Embed Size (px)
Citation preview

Journal ofOral Science & Rehabilitation
26 Volume 2 | Issue 2/2016
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s aH u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
Accelerated generation of human induced pluripotent stem cells from human oral mucosa using episomal plasmid vectors and maternal transcription factor Glis1
Takahiro Kashiwagi,a Yoshiya Hashimoto,b Masahiro Tanakac & Shunsuke Babaa
a Department of Oral Implantology, Osaka Dental University, Hirakata, Japan
b Department of Biomaterials, Osaka Dental University, Hirakata, Japan
c Department of Fixed Prosthodontics and Occlusion, Osaka Dental University, Hirakata, Japan
C o r r e s p o n d i n g a u t h o r :
Dr. Yoshiya Hashimoto
H o w t o c i t e t h i s a r t i c l e :
Kashiwagi T, Hashimoto Y, Tanaka M, Baba S. Accelerated generation of human induced pluripotent stem cells from human oral mucosa using episomal plasmid vectors and maternal transcription factor Glis1.
J Oral Science Rehabilitation.2016 Jun;2(2):26–35.
Abstract
O b j e c t i v e
Induced pluripotent stem cells (iPSCs) possess high pluripotency and dif-ferentiation potential and may constitute a possible source of autologous stem cells for clinical applications. However, the lengthy reprogramming process (up to one month) remains one of the most significant challenges facing standard virus-mediated methodology. The Gli-like transcription factor Glis1 is highly expressed in unfertilized eggs and one-cell-stage embryos. In this study, iPSCs were generated using a combination of pri-mary human oral mucosal fibroblasts (HOFs) and episomal plasmid vectors expressing transcription factors, including Glis1.
M a t e r i a l s a n d m e t h o d s
HOFs were established from oral mucosal tissue 3 mm in diameter from a 23-year-old Asian male using a skin trephine. Human iPSCs were gen-erated from the established HOFs using the following episomal plasmid vectors: pCXLE-hOCT3/4-shp53-F that expresses OCT3/4 and short- hairpin RNA (shRNA) against p53, pCXLE-hSK that expresses SOX2 and KLF4, pCXLE-hUL that expresses L-MYC and LIN28, and pCXLE-hGlis1 that expresses Glis1.
R e s u l t s
Fifty colonies of human embryonic stem (ES)-like cells were observed as early as 20 days after initial episomal plasmid vector transduction. The resulting cell lines shared several characteristics with human ES cells, including morphology, pluripotency-associated gene and protein markers, karyotype analysis and the ability to differentiate in vivo into all three germ layers.
C o n c l u s i o n
Our method, combining the use of HOFs and episomal plasmid vectors expressing OCT3/4, shRNA against p53, SOX2, KLF4, L-MYC, LIN28 and Glis1, offers a powerful tool for safely and rapidly generating bona fide human iPSCs and facilitates the application of iPSC technology to biomed-ical research.
K e y w o r d s
iPSC, integration-free plasmid vector, Glis1, human oral mucosal tissue.

Journal ofOral Science & Rehabilitation
Volume 2 | Issue 2/2016 27
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s aH u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
Introduction
The successful reprogramming of human and mouse somatic cells into induced pluripotent stem cells (iPSCs) via ectopic overexpression of pluripotency-associated transcription factors is considered a major scientific breakthrough.1–5 Similar to the characteristics of embryonic stem (ES) cells,6–8 human iPSCs can proliferate indef-initely, while retaining pluripotency, and can dif-ferentiate into all cell types found in the body. IPSCs have been generated from dermal fibro-blasts,3 peripheral blood,9 dental pulp cells,10 gingival fibroblasts,11 periodontal ligaments,12 oral mucosa13 and mesenchymal stromal cells.14
Gingival tissue is routinely resected during general dental treatments, such as tooth extrac-tion, periodontal surgery and dental implanta- tion, and generally treated as biomedical waste.15 Egusa et al. successfully derived iPSCs from human gingival fibroblasts (HGFs) using retro-viral transduction of transcription factors; they also reported that the reprogramming efficiency of mouse gingival fibroblasts was higher than that of dermal fibroblasts.11 However, retroviral integration increases the risk of tumor forma- tion, while integration-free methods decrease this potential risk.15 The development of novel approaches to generating integration-free iPSCs has eliminated the concern of integrating virus-associated genotoxicity in clinical applica-tions.16 Integration-free human iPSCs have been generated using several methods.15 Okita et al. reported a simple method that uses p53 sup-pression and nontransforming L-MYC to gene-rate human iPSCs with episomal plasmid vec-tors.15 Our recent study demonstrated that iPSCs could be generated from a combination of pri-mary HGFs and an episomal plasmid vector.17 However, the lengthy reprogramming process (up to one month) remains one of the most sig-nificant challenges facing standard virus- mediated methodology.
Maekawa et al. reported that the Gli-like transcription factor Glis1 (Glis family zinc finger 1) markedly enhances the generation of iPSCs from both mouse and human somatic fibroblasts when it is expressed together with three tran-scription factors collectively known as OSK (OCT3/4, SOX2 and KLF4) using retroviral trans-duction.18 However, little is known regarding whether Glis1 can effectively promote direct reprogramming during iPSC generation using an episomal plasmid vector. In the current study, iPSCs were generated by combining primary
human oral mucosal fibroblasts (HOFs) with episomal plasmid vectors expressing OCT3/4, short-hairpin RNA (shRNA) against p53, SOX2, KLF4, L-MYC, LIN28 and Glis1.
Materials and methods
E t h i c a l s t a t e m e n t
Approval for the sampling of human oral mucosa tissue, establishing iPSCs and genome/gene analysis was obtained from the Ethics Committee of Osaka Dental University, Hirakata, Japan (au-thorization No.: 110783; approval date: 30 Sep-tember 2013) and the DNA Recombination Ex-periment Safety Committee of Osaka Dental University (authorization No.: 54; approval date: 18 July 2014). Written informed consent was obtained from the participant. The animal exper-iments followed a protocol approved by the An-imal Committee of Osaka Dental University (au-thorization No.: 14-06002; approval date: 8 July 2014).
