Embed Size (px)
Citation preview

NO
RW
EGIA
N U
NIV
ERSITY O
F LIFE SCIEN
CES
DEPA
RTMEN
T OF IN
TERN
ATION
AL ENVIR
ON
MEN
T AND
DEVELO
PMEN
T STUD
IESM
ASTER
THESIS 30 C
RED
ITS 2007
ACARICIDE RESISTANCE IN NORWEGIAN POPULATIONSOF THE TWO-SPOTTED SPIDER MITE (Tetranychus urticae Koch) (Acari: Tetranychidae)
CASSIUS GARWO VARNEY FAHNBULLEH

i
D E C L A R A T I O N
This thesis has not previously been accepted for any degree and is not been currently
submitted in candidature of any other degree. This work is the result of my own
investigation. All other sources of information are acknowledged and a reference list
appended.
Cassius G. V. Fahnbulleh
Sgd:…………………….
Date:…………………….

ii
D E D I C A T I O N
This piece of work is dedicated to my parents (Mr. William Siaka Fahnbulleh and Ms.
Maima Gbeni Fahnbulleh), (though deceased) for their care over me. Parents, because of
your farsightedness and through God – I am whom I am. It is also dedicated to my two
children (Musukula D. Fahnbulleh and William S. S. Fahnbulleh) who toiled alongside
me and endured hardship throughout those dreadful years of exile in Sierra Leone. Their
patience, understanding and continued love are highly valued – may God illuminate the
paths on which they tread.

iii
A C K N O W L E D G E M E N T S
In my quest approximately a year ago for a suitable topic to write a thesis, I was fortunate
to have had one from Bioforsk in the area of entomology – though challenging. But
during the course of the work, I had access to many valuable, enthusiastic and inspiring
literatures and discussions on the topic from several quarters at Bioforsk lab that
broadened my horizon; for this I am grateful.
The lab work was interesting, at some point in time apprehensive and stressful but mostly
cheerful and absorbing because of the new knowledge. Several people contributed to this
work, knowingly or unknowingly in diverse ways. They are too many to allude to by
name but to mention a few: I wish to extend my sincere gratitude to my immediate
supervisor, PhD. Einar G. M. Nordhus, for his excellent guidance and immense and
valuable contributions in both the laboratory work when I have felt confused in the use of
strange gadgets, and the writing processes when the construction of my thoughts in words
became an impediment. I am particularly indebted to my main supervisor, Prof. Arild
Andersen, who readily consented to work along with PhD. Nordhus to ensure that the
work is carried out at the Bioforsk lab – to you I am thankful. Not forgetting Dr. May-
Guri Sæthre, who besides suggesting the topic for this work, made it possible for me to
meet Dr. Nina S. Johansen through whom I became acquainted with my supervisors, my
gratitude. I must equally acknowledge the willingness of Toril Sagen of the aforesaid lab
who was always ready to assist in anyway possible for this work to be successful. In
addition, a big thank you goes to all other staff members attached to Bioforsk for their

iv
helpfulness and good nature towards me during my association with them, this will
always linger in my memory.
Last, but of course not the least, my study at UMB could not have been possible without
the humanitarian gesture of the people and government of Norway, who through the UN
resettlement programme, wholeheartedly granted me and my family asylum to recreate
our devastated lives after languishing in refugee camps and slums in Sierra Leone for 13
successive years where we nearly met our demise. For this, I am sincerely grateful.
Above all, I thank Almighty God for preserving my life.
Ås, 2007
Cassius G. V. Fahnbulleh

v
C E R T I F I C A T I O N
This is to certify that this thesis work was carried out by Cassius G. V. Fahnbulleh in the
Bioforsk laboratory, The Norwegian Institute for Agricultural and Environmental
Science, Norwegian University of Life Sciences (UMB), Ås.
Sgd: ……………………….. Approved: ………………....
PhD. Einar Nordhus Prof. Arild Andersen
Thesis Supervisor Main Supervisor
Date:………………. Date:………………..

vi
ABSTRACT
Farmers and greenhouse crop growers are faced with numerous polyphagus and
phytophagus pest species, among which is the two-spotted spider mite, Tetranychus
urticae Koch (Acari: Tetranychidae). This species has become a deleterious economic
pest in many cultivated crops, and its control has posed serious threats to farmers because
of its rapid growth rate and speedy adaptation to numerous acaricides. This study is
therefore aimed at identifying and characterizing the resistance in two Norwegian
populations of the two-spotted spider mite, T. urticae compared to a susceptible strain
GSS.
Standard bioassay procedures were used to determine the resistance in T. urticae by spray
application of different rates of acaricides against the different strains of spider mite
larvae. Mortality was scored after 72 hrs. through data collection, analyzed and results
expressed in Abbott’s corrected percent mortality and the LC50 values and slopes were
estimated by probit analysis, and resistance factor (RF) calculated.
The resistance factor (RF) for NOR4 at LC50 for abamectin was twice more than the
susceptible strain (GSS). NOR4 showed low levels of resistance to hexythiazox,
clofentezin and fenazaquin respectively compared with GSS; it also showed no level of
resistance to spirodiclofen and fenpyroximate. Unlike NOR4, NOR5 showed moderate
level of resistance to abamectin but with low level of resistance to clofentezine and
hexythiazox compared to the GSS strain. But it showed no resistance to spirodiclofen
fenpyroximate and fenazaquin. This implies that they exhibited certain level of
susceptibility to these drugs.

vii
TABLE OF CONTENTS
PAGE
Declaration i
Dedication ii
Acknowledgements iii-iv
Certification v
Abstract vi
Table of Contents vii-viii
Abbreviations ix-x
1. INTRODUCTION 1
1.1 General Introduction to Two-spotted Spider Mite, Tetranychus urticae (Koch) 1-3
1.2 Resistance 3-6
1.2.1 Resistance Mechanisms 7
1.2.1.1 Metabolic Mechanisms / Detoxification Mechanisms 7-10
1.2.1.2 Target Site Insensitivity / Resistance 10-11
1.2.2 Resistance in Norway 12
2. MATERIALS AND MATHODS 12
2.1 Strains 12
2.2 Miticides 12-13
2.3 Bioassay Procedure 13-14
2.4 Statistical Calculations 14
3. RESULTS 15-26
4. DISCUSSIONS 27-29

viii
5. CONCLUSIONS 29-30
6. REFERENCES 31--44
7. APPENDIX 45-48

x
ABBREVIATIONS
AChE Acetylcholine esterase
CO Company o C Degree Celsius
Cons Concentrations
DDT dichlorodiphenyltrichloroethane
DF
EC Emulsion concentrate
GABA γ - Aminobutyric acid
GSS German susceptible strain
GST Glutathione S-transferase
g / ldH20 gram / litre distilled water
hrs hours
IPM Integrated Pest Management
IRAC Insecticide Resistance Action Committee
kdr knockdown resistance
LC50 Median lethal concentration of a pesticide expected to kill 50% of a test
organism
LC90 Lethal concentration of a pesticide expected to kill 90% of a test organism
L: D Light to darkness
LTD Limited
METI Mitochondrial electron transport inhibitor
MFO Cytochrome P450 –dependent monooxygenase (Mixed function oxidase)
NADH dehydrogenase
NIAER Norwegian Institute for Agricultural and Environmental Research
NOR4 Norwegian strain no. 4
NOR5 Norwegian strain no. 5
OP Organophosphate
RAC Resistance Action Committee
RF Resistance factor

xi
RR Recommended rate
RH Relative humidity
SC Suspension concentrate
UK United Kingdom
WHO World Health Organization

1. INTRODUCTION
1.1 Two-spotted Spider Mite, Tetranychus urticae (Koch)
The two- spotted spider mite or glasshouse red spider mite, Tetranychus urticae Koch, is
widely known as a serious pest of economic importance in agriculture. The name ‘spider
mite’ emanates from the silk webbing made by mites and not because they appear like
small spider (Zhang, 2003). They are not the only web-spinning pests to feed on plants,
but they are the most common. T. urticae is a phytophagus mite and causes significant
yield losses in many horticultural, ornamental and agronomic crops globally (Jeppson et
al., 1975; Van de Vrie et al., 1972).
Figure 1. A) A rose plant and B) Two spotted spider mite on a rose plant
The two-spotted spider mite belongs to the class Arachnida (arachnids) which contains
four pairs of legs in contrast to that of insects. It has five life stages i.e. egg, larva,
protonymph, and deutonymph (or nymphs), and adult. At emergence from the egg, the
larva is colourless and has three pairs of legs, it also has red eyes. After eating, the colour
may change to light green, brownish yellow or dark green. At this developmental stage,
two dark spots appear in the middle of the body. Upon eating enough food, the larva goes
through a rest period until it develops into protonymph; it is at this time that the four pair
of legs appears. At this stage, the legs are little bigger, and the colour of the mite differs
from light to deep green. The two spots enlarge and appear clearer than on the larva.
After further feeding, the protonymph also goes into dormancy and then develops into a
A. B.

2
deutonymph which appears bigger but with similar coloration to the protonymph. At this
stage, differences can be seen between the male and the female. The female is slightly
bigger and rounder in shape than the male which is oblong in shape. It is from the
deutonymph that the adult mite develops once the nymph has had both a feeding and a
dormancy period.
Due to its rapid proliferation and very short life cycle coupled with favourable climatic
conditions, many generation of T. urticae can be completed in a growing season
(Crooker, 1985; Helle & Sabelis, 1985a). Females can, under favorable conditions,
oviposit approximately 30-50 eggs in 24 hours on the surface of plant structures, often
hidden in fissures. The eggs take 1-2 days to hatch with an approximate temperature of
22 (±1) 0C (Zhang, 2003). Details included in Table 1. (Herbert 1981) demonstrate the
growth periods at 210 C for individual larval and nymphal stages of both sexes. This
shows that female takes longer to develop than males. Mature males will remain near the
female nymph until the mature female emerges, and copulation takes place almost
immediately after emergence.
Table 1.Development time in days for Tetranychus urticae Koch at 210 C (Herbert, 1981)
The development rates of mites are dependent upon temperature, species, host plant,
humidity and other factors. From egg to adult may vary from 5-10 days based on the
Active Quiescent Total
Larva Male 1.5 1.3 2.8 Female 1.5 1.2 2.7 Protonymph
Male 1.0 1.3 2.3
Female 1.3 1.2 2.4
Deutonymph
Male 1.0 1.4 2.5
Female 1.5 1.4 2.9

3
aforesaid condition(s). Carey & Bradley (1982) discovered the growth stages in T. urticae
from egg to adult to take only 6.2 days at 29.40 C; while the optimum temperatures for
development is found to be 30-320 C (Crooker, 1985; Zhang, 2003). This implies that the
two-spotted spider mite can complete a generation as little as 5 days under the most
favorable conditions. Mites particularly tend to increase during rainless periods. The two-
spotted spider mite multiplies rapidly during period of low humidity and any conditions
when growers reduce misting or spraying indoor (Zhang, 2003), and during periods with
low precipitation outdoors.
Both immature and mature T. urticae suck up leaf fluid and principal photosynthetic
pigment (chlorophyll) necessary for photosynthesis. This feeding habit, if severe, may
cause browning of the leaves, reduction in photosynthesis, and eventually leaf death.
Fruit set can be reduced the following year, and fruit sizes may become smaller (Polk,
1994; Barkette, 1994). In some crops, a small amount of injury can lead to high economic
loss e.g. in cut flowers (Croft & Van de Bann, 1988). The main problem with the species
in Norway is in flower production in greenhouses and strawberry production under field
condition.
1.2 Resistance
The WHO (World Health Organization) defined resistance as the inherited ability of a
strain of some organisms to survive doses of toxicant which would prove harmful to a
greater number of the individuals in a normal community of the same strain (Brown &
Pal, 1971). Yet another definition propounded by Sawicki (1987) states that: Resistance
represents a genetic modification in reaction to assortment by poisons that may hinder
control in the field. Generally, pests are known to develop resistance to chemical(s) due
to uncontrolled use and weak effect from common pesticides which are not actually
potent against them; and this resistance, in part, emanates from their genetic makeup
(Zhang, 2003).

