-end DNA breaks in cells undergoingV(D)J recombination, Mol. Cell. Biol. 18 (1998) 2029–2037.
34] C. Zhu, D.B. Roth, Characterization of coding ends in thymocytes of scidmice:implications for the mechanism of V(D)J recombination, Immunity 2(1995) 101–112.
35] A.L. Bredemeyer, G.G. Sharma, C.Y. Huang, B.A. Helmink, L.M. Walker, K.C. Khor,B. Nuskey, K.E. Sullivan, T.K. Pandita, C.H. Bassing, B.P. Sleckman, ATM stabi-lizes DNA double-strand-break complexes during V(D)J recombination, Nature(2006) 466–470.
36] D.B. Roth, J.P. Menetski, P.B. Nakajima, M.J. Bosma, M. Gellert, V(D)J recombi-nation: Broken DNA molecules with covalently sealed (hairpin) coding ends inscid mouse thymocytes, Cell 70 (1992) 983–991.
37] P.B. Nakajima, M.J. Bosma, Characterization of excised DNA intermediates asso-ciated with V(D)J recombination at the T-cell receptor delta locus., Mol. Cell.Biol. 17 (1997) 2631–2641.
38] N.S. Verkaik, R.E. Esveldt-van Lange, D. van Heemst, H.T. Bruggenwirth, J.H.Hoeijmakers, M.Z. Zdzienicka, D.C. van Gent, Different types of V(D)J recom-bination and end-joining defects in DNA double-strand break repair mutantmammalian cells, Eur. J. Immunol. 32 (2002) 701–709.
39] D.M. Weinstock, M. Jasin, Alternative pathways for the repair of RAG-inducedDNA breaks, Mol. Cell. Biol. 26 (2006) 131–139.
40] S. Rooney, J. Sekiguchi, C. Zhu, H.L. Cheng, J. Manis, S. Whitlow, J. DeVido, D.Foy, J. Chaudhuri, D. Lombard, F.W. Alt, Leaky Scid phenotype associated withdefective V(D)J coding end processing in Artemis-deficient mice, Mol. Cell. 10(2002) 1379–1390.
41] Y. Gu, K.J. Seidl, G.A. Rathbun, C. Zhu, J.P. Manis, N. van der Stoep, L. Davidson,H.L. Cheng, J.M. Sekiguchi, K. Frank, P. Stanhope-Baker, M.S. Schlissel, D.B. Roth,F.W. Alt, Growth retardation and leaky SCID phenotype of Ku70-deficient mice,Immunity 7 (1997) 653–665.
42] U. Grawunder, M.R. Lieber, A complex of RAG-1 and RAG-2 proteins persistson DNA after single-strand cleavage at V(D)J recombination signal sequences,
Nucleic Acids Res. 25 (1997) 1375–1382.43] E.J. Perkins, A. Nair, D.O. Cowley, T. Van Dyke, Y. Chang, D.A. Ramsden, Sensing ofintermediates in V(D)J recombination by ATM, Genes Dev. 16 (2002) 159–164.
44] K. Yano, D.J. Chen, Live cell imaging of XLF and XRCC4 reveals a novel viewof protein assembly in the non-homologous end-joining pathway, Cell Cycle 7(2008) 1321–1325.

NA Repair 8 (2009) 813–821 821
[
[
D. Franco, Y. Chang / D
45] K. Yano, K. Morotomi-Yano, S.Y. Wang, N. Uematsu, K.J. Lee, A. Asaithamby, E.Weterings, D.J. Chen, Ku recruits XLF to DNA double-strand breaks, EMBO Rep.9 (2008) 91–96.
46] E. Weterings, N.S. Verkaik, H.T. Bruggenwirth, J.H. Hoeijmakers, D.C. van Gent,The role of DNA dependent protein kinase in synapsis of DNA ends, NucleicAcids Res. 31 (2003) 7238–7246.
[
47] A.L. Bredemeyer, C.Y. Huang, L.M. Walker, C.H. Bassing, B.P. Sleckman, AberrantV(D)J recombination in ataxia telanglectasia mutated-deficient lymphocytesis dependent on nonhomologous DNA end joining, J. Immunol. 181 (2008)2620–2625.




