C e l l c u l t u r i n g
HOFs were established from oral mucosal tissue 3 mm in diameter obtained using a skin trephine (derma punch, Maruho, Osaka, Japan) from a 23-year-old Asian male. Human oral mucosal tissue was placed in 35 mm tissue culture dishes and cultured in Dulbecco’s Modification of Eagle’s Medium (DMEM) containing 10% fetal bovine serum at 37 °C and 5% CO2.11 The medium was replaced every three days. Once the HOFs had proliferated, the tissue was removed. When the cells reached subconfluence, they were dissoci-ated using 0.25% trypsin (Invitrogen, Carlsbad, Calif., U.S.) and transferred to 60 mm tissue cul-ture dishes (passage 1). HOFs were regularly passaged at a 1:3 ratio every three to four days.
G e n e r a t i o n o f i P S C s f r o m H O F s w i t h e p i s o m a l v e c t o r s
One microgram of an expression episomal plas-mid mixture containing pCXLE-hOCT3/4-shp53-F that expresses OCT3/4 and shRNA against p53, pCXLE-hSK that expresses SOX2 and KLF4, pCXLE-hUL that expresses L-MYC and LIN28, and pCXLE-hGlis1 that expresses Glis1 (Addgene, Cambridge, Mass., U.S.) was electro-porated into 6 × 105 primary HOFs (passage 5) with the Amaxa 4D-Nucleofector (Lonza, Basel,
Journal ofOral Science & Rehabilitation
28 Volume 2 | Issue 2/2016
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
Switzerland) according to the manufacturer’s instructions using program DT-130 (Lonza). These cells were then transferred on to mitomy-cin C-treated SNL 76/7 cells (cat. No. 07032801, lot No. 08F009; European Collection of Authen-ticated Cell Cultures, Porton Down, U.K.) at 5 × 104 cells per 100 mm dish. The following day, the culture medium was replaced with embry-onic stem cell (ESC) culture medium consisting of DMEM/F12 medium (Sigma-Aldrich, St. Louis, Mo., U.S.) supplemented with 20% Knock-Out Serum Replacement (Gibco, Grand Island, N.Y., U.S.), 2 mM L-glutamine (Nacalai Tesque, Kyoto, Japan), 1% nonessential amino acids (Gib-co), 0.1 mM 2-mercaptoethanol (Gibco) and 5 ng/mL fibroblast growth factor-2 (ReproCELL, Kanagawa, Japan). Thirty days subsequent to transduction, a number of colonies were mechan-ically picked and transferred to a 24-well plate. After several passages, ESC-like colonies were selected for further cultivation and characteriza-tion. IPSCs were generated and maintained in ESC culture medium. For routine passaging, iPSC colonies were detached with CTK solution (2.5 μg/mL trypsin, 1 mg/mL collagenase IV, 20% KSR, 1 mM CaCl2/PBS, and 70% PBS) and split at a 1:3 ratio every four to five days.
Q u a n t i t a t i v e r e a l - t i m e r e v e r s e t r a n s c r i p t i o n -
p o l y m e r a s e c h a i n r e a c t i o n
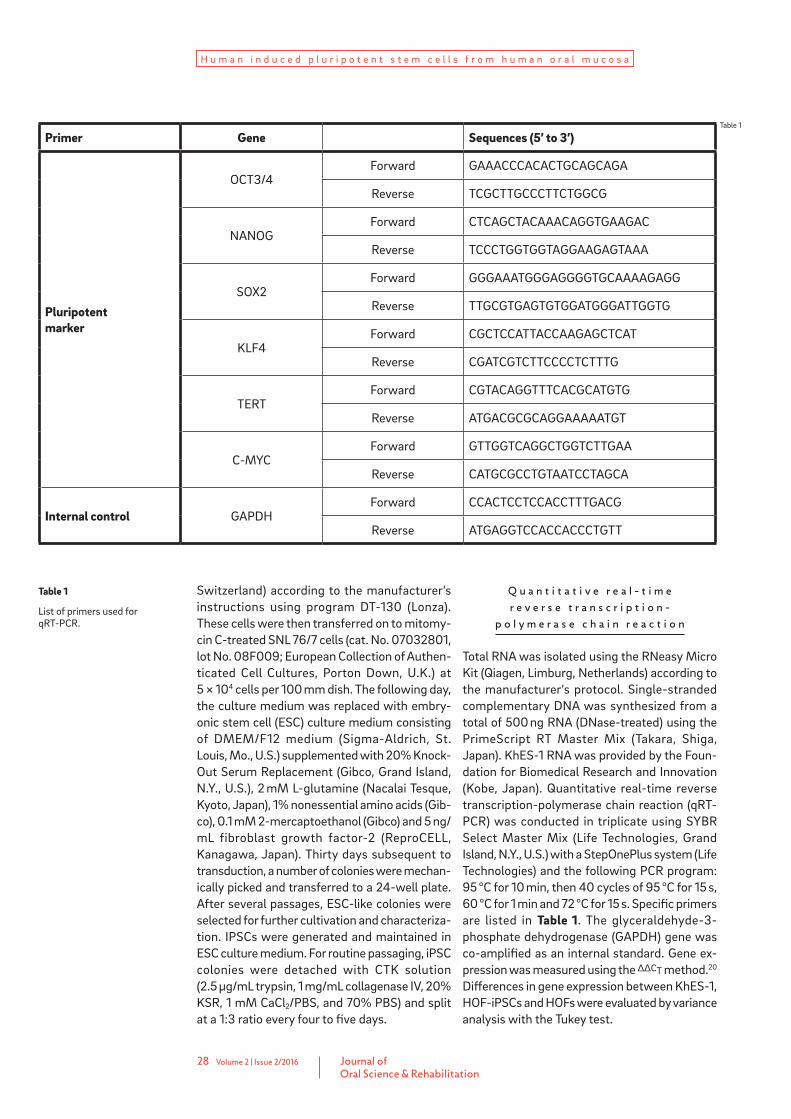
Total RNA was isolated using the RNeasy Micro Kit (Qiagen, Limburg, Netherlands) according to the manufacturer’s protocol. Single-stranded complementary DNA was synthesized from a total of 500 ng RNA (DNase-treated) using the PrimeScript RT Master Mix (Takara, Shiga, Japan). KhES-1 RNA was provided by the Foun-dation for Biomedical Research and Innovation (Kobe, Japan). Quantitative real-time reverse transcription-polymerase chain reaction (qRT-PCR) was conducted in triplicate using SYBR Select Master Mix (Life Technologies, Grand Island, N.Y., U.S.) with a StepOnePlus system (Life Technologies) and the following PCR program: 95 °C for 10 min, then 40 cycles of 95 °C for 15 s, 60 °C for 1 min and 72 °C for 15 s. Specific primers are listed in Table 1. The glyceraldehyde-3- phosphate dehydrogenase (GAPDH) gene was co-ampli fied as an internal standard. Gene ex-pression was measured using the ΔΔCT method.20
Differences in gene expression between KhES-1, HOF-iPSCs and HOFs were evaluated by variance analysis with the Tukey test.