4
The development of pesticide resistance takes a multi facet dimensions. For this reason,
worldwide cases of resistance combined with scientific and public pressures led the
pesticide industry to form various ‘resistance action committees - RACs’ including one
for insecticide (i.e. Insecticide Resistance Action Committee - IRAC). This group, not
been in total agreement with the above definition of resistance, came out with criteria to
be considered as additional reasons for resistance development (Tomlin, 1997). They
include the following: that the chemical product has a record of not been potent against
the particular pest in question; that the product lack of success is not due to poor storage,
adulteration, or application, nor caused by abnormal climatic or environmental
conditions; that the recommended dosage failure to suppress pest populations below the
level of economic threshold, and that failure to control is due to heritable change in the
susceptibility of pest population of the product. Based on the above criteria, IRAC
pointed out that the term ‘resistance’ should be used only when field failure occurs and
this situation is confirmed (Mota-Sanchez et al., 2002). Resistance can also be considered
as a form of self-defense, because pest subjection to the many environmental stresses or
dangers such as: humidity, temperature, radiation, predation/parasitism, diseases, and
pollutants in the form of pesticides or plants allelochemicals inhibit their activities for
survival. It is the apparent expression of pests’ natural response to the above stresses
through resistance mechanisms that is regarded as self-defense (Koehn & Bayne, 1989;
Scott, 1995).
Resistance can be classified as:
a) Cross-resistance i.e. the resistance to two or more types of pesticides because they
have identical or very similar mode of action. Ex. Organophosphate (OP) and carbamate
pesticides intoxicate by similar moods of action, and resistance to one usually results in
resistance to the other;
b) Multi-resistance i.e. resistance to two or more types of pesticides because of the
coexistence of two or more different resistance mechanisms. For instance, a resistant
insect may have both metabolic resistance to OPs and target-site resistance to pyrethroids
(Scott, 1995).

5
Other form of resistance development in pests include: behavioral resistance (i.e. to avoid
toxic dose of the chemical that would otherwise prove deadly) (Roush & Mckenzie,
1991).
It has been found that T. urticae has the potential to speedily develop resistance against a
large number of acaricides (Cranham & Helle, 1985; Knowles, 1997; Stumpf & Nauen,
2001). Furthermore, their high prolific rate, their rapid growth period, their polyphagus
feeding habit, coupled with their extremely dispersal behaviour (i.e. omnipresent-ness) all
help to boost resistance development in the species (Croft & Van de Bann, 1988). The
persistent exposure of T. urticae to diverse pesticides in order to contain them below
economic threshold has resulted in resistant populations found in more than 40 countries
in both greenhouses and field conditions (Georghiou & Lagunes-Tejeda, 1991), and
resistance to at least 85 different compounds has been published
(http://www.pesticideresistance.org).
There are many different classes of miticides commercially available. Some of the most
used in Norway are:
(a) METI (mitochondrial electron transport inhibitors) acaricides that can prevent and
allow mitochondrial respiration (Hollingworth & Ahammadsahib, 1995; Wood et al.,
1996). They are known to be effective against all development phases of tetranychid,
tarsonid and eriophyd mites (Tomlin, 2003). They function by hampering complex I
(NADH: ubiquinone oxidoreductase) of the mitochondrial respiratory chain (Tomlin,
2003; Hollingworth & Ahammadsahib, 1995). Resistance was first reported in
Tetranychus kanzawa (Kishida) which was collected from tea fields in Japan in 1994
(Ozawa, 1994). Resistance of T. urticae to METI acaricides has been found in England
(Devine et al., 2001; Stumpf & Nauen, 2001; Sato et al., 2004), Australia (Herron &
Rophail, 1998) and Belgium (Bylemans & Meurrens, 1997). This resistance is said to be
inborn as an incomplete dominant trait and has been associated with over-production of
detoxification enzymes like esterases (Devine et al., 2001; Stumpf & Nauen, 2001; Sato
et al., 2004).

6
(b) Mite growth inhibitors are another group of insecticides. They have little or no effect
on mature females, but can cause them to lay fewer viable eggs (Chapman & Marris,
1986). The effect of these acaricides is mostly on juveniles (nymphs), by preventing
moulting. Resistance to mite growth inhibitors has been detected in the European red
mite and the two-spotted spider mite from different countries (Nauen et al., 2001). For
example, hexythiazox is thought to disrupt chitin fusion (Flexner et al., 1995; Dekeyser &
Downer, 1994); it has been reported to be selective, and it showed no effect on adult
carnivorous mites making it well suited in integrated pest management (IPM) (Hoy &
Ouyang, 1989). Another mite growth inhibitor, clofentezine, resistance was detected for
the first time in T. uraicae in 1987 on Australian roses (Edge et al., 1987), and in 1998 on
apple (Herron & Rophail, 1998). Cross resistance between the ovo-lavicides is well
known (Stumpf & Nauen, 2001). Such cross-resistance between hexythiazox and
clofentezine is likely not the result of metabolic traits, but an alteration in the target site is
proposed to be the possible cause resulting in the high levels of resistance observed in
these populations. However, the study of target site resistance as regards to hexythiazox
and clofentezine is presently not possible because the site of action is not known (Stumpf,
2001).
(c) Another chemical group, chloride channel activators, has also been used a lot against
mites. These chemicals are antagonists of the γ -Aminobutyric acid (GABA) receptor and
increase transfer of chloride-ions which will lead to mite paralysis and death (Fritz et al.,
1979; Matsumura et al., 1987). Resistance in T. urticae to abamectin is known from
California, Florida, Netherlands (Campos et al., 1996), and Washington (Beers et al.,
1998).
(d) Spirodiclofen is a recently developed acaricide with a good potency against mites.
The compound is particularly active against the developmental stages of the mite and not
so much against the adults (Nauen et al., 2000). The mood of action is not known and no
report of resistance has been published.

7
1.2.1 Resistance Mechanisms
Although there are many different chemical pesticides and different species of insects and
mite pests, the number of processes known to cause resistance to widely-used chemical
pesticides is scarce (Scott, 1995). In light of the above, two known resistance processes
are discussed. They are metabolic resistance (detoxification mechanisms) and target site
insensitivity /resistance.
1.2.1.1 Metabolic Resistance / Detoxification Mechanisms
Metabolic resistance is the potential of pests to expel poisonous pesticides from their
body through chemically driven deterioration. Three enzyme families have been found to
be involved in this resistance mechanism: Nonspecific esterases, cytochrome P450 –
dependent monooxygenases and glutathione- S –transferases (GSTs) (Brogdon &
McAllister, 1998).
Cytochrome P450 – dependent monooxygenases
The cytochrome P450 - dependent monooxygenases belong to a vast super family of
water-repelling, hemecontaining enzymes involved in the detoxification of many
identical internally originated elements such as juvenile hormones, ecdysteroids and
pheromones, and externally originated substances such as plant allelochemicals,
insecticides and promutagenes (Eldefrawi et al., 1960; Nordhus, 2005). As for its
association with acaricide resistance, the process of fluvalinate resistance in the Varroa
mite is a good example (Hillesheim et al., 1996). Cytochrome P450 oxidizes biochemical
change in insecticides through O-, S-, and N- alkyl hydroxylation, aliphatic hydroxylation
and pixilation, aromatic hydroxylation, ester oxidation and nitrogen and thioester
oxidation (Wilkinson, 1976; Maitra et al., 1996; Brogdon & McAllister, 1998).
The first account on P450 monooxygenase resistance became apparent in the 1960’s when
it was revealed that the resistance was eliminated by the P450 suppressor desamex
(Nordhus, 2005). From that period onward, P450 monooxygenase mediated resistance has

8
been known to be in a variety of different insects, and can be seen as the most repeated
type of metabolic resistance in entomology (Scott, 1991; Nordhus, 2005). Both increased-
adjustment and amino acid replacements have been associated with increased metabolism
of insecticides by different P450. For example, a monooxygenase CYP6D1 in housefly, M.
domestica, has been associated with pyrethroid resistance through increased production
(Kasai & Scott, 2000; Nordhus, 2005). Furthermore, resistance against DDT has been
associated with amino acid replacement in P450 CYP6G1 in Drosophila strains (Berge et
al., 1998; Nordhus, 2005). Due to the high number of P450 monooxygenases in insects and
the frequent-overlapping substrate restricted to an individual species of this enzymes
group, it has been difficult to discover a good biochemical structure and molecular way
of showing the monooxygenase-mediated resistance. Some in vivo inhibition methods
have been made to study the involvement of P450 in resistance. For example, synergists
like piperonyl butoxide are used to suppress P450 activity in bioassays to elucidate
resistance involvement (Nordhus, 2005). The over-expression of P450 monooxygenases
has also been associated with resistance to a variety of categories of insecticides inclusive
of neonicotinoids, OP and growth regulators (Daborn et al., 2001; Le Goff et al., 2003;
Nordhus, 2005). Cytochrome P450-dependent monooxygenases, which is also considered
as mixed function oxidases (MFO), are one of the most vital metabolic systems (phase I)
in insects and supposedly in mites as well. The MFO-based mictrotiter plate assay using
P-nitroanisole as a substance on which an enzyme acts in biochemical reactions
developed by Rose et al. (1995), was altered for exact characterization of cytochrome
P450 - dependent monooxygenase action in spider mite microsomes. But no specific
resistance mechanisms have yet been found to be associated with it.
Glutathione- S - transferases (GSTs)
Glutathione –S – transferase is another enzymes class that has been found to be involved
in metabolic resistance (Scott, 1995). It is a diverse class of enzymes that take major part
in the detoxification of both internal and foreign substances including insecticides
(Salinas & Wong, 1999; Nordhus, 2005). They speed up the degradation of pesticides or
their metabolism (Stumpf, 2001) so as to increase water solubility and later the

9
elimination of lipophilic substances (Motoyama & Deuterman, 1975; Stumpf, 2001).
They can convert the poisonous substances of pyrethroid, organophosphate (OP), DDT,
cyclodiene and carbamate insecticides into harmless substances. An increased GST
production has been associated with resistance to all major classes of insecticides
(Prapanthadara et al., 1995; Vontas et al., 2001; Hemingway et al., 2004; Nordhus, 2005),
but the process involved in this elevated enzyme production is not well known (Anayati
et al., 2005; Nordhus, 2005). Enzymatic detoxification has the potential to confer cross-
resistance to toxins independent of their target site (Stumpf, 2001). Despite the fact that
much is known about insect GSTs and their role in the biochemical changes (metabolism)
of insecticides, little information is available concerning the enzyme in spider mite
(Stumpf, 2001) therefore, resistance mechanisms linked to GST in mites are difficult to
elucidate.
Nonspecific Esterases
Esterases are a large group of enzymes which break down a large number of different
substrates. All esterase enzymes are able to split ester bonds in the presence of water, and
because most insecticides, particularly organophosphates (OPs) and carbamates have
ester bonds, they are prone to esterase degradation. The resistance mechanisms in most
cases are a result of increased levels of esterases production (Fournier et al., 1987; Field
et al., 1988; Carlini et al., 1991; Kettermen et al., 1992; Chen & Sun, 1994; Scott, 1995).
This increased esterase production process is known in mosquitoes, houseflies, mites and
whiteflies respectively (Ahmed & Wilkins, 2002; Hemingway & Karunaratne, 1998;
Nordhus, 2005). In Myzus persicae Sulz and Culex mosquitoes, the esterases genes which
give rise to resistance are highly amplified and about 250 copies of the similar gene may
be seen in a single individual (Mouchès et al., 1986; Poiriè et al., 1992). The more the
esterase genes are ampilifed, the higher the level of resistance provided (Field et al.,
1988; Poiriè et al., 1992). Two point mutations in the region of binding and chemical
reaction of substrate of the E3 malathion carboxyl esterase in a strain of sheep blow fly,
Lucilia cuprina, cause resistance to a range of OPs (Campbell et al., 1998; Nordhus,
2005). Almost identical mutations have been noticed in several malathion resistant

10
Anopheles species (Herath et al., 1987; Hemingway, 1982; Nordhus, 2005) and in a
housefly strain (Claudianose et al., 1999; Nordhus, 2005). A considerable amount of
evidence shows that esterases are connected with acaricide resistance in many spider mite
strains. For instance, Kim and Lee (1990) obtained resistant strains of T. urticae by
sequential assortment with the acaricides: carbophenothion, ethion, dicofol, cyxehation
and bifenthrin, and separated easterase isoenzyme by polyacrylamide gel electrophoresis.
By doing so, they were able to detect different isoenzymes in the resistance and
susceptible strains. Weyda et al. (1984) suggested that there were different esterase
arrangements and particular size numbers in polyacrylamide gel electrophoresis between
strains resistance and susceptible to thiometon. The difference indicated that esterases are
associated to the resistance mechanisms of the tested acaricides. Capua et at. (1990) and
Sundukov et al. (1989) supported their finding based on their own studies.
1.2.1.2 Target site insensitivity / resistance
Target site resistance involves the modification of the structures that are the target sites
within the insect or mite which leads to a change in susceptibility, so that the target is less
reactive to pesticide action (Stumpf, 2001). At least three different target site resistance
mechanisms have been found: γ - aminobutyric acid (GABA), Acetylcholine esterase
(AchE) and Sodium channel (knockdown resistance, kdr). All of these are well known
targets of insecticides, and resistance alleles of each have been found (Scott, 1995).
A modification in the main anatomy of acetylcholinesterase (AChE) can minimize the
level of suppression by OPs and carbamates, and causes resistance in insects and other
arthropod species. An unresponsive AChE resulting in OP resistance is found in many
different insect species and has also been discovered in T. urticae strains from Germany
(Voss & Matsumura, 1964; Smissaert et al., 1970), New Zealand (Ballantyne & Harrison,
1967), and in a few other tetranychid pest species, including the carmine spider mite (T.
cinnabarinus) from Israel (Zahavi & Tahori, 1970), the kanzawa spider mite, T. kanzawa,
from Japan (Kuwahara, 1982) and Caloglyphus berlesei (Blank, 1979). Diverse
molecular studies with pests revealed that minimized responsiveness of AChE is

11
attributed to one or more point - mutations in the gene resulting in modification of
structures in the enzyme (Fournier et al., 1992; Stumpf, 2001 ); it has been revealed that
successions with several mutations can bring about higher levels of resistance (Stumpf,
2001). Two point mutations in the voltage - gated sodium channel (kdr) have been found
to increase the resistance to pyrethroids and DDT. The first mutation termed kdr produces
a 10 to 20 fold resistance, while in combination with the second mutation (super-kdr) the
resistance level can increase more than 500-fold. A point mutation in the γ - aminobutyric
acid (GABA) receptor can confer an increased resistance to cyclodiene insecticides. This
has been found in a range of different insect species (Bloomquist, 1994). None of the two
last mechanisms have until now been identified in spider mites.
1.2.2 Resistance in Norway
During several years of acricides tests to ascertain resistance status in agricultural pests,
Stenseth (1965) used a two-spotted spider mite strain (K-Strain) from a cucumber
greenhouse in Hardanger to investigate for susceptibility to acaricides. It was found that
the two-spotted spider mite (T. urticae) was resistant to dicofol and parathion. That was
the first reported case of multi-resistant mite in Norway. In another investigation,
Fjelldalen & Stenseth (1962) reported that a two-spotted spider mite strain was collected
on roses, and the chemical parathion was used to control them. But before the selection,
the strain already had a fairly high frequency (20%) of resistant spider mites. This
number increased during the trials and at the end reached an LC50 estimate of 400 times
higher than in a susceptible strain. From that time onward, not much has been done on
this species in Norway.

12
2. MATERIALS AND METHODS
2.1 Strains
The experimental work was undertaken in the laboratory of the Entomology Department
at Bioforsk, NIAER (Norwegian Institute for Agricultural and Environmental Research),
Ås, from the autumn semester of 2006 to the following autumn semester of 2007. Three
laboratory strains of the two-spotted spider mite, T. urticae species (Table 2) were used.
They were cultured on dwarf french bean plants, Phaseolus vulgaris L. The strains were
maintained at 24º C, a photoperiod of 8 - 16 h (L: D), and 60 ± 5 % RH in culture room
and 40-60 ± 5 % RH in growth room respectively. Two of the strains used were
Norwegian (i.e. NOR strains) as tested populations against a German susceptible strain
(GSS) for the investigation.
Table 2.Strains of Tetranychus urticae Tested, origin and host plant
Strain Origin Host Plant
NOR 4 Norway Bean
NOR 5 Norway Bean
GSS Germany Bean
2.2 Miticides
Six miticides were used during the experiment with different mode of actions. They were
grouped as follow:
I. Two METI acaricides:
a. Fenazaquin (Pride Ultra SC), produced by Dow AgroSciences, with recommended
dosage of 200g / L dH2O.
b. Fenpyroximate (Ortus – Akari 5SC), produced by Nihon Noyaku, with recommended
rate of 53g / L dH2O.

13
II. Two mite growth inhibitors:
a. Hexythiazox (Nissuron – Hexygon, DF) produced by Nippon Soda Co. LTD., Japan,
with, recommended rate of 65g /100 L dH2O.
b. Clofentezine (Apollo - Ovation SC), produced by Irvita Plant Protection N. V., with
recommended rate of 500g / L dH2O.
III. Abamectin (Vertimec - Avid 0.15EC), produced by Syngenta Crop Production AG,
Switzerland, with recommended rate of 35g / 100 L dH2O.
IV. Spirodiclofen (Admiral 10EC), produced by bayer CropScience, with recommended
rate of 40g / 100L dH2O.
2.3 Bioassay Procedure
Untreated bean plant (P. vulgaris) leaves were excised from the plant and placed with
the upper side of the leaf on technical solid water agar (1.3 % agar) base in 90 mm
diameter Petri-dish. The leaves-surfaces in the dishes were demarcated with a sticky
insect-glue applied with the aid of a plastic syringe. This was done to prevent mites from
possible escape during the experimental period, and to allow their observation in a
defined area.
With a binocular microscope and a fine sable / pint brush, eight to ten adult females of
the two-spotted spider mite strains, T. urticae, were transferred unto the demarcated areas
of the leaves in each petri-dish. They were covered with a ventilated dish cover and
stored in a growth room for oviposition for 24 hours. If 20 or more eggs are laid, the
female adults are removed. The numbers of eggs were counted, and the petri-dishes
stored in the growth room for hatching. The larvae hatched from the eggs after 4-5 days
were counted and results recorded. They were then immediately treated by spray-
application with acaricide(s). The application of the pesticides was done with four
replicates per concentration of each chemical, and with four concentrations plus distilled
water sprayed as control. The concentration used for each chemical and the process of
data collection and average mortality are found. An example of a chart with the necessary
information can be seen in appendix 1.

14
The six chemicals that were used were diluted into solutions of formulated acaricides
each. Immediately after the completion of the larvae count, petri-dishes containing T.
urticae nymphs of equal ages (i.e. NOR4, NOR5, and GSS) were sprayed with aqueous
solution of miticide using a constant spray rate in a Burkard – computer controlled
sprayer (Burkard Manufacturing LTD., UK.) with equal nozzle sizes firmly attached in a
fume cupboard. See section 2.2 above for the different acaricides and their recommended
dosage used in this investigation. Each of the mite populations were treated with four
different dosages of miticides (i.e. 0.25, 0.50, 1.00 and 2.00) of the above mentioned
aqueous solutions. Distilled water was used as control. The above concentrations
produced 0-100 % mortality. After 24 hours, mortality was scored and assessed. In this
investigation, the criteria used to describe mites / nymphs as dead or alive were based on
the following: ( a) if the selected mites have movement problem or cannot move, they
were considered as dead (Welthy et al., 1988); (b) if nymphs possibly walked-off from
demarcated leaf area prior to spraying, they are regarded as dead (Knight et al., 1990) and
(c) if nymphs are still in the dormant stage, they are scored as dead, but if both nymphs
and adults have no problem with locomotion, they are scored as alive. The samples were
again placed in growth room for another 48 hours (i.e. 72 hours total) after which a
second mortality count was conducted. Only the mortality after 48 hours was utilized for
the calculation of LC50 for all of the pesticides.
2.4 Statistical calculations
The percent mortality was calculated by dividing the sum of the number of dead larvae
per concentration in 48hrs by the number of units counted three days (72 hrs.) after the
application of the chemical (i.e. % mortality = ∑ of tot. dead in 48hr / # of units). All
results given in % mortality were corrected by using the “Abbot’s correction analysis”
(Abbott, 1925) (i.e. Abbot corrected = a –b /a * 100). An example is seen in Appendix 2-
a, b and c in the appendix section. The LC50 values and slopes were estimated by the use
of probit analysis, and resistance factors (RF) calculated according to the below formula
RF =LC50 or LC90 of laboratory collected population / LC50 or LC90 of susceptible
population (GSS).