Table 1
List of primers used for qRT-PCR.
Table 1
Primer Gene Sequences (5’ to 3’)
Pluripotent marker
OCT3/4Forward GAAACCCACACTGCAGCAGA
Reverse TCGCTTGCCCTTCTGGCG
NANOGForward CTCAGCTACAAACAGGTGAAGAC
Reverse TCCCTGGTGGTAGGAAGAGTAAA
SOX2Forward GGGAAATGGGAGGGGTGCAAAAGAGG
Reverse TTGCGTGAGTGTGGATGGGATTGGTG
KLF4Forward CGCTCCATTACCAAGAGCTCAT
Reverse CGATCGTCTTCCCCTCTTTG
TERTForward CGTACAGGTTTCACGCATGTG
Reverse ATGACGCGCAGGAAAAATGT
C-MYCForward GTTGGTCAGGCTGGTCTTGAA
Reverse CATGCGCCTGTAATCCTAGCA
Internal control GAPDHForward CCACTCCTCCACCTTTGACG
Reverse ATGAGGTCCACCACCCTGTT
Journal ofOral Science & Rehabilitation
Volume 2 | Issue 2/2016 29
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
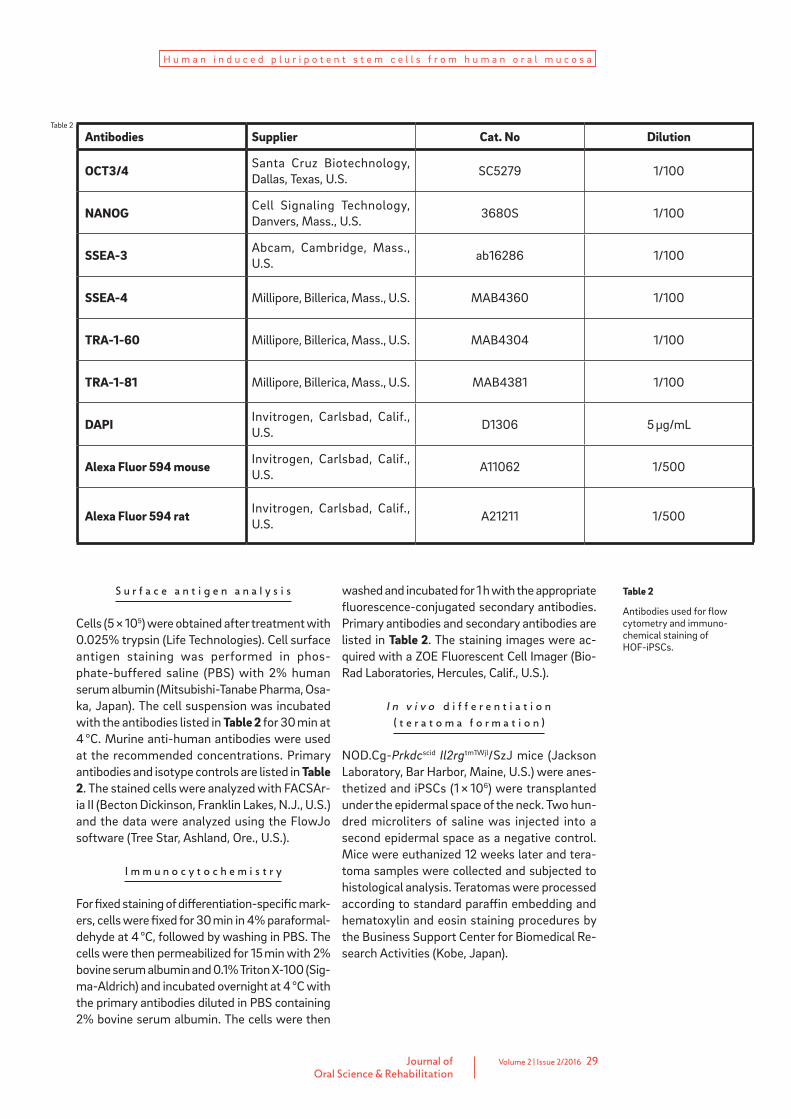
Table 2
Antibodies used for flow cytometry and immuno-chemical staining of HOF-iPSCs.
Table 2
S u r f a c e a n t i g e n a n a l y s i s
Cells (5 × 105) were obtained after treatment with 0.025% trypsin (Life Technologies). Cell surface antigen staining was performed in phos-phate-buffered saline (PBS) with 2% human serum albumin (Mitsubishi-Tanabe Pharma, Osa-ka, Japan). The cell suspension was incubated with the antibodies listed in Table 2 for 30 min at 4 °C. Murine anti-human antibodies were used at the recommended concentrations. Primary antibodies and isotype controls are listed in Table 2. The stained cells were analyzed with FACSAr-ia II (Becton Dickinson, Franklin Lakes, N.J., U.S.) and the data were analyzed using the FlowJo software (Tree Star, Ashland, Ore., U.S.).
I m m u n o c y t o c h e m i s t r y
For fixed staining of differentiation-specific mark-ers, cells were fixed for 30 min in 4% paraformal-dehyde at 4 °C, followed by washing in PBS. The cells were then permeabilized for 15 min with 2% bovine serum albumin and 0.1% Triton X-100 (Sig-ma-Aldrich) and incubated overnight at 4 °C with the primary antibodies diluted in PBS containing 2% bovine serum albumin. The cells were then
washed and incubated for 1 h with the appropriate fluorescence-conjugated secondary antibodies. Primary antibodies and secondary antibodies are listed in Table 2. The staining images were ac-quired with a ZOE Fluorescent Cell Imager (Bio-Rad Laboratories, Hercules, Calif., U.S.).