15
3. RESULTS
The virulent effect of the six acaricides used against the two strains of the two-spotted
spider mite, T. urticae, was examined by the spray application method (bioassay). The
average mortality rate observed in 48 hrs for the control never exceeded 6.82%.
Generally, resistance of the two populations (NOR4 and NOR5) was significantly
different in all of the acaricide treatments compared to the GSS. The individual analysis
showed that resistance differed between treatments negligibly, and that some of the
chemicals had serious effect on the strains more than the resistant population therefore
they showed no resistance (i.e. RF < 1) as detailed in Tables 3 – 8.
Table 3. Selection for resistance and susceptibility with fenazaquin, in population of
Tetranychus urticae strains
Strain na Slope ± SE LC50 (g / L)
(0.95 CL)
LC90 (g / L)
(0.95 CL)
RF50b RF90
b
GSS 528 0.122 ± 0.009 12.048
(11.034-13.162)
11.129
(10.058-12.309)
__ __
NOR4 654 0.119 ± 0.008 22.784
(20.949-25.101)
21.629
(19.684-24.165)
1.10 1.1
NOR5 328 0.076 ± 0.009 7.447
(5.318-9.412)
24.402
(20.948-29.677)
0.67 1.13
CL confidence limit a Number of nymphs; b
resistance factor (RF) = LC50 or LC90 of laboratory collected population / LC50 or
LC90 of susceptible population (GSS)
In table 3 above, NOR4 exhibited low level of resistance to fenazaquin with RF value of
1.10 (i.e. RF >1) compared to GSS. NOR5 showed no resistance to the chemical
compared with the control (GSS).

16
Fenazaquin
0,0
20,0
40,0
60,0
80,0
100,0
120,0
1 2 3 4 5
Cons
% m
orta
lity
NOR4 NOR5 GSS
Figure 2: Resistance monitoring in three Tetranychus urticae strains with doses of fenazaquin at different concentration rates as mentioned in table 3. In the figure above, blue = NOR4, maroon = NOR5 and cream = GSS. In fig. 2 above, NOR5 showed the highest mortality rate compared to GSS, while NOR4
exhibited low mortality compared to the resistance strain.

17
Table 4. Selection for resistance and susceptibility with fenpyroximate, in population of
Tetranychu urticae strains
CL confidence limit
a Number of nymphs; bresistance factor (RF) = LC50 or LC90 of laboratory collected population / LC50 or
LC90 of susceptible population (GSS)
When fenpyroximate was applied at the recommended rate against NOR4, the strain
showed no resistance to the acaricide. Evidently, the RF value of 0.62 at LC50 falls
below the reference strain RF value of 1. NOR5 also showed no resistance against the
chemical compared with GSS.
Strain na Slope ± SE LC50 (g / L)
(0.95 CL)
LC90 (g / L)
(0.95 CL )
RF50b RF90
b
GSS 626 0.043 ± 0.003 30.346
(27.556-33.499)
59.946
(54.519-66.99)
__ __
NOR4 474 0.08 ± 0.007 18.709
(16.877-20.795)
34.637
(31.211-39.364)
0.62 0.58
NOR5 370 0.045 ± 0.005 22.743 (19.507-
26.301)
51.046
(44.966-59.714)
0.75 0.85

18
Fenpyroximate
0,0
20,0
40,0
60,0
80,0
100,0
120,0
1 2 3 4 5
Cons
% m
orta
lity
NOR4 NOR5 GSS
Figure 3: Resistance monitoring in three Tetranychus urticae strains with doses of fenpyroximate at different concentration rates as mentioned in table 4. In the figure above, blue = NOR4, maroon = NOR5 and cream = GSS. In fig. 3, NOR4 strain showed the highest mortality rate at the recommended dosage
compared to the resistance strain GSS because the lower the RF value as shown in tab. 4,
the higher the mortality. NOR5 showed a higher mortality rate at the same recommended
dose of fenpyroximate application compared with GSS. See table 4 above.

19
Table 5. Selection for resistance and susceptibility with hexythiazox, in population of
Tetramychus urticae strains
CL Confidence limit aNumber of nymphs; b Resistance factor (RF) = LC50 or LC 90 of laboratory collected population / LC50 or
LC90 of susceptible population (GSS)
In the trial, NOR4 showed a little more of resistance factor (RF) value of 1.20 at LC50
when hexythiazox was used to control it compared with the GSS value of 1. The NOR5
strain also exhibited a slightly higher level of resistance to hexythiazox with RF value of
1.24 compared with GSS.
Strain
ηa
Slope ± SE
LC50 ( g/100L)
(0.95CL)
LC90 ( g/100L)(0.95CL)
RF50 b
RF90 b
GSS 633 17.592 ± 1.382 0.087 (0.079-0.095)
0.159 (0.146-0.178)
__ __
NOR4 429 9.884 ± 0.895 0.103 (0.088-0.118)
0.232 (0.207-0.267)
1.20 1.47
NOR5 453 10.678 ±0.950 0.108
(0.095-0.122) 0.228
(0.205-0.259) 1.24 1.43

20
Hexythiasox
0,0
20,0
40,0
60,0
80,0
100,0
120,0
1 2 3 4 5
Cons
% m
orta
lity
NOR4 NOR5 GSS
Figure 4: Resistance monitoring in three Tetranychus urticae strains with doses of hexythiazox at different
concentration rates and a control as mentioned in table 5. In the figure above, blue = NOR4, maroon =
NOR5 and cream = GSS.
In figure 4, both NOR4 and NOR5 showed a lower mortality rate then the GSS strain
when treated with hexythiazox.

21
Table 6.Selection for resistance and susceptibility with clofentezine, in population of Tetranychus
urticae strains
Strain na Slope ± SE LC50 (g / L) (0.95 CL)
LC90 (g / L) (0.95 CL)
RF50b RF90
b
GSS 518 0.084 ± 0.008 15.44 (13.833-17.413)
30.539 (27.144-5.299)
__ __
NOR4 562 0.066 ± 0.007 22.33 (19.736 25.998)
41.687 (36.046-0.295)
1.45 1.37
NOR5 480 0.071 ± 0.008 19.061
(16.924-21.88) 37.196 (32.459- 4.282)
1.23 1.22
CL confidence limit, a Number of nymphs; bresistance factor (RF) = LC50 or LC90 of laboratory collected population / LC50 or
LC90 of susceptible population (GSS)
In table 6 above, it was observed that at LC50, the NOR4 strain showed a little higher
level of resistance to clofentezine with RF value of 1.45 compared with GSS. NOR5 also
exhibited a little more of resistance with RF value of 1.23 compared to GSS.

22
Clofentezine
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
1 2 3 4 5
Cons
% m
orta
lity
NOR4 NOR5 GSS
Figure 5: Resistance monitoring in three Tetranychus urticae strains with doses of clofentezin at different concentration rates as mentioned in table 6. In the figure above, blue = NOR4, maroon = NOR5 and cream = GSS.
Figure 5 showed that the mortality rate for NOR4 was lower than GSS. Similarly the
mortality rate for NOR5 was lower than the control (GSS). The result shown in tab. 6
implies that both NOR4 and NOR5 were resistant to clofentezine than the control
because their RF values were higher than GSS.
23
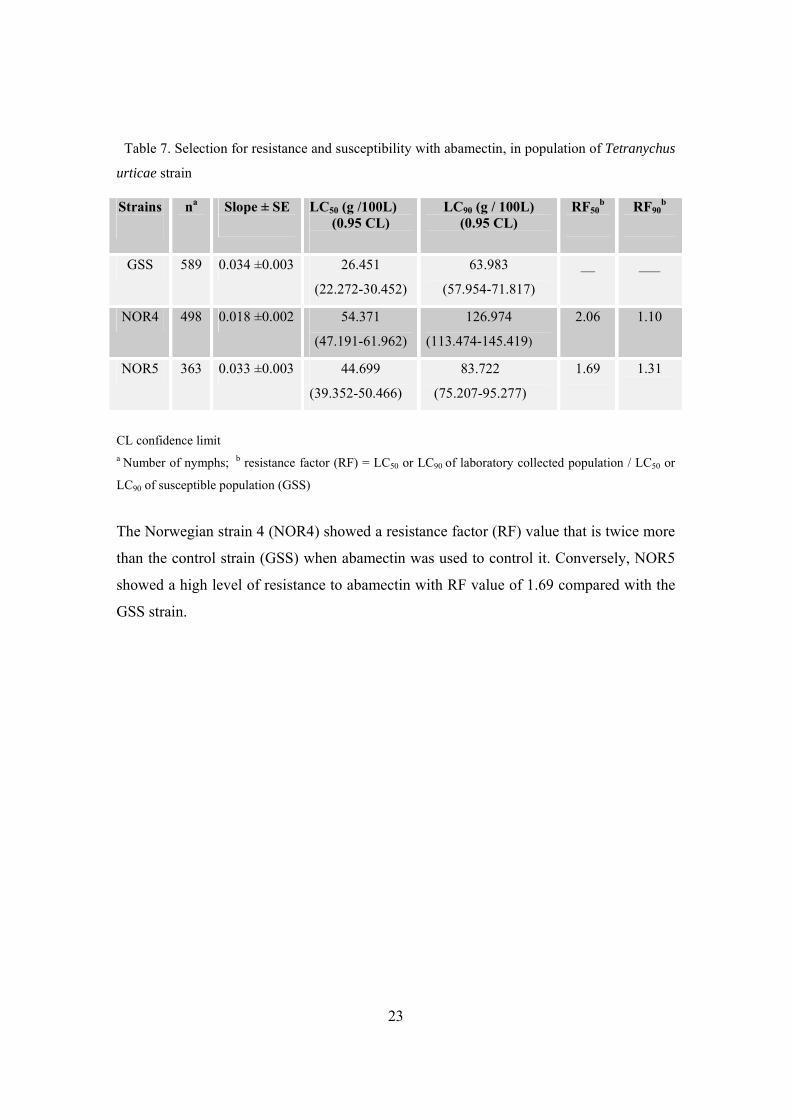
Table 7. Selection for resistance and susceptibility with abamectin, in population of Tetranychus
urticae strain
CL confidence limit a Number of nymphs; b resistance factor (RF) = LC50 or LC90 of laboratory collected population / LC50 or
LC90 of susceptible population (GSS)
The Norwegian strain 4 (NOR4) showed a resistance factor (RF) value that is twice more
than the control strain (GSS) when abamectin was used to control it. Conversely, NOR5
showed a high level of resistance to abamectin with RF value of 1.69 compared with the
GSS strain.
Strains
na
Slope ± SE
LC50 (g /100L) (0.95 CL)
LC90 (g / 100L) (0.95 CL)
RF50b
RF90
b
GSS 589 0.034 ±0.003 26.451
(22.272-30.452)
63.983
(57.954-71.817)
__ ___
NOR4 498 0.018 ±0.002 54.371
(47.191-61.962)
126.974
(113.474-145.419)
2.06 1.10
NOR5 363 0.033 ±0.003 44.699
(39.352-50.466)
83.722
(75.207-95.277)
1.69
1.31
24
Abamectin
0,0
20,0
40,0
60,0
80,0
100,0
120,0
1 2 3 4 5
Cons
% m
orta
lity
NOR4 NOR5 GSS
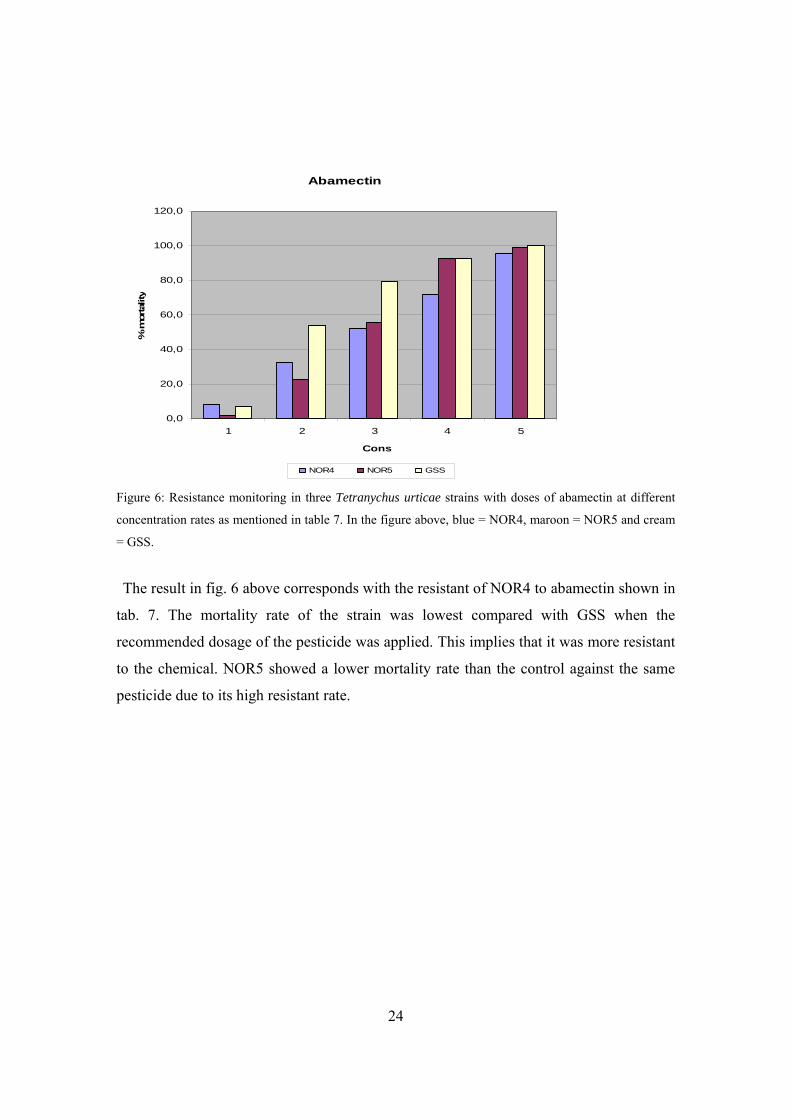
Figure 6: Resistance monitoring in three Tetranychus urticae strains with doses of abamectin at different
concentration rates as mentioned in table 7. In the figure above, blue = NOR4, maroon = NOR5 and cream
= GSS.
The result in fig. 6 above corresponds with the resistant of NOR4 to abamectin shown in
tab. 7. The mortality rate of the strain was lowest compared with GSS when the
recommended dosage of the pesticide was applied. This implies that it was more resistant
to the chemical. NOR5 showed a lower mortality rate than the control against the same
pesticide due to its high resistant rate.

25
Table 8. Selection for resistance and susceptibility with spirodiclofen, in population of
Tetranychus urticae strains
CL Confidence limit a Number of nymphs; bresistance factor (RF) = LC50 or LC90 0f laboratory collected population / LC50 or
LC90 of susceptible population (GSS)
In table 8, NOR4 showed no resistance to spirodiclofen with RF value of 0.83 compared
to the GSS. NOR5 equally showed no resistance with RF value of 0.83 at the same
concentration compared to the control.
Strain
na Slope ± SE LC50 (g / 100L)(0.95 CL)
LC90 (g / 100L(0.95 CL)
RF50b RF90
b
GSS
522 41.736 ± 10.196 0.012 (0.009-0.017)
0.042 (0.031-0.075)
__ __
NOR4 495 58.903 ± 10.395 0.01 (0.008-0.013)
0.032 (0.025-0.045)
0.83 0.76
NOR5 373 48.345 ± 11.776 0.01
(0.008-0.015) 0.037
(0.027-0.064) 0.83 0.88

26
Spirodiclofen
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
90,0
100,0
1 2 3 4 5
Cons
% m
orta
lity
NOR4 NOR5 GSS
Figure 7: Resistance monitoring in three Tetranychus urticae strains with doses of spirodiclofen at different concentration rates as mentioned in table 8. In the figure above, blue = NOR4, maroon = NOR5 and cream = GSS. In figure 7, both NOR4 and NOR5 showed higher level of mortality to spirodiclofen
when it was used against them compared with GSS. This fact is shown in the result in
tab. 8 above where both strains exhibited no resistance (RF < 1) against the chemical at
the same concentration rate.

27
4. DISCUSSIONS A standard bioassay is conducted, according to Nauen et al., 2001, by spray application
of the spider mite larvae. The method is used because it is one of the best ways of
investigating the effect of foreign substances such as drugs in living organisms when
little is known about the mechanism. The potency of acaricides differs with mite species.
The effectiveness tests of the six pesticides on two Norwegian strains of the two-spotted
spider mite T. urticae Koch (Acari: Tetranychidae) show difference in resistance. They
revealed a vast range of no and low levels of resistance in the acaricides. Both strains
exhibit low level of resistance to the mite growth inhibitors. For the METI acaricides,
they showed susceptibility or little resistance.
The two acaricides, fenazaquin and fenpyroximate, are well known METI (mitochondrial
electron inhibitor) acaricides, and are potent foreign substances that function primarily by
hindering complex I (NADH: ubiquinone oxidoreductse) of the mitochondrial respiratory
chain (Hollingworth & Ahammadsahib, 1995). They are said to be widely used because
of their high potency against many mite species. However, some strains of Tetranychus
spp from Australia – T. urticae Koch (Herron et al., 1993) and Japan – T. kanzawai
Kishide (Goka, 1998) have been reported to show over 100-fold resistance to the
compounds. Furthermore, two UK strains (TUK4 and TUK5) used in a study by Devine
et al. (2001) were said to be the first for which control failures for METI – acaricides in
Europe have been confirmed. It is therefore clear from the above studies and others that
the effectiveness of METI – acaricides is at risk, particularly where the use of pesticides
interacts with ecological and genetic factors which encourage the development of
resistance (Devine et al., 2001). In this investigation, both Norwegian strains of the two-
spotted spider mite (NOR4 and NOR5) showed no resistance to fenpyroximate. NOR4
exhibited low level of resistant to fenazaquin, while NOR5 showed no resistance in the
bioassay. The resistance factor value at LC50 for NOR4 when fenpyroximate was applied
against it was 0.62, while that of NOR5 was 0.75 indicating that the GSS had a higher
tolerance. With regards to fenazaquin, NOR4 showed a little higher level of resistance
(RF value of 1.10) at LC50 which is greater than GSS, whilst NOR5 showed no resistance

28
(RF 0.67 < 1) at LC50. The moderate level of resistance of NOR4 to fenazaquin may have
evolved from its ability to speedily adapt to pesticides when used against it for some
time. The susceptibility of NOR5 cannot easily be explained. Since the strains were
constantly kept under greenhouse condition, their adaptation to the condition as opposed
to field condition could also contribute to the resistance of NOR4 to fenazaquin, and the
no resistance of both NOR4 and NOR5 to fenpyroximate observed.
.
Hexyhtiazox and clofentezin cause little or no loss of life to adult female spider mite, but
they act as limitation to their laying of many fertile eggs (Chapman & Marris, 1986).
Nymphs hatched from the affected eggs do not undergo normal molting due to the
disruption of chitin synthesis (Flexner et al., 1995; Dekeyser & Downer, 1994). However,
resistance to these chemicals has been noticed in both the European red mite and the two-
spotted spider mite (Nauen et al., 2001). In the current investigation, both NOR4 and
NOR5 show low level of resistance to hexythiazox and clofentezin as seen in Tables 5
and 6. The resistance is due to the strains been capable of adjusting to the effect of the
mite growth inhibitor acaricides. Yet, nothing can be said concerning the mechanisms
resulting in T. urticae resistance against these acaricides (Stumpf, 2001).
Abamectin acts as nematicide, acaricide and insecticide (Putter et al., 1981). It acts on the
γ - aminobutyric acid (GABA) and glutamate-gated chloride channels, leading to
vigorousness of the chloride ion channel at higher concentrations and paralysis of the
pests (Bloomquist, 2001; Fritz et al., 1979). Spider mite, especially the two-spotted spider
mite, T. urticae, are the main aim of abamectin, and it is one of the most potent chemicals
hostile to the nymphs of a susceptible T. urticae strain (Nauen et al., 2001). This
investigation shows NOR4 and NOR5 strains to exhibit a resistance factors that were
twice more and very higher than the susceptible GSS population as seen in Table 7. The
highest resistant in the above strains may be due to the ability of spider mites to speedily
adjust to xenobiotic when applied against them. Stumpf & Nauen (2001), state that these
biological mechanisms are difficult to examine and almost nothing is known regarding
the mechanisms conferring abamectin resistance. Nauen et al. (2001) elucidate that T.
urticae are major target organisms of abamectin; but Compos et al. (1995), and Compos

29
et al. (1996) say resistance to abamectin in T. urticae is known from California, Florida
and the Netherlands. Furthermore, Stumpf et al. (2001) explain that the genetically
established resistance mechanisms in spider mites were similar to those found in insects,
and has increased degeneration of acaricides through esterases, glutathione S-
transferases (GSTs), or cytochrome P450 – dependent monooxygenases (MFO) (metabolic
resistance), and molecular changes in the target site, so that the target is no longer
sensitive to pesticide inhibition (target site resistance). Note that the two strains originate
from the greenhouse where they may have adapted to diverse conditions over a period of
time.
Spirodiclofen is one of the many new pesticides that are effective against all growth
stages and mature female of the tetranychid mite such as the two-spotted spider mite,
Tetranychus urticae. It is said not to exhibit cross-resistance to OP’s, METI’s,
hexythiazox and abamectin in well-distinguished species with high resistance to at least
one of the above chemicals (Rauch & Nauen, 2003). As a member of the tetronic acid
acaricide group, it has no adverse effect on insects of benefit and causes little harm to
predatory mites; it is different from some other pesticides in that it has short milieu
staying power (DT50 0.5-5.5 days) and a low mammalian injury (Fischer et al., 1993;
Wachendorff et al., 2000; Nauen et al., 2002). The two Norwegian strains (NOR4 and
NOR5) show no resistance when spirodiclofen was used against them.
5. Conclusion
The resistance in the two Norwegian populations of the two-spotted spider mite
(Tetranychus urticae) (NOR4 and NOR5) for the six selected acaricides (see section 2.2)
vary contingent on their mode of actions compared with the resistance strain (GSS).The
average mortality obtained after 48 hours in the control was 6.82 %.
Strain NOR4 exhibited low level of resistance to fenazaquin with RF value of 1.10
compared with GSS, and NOR5 showed no resistance to the poison with RF value of
0.67. Both NOR4 and NOR5 exhibited no resistance to fenpyroximate with RF values of