I n v i v o d i f f e r e n t i a t i o n ( t e r a t o m a f o r m a t i o n )
NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ mice (Jackson Laboratory, Bar Harbor, Maine, U.S.) were anes-thetized and iPSCs (1 × 106) were transplanted under the epidermal space of the neck. Two hun-dred microliters of saline was injected into a second epidermal space as a negative control. Mice were euthanized 12 weeks later and tera-toma samples were collected and subjected to histological analysis. Teratomas were processed according to standard paraffin embedding and hematoxylin and eosin staining procedures by the Business Support Center for Biomedical Re-search Activities (Kobe, Japan).
Antibodies Supplier Cat. No Dilution
OCT3/4 Santa Cruz Biotechnology, Dallas, Texas, U.S. SC5279 1/100
NANOG Cell Signaling Technology, Danvers, Mass., U.S. 3680S 1/100
SSEA-3 Abcam, Cambridge, Mass., U.S. ab16286 1/100
SSEA-4 Millipore, Billerica, Mass., U.S. MAB4360 1/100
TRA-1-60 Millipore, Billerica, Mass., U.S. MAB4304 1/100
TRA-1-81 Millipore, Billerica, Mass., U.S. MAB4381 1/100
DAPI Invitrogen, Carlsbad, Calif., U.S. D1306 5 μg/mL
Alexa Fluor 594 mouse Invitrogen, Carlsbad, Calif., U.S. A11062 1/500
Alexa Fluor 594 rat Invitrogen, Carlsbad, Calif., U.S. A21211 1/500
Journal ofOral Science & Rehabilitation
30 Volume 2 | Issue 2/2016
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
K a r y o t y p e a n a l y s i s
Chromosome G-band analysis was performed at Nihon Gene Research Laboratories (Sendai, Japan). At least 15 metaphases were analyzed.
Results
G e n e r a t i o n o f i P S C s f r o m H O F s u s i n g e p i s o m a l p l a s m i d v e c t o r s
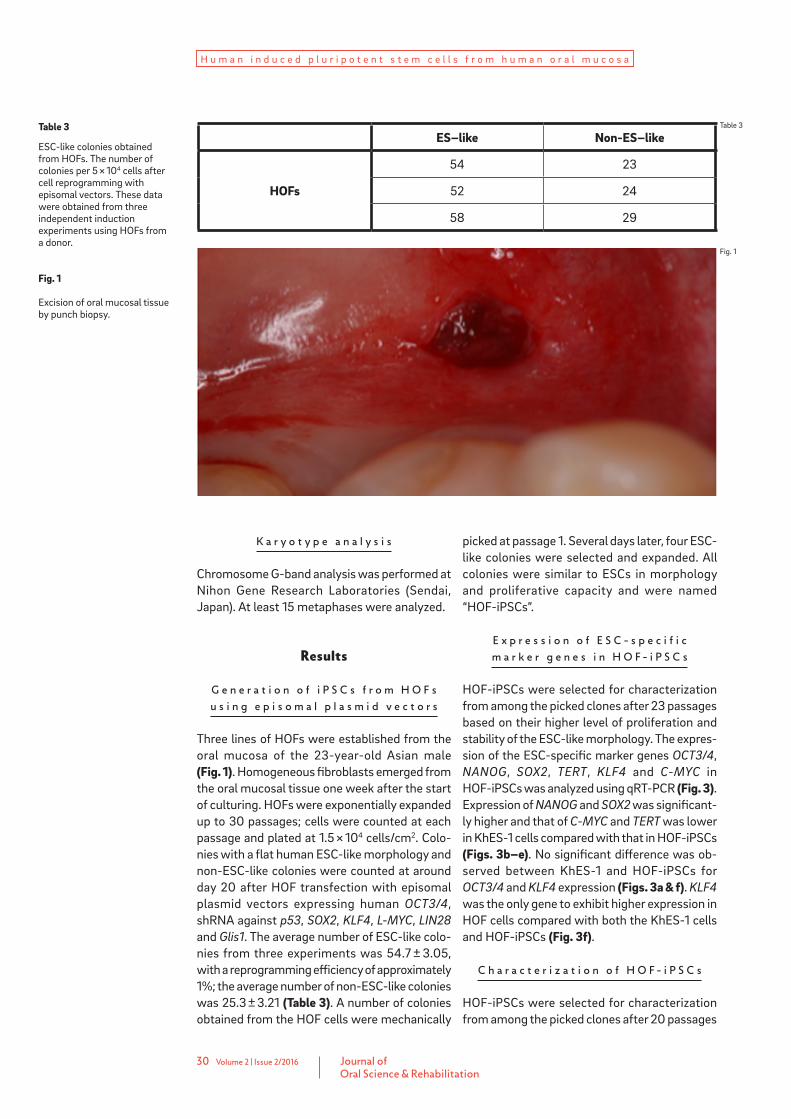
Three lines of HOFs were established from the oral mucosa of the 23-year-old Asian male (Fig. 1). Homogeneous fibroblasts emerged from the oral mucosal tissue one week after the start of culturing. HOFs were exponentially expanded up to 30 passages; cells were counted at each passage and plated at 1.5 × 104 cells/cm2. Colo-nies with a flat human ESC-like morphology and non-ESC-like colonies were counted at around day 20 after HOF transfection with episomal plasmid vectors expressing human OCT3/4, shRNA against p53, SOX2, KLF4, L-MYC, LIN28 and Glis1. The average number of ESC-like colo-nies from three experiments was 54.7 ± 3.05, with a reprogramming efficiency of approximately 1%; the average number of non-ESC-like colonies was 25.3 ± 3.21 (Table 3). A number of colonies obtained from the HOF cells were mechanically