30
0.62 and 0.75 respectively compared with GSS. The resistance factor (RFs) for NOR4
and NOR5 strains when hexythiazox was applied were at low level (i.e. 1.18 and 1.24)
with respect to GSS. NOR4 and NOR5 showed low level of resistance factor of 1.45, and
1.23 respectively to clofentazine than GSS. When abamectin was used in the trial, the Rf
value for NOR4 strain was two times more than the reference strain GSS, with a resistant
factor of 2.06 which was the highest resistance of the six meticides. NOR5 strain gave a
moderate level of RF value of 1.69 at LC50 compared with GSS. NOR4 and NOR5
showed no resistance against spirodiclofen when it was applied with RF values of 0.83
and 0.83 respectively. Both NOR4 and NOR5 showed high resistant to abamectin whilst
they exehibited no resistanc to spirodiclofen and fenpyroximate.
One can therefore suggest that the acaricides used in this trial, if used to control the two-
spotted spider mite, should be used in combination with other insecticides, or should not
be used at tall. For instance, fenpyroximate and spirodiclofen could be used with their
exact recommended rates or be included in integrated pest management (IPM)
programmes for increased potency. Biological control method could be an alternative
way of controlling the two spotted spider mite. For instance; the use of Thrips of various
species which are considered as most important predators of spider mites and eggs (Van
de Vrie et al., 1972), and the use of natural enemy fungi like Neozygites floridana that
kills the two spotted spider mite (Klingen & Westrum, 2007).

31
6. REFERENCES
Abbott, W. S. 1925. A method of computing the effectiveness of an insecticide. J. Econ.
Entomol. 18 : 265.
Ahmed, S. & R. M. Wilkins. 2002. Studies on some enzymes involved in insecticide
resistance in fenitrothion-resistance and susceptible strains of Musca domestica L. (Dipt,
Muscidae). Journal of applied Entomology-Zeitschrift fur Angewandte Entomology, 126:
510-516.
Ballantyne, G. H. & R. A. Harrison. 1967. Genetic and biochemical comparisons of
organophosphate resistance between strains of spider mites (Tetranychus species: Acari).
Entomologia experimentalis et applicata. 10: 231-239.
Barkette, L. P. 1994. Management Guide for Low-Input Sustainable Apple Production.
Diane Publiching Co.
Beers, E. H., H. Riedl & J. E. Dunley. 1998. Resistance to abamectin and reversion to
susceptibility to fenbutatin oxide in spider mite (Acari: Tetranychidae) populations in the
Pacific Northwest. J. Economic Entomology 91: 352-360.
Berge, J. B., R. Feyereisen & M. Amichot. 1998. Cytochrome P450 monooxygenases and
insecticide resistance in insects. Philosophical Transactions of the Royal Society of
London Series B Biological Sciences. 353: 1701-1705.
Blank, R. H. 1979. Studies on the non-specific esterase and acetylcholinesterase
isozymes electrophoretically separated from the mites Sancassania berlesei
(Tyroglyphidae) and Tetranychus urticae (Tetranychidae). New Zealand Journal of
Agricultural Research. 22: 497-506.

32
Bloomquest, J. R. 2001. GABA and glutamate receptors as biochemical sites for
insecticide action, in “Biochemical Sites of Insecticide Action and Resistance” (I.
Ishaaya, Ed.), pp. 17-41. Springer-Verlag, Berlin.
Bloomquist, J. R. 1994. Cyclodiene resistance at the insect GABA receptor / chloride
channel complex confer broad cross-resistance to convulsants and experimental
phenylpyrazole insecticides. Arch. Insect Biochem. Physiol. 26(1): 17-25.
Brogdon, W. G. & J. C. McAllister. 1998. Insecticide resistance and vector control.
Centers for Disease Control and Prevention, Atlanta, Georgia, USA. Vol 4: 605-613.
Brown, A. W. M. & R. Pal. 1971. Insecticide resistance in arthropods: World Health
Organization Monograph Series 38, Geneva, World Health Organization, 491.
Bylemans, D. & F. Meurrens. 1997. Anti-resistance strategies for the Two-spotted spider
mite, Tetranychus urticae (Acari: Tetranychidae) in strawberry culture. Acta. Hort., 439
(2), 869-876.
Campbell, P.M., R. Newcomb, R. J. Russell & J. G. Oakeshott. 1998. Two different
amino acid substitutions in the ali-esterase, E3, confer alternative types of
organophosphorus insecticide resistance in the sheep blowfly Luculia cuprina. Insect
Biochemistry and Molecular Biology, 28: 139-150.
Campos, F., D. A. Krupa & R. A. Dybas. 1996. Susceptibility of populations of two-
spotted spider mites (Acari: tetranychidae) from Florida, Holland and the Canary Islands
to abamectin and characterization of abamectin resistance. J. Economic entomology 89:
594-610.
Capua, S., E. Cohen & U. Gerson. 1990. Non-specific esterase in mites – a comparative
study. Comparative Pharmacology and Toxicology. 96: 125-130.

33
Carey, J. R. & J. W. Bradley. 1982. Development rates, vital schedules, sex ratios, and
life tables for Tetranychus urticae, T. turkestani and T. pacificus (Acarina:
Tetranychidae) on cotton. Acarologia, 23: 333-345.
Carlini, E. J., B. A. Mcpheron, C. M. Felland & L. A. Hull. 1991. Elevated esterase
activity in resistant tufted apple bud moth, Platynota idaeusalis (Walker) (Lepidoptera:
Tortricidae). Comp. Biochem. Physiol. 99C: 375-377.
Chapman, R. B. & J. W. M. Marris. 1986. The sterilizing effect of clofentezine and
hexythiazox on female two-spotted spider mite. Proc. 39th N. Z. Weed and Pest Control
Conf.: 237-240.
Chen, W-L. & C. N. Sun. 1994. Purification and characterization of carboxylesterases of
a rice brown planthopper, Nilaparvata lugens Ståal. Insect. Biochem. Molec. Biol. 24:
347.355.
Claudianos, C., R. J. Russell & J. Oakeshott. 1999. The same amino acid substitution in
orthologous esterases confers organophosphate resistance on the housefly and a blowfly.
Insect Biochemistry and Molecular Biology, 29: 675-686.
Compose, F., R. A. Dybas & D. A. Krupa. 1995. Susceptibility of two spotted spider mite
(Acari: Tetranychidae) populations in California to abamectin. J. Econ. Entomol. 88: 225.
Cranham, J. E. & W. Helle. 1985. Pesticide resistance in Tetranychidae. In Spider mites:
Their Biology, Natural Enemies and Control. Vol. 1B (W. Helle and M. W. Sabelis,
eds.). Elsevier, Amsterdam, pp. 405 – 421.
Croft, B. A. & H. E. van de Baan. 1988. Ecological and genetic factors influencing
evolution of pesticide resistance in tetranychid and phytoseiid mites. Exp. Appl. Acarol.
4: 277-300.

34
Crooker, A. 1985. Embryonic and Juvenile Development; in: Helle, W. & M. W. Sabelis
(eds). 1985a. Spider Mites their Biology, Natural Enemies and Control. Elsevier,
Amsterdam, vol. 1A, 403pp.
Cygler M., J. D. Schrag, J. L. Sussman, M. Harel, I. Silman, & M. K. Gentry, et al. 1993.
Relationship between sequence conservation and three-dimensional structure in a large
family of esterases, lipases and related proteins. Protein Sci ; 2: 366-82.
Daborn, P., S. Boundy, J. Yen, B. Pittendrigh & R. H. ffrench-Constant. 2001. DDT
resistance in Drosophila correlates with Cyp6g1 over-expression and confers cross
resistance to the neonicotinoid imidacloprid. Molecular Genetics and Genomics, 266:
556-563.
Dekeyser, M. A. & R. G. H. Downer. 1994. Biochemial and physiological targets for
miticides. Pestic. Sci 40: 85-101.
Devine, G. J., M. Barber & I. Denholm. 2001. Incidence and inheritance of resistance to
METI-acaricides in European strain of the two-spotted spider mite (Tetranychus urticae)
(Acari: Tetranychidae). Pest Manag. Sci. 57: 443-448.
Edge V.E., J. Rophail & D. G. James. 1987. Acaricide resistance in two-spotted mite,
Tetranychus urticae in Australian horticultural crops. In: Proceedings symposium on mite
control in horticultural crops (W. G. Thwaite. Ed), Department of Agriculture, Orange,
N. S. W., Australia, pp. 87-91.
Eldefrawi, M. E., R. Miskus & V. Sutcher. 1960. Methylenedioxyphenyl derivaties as
synergists for carbamate insecticides on susceptible, DDT- and parathion-resistant house
flies. Journal of Economic Entomology, 53: 231-234.

35
Field, L. M., A. L. Devonshire & B. G. Forde. 1988. Molecular evidence that insecticide
resistance in peach-potato aphids (Myzus persicae Sulz.) results from amplification of an
esterase gene. Biochem. J. 251: 309-312.
Fischer, R., T. Bretschneider, B. W. Krueger, J. Bachmann, C. Erdelen, U. Wachendorff-
Neumann, H. J. Santel, K. Luerssen & R. R. Schmidt. 1993. Preparation and insecticidal,
acaricidal, herbicidal and fungicidal activities of 3-aryl-4-hydroxy-3-dihydrofuranone and
–thiophenone derivatives, German Patent DE 4216814.
Fjelldalen, J. & C. Stenseth. 1962. Resistance to chemicals in Two-spotted spider mite
(Tetranychus urticae Koch) in Norway. Norwegian Plant Protection Institute. Div.of
Entomol. Vollebekk. Report No.21. 13: 267-283.
Flexner, J. L., P. H. Westigard, R. Hilton & B. A. Croft. 1995. Experimental evaluation
of resistance management for two-spotted spider mite (Acari: Tetranychidae) on Southern
Oregon pear: 1987-1993. J. Econ Entomol 88: 1517-1524.
Fournier, D., J. M. Bride, C. Mouchès, M. Raymond, M. Magnin, J-B. Bergè, N. Pasteur
& G. P. Georghiou. 1987. Biochemical characterization of the esterases A1 and B1
associated with organophosphate resistance in Culex pipiens L. complex. Pestic.
Biochem. Physiol. 27: 211-217.
Fournier, D., J. M. Bride, M. Poirie, J-B. Bergé & F. W. Plapp. 1992. Insect glutathione
S- transferases: biochemical characteristics of the major forms from houseflies
susceptible and resistant to insecticides. J. Biol. Chem. 267: 1804-1845.
Fritz, L. C., C. C. Wang, & A. Gorio. 1979. Avermectin B1 a irreversibly blocks
postsynaptic potentials at the lobster neuromuscular junction by reducing muscle
membrane resistance, Proc. Natl. Acad. Sci. USA 76(4): 2062-2066.

36
Georghiou, G. P. & A. Lagunes-Tejeda. 1991. The Occurrence of Resistance to
Pesticides in Arthropods: an Index of Cases Reported through 1989. Food and
Agricultural Organization of the United Nations, Rome.
Goka, K 1998. Mode of inheritance of resistance to three new acaricides in the kanzawa
spider mite, Tetranychus kanzawai Kishida (acari: Tetranychidae). Exp.Appl. Acar. 22
(12) 699-708.
Helle, W. & M. W. Sabelis (eds). 1985a. Spider Mites: Their Biology, Natural Enemies
and Control. vol. 1A, Elsevier, Amsterdam, 403 pp.
Hemingway, J. & S. H. P. P. Karunaratne. 1998. Mosquito carboxylesterases: a review of
the molecular biology and biochemistry of a major insecticide resistance mechanism.
Medical Veterinary Entomology, 12: 1-12.
Hemingway, J., N. J. Hawkes, L. McCarroll & H. Ranson. 2004. The molecular basis of
insecticide resistance in mosquitoes. Insect Biochemistry and Molecular Biology, 34:
653-665.
Hemingway, J. 1982. The biochemical nature of malathion resistance in Anopheles
stephensi from Pakistan. Pesticide Biochemistry and Physiology, 17: 149-155.
Herath, P. R. J., J. Hemingway, I. S. Weerashinghe & K. G. I. Jayawardena. 1987. The
detection and characterization of malathion resistance in the field populations of
Anopheles culicifacies. Pesticide Biochemistry and Physiology, 29:157-162.
Herbert, H. J. 1981 a. Biology, life tables, and innate capacity for increase of the two-
spotted spider mite, Tetranychus urticae (Acarina: Tetranychidae). Can. Entomol., 113:
371 – 378.

37
Herron, G., V. Edge, L. Wilson & J. Rophail. 1993. Clofentezine and hexythiazox
resistance in Tetranychus urticae Koch in Australia. Exp Appl Acarol 17:433-440.
Herron, G. A. & J. Rophail. 1998. Tebufenpyrad (Pyranica (R)) resistance detected in
Two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) from apples in
western Australia. Exp Appl Acarol 22: 633-641.
Hillesheim, E., W. Ritter & D. Bassand. 1996. First data on resistance mechanisms of
Varroa jacobsoni(Oud) against tau-flualinate. Experimental and Applied Acarology. 20:
283-296.
Hollingworth, R. M. & K. I. Ahammadsahib. 1995. Inhibitors of respiratory complex I:
Mechanisms, pesticidal actions and toxicology. Rev Pestic Toxicol 3: 277-302.
Hoy, M. A. & Y. L. Ouyang. 1989. Selection of the western predatory mite, Metaseiulus
occidentalis (Acari: Phytoseiidae), for resistance to abamectin. J. Econ Entomol 82(1):
35-40.
Jeppson, L. R., H. H. Keifer, & E. W. Baker. 1975. Mites injurious to economic plants.
University of California Press, Berkeley, CA, USA, 614 pp.
Kasai, S. & J. G. Scott. 2000. Over expression of cytochrome P450 CYP6D1 is associated
with monooxygenase- mediated pyrethroid in housefly from Georgia. Pesticide
Biochemistry and Physiology. 68: 34-41.
Kettermen, A. J., K. G. I. Jayawardena & J. Hemingway. 1992. Purification and
characterization of a carboxylesterase involved in insecticide resistance from the
mosquito Culex quinquefasciatus. Biochm. J. 287: 355-360.
Kim, S. S. & S. C. Lee. 1990. Korean Journal of Applied Entomology. 29: 170-175.

38
Klingen, I. & K. Westrum. 2007. The effect of pesticides used in strawberries on the
phytophagous mite Tetranychus urticae (Acari: Tetranychidae) and its fungal natural
enemy Neozygites floridana (Zygomycetes: Entomophthorales) Biological control 43:
222-230.
Knight, A. L., E. H. Beers, S. C. Hoyt & H. Rield. 1990. Acaricide bioassay with spider
mites (Acari: Tetranychidae) on Pome fruits: evaluation of methods and selection of
discrimination concentrations for resistance monitoring. J. Econ. Entomol: 83: 1752-
1760.
Knowles, C. O. 1997. Mechanisms of resistance to acaricides. In: Molecular Mechanisms
of Resistance to Agrochemicals, ed. by Sjut v. vol 13, springer-verlga, Berlin, Heidelberg,
Germany. Pp. 57 – 77.
Koehn, R. K. & B. L. Bayne. 1989. Towards a physiological and genetical understanding
of the energetics of the stress response. Biological Journal of the Linnean Society 37:
157-171.
Kuwahara, M. 1982. Insensitivity of the acetylcholinesterase from the organophosphate-
resistant Kanzawa spider mite, Tetranychus kanzawai kishidi (Acarina: Tetranychidae),
to organophosphorus and carbamate insecticides. Applied Entomology and Zoology. 17:
4,486-493.
Le Goff, G., S. Boundy, P. J. Daborn, J. L. Yen, L. Sofer, R. Lind, C. Sabourault, L.
Madi-Ravazzi & R. H. ffrench-Constant. 2003. Microarray analysis of cytochrome P450
mediated insecticide resistance in Drosophila. Insect Biochemistry and Molecular
Biology, 33: 701-708.
Maitra, S., S. M. Dombroski, L. C. Waters & R. Ganguly. 1996. Three second
chromosome-linked clustered Cyp6 genes show differential constitutive and barbital-

39
induced expression in DDT-resistant and susceptible strains of Drosophila melanogaster.
Gene; 180: 165-71.
Matsumura F., K. Tanaka & Y. Ozoe. 1987. GABA-related systems as targets for
insecticides. In: Hollingworth, R. M. & M. B. Green (eds). Sites of actions for neurotoxic
pesticides. American Chemical Society, Washington DC, pp 44–70.
Michigen State University.Athropod Pesticide Resistance Database. 2004-2007. Address:
(http://www.pesticideresistance.org/search/12/536/0/
Mota-Sanchez, D., P. S. Bills, & M. E. Whalon. 2002. Anthropod Resistance to
Pesticides: Status and Overview. Center for Integrated Plant Systems, Michigan State
University. East Lansing, Michigan, USA; In: Wheeler, W. B (eds). 2002. Pesticides in
Agriculture and the Environment. Marcel Dekker, Inc. New York. Basel. USA. correct
Motoyama, N. & W. C. Dauterman. 1975. Interstrain comparison of glutathione-
dependent reactions in susceptible and resistant houseflies. Pestic. Biochem. Physiol. 5:
489-495.
Mouchès, C., N. Pasteur, J. B. Berge, O. Hyrien, M. Raymond, et al. 1986. Amplification
of an esterase gene is responsible for insecticide resistance in a Californian Culux
mosquito. Science 233: 778-80.
Nauen, R., A. Buchholz, N. Stumpf & B. Lawson. 2002. Mode of action of the acaricide
spirodiclofen on Tetranychus urticae (Acari: Tetranychidae), abstr 10th Int Congr Chem
Crop Prot, IUPAC, Basel, 3c-12.
Nauen, R., N. Stumpf, A. Elbert, C. P. W. Zebitz, & W. Kraus. 2001. Acaricide toxicity
and resistance in larvae of different strains of Tetranychus urticae and Panonychus ulmi
(Acari: Tetranychidae), Pest Manag. Sci. 57(3): 253-261.

40
Nauen, R., N. Stumpf, & A. Elbert. 2000. Efficacy of BAJ 2740, a new acaricidal tetronic
acid derivative, against tetrantchid spider mite species resistant to conventional
acaricides. Proceedings of the Brighton Crop Protection Conference on Pests and
Diseases. pp. 453-458.
Nordhus, E. G. M. 2005. Molecular methots in apecies identification of Liriomyza spp.
and the study of insecticide resistance in Liriomyza spp., Bemisia tabacia and Myzus
persicae. PhD. Scientiarum Thesis. Norwegian University of Life Sciences, University of
miljø – og biovitenskap.
Oakeshott, J. G., E. A. van Papenrecht, T. M. Boyce, M. J. Healy & R. J. Russell. 1993.
Evolutionary genetics of Drosophila esterases. Genetica; 90: 239-68.
Ozawa, A. 1994. Acaricide susceptibility of Kanzawa spider mite, Tetranychus kanzawai
Kishida (Acarina: Tetranychidae) collected from tea fields in Chuuen and Ogasa district
in Shizuoka Prefecture. Bull Tea Res. Stn. 29: 1-14. [in Japanese].
Piorié, M., M. Raymond & N. Pasteur. 1992. Identification of two distinct amplifications
of the esterase B locus in Culex pipiens (L.) moequitoes from Mediterranean countries.
Biochem. Genet. 30: 13-26.
Polk, D. 1994. Insects and Mite Management; in: L. P. Barkette. 1994. Management
Guide for Low-Input Sustainable Apple Production. Diane Publiching Co.
Prapanthadara, L., J. Hemingway & A. J. Ketterman. 1995. DDT- resistance in Anopheles
gambiae (Diptera, Culicidae) from Zanzibar, based on insecticide DDT-
dehydrochlorinase activity of glutathione S-transferases, Bulletin of Entomological
Research, 85: 267-274.
Putter, J. G., F. A. MacConnell, A. A. Preiser, Haidri, et al. 1981. Avermectins: Novel
insecticides, acaricides and nematicides from a soil microorganism. Experientia 37, 963.

41
Rauch, N, & R. Nauen. 2003. Spirodiclofen resistance risk assessment in Tetranychus
urticae (Acari: Tetranychidae): a biological approach. Pesticide Biochemistry and
Physiology 74: 91-101.
Rose R. L., L.Barbhaiya, R. Roe, G. C. Rock & E. Hodgson. 1995. Cytochrome P450-
associated insecticide resistance and the development of biochemical diagnostic assays in
Heliothis virescens. Pestic. Biochem. Physiol. 51:178–191.
Roush, R. T. & J. A. Mckenzie. 1991. Ecologial genetics of insecticide and acaricide
resistance. Annu Rev Entomol 32: 361-380.
Salinas, A. E. & M. G. Wong. 1999. Glutathion S-transferases – A review. Current
Medical Chemistry. 6: 279-309.
Sato, M. E., T. Miyata, M. D. Silva, A. Raga & M. F. de Souza Filho. 2004. Selections
for fenpyroximate resistance and susceptibility, and inheritance, cross-resistance and
stability of fenpyroximate resistance in Tetranychius urticae Koch (Acari:
Tetranychidae). Appl.Entomol. Zool. 39: 293-302.
Sawicki, R. M. 1987. Definition, detection and documentation of insecticide resistance.
In: Combating Resistance to Xenobiotics (Ford, M. G., D. W. Holloman, B. P. S.
Khambay, & R. M. Sawicki, eds) Ellis Horwood: Chichester, England, pp 105-117.
Scott, J. A. 1995. The Molecular-genetics of resistance: Resistance as a response to
stress. Kampling-Bushland USDA Livestock Insect Research Laboratory; Agricultural
Research Service. Kerrville, TX 78028-9184. Florida Entomologist, 78: 399-414.
Scott, J. G. 1991. Insecticide resistance in insects. Handbook of Pest Management in
Agriculture (ed. By D. Pimentel), pp. 663. CRC Press, Boca Ration.