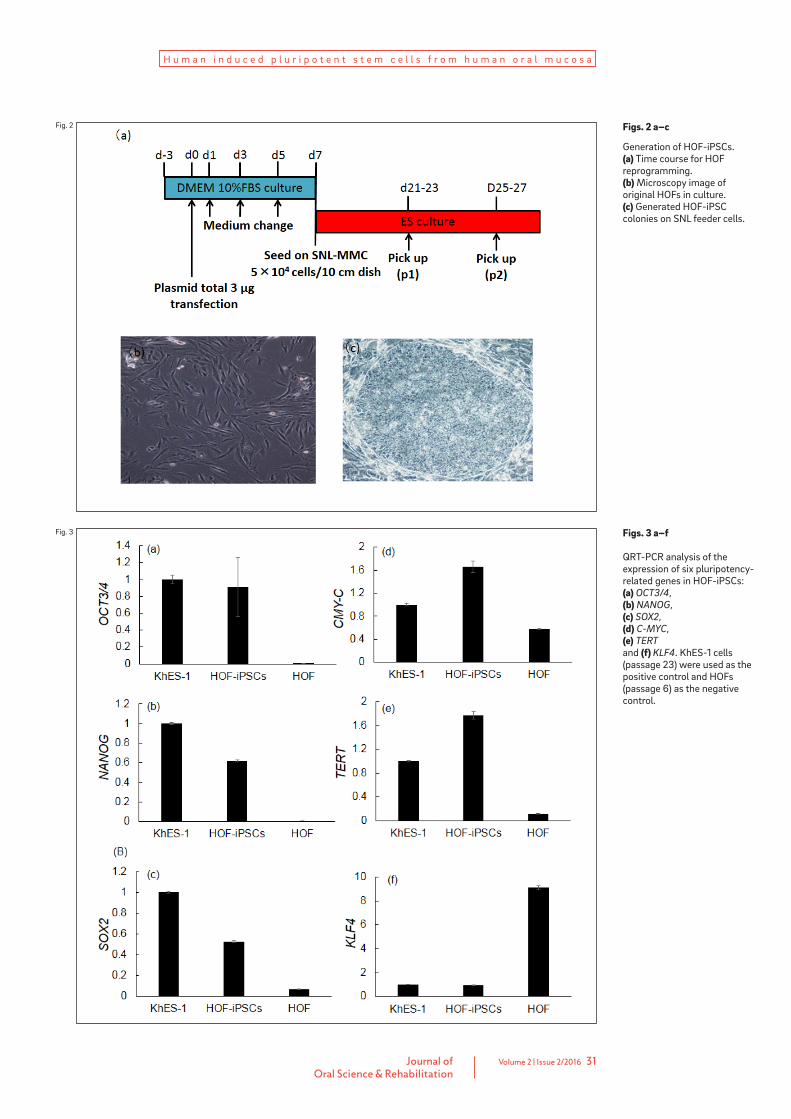
picked at passage 1. Several days later, four ESC-like colonies were selected and expanded. All colonies were similar to ESCs in morphology and proliferative capacity and were named “HOF-iPSCs”.
E x p r e s s i o n o f E S C - s p e c i f i c m a r k e r g e n e s i n H O F - i P S C s
HOF-iPSCs were selected for characterization from among the picked clones after 23 passages based on their higher level of proliferation and stability of the ESC-like morphology. The expres-sion of the ESC-specific marker genes OCT3/4, NANOG, SOX2, TERT, KLF4 and C-MYC in HOF-iPSCs was analyzed using qRT-PCR (Fig. 3). Expression of NANOG and SOX2 was significant-ly higher and that of C-MYC and TERT was lower in KhES-1 cells compared with that in HOF-iPSCs (Figs. 3b–e). No significant difference was ob-served between KhES-1 and HOF-iPSCs for OCT3/4 and KLF4 expression (Figs. 3a & f). KLF4 was the only gene to exhibit higher expression in HOF cells compared with both the KhES-1 cells and HOF-iPSCs (Fig. 3f).
C h a r a c t e r i z a t i o n o f H O F - i P S C s
HOF-iPSCs were selected for characterization from among the picked clones after 20 passages
Table 3
ESC-like colonies obtained from HOFs. The number of colonies per 5 × 104 cells after cell reprogramming with episomal vectors. These data were obtained from three independent induction experiments using HOFs from a donor.
Fig. 1
Excision of oral mucosal tissue by punch biopsy.
Table 3
Fig. 1
ES–like Non-ES–like
HOFs
54 23
52 24
58 29
Journal ofOral Science & Rehabilitation
Volume 2 | Issue 2/2016 31
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
Figs. 2 a–c
Generation of HOF-iPSCs. (a) Time course for HOF reprogramming. (b) Microscopy image of original HOFs in culture. (c) Generated HOF-iPSC colonies on SNL feeder cells.
Fig. 2
Fig. 3 Figs. 3 a–f
QRT-PCR analysis of the expression of six pluripotency -related genes in HOF-iPSCs: (a) OCT3/4, (b) NANOG, (c) SOX2, (d) C-MYC, (e) TERT and (f) KLF4. KhES-1 cells (passage 23) were used as the positive control and HOFs (passage 6) as the negative control.