42
Smissaert, H. R., S.Voerman, L. Oostenbrugge & N. Renooy. 1970.
Acetylcholinesterases of organophosphate-susceptible and resistant spider mites. Journal
of Agriculture and Food Chemistry 18: 66-75.
Stenseth, C. 1965. Multi-resistant two-spotted spider mite (Tetranychus urticae Koch.)
and its reproductive capacity compared to susceptible and parathion resistant mites.
Norwegian Plant Institute. Div. of Entmol. Vollebekk Report No. 30.
Stumpf, N. 2001. Acaricide resistance in the phytophagus spider mites Tetranychus
urticae and Panonychus ulmi: Risk assessment and biochemical mechanisms. Der
University Hohenheim, Boblingen, Germany. Doctorate Thesis.
Stumpf, N. & R. Nauen. 2001. Cross-resistance, inheritance, and biochemistry of
mitochondrial electron transport inhibitor-acaricide resistance in Tetranychus urticae
(Acari: Tetranychidae). J. Econ. Entomol. 94: 1577-1583.
Stumpf, N., C. P. W. Zebitz, W. Kraus, G. D. Moores, & R. Nauen. 2001. Resistance to
organophosphates and biochemical genotyping of acetylcholinesterases in Tetranychus
urticae (Acari: Tetranychidae). Pestic. Biochem. Physiol. 69: 131.
Sundukov, O. V., I. V. Zil-Bermints, L. S. Golovkina, & K. V. Novozhilov. 1989.
Problemy Izbiratel’nosti Deistviya Insektitsidov I Akaritsidov I Ee Znachenie V
Zashchite Rastenii. 19:64-69.
Tomlin, C. D. S. 1997. The Pesticide Manual. Farnham, UK: Br Crop Production
Council.
Tomlin, C (ed). 2003. The Pesticide Manual, 13th edn. Crop Protection Publications.
British Crop Protection Council. Farnham, Surrey, UK. 1341 pp.

43
Van de Vrie, M., J. A. McMurtry, & C. B. Huffaker. 1972. Ecology of Tetranychid mites
and their natural enemies: A review. III. Biology, ecology and pest status and host plant
relations of Tetranychids. Hilgardia. 41: 343 – 432.
Vontas, J. G., G. J. Small & J. Hemingway. 2001. Glutathione S-transferases as
antioxidant defence agents confer pyrethroid resistance in Nilaparvata lugens.
Biochemical Journal, 357:65-72.
Voss, G. & F. Matsumura. 1964. Resistance to organophosphorus compounds in the two-
spotted spider mite: two different mechanisms of resistance, Nature 202: 319-320.
Wachendorff, U., E. Bruck, A. Elbert, R. Fischer, R. Nauen, N. Stumpf & R. Tiemann.
2000. BAJ2740, a novel broad spectrum acaricide, in Proc Brighton Crop Prot Conf-
Pests Dis, BCPC, Farnham, Surrey, UK, pp53-58.
Welthy, C., W. H. Reissig, T. J. Dennehy & R. W. Weires. 1988. Comparison of residual
bioassay methods and criterir for assessing mortality of cyhexatin-resistant European red
mite (Acari: Tetranychidae) J. Econ. Entomol. 81(2): 442-448.
Weyda, F., J. Sula & M. Gresner. 1984. Sbornik Uvtiz Ochrana Rostlin. 20: 123-129.
Wilkinson, C. F. 1976. Insecticide biochemistry and physiology. New York: Plenum
Press; p. 768.
Wood, E., B. Latli & J. E. Casida. 1996. Fenazaquin acaricide specific binding sites in
NADH: ubiquinone oxidoreductase and apparently the ATP synthase stalk. Pestic
Biochem Physiol 54: 135-145.
Zahavi, M. & Tahori, A. S. 1970. Sensitivity of acetylcholinesterase in spider mites to
organophosphorus compounds. Biochemical Pharmacology. 19: 219-225.

44
Zhang, Z – Q. 2003. Mites of Greenhouses: Identification, Biology and control.CABI
Publishing, Wallingford, UK, xii+244 pp.
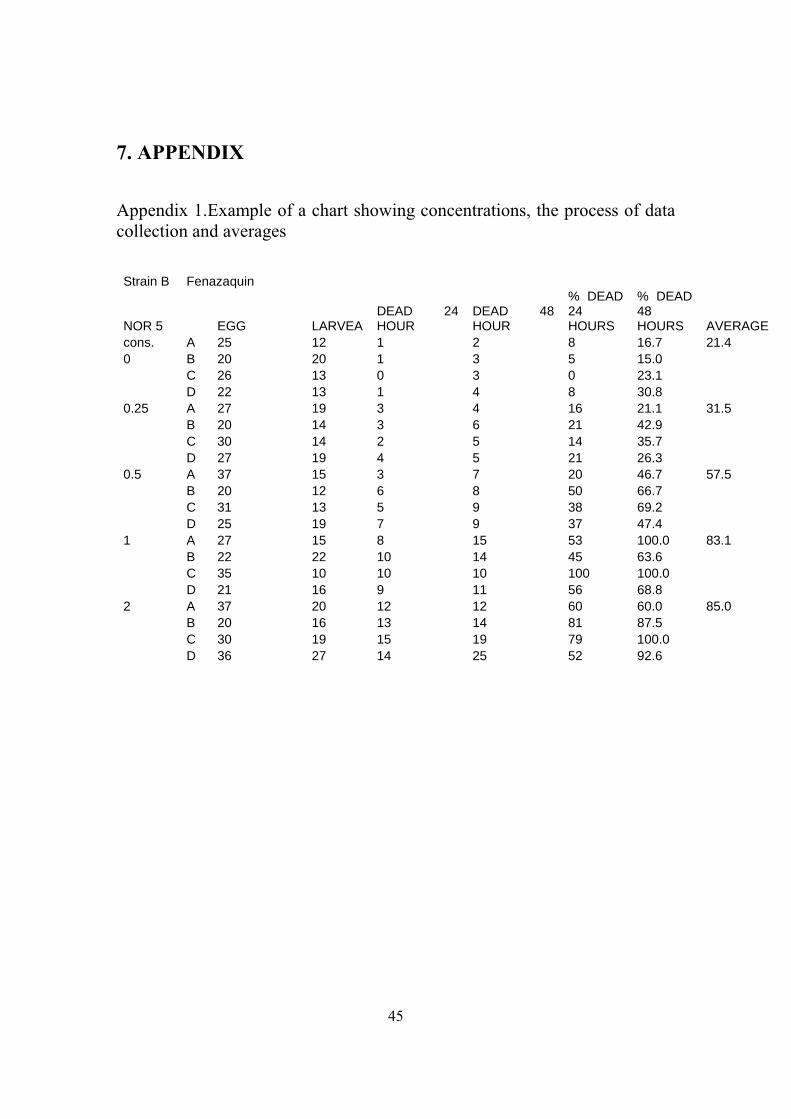
45
7. APPENDIX Appendix 1.Example of a chart showing concentrations, the process of data collection and averages
Strain B Fenazaquin
NOR 5 EGG LARVEADEAD 24 HOUR
DEAD 48 HOUR
% DEAD 24 HOURS
% DEAD 48 HOURS AVERAGE
cons. A 25 12 1 2 8 16.7 21.4 0 B 20 20 1 3 5 15.0 C 26 13 0 3 0 23.1 D 22 13 1 4 8 30.8 0.25 A 27 19 3 4 16 21.1 31.5 B 20 14 3 6 21 42.9 C 30 14 2 5 14 35.7 D 27 19 4 5 21 26.3 0.5 A 37 15 3 7 20 46.7 57.5 B 20 12 6 8 50 66.7 C 31 13 5 9 38 69.2 D 25 19 7 9 37 47.4 1 A 27 15 8 15 53 100.0 83.1 B 22 22 10 14 45 63.6 C 35 10 10 10 100 100.0 D 21 16 9 11 56 68.8 2 A 37 20 12 12 60 60.0 85.0 B 20 16 13 14 81 87.5 C 30 19 15 19 79 100.0 D 36 27 14 25 52 92.6

46
Appendix 2: Examples of Abbot’s correction (a-c) Appendix 2-a
Strain C Spirodiclofen Abbot GSS EGG LARVEA DEAD 24
HOUR DEAD 48 HOUR
% DEAD 24 % DEAD 48 AVERAGE corrected
cons. A 29 25 2 3 8 12.0 13.9 0 B 30 24 2 4 8 16.7 C 40 30 1 4 3 13.3 D 36 22 0 3 0 13.6 0.25 A 28 21 2 5 10 23.8 25.2 13.1 B 36 34 4 4 12 11.8 C 33 28 2 8 7 28.6 D 40 19 3 7 16 36.8 0.5 A 35 23 5 11 22 47.8 42.0 32.6 B 37 30 7 14 23 46.7 C 31 24 5 8 21 33.3 D 39 25 6 10 24 40.0 1 A 28 19 4 12 21 63.2 50.5 42.5 B 35 29 8 14 28 48.3 C 40 32 9 13 28 40.6 D 33 32 10 16 31 50.0 2 A 30 28 10 23 36 82.1 84.7 82.2 B 28 17 12 17 71 100.0 C 40 28 11 22 39 78.6 D 37 32 17 25 53 78.1

47
Appendix 2-b Abbot Tot. Tot. Tot. Not Mortality % not AVERAGE corrected treated dead dead % dead 13.9 101 14 87 13.9 86.1 25.2 13.1 102 24 78 25.2 74.8 42.0 32.6 102 43 59 42 58 50.5 42.5 112 55 57 50.5 49.5 84.7 82.2 105 87 18 84.7 15.3

48
Appendix 2-c Mort Dose Mort. Average 0 13.9 0.25 13.1 0.25 25.2 0.5 32.6 0.5 42 1 42.5 1 50.5 2 82.2 2 84.7








![PROPOSED EXPANSION FOR SPECIALTY CHEMICALS & …environmentclearance.nic.in/writereaddata/Online/TOR/21_Nov_2019... · (trifluoromethyl)benzene propanoate [400882-07-7] Acaricide](https://img.pdfslide.us/doc/110x75/6064b6b7a624a92d9f1c19f1/proposed-expansion-for-specialty-chemicals-trifluoromethylbenzene-propanoate.jpg)



