Journal ofOral Science & Rehabilitation
32 Volume 2 | Issue 2/2016
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
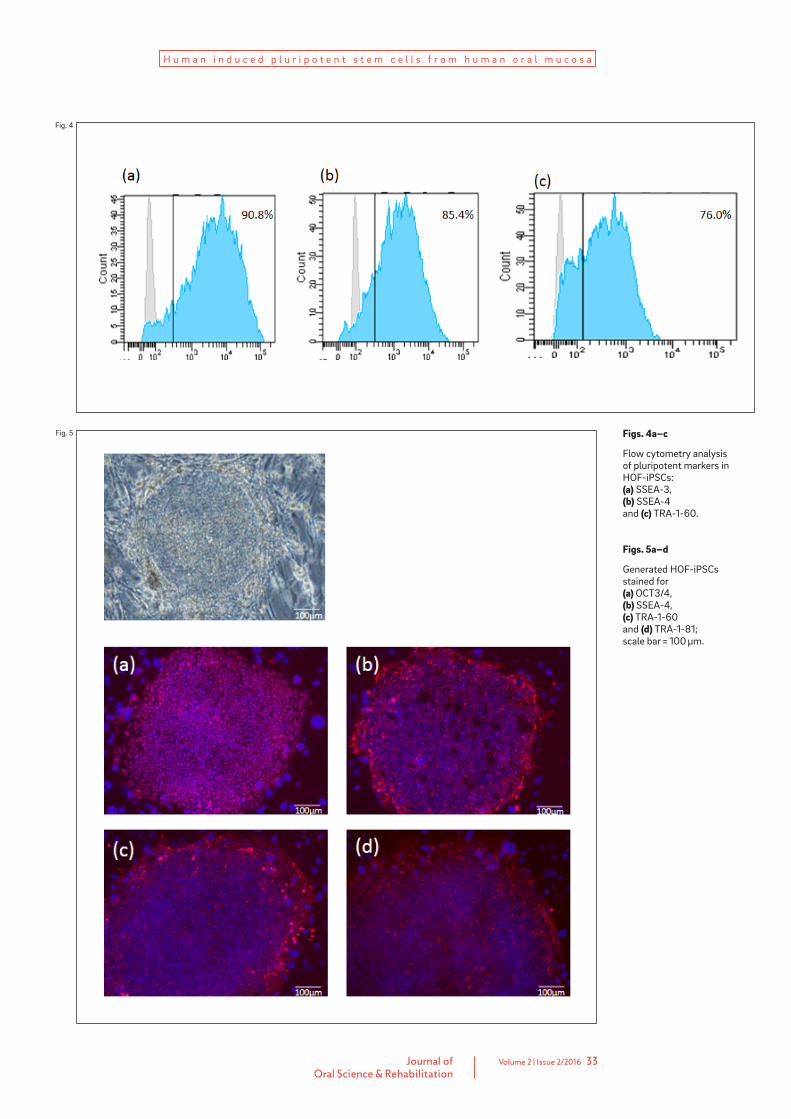
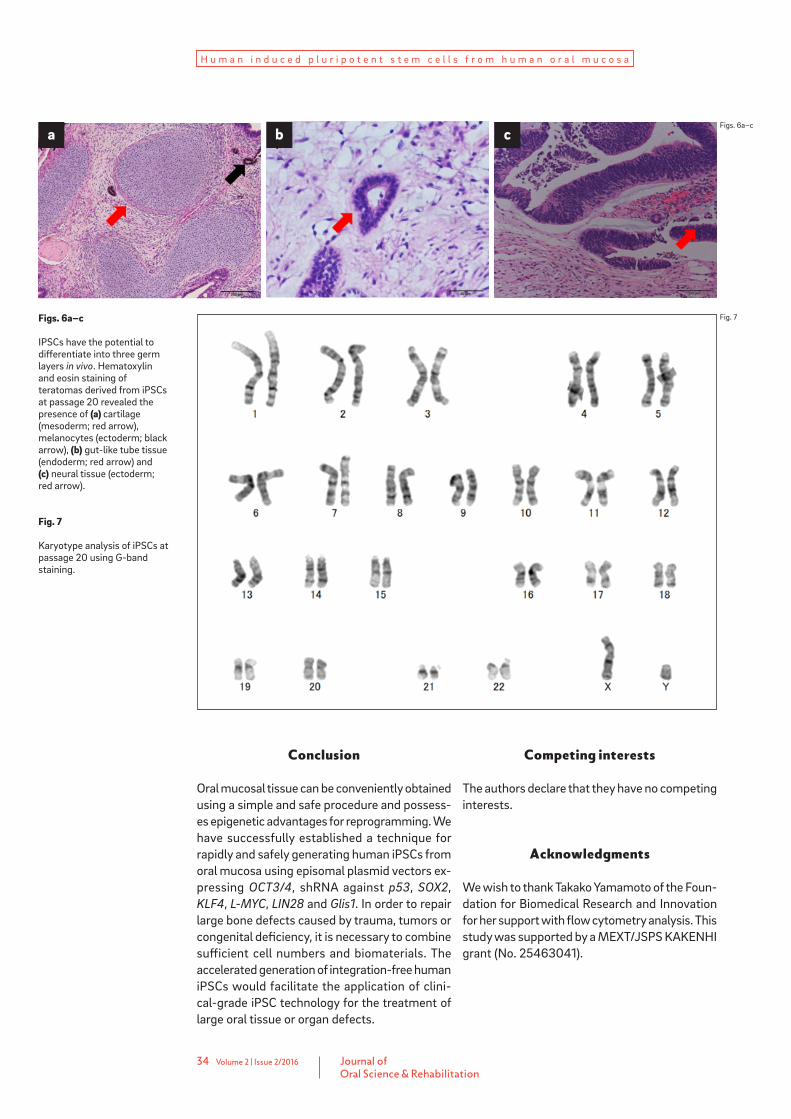
based on increased proliferation and stability of the ESC-like morphology. Expression of the ESC-specific surface markers SSEA-3, SSEA-4 and TRA-1-60 in HOF-iPSCs was analyzed using flow cytometry; all three markers were expressed (Fig. 4). HOF-iPSCs could be maintained beyond 20 passages and still demonstrated ESC-like morphology. In addition, HOF-iPSCs expressed ESC-specific surface markers, such as OCT3/4, SSEA-4, TRA-1-60 and TRA-1-81 (Fig. 5). Tumor formation was observed three months after the injection of HOF-iPSCs under the epidermal space in the neck of immunodeficient mice. His-tological examination showed that the tumor contained various tissues, including cartilage (mesoderm), melanocytes (ectoderm), gut-like tube tissue (endoderm) and neural tissue (ecto-derm; Fig. 6). Karyotype analysis of the tested clones showed a normal human karyotype (Fig. 7).
Discussion
Many strategies have been proposed for the management of large defects in oral tissue or organs such as due to congenital abnormalities, trauma or cancer treatment. Autogenous bone grafts are the gold standard for such reconstruc-tion because of their osteoconductive, osteoin-ductive and nonimmunogenic properties.20, 21 Recently, cell therapy using stem cells combined with osteoconductive biomaterials or scaffolds has become a promising alternative to autoge-nous bone grafts.22 In order for cell therapy to efficiently treat large defects in oral tissue or organs, it is important to produce a sufficient number of cells that function similarly to prima-ry islets. IPSCs, referred to as pluripotent stem cells, have been generated via retrovirus-medi-ated introduction of four transcription factors3,
4 and represent a potentially unlimited source of cells. IPSCs that can be efficiently generated from tissue easily accessible to dentists have great potential;23 iPSCs have been generated from various oral mesenchymal cells23 and these cells have been reported to possess higher reprogramming efficiency than skin fibroblasts do.10
Oral mucosal tissue is easily accessible and can be harvested by a simple and safe procedu-re. Oral mucosal wounds are characterized by rapid re-epithelialization and remodeling and are known to heal quickly compared with other skin injuries. This rapid re-epithelialization and remo-
deling is due to the increased production of ac-tive MMP-2 in oral mucosal fibroblasts compared with skin fibroblasts; MMP-2 may play an import-ant role in rapid extracellular matrix reorganiza-tion and scarless wound healing.13, 24, 25 Therefore, we hypothesized that HOFs generated from pa-tient tissue might provide a superior cell source for iPSCs. In the present study, we found that the endogenous expression level of KLF4 was higher in HOFs than in ESCs or HOF-iPSCs. Endogenous KLF4 has been shown to be expressed in gingival and periodontal fibroblasts derived from oral tissue.12 Miyoshi et al. also found that HOFs ex-press not only KLF4 and C-MYC but also NANOG and OCT4 at low levels, suggesting that HOFs possess a number of epigenetic advantages for reprogramming.13
Integrating virus-associated genotoxicity and tumor formation in iPSCs is of concern for clinical application.15 Integration-free human iPSCs have been generated using several methods.24, 26–30
Okita et al. used two of their findings to improve reprogramming efficiency using episomal plas-mids;15 iPSC generation is markedly enhanced by p53 suppression31 and L-MYC is more potent and specific than C-MYC during human iPSC genera-tion.32 In our previous study,17 iPSCs were gene-rated from HGFs using the above-mentioned method. The generated iPSCs expressed ESC-spe-cific markers, as assessed by gene analysis and immunocytochemistry. Embryoid bodies and teratomas were formed from the iPSCs, demon- strating their ability to differentiate into three germ layers. However, 50 ESC-like colonies were obtained only 30 days post-HGF transfection. This lengthy reprogramming process (up to one month) is comparable to that of the standard virus-mediated methodology.33
The maternal Gli-like transcription factor Glis1 is highly expressed in unfertilized eggs and one-cell-stage embryos.18 Maekawa et al. showed that Glis1, but not C-MYC, increased iPSC tumorige-nicity and markedly enhanced the generation of iPSCs from both mouse and human fibroblasts when expressed together with OCT3/4, SOX2 and KLF4.18 In the present study, we observed 50 co-lonies of human ES-like cells as early as 20 days after initial episomal plasmid vector transduction. These results demonstrate that Glis1 enhances the efficiency of iPSC generation using episomal plasmid vectors expressing OCT3/4, shRNA against p53, SOX2, KLF4, L-MYC and LIN28. However, iPSC generation from multiple donors will be required to establish the application of iPSC technology to biomedical research.
Journal ofOral Science & Rehabilitation
Volume 2 | Issue 2/2016 33
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
Figs. 4a–c
Flow cytometry analysis of pluripotent markers in HOF-iPSCs: (a) SSEA-3, (b) SSEA-4 and (c) TRA-1-60.
Figs. 5a–d
Generated HOF-iPSCs stained for (a) OCT3/4, (b) SSEA-4, (c) TRA-1-60 and (d) TRA-1-81; scale bar = 100 μm.
Fig. 4
Fig. 5
Journal ofOral Science & Rehabilitation
34 Volume 2 | Issue 2/2016
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
a b c
Conclusion
Oral mucosal tissue can be conveniently obtained using a simple and safe procedure and possess-es epigenetic advantages for reprogramming. We have successfully established a technique for rapidly and safely generating human iPSCs from oral mucosa using episomal plasmid vectors ex-pressing OCT3/4, shRNA against p53, SOX2, KLF4, L-MYC, LIN28 and Glis1. In order to repair large bone defects caused by trauma, tumors or congenital deficiency, it is necessary to combine sufficient cell numbers and biomaterials. The accelerated generation of integration-free human iPSCs would facilitate the application of clini-cal-grade iPSC technology for the treatment of large oral tissue or organ defects.
Competing interests
The authors declare that they have no competing interests.
Acknowledgments
We wish to thank Takako Yamamoto of the Foun-dation for Biomedical Research and Innovation for her support with flow cytometry analysis. This study was supported by a MEXT/JSPS KAKENHI grant (No. 25463041).
Figs. 6a–c
IPSCs have the potential to differentiate into three germ layers in vivo. Hematoxylin and eosin staining of teratomas derived from iPSCs at passage 20 revealed the presence of (a) cartilage (mesoderm; red arrow), melanocytes (ectoderm; black arrow), (b) gut-like tube tissue (endoderm; red arrow) and (c) neural tissue (ectoderm; red arrow).
Fig. 7
Karyotype analysis of iPSCs at passage 20 using G-band staining.
Figs. 6a–c
Fig. 7

Journal ofOral Science & Rehabilitation
Volume 2 | Issue 2/2016 35
H u m a n i n d u c e d p l u r i p o t e n t s t e m c e l l s f r o m h u m a n o r a l m u c o s a
1. Lowry WE, Richter L, Yachechko R, Pyle AD, Tchieu J, Sridharan R, Clark AT, Plath K. Generation of human induced pluripotent stem cells from dermal fibroblasts. → Proc Natl Acad Sci U S A. 2008 Feb;105(8):2883–8.
2. Park IH, Zhao R, West JA, Yabuuchi A, Huo H, Ince TA, Lerou PH, Lensch MW, Daley GQ. Reprogramming of human somatic cells to pluripotency with defined factors. → Nature. 2008 Jan;451(7175):141–6.
3. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. → Cell. 2007 Nov;131(5):861–72.
4. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. → Cell. 2006 Aug;126(4):663–76.
5. Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, Slukvin II, Thomson JA. Induced pluripotent stem cell lines derived from human somatic cells. → Science. 2007 Dec;318(5858):1917–20.
6. Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. → Nature. 1981 Jul;292(5819):154–6.
7. Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. → Proc Natl Acad Sci U S A. 1981 Dec;78(12):7634–8.
8. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. → Science. 1998 Nov;282(5391):1145–7.
9. Su RJ, Neises A, Zhang XB. Generation of iPS cells from human peripheral blood mononuclear cells using episomal vectors. In: Turksen K, Nagy A, editors. Induced pluripotent stem (iPS) cells: methods and protocols. → New York: Springer; 2016. p. 57–69. (Walker JM, editor. Methods in molecular biology; vol. 1357).
10. Tamaoki N, Takahashi K, Tanaka T, Ichisaka T, Aoki H, Takeda-Kawaguchi T, Iida K, Kunisada T, Shibata T, Yamanaka S, Tezuka K. Dental pulp cells for induced pluripotent stem cell banking. → J Dent Res. 2010 Aug;89(8):773–8.
11. Egusa H, Okita K, Kayashima H, Yu G, Fukuyasu S, Saeki M, Matsumoto T, Yamanaka S, Yatani H. Gingival fibroblasts as a promising source of induced pluripotent stem cells. → PLoS One. 2010 Sep;5(9):e12743.
12.Wada N, Wang B, Lin NH, Laslett AL, Gronthos S, Bartold PM. Induced pluripotent stem cell lines derived from human gingival fibroblasts and periodontal ligament fibroblasts. → J Periodontal Res. 2011 Aug;46(4):438–47.
13. Miyoshi K, Tsuji D, Kudoh K, Satomura K, Muto T, Itoh K, Noma T. Generation of human induced pluripotent stem cells from oral mucosa. → J Biosci Bioeng. 2010 Sep;110(3):345–50.
14. Oda Y, Yoshimura Y, Ohnishi H, Tadokoro M, Katsube Y, Sasao M, Kubo Y, Hattori K, Saito S, Horimoto K, Yuba S, Ohgushi H. Induction of pluripotent stem cells from human third molar mesenchymal stromal cells. → J Biol Chem. 2010 Sep;285(38):29270–8.
15. Okita K, Matsumura Y, Sato Y, Okada A, Morizane A, Okamoto S, Hong H, Nakagawa M, Tanabe K, Tezuka K, Shibata T, Kunisada T, Takahashi M, Takahashi J, Saji H, Yamanaka S. A more efficient method to generate integration-free human iPS cells. → Nat Methods. 2011 May;8(5):409–12.
16. Su RJ, Baylink DJ, Neises A, Kiroyan JB, Meng X, Payne KJ, Tschudy-Seney B, Duan Y, Appleby N, Kearns-Jonker M, Gridley DS, Wang J, Lau KH, Zhang XB. Efficient generation of integration-free iPS cells from human adult peripheral blood using BCL-XL together with Yamanaka factors. → PLoS One. 2013 May;8(5):e64496.
17. Umezaki Y, Hashimoto Y, Nishishita N, Kawamata S, Baba S. Human gingival integration-free iPSCs; a source for MSC-like cells. → Int J Mol Sci. 2015 Jun;16(6):13633–48.
18. Maekawa M, Yamaguchi K, Nakamura T, Shibukawa R, Kodanaka I, Ichisaka T, Kawamura Y, Mochizuki H, Goshima N, Yamanaka S. Direct reprogramming of somatic cells is promoted by maternal transcription factor Glis1. → Nature. 2011 Jun;474(7350):225–9.
19. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. → Methods. 2001 Dec;25(4):402–8.
20. Gimbel M, Ashley RK, Sisodia M, Gabbay JS, Wasson KL, Heller J, Wilson L, Kawamoto HK, Bradley JP. Repair of alveolar cleft defects: reduced morbidity with bone marrow stem cells in a resorbable matrix. → J Craniofac Surg. 2007 Jul;18(4):895–901.
21. Sakamoto F, Hashimoto Y, Kishimoto N, Honda Y, Matsumoto N. The utility of human dedifferentiated fat cells in bone tissue engineering in vitro. → Cytotechnology. 2015 Jan;67(1):75–84.
22. Levi B, Glotzbach JP, Wong VW, Nelson ER, Hyun J, Wan DC, Gurtner GC, Longaker MT. Stem cells: update and impact on craniofacial surgery. → J Craniofac Surg. 2012 Jan;23(1):319–22.
23. Egusa H, Sonoyama W, Nishimura M, Atsuta I, Akiyama K. Stem cells in dentistry—part I: stem cell sources. → J Prosthodont Res. 2012 Jul;56(3):151–65.
24. Zhou W, Freed CR. Adenoviral gene delivery can reprogram human fibroblasts to induced pluripotent stem cells. → Stem Cells. 2009 Nov;27(11):2667–74.
25.Stephens P, Davies KJ, Occleston N, Pleass RD, Kon C, Daniels J, Khaw PT, Thomas DW. Skin and oral fibroblasts exhibit phenotypic differences in extracellular matrix reorganization and matrix metalloproteinase activity. → Br J Dermatol. 2001 Feb;144(2):229–37.
26. Woltjen K, Michael IP, Mohseni P, Desai R, Mileikovsky M, Hämäläinen R, Cowling R, Wang W, Liu P, Gertsenstein M, Kaji K, Sung HK, Nagy A. piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells. → Nature. 2009 Apr;458(7239):766–70.
27. Jia F, Wilson KD, Sun N, Gupta DM, Huang M, Li Z, Panetta NJ, Chen ZY, Robbins RC, Kay MA, Longaker MT, Wu JC. A nonviral minicircle vector for deriving human iPS cells. → Nat Methods. 2010 Mar;7(3):197–9.
28. Warren L, Manos PD, Ahfeldt T, Loh YH, Li H, Lau F, Ebina W, Mandal PK, Smith ZD, Meissner A, Daley GQ, Brack AS, Collins JJ, Cowan C, Schlaeger TM, Rossi DJ. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. → Cell Stem Cell. 2010 Nov;7(5):618–30.
29. Kim D, Kim CH, Moon JI, Chung YG, Chang MY, Han BS, Ko S, Yang E, Cha KY, Lanza R, Kim KS. Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. → Cell Stem Cell.2009 Jun;4(6):472–6.
30. Fusaki N, Ban H, Nishiyama A, Saeki K, Hasegawa M. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. → Proc Jpn Acad Ser B Phys Biol Sci. 2009;85(8):348–62.
31. Hong H, Takahashi K, Ichisaka T, Aoi T, Kanagawa O, Nakagawa M, Okita K, Yamanaka S. Suppression of induced pluripotent stem cell generation by the p53-p21 pathway. → Nature. 2009 Aug;460(7259):1132–5.
32. Nakagawa M, Takizawa N, Narita M, Ichisaka T, Yamanaka S. Promotion of direct reprogramming by transformati-on-deficient Myc. → Proc Natl Acad Sci U S A. 2010 Aug;107(32):14152–7.
33. Shimada H, Hashimoto Y, Nakada A, Shigeno K, Nakamura T. Accelerated generation of human induced pluripotent stem cells with retroviral transduction and chemical inhibitors under physiological hypoxia. → Biochem Biophys Res Commun.2012 Jan;417(2):659–64.
References





























